1. Introduction
Many mushrooms have not only been prized as a valuable food for thousands of years, but also have exceptional medicinal properties and have long been used in traditional medicine [
1]. Throughout history, mushrooms have played an important role in the treatment of diseases affecting the rural populations of Eastern Europe, with
Inonotus obliquus, Fomitopsis officinalis,
Piptoporus betulinus and
Fomes fomentarius being the most commonly used [
2,
3,
4,
5,
6,
7]. These species were used in the treatment of gastrointestinal diseases, various cancers, bronchitis, asthma, night sweats, etc. In addition to the current evaluation of the nutritional value of numerous mushrooms, their pharmacological acceptability and properties are also being investigated. They represent a large, still largely untapped source of new, effective pharmaceutical preparations that are important for modern medicine. Recent studies have demonstrated the antitumor, immunomodulatory, antioxidant, antiviral, antibacterial, antiparasitic, antifungal, detoxifying, hepatoprotective and antidiabetic effects of mushrooms [
8,
9,
10,
11,
12,
13]. In addition to their medicinal effects, mushrooms are also used as novel products such as dietary supplements, functional foods, nutraceuticals, mycopharmaceuticals and designer foods that promote health through the daily consumption of mushrooms [
14]. In addition to their nutritional and health benefits, mushrooms are also a sustainable source of food and high-quality substances. Firstly, they are very efficient at utilizing waste products, and unlike traditional crops, which require large amounts of land, water and other resources to grow, they can be grown in a controlled, contained environment using waste products such as sawdust, straw and agricultural by-products. In addition, the growth cycle of mushrooms is short, so they can be grown all year round, and because they grow in a controlled environment, they are less susceptible to weather-related crop failures and traditional production.
Glycation is a non-enzymatic process of binding sugars to proteins, lipids and nucleic acids that leads to the formation of advanced glycation end products (AGEs) [
15,
16]. These compounds play a key role in the development of numerous chronic diseases, including diabetes, cardiovascular disease, neurodegenerative disorders and accelerated aging. The identification of compounds that can inhibit the formation of AGEs represents an important therapeutic strategy to mitigate the deleterious effects of these compounds [
17]. Such inhibitors offer the potential to prevent or slow the progression of age-related diseases and the debilitating complications associated with diabetes [
18]. In the search for such therapeutic agents, natural products have emerged as particularly promising alternatives to synthetic inhibitors, primarily due to their potentially more favorable safety profile and often multiple mechanisms of action. This focus on natural inhibitors addresses existing concerns regarding the side effects of certain synthetic AGE inhibitors. Therefore, the investigation of mushrooms as a rich source of various natural compounds warrants a thorough exploration of their potential to provide safe and effective AGE inhibitors.
The aim of this work was to summarize the research on the AGE-inhibitory effect of mushrooms and their extracts and to highlight their potential as a rich source of natural compounds capable of inhibiting the formation of AGEs, thus offering therapeutic promise for the treatment of AGEs-related diseases, especially diabetes and age-related diseases.
2. Advanced Glycation End-Products (AGEs) and Inhibition Strategies
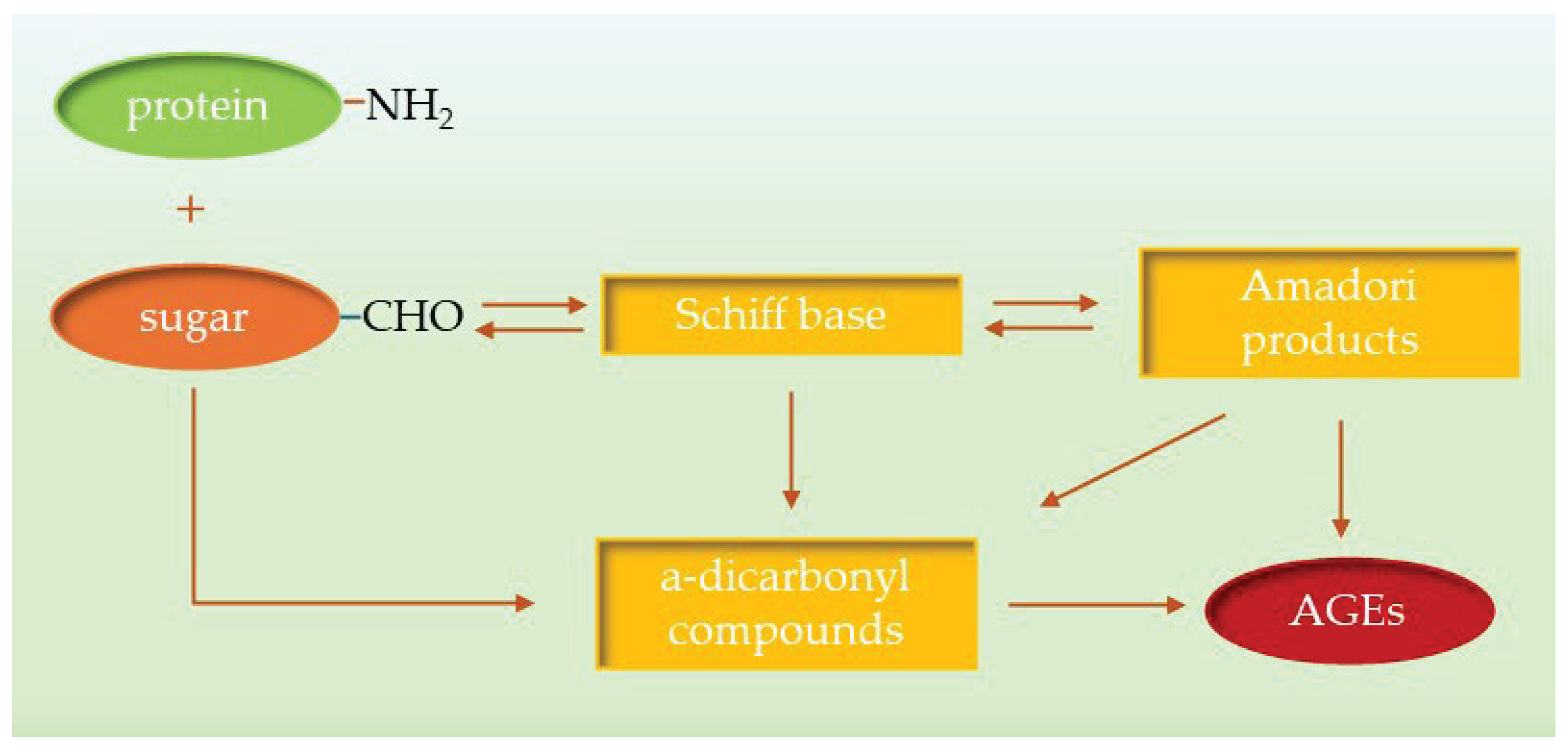
Advanced glycation end products (AGEs) are a heterogeneous group of compounds formed in non-enzymatic reactions between proteins and reducing sugars or lipids [
15,
16]. AGEs accumulate in vivo and activate several signaling pathways that are closely associated with the onset and progression of various diseases and pathological conditions, including diabetes, atherosclerosis, neuropathy, nephropathy, wound healing and Alzheimer’s disease. Decades of research have shown that AGEs are the end product of a process that begins under hyperglycemic conditions with a reaction between sugars and their autoxidation products and amino groups of biomolecules. The process of AGE formation can be divided into two basic steps; in the first, carbonyl groups of sugars react reversibly with free amino groups of proteins and nucleic acids, forming unstable Schiff bases (
Figure 1). Within a few weeks, they are converted intermolecularly into relatively stable Amadori products, which are referred to as early glycation products. In the second step, a small proportion of the Amadori products are converted directly into AGEs by irreversible oxidation or hydrolysis. The remaining Amadori products can be converted into active α- dicarbonyl compounds such as glyoxal (GO), methylglyoxal (MGO) and 3-deoxyglucosone (3-DG) in dehydration, oxidation or cyclization reactions. Reactive α-dicarbonyl compounds, also known as AGE precursors, bind covalently to proteins to form stable AGEs. As a result, AGEs cause various protein modifications such as intra- and intermolecular linkages and fragmentation, leading to a change in their activity, denaturation, and irreversible damage [
19].
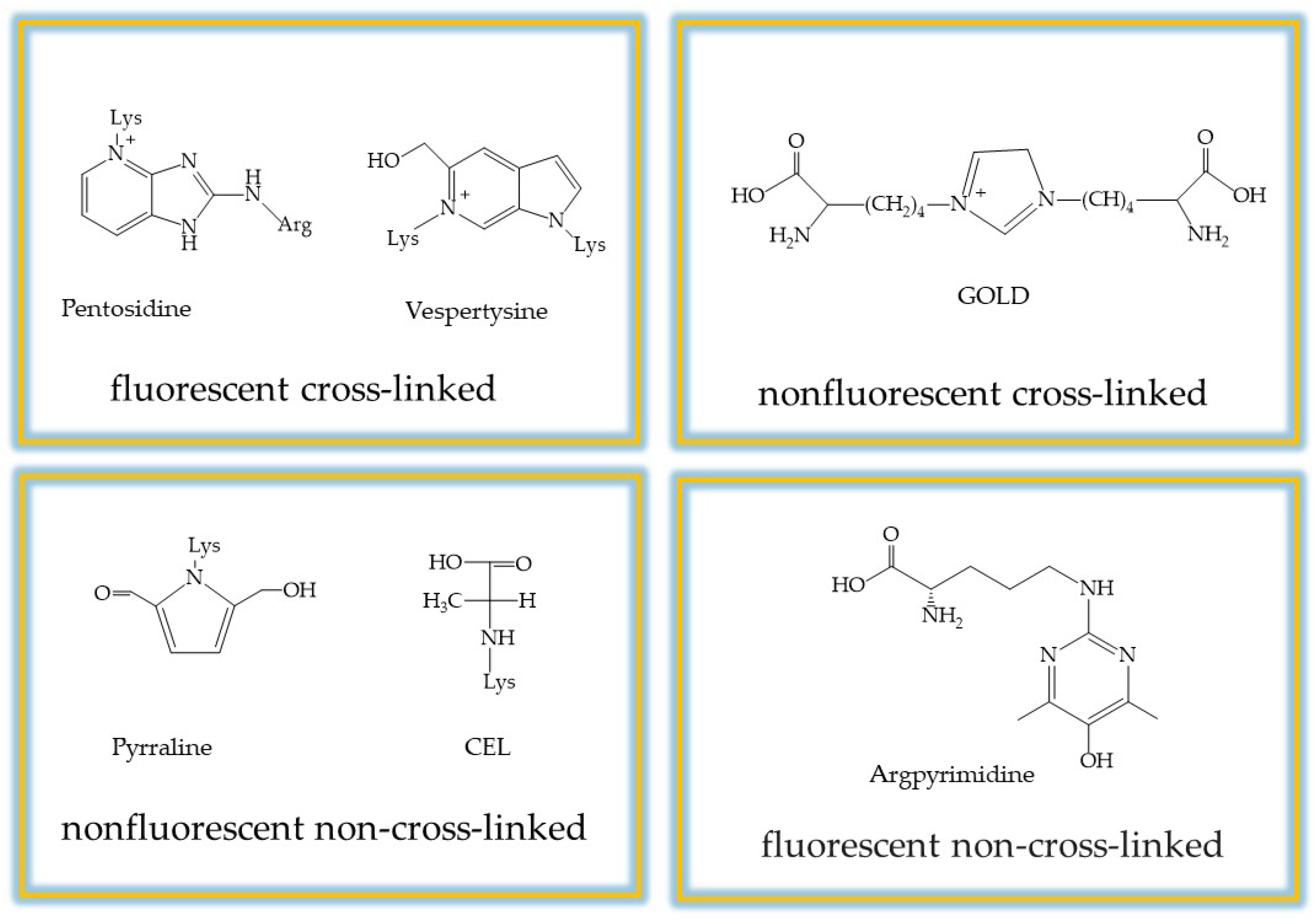
In addition, the autoxidation of glucose, the oxidative cleavage of Schiff bases and lipid peroxidation can produce dicarbonyl compounds that cause the formation of AGEs. Furthermore, glycation has been shown to contribute to oxidative stress as AGEs interact with their receptor (RAGE), leading to increased production of reactive oxygen species (ROS) that accelerate AGE formation. Consequently, under hyperglycemic conditions and in a state of oxidative stress, there is a disruption of the spatial organization of biomolecules, aggregation, and the development of glycation-related diseases. To date, more than 20 types of AGEs have been identified, which are categorized into four groups: (i) non-cross-linked, non-fluorescent; (ii) cross-linked, non-fluorescent; (iii) non-cross-linked, fluorescent; and (iv) cross-linked, fluorescent, which provide a good basis for measuring total AGEs [
20,
21] (
Figure 2).
Several strategies have been developed to inhibit the formation of AGEs, targeting different stages of the glycation process [
22]. One known mechanism is carbonyl scavenging, in which certain compounds scavenge the reactive carbonyl intermediates such as GO, MGO and 3-DG, which are precursors of AGEs [
23]. Examples of synthetic compounds that utilize this mechanism include aminoguanidine and pyridoxamine. Other approaches focus on inhibiting early glycation reactions by inhibiting the initial formation of Schiff bases and Amadori products or by inhibiting the advanced glycation reactions, thereby blocking the conversion of Amadori products into AGEs.
Antioxidant mechanisms also play a critical role in the inhibition of AGEs, as oxidative stress is known to contribute significantly to AGE formation [
21]. Natural antioxidants such as vitamin C and quercetin have demonstrated the ability to prevent AGE formation through their free radical scavenging properties [
24]. Other strategies include masking lysine residues on proteins that are primary sites for glycation, lowering blood glucose levels to reduce the availability of reactive sugars,and chelation of metal ions, as certain metal ions can catalyze the formation of AGEs [
21,
25]. Understanding these various mechanisms is fundamental to classifying how mushroom-derived compounds exert their anti-AGE effects and appreciating their potential therapeutic value.
In addition to inhibiting the formation of AGEs, some compounds known as AGE-breakers can directly break the cross-links that have already formed between proteins due to glycation. Examples of such compounds are Alagebrium (ALT-711) [
26] and its related analogs. Furthermore, in vitro studies have shown that rosmarinic acid has the potential to break existing AGE cross-links [
27]. While the focus of this work is on AGE inhibition, the possibility that certain mushroom derived compounds also have AGE-breaking activity would represent a significant therapeutic advantage, as they could potentially reverse some of the damage already caused by glycation.
The landscape of AGE inhibitors includes both natural and synthetic compounds. Synthetic inhibitors such as aminoguanidine have demonstrated efficacy in preclinical studies, but their clinical use has been limited by significant side effects [
21]. This has led to a growing interest in natural products, including plant extracts and various phytochemicals, as potential sources of safer and equally effective AGE inhibitors with different mechanisms of action [
17]. Well-known examples of natural AGE inhibitors include resveratrol [
28] and curcumin [
29]. Numerous bioactive molecules (
Figure 3) prevent or treat various pathological conditions in humans, and many plant extracts, their fractions or pure components have been thoroughly investigated as AGE inhibitors [
30,
31,
32,
33,
34].
This increasing focus on natural sources emphasizes the need to thoroughly investigate mushrooms, which are known to produce a wide range of bioactive compounds, for their potential to contribute to the development of safe and effective anti-AGE therapies.
3. Mushrooms as a Potential Source of Natural AGE Inhibitors
Mushrooms represent a rich and largely unexploited reservoir of various bioactive metabolites comprising a broad spectrum of chemical classes such as polysaccharides, proteins, peptides, terpenoids, phenolic compounds, alkaloids, vitamins, and minerals [
35]. These compounds are responsible for a broad spectrum of pharmacological activities, including significant antioxidant, anti-inflammatory, antidiabetic, anticancer, and immunomodulatory effects. The sheer diversity of chemical entities present in mushrooms greatly increases the likelihood of discovering novel molecules with potent antiglycation properties. Consequently, the systematic screening of different fungal species and their extracts represents a promising and largely unexplored opportunity for the identification of natural AGE inhibitors.
Numerous research studies have documented the significant antioxidant properties of numerous fungal compounds, particularly polysaccharides and phenolic compounds [
35]. Since oxidative stress is a critical factor in both the formation of AGEs and the mediation of their deleterious effects, the inherent antioxidant capacity of mushrooms strongly suggests their potential to mitigate the formation of AGEs and their downstream consequences. By effectively scavenging free radicals and reducing overall oxidative stress, mushroom-derived antioxidants may indirectly inhibit the glycation process [
22]. This well-established link between antioxidant activity and AGE inhibition provides a compelling rationale for focusing research efforts on mushrooms as a valuable source of anti-AGE compounds.
In addition, many species of mushrooms have long been used in traditional medicine to treat conditions such as diabetes and various age-related ailments [
36]. This ethnomedicinal tradition provides valuable initial evidence of their potential efficacy in the treatment of diseases in which the accumulation of AGEs is an important factor. The traditional use of these mushrooms for related health problems can serve as a guide for modern scientific research by highlighting specific species that are particularly rich in anti-AGE compounds. This intersection of traditional knowledge and modern scientific research offers a promising avenue for the discovery of new therapeutic agents.
Recent research has further expanded the potential of mushrooms by demonstrating that combinations of different mushroom species can exhibit synergistic effects, enhancing their overall anti-glycation and antioxidant capacity beyond what would be expected from the simple sum of their individual activities [
37]. This synergy is particularly potent at lower concentrations of mushroom extracts, highlighting the complex and beneficial interactions between their diverse bioactive compounds, such as phenolics, tannins, and flavonoids. This finding underscores the advantage of consuming a variety of mushrooms or using multi-mushroom formulations to maximize health benefits.
4. Specific Mushroom Species and Their AGE-Inhibiting Properties
Several species of fungi have been investigated for their potential to inhibit the formation of AGEs and promising results have been obtained (
Table 1). The study of
Lignosus rhinocerus (tiger’s milk mushroom) has shown that a medium molecular weight (MMW) fraction obtained from the sclerotia of
Lignosus rhinocerus has significant antiglycation activity [
38]. This fraction was found to specifically inhibit the formation of the major AGEs, namely Nε-(carboxymethyl)lysine (CML) and pentosidine. It is noteworthy that the IC
50 value of this MMW fraction for the inhibition of AGEs is remarkably low at 0.001 mg/ml, almost 520 times lower than that of the positive control, aminoguanidine hydrochloride. The observed anti-glycation activity can be attributed, at least in part, to the potent free radical scavenging activity of superoxide anions of the MMW fraction. Furthermore, the identification of genes encoding glyoxalase I in
L. rhinocerus suggests an additional mechanism by which this fungus may inhibit the formation of AGEs, namely by detoxifying methylglyoxal, an important precursor of various AGEs. These results make
L. rhinocerus a particularly promising source of potent AGE inhibitors with activity against specific AGE types, possibly acting via both antioxidant and carbonyl detoxification pathways.
Polysaccharides isolated from the mycelium of
Ganoderma capense, species that is widely used, especially in Asia, have shown both antiglycation and antiradical activities in in vitro studies [
39]. In particular, the GC70 fraction of these polysaccharides showed significant potential to inhibit AGE formation in a manner that was both time- and dose-dependent. In addition, these polysaccharides showed concentration-dependent scavenging abilities against both DPPH and hydroxyl radicals. The dual activity of
G. capense polysaccharides, namely direct inhibition of AGE formation and free radical scavenging, is particularly beneficial given the intertwining of oxidative stress and glycation in disease processes.
Ganoderma lucidum (reishi mushroom) has yielded an anti-oxidative proteoglycan named FYGL that exhibits significant anti-glycation activity [
40]. This compound inhibits glycation at every stage of the process and suppresses glycoxidation, likely due to its strong antioxidative capacity and its ability to form complexes with target proteins like bovine serum albumin. In vivo studies have confirmed that FYGL can alleviate postprandial hyperglycemia in diabetic mice and reduce AGE accumulation and vascular injury in diabetic rats. FYGL also demonstrates inhibitory effects on α-glucosidase activity, which helps slow carbohydrate metabolism and reduce postprandial hyperglycemia. This dual action against both glycation and glucose absorption makes FYGL a particularly promising compound for diabetes management.
Research indicates that polysaccharide AAP-2S extracted from
Auricularia auricula (wood ear fungus) significantly inhibits the formation of AGEs during both short- and long-term glycosylation processes [
41]. This polysaccharide has been shown to reduce oxidative damage to proteins and preserve important protein sulfhydryl groups. In addition, it reduces the carbonylation of proteins and prevents structural changes in proteins caused by glycation. Furthermore, AAP-2S also exhibits metal chelating capabilities. The mechanism of action may involve modulation of the RAGE/TGF-β/NOX4 pathway, possibly attenuating glycosylation and improving cell fibrosis. In addition, hydrolysates of polysaccharides from
Auricularia auricula-judae also improve glucose absorption in HepG2 cells and extend the lifespan of
Caenorhabditis elegans under conditions of high sugar stress [
42].
Pleurotus ostreatus (oyster mushroom) is known to contain various phenolic compounds that have been reported to have an antiglycative function. A methanolic extract of
Pleurotus ostreatus was found to have high phenolic and flavonoid content and excellent antioxidant capacity [
43]. Fraction F4 of this extract showed strong in vitro inhibition of fructose-mediated hemoglobin as well as BSA protein glycation. In addition, this fraction showed fructosamine and protein carbonylation inhibition and reduced protein aggregation and fluorescence intensity associated with AGEs.
A polyphenol-rich decoction (CPD) of
Inonotus obliquus (Chaga mushroom) has been shown to inhibit glycation of albumin and collagen gel three to four times more effectively than the known AGE inhibitor aminoguanidine [
44]. In addition, CPD exhibits significant intracellular antioxidant effects. Chaga mushroom is known to contain high levels of various antioxidants as well as proteins, minerals, fiber and vitamins [
45].
Polysaccharides obtained from water extracts of
Pholiota nameko, nutritious mushroom originating in Japan, can significantly inhibit the production of AGEs in vitro and increase the reduced cell viability and growth rate caused by glycation stress [
46]. The antiglycation effect originates from their excellent antioxidant activity and ROS scavenging ability.
Extracts from the edible mushroom
Lactarius deterrimus used separately and in combination of chestnut
Castanea sativa extracts inhibited protein glycation and AGE formation in vitro, and reduced non-enzymatic glycosylation in diabetic rats in vivo through inhibition of CML-mediated RAGE/NF-κB activation and enzymatic
O-GlcNAcylation in liver and kidney tissues [
47].
Phellinus linteus has shown strong potential as a natural inhibitor of AGEs due to the presence of various bioactive compounds [
48]. Among its phenolic constituents, one compound from the fruiting body has demonstrated exceptional efficacy in inhibiting protein cross-linking at the final stage of glycation, performing even better than aminoguanidine. Additionally, the mushroom contains potent antioxidants such as hispidin and ergothioneine, which help mitigate oxidative stress—a key driver of AGE formation—and thus indirectly reduce AGE accumulation. Furthermore, polysaccharides extracted from
P. linteus have been found to lower blood glucose levels and improve insulin sensitivity in diabetic models. This glycemic control reduces the availability of sugars that participate in glycation reactions, contributing further to the inhibition of AGE formation [
49].
Ergothioneine is an unusual amino acid derivative found in various mushrooms that has been patented for use as an anti-glycation agent [
50]. This compound acts as a powerful antioxidant and may protect proteins from glycation damage through its free radical scavenging properties. Although the patent focuses on topical application for reducing skin protein glycation, ergothioneine’s mechanism suggests potential benefits for systemic glycation processes as well.
A study by Sun et al. (2015) presents a detailed investigation of the antiglycation properties of two novel polysaccharide fractions (BSP-1b and BSP-2b) purified from the mushroom
Boletus snicus [
51]. The researchers thoroughly characterised the polysaccharides, finding that BSP-2b had a higher molecular weight (128.74 kDa) and a more complex monosaccharide composition, including a significant uronic acid content, compared to BSP-1b (59.21 kDa). In a bovine serum albumin (BSA)-glucose glycation model, both fractions showed significant, dose-dependent inhibition of AGE formation at all stages of the glycation process (initial, intermediate, and final). Notably, BSP-2b consistently exhibited stronger antiglycation activity than BSP-1b. The authors attributed this greater efficacy to the higher molecular weight and increased uronic acid content of BSP-2b, suggesting that these structural features are critical for its potent activity. This study identifies Boletus snicus as a promising source of antiglycative agents and highlights the importance of polysaccharide structure in determining bioactivity.
A crucial aspect for the practical application of mushroom-based AGE inhibitors is their stability during processing. A study on
Agaricus bisporus and
Pleurotus ostreatus demonstrated that UV-B irradiation, a common method to enhance Vitamin D₂ content, significantly reduced the overall antioxidant capacity of the mushrooms. However, the antiglycation activity of both the water-soluble and ethanol-insoluble fractions was fully retained after irradiation [
52]. This finding underscores that the bioactive compounds responsible for inhibiting AGE formation in these commercially vital species are remarkably stable, highlighting their potential for use in processed functional foods and nutraceuticals without losing efficacy.
Table 1.
Antiglycation activities of mushroom species.
Table 1.
Antiglycation activities of mushroom species.
Mushroom
Species |
Active Fraction/
Compound(s) |
Proposed
Mechanism(s) |
Ref. |
| Lignosus rhinocerus |
Medium-molecular-
weight (MMW) fraction |
Inhibition of CML and pentosidine, superoxide anion radical scavenging, glyoxalase I activity |
[38] |
| Ganoderma capense |
Polysaccharides
(GC70 fraction) |
Inhibition of AGE
formation, DPPH and
hydroxyl radical
scavenging |
[39] |
| Ganoderma lucidum |
FYGL proteoglycan |
Inhibits glycation at every stage; suppresses glycoxidation; forms protective complexes with proteins |
[40] |
| Auricularia auricula |
Polysaccharides |
Inhibition of AGE
formation,
attenuation of
oxidative damage,
preservation of
sulfhydryl groups,
metal chelation,
RAGE/TGF-β/NOX4
pathway modulation |
[41,42] |
| Pleurotus ostreatus |
Methanolic extract
(Fraction F4),
phenolic and
flavonoid
compounds |
Inhibition of glycated
hemoglobin formation,
fructosamine, protein
carbonyls, protein
aggregation,
fluorescent AGEs,
antioxidant activity |
[43] |
| Inonotus obliquus |
Polyphenol decoction
(CPD) |
Inhibition of albumin
and collagen gel
glycation,
intracellular
antioxidant activity |
[44] |
| Pholiota nameko |
Polysaccharides |
Antioxidant activity and ROS scavenging ability |
[46] |
| Lactarius deterrimus |
50% ethanolic
extract |
Reduction of the RAGE/NF-κB activation, reduction of enzymatic O-GlcNAcylation, reduction of oxidative stress |
[47] |
| Phellinus linteus |
Phenolic compounds, polysaccharides,
ergothioneine |
Carbonyl trapping, free radical scavenging, metabolic regulation |
[48,49] |
| Various mushrooms |
Ergothioneine |
Antioxidant activity; free radical scavenging |
[50] |
| Boletus snicus |
Polysaccharides
(BSP-2b fraction) |
Activity correlated with high molecular weight and high uronic acid content |
[51] |
| Agaricus bisporus, Pleurotus ostreatus |
Water-soluble and ethanol-insoluble fractions |
Retention of activity post-UV irradiation suggests the involvement of stable, non-phenolic compounds |
[52] |
Beyond the activity of individual species, the strategic combination of common edible mushrooms has emerged as a promising approach to boost anti-glycation effects. A comprehensive study evaluating pairwise combinations of
Agaricus bisporus,
Lentinula edodes,
Flammulina velutipes,
Pleurotus ostreatus, and
Pleurotus eryngii revealed that specific mixtures can produce synergistic interactions [
37]. Interestingly, the study also found that these synergistic effects were most pronounced at lower extract concentrations, while higher concentrations often led to antagonistic interactions. This suggests that the balance and ratio of bioactive compounds are critical for achieving optimal functional benefits. The anti-glycation activity of these mushroom combinations showed a significant positive correlation with their total phenolic content and radical scavenging capacity (DPPH assay), reinforcing the role of polyphenols as primary agents against AGE formation.
5. Mechanisms of Action of Mushroom-Derived AGE Inhibitors
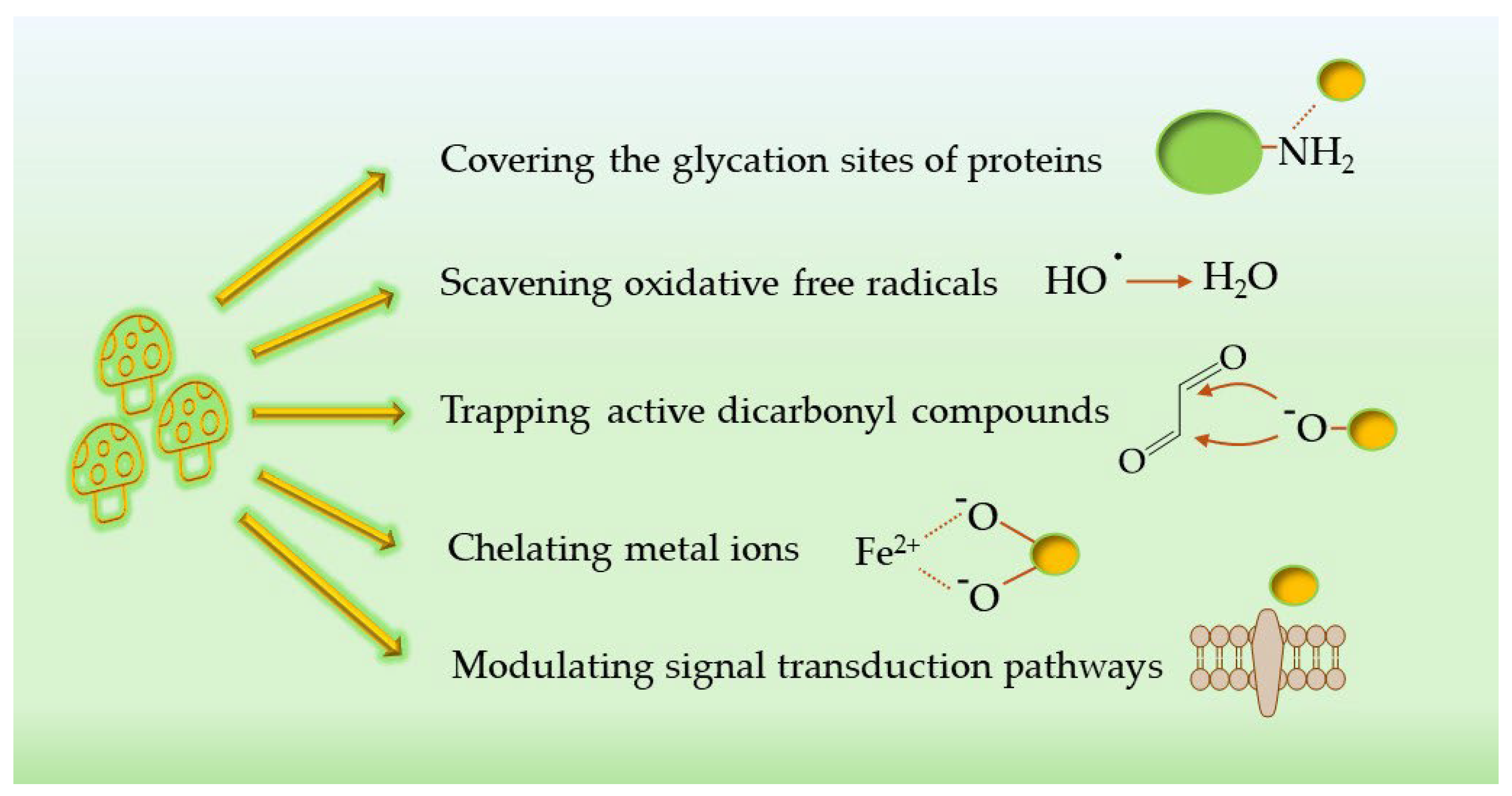
The fungal species identified in the previous section exert their AGE-inhibitory effects through a variety of mechanisms that reflect the different chemical nature of their bioactive compounds (
Figure 4).
5.1. Antioxidant Activity
A primary mechanism by which mushrooms combat AGE formation is their potent antioxidant activity. Since oxidative stress contributes to both the formation and deleterious effects of AGEs, the ability of mushroom-derived compounds to scavenge free radicals and reduce oxidative stress is critical. For example, the antiglycation activity of the MMW fraction from
Lignosus rhinocerus is associated with its strong ability to scavenge superoxide anion radicals [
38]. Similarly, the polysaccharides from
Ganoderma capense show scavenging abilities against both DPPH and hydroxyl radicals [
39]. Antiglycation activity of
Pholiota nameko is also attributed to antioxidant activity of constitutive polysaccharides [
46]. The high phenolic and flavonoid content in
Pleurotus ostreatus contributes to its excellent antioxidant capacity, which is likely involved in its anti-glycation effect [
43]. The polyphenol decoction from
Inonotus obliquus also shows significant intracellular antioxidant activity, which contributes to its ability to inhibit glycation [
44].
5.2. Scavenging of Carbonyl
Another important mechanism is the scavenging of reactive carbonyl species, which are important intermediates in the formation of AGEs. While direct evidence for scavenging of carbonyl species by specific fungal compounds is limited, the presence of glyoxalase I-related genes in
Lignosus rhinocerus suggests an ability to detoxify methylglyoxal, a major reactive carbonyl compound [
38]. This enzymatic detoxification effectively reduces the pool of reactive carbonyl compounds available for the formation of AGEs.
5.3. Interfering with the Glycation Process
Compounds derived from fungi can also interfere with the glycation process at various stages. Degraded polysaccharides from
Auricularia auricula inhibit AGE formation in both the early and advanced stages of glycation [
41]. The methanolic extract of
Pleurotus ostreatus inhibits the formation of glycated hemoglobin (an early glycation product), fructosamine (an intermediate product), and protein carbonyl (associated with advanced glycation) [
43]. The polyphenol decoction from
Inonotus obliquus effectively inhibits the glycation of albumin and collagen, suggesting an intervention in a fundamental level of protein modification by sugars [
44]. The proteoglycan FYGL isolated from
Ganoderma lucidum has been shown to inhibit glycation at multiple stages and suppress glycoxidation, possibly through formation of complexes with target proteins that protect vulnerable amino acid residues [
40]. These compounds can prevent the initial attachment of reducing sugars to proteins or interrupt subsequent rearrangement reactions that lead to AGE formation.
5.4. Modulation of Signal Transduction Pathways
Some mushroom-derived compounds may exert their anti-AGE effects by modulating relevant cellular signaling pathways. For example,
Auricularia auricula polysaccharides, can attenuate glycosylation and improve cell fibrosis by modulating the RAGE/TGF-β/NOX4 signaling pathway [
41]. Similarly,
Lactarius deterrimus extracts disrupt the cycle of CML-induced RAGE/NF-κB pathway activation by reducing protein glycation and CML formation. The RAGE signaling pathway is a key mediator of the inflammatory and deleterious effects of AGEs, and its modulation may represent an important therapeutic strategy.
5.5. Metal-Ion Chelation
The ability of
Auricularia auricula polysaccharides to chelate metal ions [
41] may also contribute to their anti-AGE activity, as certain metal ions can catalyze the glycation process and the formation of reactive oxygen species involved in AGE production. By sequestering these metal ions, the polysaccharides may help to reduce the rate of AGE formation.
5.6. Synergistic Interactions
The beneficial effects of mushroom-derived compounds can be amplified through synergistic interactions when different species are combined. These interactions are not merely additive; the combined effect exceeds the sum of individual effects, likely due to complementary mechanisms of action [
37]. For instance, the potent synergy observed in mixtures containing
Flammulina velutipes (rich in phenolics) with
Agaricus bisporus or
Lentinula edodes may arise from the interplay between different phenolic profiles, flavonoids, and tannins. Such combinations can enhance free radical scavenging, improve metal chelation, and more effectively interfere at multiple stages of the glycation pathway simultaneously. This multi-targeted approach, facilitated by synergy, represents a powerful strategy for inhibiting AGE formation.
5.7. The Critical Role of Molecular Weight and Uronic Acid Content
A comparative study of two polysaccharide fractions from Boletus snicus (BSP-1b and BSP-2b) shows a clear correlation between structural features and antiglycation efficacy. The authors demonstrated that the polysaccharide fraction with higher molecular weight and higher uronic acid content (BSP-2b) exhibited significantly stronger inhibitory activity at all stages of glycation compared to the fraction with lower molecular weight and less uronic acid (BSP-1b) [
51]. BSP-2b was more effective at inhibiting the formation of Amadori products in the initial stage, scavenging reactive dicarbonyl compounds in the intermediate stage, and preventing the formation of fluorescent AGEs in the final stage. Although the authors noted that the precise mechanistic details require further investigation, the results strongly suggest that molecular weight and, most notably, uronic acid content are critical structural features that directly enhance a polysaccharide’s ability to inhibit AGE formation. This direct comparison within the same species highlights that the antiglycation activity of polysaccharides is not generic but is highly dependent on these specific structural characteristics.
5.8. Role of Stable, High-Molecular-Weight Compounds
The resilience of antiglycation compounds to processing stresses provides valuable mechanistic insights. Research on UV-irradiated
Agaricus bisporus and
Pleurotus ostreatus showed that their antiglycation activity remained intact despite a reduction in general antioxidant power [
52]. This stability indicates that the primary antiglycative agents are likely high-molecular-weight polysaccharides or glycoproteins, which operate through robust mechanisms such as carbonyl trapping and metal chelation. These mechanisms are not easily compromised by photo-oxidation, unlike the activity of more labile, low-molecular-weight phenolic antioxidants. The stability of these compounds further confirms their suitability for development into reliable therapeutic formulations.
6. Evidence from Research Studies
The antiglycation properties of the identified fungal species are supported by various in vitro and in vivo studies. In vitro studies, usually performed in the laboratory, allow direct evaluation of the ability of mushroom extracts or isolated compounds to inhibit AGE formation under controlled conditions.
For example, the potent antiglycation activity of the MMW fraction
from Lignosus rhinocerus was demonstrated in a human serum albumin-glucose system, where it significantly inhibited the formation of CML and pentosidine [
38]. Similarly, the polysaccharides of
Ganoderma capense showed significant anti-glycation activity in vitro using non-enzymatic glycation reaction assays [
39]. The degraded polysaccharides of
Auricularia auricula judae were also effective in inhibiting AGE formation in vitro in both short-term and long-term glycosylation experiments [
42]. Furthermore, the methanolic extract of
Pleurotus ostreatus and its fraction F4 showed significant in vitro inhibitory effects on the formation of glycated hemoglobin, fructosamine and protein carbonyls [
43]. The polyphenol decoction of Chaga mushroom also showed strong in vitro antiglycation activity against albumin and collagen [
44]. Furthermore, the study on
Boletus snicus demonstrated that its purified polysaccharide fraction BSP-2b effectively inhibited AGE formation across all three stages of glycation (initial, intermediate, and final) in a BSA-glucose model [
51]. Similarly, research on UV-irradiated
Agaricus bisporus and
Pleurotus ostreatus confirmed that their antiglycation activity in the BSA-fructose system remained stable despite processing, highlighting the robustness of the active compounds [
52].
Several in vivo studies have demonstrated the potential of mushrooms to inhibit the formation and accumulation of AGEs, particularly in diabetic animal models. For instance,
Ganoderma lucidum has been shown to significantly reduce AGE accumulation in diabetic rats through the action of a proteoglycan called FYGL [
40]. This compound not only inhibited glycation at various stages but also alleviated vascular injury and postprandial hyperglycemia. Its mechanism involves strong antioxidant activity and the inhibition of α-glucosidase, which helps regulate blood glucose levels and reduces the substrate availability for glycation. Similarly, polysaccharides extracted from
Auricularia auricula have shown protective effects in vivo by reducing oxidative protein damage and preserving essential protein structures in the context of high sugar stress. These effects were also associated with the modulation of the RAGE/TGF-β/NOX4 signaling pathway, suggesting a molecular basis for the observed antiglycation activity [
41]. Another in vivo study involving
Lactarius deterrimus, particularly in combination with chestnut (
Castanea sativa) extracts, demonstrated that these mushrooms could reduce protein glycation and AGE formation in diabetic rats [
47]. The treatment inhibited the activation of the RAGE/NF-κB inflammatory pathway and also affected enzymatic O-GlcNAcylation in the liver and kidneys, processes that are typically dysregulated in diabetes.
Collectively, these studies provide compelling in vivo evidence that mushroom-derived compounds can effectively inhibit AGE formation and its associated pathologies through antioxidant effects, improved glucose metabolism, and suppression of glycation-related signaling pathways. Nevertheless, further research specifically designed to evaluate the AGE inhibitory effects of these fungal species and their isolated compounds in vivo is warranted to fully validate their therapeutic potential.
7. Potential Applications and Future Directions
The findings presented in this review highlight the significant potential of several fungal species as a source of natural compounds that can inhibit the formation of AGEs. Given the known role of AGEs in the development of numerous chronic diseases, including diabetes, cardiovascular disease and neurodegenerative disorders, these mushroom-derived inhibitors hold promise for a variety of therapeutic applications. One of the most immediate applications is in the treatment of diabetes and its complications. The accelerated formation of AGEs in diabetic patients contributes significantly to the development of micro- and macrovascular complications. Mushroom extracts and isolated compounds, particularly from Lignosus rhinocerus, Ganoderma capense and Auricularia auricula, which have shown potent antiglycation activity in vitro, could be explored as complementary therapies to conventional antidiabetic treatments. Their ability to inhibit the formation of specific AGEs such as CML and pentosidine, together with their antioxidant properties, could help to reduce the burden of diabetic complications.
In addition, the potential of fungal compounds in attenuating
age-related diseases is worth mentioning. The accumulation of AGE is a hallmark of the aging process and contributes to tissue damage and loss of function. The antioxidant and antiglycative properties of mushrooms such as
Pleurotus ostreatus and
Inonotus obliquus suggest that they may slow down the aging process and prevent or delay the onset of age-related disorders. The neuroprotective potential of some of these mushrooms, which has been pointed out in other studies [
53], supports their investigation in the context of neurodegenerative diseases in which AGEs play a role.
The possibility of incorporating mushroom extracts or isolated compounds into functional foods and nutraceuticals is also worth considering. The relatively safe profile generally associated with edible and medicinal mushrooms makes them attractive candidates for dietary interventions aimed at preventing the accumulation of AGEs and promoting overall health. For example, the degraded polysaccharides from Auricularia auricula, which have shown promising anti-AGE effects, may be developed as food additives. Also, the discovery of synergistic anti-glycation effects in mushroom combinations opens exciting new avenues for developing functional foods and nutraceuticals. Instead of relying on single-species extracts, future products could be formulated using optimized blends of mushrooms to achieve greater efficacy at lower doses. This approach aligns with dietary patterns that emphasize diversity and could lead to more potent preventive strategies. Furthermore, the stability of antiglycation compounds in mushrooms like Agaricus bisporus and Pleurotus ostreatus during UV processing, combined with the potent activity of polysaccharides from species like Boletus snicus, makes them particularly suitable candidates for the development of standardized nutraceuticals and processed functional foods.
Future research should focus on several key areas to further explore the therapeutic potential of mushroom-derived AGE inhibitors. Isolation and characterization of the specific bioactive compounds responsible for the observed anti-glycation activity are crucial. Once identified, their exact mechanisms of action need to be elucidated at the molecular level. This includes the question of whether they act as carbonyl scavengers, intervene in certain phases of glycation, modulate signaling pathways or have AGE-breaking capabilities.
Preclinical studies using in vivo models of diabetes, aging and other AGE-related diseases are essential to verify the efficacy and safety of these compounds. Such studies should evaluate the effects of mushroom extracts or isolated compounds on AGE levels in various tissues and organs and their ability to prevent or ameliorate disease progression.
Finally, human clinical trials are needed to translate the promising preclinical results into effective therapeutic interventions. These trials should demonstrate the safety and efficacy of fungal AGE inhibitors in the treatment of AGE-related diseases and to improve patient outcomes. Exploring potential synergistic effects by combining different mushroom extracts or compounds with conventional therapies could also be a valuable avenue for future research.
8. Conclusions
Analysis of the available research shows that mushrooms are a promising source of natural compounds that can inhibit the formation of advanced glycation end products (AGEs). Several species of fungi, including Lignosus rhinocerus, Ganoderma capense, Auricularia auricula, Pleurotus ostreatus, Boletus snicus and Inonotus obliquus, have shown significant anti-glycation activity in vitro, often associated with their high content of polysaccharides and phenolic compounds with potent antioxidant properties. Proposed mechanisms of action include scavenging reactive oxygen species and reactive carbonyl intermediates, interfering with different phases of the glycation process, and modulating relevant cellular signaling pathways. While in vitro studies provide strong evidence of their potential, further research, especially in vivo and clinical studies, are needed to fully validate their therapeutic efficacy in the treatment of AGE-related diseases.
Nonetheless, the results highlight the potential of fungi as a valuable resource for the development of safe and effective natural AGE inhibitors, offering a promising avenue for the prevention and treatment of a wide range of chronic diseases associated with aging and metabolic disorders.
Author Contributions
Conceptualization, M.Č.S.; methodology, M.Č.S.; investigation, M.Č.S., M.K. and F.Š.; resources, M.Č.S., M.K. and F.Š.; data curation, M.Č.S., M.K. and F.Š.; writing—original draft preparation, M.Č.S., M.K. and F.Š.; writing—review and editing, M.Č.S., M.K. and F.Š.; visualization, M.Č.S.; supervision, M.Č.S.; project administration, M.Č.S.; funding acquisition, M.Č.S. All authors have read and agreed to the published version of the manuscript.
Funding
This paper has been funded by the European Union (NextGenerationEU) under the National Recovery and Resilience Plan 2021–2026 (NRRP), through the UNIZG FFTB institutional project “Evaluation of alternative sources of bioactive compounds and proteins (BioAlter)”, approved by the Ministry of Science, Education and Youth of the Republic of Croatia (component C3.2, source 581).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing does not apply to this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wani, B.A.; Bodha, R.H.; Wani, A.H. Nutritional and medicinal importance of mushrooms. J. Med. Plant. Res. 2010, 4, 2598–2604. [Google Scholar] [CrossRef]
- Szychowski, K.A.; Skóra, B.; Pomianek, T.; Gmiński. J. Inonotus obliquus – from folk medicine to clinical use. J. Tradit. Complement. Med. 2020, 11, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Elkhateeb, W.; Daba, G.; Elnahas, M.; Thomas, P.W. Fomitopsis officinalis mushroom ancient gold mine of functional components and biological activities for modern medicine. Egypt. Pharm. J. 2019, 18, 285–289. [Google Scholar] [CrossRef]
- Fijałkowska, A.; Muszyńska, B.; Sułkowska-Ziaja, K; Kała, K.; Pawlik, A.; Stefaniuk, D.; Matuszewska, A.; Piska, K.; Pękala, E.; Kaczmarczyk, P.; Piętka, J.; Jaszek, M. Medicinal potential of mycelium and fruiting bodies of an arboreal mushroom Fomitopsis officinalis in therapy of lifestyle diseases. Sci. Rep. 2020, 10, 20081. [Google Scholar] [CrossRef]
- Papp, N.; Rudolf, K.; Bencsik, T.; Czégényi, D. Ethnomycological use of Fomes fomentarius (L.) Fr. and Piptoporus betulinus (Bull.) P. Karst. in Transylvania, Romania. Genet. Resour. Crop. Evol. 2017, 64, 101–111. [Google Scholar] [CrossRef]
- Rašeta, M.; Kebert, M.; Pintać Šarac, D.; Mišković, J.; Berežni, S.; Kulmány, Á.E.; Zupkó, I.; Karaman, M.; Jovanović-Šanta, S. Bioactive Potential of Balkan Fomes fomentarius Strains: Novel Insights into Comparative Mycochemical Composition and Antioxidant, Anti-Acetylcholinesterase, and Antiproliferative Activities. Microorganisms 2025, 13, 1210. [Google Scholar] [CrossRef]
- Ulrike, G.; Zöll, M.; Peintner, U.; Rollinger, J.M. European medicinal polypores – A modern view on traditional uses. J. Ethnopharmaco. 2014, 154, 564–583. [Google Scholar] [CrossRef]
- Gao, Y.; Zhou, S.; Chen, G.; Dai, X; Ye, J. A. Phase I/II study of a Ganoderma lucidum extract (Ganopofy) in patients with advanced cancer. Int. J. Med. Mushrooms. 2002, 4, 207–214. [Google Scholar] [CrossRef]
- Gao, Y.; Zhou, Sh.; Huang, M.; Xu, A. Antibacterial and antiviral value of the genus Ganoderma P. Karst. species (Aphyllophoromycetideae): a review. Int. J. Med. Mushrooms. 2003, 5, 235–246. [Google Scholar] [CrossRef]
- Gao, Y.; Lan, J.; Dai, X.; Ye, J.; Zhou, Sh. A phase I/II study of Ling Zhi mushroom Ganoderma lucidum (W.Curt.:Fr.) Lloyd (Aphyllophoromycetideae) extract in patients with type II diabetes mellitus. Int. J. Med. Mushrooms. 2004, 6, 96–107. [Google Scholar] [CrossRef]
- Didukh, M.Y.; Wasser, S.P.; Nevo, E. Medicinal value of species of the family Agaricaceae Cohn (higher Basidiomycetes) current stage of knowledge and future perspectives. Int. J. Med. Mushrooms. 2003, 5, 133–152. [Google Scholar] [CrossRef]
- Rowan, N.J.; Smith, J.E.; Sullivan, R. Immunomodulatory activities of mushroom glucans and polysaccharide–protein complexes in animals and humans (a review). Int. J. Med. Mushrooms. 2003, 5, 95–110. [Google Scholar] [CrossRef]
- Wasser, S.P. Medicinal mushroom science: history, current status, future trends, and unsolved problems. Int. J. Med. Mushrooms. 2010, 12, 1–16. [Google Scholar] [CrossRef]
- Cheung, P.C.K. Regulatory issues of mushrooms as functional foods and dietary supplements: safety and efficacy. In Mushrooms as functional foods; Cheung, P.C.K. Wiley: New York, 2008; pp. 199–221. [Google Scholar]
- Uceda, A.B.; Mariño, L.; Casasnovas, R.; Adrover, M. An overview on glycation: molecular mechanisms, impact on proteins, pathogenesis, and inhibition. Biophys. Rev. 2024, 16, 189–218. [Google Scholar] [CrossRef]
- Velichkova, S.; Foubert, K.; Pieters, L. Natural Products as a Source of Inspiration for Novel Inhibitors of Advanced Glycation Endproducts (AGEs) Formation. Planta Med. 2021, 87, 780–801. [Google Scholar] [CrossRef]
- Peng, X.; Ma, J.; Chen, F.; Wang, M. Naturally occurring inhibitors against the formation of advanced glycation end-products. Food Funct. 2011, 2, 289–301. [Google Scholar] [CrossRef]
- Reddy, V.P.; Aryal, P.; Darkwah, E.K. Advanced Glycation End Products in Health and Disease. Microorganisms. 2022, 10, 1848. [Google Scholar] [CrossRef]
- Lapolla, A.; Traldi, P.; Fedele, D. Importance of measuring products of non-enzymatic glycation of proteins. Clin. Biochem. 2005, 38, 103–115. [Google Scholar] [CrossRef]
- Twarda-Clapa, A.; Olczak, A.; Białkowska, A.M.; Koziołkiewicz, M. Advanced Glycation End-Products (AGEs): Formation, Chemistry, Classification, Receptors, and Diseases Related to AGEs. Cells. 2022, 11, 1312. [Google Scholar] [CrossRef]
- Perrone, A.; Giovino, A.; Benny, J.; Martinelli, F. Advanced Glycation End Products (AGEs): Biochemistry, Signaling, Analytical Methods, and Epigenetic Effects. Oxid. Med. Cell Longev. 2020, 18, 3818196. [Google Scholar] [CrossRef]
- Chen, N.; Wang, N.; Fang, Q.; Yu, Z.; Hu, Y.; Jin, J.; Yang, S. Inhibition effect of AGEs formation in vitro by the two novel peptides EDYGA and DLLCIC derived from Pelodiscus sinensis. Front. Nutr. 2025, 12, 1537338. [Google Scholar] [CrossRef] [PubMed]
- Chinchansure, A.A.; Korwar, A.M.; Kulkarni, M.J.; Joshi, S.P. Recent development of plant products with anti-glycation activity: a review. RSC Adv. 2015, 5, 31113–31138. [Google Scholar] [CrossRef]
- Chu, J.; Lin, S.; Yuan, Y.; Zhang, S.; Zhang, S. Effects of quercetin and l-ascorbic acid on heterocyclic amines and advanced glycation end products production in roasted eel and lipid-mediated inhibition mechanism analysis. Food Chem. 2024, 441, 138394. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Liu, J.; Dong, L.; Wang, X.; Zhang, X. Novel advances in inhibiting advanced glycation end product formation using natural compounds. Biomed. Pharmacother. 2021, 140, 111750. [Google Scholar] [CrossRef]
- Bakris, G.L.; Bank, A.J.; Kass, D.A.; Neutel, J.M.; Preston, R.A.; Oparil, S. Advanced glycation end-product cross-link breakers. A novel approach to cardiovascular pathologies related to the aging process. Am. J. Hypertens. 2004, 17, 23S–30S. [Google Scholar] [CrossRef]
- Jean, D.; Pouligon, M.; Dalle, C. Evaluation in vitro of AGE-crosslinks breaking ability of rosmarinic acid. Glycative Stress Research 2015, 2, 204–207. [Google Scholar] [CrossRef]
- Mizutani, K.; Ikeda, K.; Yamori, Y. Resveratrol Inhibits AGEs-Induced Proliferation and Collagen Synthesis Activity in Vascular Smooth Muscle Cells from Stroke-Prone Spontaneously Hypertensive Rats. Biochem. Biophys. Res. Comm. 2000, 274, 61–67. [Google Scholar] [CrossRef]
- Tang, Y.; Chen, A. Curcumin eliminates the effect of advanced glycation end-products (AGEs) on the divergent regulation of gene expression of receptors of AGEs by interrupting leptin signaling. Lab. Invest. 2014, 94, 503–516. [Google Scholar] [CrossRef]
- Qais, F.A.; Alam, Md.M.; Naseemb, I.; Ahmad, I. Understanding the mechanism of non-enzymatic glycation inhibition by cinnamic acid: an in vitro interaction and molecular modelling study. RSC Adv. 2016, 6, 65322–65337. [Google Scholar] [CrossRef]
- Selvakumar, G.; Venu, D.; Kuttalam, I.; Lonchin, S. Inhibition of Advanced Glycation End Product Formation in Rat Tail Tendons by Polydatin and p-Coumaric acid: an In Vitro Study. Appl. Biochem. Biotechnol. 2022, 194, 339–353. [Google Scholar] [CrossRef]
- Alam, M.M.; Ahmad, I.; Naseem, I. Inhibitory effect of quercetin in the formation of advance glycation end products of human serum albumin: An in vitro and molecular interaction study. Int. J. Biol. Macromol. 2015, 79, 336–343. [Google Scholar] [CrossRef]
- Ou, J.; Huang, J.; Wang, M.; Ou, S. Effect of rosmarinic acid and carnosic acid on AGEs formation in vitro. Food Chem. 2017, 221, 1057–1061. [Google Scholar] [CrossRef] [PubMed]
- Rashedinia, M.; Rasti Arbabi, Z.; Sabet, R.; Emami, L.; Poustforoosh, A.; Sabahi, Z. Comparison of Protective Effects of Phenolic Acids on Protein Glycation of BSA Supported by In Vitro and Docking Studies. Biochem. Res. Int. 2023, 2023, 9984618. [Google Scholar] [CrossRef] [PubMed]
- Chugh, R.M.; Mittal, P.; Mp, N.; Arora, T.; Bhattacharya, T.; Chopra, H.; Cavalu, S.; Gautam, R.K. Fungal Mushrooms: A Natural Compound with Therapeutic Applications. Front. Pharmacol. 2022, 13, 925387. [Google Scholar] [CrossRef] [PubMed]
- Bhambri, A.; Srivastava, M.; Mahale, V.G.; Mahale, S.; Karn, S.K. Mushrooms as Potential Sources of Active Metabolites and Medicines. Front. Microbiol. 2022, 13, 837266. [Google Scholar] [CrossRef]
- Tan, K.W.K.; Yong, P.H.; Ng, Z.X. Synergistic anti-glycation and antioxidant interaction among different mushroom extract combinations. Foods Raw Mater. 2026, 14, 300–320. [Google Scholar] [CrossRef]
- Yap, H.Y.; Tan, N.H.; Ng, S.T.; Tan, C.S.; Fung, S.Y. Inhibition of Protein Glycation by Tiger Milk Mushroom [Lignosus rhinocerus (Cooke) Ryvarden] and Search for Potential Anti-diabetic Activity-Related Metabolic Pathways by Genomic and Transcriptomic Data Mining. Front. Pharmacol. 2018, 9, 103. [Google Scholar] [CrossRef]
- Yan, C.; Kong, F.; Zhang, D.; Cui, J. Anti-glycated and antiradical activities in vitro of polysaccharides from Ganoderma capense. Pharmacogn. Mag. 2013, 33, 23–27. [Google Scholar] [CrossRef]
- Zhang, Y.; Pan, Y.; Li, J.; Zhang, Z.; He, Y.; Yang, H.; Zhou, P. Inhibition on α-Glucosidase Activity and Non-Enzymatic Glycation by an Anti-Oxidative Proteoglycan from Ganoderma lucidum. Molecules 2022, 27, 1457. [Google Scholar] [CrossRef]
- Gong, P.; Pei, S.; Long, H.; Yang, W.; Yao, W.; Li, N.; Wang, J.; Zhao, Y.; Chen, F.; Xie, J.; Guo, Y. Potential inhibitory effect of Auricularia auricula polysaccharide on advanced glycation end-products (AGEs). Int. J. Biol. Macromol. 2024, 262, 129856. [Google Scholar] [CrossRef]
- Shen, M.; Fang, Z.; Chen, Y.; Chen, Y.; Xiao, B.; Guo, L.; Xu, Y.; Wang, G.; Wang, W.; Zhang, Y. Hypoglycemic Effect of the Degraded Polysaccharides from the Wood Ear Medicinal Mushroom Auricularia auricula-judae (Agaricomycetes). Int. J. Med. Mushrooms. 2019, 21, 1033–1042. [Google Scholar] [CrossRef]
- Macwan, D.; Patel, H.V. Inhibitory Effect of Bioassay-Guided Fractionation of Mushroom (Pleurotus ostreatus) Extract on Fructose-induced Glycated Hemoglobin and Aggregation in vitro. Pharmacogn. Res. 2023, 15, 776–784. [Google Scholar] [CrossRef]
- Doi, N.; Araki, K.; Fukuta, Y.; Kuwagaito, Y.; Yamauchi, Y.; Sasai, Y.; Kondo, S-i.; Kuzuya, M. Anti-glycation and antioxidant effects of Chaga mushroom decoction extracted with a fermentation medium. Food Sci. Tech. Res. 2023, 29, 155–161. [Google Scholar] [CrossRef]
- Fordjour, E.; Manful, C.F.; Javed, R.; Galagedara, L.W.; Cuss, C.W.; Cheema, M.; Thomas, R. Chaga mushroom: a super-fungus with countless facets and untapped potential. Front. Pharmacol. 2023, 14, 1273786. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Lin, T.-Y.; Lin, J.-A.; Cheng, K.-C.; Santoso, S.P.; Chou, C.-H.; Hsieh, C.-W. Effect of Pholiota nameko Polysaccharides Inhibiting Methylglyoxal-Induced Glycation Damage In Vitro. Antioxidants. 2021, 10, 1589. [Google Scholar] [CrossRef]
- Jovanović, J.A.; Mihailović, M.; Uskoković, A.S.; Grdović, N.; Dinić, S.; Poznanović, G.; Mujić, I.; Vidaković, M. Evaluation of the Antioxidant and Antiglycation Effects of Lactarius deterrimus and Castanea sativa Extracts on Hepatorenal Injury in Streptozotocin-Induced Diabetic Rats. Front. Pharmacol. 2017, 8, 793. [Google Scholar] [CrossRef]
- Lee, Y.S.; Kang, Y.H.; Jung, J.Y.; Lee, S.; Ohuchi, K.; Shin, K.H.; Kang, I.J.; Park, J.H.; Shin, H.K.; Lim, S.S. Protein glycation inhibitors from the fruiting body of Phellinus linteus. Biol. Pharm. Bull. 2008, 31, 1968–1972. [Google Scholar] [CrossRef]
- Chen, W.; Tan, H.; Liu, Q.; Zheng, X.; Zhang, H.; Liu, Y.; Xu, L. A Review: The Bioactivities and Pharmacological Applications of Phellinus linteus. Molecules. 2019, 24, 1888. [Google Scholar] [CrossRef]
- Use of ergothioneine and/or its derivatives as an anti-glycation agent. Available online: https://patents.google.com/patent/US20020042438A1/en (accessed on 15 June 2025).
- Sun, L.; Su, X.; Zhuang, Y. Preparation, characterization and antiglycation activities of the novel polysaccharides from Boletus snicus. Int. J. Biol. Macromol. 2016, 92, 607–614. [Google Scholar] [CrossRef]
- Gallotti, F.; Lavelli, V. The Effect of UV Irradiation on Vitamin D2 Content and Antioxidant and Antiglycation Activities of Mushrooms. Foods 2020, 9, 1087. [Google Scholar] [CrossRef]
- Liuzzi, G.M.; Petraglia, T.; Latronico, T.; Crescenzi, A.; Rossano, R. Antioxidant Compounds from Edible Mushrooms as Potential Candidates for Treating Age-Related Neurodegenerative Diseases. Nutrients. 2023, 15, 1913. [Google Scholar] [CrossRef]
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).