Submitted:
16 December 2025
Posted:
17 December 2025
You are already at the latest version
Abstract
Slow-growing and locally invasive, chordoma is a rare malignant bone tumor that poses significant challenges because of its high recurrence rate and resistance to several standard treatment techniques. All cancers, including chordomas, have altered energy metabolism processes that contribute to their unchecked growth and survival. The significance of non-coding RNAs, particularly circular RNAs (circRNAs), as key regulators at the intersection of cellular metabolism and immune function has been highlighted by recent discoveries. By focusing on important glycolytic enzymes in tumor cells and altering metabolic reprogramming pathways, CircRNAs can influence cancer metabolic adaptability. Furthermore, via influencing immune cell functions as immunological checkpoint signaling and macrophage polarization, circRNAs influence immune evasion in the tumor microenvironment. These frequently happen via regulating important pathway signals, like PI3K/AKT/mTOR and NRF2, or by processes like miRNA sponging, creating a tumor microenvironment that is immunosuppressive and metabolically friendly. The translational pathway of circRNA-targeted therapeutics is promoted as a developing pharmacological entity in this review, which also highlights recent information on the control of circRNA-mediated immunometabolism in chordoma and examines numerous important molecular axes. There are promising opportunities to develop novel precision treatments for chordoma by considering circRNAs as dual regulators of immunological and metabolic networks.
Keywords:
immunometabolism
; chordoma
; circRNAs
; metabolic reprogramming
; immune evasion
Introduction
Chordoma: Characteristics and Pathophysiology
Chordoma is an uncommon malignant bone tumor that grows slowly and spreads locally. It starts from the remains of the basic notochord [1]. The sacrum/coccyx and the base of the skull are the most often affected areas, and it mostly affects the axial skeleton. Despite being regarded as low-grade tumors, chordomas have a dismal prognosis because of their high recurrence rates, potential for metastasis, and local aggressiveness [2]. The molecular foundations of chordoma have been further clarified by recent research, which has also highlighted the disease’s distinct cellular features and treatment difficulties. Since these embryonic remains are normally regulated for apoptosis, the notochordal origin of chordoma is crucial because they have the potential to change malignantly and cause tumor initiation [3]. This metamorphosis is frequently associated with particular genetic and epigenetic changes that promote unchecked growth and survival. Due to chordoma’s slow growth rate, diagnosis is frequently delayed, allowing the tumor to develop to a big and invasive size before being discovered. Since total surgical excision is frequently difficult, particularly in crucial anatomical regions like the base of the skull, this local invasiveness plays a significant role in its high recurrence rates [2].
Pathophysiology
Notochordal cells that continue to grow abnormally after embryonic development are recognized to be the source of chordomas, however the exact mechanisms causing them are still being investigated [3]. Normally, these cells retreat, but in chordoma, they change malignantly. Due to the tumor’s slow growth, detection is sometimes delayed, which permits it to spread far and infiltrate nearby tissues, such as soft tissue and bone (Figure 1). Depending on the location of the tumor, this local invasiveness can cause serious morbidity by compressing important structures, resulting in symptoms including pain, neurological impairments (e.g., weakness, numbness, bladder/bowel malfunction), and headaches. It is also becoming more well acknowledged that the chordoma tumor microenvironment (TME) plays a crucial role in determining the course of the tumor and the effectiveness of treatment. Immune cells, stromal cells, and extracellular matrix constituents interact intricately to create an immunosuppressive environment, which is its defining feature [4,5].
Chordomas are distinguished genetically by certain molecular changes. Consistently overexpressed in chordoma cells, brachyury is a transcription factor essential for notochord formation and is regarded as a possible therapeutic target as well as a critical diagnostic marker [6,7]. Recent studies have also discovered recurring genetic abnormalities that contribute to chordoma pathogenesis and treatment resistance, such as mutations in genes related to signaling pathways (e.g., PI3K/AKT/mTOR pathway components) and cell cycle regulation (e.g., CDKN2A) [8,9,10]. Histone changes and DNA methylation are examples of epigenetic modifications that are important in controlling gene expression and promoting the growth of chordomas [11].
Circular RNAs in Cancer Immunometabolism
Circular RNAs (circRNAs), a kind of endogenous non-coding RNA, are more stable than linear RNAs due to their covalently closed loop structure [12,13,14]. New evidence demonstrates their critical significance in various biological processes, such as carcinogenesis, tumor development, and TME modulation. Remarkably, circRNAs have a significant role in immunometabolism, the intersection of cellular metabolism and immune activity [15].
Role in Metabolic Reprogramming
Cancer cells exhibit altered metabolic pathways to support their rapid proliferation and survival, a phenomenon known as metabolic reprogramming. This metabolic plasticity allows cancer cells to adapt to fluctuating nutrient availability and hypoxic conditions within the tumor microenvironment. CircRNAs have been identified as key regulators of these metabolic adaptations, influencing various aspects of glucose, lipid, and amino acid metabolism within tumor cells [16]. For example, the Warburg effect, a metabolic shift in which cancer cells preferentially rely on glycolysis even in the presence of oxygen- is facilitated by the regulation of glycolytic enzymes, such as hexokinase 2 (HK2) and phosphofructokinase (PFK), by a variety of circRNAs [17,18]. This metabolic reprogramming gives cancer cells the building materials they need to divide and expand quickly, as well as the intermediates they need for anabolic functions. In addition to glycolysis, circRNAs also affect other metabolic processes that are essential for maintaining the high energy requirements of cancer cells in growth, such as glutaminolysis, fatty acid synthesis, and oxidative phosphorylation (Figure 2) [19].
Role in Immune Regulation
CircRNAs have a significant influence on immunological responses within the TME, which aids in immune evasion, in addition to their direct effects on tumor cell metabolism [20]. They have the ability to affect the polarization and activity of different immune cells, such as T cells, natural killer (NK) cells, and macrophages. For instance, it has been demonstrated that specific circRNAs alter macrophage polarization toward an M2-like phenotype, which is linked to angiogenesis, immunosuppression, and tumor development [21,22]. By releasing anti-inflammatory cytokines and encouraging tumor growth, these M2 macrophages help to create an immunosuppressive microenvironment. Additionally, circRNAs can control immune checkpoint signaling pathways including PD-1/PD-L1 and CTLA-4, which are important targets in cancer immunotherapy, and cause T cell depletion and malfunction, which reduces their anti-tumor efficacy (Figure 3) [23,24].
By acting as miRNA sponges, interacting with RNA-binding proteins, or directly influencing gene expression, circRNAs can orchestrate a complex network that leads to immune tolerance and evasion, thereby allowing cancer cells to escape immune surveillance and proliferate unchecked [25].
Molecular Crosstalk in Immunometabolism
The TME is shaped by the intricate and varied interactions among circRNAs, metabolism, and immunology that create a complex molecular crosstalk. CircRNAs are essential for integrating immunological responses with metabolic cues. CircRNAs, for instance, have the ability to alter important signaling pathways that are essential to immune cell function and cellular metabolism, including PI3K/AKT/mTOR and NRF2 [26]. A master regulator of cell growth, proliferation, and survival, the PI3K/AKT/mTOR pathway is frequently aberrantly activated in cancer, which affects immune cell differentiation and function and frequently results in increased glycolysis and lipid production. Likewise, NRF2 is a transcription factor that affects immune cell responses as well as tumor cell survival and is essential for metabolic adaptability and antioxidant defense. By altering these pathways, circRNAs can suppress anti-tumor immune responses in a number of ways, such as encouraging immune cell fatigue or anergy, while also giving tumor cells a metabolically favorable environment that gives them plenty of energy and building blocks [27]. CircRNAs’ dual regulatory ability makes them important participants in the immunometabolic reprogramming of cancer, providing fresh perspectives on the pathophysiology of the disease and possible treatment approaches. This is especially true for chordoma, where complex interactions are likely to have a significant impact on the course of the disease and resistance to treatment.
Specific circRNA Mechanisms in Chordoma: Bridging to Immunometabolism
Although the topic of circRNAs’ participation in chordoma’s immunometabolic reprogramming is still in its infancy, current research has started to clarify which circRNAs are implicated in the evolution of chordoma and may provide insights into their wider immunometabolic effects. Compared to more prevalent tumors like breast cancer, research directly connecting circRNAs to chordoma’s immunometabolism is still in its infancy because of the tumor’s rarity and distinctive features. However, there is a solid foundation for speculating about the activities of circRNA in chordoma based on our understanding of how it acts in other malignancies. CircRNAs may be especially important in mediating metabolic adaptations and immune evasion tactics peculiar to chordoma, given its distinct microenvironment, which is marked by its sluggish development, local invasiveness, and resistance to standard therapy [28].
The discovery of circTEAD1 as a major oncogenic driver in chordoma is the most important development in this field. By stabilizing Yap1 mRNA, our work showed that circTEAD1 is elevated in chordomas and stimulates carcinogenesis. This is accomplished by a m6A-dependent mechanism in which circTEAD1’s cytoplasmic export and the creation of a circTEAD1/IGF2BP3/Yap1 mRNA complex are facilitated by METTL3-mediated m6A alteration of circTEAD1. This discovery is significant because it establishes the first direct connection between a particular circRNA and the Hippo/Yap1 oncogenic pathway in chordoma. This makes it possible to study other circRNAs with comparable mechanisms and offers a tangible illustration of how circRNAs may work in this illness [29].
Potential Immunometabolic Roles
Given the known functions of circRNAs in the immunometabolism of general cancer, it is conceivable that the circRNAs found in chordoma—or others that have not yet been found—also play a part in the disease’s distinct immunometabolic environment. If a circRNA, for instance, stimulates chordoma cell proliferation, it probably does so by affecting metabolic pathways that supply the energy and building blocks required for quick growth. The metabolic status of immune cells within the tumor may also be impacted directly or indirectly by circRNAs that alter the TME in chordoma, such as those that influence immunological checkpoint expression (PD-1, CTLA-4, for example, are enhanced in chordoma) [30].
CircRNAs may play a role in metabolic adaptations and immune evasion. CircRNAs may further give chordoma cells metabolic flexibility in their hypoxic and nutritionally relaxed TME by controlling important enzymes or transporters involved in glucose, glutamine, or lipid metabolism [31]. Furthermore, CircRNAs will alter the actions of immune cells found in the chordoma milieu, such as T cells or macrophages: Their roles may directly impact immunological signaling pathways that result in T cell fatigue or anergy, or they may have an impact on metabolic alterations (e.g., direct macrophage metabolic changes to an M2-like phenotypic shift). One of the main ways that circRNAs coordinate the immunometabolic reprogramming in chordoma may be through their interaction with signaling pathways that are essential for both metabolism and immunity, such as PI3K/AKT/mTOR or NRF2 (Table 1) [32]. To clarify these particular chemical axes in the context of chordoma, more investigation is required.
It is essential to comprehend these particular circRNA-mediated processes in chordoma’s immunometabolism in order to find new diagnostic biomarkers and create focused treatment plans for this difficult cancer.
Therapeutic Strategies Targeting circRNAs in Chordoma
Given the emerging role of circRNAs in chordoma, several therapeutic strategies can be envisioned. These strategies can be broadly categorized into direct targeting of circRNAs and indirect targeting through the pathways they regulate [33].
Direct Targeting of circRNAs
Directly targeting oncogenic circRNAs represents a promising therapeutic avenue. This can be achieved through various approaches, such as antisense oligonucleotides (ASOs) or small interfering RNAs (siRNAs) designed to bind to the back-splice junction of the target circRNA, leading to its degradation [15,34,35]. For example, an ASO targeting circTEAD1 could be developed to reduce its expression in chordoma cells, thereby inhibiting its oncogenic functions. Another approach is to use small molecules that can bind to and disrupt the function of circRNAs or their protein complexes [36].
Indirect Targeting of circRNA-Regulated Pathways
Indirectly targeting the pathways regulated by circRNAs offers another therapeutic strategy. For example, since circTEAD1 promotes chordoma tumorigenesis by stabilizing Yap1 mRNA, inhibitors of the Yap1/Hippo pathway could be used to counteract the effects of circTEAD1. Similarly, if a circRNA is found to promote glycolysis, inhibitors of glycolytic enzymes could be used in combination with circRNA-targeted therapies (Table 2) [37].
Molecular Crosstalk and Combination Therapies
Because circRNAs, metabolism, and immunology have intricate molecular relationships, combination therapy may be particularly effective. For example, circRNA-targeted therapies may be used in combination with immune checkpoint inhibition or metabolic inhibitors to stop tumor development and overcome resistance mechanisms [47]. Further research is needed to identify the key circRNA-mediated molecular axis in chordoma immunometabolism and to develop effective delivery systems for circRNA-based therapies.
The discovery that circRNAs in chordoma function as both immunological and metabolic network regulators opens up new possibilities for the development of targeted and innovative treatment strategies, which will eventually enhance patient outcomes for this challenging disease.
Comparative Analysis of Chordoma Molecular and Immunological Studies
A comparison of recent important research that has improved our knowledge of the molecular and immunological landscape of chordoma is given in this section. Contextualizing the function of circRNAs in chordoma immunometabolism requires an understanding of the results presented in these studies (Table 3).
Synthesis and Comparative Insights
The collective findings from these studies paint a picture of chordoma as a molecularly heterogeneous and immunologically complex cancer. The 2015 review by Sun et al. laid the groundwork by establishing the importance of molecular drivers like brachyury and the involvement of non-coding RNAs, specifically microRNAs [7]. This sets the stage for investigating other non-coding RNAs like circRNAs.
The more recent papers build upon this foundation by revealing deeper layers of complexity. The work by Bai et al. (2023) and Zhang et al. (2024) independently converges on the idea of distinct molecular subtypes of chordoma [8,25]. While Bai et al. used gene expression to define subtypes based on chromatin remodeling and developmental pathways, Zhang et al. used proteogenomics to identify subtypes characterized by chromosomal instability and an ‘immune cold’ phenotype. This convergence from different ‘omics’ approaches strongly suggests that these subtypes are biologically meaningful and clinically relevant.
The most critical paper for this review is the 2024 study by Li et al., which is the first to directly implicate a specific circRNA, circTEAD1, in chordoma tumorigenesis [18]. This study is a cornerstone because it not only provides the first direct evidence of circRNA involvement but also links it to a key cancer-related pathway (Hippo/Yap1) and a crucial regulatory mechanism (m6A modification). This provides a mechanistic framework for understanding how circRNAs could contribute to the molecular subtypes identified by Bai et al. and Zhang et al.
Finally, the review by Chen and Zhang (2024) on the immune microenvironment provides the immunological context [5]. It highlights the immunosuppressive nature of the chordoma TME, which is dominated by M2 macrophages and regulatory T cells. This is consistent with the ‘immune cold’ subtype identified by Zhang et al. and provides a potential link between the molecular mechanisms driven by circRNAs and the resulting immune landscape. For example, circRNAs could be involved in the recruitment or polarization of these immunosuppressive immune cells.
In conclusion, the progression from general molecular characterization to specific circRNA functions and detailed immune profiling demonstrates a growing understanding of chordoma. The next frontier is to integrate these different facets – to understand how specific circRNAs like circTEAD1 contribute to the different molecular subtypes and how they shape the immune microenvironment to promote tumor growth and immune evasion. This review paper is positioned to do exactly that, by synthesizing these findings and proposing a model for circRNA-mediated immunometabolic reprogramming in chordoma.
circRNA-Based Vaccines
Another exciting possibility is the use of circRNAs as cancer vaccines. CircRNAs can be engineered to encode tumor antigens, and their high stability makes them attractive candidates for vaccine development [41]. Furthermore, recent advances in artificial intelligence can be leveraged to design and optimize circRNA-based vaccines for enhanced immunogenicity and efficacy [42].
Challenges and Future Perspectives
Despite the promise of circRNA-targeted therapies, several challenges need to be addressed. These include the efficient and specific delivery of therapeutic agents to tumor cells, potential off-target effects, and the development of resistance mechanisms. Future research should focus on developing novel delivery systems, identifying biomarkers to predict treatment response, and exploring combination therapies to overcome resistance.
CircRNA research is at the vanguard of chordoma therapy development due to convergent evidence from current molecular, immunological, and mechanistic investigations; nevertheless, to fully exploit this promise, several crucial research objectives need to be addressed. Building on the groundbreaking work with circTEAD1 to create a comprehensive regulatory network, it is imperative to conduct thorough circRNA profiling across chordoma molecular subtypes to find subtype-specific circRNA signatures that may direct individualized treatment approaches. Second, to mechanistically validate the immunometabolic roles of candidate circRNAs, specifically their impact on T cell exhaustion and macrophage polarization within the distinct chordoma microenvironment, functional validation studies employing sophisticated chordoma organoid models and patient-derived xenografts are crucial. Third, a crucial translation barrier that needs to be addressed before circRNA-targeted treatments can be used in clinical settings is the creation of novel delivery systems, such as lipid nanoparticles or viral vectors that are especially tailored for the anatomically difficult sites where chordomas develop. Fourth, combining machine learning and artificial intelligence techniques may speed up the discovery of circRNA-based biomarkers and forecast treatment outcomes, opening the door to precision medicine treatments for this uncommon but deadly illness. Lastly, to ensure reproducibility and make it easier to convert encouraging preclinical results into clinical trials, cooperative international efforts are required to develop standardized techniques for circRNA identification, quantification, and functional analysis in chordoma. The ultimate objective is to turn our expanding knowledge of circRNA-mediated immunometabolic reprogramming into efficient therapeutic approaches that can help chordoma patients, who presently have few treatment options and dismal prognoses despite advancements in radiation and surgery.
Conclusions
A dangerous bone tumor called chordoma necessitates urgent medical attention. Understanding circular RNAs (circRNAs) as master regulators of immune responses and cancer metabolism has recently advanced, opening up new therapeutic paths and offering insight into how chordomas could form. This review discusses the multiple roles played by circRNAs in directing the immunometabolic reprogramming of the chordoma tumor microenvironment to effect metabolic changes driving tumor growth that lead to immune evasion.
Research on the relationship between chordoma and circRNA immunometabolism is still in its early stages, but evidence from other cancers, particularly when combined with the recently published preliminary findings from chordoma, strongly suggests that non-coding RNAs will play a significant role in these systems. Because circRNAs can modulate proteins, act as miRNA sponges, and interfere with signaling pathways like PI3K/AKT/mTOR and NRF2, they will be at the center of a complex network that involves immune cells, tumor cells, and their metabolic environment.
Directly or indirectly targeting circRNAs offers chordoma patients promising treatment options. Future directions in precision medicine may involve repairing tumor-suppressive circRNAs or preventing oncogenic circRNAs. Furthermore, this strategy may offer a promising way to enhance anti-tumor immunity or reverse metabolic reprogramming. The future of chordoma treatment may lie in circRNA-targeting combination approaches with new or established metabolic and immunotherapeutic approaches. The conversion of these innovative concepts into effective clinical treatments that will significantly enhance patients’ prognoses and quality of life requires further investigation into particular circRNA-mediated molecular axes in chordoma immunometabolism.
Conflicts of Interest
The author declares that there is no conflict of interest.
References
- Walcott BP, Nahed B V., Mohyeldin A, Coumans JV, Kahle KT, Ferreira MJ. Chordoma: Current concepts, management, and future directions. Lancet Oncol 2012;13. [CrossRef]
- Pennington Z, Ehresman J, McCarthy EF, Ahmed AK, Pittman PD, Lubelski D, et al. Chordoma of the sacrum and mobile spine: a narrative review. Spine Journal 2021;21:500–17. [CrossRef]
- Salisbury JR. The pathology of the human notochord. J Pathol 1993;171:253–5. [CrossRef]
- Niu HQ, Zheng BY, Zou MX, Zheng BW. Complex immune microenvironment of chordoma: a road map for future treatment. J Immunother Cancer 2024;12. [CrossRef]
- Chen Y, Zhang H. Immune microenvironment and immunotherapy for chordoma. Front Oncol 2024;14. [CrossRef]
- Yang XR, Ng D, Alcorta DA, Liebsch NJ, Sheridan E, Li S, et al. T (brachyury) gene duplication confers major susceptibility to familial chordoma. Nat Genet 2009;41:1176–8. [CrossRef]
- Sun X, Hornicek F, Schwab JH. Chordoma: an update on the pathophysiology and molecular mechanisms. Curr Rev Musculoskelet Med 2015;8:344. [CrossRef]
- Bai J, Shi J, Zhang Y, Li C, Xiong Y, Koka H, et al. Gene Expression Profiling Identifies Two Chordoma Subtypes Associated with Distinct Molecular Mechanisms and Clinical Outcomes. Clin Cancer Res 2023;29:261–70. [CrossRef]
- Presneau, Shalaby A, Idowu B, Gikas P, Cannon SR, Gout I, et al. Potential therapeutic targets for chordoma: PI3K/AKT/TSC1/TSC2/mTOR pathway. Br J Cancer 2009;100:1406–14. [CrossRef]
- Tarpey PS, Smith R, Pleasance E, Whibley A, Edkins S, Hardy C, et al. A systematic, large-scale resequencing screen of X-chromosome coding exons in mental retardation. Nat Genet 2009;41:535–43. [CrossRef]
- Yu X, Li Z. Epigenetic deregulations in chordoma. Cell Prolif 2015;48:497–502. [CrossRef]
- Conn VM, Chinnaiyan AM, Conn SJ. Circular RNA in cancer. Nat Rev Cancer 2024;24:597–613. [CrossRef]
- Zheng Y, Ren S, Zhang Y, Liu S, Meng L, Liu F, et al. Circular RNA circWWC3 augments breast cancer progression through promoting M2 macrophage polarization and tumor immune escape via regulating the expression and secretion of IL-4. Cancer Cell Int 2022;22. [CrossRef]
- Kristensen LS, Andersen MS, Stagsted LVW, Ebbesen KK, Hansen TB, Kjems J. The biogenesis, biology and characterization of circular RNAs. Nat Rev Genet 2019;20:675–91. [CrossRef]
- Kristensen LS, Jakobsen T, Hager H, Kjems J. The emerging roles of circRNAs in cancer and oncology. Nat Rev Clin Oncol 2022;19:188–206. [CrossRef]
- Yu T, Wang Y, Fan Y, Fang N, Wang T, Xu T, et al. CircRNAs in cancer metabolism: a review. J Hematol Oncol 2019;12. [CrossRef]
- Ji X, Sun W, Lv C, Huang J, Zhang H. Circular RNAs Regulate Glucose Metabolism in Cancer Cells. Onco Targets Ther 2021;14:4005. [CrossRef]
- Li T, Xian HC, Dai L, Tang YL, Liang XH. Tip of the Iceberg: Roles of CircRNAs in Cancer Glycolysis. Onco Targets Ther 2021;14:2379. [CrossRef]
- Zhou WY, Cai ZR, Liu J, Wang DS, Ju HQ, Xu RH. Circular RNA: metabolism, functions and interactions with proteins. Mol Cancer 2020;19:1–19. [CrossRef]
- Liu T, Long K, Zhu Z, Song Y, Chen C, Xu G, et al. Roles of circRNAs in regulating the tumor microenvironment. Med Oncol 2023;40:329. [CrossRef]
- Gonzalez C, Cimini M, Cheng Z, Benedict C, Wang C, Trungcao M, et al. Role of circular RNA cdr1as in modulation of macrophage phenotype. Life Sci 2022;309:121003. [CrossRef]
- He L, Tam PKH, Deng CX. Orchestration of Tumor-Associated Macrophages in the Tumor Cell-Macrophage-CD8+ T Cell Loop for Cancer Immunotherapy. Int J Biol Sci 2025;21:4098–116. [CrossRef]
- Carlos-Reyes Á, Romero-Garcia S, Contreras-Sanzón E, Ruiz V, Prado-Garcia H. Role of Circular RNAs in the Regulation of Immune Cells in Response to Cancer Therapies. Front Genet 2022;13:823238. [CrossRef]
- Jiang W, Pan S, Chen X, Wang Z wei, Zhu X. The role of lncRNAs and circRNAs in the PD-1/PD-L1 pathway in cancer immunotherapy. Mol Cancer 2021;20:1–17. [CrossRef]
- Zhang W, Xu C, Yang Z, Zhou J, Peng W, Zhang X, et al. Circular RNAs in tumor immunity and immunotherapy. Mol Cancer 2024;23:1–15. [CrossRef]
- Geng Y, Wang M, Wu Z, Jia J, Yang T, Yu L. Research progress of circRNA in malignant tumour metabolic reprogramming. RNA Biol 2023;20:641. [CrossRef]
- Wang Y, Cui Y, Li X, Jin SH, Wang H, Gaipl US, et al. CircRNAs: functions and emerging roles in cancer and immunotherapy. BMC Med 2025;23:1–18. [CrossRef]
- Ghosh PK, Ghosh A. Dysregulation of noncoding RNA in chordoma; implications in identifying potential targets for novel therapeutic approaches. Mol Biol Rep 2024;51. [CrossRef]
- Li H, Tang Y, Ruan X, Zhang J, Liu H, Yu S, et al. N6-methyladenosine-modified circTEAD1 stabilizes Yap1 mRNA to promote chordoma tumorigenesis. Clin Transl Med 2024;14. [CrossRef]
- Niu HQ, Zheng BY, Zou MX, Zheng BW. Complex immune microenvironment of chordoma: a road map for future treatment. J Immunother Cancer 2024;12. [CrossRef]
- Mammar H, Polivka M, Belkacemi Y, Lot G, Froelich S, Carpentier A, et al. Hypoxia and Metabolism Regulation in Chordomas: Correlation Between Biology and Clinical Features for Potential Targeted Therapy. International Journal of Radiation Oncology*Biology*Physics 2015;93:E89–90. [CrossRef]
- Xu J, Shi Q, Wang B, Ji T, Guo W, Ren T, et al. The role of tumor immune microenvironment in chordoma: promising immunotherapy strategies. Front Immunol 2023;14:1257254. [CrossRef]
- Barber SM, Sadrameli SS, Lee JJ, Fridley JS, Teh BS, Oyelese AA, et al. Chordoma-Current Understanding and Modern Treatment Paradigms. J Clin Med 2021;10:1–18. [CrossRef]
- Jeoung J, Kim W, Jo H, Jeoung D. Circular RNAs as Targets for Developing Anticancer Therapeutics. Cells 2025, Vol 14, Page 1106 2025;14:1106. [CrossRef]
- Yu T, Wang Y, Fan Y, Fang N, Wang T, Xu T, et al. CircRNAs in cancer metabolism: A review. J Hematol Oncol 2019;12:1–10. [CrossRef]
- He AT, Liu J, Li F, Yang BB. Targeting circular RNAs as a therapeutic approach: current strategies and challenges. Signal Transduct Target Ther 2021;6:1–14. [CrossRef]
- Han C, Seebacher NA, Hornicek FJ, Kan Q, Duan Z, Han C, et al. Regulation of microRNAs function by circular RNAs in human cancer. Oncotarget 2017;8:64622–37. [CrossRef]
- Koch L. CRISPR–Cas13 targets circRNAs. Nat Rev Genet 2021;22:68. [CrossRef]
- He AT, Liu J, Li F, Yang BB. Targeting circular RNAs as a therapeutic approach: current strategies and challenges. Signal Transduction and Targeted Therapy 2021 6:1 2021;6:185-. [CrossRef]
- Dawoud A, Elmasri RA, Mohamed AH, Mahmoud A, Rostom MM, Youness RA. Involvement of CircRNAs in regulating The “New Generation of Cancer Hallmarks”: A Special Depiction on Hepatocellular Carcinoma. Crit Rev Oncol Hematol 2024;196:104312. [CrossRef]
- Gong Z, Hu W, Zhou C, Guo J, Yang L, Wang B. Recent advances and perspectives on the development of circular RNA cancer vaccines. NPJ Vaccines 2025;10. [CrossRef]
- Zhao Y, Wang H. Artificial intelligence-driven circRNA vaccine development: multimodal collaborative optimization and a new paradigm for biomedical applications. Brief Bioinform 2025;26. [CrossRef]
- Jiang W, Pan S, Chen X, Wang Z wei, Zhu X. The role of lncRNAs and circRNAs in the PD-1/PD-L1 pathway in cancer immunotherapy. Molecular Cancer 2021 20:1 2021;20:116-. [CrossRef]
- Allegra A, Cicero N, Tonacci A, Musolino C, Gangemi S. Circular RNA as a Novel Biomarker for Diagnosis and Prognosis and Potential Therapeutic Targets in Multiple Myeloma. Cancers (Basel) 2022;14. [CrossRef]
- Piras R, Ko EY, Barrett C, De Simone M, Lin X, Broz MT, et al. circCsnk1g3- and circAnkib1-regulated interferon responses in sarcoma promote tumorigenesis by shaping the immune microenvironment. Nature Communications 2022 13:1 2022;13:7243-. [CrossRef]
- Wei CY, Zhu MX, Lu NH, Liu JQ, Yang YW, Zhang Y, et al. Circular RNA circ_0020710 drives tumor progression and immune evasion by regulating the miR-370-3p/CXCL12 axis in melanoma. Mol Cancer 2020;19. [CrossRef]
- Dang Q, Li B, Jin B, Ye Z, Lou X, Wang T, et al. Cancer immunometabolism: advent, challenges, and perspective. Mol Cancer 2024;23:1–22. [CrossRef]
- Das U, Banerjee S, Sarkar M, Muhammad L F, Soni TK, Saha M, et al. Circular RNA vaccines: Pioneering the next-gen cancer immunotherapy. Cancer Pathogenesis and Therapy 2025;3:309–21. [CrossRef]
Figure 1.
An illustration of chordoma pathophysiology, This figure illustrates the key aspects of chordoma development and its clinical features. A) The process begins with persistent notochordal remnants from embryonic development that undergo malignant transformation into chordoma. B) Histologically, the tumor is characterized by physaliferous cells with vacuolated cytoplasm and shows aggressive local invasion into bone and soft tissue. C) Chordomas are primarily located in the skull base (35%), mobile spine (15%), and sacrococcygeal region (50%), and are defined by the overexpression of the transcription factor brachyury, a key diagnostic and diagnostic marker.
Figure 1.
An illustration of chordoma pathophysiology, This figure illustrates the key aspects of chordoma development and its clinical features. A) The process begins with persistent notochordal remnants from embryonic development that undergo malignant transformation into chordoma. B) Histologically, the tumor is characterized by physaliferous cells with vacuolated cytoplasm and shows aggressive local invasion into bone and soft tissue. C) Chordomas are primarily located in the skull base (35%), mobile spine (15%), and sacrococcygeal region (50%), and are defined by the overexpression of the transcription factor brachyury, a key diagnostic and diagnostic marker.
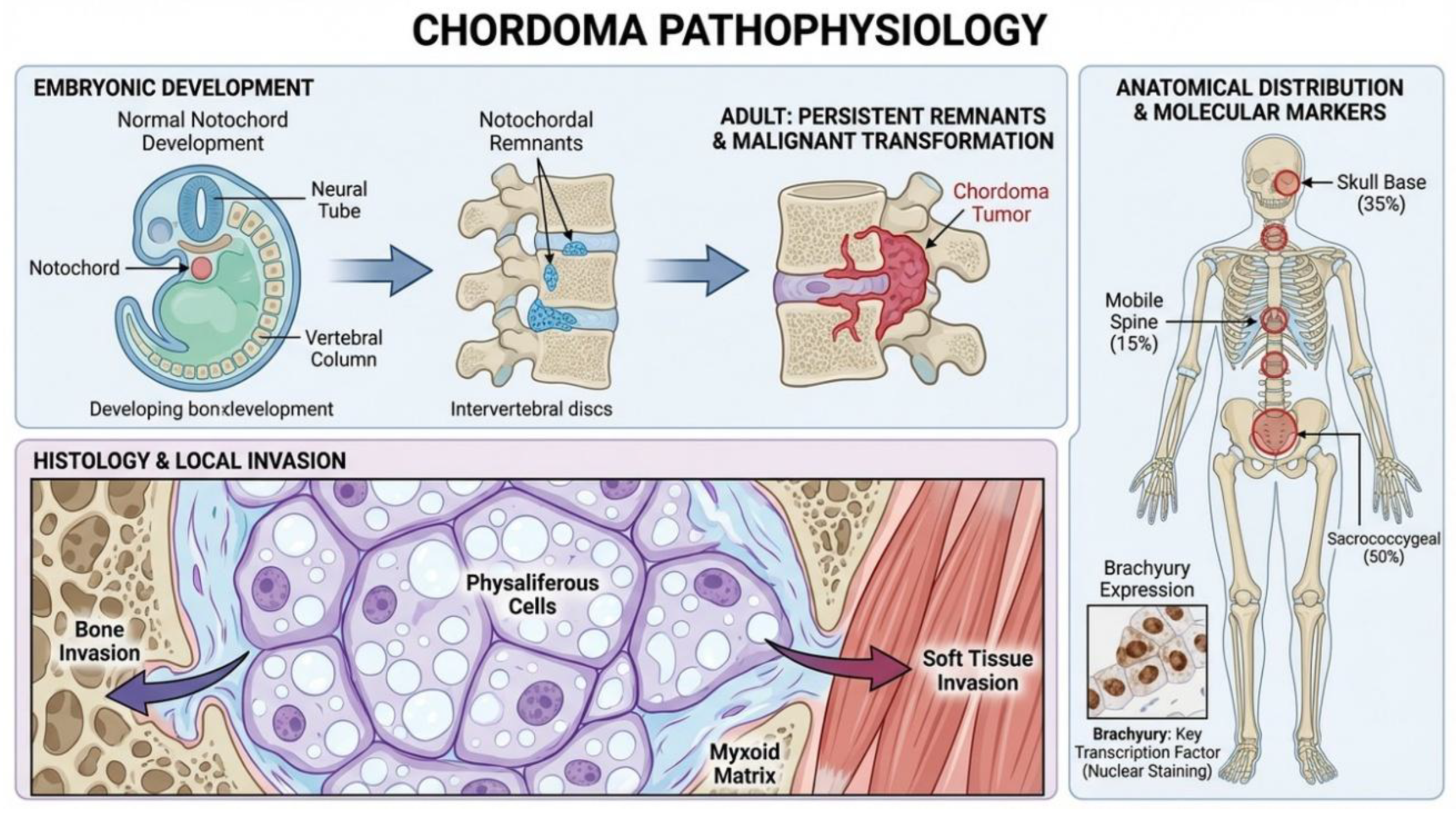
Figure 2.
A detailed diagram illustrating the key metabolic pathways, such as glycolysis, oxidative phosphorylation, and glutaminolysis, that are often reprogrammed in chordoma cells. Show the inputs, outputs, and key enzymes involved in each pathway, and highlight how they contribute to tumor growth and survival. PDH, Pyruvate dehydrogenase; GLS, Glutaminas; TCA cycle, Tricarboxylic acid cycle.
Figure 2.
A detailed diagram illustrating the key metabolic pathways, such as glycolysis, oxidative phosphorylation, and glutaminolysis, that are often reprogrammed in chordoma cells. Show the inputs, outputs, and key enzymes involved in each pathway, and highlight how they contribute to tumor growth and survival. PDH, Pyruvate dehydrogenase; GLS, Glutaminas; TCA cycle, Tricarboxylic acid cycle.
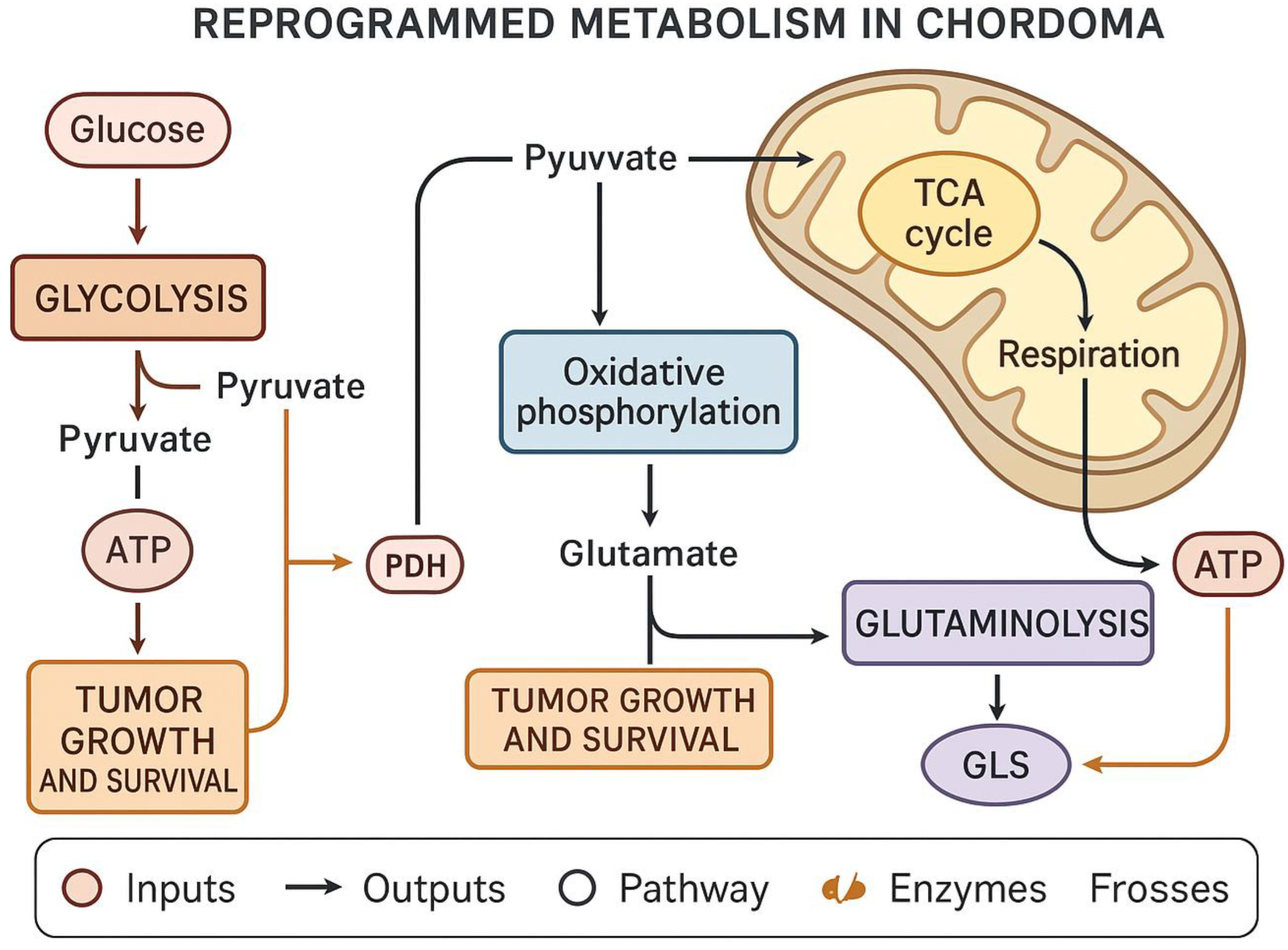
Figure 3.
A diagram illustrating how circRNAs modulate immune responses within the tumor microenvironment, contributing to immune evasion. circRNA, Circular RNA; PD-1, Programmed Cell Death Protein 1; PD-L1, Programmed Cell Death Ligand 1; CTLA-4, Cytotoxic T-Lymphocyte–Associated Protein 4; NF-κB, Nuclear Factor kappa-light-chain-enhancer of activated B cells; MAPK, Mitogen-Activated Protein Kinase; PI3K, Phosphoinositide 3-Kinase; AKT, Protein Kinase B.
Figure 3.
A diagram illustrating how circRNAs modulate immune responses within the tumor microenvironment, contributing to immune evasion. circRNA, Circular RNA; PD-1, Programmed Cell Death Protein 1; PD-L1, Programmed Cell Death Ligand 1; CTLA-4, Cytotoxic T-Lymphocyte–Associated Protein 4; NF-κB, Nuclear Factor kappa-light-chain-enhancer of activated B cells; MAPK, Mitogen-Activated Protein Kinase; PI3K, Phosphoinositide 3-Kinase; AKT, Protein Kinase B.
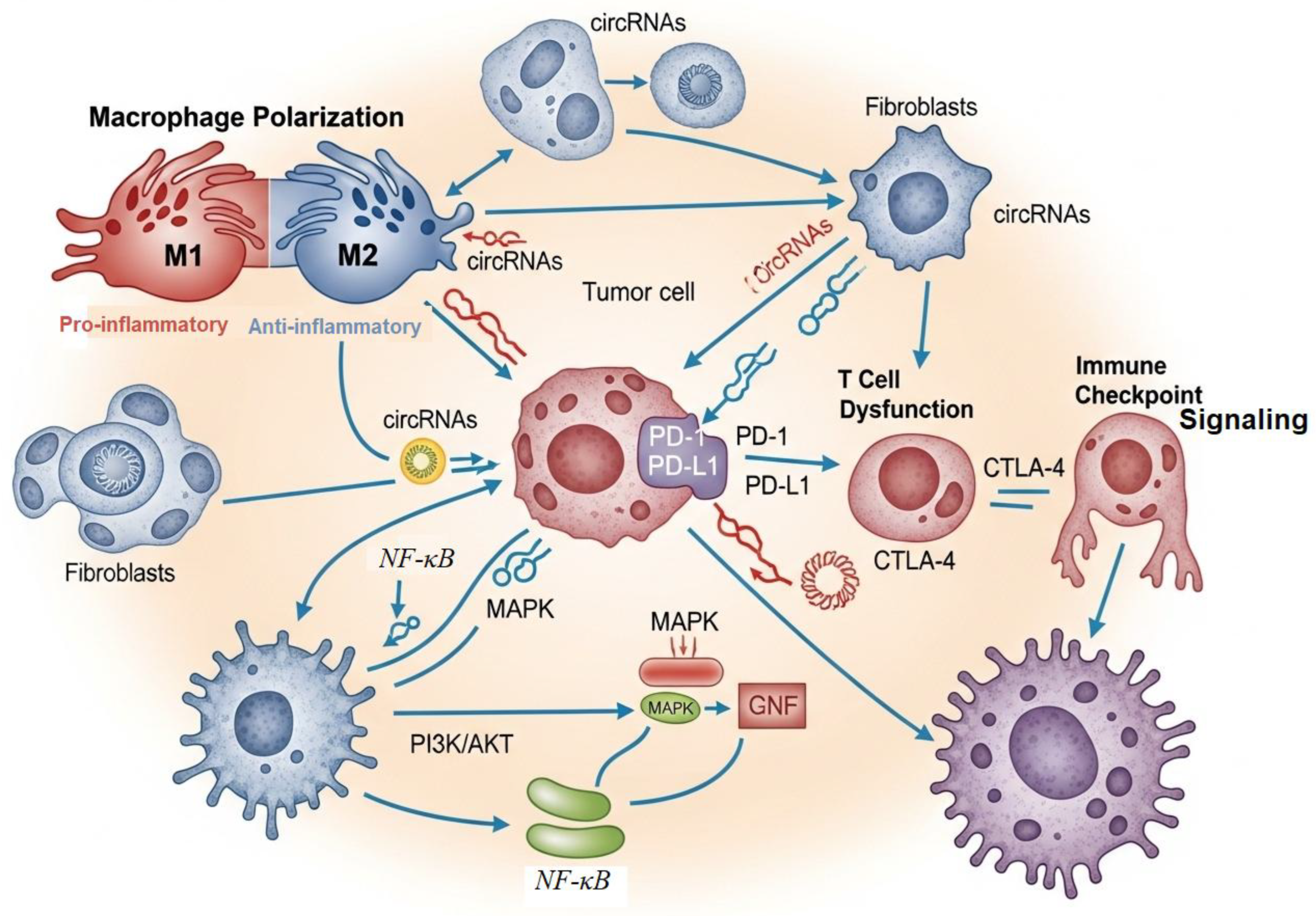

Table 1.
Immunometabolic Pathways and Their Impact in Chordoma.
| Pathway | Role in Chordoma | Associated CircRNAs | Therapeutic Implications |
|---|---|---|---|
| PI3K/AKT/mTOR | Promotes cell growth, proliferation, and survival; involved in metabolic reprogramming | circRNAs modulating PI3K/AKT/mTOR pathway | Targeting circRNAs to inhibit pathway activity and reduce tumor growth |
| NRF2 | Regulates antioxidant response and metabolic adaptation | circRNAs modulating NRF2 pathway | Targeting circRNAs to enhance oxidative stress and improve therapeutic response |
| Glycolysis (Warburg Effect) | Provides rapid ATP production and metabolic intermediates for biosynthesis | circRNAs regulating glycolytic enzymes | Targeting circRNAs to reverse metabolic reprogramming and sensitize to therapy |
| Immune Checkpoint Signaling | Contributes to immune evasion by suppressing anti-tumor immune responses | circRNAs modulating immune checkpoint molecules (e.g., PD-1, CTLA-4) | Targeting circRNAs to enhance anti-tumor immunity and improve immunotherapy efficacy |
Table 2.
Therapeutic Strategies Targeting CircRNAs in Chordoma.
| Strategy | Mechanism | Examples/Approach | Potential Benefits | Kaynak |
|---|---|---|---|---|
| Inhibition of oncogenic circRNAs | Degradation or blocking of circRNA function | Antisense oligonucleotides (ASOs), siRNAs, CRISPR-based technologies | Suppress tumor growth, reduce proliferation, overcome drug resistance | [34,38,39,40] |
| Restoration of tumor-suppressive circRNAs | Re-expression or upregulation of circRNA | Gene therapy, small molecules | Inhibit tumor growth, promote apoptosis, enhance chemosensitivity | [41,42] |
| Indirect targeting via immunometabolism | Modulating circRNAs that influence metabolic pathways or immune checkpoints | Targeting circRNAs regulating glycolytic enzymes, macrophage polarization, immune checkpoint molecules | Reverse metabolic reprogramming, enhance anti-tumor immunity, improve immunotherapy efficacy | [43,44,45,46] |
| Combination therapies | Combining circRNA-targeted therapies with other treatments | CircRNA inhibition + metabolic inhibitors, CircRNA inhibition + immune checkpoint blockade | Synergistic anti-tumor effects, overcome resistance mechanisms | [47,48] |
Table 3.
Summary of Key Research Findings.
| Paper Title | Year | Key Focus | Main Findings | Relevance to circRNA Review |
|---|---|---|---|---|
| Chordoma: an update on pathophysiology and molecular mechanisms | 2015 | Pathophysiology, molecular mechanisms | - Brachyury as a key driver- Role of chromosomal alterations, DNA methylation, and microRNAs | Foundational understanding of chordoma molecular biology, including non-coding RNAs. |
| Gene Expression Profiling Identifies Two Chordoma Subtypes Associated with Distinct Molecular Mechanisms and Clinical Outcomes | 2023 | Molecular subtyping, clinical outcomes | - Identified two subtypes with distinct molecular mechanisms (chromatin remodeling vs. EMT/Hedgehog pathways)- Subtypes correlate with clinical outcomes | Highlights molecular heterogeneity and the importance of RNA-based classification. |
| N6-methyladenosine-modified circTEAD1 stabilizes Yap1 mRNA to promote chordoma tumorigenesis | 2024 | circRNA function in chordoma | - First study on circRNAs in chordoma- circTEAD1 is upregulated and promotes tumorigenesis- m6A modification is crucial for circTEAD1 function- circTEAD1 stabilizes Yap1 mRNA via an RNA-protein complex | Core paper for the review, providing direct evidence of circRNA involvement in chordoma. |
| Immune microenvironment and immunotherapy for chordoma | 2024 | Immune microenvironment, immunotherapy | - Chordoma TME is characterized by high infiltration of M2 macrophages and regulatory T cells- Reviews current immunotherapy trials and their limitations | Provides the immunological context for the review, highlighting the immunosuppressive microenvironment that circRNAs may influence. |
| Proteogenomic characterization of skull-base chordoma | 2024 | Proteogenomics, chromosome instability | - Chromosome instability is a key prognostic factor- Identified immune cold subtype linked to chromosome 9p/10q loss- Proteomics-based classification reveals subtypes with high CIN and immune cold features | Reinforces the concept of molecular subtypes and links genomic instability to the immune landscape. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



