
Submitted:
23 December 2025
Posted:
24 December 2025
You are already at the latest version
Abstract
Despite over two centuries of debate, the morphological nature of the grass embryo’s parts remains unresolved. The bipartite interpretation of the grass cotyledon suggests that it is composed of two parts: the scutellum and the coleoptile. According to Wu et al. (2024, 2025), the cotyledon in maize is organized similarly to the vegetative leaf, comprising scutellum and coleoptile, homologous to the leaf blade and sheath, respectively. This view, which Wu et al. (2024, 2025) erroneously associate with the bipartite model of grass cotyledon, is morphologically impossible because within the embryo, the coleoptile is located above the scutellum and therefore cannot be homologous to the latter’s sheath that is suppressed. Within a bipartite context, the coleoptile may only be homologous to the leaf ligule or represent a de novo outgrowth of the scutellum’s reduced sheath, thus lacking homology with mature leaf structures. Molecular evidence from the mesocotyl and the leaf collar region is essential for evaluating the bipartite nature of the grass cotyledon. The genetic results of Wu et al. (2024) do, in fact, support the view that the coleoptile is the next leaf on the embryo’s axis after the scutellum, thus disproving the bipartite view of the cotyledon in Poaceae.
Keywords:
grass embryo
; coleoptile
; mesocotyl
; ligule
; scutellum
; plant morphology
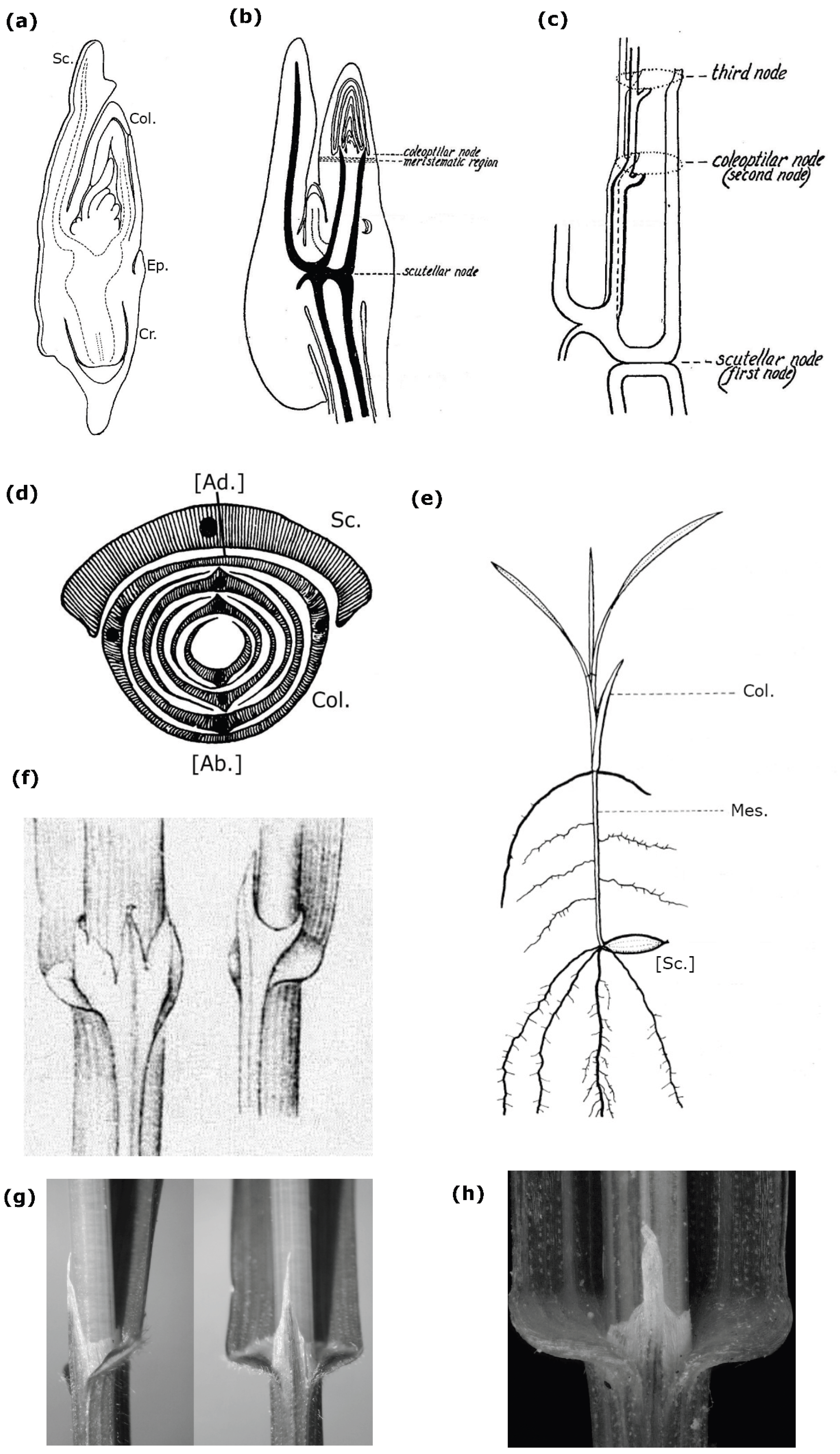
Both grass embryos and seedlings consist of the following parts: the scutellum, which connects the embryo to the seed’s endosperm; the coleoptile, a two-keeled cap that covers the plumule (embryo’s bud); the coleorhiza, sometimes described as a “root sheath”; and the epiblast, an embryonic scale located on the side of the embryo opposite the scutellum (Figure 1a). Not all these structures are equally developed across the various taxa within the Poaceae, and there is still no unified viewpoint regarding the morphological nature of the grass embryo’s parts (Brown, 1960; Kaplan & Specht, 2022, but see Tillich, 2007).
Drawing on genetic and extensive transcriptomic evidence, Wu et al. (2024, 2025) argue that the bipartite interpretation of the grass cotyledon (hereinafter BICG)—which regards the scutellum and coleoptile as its two components—best aligns with their findings. BICG was supported, at least at some stage of their careers, by nearly all leading late-19th- to 21st-century morphologists who wrote on the topic (e.g., Van Tieghem, 1872; Čelakovský, 1897; Goebel, 1900; Arber, 1934; Johansen, 1950; Tillich, 2007; Kaplan & Specht, 2022; see also Brown's (1960) review). Below, we examine BICG as it was explicitly formulated in the works of the listed morphologists and, in this formulation, became part of the history of morphology. We leave aside the rare contrasting interpretations of the bipartite cotyledon in Poaceae (e.g., Bugnon, 1921; see also Brown, 1960), which warrant separate study and are only distantly related to the main topic of this note.
In a paragraph "Homology of the maize cotyledon", Wu et al. (2024) concluded that, “… In the same way that the grass leaf comprises a distal blade fused to a proximal sheath to form a single lateral organ (phytomer), we propose that the grass cotyledon comprises a distal scutellum and a proximal coleoptile fused to form a single cotyledon.” They summarized: "Genetic, RNA-targeting, and morphological evidence supports models for cotyledon evolution where the scutellum and coleoptile, respectively, comprise the distal and proximal domains of the highly modified, single grass cotyledon.” (Wu et al., 2024). Wu et al. (2025) pointed out that their “… co-expression analyzes, RNA targeting assays, and genetic analyzes of meristem mutants collectively support the hypothesis that the cotyledon is a bipartite organ comprising an apical, haustorial scutellum fused to a basal, emergent and sheathing coleoptile”. The latter statements contradict BIGC, and the conclusions of Wu et al. (2024, 2025) must be reconsidered.
In the seedling of any hypogeal monocotyledon plant, the embryonic bud is wrapped with the expanded base of the cotyledon, the cotyledonary sheath (Velenovský, 1907; Sargant & Arber, 1915; Serebryakov, 1952), the apex of the cotyledon becomes a haustorium (sucker), absorbing endosperm nutrients (Velenovský, 1907; Sargant & Arber, 1915; Serebryakov, 1952). The sucker and sheath are connected by a stalk, which may be completely reduced (Velenovský, 1907; Sargant & Arber, 1915; Serebryakov, 1952). This view of monocot seedling morphology does not imply definitive claims about the homology of embryonic structures (Sargant & Arber, 1915) and can be considered primarily functional and descriptive. Therefore, the descriptive statement that the haustorial scutellum and protective coleoptile represent the “distal” and “proximal” regions of a unified grass cotyledon (Wu et al. 2024) is not a theoretical model requiring validation (Wu et al. 2024, 2025), but rather a straightforward observational conclusion. As already stressed by Sargant & Arber (1915, p. 164, bold is mine), "... botanists would probably have found no difficulty in considering the scutellum as the sessile sucker of the cotyledon, and the coleoptile as its sheath." Therefore, no genetic or extensive transcriptomic data are required to prove the latter observation.
Although the coleoptile is typically described as a “cotyledonary sheath” (Van Tieghem, 1872; Čelakovský, 1897; Goebel, 1900; Velenovský, 1907; Serebryakov, 1952; Roth, 1955; Kaplan & Specht, 2022), which is perhaps clear from the etymology of the term “coleoptile” (Kaplan & Specht, 2022), it is positioned above the scutellum (Figure 1b, c, d), specifically on the same side of the embryonic axis as the scutellum (Figure 1d), rather than above and opposite to it (Čelakovský, 1897; Goebel, 1900; Serebryakov, 1952; Roth, 1955; Skvortsov, 1977; Kaplan & Specht, 2022). The latter observation creates a challenge for interpreting the coleoptile as a leaf of axis next to scutellum, as its position disrupts the embryo's distichous phyllotaxis (Figure 1d) (Čelakovský, 1897; Goebel, 1900; Kaplan & Specht, 2022, see also Wu et al., 2024), particularly if the epiblast is ignored (Roth, 1955). The BIGC offers a solution to this issue: if the coleoptile, which lies above the scutellum, is still a part of the cotyledon, then the embryo distichy remains saved because the first leaf of the plumule is situated on the opposite side of the morphological axis of the seedling relative to the scutellum (Figure 1d) (Čelakovský, 1897; Goebel, 1900; Serebryakov, 1952; Roth, 1955; Kaplan & Specht, 2022). Thus, the position of the coleoptile above the scutellum, rather than below it, provides one of the key morphological premises for BIGC itself. Therefore, within BIGC, the location of the coleoptile above the scutellum cannot be disputed. The position of the coleoptile within the embryo can also be explained by its function as a protective cap for the plumule, which must cover the first plumule leaf and, for this reason, lies above the scutellum rather than below it, but below the first plumole leaf (Figure 1b, c). In other words, it is difficult to imagine how a leaf sheath located below the apical bud could simultaneously serve as a protective cap for the latter.
From that, drawing an exact parallel between the coleoptile and a leaf sheath (Wu et al. 2024, 2025) is morphologically impossible, as the sheath of the leaf cannot be positioned above its blade. If the coleoptile is a modified scutellum’s sheath (Wu et al. 2024, 2025), then in grass seedlings with a developed mesocotyl, which is the segment of the embryo’s axis between the scutellum and the coleoptile (Čelakovský, 1897; Goebel, 1900; Velenovský, 1907; Sargant & Arber, 1915; Arber, 1934; Serebryakov, 1952; Roth, 1955; Kaplan & Specht, 2022) (Figure 1e), the sheath may be positioned above the leaf blade at a distance up to three dozen of centimeters (Arber, 1934). Regrettably, the issue of mesocotyl development, perhaps the key problem of BICG (Van Tieghem, 1872; Velenovský, 1907; Sargant & Arber, 1915; Roth, 1955; Serebryakov, 1952), was not even mentioned by Wu et al. (2024, 2025), even though in maize mesocotyl is well developed (Arber, 1934; Hoshikawa, 1969), and no comparative molecular data from the mesocotyl, stem nodes, or internodes have been generated by them.
Thus, within BIGC, the scutellum (the modified unifacial cotyledonary blade) is indeed homologous to the blade of the mature leaf (Čelakovský, 1897; Goebel, 1900; Arber, 1934; Johansen, 1950) while the coleoptile, the cotyledonary sheath, is homologous not to the leaf sheath, but to the ligule of the same leaf (Figure 1f) (Čelakovský, 1897; Goebel, 1900; Arber, 1934; Johansen, 1950) whose morphological sheath is suppressed (Čelakovský, 1897; Goebel, 1900; Arber, 1934; Johansen, 1950). “Ligulartheorie” is simply another name for BIGC (Roth, 1955).
Johansen (1950) offered a classically clear summary of the core idea behind the BIGC: “Most workers agree that the coleoptile, scutellum, and epiblast are all parts of one structure, the cotyledon, and that they represent, respectively, the ligule, lamina, and auricles of the vegetative leaf. The sheathing leaf base disappears, or at last is merely indicated, in the earliest embryonic stage of development” (Johansen, 1950, p. 270; see also Čelakovský, 1897, bold is mine). Therefore, it is precisely the correspondence between the coleoptile and the leaf ligule (Figure 1f-h) that should have been examined first and foremost using genetic and transcriptomic analyses, if the study aims to validate the correctness of the BIGC. However, Wu et al. (2024, 2025) do not present any relevant data for the leaf collar region of maize.
An alternative and more recent viewpoint within BIGC suggest that the coleoptile is not a ligule, but a closed de novo tubular outgrowth of the marginal tissues of the reduced sheath of the scutellum (Skvortsov, 1977; Tillich, 2007). Within this alternative, the coleoptile must be considered an organ that lacks homologous among the morphological structures of the mature grass leaf (Skvortsov, 1977). This interpretation of coleoptile, which does not mean or imply that the latter is homologous to the sheath itself, was also not considered in the studies by Wu et al. (2024, 2025).
Wu et al. (2024) present data supporting the homology between the coleoptile and the leaf sheath, without considering that, if the coleoptile is indeed homologous to a latter, the corresponding leaf can only be the one following the scutellum - and certainly not the sheath of the scutellum itself, which, as previously noted, is suppressed (Čelakovský, 1897; Goebel, 1900; Arber, 1934; Johansen, 1950; Tillich, 2007). Consequently, any arguments in favor of the homology between the coleoptile and the leaf sheath (Wu et al., 2024) simultaneously serve as arguments not for, but rather against the BIGC. Therefore, genetic results of Wu et al. (2024) in fact support the classical statment that the scutellum represents the first leaf of the embryo (the cotyledon) and the coleoptile (the cotyledonary sheath) is the second or third (following the epiblast) phyllome of the embryo’s axis (Bruns, 1892; Velenovský, 1907, 1913; Avery, 1930; Serebryakov, 1952; Roth, 1955; Guignard & Mestre 1969), thus arguing against BIGC. The cluster (scutellum + coleoptile) identified in the WGCNA analyses in Wu et al. (2025) is well explained by the functional specialization of these structures as embryonic organs and by their development from the grass proembryo (reviewed in Johansen, 1950) rather than from the shoot apex meristem.
The discussion of Wu et al.’s (2024, 2025) results in the context of grass embryo interpretations other than BIGC raises concerns about Wu et al.’s (2024, 2025) limited consideration of these alternatives. Fundamentally, the latter studies focused on reconciling the obtained genetic and transcriptomic results with the BIGC framework as it was erroneously sketched by them, rather than addressing the necessary methodological question, namely, which of the available interpretations of the grass embryo (Roth, 1955; Brown, 1960) best corresponds with the obtained outcomes.
In conclusion, Wu et al. (2024, 2025) conflate the descriptive and morphological meanings of the term "cotyledonary sheath” and therefore lack both interpretative context and necessary data to test the validity of BIGC. The central structural proposition of the Wu et al. (2024, 2025) studies, that cotyledon in Poaceae is organized similarly to the mature grass leaf, comprising a scutellum and a coleoptile, homologous to the modified leaf blade and sheath, respectively is morphologically impossible and does not reflect the meaning of BIGC.
Abbreviations
Ab., abaxial, and Ad., adaxial (coleoptile, Col.).; Cr., coleorhiza; Ep., epiblast; Mes., mesocotyl; Sc., scutellum.
References
- Arber, A. The Gramineae: A Study of Cereal, Bamboo and Grass; Cambridge University Press: Cambridge, 1934. [Google Scholar]
- Avery, GS, Jr. Comparative anatomy and morphology of embryos and seedlings of maize, oats, and wheat. Botanical Gazette 1930, 89, 1–39. [Google Scholar] [CrossRef]
- Brown, WV. The morphology of the grass embryo. Phytomorphology 1960, 10, 215–223. [Google Scholar]
- Bruns, E. Der Grasembryo; Munich: Val. Höfling, 1892. [Google Scholar]
- Bugnon, P. La feuille chez les Gramineae. Mémoires de la Société Linnéenne de Normandie 1921, 21, 1–107. [Google Scholar]
- Čelakovský, LJ. Über die Homologien des Grasembryos. Botanische Jahrbücher 1897, 55, 141–174. [Google Scholar]
- Goebel K von. Organographie der Pflanzen, insbesondere der Archegoniaten und Samenpflanzen; G. Fischer: Jena, 1900. [Google Scholar]
- Guignard, JL; Mestre, JC. L’origine du cotylédon et du cône végétatif de la tige chez les Monocotylédones. Bulletin de la Société Botanique de France 1969, 116, 207–214. [Google Scholar] [CrossRef]
- Hoshikawa, K. Underground organs of the seedlings and the systematics of Gramineae. Botanical Gazette 1969, 130, 192–203. [Google Scholar] [CrossRef]
- Johansen, DA. Plant Embryology: Embryogeny of the Spermatophyta; Chronica Botanica Co: Waltham, MA, 1950. [Google Scholar]
- Kaplan, D; Specht, CD. Kaplan's Principles of Plant Morphology; CRC Press: New York, 2022. [Google Scholar]
- Roth, I. Zur morphologischen Deutung des Grasembryos und verwandter Embryotypen. Flora 1955, 142, 564–600. [Google Scholar] [CrossRef]
- Sargant, E; Arber, A. The comparative morphology of the embryo and seedling in the Gramineae. Annals of Botany 1915, 29, 161–222. [Google Scholar] [CrossRef]
- Serebryakov, IG. Morphology of the Vegetative Organs of Higher Plants; Soviet Science: Moscow, 1952. [Google Scholar]
- Skvortsov, AK. Once more on the morphological nature of the parts of grass embryo and seedling. Bulletin of the Moscow Society of Naturalists, Biological Series 1977, 82, 96–111. [Google Scholar]
- Tillich, H-J. Seedling diversity and the homologies of seedling organs in the order Poales (Monocotyledons). Annals of Botany 2007, 100, 1413–1429. [Google Scholar] [CrossRef] [PubMed]
- Van Tieghem, P. Observations anatomiques sur le cotylédon des Graminées. Annales des Sciences Naturelles, Botanique 1872, 15, 236–276. [Google Scholar]
- Velenovský, J. Vergleichende Morphologie der Pflanzen, Bds. II & IV (Supplement); F. Řivnáč: Praga, 1907, 1913. [Google Scholar]
- Wu, H; Zhang, R; Scanlon, MJ. Genetic analyses of embryo homology and ontogeny in the model grass Zea mays subsp. mays. New Phytologist 2024, 243, 1610–1619. [Google Scholar] [CrossRef] [PubMed]
- Wu, H; Zhang, R.; Niklas, KJ; Scanlon, MJ. Multiplexed transcriptomic analyses of the plant embryonic hourglass. Nature Communications 2025, 16, 802. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(a) Median longitudinal section through a wheat embryo. Modified from Roth (1955, Figure 1). (b) Zea mays L.: Germinating embryo, 24 hours old, showing elongation of the axis between the level of attachment of the scutellum (“scutellar node”) and that of the coleoptile (“coleoptilar node”), which is positioned above the scutellum. Modified from Avery (1930, Figure 19). The expression “level of attachment,” the term “node,” and the original handwriting are from Avery (1930). (c) Z. mays: Sequence of attachment levels (“nodes”) of the scutellum, coleoptile, and the leaf adjacent to the coleoptile in the seedling, reconstructed from the procambial vascular system of the latter. Only those bundles of the seedling vascular system that originate, at least in part, from the scutellar bundle are shown. Similarly to (b), diagram (c) illustrates that the coleoptile, functioning as a bud-protective cotyledonary sheath, is positioned above the scutellum - the only spatial arrangement that allows it to serve as a protective cup for the emerging plumule. Modified from Avery (1930, Figure 39). See the clarifying note in legend (b). (d) Diagram showing the leaf arrangement in the grass embryo plumule, modified from Skvortsov (1977, Figure 3.4). Relative to the scutellum, the first leaf of the plumule is positioned on the side opposite the embryo axis. (e) Schematic illustration of a grass seedling with a well-developed mesocotyl. Modified from Hoshikawa (1969, Figure 1). The scutellum (Sc.), functioning as a sucker, is located inside the seed. (f)–(h) Closed ligule of Melica L. (Poaceae) species, a tubular structure that encircles the stem, and interpreted as one of the presumed homologues of the coleoptile within the BICG (Čelakovský, 1897). (f) Ligules of M. alsissima L. (left-hand illustration) and M. uniflora Retz. (right-hand illustration); image modified from the original drawing by Čelakovský (1897, Table 4.16, 17). (g) and (h) Ligule of M. uniflora; (g) modified from photo by S. Lefnaer https://commons.wikimedia.org/wiki/File:Melica_uniflora_sl7.jpg ; (h) modified from photo by M. Lučanova (https://www.botanickafotogalerie.cz/ ). Although ligules can be considered outgrowths on the adaxial surface of a bifacial leaf sheath (Tillich, 2007), they still can develop as a pipe-like formation encircling the axis (Čelakovský, 1897).
Figure 1.
(a) Median longitudinal section through a wheat embryo. Modified from Roth (1955, Figure 1). (b) Zea mays L.: Germinating embryo, 24 hours old, showing elongation of the axis between the level of attachment of the scutellum (“scutellar node”) and that of the coleoptile (“coleoptilar node”), which is positioned above the scutellum. Modified from Avery (1930, Figure 19). The expression “level of attachment,” the term “node,” and the original handwriting are from Avery (1930). (c) Z. mays: Sequence of attachment levels (“nodes”) of the scutellum, coleoptile, and the leaf adjacent to the coleoptile in the seedling, reconstructed from the procambial vascular system of the latter. Only those bundles of the seedling vascular system that originate, at least in part, from the scutellar bundle are shown. Similarly to (b), diagram (c) illustrates that the coleoptile, functioning as a bud-protective cotyledonary sheath, is positioned above the scutellum - the only spatial arrangement that allows it to serve as a protective cup for the emerging plumule. Modified from Avery (1930, Figure 39). See the clarifying note in legend (b). (d) Diagram showing the leaf arrangement in the grass embryo plumule, modified from Skvortsov (1977, Figure 3.4). Relative to the scutellum, the first leaf of the plumule is positioned on the side opposite the embryo axis. (e) Schematic illustration of a grass seedling with a well-developed mesocotyl. Modified from Hoshikawa (1969, Figure 1). The scutellum (Sc.), functioning as a sucker, is located inside the seed. (f)–(h) Closed ligule of Melica L. (Poaceae) species, a tubular structure that encircles the stem, and interpreted as one of the presumed homologues of the coleoptile within the BICG (Čelakovský, 1897). (f) Ligules of M. alsissima L. (left-hand illustration) and M. uniflora Retz. (right-hand illustration); image modified from the original drawing by Čelakovský (1897, Table 4.16, 17). (g) and (h) Ligule of M. uniflora; (g) modified from photo by S. Lefnaer https://commons.wikimedia.org/wiki/File:Melica_uniflora_sl7.jpg ; (h) modified from photo by M. Lučanova (https://www.botanickafotogalerie.cz/ ). Although ligules can be considered outgrowths on the adaxial surface of a bifacial leaf sheath (Tillich, 2007), they still can develop as a pipe-like formation encircling the axis (Čelakovský, 1897).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



