
Submitted:
18 December 2025
Posted:
18 December 2025
You are already at the latest version
Abstract
Soil salinity is recognized as a critical abiotic stress that limits plant growth on marginal lands. The cup plant (Silphium perfoliatum L.), a perennial bioenergy species with high biomass potential, has been proposed for cultivation on saline-degraded soils; however, its responses to salt stress remain insufficiently characterized. In the present study, the physiological responses of cup plant to neutral (NaCl) and alkaline (NaHCO₃) salt stress at concentrations of 100, 200, and 300 mM were evaluated in a pot experiment conducted under controlled conditions. The assessed indicators included relative chlorophyll content (CCI), chlorophyll a fluorescence parameters (Fv/Fm, Fv/Fo, PI), and gas ex-change characteristics, namely net photosynthetic rate (Pn), stomatal conductance (gs), transpiration rate (E), and intercellular CO₂ concentration (Ci). Dose-dependent de-creases in all measured physiological parameters were induced by salinity. A more pronounced negative effect was observed under alkaline salt stress than under neutral salt stress. For instance, at 300 mM NaHCO₃, chlorophyll content and photosynthetic rate were substantially reduced compared with those recorded under the corresponding NaCl treatment. Even at the moderate salinity level of 100 mM, reductions in photo-synthetic performance were detected relative to the control. Overall, photosynthetic efficiency and gas exchange in cup plant were markedly impaired by salinity, particu-larly under conditions of high bicarbonate concentration. These findings provide new insight into the physiological limitations of S. perfoliatum under acute salt stress and demonstrate that alkaline salinity, associated with elevated pH due to HCO₃⁻, exacer-bates stress effects beyond the osmotic and ionic impacts of neutral salinity.
Keywords:
Silphium perfoliatum L.
; soil salinity
; relative chlorophyll content
; chlorophyll fluorescence
; gas exchange
1. Introduction
In response to the growing challenges of climate change, soil degradation, and the need to reduce greenhouse gas emissions, the transformation of global energy systems has become a strategic priority for many countries and international organizations [1,2]. Achieving climate neutrality targets requires diversification of energy sources and a shift away from fossil fuels toward low-emission renewable technologies. In this context, biomass plays a particularly important role, as it can be locally sourced, integrated with agricultural systems, and relatively easily converted into heat, electricity, and biofuels [3,4,5]. Among biomass feedstocks, energy crops are gaining increasing interest because their cultivation allows for stable and predictable raw material production in areas with limited agricultural suitability [6,7].
A particularly promising species in this group is cup plant (Silphium perfoliatum L.), a perennial herb with high biomass productivity, well adapted to variable environmental conditions, and relatively low agronomic requirements. Previous studies have demonstrated its high yield potential on marginal soils [8,9]. S. perfoliatum exhibits considerable habitat flexibility and a strong ability to adapt to less fertile or marginal soils. Field trials conducted on heavy soil under Central European climatic conditions have shown that this species can produce substantial yields of both green and dry biomass even under limited soil resources and variable water availability [9]. Thus, cup plants represent a promising alternative to traditional energy crops such as giant miscanthus or willow, particularly in areas unsuitable for intensive agriculture. The species can also be integrated into agro energy systems, contributing to soil quality improvement and carbon sequestration [8,10]. Research has shown that perennial cultivation of S. perfoliatum increases water retention, reduces erosion, and supports soil organism development (Auerswald et al., 2025). Moreover, its extended flowering period benefits pollinator populations, aligning with biodiversity conservation efforts in agricultural landscapes. Therefore, integrating cup plant into agricultural practices can contribute both to renewable energy production and environmental enhancement [11]. Additionally, S. perfoliatum biomass can be utilized in various conversion technologies, including agricultural biogas plants, combustion, or as a component in solid fuels, increasing its market potential [12].
Excessive soil salinity is one of the most severe stress factors limiting plant growth and development [13,14]. Soil salinity induces osmotic stress, preventing water uptake from the substrate, leading to cellular dehydration, reduced turgor, and inhibited shoot elongation. Simultaneously, ionic stress arises from excessive sodium (Na⁺) and chloride (Cl⁻), potentially causing ion toxicity, nutrient imbalances, and deficiencies in essential elements such as potassium (K⁺) and calcium (Ca²⁺) [15,16]. At the physiological level, salt stress significantly affects plant water relations, including reduced hydraulic conductivity and limited root water uptake (dos Santos et al., 2022). Photosynthesis is also impaired due to decreased chlorophyll content, reduced photosystem activity, and lower CO₂ assimilation rates. Salt stress additionally influences metabolic enzyme activity and the production of reactive oxygen species (ROS), which may damage cell membranes, chloroplasts, and mitochondria [17,18]. Another adaptive mechanism involves modulation of plant hormone levels. Elevated abscisic acid (ABA) promotes stomatal closure and reduces transpiration, while changes in cytokinin and auxin concentrations regulate growth and cell division [19,20]. Prolonged salinity can also modify osmolyte composition, including accumulation of proline and soluble sugars, which protect cells under osmotic stress conditions [21,22].
Despite the cup plant’s broad environmental adaptability, detailed information on its tolerance to soil salinity particularly under different types of salt stress remains limited. Most research on plant salinity stress has focused on neutral salts (primarily NaCl), whereas many salt-affected soils also contain alkaline salts (such as NaHCO₃) that raise soil pH. How S. perfoliatum copes with alkaline versus neutral salinity is an open question and represents a clear research gap. Therefore, the aim of this study was to evaluate the physiological responses of cup plant to salt stress induced by two distinct salt types (neutral NaCl and alkaline NaHCO₃) applied at equal concentrations. We formulated a specific hypothesis that salt type would significantly influence the severity of plant stress responses. In particular, we hypothesized that alkaline salinity (NaHCO₃) would exert more deleterious effects on cup plant physiology than an equivalent concentration of neutral salinity (NaCl), due to the additional high-pH (bicarbonate) stress. Furthermore, a proportional, concentration-dependent decline in plant performance was expected under both salt types, with higher salt levels causing more pronounced reductions in chlorophyll content, photosynthetic efficiency, and gas exchange rates.
2. Materials and Methods
2.1. Pot Experiment
The pot experiment was conducted at the University of Rzeszów, Poland. Seeds of Silphium perfoliatum L. were obtained from the Institute of Plant Breeding and Acclimatization – National Research Institute, Bydgoszcz, Poland. The experiment was conducted in a growth chamber (model GC-300/1000, JEIO Tech Co., Ltd., South Korea) under the following conditions: temperature 22 ± 2 °C, relative humidity 60 ± 3%, photoperiod 16/8 h (day/night), and maximum light intensity of approximately 300 µE m⁻² s⁻¹. Seeds were sown in seed trays, and after germination, the seedlings were transplanted into 15 × 15 cm plastic pots, three per pot. The pots were filled with a slightly acidic substrate (pH: KCl 6.35; H₂O 6.52) with a sandy-loam consistency. Substrate moisture was maintained at 60% of field water capacity. One month after transplanting (plant height 8-12 cm), salt stress was induced by watering the soil with neutral (NaCl) or alkaline (NaHCO₃) salt solutions. Salt solutions were applied at concentrations of 100, 200, and 300 mM, at 100 ml per pot. Control plants were watered with the same volume of deionized water.
The salt concentrations of 100, 200, and 300 mM were selected to encompass a range from moderate to severe salinity stress in a short-term pot study. These levels ensured that both threshold effects (at the lower end) and extreme stress responses (at the higher end) could be observed. It is noted that 300 mM represents an unusually high salinity that would rarely occur homogeneously in field soils; however, such a concentration serves as a model stress condition to probe the upper limits of the plant's physiological tolerance in controlled settings. Using NaCl and NaHCO₃ as salt sources allowed us to simulate two distinct types of saline soils: neutral salinity (dominated by chloride, with minimal direct effect on soil pH) versus alkaline salinity (dominated by bicarbonate, causing elevated soil pH). By applying equal molar concentrations of Na⁺ in these two chemical forms, we could directly compare plant responses to neutral vs. alkaline salt stress under equivalent ionic strength.
The experiment was conducted with four biological replicates per treatment; each replicate consisted of three plants grown in a separate pot. All measurements were performed on each replicate pot (or its soil) with an appropriate technical replicate.
2.2. Physiological Measurements
Physiological measurements of the plants were taken four times on the first or second fully developed leaves at intervals of 7, 14, 21, and 28 days after the application of the salt solution to the soil. For physiological traits, repeated measurements within a pot were treated as technical replicates (subsamples), while pots were treated as biological replicates. Specifically, relative chlorophyll content and chlorophyll fluorescence parameters were recorded five times for each pot during each sampling period. Gas exchange parameters were recorded three times for each pot during each sampling period. Physiological parameters were assessed using methods previously described by Jańczak-Pieniążek et al. [23].
2.2.1. Relative Chlorophyll Content
Relative chlorophyll content was measured using a CCM-200 plus chlorophyll content meter (Opti-Sciences, Hudson, NH, USA). Measurements were performed five times per pot (on fully developed leaves), and the mean value was calculated for each pot and sampling term.
2.2.2. Chlorophyll Fluorescence
Chlorophyll fluorescence was measured using a Pocket PEA fluorimeter (Hansatech Instruments, King’s Lynn, Norfolk, UK) equipped with dark-adaptation clips. Before measurement leaves were dark-adapted for 30 minutes using these clips, which were attached to the upper part of the leaf blade while avoiding the main vein [24]. Fluorescence was recorded under red actinic light with a peak wavelength of 627 nm, applied for 1 s at a maximum intensity of 3500 μmol m⁻² s⁻¹ PAR. The following parameters were evaluated: maximum efficiency of water-splitting on the donor side of PSII (Fv/Fo), maximum photochemical efficiency of PSII (Fv/Fm), and the performance index (PI). Measurements were performed five times per pot (on fully developed leaves), and the mean value was calculated for each pot and sampling term.
2.2.3. Gas Exchange
Gas exchange parameters were measured using an LCpro-SD photosynthesis system (ADC Bioscientific Ltd., Herts, UK). The following parameters were recorded: net photosynthetic rate (Pn), stomatal conductance (gs), transpiration rate (E), and intercellular CO₂ concentration (Ci). During measurements, PAR intensity was set to 300 μmol m⁻² s⁻¹ and chamber temperature was maintained at 23 °C. Measurements were taken three times per pot (on fully developed leaves), and the mean value was calculated for each pot and sampling term.
2.3. Soil Analysis
Soil samples were air-dried, homogenized, and sieved through a 2 mm mesh. Representative subsamples were collected for the determination of soil pH and electrical conductivity (EC). Soil pH was measured potentiometrically in distilled water and in 1 M KCl solution, representing active and exchangeable acidity, respectively [25]. Soil suspensions were prepared at a 1:5 (w/w) ratio (soil:solution). Briefly, 5 g of soil were placed in 100 cm³ beakers and mixed with 25 cm³ of distilled water or 1 M KCl. The suspensions were stirred thoroughly and left to equilibrate for 24 h. After re-mixing, pH was measured using a pH meter (HI 4221, Hanna Instruments) at 20 °C. EC was measured in soil-water suspensions using a conductivity meter (HI 2316, Hanna Instruments). EC values, expressed in mS, were used as an indicator of soil salinity [26]. For each pot, soil pH was measured three times both in distilled water (pHH₂O) and in 1 M KCl (pHKCl). Soil electrical conductivity (EC) was also measured three times per pot, and mean values were used for further analyses.
2.4. Statistical Analysis
Statistical analysis was performed using TIBCO Statistica 13.3.0 software (TIBCO Software Inc., Palo Alto, California, USA). Normality of distribution was assessed using the Shapiro-Wilk test (α=0.05), and homogeneity of variance was confirmed using Levene's test (α=0.05). A two-way analysis of variance (ANOVA) with repeated measures (including time as a factor) was performed. Salinity relationships were also assessed using one-way analysis of variance (ANOVA). Significant differences between means were identified using Tukey's post hoc test at α≤0.05.
3. Results
3.1. Relative Chlorophyll Content
Chlorophyll content decreased under salt stress (Figure 1). Irrigation with 100 mM NaCl caused a significant reduction in chlorophyll content only in Term III. On the other terms, this concentration did not significantly affect the parameter compared to the control. Higher NaCl concentrations (200 and 300 mM) led to a significant decrease in chlorophyll content at all terms. Differences between these two concentrations were observed only in Terms II and IV. Application of NaHCO₃ also reduced chlorophyll content relative to the control, but the effect was less pronounced than that of NaCl. The lowest chlorophyll values were recorded at 200 and 300 mM NaHCO₃. At 100 mM NaHCO₃, chlorophyll content was similar to that observed at 300 mM NaCl. In Term I, all treatments except the control showed the lowest chlorophyll content. This value increased in Term II, but statistically significant differences were detected only for 200 mM NaHCO₃. In Term III, chlorophyll content decreased compared to Term IV; however, the difference was not statistically significant.
3.2. Chlorophyll Fluorescence
3.2.1. Maximum Efficiency of Water-splitting on the Donor Side of PSII (Fv/Fo)
Under salt stress, a decrease in the Fv/F₀ parameter was observed (Figure 2). Irrigation with neutral salt (NaCl) caused a significant reduction in Fv/Fo only at concentrations of 200 and 300 mM, which was observed across all terms. Alkaline salt induced a stronger decline in Fv/F₀ compared to neutral salt. No significant differences in Fv/Fo were detected at 300 mM NaCl or 100 mM NaHCO₃. The greatest decrease in Fv/Fo was observed at 200 and 300 mM for both salts; however, statistically significant differences compared to 100 mM were only found for 300 mM. In Term III, the lowest Fv/Fo values were recorded across all salinity treatments, but statistical significance was observed only for 100 mM NaCl compared to the other terms. The highest Fv/Fo values were recorded in Term II, yet significant differences were found only for 200 mM NaHCO₃ relative to the other terms.
3.2.2. Maximum Photochemical Efficiency of PSII (Fv/Fm)
Salt stress resulted in a reduction of the Fv/Fm parameter (Figure 3). Irrigation with neutral salt (NaCl) at 100 and 200 mM did not cause statistically significant differences compared to the control. Only the application of 300 mM NaCl led to a significant decrease in Fv/Fm across all terms. Alkaline salt further reduced Fv/Fm values. Irrigation with 100 and 200 mM NaHCO₃ did not differ from 300 mM NaCl. A significant reduction in Fv/Fm was observed only after the application of 300 mM NaHCO₃, specifically in Terms I, III, and IV. In Term II, the highest Fv/Fm values were recorded; however, statistically significant differences were detected only for 200 and 300 mM NaHCO₃ compared to the other terms.
3.2.3. Performance Index (PI)
As a result of applying salt stress, a decrease in the PI parameter value was observed relative to the control (Figure 4), except for the 100 mM and 200 mM concentrations of NaCl (Term II). A significant decrease was recorded when using the 300 mM concentration of neutral salt compared to the 100 mM concentration. A further reduction in the PI parameter value was found following the application of NaHCO₃, however, differences compared with the 200 mM neutral salt concentration were observed only at 300 mM. In Term II, a significantly highest PI value was recorded compared to the remaining Terms in all variants, except for the control, in which the value of this parameter did not differ between the Terms.
3.3. Gas Exchange
3.3.1. Net Photosynthetic Rate (Pn)
Soil salinity resulted in a reduction of the gas-exchange parameter Pn (Figure 5) compared to plants not subjected to this stress (control), except for the use of NaCl at a concentration of 100 mM (Term I and Term II). The application of NaHCO₃ caused a further decrease in Pn values. However, no statistically significant differences were found between the use of 300 mM neutral salt and the concentrations of 100 mM and 200 mM alkaline salt. The application of 300 mM alkaline salt resulted in a significantly lowest Pn value compared to the remaining salinity variants, except for the alkaline-salt concentrations of 100 mM (Term I) and 200 mM (Term I and Term III). In Term II, the highest value of the analyzed parameter was observed only in the variants with 100 mM and 200 mM alkaline salt.
3.3.2. Stomatal Conductance (gs)
Under salt stress, a decrease in the gs parameter value (Figure 6) was recorded relative to the control. However, when 100 mM NaCl was applied, no significant effect of salinity on the value of the parameter analyzed was observed. The use of 200 mM and 300 mM concentrations caused a significant decrease in gs compared to the control (except for 200 mM in Term III and Term IV). The application of NaHCO₃ led to a further reduction in gs values. The 300 mM neutral salt and 100 mM alkaline salt concentrations did not differ from each other. The lowest value of the analyzed parameter was recorded under the 300 mM alkaline salt concentration, compared to all other salt treatments except for the alkaline-salt concentrations of 200 mM and 100 mM (Term II). The highest gs value was observed in Term II, although it did not differ significantly from the values recorded in the remaining Terms in any of the experimental variants.
3.3.3. Transpiration Rate (E)
Soil salinity caused a decrease in the value of the E parameter (Figure 7) compared with the control, except for the variant with 100 mM NaCl (Term I and Term II). Higher concentrations of this salt (200 mM and 300 mM) led to a further decline in E relative to the control; however, no significant differences were found between these concentrations. No significant effect on the value of the analyzed parameter was observed between 300 mM neutral salt and 100 mM or 200 mM NaHCO₃. A significantly lowest E value was observed only under the 300 mM concentration, which was lower than the 100 mM concentration (except in Term II). In Term II, the lowest E value was recorded compared with the remaining Terms in all salinity variants, except for the 200 mM neutral salt concentration.
3.3.4. Intercellular CO₂ Concentration (Ci)
Under soil salinity, an increase in the Ci parameter value (Figure 8) was observed compared with the control, except for the variant in which 100 mM NaCl was applied (Term II). The application of 300 mM neutral salt caused a significant increase in Ci compared with the 200 mM concentration, but only in Term I and Term III. The use of 300 mM neutral salt did not result in changes compared with 100 mM NaHCO₃. The higher alkaline-salt concentration caused an increase in the value of the analyzed parameter compared with the lower concentrations. Under the 300 mM concentration, a significant increase in Ci was recorded compared with the 100 mM concentration, except in Term II. In Term III, the highest Ci value was observed across all salinity treatments; however, this was statistically confirmed only for the variants with 200 mM and 300 mM neutral salt and 100 mM alkaline salt.
3.4. Soil Analysis
3.4.1. Soil pH
Soil pH and salinity levels changed in response to the salt treatments, with clear differences between neutral and alkaline salt addition (Figure 10, Figure 11). NaHCO₃ tended to increase soil pH (measured in H₂O), whereas NaCl had little effect on pH. In fact, only the highest NaHCO₃ treatment (300 mM) caused a statistically significant rise in pHH₂O compared to the control (no-salt) soil, while 100 mM and 200 mM NaHCO₃ produced slight, non-significant pH increases. All NaCl-treated soils had pHH₂O values similar to the control. Soil pH measured in KCl (pHKCl) was consistently lower than pHH₂O and showed smaller absolute changes. Addition of NaCl caused no significant pHKCl change versus control, and NaHCO₃ caused only a modest increase. A significant elevation in pHKCl was observed only at 200 mM and 300 mM NaHCO₃, by approximately +4.8% and +6.7% relative to control, respectively.
3.4.2. Soil Electrical Conductivity (EC)
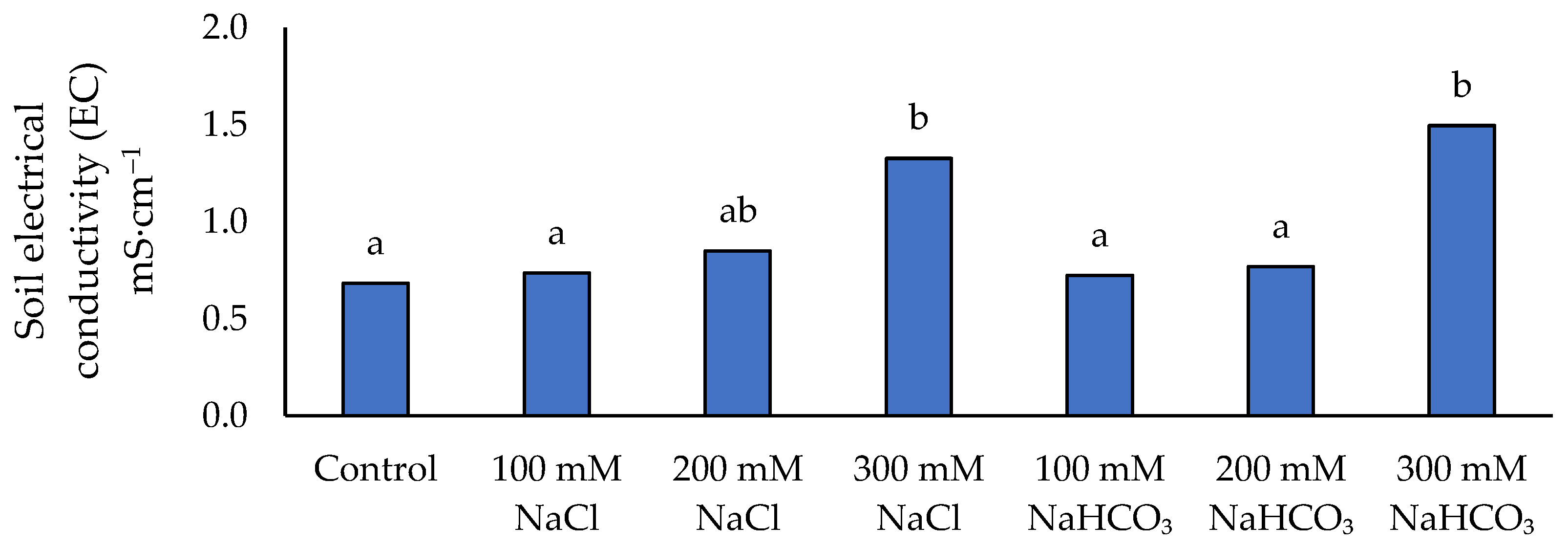
Soil electrical conductivity increased as a result of soil salinization (Figure 11). EC increased with increasing salt concentration for both salt types, reflecting higher soluble salt content. However, a statistically significant jump in EC (compared to control) was detected only at the highest concentration (300 mM). At 300 mM NaCl, the soil EC was about 92.8% above the control value, whereas 300 mM NaHCO₃ led to an EC increase of approximately 117.4% over control. These results indicate that substantial salinization of the soil occurred only under the most severe treatment, with NaHCO₃ contributing slightly more to EC rise than NaCl at the same molarity.
4. Discussion
In the conducted study, the effects of salt stress induced by neutral and alkaline salts at concentrations of 100, 200, and 300 mM on the physiological responses of Silphium perfoliatum L. were evaluated. This species, due to its high biomass productivity and perennial growth habit, is considered a crop with strong potential for bioenergy systems [10]. Therefore, assessing its response to soil salinity is important in the context of future cultivation on degraded, marginal, and salinizing lands [27]. The results demonstrated that salt stress induced by both NaCl and NaHCO₃ significantly reduced the physiological parameters analyzed. Soil salinity, as a typical abiotic stressor, disrupts key physiological and biochemical processes in plants. Excessive accumulation of Na⁺ ions lead to ionic imbalance, a decrease in soil water potential, and difficulties in water uptake, as confirmed by previous studies [28,29]. High salt concentrations also adversely affect photosynthesis. Components necessary for the functioning of the photosynthetic apparatus, such as chlorophylls, photosystems, and carbon metabolism enzymes, undergo degradation under salt stress [30].
In the present study, reductions in chlorophyll content and fluorescence parameters were observed, indicating impaired photosystem II efficiency and disturbances in photochemical processes. At the same time, salinity decreased gas exchange parameters—net photosynthesis (Pn), stomatal conductance (gs), and transpiration (E)—confirming constraints on CO₂ uptake and plant water regulation [31,32]. These effects intensified with increasing salt concentrations, particularly under NaHCO₃ treatment, consistent with previous findings that alkaline salts are more toxic than neutral [33,34]. The negative impact of salinity on photosynthesis is multilevel, encompassing osmotic, ionic, and biochemical stress [21,29,35]. Osmotic stress limits water availability and induces stomatal closure, leading to reduced gs and Pn [19,21,22]. Ionic stress, caused by excess Na⁺ and Cl⁻, disrupts the transport of ions essential for chlorophyll biosynthesis, destabilizes cell membranes, and damages thylakoids, thereby reducing the efficiency of light energy conversion [37,38]. Similar damage to the photosynthetic apparatus under high salinity has been reported in seedlings of Robinia pseudoacacia [39]. Alkaline salts such as NaHCO₃ are more toxic than neutral salts because they induce both ionic stress and substrate alkalization. Elevated soil pH reduces the availability of key ions (Mg²⁺, Fe²⁺, Mn²⁺) necessary for chlorophyll biosynthesis and photosystem functioning. Consequently, chlorophyll content, chlorophyll fluorescence, and CO₂ assimilation decrease, as confirmed by other studies [32,33]. Additionally, the HCO₃⁻ ion can destabilize cellular and thylakoid membranes, contributing to pigment degradation and reduced photosystem efficiency [32]. Under alkaline stress, increased production of ROS leads to lipid peroxidation, chlorophyll degradation, and damage to Calvin cycle enzymes, including RuBisCO [18,39]. These further decreases photosynthetic parameters (Pn, gs, E). Stomatal closure, a protective response, additionally limits CO₂ uptake, intensifying the decline in photosynthetic performance [33,38].
At later measurement dates, a partial improvement in physiological parameters was observed, indicating the activation of multilayered adaptive mechanisms that enable plant functioning under prolonged salt stress. In the adaptive phase, plants enhance the activity of enzymatic components of the antioxidant system, such as superoxide dismutase (SOD) and catalase (CAT). These enzymes effectively neutralize excess ROS, which accumulate rapidly during the initial phase of stress and cause damage to membrane lipids, proteins, and photosynthetic structures [40]. Earlier studies by Rohman et al. [41] and Rohman et al. [42] also reported increased SOD activity under salt stress, confirming the activation of protective mechanisms aimed at reducing oxidative damage. Simultaneously, plants intensify the synthesis of osmolytes proline, glycine betaine, soluble sugars, and osmoregulatory proteins. These compounds decrease the osmotic potential of the cytoplasm, allowing the maintenance of turgor and continuation of metabolic processes despite limited water availability [43,44]. During later measurement periods, a partial recovery of physiological parameters was again observed, suggesting further activation of adaptive mechanisms enabling plant functioning under prolonged salt exposure. A similar trend was reported by Jańczak-Pieniążek et al. [45] in wheat seedlings, where improved physiological parameters at later stages were attributed to repair processes. Together, these mechanisms indicate that plants not only respond defensively to salt stress but also undergo physiological and biochemical adaptation that enables relatively stable functioning despite persistent environmental stressors [46].
In all treatments, soil pH measured in KCl was lower than pH in H₂O. This is a well-known effect of using a saline extract, which displaces exchangeable H⁺ from soil colloids and thus produces a lower pH reading. Notably, adding NaHCO₃ significantly increased soil pH in H₂O, while causing only a small increase in pHKCl. This indicates that much of the added alkalinity was buffered by exchangeable soil acidity (H⁺ ions at cation exchange sites), so the net increase in exchangeable pH was limited. Electrical conductivity (EC) values reflected the degree of salinity: only the 300 mM treatments caused a sharp increase in EC, confirming that these soils had become highly saline. Such a high EC corresponds to a very low soil water potential, effectively inducing osmotic drought for plants [47]. This means that at a concentration of 300 mM, in addition to specific ionic effects, plants likely experienced severe limitations in water uptake due to the high salt concentration in the soil solution. Besides osmotic stress, the type of salt imposed qualitatively different stresses on plants. Neutral salt (NaCl) supplies an excess of Na⁺ and Cl⁻ ions, which can lead to ion-specific toxicity and nutrient imbalances (e.g., Na⁺ competes with K⁺ uptake, Cl⁻ antagonizes NO₃⁻ uptake) [48,49], along with the osmotic effects of elevated solute concentrations. Alkaline salt (NaHCO₃) also introduces Na⁺ but also drastically raises the pH of the soil solution. High environmental pH can cause precipitation or reduced availability of essential nutrients such as iron, calcium, and magnesium. For example, Fe²⁺ can precipitate as Fe(OH)₃ under alkaline conditions [50]. This induced nutrient deficiency (often manifested by iron chlorosis and other symptoms) adds an additional layer of stress to the plant. Consequently, NaHCO₃ treatments were more detrimental to the plant than NaCl treatments at equivalent salt levels. In our results, chlorophyll content and photosynthetic efficiency decreased more under NaHCO₃ treatment, consistent with the combination of stresses (salinity and alkalinity) in these treatments. In summary, the presence of bicarbonate and the resulting high soil pH intensified the negative effects of salinity, making alkaline salt stress significantly more detrimental to S. perfoliatum than neutral salt stress under the conditions of this experiment.
The limitations of this study should also be noted. This study was conducted in a growth-chamber pot environment, which, while allowing tight control of variables, does not fully replicate field conditions. The controlled setting (uniform soil medium, constant climate) lacks the complexity of natural habitats. Factors such as rainfall variability, soil heterogeneity, and microbial interactions were not represented. The salt levels used (100-300 mM NaCl or NaHCO₃) were relatively high and imposed acutely; these concentrations effectively induced stress responses, but such extreme salinity might seldom occur homogeneously in open-field soils. The experiment also focused on short-term physiological responses and did not assess longer-term adaptation, growth, or reproductive effects under salinity. Additionally, we measured primarily physiological and growth indicators; other parameters like tissue ion accumulation, osmolyte levels, or antioxidant enzyme activities were not examined and could be topics for future research to elucidate the plant’s internal tolerance mechanisms. These limitations should be kept in mind when extrapolating our findings. Nonetheless, the controlled approach and replicated design provided clear, comparative insights into how neutral vs. alkaline salinity affect cup plant physiology. The results serve as a valuable baseline for understanding S. perfoliatum responses to salt stress and will inform subsequent studies under more complex or long-term conditions.
5. Conclusions
This experiment demonstrated that soil salinity whether caused by NaCl or NaHCO₃ markedly impairs the photosynthetic performance and physiological status of cup plant (Silphium perfoliatum). All measured indicators of plant function (chlorophyll content, chlorophyll fluorescence parameters, gas exchange rates, etc.) showed significant declines with increasing soil salt concentration, confirming that salinity stress directly limits the species’ photosynthetic efficiency and overall vigor in a controlled setting. Importantly, the type of salinity influenced the severity of the response: alkaline salt (NaHCO₃) consistently caused more pronounced physiological disruptions than an equivalent concentration of neutral salt (NaCl). In other words, high bicarbonate levels (alkaline pH) exacerbated the detrimental effects of salinity beyond the osmotic and ionic stress imposed by NaCl alone. Overall, S. perfoliatum exhibited substantial sensitivity to elevated salinity, especially under alkaline conditions, as evidenced by sharp reductions in chlorophyll content, photosynthetic gas exchange, and photochemical efficiency at high salt levels. These conclusions delineate the short-term physiological limits of cup plant under saline conditions. Future studies should build on this work by examining longer-term and field-scale responses to salinity in S. perfoliatum, in order to fully assess its capacity to maintain growth and productivity on salt-affected soils.
Author Contributions
Conceptualization, M.J.-P. and D.M.; methodology, M.J.-P., K.S.; and D.M.; formal analysis, M.J.-P. and D.M.; investigation, M.J.-P.; M.K.; K.S. and D.M; writing—original draft preparation, M.J.-P.; M.K.; K.S. and D.M.; writing—review and editing, M.J.-P.; K.S. and D.M.; supervision, M.J.-P.; funding acquisition, M.J.-P. and D.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Minister of Science of the Republic of Poland under the Programme “Regional initiative of excellence”, Agreement No. RID/SP/0010/2024/1, and from the funds of the Minister of Science of the Republic of Poland allocated to the activities of the Faculty of Technology and Life Sciences of the University of Rzeszów.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available from the corresponding author upon reasonable request.
Acknowledgments
The authors gratefully acknowledge dr Włodzimierz Majtkowski (Institute of Plant Breeding and Acclimatization – National Research Institute, Bydgoszcz, Poland) for providing plant material of cup plant (Silphium perfoliatum L.).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| Fv/Fm | maximum photochemical efficiency of PSII |
| Fv/Fo | maximum efficiency of water-splitting on the donor side of PSII |
| PI | PSII performance index |
| PN | net photosynthetic rate |
| Fv/Fm | maximum photochemical efficiency of PSII |
| gs | stomatal conductance |
| Ci | intercellular CO₂ concentration |
| gs | stomatal conductance |
References
- Cronin, J.; Anandarajah, G.; Dessens, O. Climate change impacts on the energy system: a review of trends and gaps. Clim. Change 2018, 151, 79–93. [Google Scholar] [CrossRef]
- Zhou, P.; Lv, Y.; Wen, W. The low-carbon transition of energy systems: A bibliometric review from an engineering management perspective. Eng. 2023, 29, 147–158. [Google Scholar] [CrossRef]
- Yang, C.; Kwon, H.; Bang, B.; Jeong, S.; Lee, U. Role of biomass as low-carbon energy source in the era of net zero emissions. Fuel 2022, 328, 125206. [Google Scholar] [CrossRef]
- Mignogna, D.; Szabó, M.; Ceci, P.; Avino, P. Biomass energy and biofuels: Perspective, potentials, and challenges in the energy transition. Sustainability 2024, 16, 7036. [Google Scholar] [CrossRef]
- Katerla, J.; Sornek, K. Biomass for residential heating: A review of technologies, applications, and sustainability aspects. Energies 2025, 18, 5875. [Google Scholar] [CrossRef]
- Gerwin, W.; McBratney, A.; Tóth, G.; Montanarella, L.; Gardi, C.; van der Meijden, G. Assessment and quantification of marginal lands for biomass production in Europe using soil quality indicators. Soil Discuss. 2018, 4, 267–287. [Google Scholar] [CrossRef]
- Cossel, M.V.; Lewandowski, I.; Elbersen, B.; Staritsky, I.; Van Eupen, M.; Iqbal, Y.; Mantel, S.; Scordia, D.; Testa, G.; Cosentino, S.L.; et al. Marginal agricultural land low-input systems for biomass production. Energies 2019, 12, 3123. [Google Scholar] [CrossRef]
- Bury, M.; Możdżer, E.; Kitczak, T.; Siwek, H.; Włodarczyk, M. Yields, calorific value and chemical properties of cup plant Silphium perfoliatum L. biomass, depending on the method of establishing the plantation. Agronomy 2020, 10, 851. [Google Scholar] [CrossRef]
- Tóth, Š. The yield of green phytomass of Silphium perfoliatum L., newly-introduced energy crop tested on marginal heavy soils under Central European continental climate. J. Cent. Eur. Agric. 2023, 24, 374–390. [Google Scholar] [CrossRef]
- Peni, D.; Stolarski, M.J.; Bordiean, A.; Krzyżaniak, M.; Dębowski, M. Silphium perfoliatum—A herbaceous crop with increased interest in recent years for multi-purpose use. Agriculture 2020, 10, 640. [Google Scholar] [CrossRef]
- Gansberger, M.; Montgomery, L.F.R.; Liebhard, P. Botanical characteristics, crop management and potential of Silphium perfoliatum L. as a renewable resource for biogas production: A review. Ind. Crops Prod. 2015, 63, 362–372. [Google Scholar] [CrossRef]
- Koniuszy, A.; Hawrot-Paw, M.; Podsiadło, C.; Sędłak, P.; Możdżer, E. Gasification of cup plant (Silphium perfoliatum L.) biomass–Energy recovery and environmental impacts. Energies 2020, 13, 4960. [Google Scholar] [CrossRef]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef]
- Balasubramaniam, T.; Shen, G.; Esmaeili, N.; Zhang, H. Plants’ response mechanisms to salinity stress. Plants 2023, 12, 2253. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of plant responses to salt stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef]
- dos Santos, T.B.; Ribas, A.F.; de Souza, S.G.H.; Budzinski, I.G.F.; Domingues, D.S. Physiological responses to drought, salinity, and heat stress in plants: A review. Stresses 2022, 2, 113–135. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genomics 2014, 701596. [Google Scholar] [CrossRef] [PubMed]
- Hameed, A.; Ahmed, M.Z.; Hussain, T.; Aziz, I.; Ahmad, N.; Gul, B.; Nielsen, B.L. Effects of salinity stress on chloroplast structure and function. Cells 2021, 10, 2023. [Google Scholar] [CrossRef]
- Liu, C.; Jiang, X.; Yuan, Z. Plant responses and adaptations to salt stress: A review. Horticulturae 2024, 10, 1221. [Google Scholar] [CrossRef]
- Mahajan, M.; Poor, P.; Kaur, H.; Aher, R.R.; Palakolanu, S.R.; Khan, M.I.R. Salt stress tolerance and abscisic acid in plants: Associating role of plant growth regulators and transcription factors. Plant Physiol. Biochem. 2025, 228, 110303. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.; Wang, Y.; Yan, Y.; Liu, Y.; Wang, J.; Chen, S. A review on plant responses to salt stress and their mechanisms of salt resistance. Horticulturae 2021, 7, 132. [Google Scholar] [CrossRef]
- Singh, P.; Choudhary, K.K.; Chaudhary, N.; Gupta, S.; Sahu, M.; Tejaswini, B.; Sarkar, S. Salt stress resilience in plants mediated through osmolyte accumulation and its crosstalk mechanism with phytohormones. Front. Plant Sci. 2022, 13, 1006617. [Google Scholar] [CrossRef] [PubMed]
- Jańczak-Pieniążek, M.; Migut, D.; Piechowiak, T.; Balawejder, M. Enhancement of photosynthetic efficiency and antioxidant response in wheat under drought stress by quercetin–copper complex. Int. J. Mol. Sci. 2025, 26, 10365. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—a practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Zondo, N.; Ntuli, N.R.; Mavengahama, S.; Van Jaarsveld, C.M. Improving growth and yield of Cucurbita argyrosperma with goat manure. Front. Plant Sci. 2025, 16, 1658365. [Google Scholar] [CrossRef] [PubMed]
- Hmidi, O.; Srarfi, F.; Brahim, N.; Bambina, P.; Lo Papa, G. Predicting Soil Electrical Conductivity of Saturated Paste Extract Using Pedotransfer Functions in Northeastern Tunisia. Sustainability 2025, 17, 9177. [Google Scholar] [CrossRef]
- Gazoulis, I.; Pyliou, K.; Kokkini, M.; Danaskos, M.; Kanatas, P.; Travlos, I. Cup plant (Silphium perfoliatum): Agronomy, uses, and potential role for land restoration. Land 2025, 14, 1307. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef]
- Shelar, P.V.; Mankar, G.D.; Sontakke, O.P.; Wayase, U.R.; Bhosale, K.S.; Nikalje, G.C.; Ahire, M.L.; Nikam, T.D.; Barmukh, R.B. A review on physio-biochemical and molecular mechanisms of salt tolerance in crops. Curr. Agri Res 2024, 12, 2. [Google Scholar] [CrossRef]
- Tsai, Y.C.; Chen, K.C.; Cheng, T.S.; Lee, C.; Lin, S.H.; Tung, C.W. Chlorophyll fluorescence analysis in diverse rice varieties reveals the positive correlation between the seedlings salt tolerance and photosynthetic efficiency. BMC Plant Biol. 2019, 19, 403. [Google Scholar] [CrossRef]
- Hnilickova, H.; Kraus, K.; Vachova, P.; Hnilicka, F. Salinity stress affects photosynthesis, malondialdehyde formation, and proline content in Portulaca oleracea L. Plants 2021, 10, 845. [Google Scholar] [CrossRef]
- Shen, C.; Yang, W.; Kang, Y.; Qin, S.; Zhang, W.; Liu, Y.; Qian, S.; Han, Y. Effect of alkaline salt stress on photosynthetic activities of potato plants (Solanum tuberosum L.). Plants 2025, 14, 2979. [Google Scholar] [CrossRef]
- Guo, R.; Shi, L.; Yan, C.; et al. Ionomic and metabolic responses to neutral salt or alkaline salt stresses in maize (Zea mays L.) seedlings. BMC Plant Biol. 2017, 17, 41. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Yu, L.; Hou, Y.; Zhang, Y.; Guo, W.; Xue, Y. Contrasting effects of NaCl and NaHCO3 stresses on seed germination, seedling growth, photosynthesis, and osmoregulators of the common bean (Phaseolus vulgaris L.). Agronomy 2019, 9, 409. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Z.; Sui, N. Sensitivity and responses of chloroplasts to salt stress in plants. Front. Plant Sci. 2024, 15, 1374086. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kaiser, E.; Li, T.; Marcelis, L.F.M. NaCl affects photosynthetic and stomatal dynamics by osmotic effects and reduces photosynthetic capacity by ionic effects in tomato. J. Exp. Bot. 2022, 73, 3637–3650. [Google Scholar] [CrossRef] [PubMed]
- Shu, S.; Guo, S.R.; Sun, J.; Yuan, L.Y. Effects of salt stress on the structure and function of the photosynthetic apparatus in Cucumis sativus and its protection by exogenous putrescine. Physiol. Plant. 2012, 146, 285–296. [Google Scholar] [CrossRef]
- Lu, X.; Ma, L.; Zhang, C.; et al. Grapevine (Vitis vinifera) responses to salt stress and alkali stress: Transcriptional and metabolic profiling. BMC Plant Biol. 2022, 22, 528. [Google Scholar] [CrossRef]
- Wang, X.; Wang, W.; Huang, J.; Peng, S.; Xiong, D. Diffusional conductance to CO2 is the key limitation to photosynthesis in salt-stressed leaves of rice (Oryza sativa). Physiol. Plant. 2018, 163, 45–58. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Rohman, M.M.; Talukder, M.Z.A.; Hossain, M.G.; Uddin, M.S.; Amiruzzaman, M.; Biswas, A.; Ahsan, A.F.M.S.; Chowdhury, M.A.Z. Saline sensitivity leads to oxidative stress and increases the antioxidants in presence of proline and betaine in maize (Zea mays L.) inbred. Plant Omics J. 2016, 9, 35–47. [Google Scholar]
- Rohman, M.M.; Islam, M.R.; Monsur, M.B.; Amiruzzaman, M.; Fujita, M.; Hasanuzzaman, M. Trehalose protects maize plants from salt stress and phosphorus deficiency. Plants 2019, 8, 568. [Google Scholar] [CrossRef] [PubMed]
- Parvin, K.; Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Nahar, K.; Mohsin, S.M.; Fujita, M. Comparative physiological and biochemical changes in tomato (Solanum lycopersicum L.) under salt stress and recovery: Role of antioxidant defense and glyoxalase systems. Antioxidants 2019, 8, 350. [Google Scholar] [CrossRef]
- Wang, D.; Gao, Y.; Sun, S.; Lu, X.; Li, Q.; Li, L.; Wang, K.; Liu, J. Effects of salt stress on the antioxidant activity and malondialdehyde, solution protein, proline, and chlorophyll contents of three Malus species. Life 2022, 12, 1929. [Google Scholar] [CrossRef]
- Jańczak-Pieniążek, M.; Migut, D.; Piechowiak, T.; Balawejder, M. Assessment of the impact of the application of a quercetin–copper complex on the course of physiological and biochemical processes in wheat plants (Triticum aestivum L.) growing under saline conditions. Cells 2022, 11, 1141. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Yu, Y.; Chen, H.; Feng, Y.; Chen, S.; Zhang, H.; Zhou, H.; Meng, L.; Wang, Y. Response of photosynthetic characteristics and antioxidant system in the leaves of safflower to NaCl and NaHCO3. Plant Cell Rep. 2024, 43, 146. [Google Scholar] [CrossRef]
- Wang, G.; Shen, W.; Zhang, Z.; Guo, S.; Hu, J.; Feng, R.; Zhao, Q.; Du, J.; Du, Y. The Effect of Neutral Salt and Alkaline Stress with the Same Na+ Concentration on Root Growth of Soybean (Glycine max (L.) Merr.) Seedlings. Agronomy 2022, 12, 2708. [Google Scholar] [CrossRef]
- Guo, J.; Liu, L.; Du, M.; Tian, H.; Wang, B. Cation and Zn Accumulation in Brown Seeds of the Euhalophyte Suaeda salsa Improves Germination Under Saline Conditions. Front. Plant Sci. 2020, 11, 602427. [Google Scholar] [CrossRef]
- Lin, J.; Hua, X.; Peng, X.; Dong, B.; Yan, X. Germination Responses of Ryegrass (Annual vs. Perennial) Seed to the Interactive Effects of Temperature and Salt-Alkali Stress. Front. Plant Sci. 2018, 9, 1458. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, Q.; Cai, H.; Xu, F. Ethylene Mediates Alkaline-Induced Rice Growth Inhibition by Negatively Regulating Plasma Membrane H(+)-ATPase Activity in Roots. Front. Plant Sci. 2017, 8, 1839. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Impact of NaCl and NaHCO₃ salt stress and sampling time on relative chlorophyll content. Different uppercase letters indicate significant differences among sampling terms within each treatment, while different lowercase letters indicate significant differences among treatments within the same sampling term, based on ANOVA followed by Tukey’s HSD test (p ≤ 0.05).
Figure 1.
Impact of NaCl and NaHCO₃ salt stress and sampling time on relative chlorophyll content. Different uppercase letters indicate significant differences among sampling terms within each treatment, while different lowercase letters indicate significant differences among treatments within the same sampling term, based on ANOVA followed by Tukey’s HSD test (p ≤ 0.05).
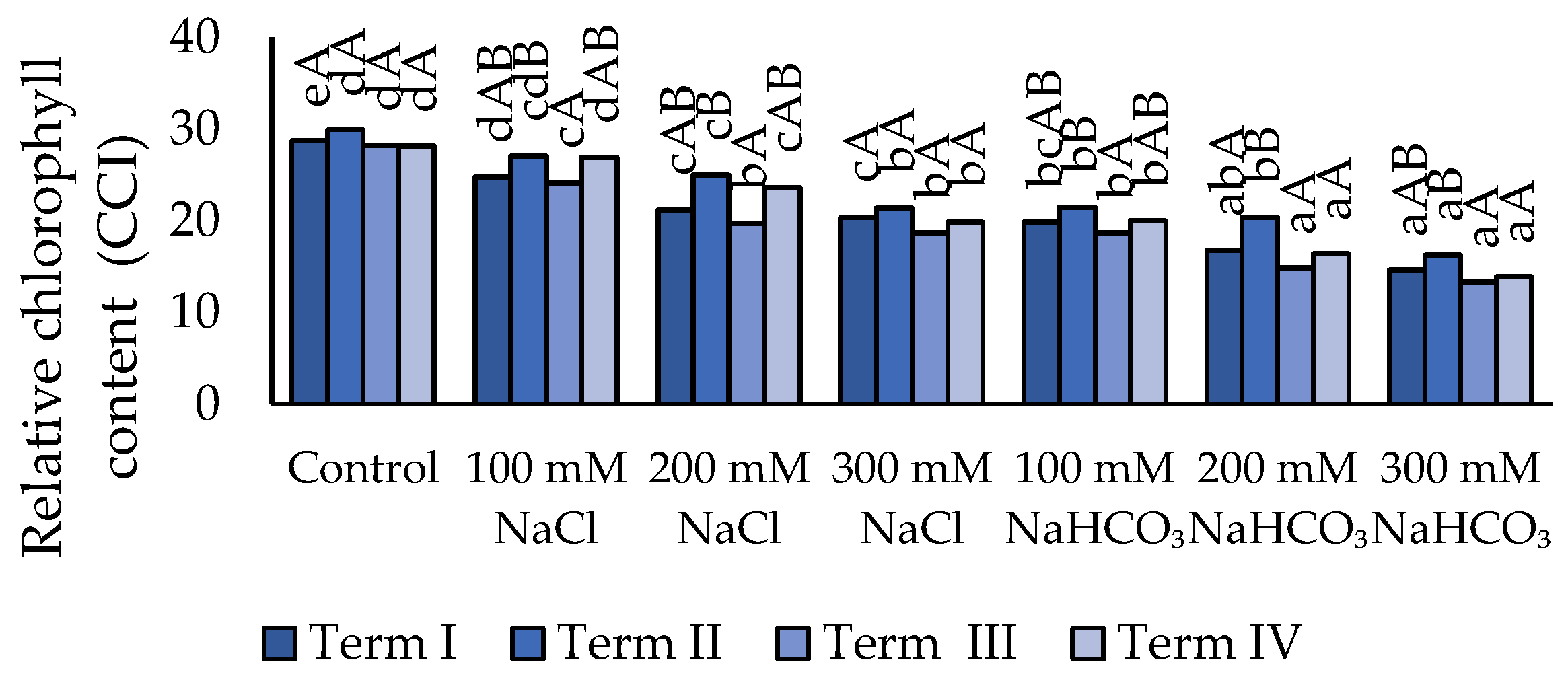
Figure 2.
Impact of NaCl and NaHCO₃ salt stress and sampling time on maximum efficiency of water-splitting on the donor side of PSII (Fv/Fo). Different uppercase letters indicate significant differences among sampling terms within each treatment, while different lowercase letters indicate significant differences among treatments within the same sampling term, based on ANOVA followed by Tukey’s HSD test (p ≤ 0.05).
Figure 2.
Impact of NaCl and NaHCO₃ salt stress and sampling time on maximum efficiency of water-splitting on the donor side of PSII (Fv/Fo). Different uppercase letters indicate significant differences among sampling terms within each treatment, while different lowercase letters indicate significant differences among treatments within the same sampling term, based on ANOVA followed by Tukey’s HSD test (p ≤ 0.05).
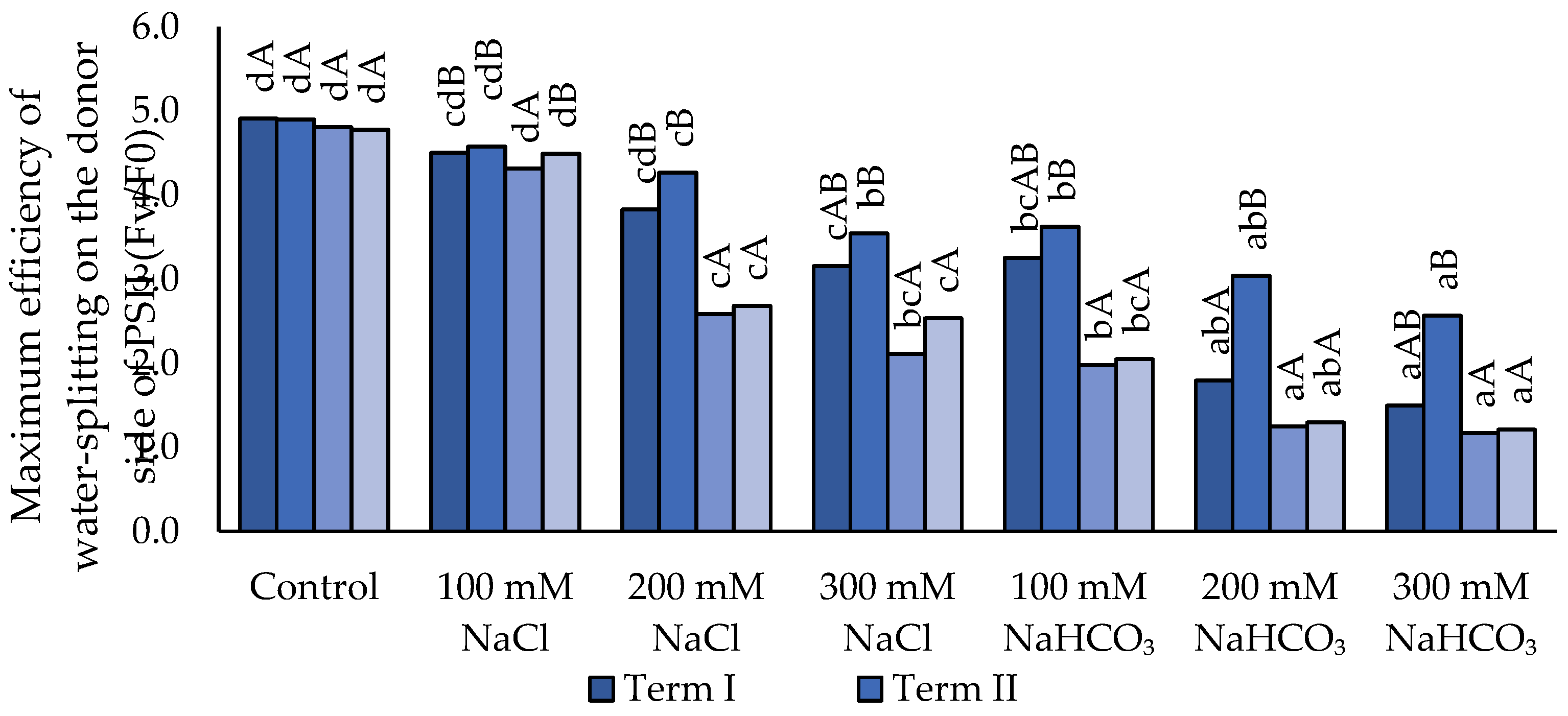
Figure 3.
Impact of NaCl and NaHCO₃ salt stress and sampling time on maximum photochemical efficiency of PSII (Fv/Fm). Different uppercase letters indicate significant differences among sampling terms within each treatment, while different lowercase letters indicate significant differences among treatments within the same sampling term, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
Figure 3.
Impact of NaCl and NaHCO₃ salt stress and sampling time on maximum photochemical efficiency of PSII (Fv/Fm). Different uppercase letters indicate significant differences among sampling terms within each treatment, while different lowercase letters indicate significant differences among treatments within the same sampling term, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
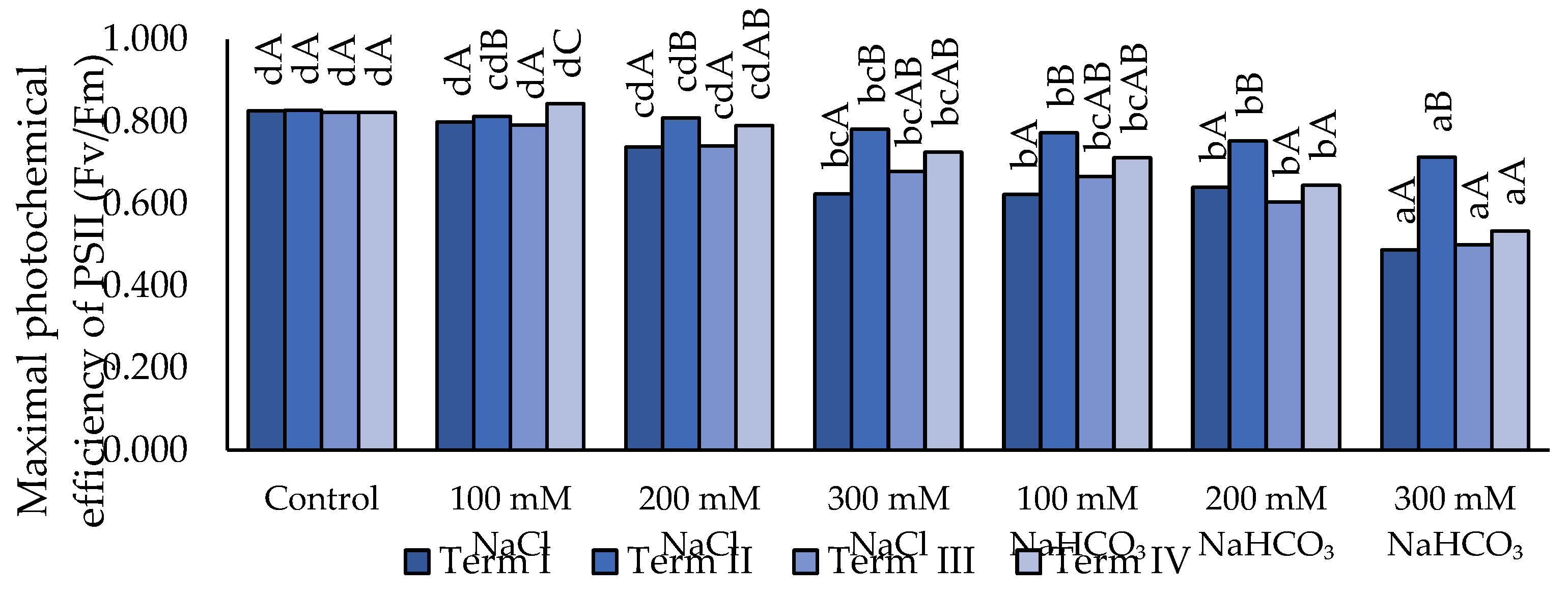
Figure 4.
Impact of NaCl and NaHCO₃ salt stress and sampling time on performance index (PI). Different uppercase letters indicate significant differences among sampling terms within each treatment, while different lowercase letters indicate significant differences among treatments within the same sampling term, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
Figure 4.
Impact of NaCl and NaHCO₃ salt stress and sampling time on performance index (PI). Different uppercase letters indicate significant differences among sampling terms within each treatment, while different lowercase letters indicate significant differences among treatments within the same sampling term, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
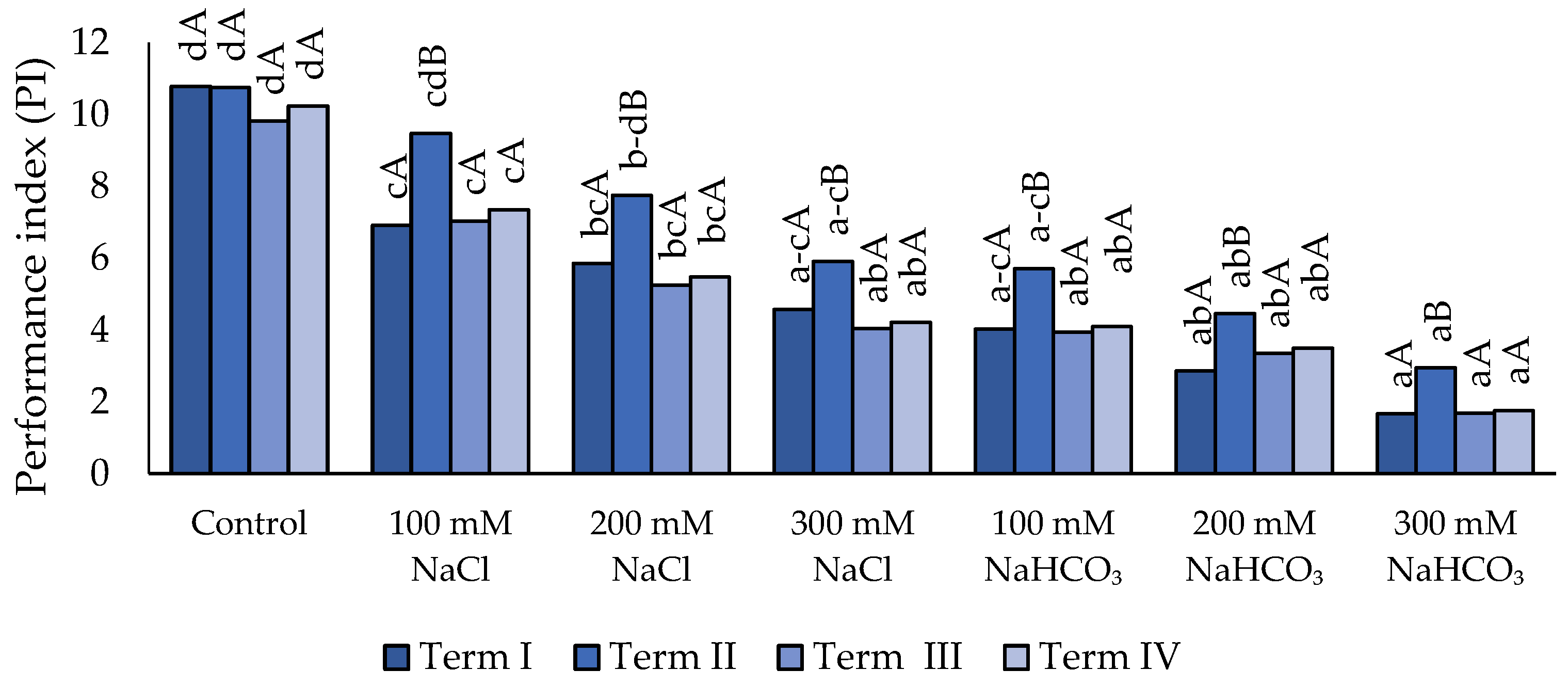
Figure 5.
Impact of NaCl and NaHCO₃ salt stress and sampling time on net photosynthetic rate (Pn). Different uppercase letters indicate significant differences among sampling terms within each treatment, while different lowercase letters indicate significant differences among treatments within the same sampling term, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
Figure 5.
Impact of NaCl and NaHCO₃ salt stress and sampling time on net photosynthetic rate (Pn). Different uppercase letters indicate significant differences among sampling terms within each treatment, while different lowercase letters indicate significant differences among treatments within the same sampling term, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
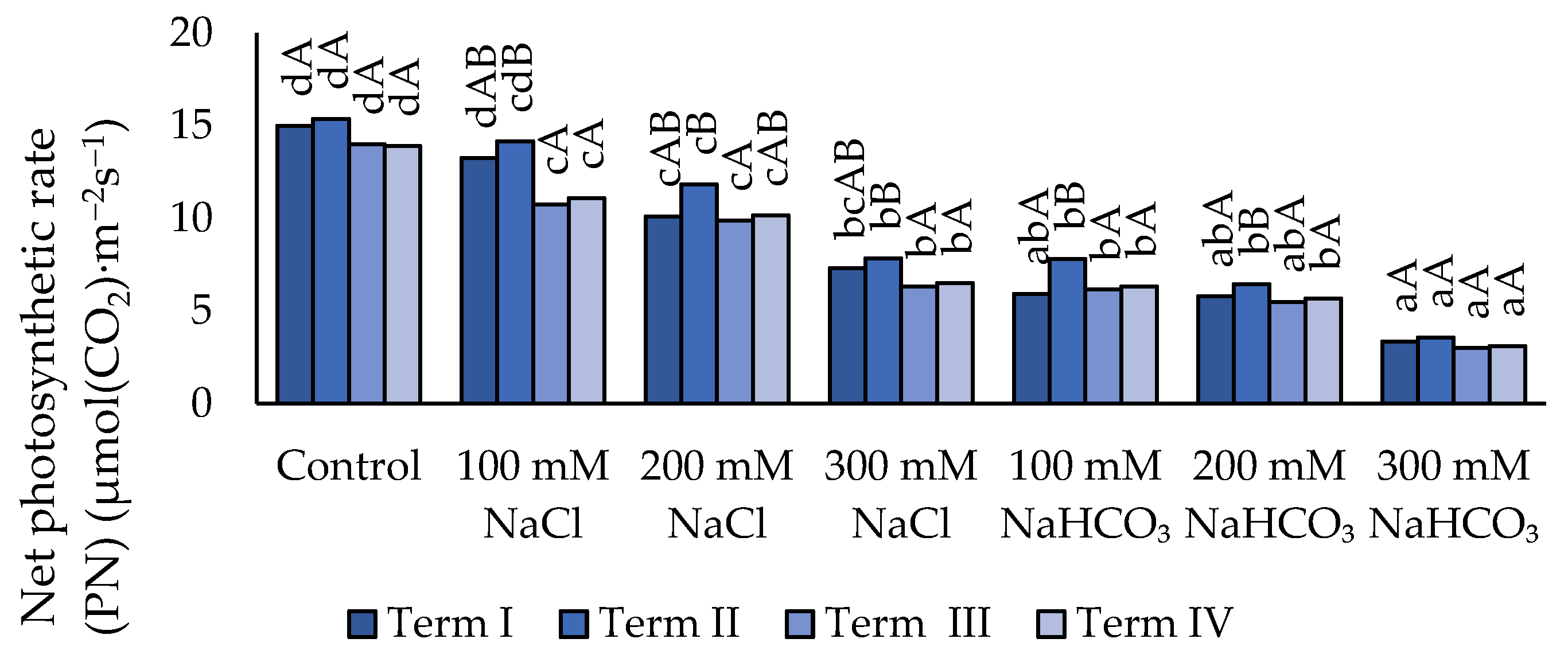
Figure 6.
Impact of NaCl and NaHCO₃ salt stress and sampling time on stomatal conductance (gs). Different uppercase letters indicate significant differences among sampling terms within each treatment, while different lowercase letters indicate significant differences among treatments within the same sampling term, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
Figure 6.
Impact of NaCl and NaHCO₃ salt stress and sampling time on stomatal conductance (gs). Different uppercase letters indicate significant differences among sampling terms within each treatment, while different lowercase letters indicate significant differences among treatments within the same sampling term, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
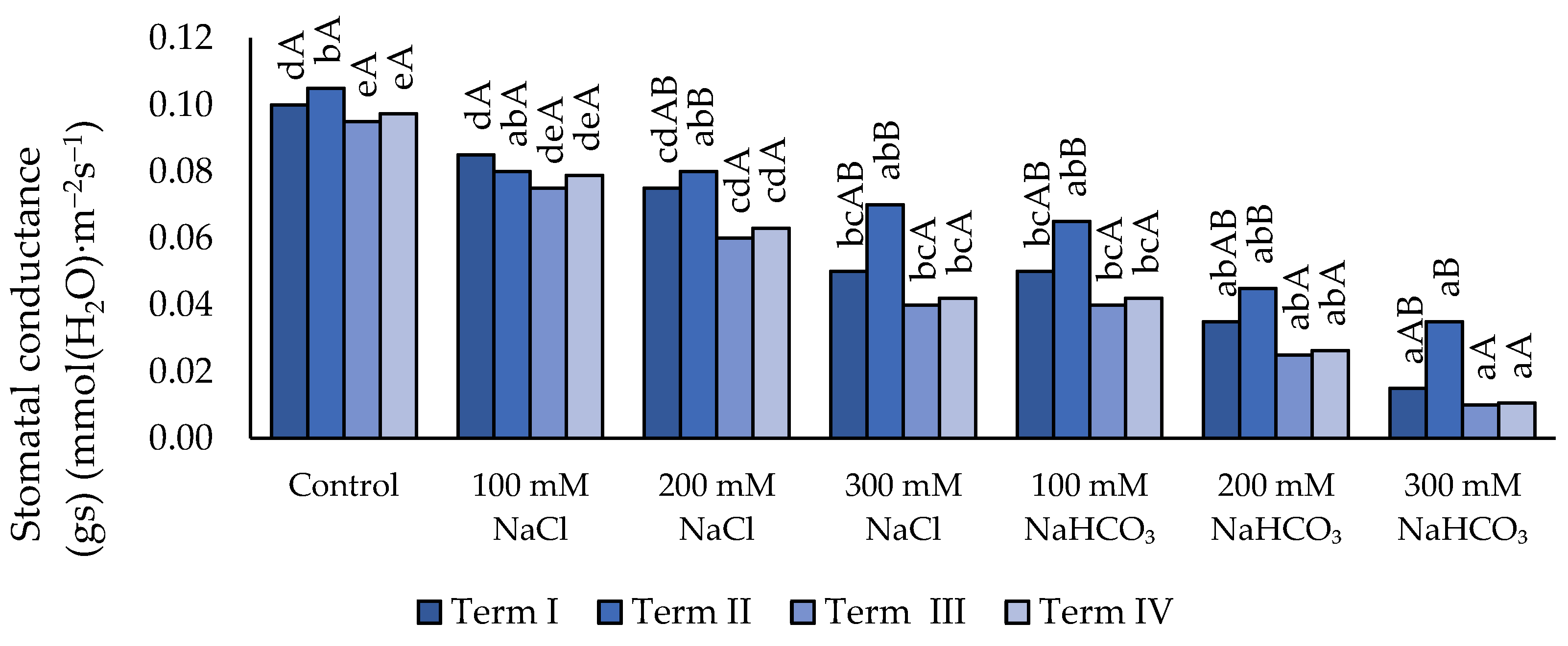
Figure 7.
Impact of NaCl and NaHCO₃ salt stress and sampling time on transpiration rate (E). Different uppercase letters indicate significant differences among sampling terms within each treatment, while different lowercase letters indicate significant differences among treatments within the same sampling term, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
Figure 7.
Impact of NaCl and NaHCO₃ salt stress and sampling time on transpiration rate (E). Different uppercase letters indicate significant differences among sampling terms within each treatment, while different lowercase letters indicate significant differences among treatments within the same sampling term, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
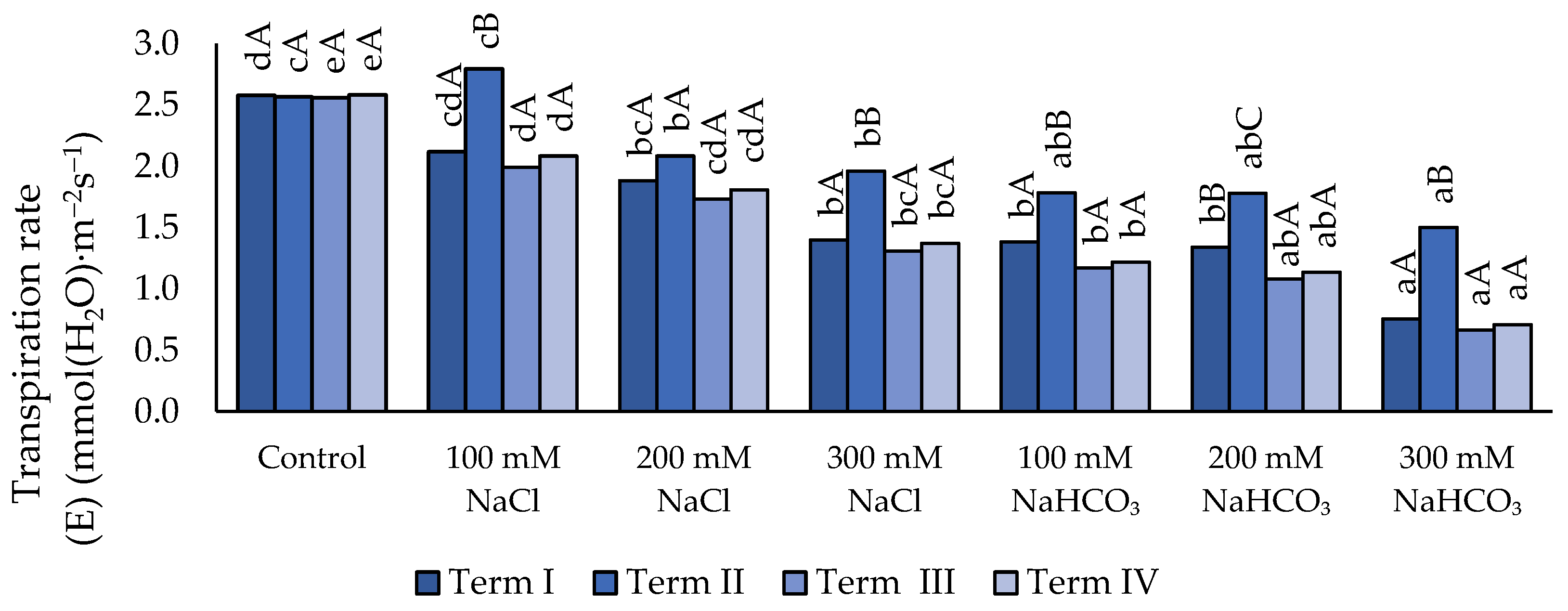
Figure 8.
Impact of NaCl and NaHCO₃ salt stress and sampling time on intercellular CO₂ concentration (Ci). Different uppercase letters indicate significant differences among sampling terms within each treatment, while different lowercase letters indicate significant differences among treatments within the same sampling term, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
Figure 8.
Impact of NaCl and NaHCO₃ salt stress and sampling time on intercellular CO₂ concentration (Ci). Different uppercase letters indicate significant differences among sampling terms within each treatment, while different lowercase letters indicate significant differences among treatments within the same sampling term, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
Figure 9.
Soil pH in H2O on the applied NaCl and NaHCO₃ salt solutions. Different lowercase letters indicate significant differences among treatments, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
Figure 9.
Soil pH in H2O on the applied NaCl and NaHCO₃ salt solutions. Different lowercase letters indicate significant differences among treatments, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
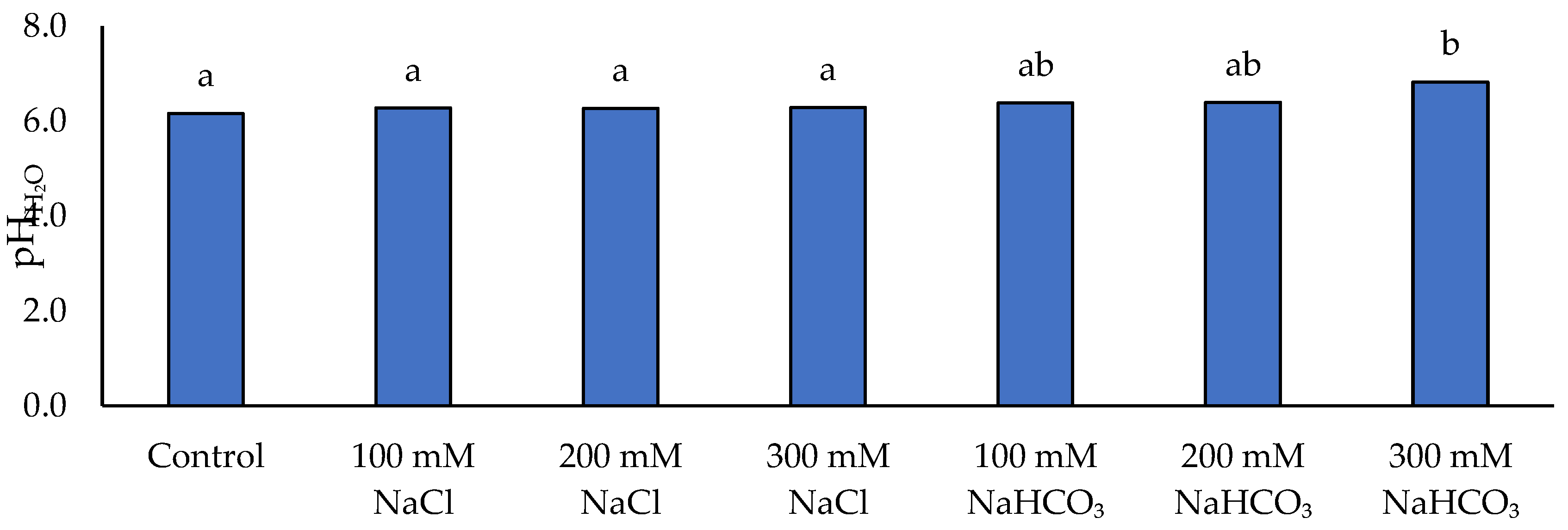
Figure 10.
Soil pH in KCl on the applied NaCl and NaHCO₃ salt solutions. Different lowercase letters indicate significant differences among treatments, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
Figure 10.
Soil pH in KCl on the applied NaCl and NaHCO₃ salt solutions. Different lowercase letters indicate significant differences among treatments, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
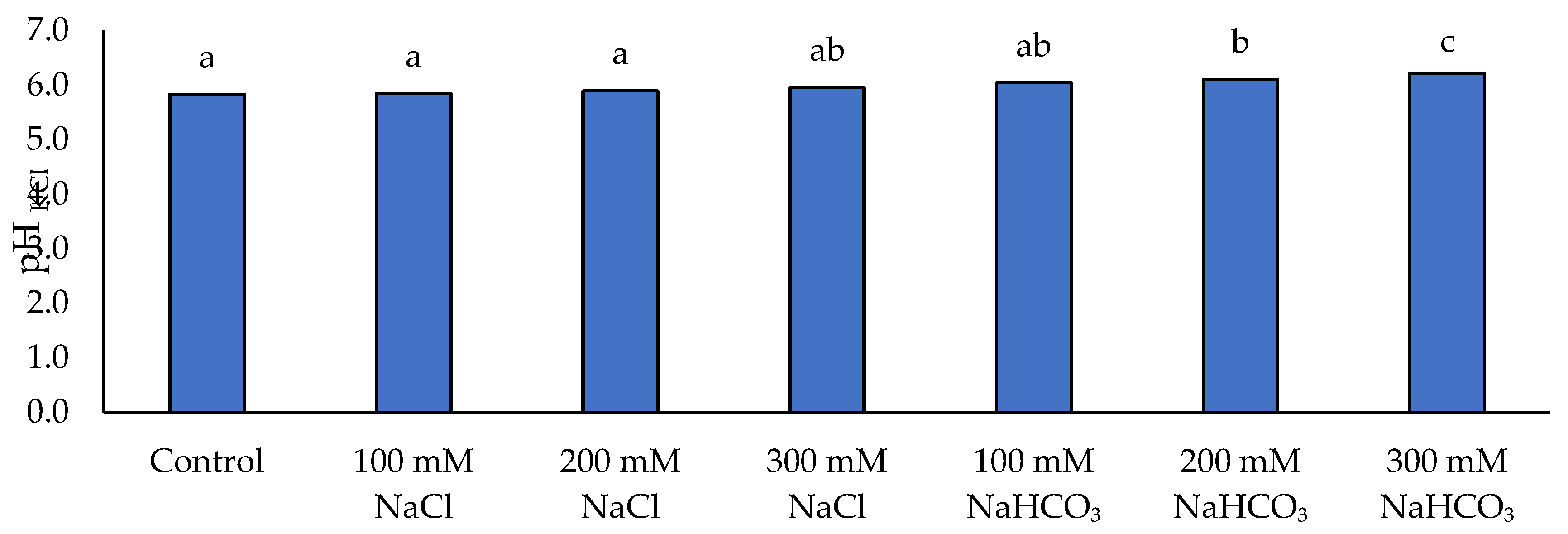
Figure 11.
Soil electrical conductivity depending on the applied NaCl and NaHCO₃ salt solutions. Different lowercase letters indicate significant differences among treatments, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
Figure 11.
Soil electrical conductivity depending on the applied NaCl and NaHCO₃ salt solutions. Different lowercase letters indicate significant differences among treatments, based on ANOVA followed by Tukey’s HSD test (p≤0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



