
Submitted:
16 December 2025
Posted:
17 December 2025
You are already at the latest version
Abstract
Soil salinity is a major constraint to strawberry (Fragaria × ananassa) cultivation, adversely affecting plant growth, yield, and fruit quality. Salinity stress triggers complex physiolog-ical and biochemical responses, including osmotic adjustment, antioxidant defense, ion homeostasis, and shifts in metabolite accumulation. Genotype-specific variability in tol-erance highlights the potential for breeding salt-resilient cultivars. This review summa-rizes current knowledge on strawberry responses to salinity, emphasizing on the impacts on growth, photosynthesis, water relations, and fruit quality, as well as the underlying mechanisms of tolerance. In addition, it reviews biologically based approaches, including biostimulants, small signaling molecules, and plant–microbe interactions, that help alle-viate salinity stress and strengthen plant resilience. By integrating these physiological in-sights with advances in biological and breeding-based approaches, the review provides a comprehensive framework for improving strawberry performance under saline conditions and guiding future cultivation and genetic improvement strategies.
Keywords:
strawberry
; soil salinity
; breeding
; tolerance
; Fragaria
; biostimulants
1. Introduction
Agriculture, as the backbone of global food production, faces numerous challenges that threaten its sustainability and capacity to meet the nutritional demands of a growing population. Among these challenges, soil salinity has emerged as one of the most pressing abiotic stresses that hinder crop productivity worldwide. Soil salinization, the process by which soluble salts accumulate in the soil profile, stands as a major form of land degradation that has profound consequences for agriculture, specifically plant growth, yield, soil health, and long-term sustainability [1,2]. Climate change, intensive irrigation practices, and the overexploitation of natural resources contribute to the acceleration of salt accumulation in agricultural [3,4].
1.1. Soil Salinity and Its Impact in Agriculture
Soil salinity is a multifaceted phenomenon encompassing saline, sodic, and alkaline soil types, each defined by distinct types and concentrations of soluble salts and exchangeable cations. Saline soils contain high levels of soluble salts, resulting to reduction of the osmotic potential of the soil solution, thereby impairing plant water uptake, while sodic soils are dominated by sodium ions (Na⁺), which degrade soil structure, decrease aeration, and restrict water infiltration and movement [1]. Moreover, alkaline soils, often associated with carbonate ion accumulation, exhibit high pH values that further limit nutrient availability and root development [5]. In all cases, the presence of excessive salts in the rhizosphere alters plant–soil–water relationships, ultimately reducing crop yield and quality.
Globally, the magnitude of the problem is striking. According to recent estimates by the Food and Agriculture Organization [4], approximately 424 million hectares of surface soils and 833 million hectares of subsoils are affected by salinity worldwide. Within Europe, salinity affects nearly 15 million hectares of cultivated land, with saline, sodic, and saline-sodic soils coexisting in varying proportions. Salinity is mainly affecting riparian, semi-arid and arid areas [6,7]. According to Shokat et al., 2019 [8], the implementation of sustainable cultivation practices and the cultivation of salinity-resistant genotypes has contributed to the restoration and productive use of problematic soils in developed countries. In contrast, in developing regions, short-term management strategies have failed to yield effective or lasting solutions, thereby exacerbating food insecurity. Mediterranean countries, such as Greece, are particularly vulnerable to salinity due to their semi-arid climate, dependence on irrigation, and overexploitation of groundwater resources [9,10]. In coastal areas, seawater intrusion into aquifers has further aggravated salinity problems, compromising both water quality and agricultural productivity. Climatic projections for the region indicate rising temperatures, reduced precipitation, and increased evapotranspiration, mainly driven by climate change. These developments are expected to accelerate salt accumulation in irrigated soils, ultimately destabilizing yields and compromising crop quality [3,11,12].
The impact of salinity stress or injury to plants (salt injury) depends on the plant species and variety, growth stage, environmental conditions and salt composition [13]. A comprehensive understanding of the physiological, biochemical, and molecular mechanisms that govern plant responses to stress is essential for developing effective mitigation strategies and guiding breeding programs. Such knowledge forms the foundation of sustainable agriculture, which depends on integrating improved management practices with the cultivation of stress-resilient crop varieties [14,15].
1.2. Natural and Human-Induced Causes of Soil Salinity
The development of soil salinity results from both natural and anthropogenic factors operating across varying temporal and spatial scales. Naturally occurring salts derive from the geochemical weathering of parent rock material, which releases ions such as sodium (Na⁺), calcium (Ca²⁺), magnesium (Mg²⁺), and chloride (Cl⁻) into the soil solution [16]. In coastal environments and river deltas, seawater intrusion constitutes an additional pathway for salt enrichment. Excessive pumping of groundwater resources intensifies this process, as the decline in freshwater levels allows seawater to encroach into aquifers, leading to salinization of irrigation water and, subsequently, agricultural soils [17].
The use of low-quality water for irrigation threatens long-term agricultural sustainability by promoting salt accumulation in the root zone and soil degradation [18]. This process, commonly referred to as secondary salinization, is closely linked to irrigation and land management practices, as inadequate drainage fosters salt buildup, thereby reducing soil fertility and crop productivity. In arid and semi-arid regions, inappropriate irrigation scheduling and insufficient drainage exacerbate this process [19]. Additionally, groundwater interactions, such as seawater intrusion and water–rock reactions in coastal zones, further intensify salinity risks [20,21]. This salt accumulation also induces osmotic stress by reducing the plant’s ability to take up water due to lowered soil water potential, thereby further exacerbating the negative impacts of salinity on plant growth [22].
Human activities exacerbate these natural processes, as inefficient irrigation practices, particularly the use of brackish water combined with inadequate drainage systems, accelerate salt accumulation in the root zone [16]. Over-irrigation promotes waterlogging and reduces leaching efficiency, causing salt to rise into the root zone through capillary action. Once established, excessive salinity is difficult to remediate, as soil restoration is both costly and resource-intensive [23] (Figure 1).
2. Salinity Stress and Strawberry Cultivation
2.1. Strawberry Cultivation and Salinity Stress
Strawberry (Fragaria × ananassa Duch.) represents a particularly important case study for salinity tolerance, due to its high economic value, widespread cultivation, and pronounced sensitivity to soil and water salinity [24,25].
Belonging to the Rosaceae family, strawberries are prized for their unique flavor, nutritional value, and bioactive compounds, including anthocyanins, polyphenols, vitamins, and minerals, which contribute to their recognized health-promoting properties [26,27].
Global demand for strawberries has increased steadily over the past decades, driven by changing consumer preferences and the expanding role of strawberries in functional foods and nutraceuticals. In response to strong market demand, global production has expanded over the past decade, extending into arid and saline regions where irrigation water quality is often poor [28]. However, strawberry plants as glycophytes, are highly sensitive to abiotic stresses, particularly salinity, and exhibit a low threshold of tolerance, with yield reductions observed at soil electrical conductivity (ECe) values as low as 1.0 dS m⁻¹ [25]. For each additional unit increase in ECe, yield declines by approximately 33%, making salinity one of the most severe constraints on strawberry cultivation in irrigated regions [16]. Beyond reducing fruit yield, salinity compromises key quality traits such as sugar content, acidity, firmness, and shelf life, attributes that directly influence marketability and consumer acceptance [25,29].
Strawberries are among the irrigated crops increasingly exposed to salt stress because of secondary salinization, whose prevalence has risen with the intensification of agriculture in semi-arid and arid regions. For strawberries, a shallow-rooted crop, the consequences of secondary salinization are especially severe, leading to physiological stress [17,30]. Exposure to elevated salt levels disrupts strawberry growth and metabolism by reducing water uptake due to osmotic effects, while sodium and chloride ions accumulate in tissues, competing with essential nutrients such as potassium and calcium [31]. This ionic imbalance weakens physiological functions such as photosynthesis, stomatal regulation, and enzyme activity. The oxidative stress induced by excess salts promotes the overproduction of reactive oxygen species, which damage cellular membranes and proteins.
Although strawberries possess antioxidant defence mechanisms, their protective capacity is often insufficient under sustained salinity stress, resulting in leaf chlorosis, growth retardation, and overall yield decline. Moreover, the agronomic impacts of salinity in strawberry cultivation extend beyond yield losses, as previously discussed. Secondary salinization alters strawberry fruit quality by modifying sugar and acid balance, as well as affecting secondary metabolites, which are crucial for flavour and nutritional properties. These changes reduce market value and consumer acceptability [32]. An overview of how salt stress influences the growth, development, and productivity of strawberry plants is provided in Figure 2.
The severity of yield and quality losses can escalate when salinity is combined with shallow groundwater tables and waterlogging, which together intensify stress conditions and limit root aeration [19,33]. For strawberry cultivation, strategies such as soil flushing, adoption of salt-tolerant cultivars, and precision irrigation techniques can help mitigate stress and sustain production.
Research on strawberry salinity tolerance has expanded considerably in recent years, addressing genotypic differences as well as the morphological, physiological, biochemical, and molecular responses underlying stress adaptation [34,35,36,37,38,39,40]. Salinity stress is known to induce pronounced changes in plant morphology, leaf gas exchange, antioxidant activity, and ion homeostasis, reflecting the complexity of the strawberry stress-response network. Advances in molecular biology and genomics have further identified key genes and regulatory pathways associated with salinity tolerance, providing promising targets for breeding efforts and biotechnological applications [41,42,43]. Research has also focused on the effects of salinity on fruit quality and yield, highlighting significant physiological and metabolic adjustments that influence consumer-relevant traits [25,29,44].
Complementing genetic and molecular approaches, a wide range of agronomic strategies have been evaluated to mitigate the negative effects of salinity on strawberry growth and fruit quality. These strategies include the application of biostimulants [45], plant hormones and growth regulators [46,47], arbuscular mycorrhizal fungi [48], nanoparticles [49] organic fertilizers [50], and plant growth–promoting rhizobacteria [51,52]. Furthermore, the use of complementary light spectra and controlled-environment cultivation has been shown to enhance both stress tolerance and fruit quality [53]. Collectively, these multidimensional strategies reinforce the importance of integrating physiological, genetic, and agronomic methodologies.
Given the high economic and nutritional value of strawberries and their well-documented sensitivity to salinity, there is an urgent need to develop cultivars with enhanced tolerance while promoting sustainable agricultural practices that preserve soil and water resources. By deepening our understanding of strawberry responses to salinity and implementing interdisciplinary approaches, it becomes possible to support resilient and sustainable production systems under increasingly challenging environmental conditions.
2.2. Variation in Salinity Responses Across Different Strawberry Species and Cultivars
Strawberry species and their wild ancestors exhibit distinct responses to salinity stress, largely due to differences in genetic background and genome complexity. The genus Fragaria comprises 16 priority crop wild relatives (CWRs) that can be exploited in plant breeding to enhance cultivated strawberries, offering opportunities to improve fruit quality, tolerance to abiotic stresses, and resistance to pests and diseases [54]. Comparative analyses of salinity tolerance mechanisms across Fragaria species are therefore essential for identifying key regulatory nodes and molecular pathways underpinning adaptive responses to salinity stress.
Among these CWRs, the Chilean strawberry (Fragaria chiloensis), an octoploid wild ancestor of the cultivated strawberry (F. × ananassa), comprises four subspecies: F. chiloensis subsp. chiloensis, F. chiloensis subsp. lucida, F. chiloensis subsp. pacifica, and F. chiloensis subsp. sandwicensis. This species is widely regarded as tolerant of abiotic stresses, particularly salinity and drought, and represents a valuable genetic resource for strawberry breeding [55,56,57].
Consistent with the diversity of stress responses observed among Fragaria CWRs, Nikoloudi [58], demonstrated that F. virginiana and F. chiloensis differ markedly in their responses to salinity stress. Although both species are wild ancestors of the cultivated strawberry, F. chiloensis exhibited a substantially greater capacity to withstand saline conditions, highlighting functional divergence within the genus and its relevance for breeding programs.
Subsequent comparative studies involving cultivated varieties and CWRs further substantiated these findings. Garriga et al. in 2015 [59], evaluated salinity effects on growth performance, physiological responses, productivity, and fruit quality in three strawberry genotypes: the commercial cultivar ‘Camarosa’, the cultivated Chilean strawberry Bau (F. chiloensis subsp. chiloensis f. chiloensis), and the wild Chilean strawberry Cucao (F. chiloensis subsp. chiloensis f. patagonica). Notably, F. chiloensis f. patagonica was largely unaffected by salt stress, indicating superior salinity tolerance among the genotypes examined.
Plant responses to salinity also vary substantially among cultivars within the same species. Because tolerance alone is insufficient for commercial production, breeding efforts increasingly aim to identify cultivars that combine stress resistance with high yield, superior fruit quality, and early production [25,60]. Controlled-environment studies consistently reveal pronounced cultivar-dependent variation in salinity responses. Denaxa et al. [34] reported that ‘Camarosa’ and ‘Rociera’ exhibit moderate salinity tolerance through distinct physiological and biochemical mechanisms, while Demiral et al. [61] identified significant cultivar × salinity interactions in ‘Camarosa’ and ‘Sweet Charlie’, attributable to contrasting Na⁺ regulation strategies. Specifically, ‘Camarosa’ predominantly restricts Na⁺ uptake at the root level, whereas ‘Sweet Charlie’ limits Na⁺ translocation to the shoots.
Morphological traits further contribute to differential tolerance. For example, Elsanta’ exhibits lower stomatal density than ‘Elsinore’ [62], a characteristic associated with improved water-use efficiency under salinity stress [63]. Additional physiological differentiation was documented by Turhan et al. [64], who, building on earlier work [65,66,67], demonstrated marked differences between ‘Camarosa’ and ‘Chandler’ in stomatal regulation, LT₅₀ values, and antioxidant enzyme activity. Similarly, Ghaderi [35] reported greater salinity tolerance in ‘Kurdistan’ compared with ‘Queen Elisa’, associated with enhanced antioxidant capacity and higher relative water content (RWC).
Furthermore, Kortekamp et al. [68] assessed salinity tolerance in an F₂ population derived from a cross between the salt-sensitive cultivar ‘Senga Sengana’ and a salt-tolerant F. chiloensis ssp. lucida accession. Under greenhouse salt stress, F₂ individuals exhibited variable necrotic damage and growth reduction, revealing genetic variation in salt tolerance and identifying genotypes that could serve as a valuable resource for breeding strawberries with enhanced stress resilience.
Beyond whole-plant physiology, cultivars also differ in biochemical, metabolic, and fruit-quality responses to salinity. Alnayef et al. [69] observed increased antioxidant capacity and phenolic accumulation in ‘Elsanta’ and ‘Elsinore’ under saline conditions, with mild stress (≤20 mM) even improving fruit quality when combined with appropriate fertilization. At the cellular level, Hussein et al. [37] documented reductions in chlorophyll content, biomass, and growth in ‘Fortuna’ and ‘Festival’, accompanied by elevated proline accumulation and antioxidant activity. Similar osmoprotective responses were reported by Pirlak et al. [70], who observed proline accumulation alongside changes in Na⁺ and Cl⁻ concentrations in ‘Fern’ and ‘Camarosa’. Under hydroponic conditions, Rahimi et al. in 2011 [71] further demonstrated cultivar-specific ionic imbalances in ‘Camarosa’, characterized by increased Na⁺ accumulation, reduced Ca²⁺ and Mg²⁺ uptake, and concomitant growth inhibition.
Metabolic studies further revealed cultivar-dependent sensitivity. Keutgen [32] reported increased malondialdehyde (MDA), amino acids, and proline in ‘Korona’ and ‘Elsanta’, with ‘Elsanta’ exhibiting greater sensitivity and reduced fruit quality. Ondrašek et al. [72] similarly documented declines in fruit yield, number, and size under salinity, along with accelerated leaf senescence and shortened growth duration. Long-term experiments by Keutgen and Pawelzik [31] showed that although salinity reduced yield in both ‘Elsanta’ and ‘Korona’, the latter maintained more stable flavor-related traits. Notably, Galli et al. [28] demonstrated that moderate salinity in ‘Camarosa’ increased sugar content, anthocyanins, phenolics, and abscisic acid (ABA) without reducing yield, suggesting that controlled salinity may enhance nutritional quality.
Taken together, these studies demonstrate substantial genotypic variation in strawberry responses to salinity stress, reflecting complex and genotype-dependent physiological, biochemical, and morphological adaptation strategies. Traits such as efficient ionic regulation, enhanced antioxidant defense, osmotic adjustment, and modified stomatal behavior play key roles in determining tolerance. In particular, the capacity to balance Na⁺ exclusion with Cl⁻ compartmentalization, maintain relative water content, and sustain metabolic activity under moderate salinity appears critical for preserving productivity and fruit quality. Collectively, these findings underscore the importance of integrating physiological screening with molecular and agronomic approaches to identify and develop strawberry cultivars capable of thriving under saline conditions,
3. Mitigation Mechanisms in Strawberry under Salinity Stress
3.1. The Role of Small Signaling Molecules and Biostimulants in Mitigating Salinity Stress in Strawberry
The external application of small signaling molecules, classified as biostimulants, has been shown to significantly mitigate the adverse effects of various abiotic stresses, including salinity [73]. Among these, 5-Aminolevulinic acid (ALA) serves as the primary precursor for tetrapyrrole compounds, namely chlorophyll, phytochrome and heme [74,75,76]. Wu et al. (2018) [76] reported that ALA enhance salt stress tolerance by promoting chlorophyll biosynthesis, improving light-harvesting efficiency and photosynthetic performance, and inhibiting thylakoid membrane degradation. Subsequently, they demonstrated that ALA appears to promote root-specific retention of Na⁺ in strawberry plants, thereby limiting its translocation to the shoots [75,76]. ALA sequentially upregulated the expression of the NHX1 (Na+/H+ exchanger), HKT1 (High-affinity K+ uptake), and SOS1 (Salt overly sensitive 1) genes and increased the hydrogen peroxide (H2O2) content in the roots, thus enhancing the salt tolerance of Fragaria x ananassa cv. ‘Benihoppe’ strawberry. These findings highlight ALA’s potential as an effective agent for improving salt tolerance of strawberry plants under saline conditions.
Melatonin (N-acetyl-5-methoxy-tryptamine) is another important small signaling molecule, which plays a vital role as an antioxidant in plant stress responses. During salt stress conditions, melatonin accumulation is commonly observed, functioning as a ROS scavenger and enhancing the cellular antioxidant capacity of plant cells [77,78,79]. In the study of Zahedi et al. [47], the foliar applications of melatonin have been shown to mitigate the adverse effects of salinity in strawberry leaves, namely increasing the leaf photosynthetic pigments and reducing the changes in macronutrients concentrations induced by salinity. At the same time these melatonin foliar sprays improved fruit yield, size, firmness and fruit chemical parameters. Georgiadou et al. [80], have also demonstrated that preharvest application of melatonin with sodium alginate on strawberry fruit enhanced ellagitannins and ellagic acid derivatives, confirming the beneficial effect of melatonin on the antioxidant mechanism of plants. Continued investigation into the metabolic and regulatory networks of melatonin is essential to optimize its application for enhancing plant resilience to salt stress.
Salicylic acid (SA) is a phenolic compound which is also acknowledged as a signaling molecule that participates in regulating plant responses to various abiotic stress conditions [81]. Karlidag et al. [82], investigated the impact of various SA concentrations on the growth, chlorophyll content, and mineral composition of strawberry plants subjected to salt stress. Regarding growth parameters and nutrient content, SA treatments resulted in higher fresh and dry weight of both shoots and roots along with a general enhancement of nutrient concentrations in both the leaves and roots of strawberry plants exposed to salt stress. Jamali and Eshghi [83], proposed that the time of SA application is a significant parameter to consider, since the pretreatment of strawberry plants with SA before NaCl induced salt stress showed higher levels of root Mg and shoot Ca in comparison to salt-stressed plants treated with the same SA concentration one week after salt exposure. In cv. ‘Gaviota’ strawberries under salt stress, foliar application of 100 μM SA effectively alleviated damage by enhancing PSII efficiency, promoting the accumulation of compatible solutes such as trehalose, and increasing antioxidant enzyme activity and phenolic content [84]. This led to reduced lipid peroxidation and membrane damage, highlighting the role of SA in improving photosynthetic performance and activating defense mechanisms under salinity stress. The results of Roshdy et al. [85], indicate that foliar application of salicylic acid at 90 ppm can mitigate the adverse effects of salinity on the growth of the strawberry cv. ‘Camarosa’. The improvement in key indicators of salinity response, such as Salt Tolerance Index (STI), catalase (CAT), peroxidase (POD) and proline (PRO), following foliar application of SA in strawberries further supports its potential to alleviate the detrimental effects of salinity on the growth of strawberry plants.
Humic acid (HA) has been reported to confer beneficial effects under abiotic stress by improving soil physicochemical properties, such as osmotic and ionic balance, cation exchange capacity, and microbial activity, while also enhancing plant morphological traits, physiological processes (including photosynthesis, chlorophyll concentration, and CO₂ assimilation), and overall biochemical responses [22,86,87]. Saidimoradi et al. [50], reported that HA application in two commercial strawberry cultivars under salinity stress stimulated antioxidant enzyme activity and promoted effective osmotic adjustment, thereby enhancing stress tolerance. The addition of humic acid to the nutrient solution significantly reduced Na⁺ accumulation in both roots and leaves, while simultaneously increasing chlorophyll content in salinity-stressed plants. Moreover, humic acid treatment resulted in increased leaf area and fruit number compared with plants exposed to NaCl alone.
The foliar application of 1mM of acetic acid, one week before the NaCl treatment in strawberry plants enhanced their salinity tolerance [88]. Acetic acid serves as a precursor to acetyl-CoA, a central metabolic intermediate essential for macromolecule biosynthesis and energy production that supports plant cell growth [89]. Treatment with acetic acid under NaCl stress benefited strawberry plants by protecting chlorophyll from degradation, reducing toxic ion accumulation in leaves, and increasing osmoprotectants like soluble carbohydrates and proline, along with non-enzymatic antioxidants such as phenols and ascorbic acid in fruits.
The study of Moradi et al. [41], highlights the promising potential of silicon nanoparticles (SiNPs) and methyl jasmonate (MeJA) in enhancing the physiological responses of strawberry plants under salinity stress. While SiNPs were found to significantly improve chlorophyll content and reduce oxidative damage by enhancing antioxidant enzyme activity, the study suggests that the mechanisms by which SiNPs modulate stress-related gene expression, such as for SOS1 and HKT1, still require further exploration. Similarly, MeJA’s role in improving plant stress tolerance through upregulation of stress-related genes is well-supported, but optimal concentrations and application methods for maximizing its benefits remain unclear.
3.2. Microorganisms and Plant-Microbe Interactions under Salinity
In recent years, increasing attention has been given to the role of beneficial soil microorganisms, particularly arbuscular mycorrhizal fungi (AMF), in enhancing plant tolerance to salinity. These symbiotic fungi form associations with plant roots, facilitating improved water and nutrient acquisition, especially phosphorus and micronutrients, and modulating physiological responses under salt stress [90,91]. Inoculation of strawberry plants with the arbuscular mycorrhizal fungus Gigaspora margarita significantly improved salt tolerance by enhancing shoot and root biomass, maintaining chlorophyll content, and reducing leaf browning under 200 mM NaCl stress [48]. Mycorrhizal plants showed lower Na⁺ accumulation and a reduced Na⁺/K⁺ ratio in both root and shoot tissues compared to non-inoculated controls. SEM-EDX analysis revealed that excess sodium was primarily localized around the vascular bundles, with mycorrhizal plants accumulating less Na⁺ overall. Additionally, increased cellulose and lignin content in the roots of mycorrhizal plants suggested the formation of a stronger apoplastic barrier, contributing to the suppression of Na⁺ uptake and improved physiological resilience under salinity stress.
A study by Redondo-Gómez et.al, examined whether Plant Growth Promoting Rhizobacteria (PGPR) isolated from halophytes could improve the growth of eight major crops, including strawberry, commonly cultivated in salinized soils. Using environmental conditions that mimicked future climate scenarios (elevated CO₂, higher temperature, and salinity), they indicated that these biofertilizers generally enhanced crop growth and mitigated the negative effects of salt stress. Strawberry and rice demonstrated the strongest positive responses, with notably higher biomass when inoculated with PGPR under saline conditions. Overall, the study demonstrated that halotolerant PGPR can boost crop performance under current and future climate-related stress conditions [92].
In a similar context, a recent study focused on the effects of hydrogen-rich water (HRW) on alleviating salt stress in strawberry ‘Benihoppe’ seedlings. They presented that HRW improved plant growth, increased root biomass, enhanced antioxidant activity, promoted K⁺ uptake while it reduced Na⁺ accumulation, and modulated key metabolites. Transcriptomic and metabolomic analyses revealed that HRW activated genes and pathways related to ion transport, antioxidant defense, and cell wall biosynthesis, with phenylpropanoid biosynthesis and amino/nucleoside sugar metabolism proved particularly important for mitigating salt stress [93].
GABA (γ-aminobutyric acid) is a non-protein amino acid that acts as a signaling molecule in plants. It helps plants cope with salinity by regulating osmolytes, enhancing antioxidant defenses, reducing oxidative stress, and modulating gene expression related to stress tolerance, photosynthesis, and ion balance. Exploiting that knowledge, recently, Zhang et al. studied the effects of γ-aminobutyric acid (GABA) on salt tolerance in strawberry seedlings. GABA treatment under salt stress improved osmolyte accumulation, antioxidant capacity, and photosynthesis, while reducing ROS and MDA levels. Transcriptomic analysis demonstrated upregulation of genes related to chlorophyll metabolism and photosynthesis, while 38 transcription factors (WRKY, AP2/ERF, MYB) were identified as potentially contributing to GABA-induced salt tolerance. Overall, GABA enhanced strawberry salt resistance and could be useful in breeding programs by revealing physiological markers and transcriptional targets linked to salt tolerance [42].
4. Genetic and Molecular Approaches for Improving Salinity Tolerance in Strawberry
Gene Identification and Functional Annotation for Breeding Targets
Recent studies have broadened the genetic understanding of salinity tolerance in strawberry by uncovering novel regulatory genes and elucidating previously unknown stress-response mechanisms [94]. Gene identification and functional annotation provide a critical framework for understanding the molecular mechanisms underlying complex stress-adaptive traits. Such approaches enable the identification of key candidate genes that can be targeted to enhance stress tolerance through breeding [95].
Devinder Sandhu [96], explored the relationship between salinity tolerance and the expression of seven genes associated with Na⁺ transport (SOS1, SOS2, SOS3, NHX1, NHX2, AKT1, SAL1) and four genes involved in Cl⁻ transport (CLC_G, CLC_C, SLAH3, ALMT12). The study assessed biomass production, ion accumulation (particularly Na⁺ and Cl⁻), and a range of physiological and genetic parameters across eight genotypes of F. vesca. Notably, the upregulation of genes involved in Na⁺ and Cl⁻ transport was strongly associated with the plant’s ability to sequester or exclude toxic ions, thereby influencing overall performance and survivability under saline conditions. The findings underscore the importance of deciphering the genetic mechanisms that mediate salt tolerance, as such insights may inform the development of commercially viable, salt-tolerant strawberry cultivars.
Several studies have employed similar strategies to explore other stress-related regulatory networks. Li et al. [97], focused on the Growth-Regulating Factor (GRF) gene family in F. vesca). Ten FvGRF genes were identified and classified into five subfamilies, with several members, specifically FvGRF3, FvGRF6, and FvGRF8, showing significant upregulation in response to salt stress. These GRF proteins, characterized by the conserved Glutamine–Leucine–Glutamine (QLQ) and Tryptophan–Arginine–Cysteine (WRC) DNA-binding domains, are implicated in chromatin remodeling and transcriptional regulation. Their putative involvement in ABA signaling pathways further supports a role in the plant’s abiotic stress response network; however, functional validation in transgenic strawberry lines is still required.
Husaini et al. [98], reported that transgenic strawberry plants expressing the osmotin gene showed enhanced tolerance to salinity stress compared with control plants. These plants accumulated higher levels of proline and total soluble proteins, maintained greater chlorophyll content, and performed better under high salinity conditions, tolerating electrical conductivity levels up to 20 dS m⁻¹. The improved stress tolerance was associated with elevated osmotin expression, indicating that this gene plays a key role in salinity adaptation and represents a promising target for improving salt tolerance in strawberry.
Through a targeted analysis of superoxide dismutase (SOD) genes in octoploid strawberry cv. ‘Benihoppe’, Zhang et al. [99], identified specific isoforms, such as Manganese Superoxide Dismutase 5 (FaMSOD5), with salinity-induced expression patterns, highlighting their potential role in stress mitigation. However, the absence of functional validation limits their immediate use as breeding targets, underscoring the need for further molecular and physiological studies to confirm their contribution to salt tolerance.
Broadening the molecular understanding of salinity responses, Kong et al. [100], explored enzymatic responses associated with salt-induced oxidative stress in woodland strawberry (F. vesca). Among 41 identified Laccase (FvLAC) genes, 20 representative members were analyzed under salt treatment using qRT-PCR. The expression of FvLAC7, FvLAC24, FvLAC32, and FvLAC51 increased steadily over 12 hours of NaCl exposure. Functional validation in transgenic yeast revealed contrasting roles: FvLAC51 enhanced salt tolerance, while FvLAC24 and FvLAC32 reduced it. These results suggest that members of the laccase family play differential roles in the regulation of redox balance and cell wall remodeling under saline conditions, with FvLAC51 acting as a potential positive regulator of salt tolerance in strawberries.
Transcription factors (TFs) are gene families which play many roles in plants’ biological processes including regulation of salinity stress tolerance [101]. Indicatively, the NAC gene family includes several transcription factors, which are known to regulate various aspects of plant development, tissue differentiation, and responses to abiotic stress [102]. Zhang et al. in 2018 [103], investigated the NAC transcription factor family in woodland strawberry. They identified 37 FvNAC genes and systematically analyzed their gene structures, conserved motifs, and expression patterns across different tissues. Expression analyses under abiotic stresses (cold, heat, drought, and salt), biotic stresses (pathogen infection), and exposure to signaling molecules revealed that several FvNAC genes responded strongly to stress, particularly under cold and H₂O₂ treatments. In addition, some FvNAC genes were either upregulated or downregulated during pathogen infection. More specifically, for the fungal Colletotrichum gloeosporioides, genes FvNAC04, FvNAC28, FvNAC29 found upregulated, while FvNAC09, FvNAC18, FvNAC20 presented down regulated. Regarding the Ralstonia solanacearum bacteria, significant down regulation was detected for FvNAC02, FvNAC13, FvNAC14, FvNAC21, FvNAC26, and FvNAC32 genes. These findings highlight stress-responsive regulation of specific FvNAC genes and identify them as potential candidates for improving salinity stress tolerance in woodland strawberry.
More targeted research on the FvNAC gene family, identified FaNAC2, a transcription factor from the ATAF sub-family in ‘Benihoppe’ strawberry, as a key regulator in stress responses [104]. FaNAC2 expression is induced under various abiotic stresses and is predominantly localized in the shoot apical meristem and older leaves. Its overexpression in Nicotiana benthamiana enhanced tolerance to salt, cold, and drought stress, likely through upregulation of proline biosynthesis and ABA biosynthesis genes. These results suggest that FaNAC2 plays a beneficial role in enhancing abiotic stress tolerance and represents a promising target for improving stress tolerance in strawberry.
In a more recent study, Wenhui Li et al. [105], cloned the FvNAC29 gene from F. vesca. The study indicated a potential link between FvNAC29 expression and the plant’s molecular mechanisms for coping with high salinity stress. Transgenic lines exhibited reduced accumulation of reactive oxygen species (ROS) and malondialdehyde (MDA) along with elevated levels of proline and chlorophyll. In addition, there was an up regulation in the activities of key antioxidant enzymes: CAT, SOD, and POD. These findings suggest that FvNAC29 may contribute to enhanced salinity tolerance by modulating both oxidative stress responses and osmoprotective pathways.
In a follow-up study, Wenhui Li et al., cloned the FaTINY2 gene from the octoploid strawberry. FaTINY2 is a member of the APETALA2/Ethylene Responsive Factor (AP2/ERF) transcription factor family, which is involved in diverse biological functions, including plant morphogenesis, stress signaling pathways, hormone regulation, and secondary metabolite biosynthesis [106]. The expression of FaTINY2 in transgenic lines was associated with enhanced antioxidant defense, as evidenced by reduced levels of ROS and MDA. In addition, transgenic plants exhibited increased concentrations of proline and chlorophyll, along with elevated enzymatic activities of CAT, SOD, and POD. Collectively, these findings indicate that FaTINY2 plays an important regulatory role in salinity tolerance by coordinating antioxidant defense mechanisms and osmotic adjustment [107].
Building on the understanding of transcriptional and metabolic responses to salinity, Li et al. [108], analyzed two contrasting strawberry cultivars, the salt-tolerant ‘Sweet Charlie’ and the salt-sensitive ‘Benihoppe’ using integrated transcriptomic and metabolomic profiling. The tolerant cultivar showed a more robust response to salt stress, with a higher number of differentially expressed genes and metabolites compared to the sensitive one. Notably, genes associated with cell wall restructuring and ion regulation were more frequently upregulated in ‘Sweet Charlie’, suggesting these pathways play a central role in its adaptive capacity. A distinct pattern was also observed for genes encoding ATP-binding cassette (ABC) transporters, which were linked to coordinated changes in metabolite accumulation, particularly in pathways involving secondary metabolism. These differences indicate that enhanced activation of transport systems and reinforcement of cell wall architecture may underpin varietal differences in salt stress tolerance, providing potential targets for breeding programs focused on stress resilience in cultivated strawberry.
The MYB transcription factor family represents a promising target associated with salt stress tolerance in strawberry [109]. A novel MYB transcription factor gene FvMYB82, isolated from F. vesca, has been characterized by Wenhui Li et al. [110], characterized. Functional analyses in transgenic lines revealed elevated proline and chlorophyll levels, together with enhanced activities of key antioxidant enzymes, including SOD, POD, and CAT. These results suggest that FvMYB82 plays a central regulatory role in the strawberry salt stress response by integrating osmotic adjustment and antioxidant defense pathways.
In a complementary study on the MYB transcription factor family, Wang et al. in 2021 [109], characterized FvMYB24, an R2R3-MYB transcription factor from F. vesca and demonstrated its functional role in salinity tolerance using transgenic Arabidopsis thaliana lines. Plants overexpressing FvMYB24 exhibited improved performance under salt stress, including increased germination rates, root elongation, fresh biomass, and chlorophyll content. Physiological assays revealed elevated proline levels and antioxidant enzyme activities (SOD, POD, CAT), alongside reduced malondialdehyde (MDA) content, indicating enhanced cellular redox regulation and oxidative stress mitigation. Gene expression analysis showed that key salt-responsive genes, including AtSOS1, AtNHX1, AtLEA3, were upregulated in the transgenic lines. Importantly, FvMYB24 was shown to bind the SOS1 promoter, acting as a positive regulator of SOS1-mediated Na⁺ transport.
Kula et al. [111], compared the responses of two strawberry cultivars, ‘Petaluma’ and ‘Cabrillo’, to 0, 150, and 300 mM NaCl for ten days, combining physiological assays with gene expression analysis (qRT-PCR) to associate transcription factor activity with stress tolerance. Their findings revealed that ‘Petaluma’ displayed superior tolerance, maintaining growth parameters more effectively, experiencing lower oxidative damage, and significantly increasing anthocyanin accumulation compared with ‘Cabrillo’. At the molecular level, Petaluma showed strong upregulation of the MYB5 transcription factor, which correlated almost perfectly with anthocyanin accumulation, and early induction of ATF1-2, a stress-responsive regulator. These coordinated physiological and molecular adjustments underpin Petaluma’s enhanced ability to withstand high salinity, highlighting the importance of transcription factor regulation in strawberry salt tolerance.
In addition to gene expression and protein regulation, salt stress influences epigenetic mechanisms that control gene activity in strawberry. In this context, López et al. [112], found that epigenetic modifications, particularly DNA methylation, mediate stress responses in F. vesca. More specifically, salt stress triggered widespread hypomethylation, particularly in the CHG and CHH sequence contexts, with differentially methylated regions (DMRs) primarily located in promoter and intergenic regions. RNA-seq analysis revealed over 5,000 salt-responsive genes, enriched in functions related to transport, homeostasis, hydrolase and motor activity, as well as pyrophosphatase activity. Notably, MYB transcription factors represented the most enriched regulatory group among these DEGs. A subset of these salt-responsive genes showed concurrent methylation and expression changes, suggesting that DNA methylation acts as a context-specific modulator of gene activity. These findings indicate that epigenetic remodeling under salt stress may influence the expression of genes involved in key physiological processes, contributing to adaptive responses in F. vesca.
Chu et al. [113], isolated and characterized FaTEDT1L, a transcription factor from octoploid cultivated strawberry, which shares high homology with AtEDT1/HDG11 of Arabidopsis. Functional validation in transgenic Arabidopsis demonstrated enhanced tolerance to both drought and salt stress, with transgenic lines showing increased root growth, decreased water loss, and upregulated expression of SOS pathway genes (SOS1, SOS2, and SOS3). These findings suggest that FaTEDT1L may act as a master regulator of osmotic stress response and could be a strong candidate for engineering salinity-tolerant strawberry cultivars.
A genome-wide association study (GWAS) is a genomic approach that scans genetic variation across an organism’s entire genome to identify associations between specific genetic variants and phenotypic traits. Unlike studies limited to a few candidate genes, GWAS surveys the whole genome to find variants correlated with trait variation, providing researchers with a deeper understanding of the species under study [114].
Lei et al. [115], performed GWAS of the glucose-6-phosphate dehydrogenase (G6PDH) gene family in strawberry, identifying 19 members involved in redox regulation via the pentose phosphate pathway. Several G6PDH genes responded to salt stress, indicating their role in protecting cells from oxidative damage through NADPH production. G6PDH genes contribute to cellular detoxification and redox homeostasis under salinity conditions [116]. The integration of transcriptional and metabolic strategies appears critical for engineering effective salinity tolerance in strawberry.
Crizel et al. [117], conducted genome-wide identification and characterization of the calcium-dependent protein kinase (CDPK) gene family in Fragaria × ananassa cv. Camarosa, identifying 35 FaCDPK genes with diverse structural domains and expression patterns. Under salt stress, several members, particularly FaCDPK11, FaCDPK15, and FaCDPK27, showed pronounced upregulation, indicating their active involvement in salinity tolerance. These findings highlight the pivotal role of CDPK-mediated calcium signaling in the salt stress response, wherein Ca²⁺-dependent phosphorylation pathways activate specific downstream genes and physiological processes that collectively enhance the plant’s ability to maintain ion homeostasis and mitigate cellular damage under saline conditions.
Recent studies have provided additional insights into the molecular mechanisms underlying salinity tolerance. Li et al. [118], examined another important gene family involved in stress regulation. They conducted a comprehensive GWAS of the C2H2-Zing Finger Proteins (C2H2-ZFPs) C1-2i subclass in ‘Benihoppe’ cultivar. C2H2-ZFPs are crucial regulators of plant responses to diverse environmental stresses. Under salt stress, the expression levels of nineteen variants of FaZAT8, FaZAT10, FaZAT11 and FaZAT12 were elevated in roots relative to untreated control. Transcriptome data highlighted FaZAT10 as exhibiting the highest expression in roots under these conditions, indicating that it may serve as a key regulator in stress signal transduction.
Focusing on a different aspect of salt stress response, Jiang et al. [119], performed GWAS of the U-box E3 ubiquitin ligase gene family. Among the 155 identified FaU-box genes, several exhibited marked transcriptional responses to salt stress. qRT-PCR analysis revealed that FaU-box9, FaU-box18, FaU-box52, and FaU-box65 were rapidly induced within three hours of NaCl treatment, whereas FaU-box3, FaU-box83, and FaU-box142 showed gradual upregulation, peaking at 12 hours before slightly declining. The authors suggested that U-box E3 ubiquitin ligases may regulate salinity tolerance through targeted ubiquitination of specific signaling proteins, influencing their stability and degradation to adjust cellular responses under stress.

Table 1.
Key genes and proteins involved in strawberry salinity tolerance related to ion homeostasis, osmotic adjustment, and stress signaling pathways.
Table 1.
Key genes and proteins involved in strawberry salinity tolerance related to ion homeostasis, osmotic adjustment, and stress signaling pathways.
| Gene / Protein | Function / Role In SalinityTolerance | Mechanism / Pathway | Reference |
|---|---|---|---|
| SOS1 | Plasma membrane Na⁺/H⁺ antiporter; removes Na⁺ from cytoplasm | Na⁺ transport / ionhomeostasis | [96,120] |
| SOS2 | Part of SOS pathway; regulates Na⁺ homeostasis | Na⁺ transport / signaltransduction | [96] |
| SOS3 | Part of SOS pathway; regulates Na⁺ homeostasis | Na⁺ transport / signaltransduction | [96] |
| NHX1 | Vacuolar Na⁺/H⁺ antiporter; sequesters Na⁺ into vacuoles | Na⁺ compartmentalization / ion homeostasis | [96,120] |
| NHX2 | Vacuolar Na⁺/H⁺ antiporter; ion compartmentalization | Na⁺ compartmentalization / ion homeostasis | [96] |
| HKT1 | Na⁺ transporter; regulates Na⁺ uptake and distribution | Na⁺ transport / ionhomeostasis | [96,120] |
| AVP1 | Vacuolar H⁺-PPase; generates proton gradient for ion sequestration | Proton gradient / ioncompartmentalization | [120] |
| Osmotin | Enhances proline, total soluble protein, chlorophyllcontent; improves salinitytolerance | Osmotic adjustment / stress-responsive protein | [98,121] |
| FvGRF3 | Growth-Regulating Factor; involved in chromatin remodeling and ABA-mediated stress response | Transcriptional regulation / ABA signaling | [97] |
| FvGRF6 | Growth-Regulating Factor; involved in chromatin remodeling and ABA-mediated stress response | Transcriptional regulation / ABA signaling | [97] |
| FvGRF8 | Growth-Regulating Factor; involved in chromatin remodeling and ABA-mediated stress response | Transcriptional regulation / ABA signaling | [97] |
| FvMYB82 | MYB transcription factor; increases proline, chlorophyll, and antioxidant enzyme activity | Transcriptional regulation / antioxidant defense | [110] |
| FaNAC2 | NAC transcription factor; regulates proline and ABA biosynthesis under stress | Transcriptional regulation / osmotic adjustment | [104] |
| FvNAC29 | NAC transcription factor; reduces ROS and MDA, enhances antioxidant enzymes and osmotic adjustment | Transcriptional regulation / oxidative stress & osmoprotection | [105] |
| FaTINY2 | AP2/ERF transcription factor; coordinates osmotic adjustment and antioxidant defense | Transcriptional regulation / stress signaling | [107] |
| FaTEDT1L | HDG11-like transcriptionfactor; enhances root growth, reduces waterloss, upregulates SOS genes | Transcriptional regulation / osmotic stress response | [113] |
| G6PDH gene family | Redox regulation via pentosephosphate pathway; protects cells from oxidative stress and contributes to NADPH production under salinity | Redox homeostasis / ROS detoxification | [115] |
| AKT1 | K⁺ channel; maintains ionbalance under salinity stress | K⁺ transport / ionhomeostasis | [96] |
| SAL1 | Phosphatase involved in stress signaling and ionhomeostasis | Signal transduction / stress signaling | [96] |
| CLC_G | Chloride channel; regulates Cl⁻ transport and homeostasis | Cl⁻ transport / ionhomeostasis | [96] |
| CLC_C | Chloride channel; regulates Cl⁻ transport and homeostasis | Cl⁻ transport / ionhomeostasis | [96] |
| SLAH3 | Anion channel; contributesto Cl⁻ homeostasis under salt stress | Cl⁻ transport / ionhomeostasis | [96] |
| ALMT12 | Malate transporter; contributes to ion balance and stress response | Organic acid transport / ion balance | [96] |
5. Future Prospects
Future research on strawberry salinity tolerance will benefit from integrating multilevel approaches that span genetics, physiology, molecular biology, and agronomy. The rapid identification of key regulatory genes, such as members of transcription factors (NACs, AP2/ERFs, MYBs), ion-transport regulators and redox-related genes, provides promising targets for breeding programs aimed at improving salt tolerance while preserving fruit quality. These findings highlight the need for functional validation, gene pyramiding, and the translation of candidate genes into marker-assisted selection, genomic selection, and CRISPR/Cas-based engineering to accelerate the development of elite cultivars. The genetic diversity of wild relatives, particularly F. chiloensis, offers additional reservoirs of tolerance traits that can be introgressed into commercial cultivars.
At the same time, advances in biostimulants and small signaling molecules (e.g., ALA, melatonin, salicylic acid, silicon nanoparticles, acetic acid) show strong potential for mitigating salinity effects by enhancing antioxidant capacity, osmotic adjustment, ion homeostasis, and stress-responsive gene expression. Future studies should establish standardized application protocols and test these compounds under field conditions. Similarly, plant–microbe interactions, including arbuscular mycorrhizal fungi and plant growth–promoting rhizobacteria, represent eco-friendly tools that can complement genetic improvement by enhancing nutrient uptake, reducing Na⁺ toxicity, and reinforcing root structural barriers. Integrating these biological approaches with precision irrigation, soil salinity monitoring, controlled-environment horticulture, and sustainable water management will allow growers to maintain salinity within manageable thresholds—especially given evidence that mild salinity can enhance fruit nutritional quality in certain cultivars.
Collectively, future progress will depend on bridging molecular insights with practical cultivation strategies, combining advanced genomic resources with optimized agronomic and microbial interventions. Such a holistic perspective will be essential for developing resilient strawberry production systems capable of sustaining yield and fruit quality under increasingly saline and resource-constrained environments.
6. Conclusions
Soil salinity is a major abiotic constraint that severely limits both the productivity and nutritional quality of strawberry crops. This review synthesizes current knowledge of strawberry responses to salinity stress, with emphasis on the underlying physiological, biochemical, and molecular mechanisms. Salinity impairs plant growth, photosynthetic performance, water relations, and fruit quality, while inducing osmotic adjustment, activation of antioxidant defenses, and shifts in primary and secondary metabolite profiles. Significant genotypic variation shapes the degree of tolerance, underscoring the need for identifying and developing salt-tolerant cultivars. By integrating evidence from diverse studies, this review provides a framework for breeding strategies aimed at enhancing strawberry resilience to saline environments and offers practical insights to support growers managing production in salt-affected regions.
Author Contributions
“Conceptualization, K.P., E.T., and V.P.; writing—original draft preparation, K.P., M.Go., M.Ge.; writing—review and editing, K.P., M.Ge., E.T., and V.P.; visualization, M.Ge.; supervision, E.T. and V.P.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Pisinaras, V.; Tsihrintzis, V.A.; Petalas, C.; Ouzounis, K. Soil Salinization in the Agricultural Lands of Rhodope District, Northeastern Greece. Environmental monitoring and assessment 2010, 166, 79–94. [Google Scholar] [CrossRef]
- Hardie, M.; Doyle, R. Measuring Soil Salinity. In Plant Salt Tolerance;Methods in Molecular Biology; Shabala, S., Cuin, T.A., Eds.; Humana Press: Totowa, NJ, 2012; Vol. 913, pp. 415–425. ISBN 978-1-61779-985-3. [Google Scholar]
- Daliakopoulos, I.N.; Tsanis, I.K.; Koutroulis, A.; Kourgialas, N.N.; Varouchakis, A.E.; Karatzas, G.P.; Ritsema, C.J. The Threat of Soil Salinity: A European Scale Review. Science of the total environment 2016, 573, 727–739. [Google Scholar] [CrossRef]
- FAO Global Map of Salt-Affected Soils 2021.
- Tóth, T. Classification and Mitigation of Soil Salinization. In Oxford Research Encyclopedia of Environmental Science; 2017. [Google Scholar]
- Singh, M.; Nara, U.; Kumar, A.; Choudhary, A.; Singh, H.; Thapa, S. Salinity Tolerance Mechanisms and Their Breeding Implications. Journal of Genetic Engineering and Biotechnology 2021, 19, 173. [Google Scholar] [CrossRef] [PubMed]
- Pessarakli, M.; Szabolcs, I. Soil Salinity and Sodicity as Particular Plant/Crop Stress Factors. In Handbook of Plant and Crop Stress, Fourth Edition ed; CRC press, 2019; pp. 3–21. [Google Scholar]
- Shokat, S.; Großkinsky, D.K. Tackling Salinity in Sustainable Agriculture—What Developing Countries May Learn from Approaches of the Developed World. Sustainability 2019, 11, 4558. [Google Scholar] [CrossRef]
- Petalas, C.; Pisinaras, V.; Gemitzi, A.; Tsihrintzis, V.A.; Ouzounis, K. Current Conditions of Saltwater Intrusion in the Coastal Rhodope Aquifer System, Northeastern Greece. Desalination 2009, 237, 22–41. [Google Scholar] [CrossRef]
- Karamanos, A.; Aggelides, S.; Londra, P. Water Use Efficiency and Water Productivity in Greece. Options Méditerranéennes 2005, 57, 92–100. [Google Scholar]
- Yuan, X.; Li, S.; Chen, J.; Yu, H.; Yang, T.; Wang, C.; Huang, S.; Chen, H.; Ao, X. Impacts of Global Climate Change on Agricultural Production: A Comprehensive Review. Agronomy 2024, 14, 1360. [Google Scholar] [CrossRef]
- Terán, F.; Vives-Peris, V.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Facing Climate Change: Plant Stress Mitigation Strategies in Agriculture. Physiologia Plantarum 2024, 176, e14484. [Google Scholar] [CrossRef]
- Zaman, M.; Shahid, S.A.; Heng, L. Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Springer Nature, 2018. [Google Scholar]
- Negrão, S.; Schmöckel, S.M.; Tester, M. Evaluating Physiological Responses of Plants to Salinity Stress. Annals of botany 2017, 119, 1–11. [Google Scholar] [CrossRef]
- Xiong, W.; Reynolds, M.; Xu, Y. Climate Change Challenges Plant Breeding. Current Opinion in Plant Biology 2022, 70, 102308. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R.; Maas, E.V. Plant Salt Tolerance. ASCE manual and reports on engineering practice 2012, 71, 405–459. [Google Scholar]
- Maas, E.V.; Grattan, S.R. Crop Yields as Affected by Salinity. In Agronomy Monographs; Skaggs, R.W., Schilfgaarde, J., Eds.; Wiley, 1999; Vol. 38, pp. 55–108. ISBN 978-0-89118-141-5. [Google Scholar]
- Singh, A. Salinization and Drainage Problems of Agricultural Land. Irrigation and Drainage 2020, 69, 844–853. [Google Scholar] [CrossRef]
- Endo, T.; Yamamoto, S.; Larrinaga, J.A.; Fujiyama, H.; Honna, T. Status and Causes of Soil Salinization of Irrigated Agricultural Lands in Southern Baja California, Mexico. Applied and Environmental Soil Science 2011, 2011, 1–12. [Google Scholar] [CrossRef]
- Rengasamy, P. World Salinization with Emphasis on Australia. J Exp Bot 2006, 57, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Bouzourra, H.; Bouhlila, R.; Elango, L.; Slama, F.; Ouslati, N. Characterization of Mechanisms and Processes of Groundwater Salinization in Irrigated Coastal Area Using Statistics, GIS, and Hydrogeochemical Investigations. Environmental Science and Pollution Research 2015, 22, 2643–2660. [Google Scholar] [CrossRef]
- Nabi, F.; Sarfaraz, A.; Kama, R.; Kanwal, R.; Li, H. Structure-Based Function of Humic Acid in Abiotic Stress Alleviation in Plants: A Review. Plants 2025, 14, 1916. [Google Scholar] [CrossRef]
- Gkiougkis, I.; Kallioras, A.; Pliakas, F.; Pechtelidis, A.; Diamantis, V.; Diamantis, I.; Ziogas, A.; Dafnis, I. Assessment of Soil Salinization at the Eastern Nestos River Delta, NE Greece. Catena 2015, 128, 238–251. [Google Scholar] [CrossRef]
- Hancock, J.F. Temperate Fruit Crop Breeding: Germplasm to Genomics; Springer Science & Business Media, 2008. [Google Scholar]
- Ferreira, J.F.; Liu, X.; Suarez, D.L. Fruit Yield and Survival of Five Commercial Strawberry Cultivars under Field Cultivation and Salinity Stress. Scientia Horticulturae 2019, 243, 401–410. [Google Scholar] [CrossRef]
- Mazzoni, L.; Perez-Lopez, P.; Giampieri, F.; Alvarez-Suarez, J.M.; Gasparrini, M.; Forbes-Hernandez, T.Y.; Quiles, J.L.; Mezzetti, B.; Battino, M. The Genetic Aspects of Berries: From Field to Health. J Sci Food Agric 2016, 96, 365–371. [Google Scholar] [CrossRef]
- Giampieri, F.; Alvarez-Suarez, J.M.; Battino, M. Strawberry and Human Health: Effects beyond Antioxidant Activity. J. Agric. Food Chem. 2014, 62, 3867–3876. [Google Scholar] [CrossRef]
- Galli, V.; da Silva Messias, R.; Perin, E.C.; Borowski, J.M.; Bamberg, A.L.; Rombaldi, C.V. Mild Salt Stress Improves Strawberry Fruit Quality. Lwt 2016, 73, 693–699. [Google Scholar] [CrossRef]
- Wu, Y.; Li, L.; Li, M.; Zhang, M.; Sun, H.; Sigrimis, N. Optimal Fertigation for High Yield and Fruit Quality of Greenhouse Strawberry. PLoS One 2020, 15, e0224588. [Google Scholar] [CrossRef] [PubMed]
- Shannon, M.C.; Grieve, C.M. Tolerance of Vegetable Crops to Salinity. Scientia horticulturae 1998, 78, 5–38. [Google Scholar] [CrossRef]
- Keutgen, A.J.; Pawelzik, E. Quality and Nutritional Value of Strawberry Fruit under Long Term Salt Stress. Food chemistry 2008, 107, 1413–1420. [Google Scholar] [CrossRef]
- Keutgen, A.J.; Pawelzik, E. Impacts of NaCl Stress on Plant Growth and Mineral Nutrient Assimilation in Two Cultivars of Strawberry. Environmental and experimental botany 2009, 65, 170–176. [Google Scholar] [CrossRef]
- Singh, A. Soil Salinization and Waterlogging: A Threat to Environment and Agricultural Sustainability. Ecological indicators 2015, 57, 128–130. [Google Scholar] [CrossRef]
- Denaxa, N.-K.; Nomikou, A.; Malamos, N.; Liveri, E.; Roussos, P.A.; Papasotiropoulos, V. Salinity Effect on Plant Growth Parameters and Fruit Bioactive Compounds of Two Strawberry Cultivars, Coupled with Environmental Conditions Monitoring. Agronomy 2022, 12, 2279. [Google Scholar] [CrossRef]
- Ghaderi, N.; Hatami, M.R.; Mozafari, A.; Siosehmardeh, A. Change in Antioxidant Enzymes Activity and Some Morpho-Physiological Characteristics of Strawberry under Long-Term Salt Stress. Physiology and molecular biology of plants 2018, 24, 833–843. [Google Scholar] [CrossRef]
- Mozafari, A.; Ghaderi, N.; Havas, F.; Dedejani, S. Comparative Investigation of Structural Relationships among Morpho-Physiological and Biochemical Properties of Strawberry (Fragaria\times Ananassa Duch.) under Drought and Salinity Stresses: A Study Based on in Vitro Culture. Scientia Horticulturae 2019, 256, 108601. [Google Scholar] [CrossRef]
- Hussein, A.E.; El-Kerdany, A.Y.; Afifi, K.M. Effect of Drought and Salinity Stresses on Two Strawberry Cultivars during Their Regeneration in Vitro. International Journal of Innovative Science Engineering and Technology 2016, 4, 83–93. [Google Scholar]
- Akbari, M.; Mahna, N.; Ramesh, K.; Bandehagh, A.; Mazzuca, S. Ion Homeostasis, Osmoregulation, and Physiological Changes in the Roots and Leaves of Pistachio Rootstocks in Response to Salinity. Protoplasma 2018, 255, 1349–1362. [Google Scholar] [CrossRef] [PubMed]
- Sikder, R.K.; Wang, X.; Zhang, H.; Gui, H.; Dong, Q.; Jin, D.; Song, M. Nitrogen Enhances Salt Tolerance by Modulating the Antioxidant Defense System and Osmoregulation Substance Content in Gossypium Hirsutum. Plants 2020, 9, 450. [Google Scholar] [CrossRef] [PubMed]
- Asghar, R.; Biglarifard, A.; Mirdehghan, H.; Borghei, S.F. Influence of NaCl Salinity on Growth Analysis of Strawberry Cv. Camarosa. Journal of Stress Physiology & Biochemistry 2011, 7, 145–156. [Google Scholar]
- Moradi, P.; Vafaee, Y.; Mozafari, A.A.; Tahir, N.A. Silicon Nanoparticles and Methyl Jasmonate Improve Physiological Response and Increase Expression of Stress-Related Genes in Strawberry Cv. Paros under Salinity Stress. Silicon 2022, 14, 10559–10569. [Google Scholar] [CrossRef]
- Zhang, Y.; Deng, M.; Lin, B.; Tian, S.; Chen, Y.; Huang, S.; Lin, Y.; Li, M.; He, W.; Wang, Y.; et al. Physiological and Transcriptomic Evidence Revealed the Role of Exogenous GABA in Enhancing Salt Tolerance in Strawberry Seedlings. BMC Genomics 2025, 26, 196. [Google Scholar] [CrossRef]
- Golnari, S.; Vafaee, Y.; Nazari, F.; Ghaderi, N. Gamma-Aminobutyric Acid (GABA) and Salinity Impacts Antioxidative Response and Expression of Stress-Related Genes in Strawberry Cv. Aromas. Brazilian Journal of Botany 2021, 44, 639–651. [Google Scholar] [CrossRef]
- Keutgen, A.; Pawelzik, E. Modifications of Taste-Relevant Compounds in Strawberry Fruit under NaCl Salinity. Food chemistry 2007, 105, 1487–1494. [Google Scholar] [CrossRef]
- Sangiorgio, D.; Cellini, A.; Spinelli, F.; Donati, I. Promoting Strawberry (Fragaria\times Ananassa) Stress Resistance, Growth, and Yield Using Native Bacterial Biostimulants. Agronomy 2023, 13, 529. [Google Scholar] [CrossRef]
- Faghih, S.; Ghobadi, C.; Zarei, A. Response of Strawberry Plant Cv.‘Camarosa’to Salicylic Acid and Methyl Jasmonate Application under Salt Stress Condition. Journal of Plant Growth Regulation 2017, 36, 651–659. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Hosseini, M.S.; Abadía, J.; Marjani, M. Melatonin Foliar Sprays Elicit Salinity Stress Tolerance and Enhance Fruit Yield and Quality in Strawberry (Fragaria\times Ananassa Duch.). Plant Physiology and Biochemistry 2020, 149, 313–323. [Google Scholar] [CrossRef]
- Haque, S.I.; Matsubara, Y. Salinity Tolerance and Sodium Localization in Mycorrhizal Strawberry Plants. Communications in Soil Science and Plant Analysis 2018, 49, 2782–2792. [Google Scholar] [CrossRef]
- Zeid, I.M.A.; Mohamed, F.H.; Metwali, E.M. Responses of Two Strawberry Cultivars to NaCl-Induced Salt Stress under the Influence of ZnO Nanoparticles. Saudi Journal of Biological Sciences 2023, 30, 103623. [Google Scholar] [CrossRef] [PubMed]
- Saidimoradi, D.; Ghaderi, N.; Javadi, T. Salinity Stress Mitigation by Humic Acid Application in Strawberry (Fragaria x Ananassa Duch.). Scientia Horticulturae 2019, 256, 108594. [Google Scholar] [CrossRef]
- Karlidag, H.; Yildirim, E.; Turan, M.; Pehluvan, M.; Donmez, F. Plant Growth-Promoting Rhizobacteria Mitigate Deleterious Effects of Salt Stress on Strawberry Plants (Fragaria\times Ananassa). HortScience 2013, 48, 563–567. [Google Scholar] [CrossRef]
- Yavuz, A.; Erdogan, U.; Turan, M.; Argın, S.; Kocaman, A. Synergistic Strategies for Overcoming Salt Stress in Strawberry Farming: The Use of Organic Fertilizers and Plant Growth Promoting Rhizobacteria (PGPR). Applied Fruit Science 2024, 66, 1787–1797. [Google Scholar] [CrossRef]
- Malekzadeh Shamsabad, M.R.; Esmaeilizadeh, M.; Roosta, H.R.; Dąbrowski, P.; Telesiński, A.; Kalaji, H.M. Supplemental Light Application Can Improve the Growth and Development of Strawberry Plants under Salinity and Alkalinity Stress Conditions. Scientific Reports 2022, 12, 9272. [Google Scholar] [CrossRef]
- Hu, Y.; Feng, C.; Yang, L.; Edger, P.P.; Kang, M. Genomic Population Structure and Local Adaptation of the Wild Strawberry Fragaria Nilgerrensis. Horticulture Research 2022, 9, uhab059. [Google Scholar] [CrossRef]
- Bringhurst, R.S.; Hancock, J.F.; Voth, V. The Beach Strawberry, an Important Natural Resource; 1977. [Google Scholar]
- VanDerZanden, A.M.; Cameron, J.S. Effect of Water Deficit Stress on 11 Native Fragaria Chiloensis Clones Selected as Ornamental Groundcovers. Scientia horticulturae 1996, 66, 241–253. [Google Scholar] [CrossRef]
- Carrasco, B.; Garcés, M.; Rojas, P.; Saud, G.; Herrera, R.; Retamales, J.B.; Caligari, P.D. The Chilean Strawberry [Fragaria Chiloensis (L.) Duch.]: Genetic Diversity and Structure. Journal of the American Society for Horticultural Science 2007, 132, 501–506. [Google Scholar] [CrossRef]
- Nikoloudi, A. Salinity Tolerance in Strawberry (Fragaria Spp) as Influenced by Genotype; Michigan State University, 2003. [Google Scholar]
- Garriga, M.; Muñoz, C.A.; Caligari, P.D.; Retamales, J.B. Effect of Salt Stress on Genotypes of Commercial (Fragaria x Ananassa) and Chilean Strawberry (F. Chiloensis). Scientia Horticulturae 2015, 195, 37–47. [Google Scholar] [CrossRef]
- Suarez, D.L.; Grieve, C.M. GROWTH, YIELD, AND ION RELATIONS OF STRAWBERRY IN RESPONSE TO IRRIGATION WITH CHLORIDE-DOMINATED WATERS. Journal of Plant Nutrition 2013, 36, 1963–1981. [Google Scholar] [CrossRef]
- Demiral, M.A. Effect of Salt Stress on Concentration of Nitrogen and Phosphorus in Root and Leaf of Strawberry Plant. Eurasian Journal of Soil Science 2017, 6, 357–364. [Google Scholar] [CrossRef]
- Orsini, F.; Alnayef, M.; Bona, S.; Maggio, A.; Gianquinto, G. Low Stomatal Density and Reduced Transpiration Facilitate Strawberry Adaptation to Salinity. Environmental and Experimental Botany 2012, 81, 1–10. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Zhou, M.; Shabala, S. How Does Stomatal Density and Residual Transpiration Contribute to Osmotic Stress Tolerance? Plants 2023, 12, 494. [Google Scholar] [CrossRef]
- Turhan, E.; Eris, A. Changes of Growth, Amino Acids, and Ionic Composition in Strawberry Plants under Salt Stress Conditions. Communications in Soil Science and Plant Analysis 2009, 40, 3308–3322. [Google Scholar] [CrossRef]
- Turhan, E.; Eris, A. Changes of Micronutrients, Dry Weight, and Chlorophyll Contents in Strawberry Plants Under Salt Stress Conditions*. Communications in Soil Science and Plant Analysis 2005, 36, 1021–1028. [Google Scholar] [CrossRef]
- Turhan, E.; Eris, A. Effects of Sodium Chloride Applications and Different Growth Media on Ionic Composition in Strawberry Plant. Journal of Plant Nutrition 2005, 27, 1653–1665. [Google Scholar] [CrossRef]
- Gulen, H.; Turhan, E.; Eris, A. Changes in Peroxidase Activities and Soluble Proteins in Strawberry Varieties under Salt-Stress. Acta Physiologiae Plantarum 2006, 28, 109–116. [Google Scholar] [CrossRef]
- Kortekamp, T.; Chen, T.-W.; Wagner, H.; Olbricht, K. Evaluation of Salinity Tolerance in a Fragaria F2 Population. In Proceedings of the XVI EUCARPIA Symposium on Fruit Breeding and Genetics 1412, 2023; pp. 111–116. [Google Scholar]
- Alnayef, M.; Hartley, J.; Orsini, F.; Di Silvestro, R.; Sanoubar, R.; Marotti, I.; Gianquinto, G.; Dinelli, G.; Puniran-Hartley, N. Using Salinity to Improve Nutritional and Market Value of Strawberries. Australian Journal of Crop Science 2022, 16, 7–S3. [Google Scholar] [CrossRef]
- Pirlak, L.; Eşitken, A. Salinity Effects on Growth, Proline and Ion Accumulation in Strawberry Plants. Acta Agriculturae Scandinavica, Section B - Soil & Plant Science 2004, 54, 189–192. [Google Scholar] [CrossRef]
- Rahimi, A.; Biglarifard, A. Influence of NaCl Salinity and Different Substracts on Plant Growth, Mineral Nutrient Assimilation and Fruit Yield of Strawberry. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 2011, 39, 219–226. [Google Scholar] [CrossRef]
- Ondrašek, G.; Romić, D.; Romić, M.; Duralija, B.; Mustač, I. Strawberry Growth and Fruit Yield in a Saline Environment. Agriculturae conspectus scientificus 2006, 71, 155–158. [Google Scholar]
- Chan, Z.; Shi, H. Improved Abiotic Stress Tolerance of Bermudagrass by Exogenous Small Molecules. Plant Signaling & Behavior 2015, 10, e991577. [Google Scholar] [CrossRef] [PubMed]
- Averina, N.G.; Gritskevich, E.R.; Vershilovskaya, I.V.; Usatov, A.V.; Yaronskaya, E.B. Mechanisms of Salt Stress Tolerance Development in Barley Plants under the Influence of 5-Aminolevulinic Acid. Russian Journal of Plant Physiology 2010, 57, 792–798. [Google Scholar] [CrossRef]
- Wu, Y.; Liao, W.; Dawuda, M.M.; Hu, L.; Yu, J. 5-Aminolevulinic Acid (ALA) Biosynthetic and Metabolic Pathways and Its Role in Higher Plants: A Review. Plant Growth Regul 2019, 87, 357–374. [Google Scholar] [CrossRef]
- Wu, Y.; Jin, X.; Liao, W.; Hu, L.; Dawuda, M.M.; Zhao, X.; Tang, Z.; Gong, T.; Yu, J. 5-Aminolevulinic Acid (ALA) Alleviated Salinity Stress in Cucumber Seedlings by Enhancing Chlorophyll Synthesis Pathway. Frontiers in plant science 2018, 9, 635. [Google Scholar] [CrossRef]
- Wei, W.; Li, Q.-T.; Chu, Y.-N.; Reiter, R.J.; Yu, X.-M.; Zhu, D.-H.; Zhang, W.-K.; Ma, B.; Lin, Q.; Zhang, J.-S. Melatonin Enhances Plant Growth and Abiotic Stress Tolerance in Soybean Plants. Journal of experimental botany 2015, 66, 695–707. [Google Scholar] [CrossRef]
- Li, J.; Liu, J.; Zhu, T.; Zhao, C.; Li, L.; Chen, M. The Role of Melatonin in Salt Stress Responses. International journal of molecular sciences 2019, 20, 1735. [Google Scholar] [CrossRef]
- Pardo-Hernández, M.; López-Delacalle, M.; Rivero, R.M. ROS and NO Regulation by Melatonin under Abiotic Stress in Plants. Antioxidants 2020, 9, 1078. [Google Scholar] [CrossRef]
- Georgiadou, E.C.; García, C.J.; Taliadorou, A.M.; Gedeon, S.; Valanides, N.; Varaldo, A.; Gohari, G.; Balsells-Llauradó, M.; Alcázar, R.; Hertog, M.L. Pre-Harvest Application of Sodium Alginate Functionalized with Melatonin Enhances Secondary Metabolism in Strawberry Fruit. Current Plant Biology 2025, 43, 100515. [Google Scholar] [CrossRef]
- Bandurska, H. Salicylic Acid: An Update on Biosynthesis and Action in Plant Response to Water Deficit and Performance Under Drought. In SALICYLIC ACID; Hayat, S., Ahmad, A., Alyemeni, M.N., Eds.; Springer Netherlands: Dordrecht, 2013; pp. 1–14. ISBN 978-94-007-6427-9. [Google Scholar]
- Karlidag, H.; Yildirim, E.; Turan, M. Melhora Do Efeito de Estresse Salino Em Morango Por Adição de Ácido Salicílico. Scientia Agricola 2009, 66, 180–187. [Google Scholar] [CrossRef]
- Jamali, B.; Eshghi, S. Salicylic Acid–Induced Salinity Redressal in Hydroponically Grown Strawberry. Communications in Soil Science and Plant Analysis 2015, 46, 1482–1493. [Google Scholar] [CrossRef]
- Samadi, S.; Habibi, G.; Vaziri, A. Effects of Exogenous Salicylic Acid on Antioxidative Responses, Phenolic Metabolism and Photochemical Activity of Strawberry under Salt Stress. Iranian Journal of Plant Physiology 2019, 9, 2685–2694. [Google Scholar]
- Roshdy, A.E.-D.; Alebidi, A.; Almutairi, K.; Al-Obeed, R.; Elsabagh, A. The Effect of Salicylic Acid on the Performances of Salt Stressed Strawberry Plants, Enzymes Activity, and Salt Tolerance Index. Agronomy 2021, 11, 775. [Google Scholar] [CrossRef]
- Canellas, L.P.; Da Silva, R.M.; Busato, J.G.; Olivares, F.L. Humic Substances and Plant Abiotic Stress Adaptation. Chem. Biol. Technol. Agric. 2024, 11, 66. [Google Scholar] [CrossRef]
- Mora, V.; Olaetxea, M.; Bacaicoa, E.; Baigorri, R.; Fuentes, M.; Zamarreño, A.M.; Garcia-Mina, J.M. Abiotic Stress Tolerance in Plants: Exploring the Role of Nitric Oxide and Humic Substances. In Nitric Oxide in Plants: Metabolism and Role in Stress Physiology; Khan, M.N., Mobin, M., Mohammad, F., Corpas, F.J., Eds.; Springer International Publishing: Cham, 2014; pp. 243–264. ISBN 978-3-319-06709-4. [Google Scholar]
- Mirfattahi, Z.; Eshghi, S.; Gharaghani, A.; Etemadi, M.; Moghadam, A. Time and Rate of Acetate Foliar Spray Can Ameliorate Adverse Effect of NaCl Stress on Strawberry. South African Journal of Botany 2022, 150, 797–805. [Google Scholar] [CrossRef]
- Rahman, Md.M.; Keya, S.S.; Sahu, A.; Gupta, A.; Dhingra, A.; Tran, L.-S.P.; Mostofa, M.G. Acetic Acid: A Cheap but Chief Metabolic Regulator for Abiotic Stress Tolerance in Plants. Stress Biology 2024, 4, 34. [Google Scholar] [CrossRef] [PubMed]
- Evelin, H.; Kapoor, R.; Giri, B. Arbuscular Mycorrhizal Fungi in Alleviation of Salt Stress: A Review. Annals of botany 2009, 104, 1263–1280. [Google Scholar] [CrossRef] [PubMed]
- Boorboori, M.R.; Lackóová, L. Arbuscular Mycorrhizal Fungi and Salinity Stress Mitigation in Plants. Frontiers in Plant Science 2025, 15, 1504970. [Google Scholar] [CrossRef]
- Redondo-Gómez, S.; Mesa-Marín, J.; Pérez-Romero, J.A.; López-Jurado, J.; García-López, J.V.; Mariscal, V.; Molina-Heredia, F.P.; Pajuelo, E.; Rodríguez-Llorente, I.D.; Flowers, T.J. Consortia of Plant-Growth-Promoting Rhizobacteria Isolated from Halophytes Improve Response of Eight Crops to Soil Salinization and Climate Change Conditions. Agronomy 2021, 11, 1609. [Google Scholar] [CrossRef]
- Wang, R.; Chu, S.; Zhang, D.; Hayat, K.; Zhang, X.; Chi, Y.; Ma, X.; Chen, X.; Yang, H.; Ding, W.; et al. Alleviation of Salt Stress in Strawberries by Hydrogen-rich Water: Physiological, Transcriptomic and Metabolomic Responses. Physiologia Plantarum 2025, 177, e70151. [Google Scholar] [CrossRef]
- Ullah, I.; Toor, M.D.; Yerlikaya, B.A.; Mohamed, H.I.; Yerlikaya, S.; Basit, A.; Rehman, A. High-Temperature Stress in Strawberry: Understanding Physiological, Biochemical and Molecular Responses. Planta 2024, 260, 118. [Google Scholar] [CrossRef] [PubMed]
- Roychowdhury, R.; Das, S.P.; Gupta, A.; Parihar, P.; Chandrasekhar, K.; Sarker, U.; Kumar, A.; Ramrao, D.P.; Sudhakar, C. Multi-Omics Pipeline and Omics-Integration Approach to Decipher Plant’s Abiotic Stress Tolerance Responses. Genes 2023, 14, 1281. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, D.; Pudussery, M.V.; Ferreira, J.F.; Liu, X.; Pallete, A.; Grover, K.K.; Hummer, K. Variable Salinity Responses and Comparative Gene Expression in Woodland Strawberry Genotypes. Scientia Horticulturae 2019, 254, 61–69. [Google Scholar] [CrossRef]
- Li, Z.; Xie, Q.; Yan, J.; Chen, J.; Chen, Q. Genome-Wide Identification and Characterization of the Abiotic-Stress-Responsive GRF Gene Family in Diploid Woodland Strawberry (Fragaria Vesca). Plants 2021, 10, 1916. [Google Scholar] [CrossRef]
- Husaini, A.M.; Abdin, M.Z. Development of Transgenic Strawberry (Fragaria x Ananassa Duch.) Plants Tolerant to Salt Stress. Plant Science 2008, 174, 446–455. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Y.; Gao, W.; Tian, S.; Lin, B.; Gu, X.; Lin, Y.; Li, M.; Wang, Y.; He, W. Characterization of Superoxide Dismutase (SOD) Gene Family and Their Responses to Salinity Stress and Fruit Development in Octoploid Strawberry. Agronomy 2024, 14, 2514. [Google Scholar] [CrossRef]
- Kong, J.; Xiong, R.; Qiu, K.; Lin, X.; Li, D.; Lu, L.; Zhou, J.; Zhu, S.; Liu, M.; Sun, Q. Genome-Wide Identification and Characterization of the Laccase Gene Family in Fragaria Vesca and Its Potential Roles in Response to Salt and Drought Stresses. Plants 2024, 13, 3366. [Google Scholar] [CrossRef]
- Fernando, V.D. Major Transcription Factor Families Involved in Salinity Stress Tolerance in Plants. Transcription factors for abiotic stress tolerance in plants 2020, 99–109. [Google Scholar]
- Shao, H.; Wang, H.; Tang, X. NAC Transcription Factors in Plant Multiple Abiotic Stress Responses: Progress and Prospects. Frontiers in plant science 2015, 6, 902. [Google Scholar] [CrossRef]
- Zhang, H.; Kang, H.; Su, C.; Qi, Y.; Liu, X.; Pu, J. Genome-Wide Identification and Expression Profile Analysis of the NAC Transcription Factor Family during Abiotic and Biotic Stress in Woodland Strawberry. PLoS One 2018, 13, e0197892. [Google Scholar] [CrossRef]
- Liang, J.; Zheng, J.; Wu, Z.; Wang, H. Strawberry FaNAC2 Enhances Tolerance to Abiotic Stress by Regulating Proline Metabolism. Plants 2020, 9, 1417. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, H.; Wei, Y.; Han, J.; Wang, Y.; Li, X.; Zhang, L.; Han, D. Overexpression of a Fragaria Vesca NAM, ATAF, and CUC (NAC) Transcription Factor Gene (FvNAC29) Increases Salt and Cold Tolerance in Arabidopsis Thaliana. International Journal of Molecular Sciences 2024, 25, 4088. [Google Scholar] [CrossRef] [PubMed]
- Licausi, F.; Ohme-Takagi, M.; Perata, P. APETALA 2/Ethylene Responsive Factor ( AP 2/ ERF ) Transcription Factors: Mediators of Stress Responses and Developmental Programs. New Phytologist 2013, 199, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, W.; Li, H.; Yao, A.; Ma, Z.; Kang, R.; Guo, Y.; Li, X.; Yu, W.; Han, D. Overexpression of a Fragaria\times Ananassa AP2/ERF Transcription Factor Gene (FaTINY2) Increases Cold and Salt Tolerance in Arabidopsis Thaliana. International Journal of Molecular Sciences 2025, 26, 2109. [Google Scholar]
- Li, S.; Chang, L.; Sun, R.; Dong, J.; Zhong, C.; Gao, Y.; Zhang, H.; Wei, L.; Wei, Y.; Zhang, Y. Combined Transcriptomic and Metabolomic Analysis Reveals a Role for Adenosine Triphosphate-Binding Cassette Transporters and Cell Wall Remodeling in Response to Salt Stress in Strawberry. Frontiers in Plant Science 2022, 13, 996765. [Google Scholar] [CrossRef]
- Wang, S.; Shi, M.; Zhang, Y.; Xie, X.; Sun, P.; Fang, C.; Zhao, J. FvMYB24, a Strawberry R2R3-MYB Transcription Factor, Improved Salt Stress Tolerance in Transgenic Arabidopsis. Biochemical and Biophysical Research Communications 2021, 569, 93–99. [Google Scholar] [CrossRef]
- Li, W.; Zhong, J.; Zhang, L.; Wang, Y.; Song, P.; Liu, W.; Li, X.; Han, D. Overexpression of a Fragaria Vesca MYB Transcription Factor Gene (FvMYB82) Increases Salt and Cold Tolerance in Arabidopsis Thaliana. International Journal of Molecular Sciences 2022, 23, 10538. [Google Scholar] [CrossRef]
- Kula, N.; Öztürk Erdem, S.; Ünal, D. Salt Tolerance Mechanisms in Strawberry Cultivars: Comparing the Physiological and Molecular Responses of Petaluma and Cabrillo. Applied Fruit Science 2025, 67, 379. [Google Scholar] [CrossRef]
- López, M.-E.; Roquis, D.; Becker, C.; Denoyes, B.; Bucher, E. DNA Methylation Dynamics during Stress Response in Woodland Strawberry (Fragaria Vesca). Horticulture research 2022, 9, uhac174. [Google Scholar] [CrossRef]
- Chu, C.-Y.; Lin, L.-F.; Lai, S.-C.; Yang, J.-H.; Chou, M.-L. FaTEDT1L of Octoploid Cultivated Strawberry Functions as a Transcriptional Activator and Enhances Abiotic Stress Tolerance in Transgenic Arabidopsis. International Journal of Molecular Sciences 2024, 25, 10091. [Google Scholar] [CrossRef] [PubMed]
- Tibbs Cortes, L.; Zhang, Z.; Yu, J. Status and Prospects of Genome-wide Association Studies in Plants. The Plant Genome 2021, 14, e20077. [Google Scholar] [CrossRef] [PubMed]
- Lei, D.; Lin, Y.; Luo, M.; Zhao, B.; Tang, H.; Zhou, X.; Yao, W.; Zhang, Y.; Wang, Y.; Li, M. Genome-Wide Investigation of G6PDH Gene in Strawberry: Evolution and Expression Analysis during Development and Stress. International Journal of Molecular Sciences 2022, 23, 4728. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-H.; Tjong, W.-Y.; Yang, H.-C.; Liu, H.-Y.; Stern, A.; Chiu, D.T.-Y. Glucose-6-Phosphate Dehydrogenase, Redox Homeostasis and Embryogenesis. International Journal of Molecular Sciences 2022, 23. [Google Scholar] [CrossRef]
- Crizel, R.L.; Perin, E.C.; Vighi, I.L.; Woloski, R.; Seixas, A.; da Silva Pinto, L.; Rombaldi, C.V.; Galli, V. Genome-Wide Identification, and Characterization of the CDPK Gene Family Reveal Their Involvement in Abiotic Stress Response in Fragaria x Ananassa. Scientific reports 2020, 10, 11040. [Google Scholar] [CrossRef]
- Li, H.; Yue, M.; Jiang, L.; Liu, Y.; Zhang, N.; Liu, X.; Ye, Y.; Lin, X.; Zhang, Y.; Lin, Y. Genome-Wide Identification of Strawberry C2H2-ZFP C1-2i Subclass and the Potential Function of FaZAT10 in Abiotic Stress. International Journal of Molecular Sciences 2022, 23, 13079. [Google Scholar] [CrossRef]
- Jiang, L.; Lin, Y.; Wang, L.; Peng, Y.; Yang, M.; Jiang, Y.; Hou, G.; Liu, X.; Li, M.; Zhang, Y. Genome-Wide Identification and Expression Profiling Reveal the Regulatory Role of U-Box E3 Ubiquitin Ligase Genes in Strawberry Fruit Ripening and Abiotic Stresses Resistance. Frontiers in Plant Science 2023, 14, 1171056. [Google Scholar] [CrossRef]
- Zhang, J.-L.; Shi, H. Physiological and Molecular Mechanisms of Plant Salt Tolerance. Photosynthesis research 2013, 115, 1–22. [Google Scholar] [CrossRef]
- Singh, N.K.; Nelson, D.E.; Kuhn, D.; Hasegawa, P.M.; Bressan, R.A. Molecular Cloning of Osmotin and Regulation of Its Expression by ABA and Adaptation to Low Water Potential. Plant physiology 1989, 90, 1096–1101. [Google Scholar] [CrossRef]
Figure 1.
Overview of the major physiological and biochemical stress pathways triggered by soil salinity in plants from both natural and anthropogenic factors.
Figure 1.
Overview of the major physiological and biochemical stress pathways triggered by soil salinity in plants from both natural and anthropogenic factors.
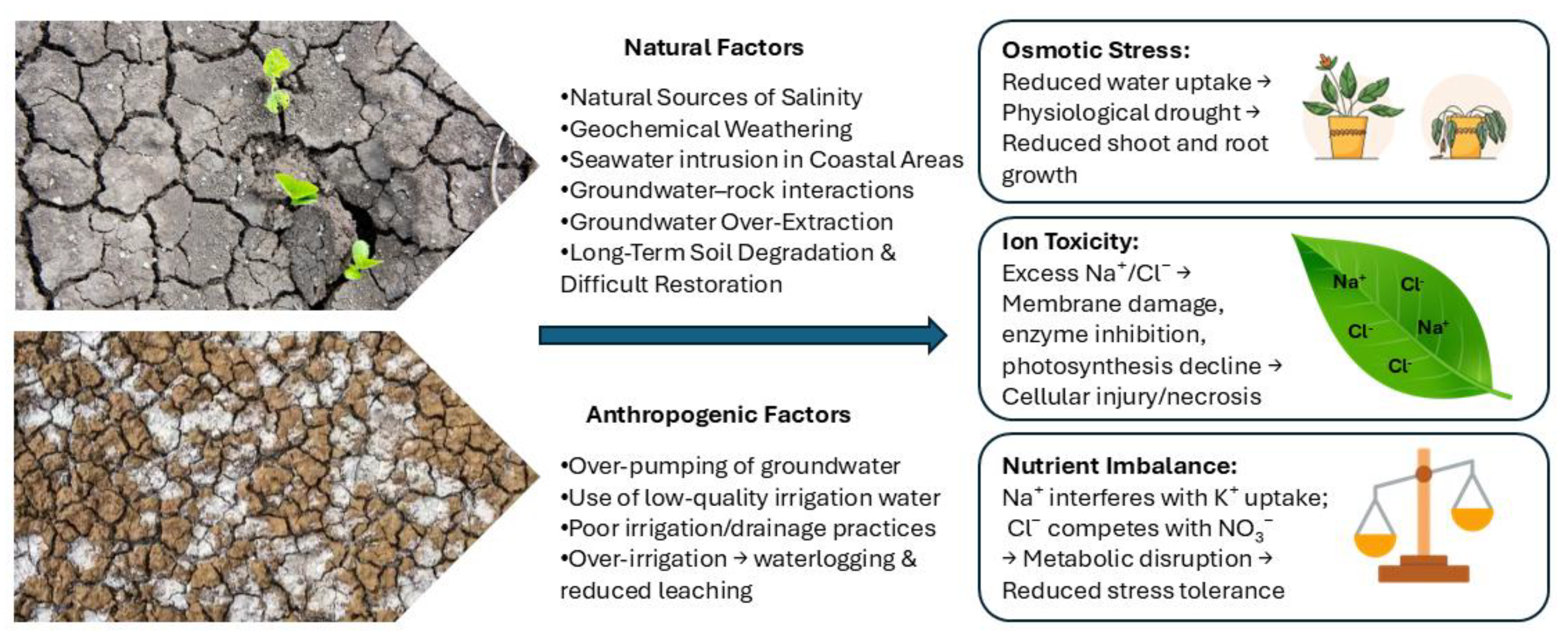
Figure 2.
Figure 2. Secondary salinization effects on strawberry plants.
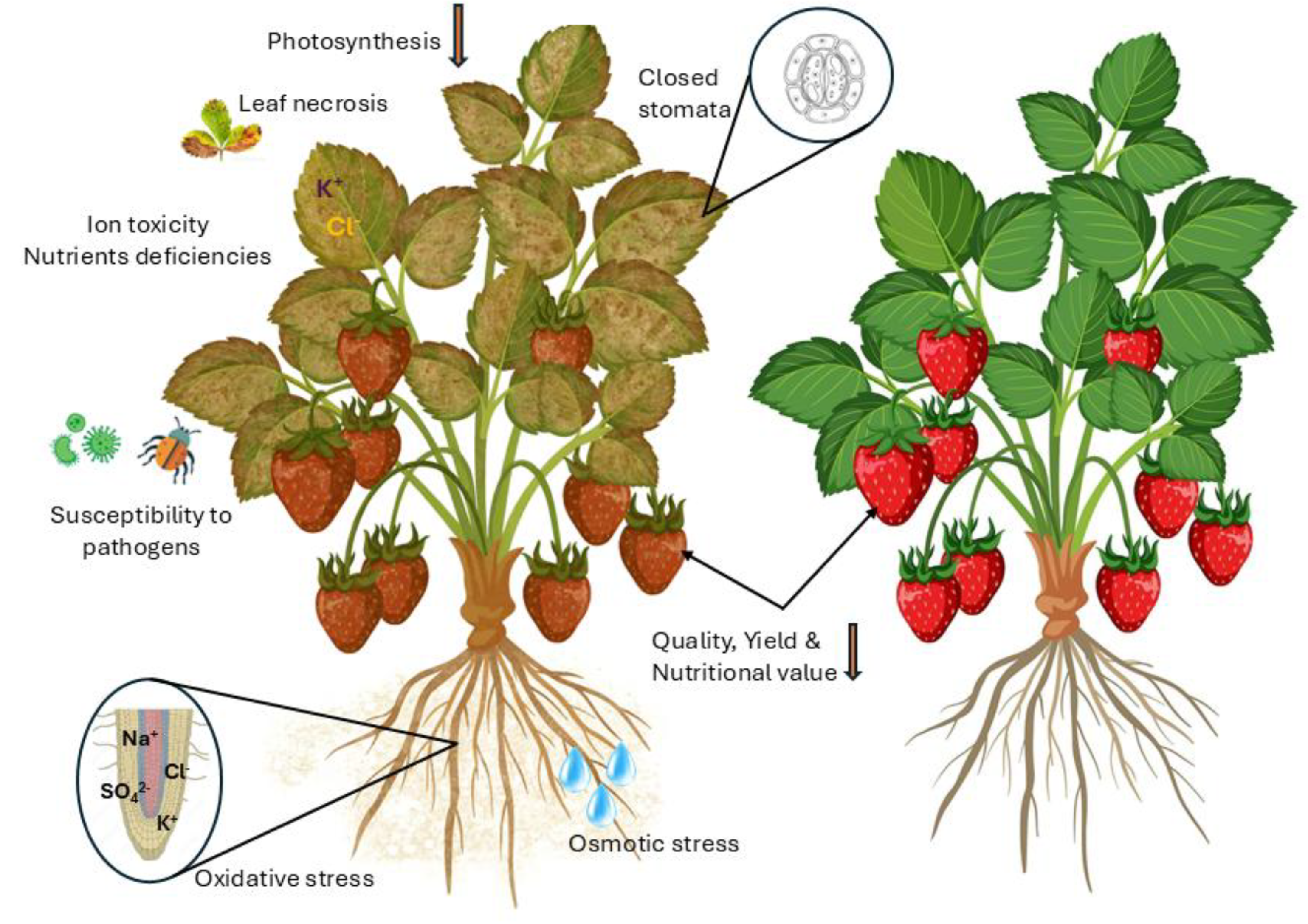
Figure 3.
Overview of the beneficial effects though the exogenous application of small signaling molecules, biostimulants, and plant–microbe interactions (AMF: Arbuscular Mycorrhizal Fungi; PGPR: Plant Growth Promoting Rhizobacteria; HRW: Hydrogen-rich Water; GABA: γ-Aminobutyric Acid) enhance salinity tolerance in strawberry.
Figure 3.
Overview of the beneficial effects though the exogenous application of small signaling molecules, biostimulants, and plant–microbe interactions (AMF: Arbuscular Mycorrhizal Fungi; PGPR: Plant Growth Promoting Rhizobacteria; HRW: Hydrogen-rich Water; GABA: γ-Aminobutyric Acid) enhance salinity tolerance in strawberry.
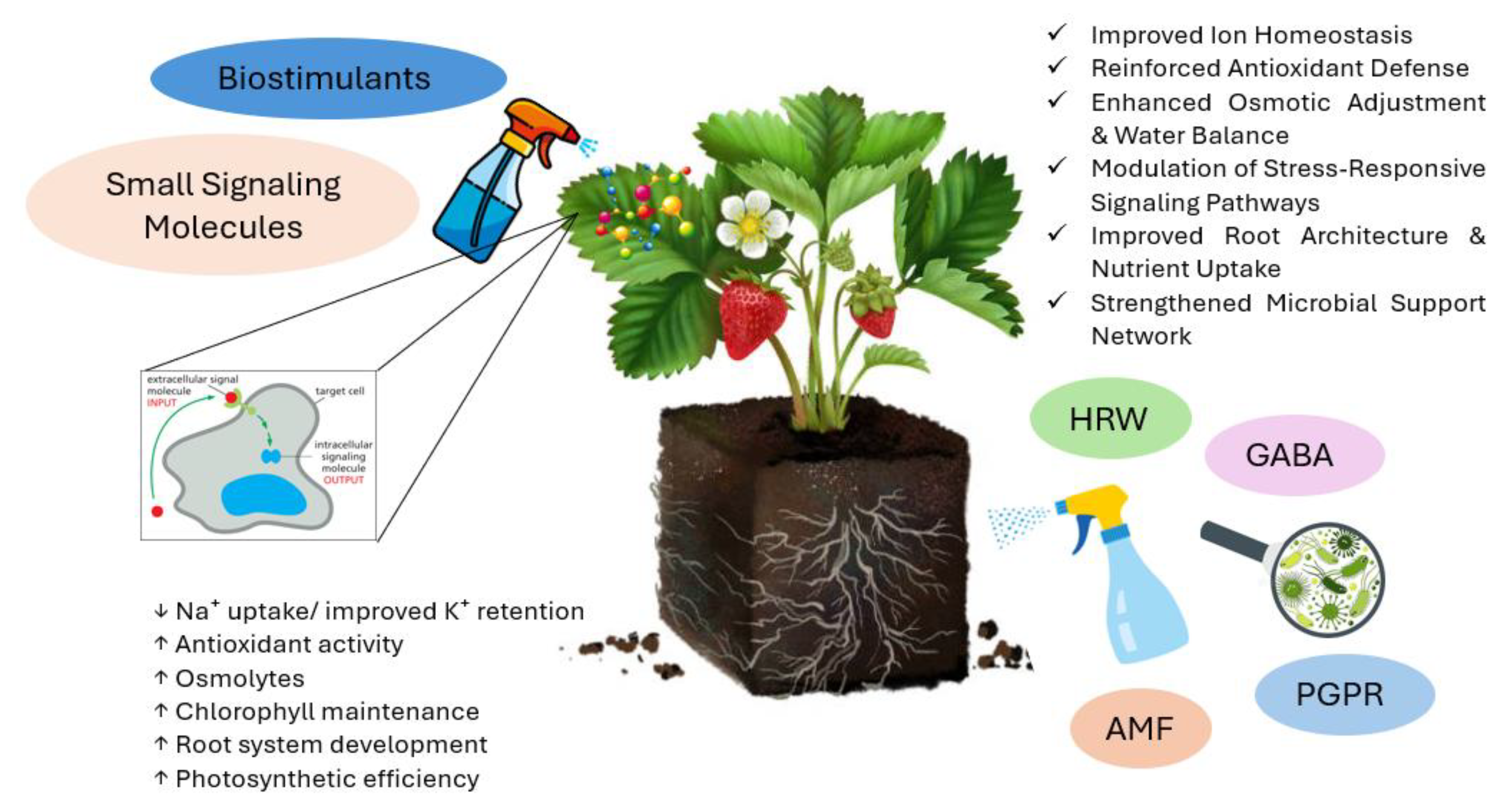
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



