Submitted:
22 December 2025
Posted:
24 December 2025
You are already at the latest version
Abstract
Erythropoietin (EPO), the master regulator of erythropoiesis, is emerging as a pivotal mediator of brain repair. While its capacity to mitigate neural damage is well-documented, we posit that its most profound potential lies in actively orchestrating functional restoration. In the present review we summarize the molecular biology of EPO and the evidence establishing EPO as a potent modulator of neuroplasticity. We use an experimental strategy in which a specific behavioral task marks experience-activated neural circuits, and a subsequent, temporally precise administration of EPO provides a surge of plasticity-related proteins. This creates a synergistic interaction where the proteins are selectively captured by the activated synapses, directing plastic changes with high specificity. We present experimental evidence demonstrating that this synchronized protocol enables the recovery of spatial memory, reinstates synaptic plasticity, and activates genetic programs for plasticity in rodent models of brain injury. Furthermore, we show that endogenous EPO signaling is itself activity-dependent and integral to memory formation. This redefines EPO as a precision tool for neurorestoration, a potential now being pursued with engineered, non-erythropoietic variants of EPO in clinical trials for neurological and psychiatric disorders
Keywords:
erythropoietin
; neuroplasticity
; synaptic tagging and capture
; circuit rewiring
; neurorestoration
; cognitive recovery
1. Introduction
The XXth century witnessed a major transformation in the way we understand the nervous system. In the early years the brain was understood as an immutable organ, able to be damaged but not to recover (1). However, cumulating evidence during the second half of the century turned the concept; the nervous system is a constantly changing system, both functionally and structurally (2) under the influence of experience.
The experiments demonstrating thicker cortex in animals raised in complex environments (3), the discovery of a relationship between neural function and metabolism (4, 5) and that neurogenesis can still take place in the adult brain (6, 7) paved the way to a new understanding of the nervous system under the concept of neural plasticity.
Plasticity can take place at different levels, from the molecular to the systemic level, but one of the more relevant is the one modifying synaptic function. Synaptic plasticity can increase efficacy of synaptic transmission, as in Long-Term-Potentiation (8, 9). Since its discovery LTP has been considered a cellular mechanism of long-term memory (10, 11) showing mechanistic coincidences that support this assumption (12-15). Synaptic plasticity has also been claimed as a mechanism of recovery after brain lesion (16).
Drugs, neurotransmitters, trophic factors and hormones can modulate synaptic plasticity (17, 18). Erythropoietin (EPO) is a pleiotropic renal hormone that affects not only the production of red blood cells (19) but nerve cells function (20).
The interactions of plasticity influencing factors are time dependent. Studying LTP reinforcement in weakly activated synapses the existence of a time window for the modulation of previously “tagged” synapses was demonstrated (21-23).
The discovery of functional receptors for erythropoietin (EPO) on neurons (24-26) opened an unexpected frontier, prompting investigation into the roles of this classical hormone beyond its hematopoietic domain. A growing body of work from both experimental models and clinical trials has since substantiated that EPO possesses significant neuroprotective and plasticity-promoting effects.
In this review, we will delineate the molecular architecture of EPO signaling, evaluate its role in neuroprotection, and present an expanded focus on its capacity to drive neuroplasticity. We will argue that EPO’s most transformative application lies in its ability to promote the functional rewiring of neural circuits after injury or degeneration. We will synthesize evidence that positions EPO not as a mere shield against damage, but as a precision tool that can be strategically deployed to guide and reinforce the brain’s innate restorative processes.
Our work suggests that functional restoration based on synaptic plasticity may require two timely coordinated signals: first, a behavioral experience that activates synaptic circuits relevant for function, followed by a systemic plasticity-reinforcing factor, such as EPO, delivered within a critical time window. This two-signal strategy offers a powerful new perspective for targeted interventions in restorative neurology and psychiatry.
2. Erythropoietin: From Oxygen Sensing to Pleiotropic Signaling
In 2019, the Swedish Academy announced the awarding of the Nobel Prize in Physiology or Medicine to William G. Kaelin Jr., Sir Peter J. Ratcliffe, and Gregg L. Semenza “for their discoveries of how cells sense and adapt to oxygen availability” (sic: https://www.nobelprize.org/prizes/medicine/2019/press-release/). This is undoubtedly a contribution of enormous significance to understanding a key mechanism in homeostasis. (See Figure 1).
The physiological link between hypoxia, red blood cell production, and a humoral factor was first suggested over a century ago by Carnot and Deflandré. Their experiments revealed that plasma from anemic rabbits could stimulate erythropoiesis in healthy recipients, a factor they termed ‘hemopoietin’. This entity was later renamed erythropoietin following the work of Bonsdorf and Jalavisto (27, 28).
EPO orchestrates red blood cell production via a classic negative feedback loop (29): renal EPO release, triggered by tissue hypoxia, stimulates erythropoiesis in the bone marrow to improve oxygen delivery. In recent decades, however, it has become unequivocally clear that EPO is a pleiotropic cytokine with receptors and functions extending far beyond the hematopoietic system, including profound effects on the nervous system. Both animal studies and clinical investigations have supported its potential as a neuroprotective and neurorestorative agent (28, 30-46).
While the preponderance of research has focused on EPO’s capacity to prevent neural damage, this review emphasizes a different view. We present an updated synthesis that highlights the molecular biology of EPO and its emerging role in neuroplasticity, the nervous system’s inherent capacity for adaptive structural and functional change in response to experience, learning, or injury. This plasticity-centric view expands EPO’s therapeutic promise from merely preventing damage to actively restoring function after injury or in the context of neurodegeneration.
3. Molecular Biology of EPO and Its Central Nervous System Expression
Renal interstitial fibroblasts are the primary source of systemic EPO in response to hypoxia. EPO is a 30 kDa glycoprotein of 165 amino acids (27-29), whose circulatory half-life is extended by terminal sialic acid residues that shield it from recognition by galactose receptors on hepatocytes, the cells responsible for its clearance (47). Its erythropoietic effect stems from an anti-apoptotic action on erythroid progenitor cells expressing the EPO receptor (EPOR). Ligand binding triggers intracellular cascades, beginning with the activation of Janus tyrosine kinase 2 (JAK2), which in turn recruits pathways including MAPK, ERK, and PI3K/Akt, culminating in the upregulation of anti-apoptotic proteins like BCL-XL (48). This JAK2/STAT5 signaling pathway, upon EPO binding, transmits a potent survival signal to the cell nucleus, inhibiting programmed cell death (apoptosis) and promoting cellular growth and proliferation.
The EPO gene resides on the long arm of chromosome 7 (q11-q22). Its five exons encode a 193-amino acid prohormone, which is proteolytically processed upon secretion to yield the active circulating form. EPO gene transcription is regulated by a complex interplay of factors. A promoter in the 5’ region is activated by GATA-4 and repressed by GATA-2 and contains binding sites for Nuclear Factor-kappa B (NF-κB). The hypoxic induction of EPO is primarily mediated by a Hypoxia Response Element (HRE) in the 3’ region, which serves as a binding site for the Hypoxia-Inducible Factor (HIF) heterodimer (HIF-α/β). The HIF (Hypoxia-Inducible Factor) complex acts as a master regulator of the cellular response to low oxygen, and upon binding to the HRE, it activates the transcription of not only EPO but also a battery of genes involved in angiogenesis, glycolysis, and cell survival, such as Vascular Endothelial Growth Factor (VEGF), glucose transporters, and several glycolytic enzymes (28, 32). See Figure 2.
4. EPO and Neuroprotection: An Established yet Limited Frontier
The clinical use of recombinant human EPO (rhEPO) to correct the anemia of chronic kidney failure provided the impetus and tools to explore its extra-hematopoietic effects (49). Critical discoveries followed: EPO has actions beyond erythropoiesis (28), neurons possess functional EPO receptors (32), and the hormone can cross the blood-brain barrier (34), a highly selective semipermeable border of endothelial cells that prevents most substances in the blood from entering the brain, which is a critical requirement for any systemically administered neurotherapeutic. Furthermore, intrinsic production of EPO occurs within the brain itself, with neurons and, most prominently, astrocytes expressing the EPO gene, as demonstrated in humans, non-human primates, and rodents (50). See Figure 2.
These findings catalyzed intense research, revealing in models of cerebral hypoperfusion that EPO attenuates damage and reduces infarct volume (47), while also stimulating axonal sprouting (48). Comparable neuroprotection has been documented in traumatic brain injury models (51, 52), effects attributable, at least in part, to the hormone’s vasogenic action, its ability to promote the formation of new blood vessels (angiogenesis), thereby improving perfusion and oxygen delivery to compromised neural tissue (53, 54). This protective efficacy extends to neonatal models of hypoxia-ischemia (55) and has shown promise in clinical studies of hypoxic-ischemic encephalopathy in newborns (56).
A diverse and compelling body of evidence points to potential applications for EPO in a wide spectrum of disorders, including optic neuropathy (57), Friedreich’s ataxia (58), multiple sclerosis (59), sleep apnea (60), Alzheimer’s dementia (61), sudden unexpected death in epilepsy (SUDEP) (62), affective disorders (63, 64), and, in both animal models (65) and patients of Parkinson’s disease (66, 67). Its potential as a neuroprotectant in preterm neonates continues to garner significant attention (68).
Notwithstanding this promising pre-clinical landscape, the translation of EPO into routine clinical practice for neuroprotection faces substantial hurdles. Meta-analyses of existing studies frequently yield inconclusive results (69). Furthermore, definitive, rigorous clinical trials in critically ill neurological patients are lacking (70) and the administration of EPO to non-anemic individuals carries risks, including significant alterations in blood viscosity and peripheral resistance (71). In summary, the future of EPO as a neuroprotective agent remains a vigorously open and critical area for both clinical and basic research. A more comprehensive summary can be found in the review by Hemani and Lane (71).
However, the neuroprotective paradigm focused on mitigating acute damage represents a limited frontier. It often overlooks a fundamental challenge: for patients living with established neurological deficits, true recovery hinges on the brain’s capacity for adaptive neuroplasticity to restore lost function. It is here where we propose to move from the passive concept of protection to the active strategy of guided restoration.
5. EPO and Neuroplasticity: Introducing a Strategy for Tag-Guided Circuit Rewiring
Our approach fundamentally reorients the application of Erythropoietin from a generalized neuroprotectant to a targeted neurorestorative tool. The critical innovation lies not in EPO itself, but in its strategic application within a synchronized protocol directly inspired by the “synaptic tagging and capture” (STC) hypothesis.
Figure 3.
Graphic summary of the neurorestorative effects of EPO. bdnf: gene for brain-derived growth factor; arc: gene for Activity-regulated cytoskeleton-associated protein; epo: gene fpr erythropoietin; epor: gene for erythropoietin receptor.
Figure 3.
Graphic summary of the neurorestorative effects of EPO. bdnf: gene for brain-derived growth factor; arc: gene for Activity-regulated cytoskeleton-associated protein; epo: gene fpr erythropoietin; epor: gene for erythropoietin receptor.
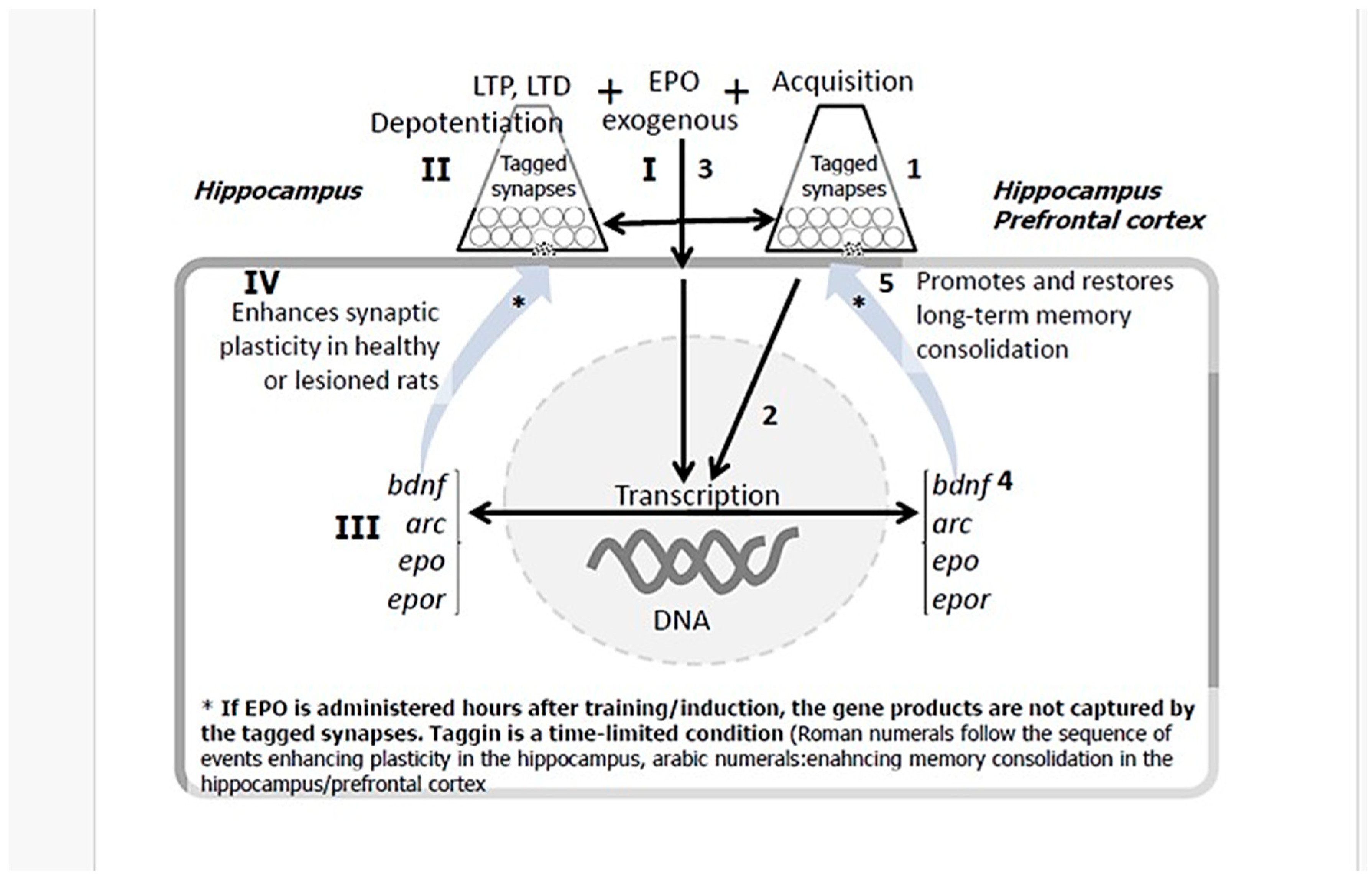
In this protocol, a specific behavioral task (e.g., spatial training) serves as the initial stimulus, tagging activated neural circuits that might be relevant for recovery. The subsequent, temporally precise administration of EPO acts as a powerful facilitator that provides a robust, exogenous surge of plasticity-related proteins (PRPs). This temporally precise coupling ensures that plastic changes are directed with high specificity to the activated circuits, enabling the de novo consolidation of long-term memories and the precise repair of neural pathways that underlie lasting neurological recovery.
This method moves beyond passive protection to active, circuit-specific rewiring. While the clinical and experimental evidence for EPO’s neuroprotective effect is substantial, the capacity for true functional recovery from established brain damage hinges on neuroplasticity. To directly investigate this capacity, our research group focused on cognitive processes and their cellular substrates, memory and synaptic plasticity, in both healthy animals and models of brain injury.
6. EPO Promotes Spatial Memory Recovery and Extends Memory Duration
The synchronized protocol was first tested in a model of established brain damage using a fimbria-fornix (FF) lesion. This tract connects the hippocampus with essential subcortical cholinergic, noradrenergic, and serotonergic inputs, and its injury causes a severe and persistent spatial memory deficit (72, 73), modeling aspects of Alzheimer’s dementia (74-76). Crucially, daily administration of EPO 10 min after training in the Morris water maze, a timing aligned with synaptic tagging mechanisms, produced significant recovery of spatial learning in FF-lesioned rats. Retention tests confirmed a stronger memory trace in EPO-treated injured animals. The specificity of this effect is paramount: administration 5 h post-training was ineffective, as was immediate post-injury administration, which would represent a purely neuroprotective strategy (77).
This finding was replicated and extended using the object place recognition test, a low-stress, single-trial spatial memory task. Again, a single EPO dose administered 10 min after acquisition facilitated learning in injured animals and, remarkably, prolonged memory duration from 24 h to 72 h in uninjured animals. The 5-h delay once again abolished the effect (78). Most recently, we demonstrated that this single, correctly timed EPO dose can prolong memory for at least 21 days (79), suggesting it promotes not only synaptic consolidation but also systemic remote memory, a process where memories, initially dependent on the hippocampus, become stabilized and stored in cortical networks for the long-term (80, 81).The consistent efficacy of the 10-min post-tagging window, and the consistent failure of the 5-h delay, underscores the temporal precision required for EPO to effectively engage the tagging mechanism.
7. EPO Expands the Boundaries of Synaptic Plasticity
Given that learning and recovery are ultimately mediated by changes in synaptic strength, we directly investigated EPO’s effects on synaptic plasticity in the dentate gyrus, focusing on long-term potentiation (LTP) and long-term depression (LTD). LTP and LTD are considered the primary cellular models for information storage in the brain, representing a long-lasting increase or decrease, respectively, in synaptic strength between neurons (8, 9, 82). In healthy animals, we found that EPO administration alone induces a slow, progressive potentiation of synaptic transmission (83), akin to the effects of brain-derived neurotrophic factor (BDNF) (84-86). Furthermore, EPO pretreatment lowered the threshold for inducing both LTP (with high-frequency stimulation) and LTD (with low-frequency stimulation) and prevented the reversal of LTP (depotentiation) (83). This collective evidence indicates that EPO induces a metaplastic state, expanding the dynamic range and stability of synaptic plasticity irrespective of the direction of change, which has profound implications for learning and neurological restoration (83).
The question arises whether EPO could restore this fundamental capacity in a compromised brain. Knowing that FF lesions severely impair synaptic plasticity (87), we demonstrated that a single systemic dose of EPO is sufficient to fully restore long-lasting LTP in the dentate gyrus of lesioned animals (88, 99). As LTP and LTD are established cellular mechanisms of memory (82, 90-92), this restoration of synaptic malleability provides a plausible substrate for the observed recovery of spatial memory, enabling the formation and stabilization of memory traces (engrams). The finding that EPO facilitates both LTP and LTD suggests it fine-tunes the homeostatic balance of neural networks, enhancing their capacity for information encoding and storage.
8. EPO Induces Molecular Mediators of Plasticity in Memory Circuits
The above-mentioned effects imply that EPO must engage the molecular machinery of plasticity. Our data confirm that a single EPO dose rapidly upregulates the expression of two key plasticity-related genes, bdnf and arc, in the prefrontal cortex of intact animals (77). BDNF (Brain-Derived Neurotrophic Factor) is a key protein that promotes neuronal survival, differentiation, and synaptic strengthening (77), while Arc (Activity-Regulated Cytoskeleton-associated protein) is crucial for the cytoskeletal remodeling that underlies long-term synaptic changes and memory consolidation (77). In subsequent studies, both the daily post-training EPO regimen in the Morris water maze and the single-dose protocol in object recognition training increased the expression of these genes in the hippocampus and prefrontal cortex following memory retrieval.
The hippocampus and prefrontal cortex form a critical circuit for explicit memory processing (93, 94), with well-defined roles in spatial and object recognition memory (93, 94). Given the established functions of BDNF and Arc in memory consolidation (93, 94), it is reasonable to posit that their induction by EPO manages the neuroplastic mechanisms supporting memory recovery and prolongation. Independent work has corroborated that EPO increases bdnf and arc expression (93, 94). The critical time-dependence of our behavioral effects is mirrored molecularly: animals that received EPO 5 h post-training, which showed no memory recovery, similarly failed to show this specific molecular signature. This supports the hypothesis that EPO modulates neural plasticity within a privileged time window after a salient event, a concept aligned with the ‘synaptic tagging and capture’ hypothesis, where EPO could act as a plasticity-related protein that selectively stabilizes synapses “tagged” by prior learning activity (78, 95-99).
Notably, the administration of BDNF itself can induce slow, spontaneous potentiation similar to what we observed after EPO (84-86, 100-102), suggesting that BDNF induction might be a key mediator of EPO’s effects. Furthermore, we found that the expression patterns of bdnf and arc shift between 24 h and 21 days post-training, indicating that EPO may be accelerating and strengthening the natural process of systems consolidation, with immediate support in the prefrontal cortex leading to more robust long-term storage in the hippocampus.
9. Endogenous EPO Signaling is an Activity-Dependent Component of Memory Formation
To determine if endogenous EPO is a physiological participant in memory processes, we examined its expression following learning. We found that just 3 min of exploration in a spatial object recognition task, which produces short-term memory, significantly increased expression of the EPO gene and its receptor EPO-r, in the hippocampus, with EPO-r expression also rising in the prefrontal cortex. Extending acquisition to 5 min, which extends memory duration to 24 h, produced a much more robust increase in both gene and receptor expression in both brain regions. This suggests that EPO/EPO-r expression is activity-dependent and tuned to the strength of the memory being formed, a finding consistent with reports that motor learning increases EPO signaling (103) and that higher endogenous EPO levels in humans correlate with superior cognitive performance (103). This positions the endogenous EPO system as a natural, activity-dependent modulator of plasticity, which our synchronized protocol seeks to amplify with exogenous, timed administration.
10. The Promise of EPO in Neuropsychiatry
The exploration of EPO’s role in psychiatric disorders, while less advanced, is gaining momentum. A decade-long retrospective search for EPO and schizophrenia yields little, but pioneering work primarily from the group of Kamila Miskowiak providing evidence for its potential use in mood disorders. Their studies indicate that EPO can ameliorate cognitive deficits in depression and counteract cognitive side-effects of electroconvulsive therapy (46, 63, 104-107). Recent pre-clinical evidence further confirms that carbamoylated EPO (a non-erythropoietic variant) is effective in animal models of stress-induced depression and anxiety (108, 109). These findings, though preliminary, open a promising avenue for EPO in treating the cognitive deficits often associated with mood disorders, which are frequently resistant to conventional antidepressants. The synchronized protocol suggests a strategic extension: combining cognitive remediation therapy (as the circuit-specific stimulus) with timed administration of non-erythropoietic EPO variants could potentially enhance the efficacy of treating these stubborn cognitive deficits.
11. Final Considerations and Conclusions
The collective evidence points toward a transformative potential for EPO in treating acquired and degenerative neurological conditions. The experimental foundation provides a robust mechanistic rationale for its use in neurorestoration. We propose that this synchronized, two-signal strategy—coordinating behavioral activation with timed EPO administration—could evolve into a powerful therapeutic tool, representing a conceptual shift in neurological restoration: from a passive, pharmacological shield against damage to an active process of guiding the brain’s innate repair mechanisms to rebuild function. This approach is grounded in a testable hypothesis for future formalization: that the efficacy of EPO in promoting recovery is not merely dose-dependent but is fundamentally “context-dependent,” requiring the prior establishment of a transient, activity-tagged neural circuit. We hypothesize that EPO’s primary restorative role is to broadly supply plasticity-related proteins that are then selectively captured and utilized to stabilize those synaptic pathways that have been functionally “tagged” by a specific experience, thereby achieving a level of precision in circuit rewiring previously unattainable with standalone pharmacological interventions.
However, the clinical translation of conventional EPO remains constrained, primarily due to safety concerns associated with its erythropoietic effects in non-anemic patients.The solution to this impasse may lie in biotechnological innovation. The production of recombinant EPO generates molecular variants, including asialoEPO, which lacks terminal sialic acid residues. This truncated form retains the neuroprotective and plasticity-promoting properties but, due to its rapid clearance from the bloodstream, minimizes the systemic erythropoietic effects that raise safety concerns in non-anemic patients. This molecule, combined with intranasal delivery to bypass the bloodstream and target the CNS directly, forms the basis of a new therapeutic strategy. The Center for Molecular Immunology in Cuba is currently conducting clinical trials with this intranasal formulation, termed NeuroEPO, in patients with Alzheimer’s disease. It is such targeted clinical efforts, leveraging engineered molecules and precise delivery that will ultimately define the place of EPO in the neurologist’s and psychiatrist’s arsenal.
In conclusion, EPO has transcended its classical role in erythropoiesis to emerge as a pleiotropic cytokine with profound effects on the nervous system. The pre-clinical evidence, including that presented here, demonstrates that beyond its well-established neuroprotective role, EPO is a potent promoter of neuroplasticity. It facilitates memory consolidation, expands the capacity of synapses for both potentiation and depression, and activates pro-plasticity genetic programs in hippocampus-cortical circuits. These mechanisms underline their ability to restore function after established injury, positioning it as a unique agent in the field of restorative neurology. Biotechnological developments like nasally administered NeuroEPO, designed to minimize systemic effects, are paving the way for the much-needed definitive clinical trials to validate this therapeutic potential in neurodegenerative diseases, brain injury sequelae, and neuropsychiatric disorders. By shifting the paradigm from broad-spectrum neuroprotection to timely-precise, circuit-specific rewiring, EPO and its engineered derivatives stand poised to redefine therapeutic strategies in restorative neurology and psychiatry.
Author Contributions
William Almaguer Melían: Participated in the conceptualization, writing, and review of the text and figures of this article. Daymara Mercerón: Participated in the conceptualization, writing, and review of the text and figures of this article. Briceida Bergado Acosta: Participated in the conceptualization, writing, and revision of the text and figures of this article. Jorge A. Bergado: Participated in the conceptualization, writing, and revision of the text and figures of this article. Corresponding author. All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
No new data were created or analyzed in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ramón y Cajal S. Degeneration and Regeneration of the Nervous System. New York: Harper Press; 1969.
- Bergado-Rosado JA, Almaguer-Melian W. [Cellular mechanisms of neuroplasticity]. Rev. Neurol. 2000; 31: 1074-95. PMID: 11190877.
- Rosenzweig MR, Bennett EL. Psychobiology of plasticity: Effects of training and experience on brain and behavior. Behav. Brain Res. 1996;78: 57-65. [CrossRef]
- Flexner JB, Flexner LB, Stellar E, De La HABA, Roberts RB. Inhibition of protein synthesis in brain and learning and memory following puromycin. J. Neurochem. 1962; 9:595-605. [CrossRef]
- Flexner LB, Flexner JB, Stellar E. Memory and cerebral protein synthesis in mice as affected by graded amounts of puromycin. Exp. Neurol. 1965; 13: 264-72. [CrossRef]
- Nottebohm F. Neuronal replacement in adulthood. Ann. N.Y. Acad. Sci. 1985; 457. [CrossRef]
- Nottebohm F. From bird song to neurogenesis. Sci. Am. 1989;260(2):74-9. [CrossRef]
- Bliss TV, Lomo T. Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J. Physiol.(Lond). 1973; 232:331-56. [CrossRef]
- Bliss TV, Gardner-Medwin AR. Long-lasting potentiation of synaptic transmission in the dentate area of the unanaestetized rabbit following stimulation of the perforant path. J. Physiol. (Lond). 1973; 232:357-74. [CrossRef]
- Levy WB, Steward O. Synapses as associative memory elements in the hippocampal formation. Brain Res. 1979; 175:233-45. [CrossRef]
- Teyler TJ, DiScenna P. Long-term potentiation as a candidate mnemonic device. Brain Res. 1984; 319:15-28. [CrossRef]
- Krug M, Loessner B, Ott T. Anisomycin blocks the late phase of long-term potentiation in the dentate gyrus of freely moving rats. Brain Res.Bull. 1984; 13:39-42. [CrossRef]
- Loessner B, Schweigert C, Pchalek V, Krug M, Frey S, Matthies H. Training- and LTP-induced changes of protein synthesis in rat hippocampus. Neurosci. Suppl. 1987; 22:9512.
- Frey U, Krug M, Reymann KG, Matthies H. Anisomycin, an inhibitor of protein synthesis, blocks late phases of LTP phenomena in the hippocampal CA1 region in vitro. Brain Res. 1988; 452:57-65. [CrossRef]
- Krug M, Ruethrich H, Bergado J. The nootropic substance methylglucamine orotate prolongs both, postconditioning potentiation and posttetanic LTP in the dentate gyrus of freely moving rats. Act.Nerv.Sup.(Prague). 1988;30(3):232-3.
- Dąbrowski J, Czajka A, Zielińska-Turek J, Jaroszyński J, Furtak-Niczyporuk M, Mela A, et al. Brain Functional Reserve in the Context of Neuroplasticity after Stroke. Neural Plast. 2019; 2019:9708905. [CrossRef]
- Wadan AS, Raza ML, Moradikor N. Synaptic modulation by coffee compounds: Insights into neural plasticity. Prog Brain Res. 2024; 289:181-91. [CrossRef]
- Colucci-D’Amato L, Speranza L, Volpicelli F. Neurotrophic Factor BDNF, Physiological Functions and Therapeutic Potential in Depression, Neurodegeneration and Brain Cancer. Int. J. Mol. Sci. 2020;21(20). [CrossRef]
- Haase VH. Regulation of erythropoiesis by hypoxia-inducible factors. Blood Rev. 2013; 27:41-53. [CrossRef]
- Noguchi CT, Asavaritikrai P, Teng R, Jia Y. Role of erythropoietin in the brain. Crit. Rev. Oncol. Hematol. 2007; 64:159-71. [CrossRef]
- Frey U, Morris RGM. Synaptic tagging: implications for late maintenance of hippocampal long-term potentiation. Trends Neurosci. 1998; 21:181-8. [CrossRef]
- Frey U, Morris RGM. Synaptic tagging and long-term potentiation. Nature. 1997;385(6616):533-6. [CrossRef]
- Seidenbecher T, Reymann KG, Balschun D. A post-tetanic time window for the reinforcement of long-term potentiation by appetitive and aversive stimuli. Proc. Natl. Acad. Sci. USA. 1997; 94: 1494-9. [CrossRef]
- Masuda S, Nagao M, Takahata K, Konishi Y, Gallyas F, Tabira T, et al. Functional Erythropoietin Receptor of the Cells with Neural Characteristics. Comparison with receptor properties of erythroid cells. J. Biol. Chem. 1993;268(15):11208-16.
- Morishita E, Masuda S, Nagao M, Yasuda Y, Sasaki R. Erythropoietin receptor is expressed in rat hippocampal and cerebral cortical neurons, and erythropoietin prevents in vitro glutamate-induced neuronal death. Neuroscience. 1997;76: 105-16. [CrossRef]
- Sargin D, El-Kordi A, Agarwal A, Muller M, Wojcik SM, Hassouna I, et al. Expression of constitutively active erythropoietin receptor in pyramidal neurons of cortex and hippocampus boosts higher cognitive functions in mice. BMCBiol. 2011; 9: 27. [CrossRef]
- Miyake T, Kung CK, Goldwasser E. Purification of human erythropoietin. J. Biol. Chem. 1977; 252: 5558-64.
- Suresh S, Rajvanshi PK, Noguchi CT. The Many Facets of Erythropoietin Physiologic and Metabolic Response. Sci. Translat. Med. 2019; 10:1534. [CrossRef]
- Tan CC, Eckardt K, Firth JD, Ratcliffe PJ. Feedback modulation of renal and hepatic erythropoietin mRNA in response to graded anemia and hypoxia. Am. J. Physiol. 1992;263: F474-81. [CrossRef]
- Konishi Y, Chui DH, Hirose H, Kunishita H, Tabira T. Trophic effect of erythropoietin and other hematopoietic factors on central cholinergic neurons in vivo and in vivo. Brain Res. 1993; 609:29-35. [CrossRef]
- Shingo T, Sorokan ST, Shimazaki T, Weiss S. Erythropoietin regulates the in vitro and in vivo production of neuronal progenitors by mammalian forebrain neural stem cells. J. Neurosci. 2001; 21: 9733-43. [CrossRef]
- Sirén AL, Knerlich F, Poser W, Gleiter CH, Brück W, Ehrenreich H. Erythropoietin and erythropoietin receptor in human ischemic/hypoxic brain. Acta Neuropathol. 2001; 101: 271-6. [CrossRef]
- Kumral A, Uysal N, Tugyan K, Sonmez A, Yilmaz O, Gokmen N, et al. Erythropoietin improves long-term spatial memory deficits and brain injury following neonatal hypoxia-ischemia in rats. Behav. Brain Res. 2004;153(1):77-86. [CrossRef]
- Brines M, Cerami A. Emerging biological roles for erythropoietin in the nervous system. Nature. 2005; 6:484-94. [CrossRef]
- Chen J, Chopp M. Neurorestorative treatment of stroke: cell and pharmacological approaches. NeuroRx. 2006;3: 466-73. [CrossRef]
- Adamcio B, Sargin D, Stradomska A, Medrihan L, Gertler C, Theis F, et al. Erythropoietin enhances hippocampal long-term potentiation and memory. BMC Biol. 2008; 6:37. [CrossRef]
- Bartels C, Spate K, Krampe H, Ehrenreich H. Recombinant Human Erythropoietin: Novel Strategies for Neuroprotective/Neuro-regenerative Treatment of Multiple Sclerosis. Ther. Adv. Neurol. Disord. 2008; 1:193-206. [CrossRef]
- Ehrenreich H, Bartels C, Sargin D, Stawicki S, Krampe H. Recombinant human erythropoietin in the treatment of human brain disease: focus on cognition. J. Ren. Nutr. 2008; 18:146-53. [CrossRef]
- Jing M, Shingo T, Yasuhara T, Kondo A, Morimoto T, Wang F, et al. The combined therapy of intrahippocampal transplantation of adult neural stem cells and intraventricular erythropoietin-infusion ameliorates spontaneous recurrent seizures by suppression of abnormal mossy fiber sprouting. Brain Res. 2009; 1295:203-17. [CrossRef]
- Kamal A, Al ST, Ramakers G. Erythropoietin decreases the excitatory neurotransmitter release probability and enhances synaptic plasticity in mice hippocampal slices. Brain Res. 2011; 1410:33-7. [CrossRef]
- Sedriep S, Xia X, Marotta F, Zhou L, Yadav H, Yang H, et al. Beneficial nutraceutical modulation of cerebral erythropoietin expression and oxidative stress: an experimental study. J. Biol. Regul. Homeost. Agents. 2011;25(2):187-94. PMID: 21880207.
- Arabpoor Z, Hamidi G, Rashidi B, Shabrang M, Alaei H, Sharifi MR, et al. Erythropoietin improves neuronal proliferation in dentate gyrus of hippocampal formation in an animal model of Alzheimer’s disease. Adv. Biomed. Res. 2012; 1:50. [CrossRef]
- Mengozzi M, Cervellini I, Villa P, Erbayraktar Z, Gokmen N, Yilmaz O, et al. Erythropoietin-induced changes in brain gene expression reveal induction of synaptic plasticity genes in experimental stroke. Proc. Natl. Acad. Sci. USA. 2012; 109:9617-22. [CrossRef]
- Wobrock T, Hasan A, Falkai P. Innovative treatment approaches in schizophrenia enhancing neuroplasticity: aerobic exercise, erythropoetin and repetitive transcranial magnetic stimulation. Curr. Pharm. Biotechnol. 2012; 13:1595-605. [CrossRef]
- Esmaeili TP, Moosavi SM, Shabani M, Haghani M. Erythropoietin improves synaptic plasticity and memory deficits by decrease of the neurotransmitter release probability in the rat model of Alzheimer’s disease. Pharmacol. Biochem. Behav. 2015; 130:15-21. [CrossRef]
- Miskowiak KW, Petersen JZ, Macoveanu J, Ysbæk-Nielsen AT, Lindegaard IA, Cramer K, et al. Effect of erythropoietin on cognitive side-effects of electroconvulsive therapy in depression: A randomized, double-blind, placebo-controlled trial. Europ. Neuropsychopharmacol. 2024; 79:38-48. [CrossRef]
- Sakanaka M, Wen TC, Matsuda S, Masuda S, Morishita E, Nagao M, et al. In vivo evidence that erythropoietin protects neurons from ischemic damage. Proc. Natl. Acad. Sci. USA. 1998; 95:4635-40. [CrossRef]
- Li SJ, Cui KF, Fu JJ, Fu XJ, Gao YF, Zhang D, et al. EPO promotes axonal sprouting via upregulating GDF10. Genet. Mol. Biol. 2019; 711:134412. [CrossRef]
- Ben Attia M, N’Gouemo P, Belaidi M, Rondouin G, Chicheportiche R. Kindling and electrode effects on the benzodiazepine receptors density of olfactory bulb and hippocampus after olfactory bulb kindling. Neurosci. Lett. 1992; 143:74-8. [CrossRef]
- Marti HH, Gassmann M, Wenger RH, Kvietikova I, Morganti-Kossmann MC, Kossmann T, et al. Detection of erythropoietin in human liquor: intrinsic erythropoietin production in the brain. Kidney internat. 1997;51:416-8. [CrossRef]
- Peng W, Xing Z, Yang J, Wang Y, Wang W, Huang W. The efficacy of erythropoietin in treating experimental traumatic brain injury: a systematic review of controlled trials in animal models. J. Neurosurg. 2014; 121: 653-64. [CrossRef]
- Blixt J, Song Y, Wanecek M, Gunnarson E. EPO has multiple positive effects on astrocytes in an experimental model of ischemia. Brain Res. 2023; 1802:148207. [CrossRef]
- Xiong Y, Zhang Y, Mahmood A, Meng Y, Qu C, Chopp M. Erythropoietin mediates neurobehavioral recovery and neurovascular remodeling following traumatic brain injury in rats by increasing expression of vascular endothelial growth factor. Translat. Stroke Res. 2011; 2:619-32. [CrossRef]
- Ureña-Guerrero ME, Castañeda-Cabral JL, Rivera-Cervantes MC, Macias-Velez RJ, Jarero-Basulto JJ, Gudiño-Cabrera G, et al. Neuroprotective and Neurorestorative Effects of Epo and VEGF: Perspectives for New Therapeutic Approaches to Neurological Diseases. Curr. Pharmaceut. Design. 2020;2 6:1263-76. [CrossRef]
- Demers EJ, McPherson RJ, Juul SE. Erythropoietin protects dopaminergic neurons and improves neurobehavioral outcomes in juvenile rats after neonatal hypoxia-ischemia. Pediat. Res. 2005; 58:297-301. [CrossRef]
- Xu Y, Huang L, Han J, Zhou Y. Effects of EPO combined with mild hypothermia on oxidative stress and neuroprotection in neonates with hypoxic-ischemic encephalopathy. Cell. Mol. Biol. (Noisy-le-Grand). 2022; 68:36-45. [CrossRef]
- Lai YF, Lin TY. Erythropoietin in Optic Neuropathies: Current Future Strategies for Optic Nerve Protection and Repair. 2022; 23(13). [CrossRef]
- Boesch S, Indelicato E. Erythropoietin and Friedreich Ataxia: Time for a Reappraisal? Front. Neurosci. 2019; 13:386. [CrossRef]
- Bartels C, Späte K, Krampe H, Ehrenreich H. Recombinant Human Erythropoietin: Novel Strategies for Neuroprotective/Neuro-regenerative Treatment of Multiple Sclerosis. Ther. Adv. Neurol. Dis. 2008; 1:193-206. [CrossRef]
- Dayyat EA, Zhang SX, Wang Y, Cheng ZJ, Gozal D. Exogenous erythropoietin administration attenuates intermittent hypoxia-induced cognitive deficits in a murine model of sleep apnea. BMC Neurosci. 2012; 13:77. [CrossRef]
- Maurice T, Mustafa MH, Desrumaux C, Keller E, Naert G, de la CG-BM, et al. Intranasal formulation of erythropoietin (EPO) showed potent protective activity against amyloid toxicity in the Aβ25₋35 non-transgenic mouse model of Alzheimer’s disease. J. Psychopharmacol. 2013; 27:1044-57. [CrossRef]
- Auzmendi J, Puchulu MB, Rodríguez JCG, Balaszczuk AM, Lazarowski A, Merelli A. EPO and EPO-Receptor System as Potential Actionable Mechanism for the Protection of Brain and Heart in Refractory Epilepsy and SUDEP. Neurochem. Res. 2020;26(12):1356-64. [CrossRef]
- Miskowiak KW, Macoveanu J, Vinberg M, Assentoft E, Randers L, Harmer CJ, et al. Effects of erythropoietin on memory-relevant neurocircuitry activity and recall in mood disorders. Acta Psychiat. Scand. 2016; 134:249-59. [CrossRef]
- Vinberg M, Weikop P, Olsen NV, Kessing LV, Miskowiak K. Effect of recombinant erythropoietin on inflammatory markers in patients with affective disorders: A randomised controlled study. Brain Behav. Immun. 2016; 57:53-7. [CrossRef]
- Qi C, Xu M, Gan J, Yang X, Wu N, Song L, et al. Erythropoietin improves neurobehavior by reducing dopaminergic neuron loss in a 6-hydroxydopamine-induced rat model. Internat. J. Mol. Med. 2014; 34: 440-50. [CrossRef]
- Rey F, Ottolenghi S. Mitochondrial Metabolism as Target of the Neuroprotective Role of Erythropoietin in Parkinson’s Disease. Antioxidants (Basel) 2021;10(1). [CrossRef]
- Pedroso I, Bringas ML, Aguiar A, Morales L, Alvarez M, Valdés PA, et al. Use of Cuban recombinant human erythropoietin in Parkinson’s disease treatment. MEDICC Review. 2012; 14:11-7. [CrossRef]
- Fischer HS, Reibel NJ, Bührer C, Dame C. Prophylactic Erythropoietin for Neuroprotection in Very Preterm Infants: A Meta-Analysis Update. Front. Pediat. 2021; 9:657228. [CrossRef]
- Ma Y, Zhou Z, Yang GY, Ding J, Wang X. The Effect of Erythropoietin and Its Derivatives on Ischemic Stroke Therapy: A Comprehensive Review. Front. Pharmacol. 2022; 13:743926. [CrossRef]
- Robertson C, Sadrameli S. Erythropoietin in the neurology ICU. Curr. Treat. Options Neurol. 2013; 15:104-12. [CrossRef]
- Hemani S, Lane O. Systematic Review of Erythropoietin (EPO) for Neuroprotection in Human Studies. Neurochem. Res. 2021; 46:732-9. [CrossRef]
- Cassel J-C, Duconseille E, Jeltsch H, Will B. The fimbria-fornix/cingular bundle pathways: a review of neurochemical and behavioural approaches using lesions and transplantation techniques. Prog. Neurobiol. 1997; 51:663-716. [CrossRef]
- Vizi ES, Kiss JP. Neurochemistry and pharmacology of the major hippocampal transmitter systems: synaptic and non-synaptic interaction. Hippocampus. 1998; 8:566-607. [CrossRef]
- Liu J, Zhang Z, Li JT, Zhu YH, Zhou HL, Liu S, et al. Effects of NT-4 gene modified fibroblasts transplanted into AD rats. Neurosci. Lett. 2009; 466:1-5. [CrossRef]
- Xuan AG, Luo M, Ji WD, Long DH. Effects of engrafted neural stem cells in Alzheimer’s disease rats. Neurosci. Lett. 2009; 450:167-71. [CrossRef]
- Xuan AG, Long DH, Gu HG, Yang DD, Hong LP, Leng SL. BDNF improves the effects of neural stem cells on the rat model of Alzheimer’s disease with unilateral lesion of fimbria-fornix. Neurosci. Lett. 2008; 440:331-5. [CrossRef]
- Almaguer-Melian W, Merceron-Martinez D, Pav¢n-Fuentes N, Alberti Amador E, Leon-Martinez R, Ledon N, et al. Erythropoietin promotes neural plasticity and spatial memory recovery in fimbria-fornix-lesioned ats. Neurorehabil. Neural Repair. 2015; 29: 979-88. [CrossRef]
- Almaguer-Melian W, Mercerón-Martinez D, Delgado-Ocaña S, Alberti-Amador E, Gonzalez-Gómez R, Bergado JA. Erythropoietin improves object placement recognition memory in a time dependent manner in both, uninjured animals and fimbria-fornix-lesioned male rats. Horm. Behav. 2018; 100:6. [CrossRef]
- Almaguer-Melian W, Mercerón-Martinez D, Alberti-Amador E, Alacán-Ricardo L, de Bardet JC, Orama-Rojo N, et al. Learning induces EPO/EPOr expression in memory relevant brain areas, whereas exogenously applied EPO promotes remote memory consolidation. Synapse. 2024;78:e22282. [CrossRef]
- Tse D, Langston RF, Kakeyana M, Bethus I, Spooner PA, Wood ER, et al. Schemas and Memory Consolidation. Science. 2007; 316:76-82. [CrossRef]
- Dudai Y. Molecular basis of long-term memories: a question of persistence. Curr. Op. Neurobiol. 2002; 12:211-6. [CrossRef]
- Malenka RC, Bear MF. LTP and LTD; An Embarrassment of Riches. Neuron. 2004; 44:5-21. [CrossRef]
- Almaguer-Melian W, Mercerón-Martínez D, Delgado-Ocaña S, Pavón-Fuentes N, Ledón N, Bergado JA. EPO Induces Changes in Synaptic Transmission and Plasticity in the Dentate Gyrus of Rats. Synapse. 2016; 70: 240-52. [CrossRef]
- Bramham CR, Messaoudi E. BDNF function in adult synaptic plasticity: The synaptic consolidation hypothesis. Prog. Neurobiol. 2005; 76: 99-125. [CrossRef]
- Soule J, Messaoudi E, Bramham CR. Brain-derived neurotrophic factor and control of synaptic consolidation in the adult brain. Biochem. Soc. Trans. 2006; 34 (Pt 4): 600-4. [CrossRef]
- Ying SW, Futter M, Rosenblum K, Webber MJ, Hunt SP, Bliss TV, et al. Brain-derived neurotrophic factor induces long-term potentiation in intact adult hippocampus: requirement for ERK activation coupled to CREB and upregulation of Arc synthesis. J. Neurosci. 2002; 22:1532-40. [CrossRef]
- Buzsaki G, Gage FH. Absence of long-term potentiation in the subcortically deafferented dentate gyrus. Brain Res. 1989; 484:94-101. [CrossRef]
- Almaguer-Melian W, Mercerón-Martínez D, Bergado-Rosado J. A unique erythropoietin dosage induces the recovery of long-term synaptic potentiation in fimbria-fornix lesioned rats. Brain Res. 2023; 1799:148178. [CrossRef]
- Bear MF, Malenka RC. Synaptic plasticity: LTP and LTD. Curr. Op. Neurobiol. 1994; 4:389-99. [CrossRef]
- Malenka RC. Synaptic Plasticity in the Hippocampus: LTP and LTD. Cell. 1994; 78:535-8. [CrossRef]
- Bliss TVP, Collingridge GL. A synaptic model of memory: Long-term potentiation in the hippocampus. Nature. 1993; 361:31-9. [CrossRef]
- Matthies H. Neurobiological aspects of learning and memory. Ann. Rev. Psychol. 1989; 40:381-404. [CrossRef]
- McCormick DA, Nestvogel DB, He BJ. Neuromodulation of Brain State and Behavior. Ann. Rev. Neurosci. 2020; 43:391-415. [CrossRef]
- Rolls ET. The hippocampus, ventromedial prefrontal cortex, and episodic and semantic memory. Prog. Neurobiol. 2022; 217:102334. [CrossRef]
- Frey U, Morris RGM. Synaptic tagging and long-term potentiation. Nature. 1997; 385:533-6. [CrossRef]
- Frey U, Morris RGM. Synaptic tagging: implications for late maintenance of hippocampal long-term potentiation. Trends Neurosci. 1997; 21:181-8. [CrossRef]
- McGaugh JL. Memory-a century of consolidation. Science. 2000; 287:248-51. [CrossRef]
- Ballarini F, Moncada D, Mart¡nez MC, Alen N, Viola H. Behavioral tagging is a general mechanism of long-term memory formation. Proc. Nat. Acad. Sci. USA. 2009; 106:14599-604. [CrossRef]
- Moncada D, Ballarini F, Viola H. Behavioral Tagging: A Translation of the Synaptic Tagging and Capture Hypothesis. Neural Plast. 2015; 2015: 650780. [CrossRef]
- Bramham CR, Worley PF, Moore MJ, Guzowski JF. The immediate early gene arc/arg3.1: regulation, mechanisms, and function. J. Neurosci. 2008;28: 11760-7. [CrossRef]
- Bramham CR. Opioid receptor dependent long-term potentiation: Peptidergic regulation of synaptic plasticity in the hippocampus. Neurochemistry International. 1992; 20: 441-55. [CrossRef]
- Tyler WJ, Alonso M, Bramham CR, Pozzo-Miller LD. From acquisition to consolidation: on the role of brain-derived neurotrophic factor signaling in hippocampal-dependent learning. Learn. Mem. 2002; 9: 224-37. [CrossRef]
- Wakhloo D, Scharkowski F. Functional hypoxia drives neuroplasticity and neurogenesis via brain erythropoietin. Nat. Commun. 2020;11: 1313. [CrossRef]
- Miskowiak KW, Vinberg M, Macoveanu J, Ehrenreich H, Køster N, Inkster B, et al. Effects of Erythropoietin on Hippocampal Volume and Memory in Mood Disorders. Biol Psychiatry. 2015;78: 270-7. [CrossRef]
- Miskowiak KW, Vinberg M, Glerup L, Paulson OB, Knudsen GM, Ehrenreich H, et al. Neural correlates of improved executive function following erythropoietin treatment in mood disorders. Psychol Med. 2016;46: 1679-91. [CrossRef]
- Miskowiak KW, Rush AJ, Jr., Gerds TA, Vinberg M, Kessing LV. Targeting Treatments to Improve Cognitive Function in Mood Disorder: Suggestions From Trials Using Erythropoietin. The Journal of clinical psychiatry. 2016;77: e1639-46. [CrossRef]
- Miskowiak KW, Petersen NA, Harmer CJ, Ehrenreich E, Kessing LV. Neural correlates of improved recognition of happy faces after erythropoietin treatment in bipolar disorder. 2018;138: 336-47. [CrossRef]
- Yaeger JDW, John MM, Ledesma LJ, Krupp KT, Booth CD, Jones NT, et al. Acute carbamoylated erythropoietin reduces social stress-induced anxiety and depression-related behaviors. Neuropharmacology. 2025:110558. [CrossRef]
- Martin L, Garcia Rodriguez JC, Audran M, Ericsson M, Maurice T. Detection of a nonerythropoietic erythropoietin, Neuro-EPO, in blood after intranasal administration in rat. Drug Test Anal. 2020;12: 1605-13. [CrossRef]
Figure 1.
Mechanism of EPO expression-induction in erythropoiesis. On the left, under normal oxygenation conditions (NOR) Hif-1α is destroyed by the ubiquitin mechanism. On the right, under hypoxic (HIP) conditions dimerization of Hif-1α and Hif-1β occurs. Hif: Hipoxia inducible factor (1α y 1β). VHL: von Hippel-Lindau antitumoral factor. EPO: erythropoietin. VEGF: vascular growth factor. TCA: Tricarboxylic Acids cycle. See the text for more details.
Figure 1.
Mechanism of EPO expression-induction in erythropoiesis. On the left, under normal oxygenation conditions (NOR) Hif-1α is destroyed by the ubiquitin mechanism. On the right, under hypoxic (HIP) conditions dimerization of Hif-1α and Hif-1β occurs. Hif: Hipoxia inducible factor (1α y 1β). VHL: von Hippel-Lindau antitumoral factor. EPO: erythropoietin. VEGF: vascular growth factor. TCA: Tricarboxylic Acids cycle. See the text for more details.
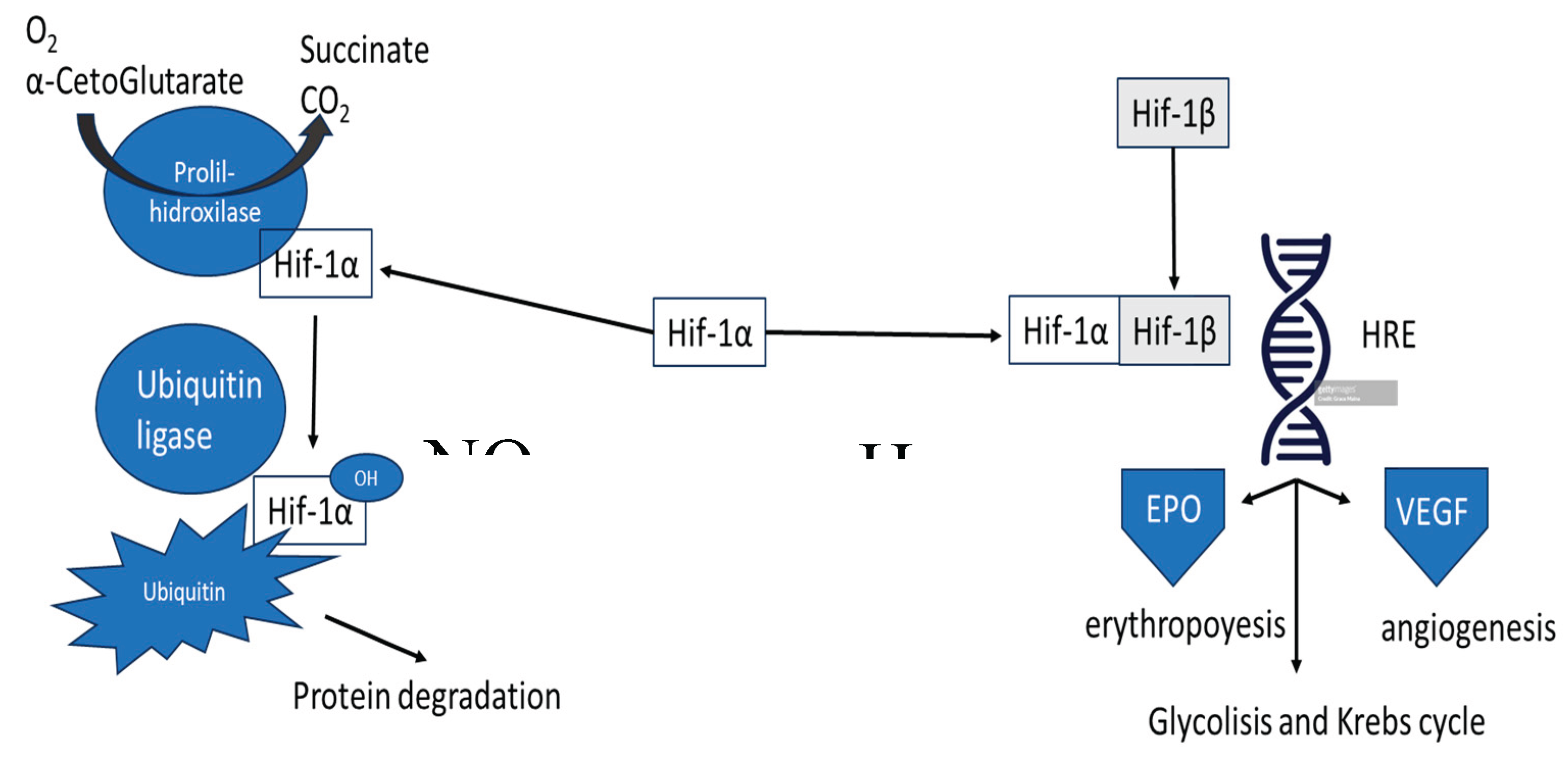
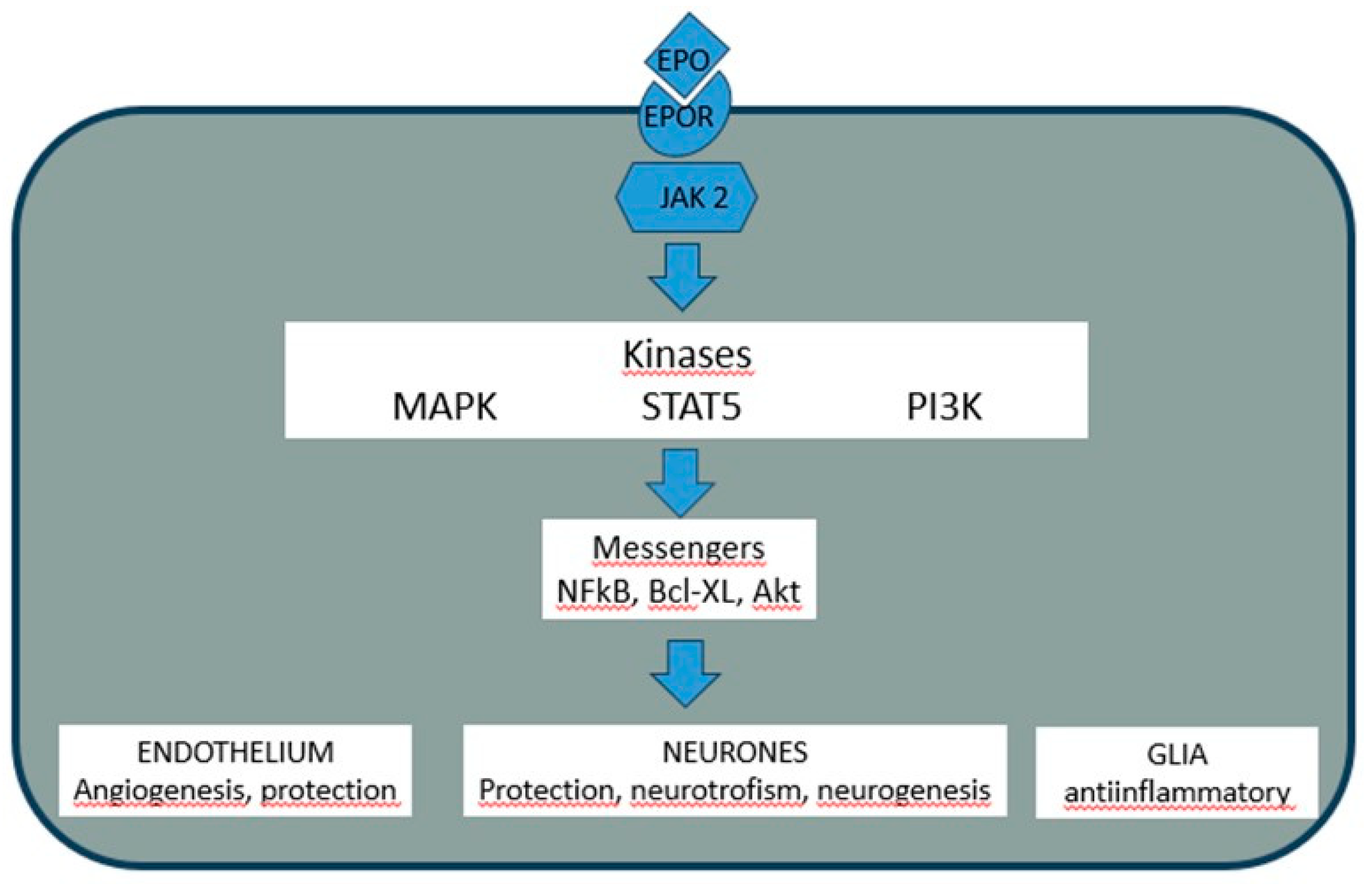
Figure 2.
Mechanism of EPO’s effects on sensitive cells bearing the EPO receptor. The molecular cascades activated by EPO-receptor (EPOR) binding have promoting effects on endothelial cells, neurons, and glial cells. EPO: erythropoietin; EPOR: EPO Receptor; JAK2: Janus kinase 2; MAPK: mitogen-activated kinase; STAT5: Transcription activator; PI3K: Phosphatidil inositol 3 kinase; NFkB: kappaB nuclear factor; Bcl-xL: B cells lymphoma factor; Akt: proteinkinase B.
Figure 2.
Mechanism of EPO’s effects on sensitive cells bearing the EPO receptor. The molecular cascades activated by EPO-receptor (EPOR) binding have promoting effects on endothelial cells, neurons, and glial cells. EPO: erythropoietin; EPOR: EPO Receptor; JAK2: Janus kinase 2; MAPK: mitogen-activated kinase; STAT5: Transcription activator; PI3K: Phosphatidil inositol 3 kinase; NFkB: kappaB nuclear factor; Bcl-xL: B cells lymphoma factor; Akt: proteinkinase B.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



