
Submitted:
15 December 2025
Posted:
16 December 2025
You are already at the latest version
Abstract
This study evaluated the fermentative potential of eight industrial strains of Saccharomyces cerevisiae for producing mead from honeys originating from the Caatinga Biome in the semi-arid region of Pernambuco, Brazil. The fermentations revealed preferential consumption of glucose and fructose, with limited utilization of apparent sucrose. Despite presenting similar ethanol yields (38% mass conversion), the strains differed in fermentation rate, residual sugar profile, and metabolic composition of the final products. Strains TR313 and JP1 were selected for more detailed analyses, with JP1 standing out for its higher volumetric productivity (0.23 g·L⁻¹·h⁻¹) and shorter fermentation time. The mead produced by JP1 exhibited a higher content of amino acids, particularly alanine and proline, as well as bioactive compounds such as dehydroascorbic acid, glycerate, and GABA, which may confer antioxidant and nutraceutical properties. Analysis of volatile compounds indicated a predominance of 2-phenylethanol and tyrosol, with floral and fruity notes intensified by the presence of anisic acids and alcohols. Technological tests demonstrated that increasing biomass, supplementing with nitrogen, or cell immobilization accelerates fermentation without compromising yield. Thus, the JP1 strain shows promise as a ferment for producing regionally identified mead, combining autochthonous yeasts and honeys typical of the Caatinga biome, which can add socioeconomic value to beekeeping in the semi-arid Northeast of Brazil.
Keywords:
honey
; beverage
; mead
1. Introduction
Mead is an alcoholic beverage obtained from the fermentation of a honeybee solution and is considered one of the oldest fermented beverages. It has been more popular in some European countries since the Middle Ages [1,2]. In recent years, public interest in this beverage has returned, even in Western Europe and North America, and more recently in Latin America, due to its potential health benefits [3,4,5,6].
In Brazil, apiculture is practiced across all natural biomes, including the Amazon rainforest, Atlantic Forest, Pantanal tropical wetland, Pampa (grassland biome), and Caatinga biome (semiarid area in the hinterland). The last one is exclusive to Brazil and encompasses xerophytic plants. Most honeys are produced in semiarid areas of Pernambuco, which comprise the geopolitical mesoregion called “Sertão”, a semiarid area in the hinterland. The harsh edaphoclimatic conditions impose on the vegetal cover the need to produce protective molecules, such as phenolic derivatives, which are taken up by bees with the nectar and concentrated during honey preparation [7,8].
We recently reported the high content of flavonols and flavonoids, which confer significant antioxidant activity and, as observed, a protective effect on the probiotic yeast Saccharomyces boullardi during in vitro simulation of the gastrointestinal tract [9]. Together, these characteristics made the honey from the Sertão a product with high nutraceutical properties. Hence, the honey from that area fulfils two important requirements: high nutraceutical properties and the absence of chemical contaminants. In this context, using the right strain of yeast can yield a high-quality mead, combining the characteristics of the honey from the Sertão with a strain well adapted to this environment that can produce key compounds that enhance the mead's sensorial properties.
However, there is a scarcity of studies evaluating the fermentation of these honeys with industrial yeasts, as well as systematic investigations into atypical residual sugars and the resulting metabolic profile. Undesirable events more frequently cited in honey fermentation are delayed or arrested fermentation and unpleasant sensory attributes that severely affect the quality of the final product [10]. The fermentations can last from 2 weeks to 1 month, making them susceptible to bacterial contamination when non-specific strains are used [11,12]. Therefore, adjustments can be made by using a high-quality substrate and an adapted yeast strain. In fermentations for other types of beverages, such as wine, cider, and beer, the yeast strain is one of the determining factors of their sensory characteristics, since the biosynthesis of sensorial molecules depends on each strain's metabolic potential [13]. However, there is no specific yeast for mead on the market so far, and producers have chosen strains used to produce red, white, and sparkling wines. Between grape wort and honey, there are quite different chemical compositions that can be offered to the yeast cells to produce sensorial molecules. For example, grape wort has approximately 100 times more assimilable nitrogen than honey wort, which is essential to produce higher alcohols and their equivalent esters [12,14,15]. Besides, nitrogen is essential for the proper functioning of key intracellular processes, and its scarcity can disrupt fermentative metabolism, leading to incomplete fermentations [16]. Given this scenario, this work proposes to evaluate the fermentative performance of different industrial strains of S. cerevisiae in the fermentation of honeys extracted from the semi-arid region of Pernambuco, with emphasis on (i) ethanol yield and sugar consumption, (ii) identification of residual sugars and potential unconventional disaccharides, (iii) characterization of the metabolic profile (organic compounds, amino acids and volatiles) of the fermented products and (iv) evaluation of technological strategies (nitrogen supplementation, high inoculation, cell immobilization) for process optimization. It is expected that the combination of robust yeasts and regional honeys will allow the production of a mead with local identity, sensory attributes, and potentially superior functional properties.
2. Materials and Methods
2.1. Bee Honeys
The bee honeys used in the present study were collected by the local producers directly from hives of Africanized Apis mellifera in apiaries of the municipalities of Triunfo (07°50′16″S 38°06′07″W) and Serra Talhada (07°59'31"S 38°17'54"W), both in the “Sertão do Pajeú”, and from the municipality of Ibimirim (8°32′26″S 37°41′25″W), in the “Sertão do Moxotó”, all in the state of Pernambuco. The word “Sertão” refers to any semiarid hinterland area in the northeastern political region of Brazil, which comprises nine states of the federation. This word is always accompanied by a geographic designation. In this case, “Pajeú” refers to the Pajeú River basin while “Moxotó” refers to the Moxotó River basin. These honeys and their respective musts will be designated from now on in this work by the acronym TF/P, ST/P and IB/M to identify their geographic origins from Triunfo/Pajeú, Serra Talhada/Pajeú and Ibimirim/Moxotó, respectively. The physical and chemical characteristics of these honeys were recently described [6,9].
2.2. Yeast Strain and Maintenance
The wine strains used in the present study are described in Table 1, along with their suppliers, product types, and sensorial characteristics.
The CA-11 strain is used for cachaça production, while the JP1 strain is commonly used to produce fuel ethanol and cachaça. It is worth noting that wine strains are used to ferment nitrogen-rich grape wort while CA-11 and JP1 are used to ferment nitrogen-poor sugarcane juice or molasses. The cells were maintained in YPD medium containing 10 g/L yeast extract, 20 g/L peptone, and 20 g/L glucose. For solid medium in plates, agar was added at a concentration of 20 g/L.
2.3. Yeast Inoculum and Wort Preparation and Mead Fermentation
The yeast cells were purchased in lyophilized form from Indupropil (Brazil), a reseller of the companies listed in Table 1, except for the Fermol Distiller JP1 strain, which was purchased from Fermenta Biotecnologia (Brazil). The cells were resuspended in YPD medium and incubated at 30 °C in an orbital shaker incubator at 160 rpm for 24 h to reactivate. One volume of the culture was used to re-inoculate nine volumes of fresh YPD medium, and the re-inoculum was incubated until the end of the exponential phase of growth. This culture was called an inoculum.
Fermentation wort was prepared by diluting honey with sterile mineral water to maintain the necessary minerals for yeast metabolism. The initial sugar content in the wort was between 24 and 26 °Brix. Then, the wort was inoculated with the inoculum culture to an initial yeast cell concentration of 1.25-2.5 × 106 cells/mL (standard condition). This initial biomass was increased whenever necessary.
2.4. Physicochemical and Microbiological Analysis
Cell counting was done under light microscopic inspection using the Neubauer chamber, and methylene blue solution (0.02% w/v) was used to identify viable (not stained) from non-viable (blue-stained) cells.
Honey, honey wort, and mead were analysed according to previously described [9]. For sugar content, samples were diluted with deionized water and filtered using 0.22 µm filters and sugars (glucose, fructose and sucrose), glycerol, acetic acid and ethanol were quantified by HPLC (High Performance Liquid Chromatography) equipped with an HPX-87H+ ion exchange column (Aminex® HPX-87H+, Bio-Rad, USA) and a refractive index detector (RID) (Agilent Technologies 1200 Series). The mobile phase consisted of 5 mM H2SO4 at a flow rate of 0.6 mL/min, maintained at 35 °C. The metabolites were identified by their respective retention times and quantified using calibration curves for each metabolite [14,17]. Whenever necessary, the content of reduced sugar was quantified by the DNS (3,5-dinitrosalicilic acid) method [18]. The total soluble solids were quantified using a manual refractometer. pH and titratable free acidity parameters were also measured using a 0.05 mol/L NaOH solution to raise the pH of the sample to 8.3 [19].
The physiological data were used to calculate the overall distribution of sugar mass in central yeast metabolism, which produces the main fermentation products. Ethanol and glycerol were directly quantified by HPLC analysis. CO2 was indirectly calculated, taking into consideration that the production of every ethanol molecule is accompanied by the production of one molecule of this gas, and that the molecular mass of CO2 (44) is 95% of the ethanol molecular mass (46). The biomass and biomass-associated CO2 were calculated according to Equation 2 previously described for S. cerevisiae oxidative metabolism [20].
2.5. Quantification of Organic Compounds
Mead samples were submitted to analysis by mass spectrometry using a Quadrupole 5975C device (Agilent, Palo Alto, CA, USA) coupled to an Agilent gas chromatograph HP 7890A equipped with an Agilent J&W non-polar HP-5ms column (30 m x 0.25 mm id.; 0.25 mm film thickness) and a thermal separation probe. Samples of fermented wort were subjected to liquid-liquid extraction by mixing the wort with cooled ethyl ether (4°C) in a 1:1 ratio. The tubes were agitated in a vortex for 1 minute, then centrifuged for 5 minutes at 11,180 x g at 4°C to separate the phases. Aliquots (1 μL) of the organic phase were placed in a quartz microvial, which was loaded into the probe and inserted into the modified GC injector. The GC injector was set to 250°C in split mode (1:10). The GC oven temperature was initially set to 60°C for 2 minutes, then increased to 300°C at 10°C/min and held for 5 minutes. Electronic flow control maintained a constant helium carrier gas flow of 1.5 mL/min. Helium (He) carrier gas flow was maintained at 7.0 psi. The MS Source and quadrupole temperatures were set at 230°C and 150°C, respectively. Mass spectra were obtained at 70 eV (EI mode) with a scan rate of 1.0 scans per second from m/z 30 to 450. Compounds detected in the samples were identified by comparing their mass spectra and retention indices with those from commercially available mass spectral libraries (MassFinder 4, NIST11, Adams, and Wiley Registry 9th Edition), integrated into the Agilent MSD Productivity ChemStation software (Agilent Technologies, Palo Alto, USA). Confirmation of the identity of some compounds was obtained by comparing their mass spectra and GC retention indices with those of authentic standards in our compound collection. The peak areas on the chromatograms were integrated to obtain the total ion signal, and their values were used to determine the relative amount (percentages) of each compound in the mead samples.
2.6. Statistical Analysis
Physiological data were expressed as the mean of at least triplicate results (±standard deviation). To evaluate significant differences amongst physicochemical characteristics of meads, a one-factor analysis of variance (ANOVA) was conducted. All statistical tests were performed at a 5% significance level. The analysis was processed in Excel software for Windows.
The VOCs MS chemical profiles of the different yeast strains were compared using both qualitative (presence/absence of compounds) and semi-quantitative (relative quantity of compounds with respect to total peak area) approaches. For qualitative and semi-quantitative comparisons, we calculated the Sørensen and Bray-Curtis’s similarity indices, respectively. These indices determine pairwise similarities among the individual samples. The relative ratios of compounds were transformed to their square root for the semi-quantitative analysis. After that and based on the obtained similarity matrices (individual-based matrices), analyses of similarities (ANOSIM, 10,000 permutations) were performed to test for differences in the scent profile of the strains. ANOSIM is a commonly used multivariate procedure roughly analogous to ANOVA/MANOVA that operates directly on a (dis)similarity matrix. It yields a test statistic R, which is a relative measure of separation among a priori-defined groups. It is based on differences of mean ranks among and within groups. A R value of ‘0’ indicates completely random grouping, whereas a value of ‘1’ indicates that samples within groups are more like each other than to any sample from a different group. Non-metric multidimensional scaling (NMDS) based on the generated similarity matrices was used to graphically display the qualitative and semi-quantitative differences in the scent profile of the yeast strain. Primer 6.1.6 was used to calculate the Sørensen and Bray-Curtis’s similarity indices and to perform ANOSIM and NMDS analyses [21].
3. Results
3.1. Fermentative Potential of Industrial Yeast Strains
Eight commercial strains of S. cerevisiae, from bread to fuel ethanol strains (Table 1), were used for the fermentation of bee honeys from the semiarid area of the state of Pernambuco, Brazil. The fermentation wort from ST/P honey was composed, on average, of fructose (52%) and glucose (38.8%). The remaining sugar was designated “apparent sucrose”. The kinetic data for total sugar consumption and ethanol production were monitored over a ten-day fermentation period (Figure 1). Except for the Côte des Blancs and Premier Cru strains, all six other strains presented nearly identical sugar consumption profiles (Figure 1A, C). Regarding the types of sugars assimilated, TR313 and JP1 strains consumed most of the glucose and fructose, while the CA-11 strain left the highest levels of these monosaccharides in the wort (data not shown). On average, only 13% of the initial apparent sucrose was used by cells from the eight strains. Later, we will return to the issue of sucrose.
TR313, Côte des Blancs, Premier Classic, and JP1 strains showed slower ethanol production kinetics than the other strains (Figure 1B, D). However, at the end of fermentation, the strain JP1, along with strains Diamond and Lalvin, showed the highest alcohol content (Table 2).
The fermentation yield was the same for all eight strains, indicating a constant physiological parameter in the conversion efficiency of 38% of the mass of honey wort sugars into ethanol (Table 2). Glycerol production (5.9±0.7 g/L) was also similar between strains, meaning that 3% of the mass of consumed sugar was converted to glycerol by cells. These values indicate that the distribution of carbon through the central metabolism of these strains was very similar, reaching constant conversion values to ethanol and glycerol. Therefore, the most relevant difference among them, although small, seems to be the differential consumption capacity of the sugars present in the wort.
Therefore, given the constancy of the yield data, we took into consideration the stoichiometry of biomass/CO2 formation for S. cerevisiae [20] to define the general calculation of the yields for the main products generated by the consumption of sugars at the end of fermentation of the bee honey. This calculation indicated the following mean stoichiometric equation for mead production:
100 g consumed honey sugar = 39 g ethanol + 3 g glycerol + 45 g CO2 + 13 g yeast biomass
Therefore, this equation served as a reference for the selection of yeasts with minimal performance for mead production when using honey from that origin. The maximal theoretical yield for ethanol is 51.1 g of ethanol per 100 g of consumed sugar. Thus, this equation indicated that the fermentation efficiency of this type of honey achieved only 76% of the maximum. This low efficiency can be explained by the presence of phenolic compounds in the honey must, which would cause some metabolic diversion of carbon to non-fermentative pathways. However, it should be noted that the alcohol content achieved by fermentation with the different strains is within the expected range for this type of alcoholic beverage.
The analysis of organic molecules in the fermented must revealed the presence of TCA cycle derivatives, indicating an overflow of central metabolism due to the high amount of glucose taken up by the yeast cells (Figure 2). All the commercial strains secreted practically the same amount of citrate and succinate. Strain TR313 produced more 2-oxoglutarate, while the Lavin strain produced more fumarate (Figure 2). In addition, TR313 and JP1 produced less malate than the other strains (Figure 2). This represents an advantage, as malate might impart a tart taste to beverages and must be converted to lactate during malolactic fermentation in the wine process [22]. This conversion is catalysed by the malic enzyme produced by lactic acid bacteria present in the grape must during the second fermentation stage of the winemaking process [23]. Therefore, the lower malate production can be considered as a superior characteristic among the different strains of S. cerevisiae, in this case, strains TR313 and JP1. Furthermore, the high citrate production by the JP1 strain (Figure 2) can confer a refreshing character to the mead produced by this yeast.
The screening data showed that, despite very close ethanolic yields, the final products fermented by the different yeasts must present different characteristics, with sweetness as the main one. Therefore, a classification of the tested yeasts can be established using this characteristic as follows. Lalvin k1v-1116 strain produced a drier, more ethanolic beverage. According to the manufacturer, this strain is marketed for aromatic red wine with a very floral bouquet, as well as for whites, sparkling wines, and late-harvest wines, and is recommended for fermentation under so-called difficult conditions (https://www.lallemandbrewing.com). However, the drier content can make consumption somewhat difficult in warmer regions, such as the tropics. On the other hand, the LalBrew Belle Saison strain produced a less alcoholic and sweeter beverage. According to the manufacturer, the yeast S. cerevisiae var. diastaticus is suitable for fruity, spicy, and refreshing Belgian-style beer (https://www.lallemandbrewing.com/). This product, although it may have strong market acceptance, may be harmed by the trend towards reducing sugar consumption among populations.
In this context, the Renascence TR313 strain also presented the same alcohol content with less residual sugar than the LalBrew Belle saison. This yeast is suitable for the production of white wines, with a pronounced flavour of passion fruit, guava, and cherries, and the production of mercaptan volatiles, as well as low nitrogen requirements (https://renaissanceyeast.com/). The passion fruit touch is generally well-received in warmer regions, such as the tropics, leading to strong consumer acceptance of the product.
In contrast, Fermol Distiller JP1 strain produced a product with a high alcohol content and a moderate residual sugar content. This yeast was isolated from the fermentation of sugarcane juice to produce fuel ethanol [24,25] and marketed basically for this purpose in the state of Pernambuco and other states in the Northeast region of Brazil due to its high-capacity persistence in fermentation tanks and its high tolerance to variations in process temperature (https://www.aeb-group.com/). This strain was also tested for cachaça production from sugarcane juice, demonstrating high efficiency in producing medium-chain alcohols (amyl, isoamyl, and isobutyric) [15]. Based on these studies, the JP1 strain has also been commercialized to produce cachaça in the State of Pernambuco and other states in the Northeast region of Brazil. The characteristics of this strain, combined with its geographic origin and the origin of the honey used, can give the mead produced a very distinctive identity as the first genuinely mead from this region of the world, with promising market value.
3.2. Fermentation Profile of Selected Strains
Given the above factors, the JP1 and TR313 strains were selected to better evaluate mead production with ST-P honey. The fermentative kinetics data showed that the JP1 strain consumed all the glucose within 16 days of fermentation but was unable to exhaust the fructose even after 30 days (Figure 3A). On the other hand, the TR313 strain consumed glucose within 20 days of fermentation and fructose after 30 days (Figure 3B). This result revealed the physiological difference between these S. cerevisiae strains, as noted above.
Another interesting aspect was that ethanol production began earlier with JP1 than with TR313, coinciding with the strain's fructose consumption (Figure 3). This meant that fermentation practically ended after 22 days of incubation with JP1 (Figure 2A), whereas TR313 continued producing ethanol until the 30th day due to fructose consumption (Figure 3B). As a result, the volumetric productivity calculated from the most linear range of the ethanol production curves was 0.23 g of ethanol/L.h for JP1 (between zero and 12 days) and 0.17 g of ethanol/L.h for TR313 (between 2 and 22 days). Therefore, it is concluded that the production of mead by the JP1 strain must occur in a shorter time than by TR313, which implies a reduction in production costs.
An important aspect noted in all mead fermentations was the high residual sucrose. Figure 3 showed no mobilization of this sugar during fermentation by either strain tested. This is very intriguing, since the yeast S. cerevisiae normally secretes invertase for extracellular hydrolysis of sucrose and the consumption of the resulting monosaccharides, glucose and fructose. The three hypotheses to explain this result are: the yeasts are not producing invertase, the honey contains some invertase-inhibiting substance, or this sugar is another polysaccharide with a retention time similar to sucrose under the HPLC conditions used. To test the first two hypotheses, JP1 cells were cultivated in a synthetic mineral medium with sucrose to induce invertase production. The supernatant was collected and tested for enzyme activity (Figure 4). When added to a solution containing sucrose, it was possible to observe the disappearance of the disaccharide. This indicates that JP1 cells can produce invertase. However, the supposed sucrose in the honey wort remained constant, suggesting that the enzyme produced was not capable of hydrolysing it. When sucrose was added to honey must, its hydrolysis and the intact residual of supposed sucrose were observed. Therefore, it is concluded that there are no invertase inhibitors in honey wort and that this sugar must be of another type. It is interesting to note that even the LalBrew Belle Saison strain, which produces β-glucosidase in addition to invertase, was not able to consume this sugar (Figure 1A). This recalcitrant sugar must be a di- or trisaccharide that does not have β1,4 bonds in its structure, such as isomaltose that is found in honey [26,27,28].
Another possibility for this apparent sucrose is an analogue called turanose. This is a dimer of glucose and fructose with an α-1,3 glycosidic bond, whereas in sucrose this bond is α-1,2 [29]. Other structural analogues of sucrose include trehalulose, leucrose, maltulose, and isomaltulose, all of which have great potential as low-calorie sweeteners [30]. Turanose has recently gained attention for its potential use in biotechnological processes in the food and pharmaceutical industries, as well as serving as a molecular marker of honey authenticity [29]. This molecule is a byproduct of the synthesis of linear α-1,4 glucan structures by plant amylosucrase, and it appears in European nectar and honey at concentrations so low that they prevent commercial extraction [31]. However, its residual concentration ranged from 20 g/L in the must to 20 g/L in the final mead (Figure 3). This means that the turanose concentration in honey should be around 60 g/L, which would represent a very important characteristic of the nectar of plants in the Caatinga biome, exclusive to the semiarid region of northeastern Brazil. In this scenario, fermentation of the honey must remove all glucose and fructose, thereby increasing the purification efficiency and the concentration of turanose for industrial use.
Nevertheless, its identification is beyond the scope of this work. Here, it is only important to note that this sugar will impart a sweet taste to the mead, which can alleviate the alcoholic sensation and increase consumer acceptability.
3.3. Extracellular Metabolomics of the Selected Strains
Amino acids and aliphatic organic acids were quantified from the supernatant of the mead produced by TR313 and JP1 strains (Figure 5). The beverage resulting from JP1 fermentation had higher content of almost all detected amino acids compared to TR313, while the fermented product of TR313 contained more leucine (Figure 5A,B). It is worth noting the high content of alanine and proline in the meads of both strains.
The amino acid-rich mead produced by JP1, especially in alanine, valine, phenylalanine, and proline, is a relevant characteristic of this product. Pollinating insects, such as bees, prefer proline-rich nectars because proline is oxidized in the insect's flight muscle, thereby protecting this tissue from oxidative stress, especially during the first 30 s of flight [32]. Therefore, it appears that there has been a process of plant-insect coevolution toward the provision of proline-rich nectars as a mechanism to attract visiting pollinators [33]. This may explain the higher proline content relative to other amino acids in fermented musts derived from diluted honey (Figure 5B). Proline is an important amino acid with neurotransmitter activity in the central nervous system, influencing cognitive and behavioural functions [34]. In controlled doses, proline may exhibit neuroprotective effects that help mitigate the symptoms of various neurological diseases and psychiatric disorders [34].
Regarding the other compounds, 54 chromatography peaks were detected, of which 31 were identified, and 23 remained unidentified. Among the identified compounds, 10 were significantly present in the replicates (Figure 5C,D). However, only a few are worthy of consideration due to their representation in the samples, which are more abundant in the supernatant of those fermented with the JP1 strain: glyceric acid, 2-aminobutyric acid, and dehydroascorbic acid (Figure 5C,D).
Glyceric acid is an oxidated product of glycerol that is present in several fruit like apple, grape, banana and tomato, which has been used as raw material to produce pharmaceutical s and cosmetic products [35], and as a moisturizing agent by the skincare industry as well as a flavouring agent by the food industry [36].
3-aminobutyric acid, also known as gamma-aminobutyric acid (GABA), is the product of glutamine decarboxylation that occurs in the glutamine degradation pathway across the eukaryotic domain [37]. In S. cerevisiae, this reaction occurs in the cytoplasm via glutamine decarboxylase encoded by the GAD1 gene in response to oxidative stress [38]. In animals, specifically in the mammalian brain, GABA acts as a neurotransmitter involved in a series of metabolic activities, and is pharmaceutically explored for the treatment of epilepsy, diabetes, hypertension, etc. [37]. Notably, this molecule accounts for a significant fraction of the extracellular metabolome of JP1-produced mead, potentially conferring certain pharmaceutical properties to the mead.
Finally, dehydroascorbic acid (DHAA) is the oxidized form of ascorbic acid, resulting from its action in the oxidative stress response [39]. Under oxidative stress, the intracellular glutathione pool is mobilised to reduce and detoxify oxidant molecules. As a result, two molecules of oxidised glutathione form a disulfide bridge, which is broken in a reduction reaction using ascorbic acid as an electron donor. Ultimately, glutathione is restored to its reduced form, while ascorbic acid is oxidized to DHAA [39]. S. cerevisiae does not synthesize ascorbic acid nor DHAA under physiological conditions, but its analogous D-erythroascorbic acid from D-arabinose [40]. Thus, we concluded that DHAA found in the mead supernatant came from the honey and, consequently, from plant nectar. The antioxidant potential of DHAA has been reported, either directly or via the regeneration of ascorbic acid, a well-established potent antioxidant [41]. Thus, its presence in the mead may increase the antioxidant potential of the final product, in conjunction with the phenolic agents highly present in the honeys of Sertão do Pajeú. Moreover, DHAA and other lactone derivatives have been reported for their anticancer potential [39]. The higher concentration of DHAA in the JP1 fermentation supernatant may reflect a more active degradation mechanism in the TR313 cells.
Extracellular metabolomics also revealed the presence of volatile organic compounds (VOCs) relevant to the sensorial characteristics of the meads (Figure 6). 2,3-butanediol, a reduced form of diacetyl (2,3-butanedione), is a well-known metabolite produced by yeast during fermentation. In beer and wine, diacetyl is an important off-flavour that imparts a buttery or butterscotch-like flavour to the beverage [42]. The diacetyl is formed from the condensation of pyruvate and lactate to α-acetolactate, followed by decarboxylation of this product. However, during prolonged fermentation or maturation, diacetyl can be reduced to acetoin, which can then be reduced to 2,3-butanediol [42]. Therefore, the fermentation period appears to have allowed the JP1 strain's cells to remove diacetyl more efficiently (Figure 6).
The aromatic alcohol 2-phenylethanol (2-PE) is one of the most important rose-flavour metabolites present in beverages, foods, and cosmetics [43]. In beverages, it is produced by yeast cells during fermentation from the catabolism of phenylalanine present in the must via the Ehrlich pathway [14,44,45]. This molecule was detected in the meads produced by both S. cerevisiae strains in the same amount, accounting for almost 40% of the VOCs (Figure 6).
p-hydroxyphenethyl alcohol (tyrosol) was another compound that was relatively abundant in both meads, between 6% to 9% of the total VOCs in the supernatants (Figure 6). Tyrosol is a phytochemical found in natural products such as olive oil and wine, and is described as a sweet, fruity-floral odour [46]. It is also the product of the Ehrlich pathway for the catabolism of tyrosine in S. cerevisiae cells during fermentation [47]. Olive oil is the natural source from which tyrosol is extracted. This compound is described as having antioxidant and anti-inflammatory properties, among others, and acts as a free radical scavenger, together with other phenolic compounds [48]. Therefore, the presence of tyrosol may aggregate the final antioxidant panel of the meads, together with the flavonoids and flavonols already present at high concentration in the honeys from the Sertão do Pajeú [9].
p-Anisic acid (ANAc), also known as 4-methoxybenzoic acid, is a p-alkoxy benzoic acid originally purified from Pimpinella anisum, a Mediterranean herbal plant cultivated for its flavouring properties for foods and alcoholic drinks, for its traditional medicine use, and as a source of biopharmaceutical molecules [49]. Another plant, Acacia mearnsii, cultivated in Brazil and South Africa as a source of tannins and other bioactive molecules, contains a high level of this phenolic acid [50]. ANAc is the oxidised product of p-anisaldehyde (pANAd), while the reduction of pANAd produces p-anisyl alcohol (pANOH). Both ANAc and pANAd were reported as volatile compounds in Manuka and Kanuka honeys, respectively [52]. These compounds are natural fragrances produced by plants' secondary metabolism, with a sweet, floral bouquet ANAc, pANAd and pANOH, together with some coumarin derivatives, compose the so-called hawthorn note, which is very appreciated by the cosmetic and perfumery industry [52]. Moreover, ANAc is described to have antioxidant, antitumor, antiseptic, anti-diabetic and insulinotropic properties [53,54]. In microbial metabolism, genes such as p-anisaldehyde dehydrogenase (PAADH) and trans-anethole oxygenase (TAO) are found in the bacterium Pseudomonas putida, which makes ANAc production from vanillin [55]. However, homologous of those genes are not found in S. cerevisiae. Even so, the two yeast strains showed a significant difference in the relative concentration of these compounds in the mead (Figure 6). The mead produced by the JP1 strain contained more of the three molecules than the mead of the TR313 strain. This leads to the conclusion that these two strains interact differently with these compounds from honey.
Overall, despite the VOCs’ chemical profiles being similar, the mead produced by JP1 seems to have a more pronounced sensorial character. It is important to note that TR313 is a selected yeast for wine production, while JP1 is an industrial yeast from fuel-ethanol production. This ecological difference further highlights JP1's potential for mead production, especially when using honey extracted from plants in the semiarid region of Brazil. The advantages of a mead produced by JP1 over the TR313-fermented product were shown, considering the presence of metabolites with different sensorial and pharmaceutical properties. The combination of an exclusive hinterland honey with local fermenting yeast may confer the originality of a mead brand that would aid in increasing the product's added value and, consequently, increase the incoming of small honey producers of that region.
3.4. Fermol Distiller JP1 Strain as Mead-Fermenter Yeast
All biochemical and physiological parameters evaluated above indicated the potential of the JP1 strain for fermenting honey and producing mead. To expand this hypothesis, fermentation assays were conducted using different honeys from other regions in the semiarid area of the state of Pernambuco, which had been chemically characterized in recent works [6,9]. Worts were prepared by diluting all the bee honeys to 28% (w/v) of the initial sugar. Despite the small differences, the fermentation profiles were very similar (Table 3). Meads produced from IB/M and ST/P honeys were sweeter than the mead from the TF/P honey, while meads from ST/P and TF/P showed higher ethanol content. However, the higher glycerol concentration in TF/P mead should give this drink greater smoothness, helping to alleviate the higher alcohol content. All preparations reached the minimum alcoholic yield of 38% established above for the efficiency of the fermentation process. Therefore, the JP1 strain is efficient at fermenting must prepared with honeys from different locations in the semiarid hinterland of the state of Pernambuco.
Given the above, we tested three technological improvements to evaluate their potential impact on the fermentation process (Figure 7). The first was supplementing the honey must with ammonium sulphate to increase nitrogen availability for the yeast cells (Figure 7A). This supplementation accelerated fermentation, with increased hexose consumption and an anticipation of peak ethanol production by 10 days (Figure 7A), half the time observed in the must without supplementation (Figure 3A). Glycerol production and maintenance of apparent sucrose were similar between the two conditions. The second was a twofold increase in initial yeast biomass (Figure 7B). In this case, the fermentation profile resembled that of ammonium supplementation (Figure 7A), indicating that the added nitrogen doubled biomass in the first hours of the fermentation trial. The third technological improvement involved the use of cells immobilized in alginate (Figure 7C). In this case, the fermentation profile was like the first two, indicating that immobilization likely led to a twofold increase in the process's initial biomass without disturbing the cells' metabolic activity. Even though immobilization provides more efficient cell removal from the final product, leaving it clearer, it also incurs a cost. Therefore, the producer can choose any of the three technological modifications based on the cost-benefit ratio.
4. Conclusions
Mead is an ancient beverage that faces several challenges in production, as the quality of the honey and the yeast strain are not well-suited to this substrate. The results showed that, even with a similar fermentative profile, the rate of sugar consumption can be important in choosing the better strain for fermentation. Saccharomyces cerevisiae JP1 seems to be a good candidate for the process, even though it is a yeast isolated from the sugarcane industry for fuel-ethanol production in the Northeast of Brazil, especially for its rapid carbohydrate assimilation in the medium. In addition, S. cerevisiae JP1 can impart a more complex sensorial profile and bioactive compounds to the mead compared to other strains. The use of this strain with the honey of the Sertão can characterize the regional product and increase its value. Otherwise, more studies are necessary to increase productivity without affecting sensorial quality and to understand the presence of carbohydrates that cannot be assimilated during the fermentative process, as well as the potential nutraceutical effects of this mead.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
RKS, JDF and HAFM: Methodology, Investigations, Formal analysis, Data curation, Conceptualization, Writing-original draft. JDF: Methodology, Investigations, formal analysis, Data curation. WPPN and LLALA: Investigations. AMPS: Investigations, formal analysis, Data curation, Supervision. HFM: Methodology, Investigations, formal analysis, Data curation, Conceptualization. DMAFN and PMP: Investigations, formal analysis, Data curation. MAMJ and RBS: Formal analysis, Data curation, Conceptualization, Supervision, Project administration, Funding acquisition, Writing-original draft, Writing-review and editing.
Funding
This work was supported by Fundação de Amparo à Pesquisa do Estado de Pernambuco (FACEPE), which financed this research by the Local Productive Arrangements grant number APQ-0434-2.12/20 and by the Innovation Locus program 02/2022 grant number APQ-0161-9.26/22.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are available on this work.
Acknowledgments
The authors are grateful to the Fundação de Amparo à Pesquisa do Estado de Pernambuco (FACEPE) to the Beekeepers Association of Serra Talhada, Beekeepers Association of Triunfo and Beekeepers Association of Ibimirim for providing the honey samples for this study.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| HPLC | High-Performance Liquid Chromatography |
| TF/P | Triunfo/Pajeú |
| ST/P | Serra Talhada/Pajeú |
| IB/M | Ibimirim/Moxotó |
| MRS | Man, Rogosa & Sharpe medium |
| RID | Refractive Index Detector |
References
- Gupta, J.K.; Sharma, R. Production technology and quality characteristics of mead and fruit-honey wines: a review. Nat. Prod. Radiance 2009, 8, 345–355. [Google Scholar]
- Ramalhosa, E.; Gomes, T.; Pereira, A.P.; Dias, T.; Estevinho, L.M. Mead production: tradition versus modernity. Adv. Food Nutr. Res. 2011, 101–118. [Google Scholar]
- Barbosa, A.B.; Martins, E.A. Produção artesanal de hidromel. Jornada Cient. Tecnol. FATEC Botucatu 2017. [Google Scholar]
- Simão, L.; Wanderley, B.R.D.S.M.; Tavares Vieira, M.P.; Silva Haas, I.C.; Amboni, R.D.M.C.; Fritzen-Freire, C.B. How do different ingredients and additives affect mead production steps and bioactive potential? Food Technol. Biotechnol. 2023, 61, 179–190. [Google Scholar] [CrossRef]
- Jose-Salazar, J.A.; Ballinas-Cesatti, C.B.; Hernández-Martínez, D.M.; Cristiani-Urbina, E.; Melgar-Lalanne, G.; Morales-Barrera, L. Kinetic evaluation of mead production from a non-Saccharomyces strain. Foods 2024, 13, 1948. [Google Scholar] [CrossRef]
- Pinto Neto, W.P.; Lucena, T.M.C.; Paixão, G.A.; Shinohara, N.K.S.; Pinheiro, A.C.; Vicente, A.A.; Souza, R.B.; Morais Junior, M.A. Symbiotic honey beverages: a matrix which tells a story of survival and protection of human health. Int. J. Gastron. Food Sci. 2025, 40, 101183. [Google Scholar] [CrossRef]
- Galvíncio, J.D.; Badarau, M.S.; Barbosa, V.V.; Freire, F.J.; Freire, M.B.G.S.; Souza, W.M. Avaliação do estágio de regeneração do bioma Caatinga na microrregião do Araripe com sensoriamento remoto. Rev. Bras. Geogr. Fís. 2016, 9, 986–996. [Google Scholar] [CrossRef]
- Vidal, M.F. Produção de mel na área de atuação do BNB entre 2011 e 2016. In Caderno Setorial ETENE; 2018. [Google Scholar]
- Pinto-Neto, W.P.; Silva, R.K.; Lima, B.S.; Acioli, G.F.S.; Paixão, G.A.; Muniz, B.C.; Silva, P.K.N.; Costa, R.M.P.B.; Silva, F.S.B.; Melo, H.F.; Souza, R.B.; Morais Junior, M.A. Bee honey of the Pajeú hinterland, Pernambuco, Brazil: physicochemical characterization and biological activity. Food Biosci. 2024, 60, 104289. [Google Scholar] [CrossRef]
- Iglesias, A.; Pascoal, A.; Choupina, A.B.; Carvalho, C.A.; Feás, X.; Estevinho, L.M. Developments in the fermentation process and quality improvement strategies for mead production. Molecules 2014, 19, 12577–12590. [Google Scholar] [CrossRef] [PubMed]
- Sroka, P.; Tuszyński, T. Changes in organic acid contents during mead wort fermentation. Food Chem. 2007, 104, 1250–1257. [Google Scholar] [CrossRef]
- Pereira, A.P.; Dias, T.; Andrade, J.; Ramalhosa, E.; Estevinho, L.M. Mead production: selection and characterization assays of Saccharomyces cerevisiae strains. Food Chem. Toxicol. 2009, 47, 2057–2063. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its importance to wine aroma: a review. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef]
- Vidal, E.E.; de Billerbeck, G.M.; Simões, D.A.; Schuler, A.; François, J.M.; de Morais Junior, M.A. Influence of nitrogen supply on higher alcohol and ester formation in cachaça fermentation. Food Chem. 2013, 138, 701–708. [Google Scholar] [CrossRef]
- Vidal, E.E.; de Morais Junior, M.A.; François, J.M.; de Billerbeck, G.M. Biosynthesis of higher alcohol flavour compounds by Saccharomyces cerevisiae. Yeast 2015, 32, 47–56. [Google Scholar]
- Schwarz, L.V.; Marcon, A.R.; Delamare, A.P.L.; Agostini, F.; Moura, S.; Echeverrigaray, S. Selection of low-nitrogen-demand yeast strains and their impact on mead composition. J. Food Sci. Technol. 2020, 57, 2840–2851. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Neto, W.P.; Silva, A.K.L.F.; Araújo, K.S.; Pinheiro, I.O.; Souza, R.B.; Morais Junior, M.A. Optimizing wheat beer production by recycling yeast biomass. Food Sci. Technol. 2023, 43. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Instituto Adolfo Lutz. Métodos físico-químicos para análise de alimentos; São Paulo, 2008. [Google Scholar]
- Leite, F.C.; Basso, T.O.; Pita, W.B.; Gombert, A.K.; Simões, D.A.; de Morais Junior, M.A. Quantitative aerobic physiology of Dekkera bruxellensis. FEMS Yeast Res. 2013, 13, 34–43. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: user manual/tutorial; PRIMER-E: Plymouth, 2006. [Google Scholar]
- Fu, J.; Wang, L.; Sun, J.; Ju, N.; Jin, G. Malolactic fermentation: new approaches to old problems. Microorganisms 2022, 10, 2363. [Google Scholar] [CrossRef]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Implications of new research for malolactic fermentation in wine. Appl. Microbiol. Biotechnol. 2014, 98, 8111–8114. [Google Scholar] [CrossRef]
- Silva-Filho, E.A.; Santos, S.K.B.; Resende, A.M.; Simões, D.A.; de Morais Junior, M.A. Yeast population dynamics in industrial ethanol fermentation. Antonie Van Leeuwenhoek 2005, 88, 13–23. [Google Scholar] [CrossRef]
- Silva-Filho, E.A.; Melo, H.F.; Antunes, D.F.; Santos, S.K.; Resende, A.M.; Simões, D.A.; de Morais Junior, M.A. Isolation of a fermentative Saccharomyces cerevisiae strain. J. Ind. Microbiol. Biotechnol. 2005, 32, 481–486. [Google Scholar] [CrossRef]
- Pascual-Maté, A.; Osés, S.M.; Marcazzan, G.L.; Gardini, S.; Fernández-Muiño, M.A.; Sancho, M.T. Sugar composition and related parameters of honeys. J. Food Compos. Anal. 2018, 74, 34–43. [Google Scholar] [CrossRef]
- Loza, J.P.; Chailyan, A.; Förster, J.; Katz, M.; Mortensen, U.H.; Garcia Sanchez, R. Improving the utilization of isomaltose and panose by lager yeast. Fermentation 2021, 7, 107. [Google Scholar] [CrossRef]
- Xu, X.; Asai, K.; Kato, D.; Ishiuchi, K.; Ding, K.; Tabuchi, Y.; Ota, M.; Makino, T. Honey isomaltose and G-CSF induction. Sci. Rep. 2020, 10, 15178. [Google Scholar] [CrossRef]
- Ponnurangam, M.; Balaji, S. Tune in to the terrific applications of turanose. Eur. Food Res. Technol. 2024, 250, 375–387. [Google Scholar] [CrossRef]
- Tian, Y.; Deng, Y.; Zhang, W.; Mu, W. Sucrose isomers as alternative sweeteners. Appl. Microbiol. Biotechnol. 2019, 103, 8677–8687. [Google Scholar] [CrossRef]
- Kaškonienė, V.; Venskutonis, P.; Čeksterytė, V. Carbohydrate composition and electrical conductivity of honeys. LWT Food Sci. Technol. 2010, 43, 801–807. [Google Scholar] [CrossRef]
- Bertazzini, M.; Medrzycki, P.; Bortolotti, L.; Maistrello, L.; Forlani, G. Amino acid content and nectar choice by honeybees. Amino Acids 2010, 39, 315–318. [Google Scholar] [CrossRef]
- Carter, C.; Shafir, S.; Yehonatan, L.; Palmer, R.G.; Thornburg, R. A novel role for proline in floral nectar. Naturwissenschaften 2006, 93, 72–79. [Google Scholar] [CrossRef]
- Carvalho, G.A.; Cavalcante, D.P.; Parreira, R.C.; Chiareli, R.A.; Ortiz Leoncini, G.; Gomez, R.S.; Ulrich, H.; Ferreira Caixeta, L.; Oliveira-Lima, O.C.; Pinto, M.C.X. Neurobiology of L-proline. Neuroscience 2025, 568, 116–129. [Google Scholar] [CrossRef]
- Habe, H.; Fukuoka, T.; Kitamoto, D.; et al. Biotechnological production of d-glyceric acid. Appl. Microbiol. Biotechnol. 2009, 84, 445–452. [Google Scholar] [CrossRef]
- Matter, Chemical. Glyceric acid. Available online: https://chemicalmatter.com/glyceric-acid (accessed on 16 July 2025).
- Zhang, Q.; Zhu, L.; Li, H.; et al. Insights and progress on biosynthesis and functions of GABA. PeerJ 2024, 12, e18712. [Google Scholar] [CrossRef]
- Coleman, S.T.; Fang, T.K.; Rovinsky, S.A.; Turano, F.J.; Moye-Rowley, W.S. Glutamate decarboxylase and oxidative stress tolerance in yeast. J. Biol. Chem. 2001, 276, 244–250. [Google Scholar] [CrossRef]
- Toohey, J.I. Dehydroascorbic acid as an anti-cancer agent. Cancer Lett. 2008, 263, 164–169. [Google Scholar] [CrossRef]
- Amako, K.; Fujita, K.; Shimohata, T.A.; Hasegawa, E.; Kishimoto, R.; Goda, K. NAD+-specific D-arabinose dehydrogenase in yeast. FEBS Lett. 2006, 580, 6428–6434. [Google Scholar] [CrossRef]
- Wilson, J.X. The physiological role of dehydroascorbic acid. FEBS Lett. 2002, 527, 5–9. [Google Scholar] [CrossRef]
- Gibson, B.; Vidgren, V.; Peddinti, G.; Krogerus, K. Diacetyl control in brewery fermentation via adaptive engineering. J. Ind. Microbiol. Biotechnol. 2018, 45, 1103–1112. [Google Scholar] [CrossRef]
- Mitri, S.; Koubaa, M.; Maroun, R.G.; Rossignol, T.; Nicaud, J.M.; Louka, N. Bioproduction of 2-phenylethanol. Foods 2022, 11, 109. [Google Scholar] [CrossRef]
- Vuralhan, Z.; Morais, M.A.; Tai, S.L.; Piper, M.D.; Pronk, J.T. Phenylpyruvate decarboxylase genes in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2003, 69, 4534–4541. [Google Scholar] [CrossRef]
- Bernardino, A.R.S.; Torres, C.A.V.; Crespo, J.G.; Reis, M.A.M. Biotechnological 2-phenylethanol production. Molecules 2024, 29, 5761. [Google Scholar] [CrossRef]
- Gutiérrez-Escobar, R.; Aliaño-González, M.J.; Cantos-Villar, E. Wine polyphenol content and its influence on wine properties. Molecules 2021, 26, 718. [Google Scholar] [CrossRef]
- Li, J.; Zhang, S.; Li, H.; et al. Engineering phenylpyruvate decarboxylase for biosynthesis of aromatic derivatives. Metab. Eng. 2025, 91, 466–479. [Google Scholar] [CrossRef]
- Rocha, J.; Borges, N.; Pinho, O. Table olives and health: a review. J. Nutr. Sci. 2020, 9, e57. [Google Scholar] [CrossRef]
- Wu, J.; Cao, Z.; Hassan, S.S.U.; et al. Emerging biopharmaceuticals from Pimpinella genus. Molecules 2023, 28, 1571. [Google Scholar] [CrossRef]
- Da Silva, G.F.; de Souza Júnior, E.T.; Almeida, R.N.; Fianco, A.L.B.; do Espírito Santo, A.T.; Lucas, A.M.; Vargas, R.M.F.; Cassel, E. Supercritical CO₂ extraction of p-anisic acid from Acacia mearnsii. Molecules 2022, 27, 970. [Google Scholar]
- Beitlich, N.; Koelling-Speer, I.; Oelschlaegel, S.; Speer, K. Differentiation of manuka and kanuka honeys. J. Agric. Food Chem. 2014, 62, 6435–6444. [Google Scholar] [CrossRef]
- Bouville, A.S.; Fonte, K.; Portes, P.; et al. Forgotten perfumery plants: hawthorn case study. Chem. Biodivers. 2024, 21, e202301020. [Google Scholar] [CrossRef] [PubMed]
- Papageorgiou, S.; Varvaresou, A.; Tsirivas, E.; Demetzos, C. New alternatives to cosmetic preservation. J. Cosmet. Sci. 2010, 61, 107–123. [Google Scholar]
- Vora, V.; Tirgar, P.; Raval, K. Anti-diabetic and insulinotropic effects of p-anisic acid. Chem. Biodivers. 2024, 21, e202401575. [Google Scholar] [CrossRef]
- Han, D.; Kurusarttra, S.; Ryu, J.Y.; Kanaly, R.A.; Hur, H.G. Production of aromatic acids by engineered Escherichia coli. J. Agric. Food Chem. 2012, 60, 11972–11979. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Kinetics profile of sugar consumption (A, C) and ethanol production (B, D) by Saccharomyces cerevisiae industrial strains fermenting a wort of diluted bee honey from Serra Talhada, in the Pajeú semiarid hinterland, Pernambuco, Brazil. Panels A and B are shown Saf-Instant for baking (Sc01), CA-11 for cachaça (Sc02), Diamond for German Lager beer (Sc03), Red Star Premier for white wine (Sc04), and LalBrew Belle saison for Belgian beer (Sc05). Panels C and D are shown TR313 for white wine (Sc06), Lalvin k1v-1116 for red wine and mead (Sc07), Côte de blancs for white wine and mead (Sc08), Premier classic for white wine and mead (Sc09) and Fermol Distiller JP1 for fuel-ethanol and cachaça (Sc10).
Figure 1.
Kinetics profile of sugar consumption (A, C) and ethanol production (B, D) by Saccharomyces cerevisiae industrial strains fermenting a wort of diluted bee honey from Serra Talhada, in the Pajeú semiarid hinterland, Pernambuco, Brazil. Panels A and B are shown Saf-Instant for baking (Sc01), CA-11 for cachaça (Sc02), Diamond for German Lager beer (Sc03), Red Star Premier for white wine (Sc04), and LalBrew Belle saison for Belgian beer (Sc05). Panels C and D are shown TR313 for white wine (Sc06), Lalvin k1v-1116 for red wine and mead (Sc07), Côte de blancs for white wine and mead (Sc08), Premier classic for white wine and mead (Sc09) and Fermol Distiller JP1 for fuel-ethanol and cachaça (Sc10).
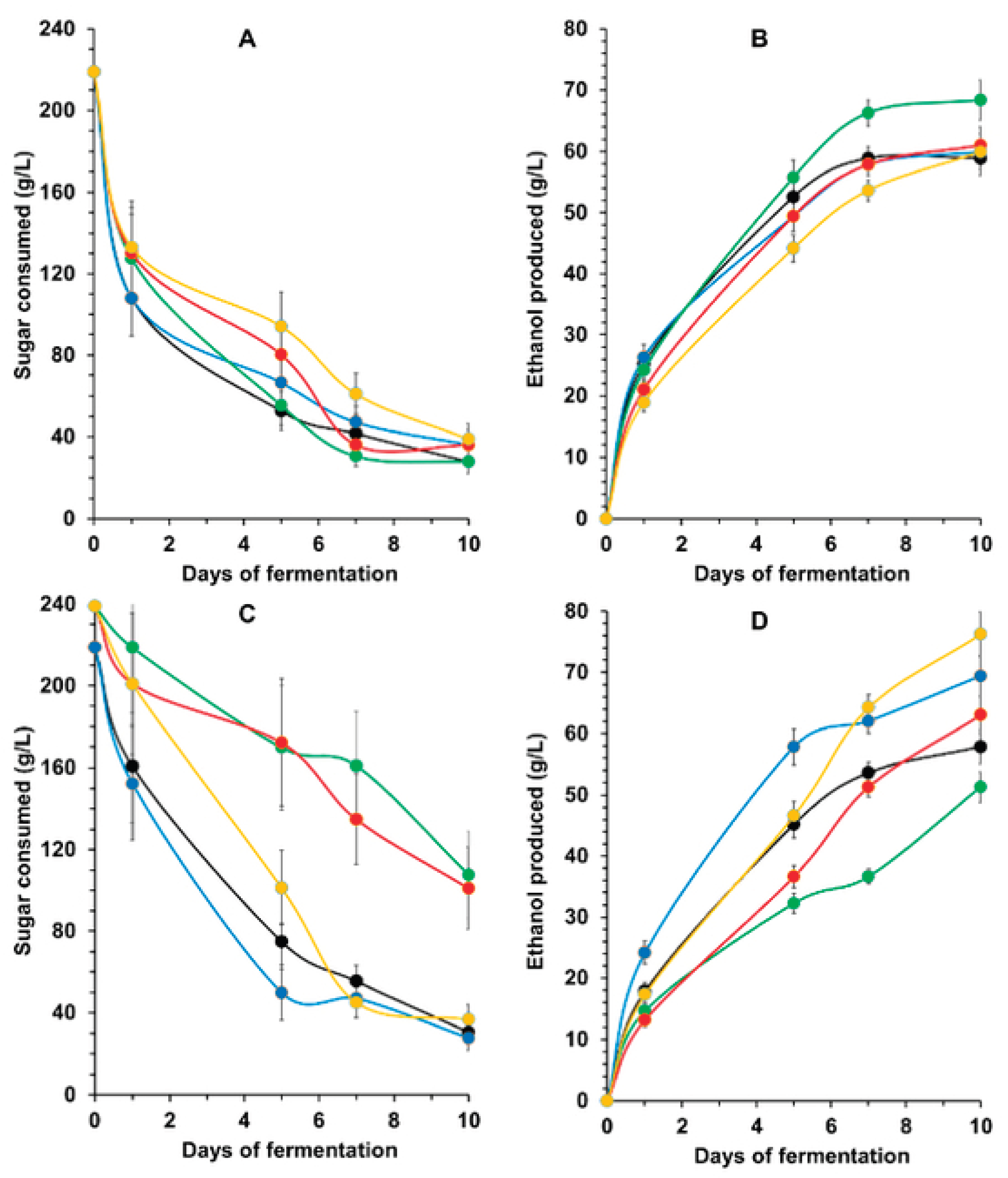
Figure 2.
Quantification of intermediates of the Tricarboxylic Acid cycle in the supernatant of honey-fermented must by different industrial Saccharomyces cerevisiae strains.
Figure 2.
Quantification of intermediates of the Tricarboxylic Acid cycle in the supernatant of honey-fermented must by different industrial Saccharomyces cerevisiae strains.
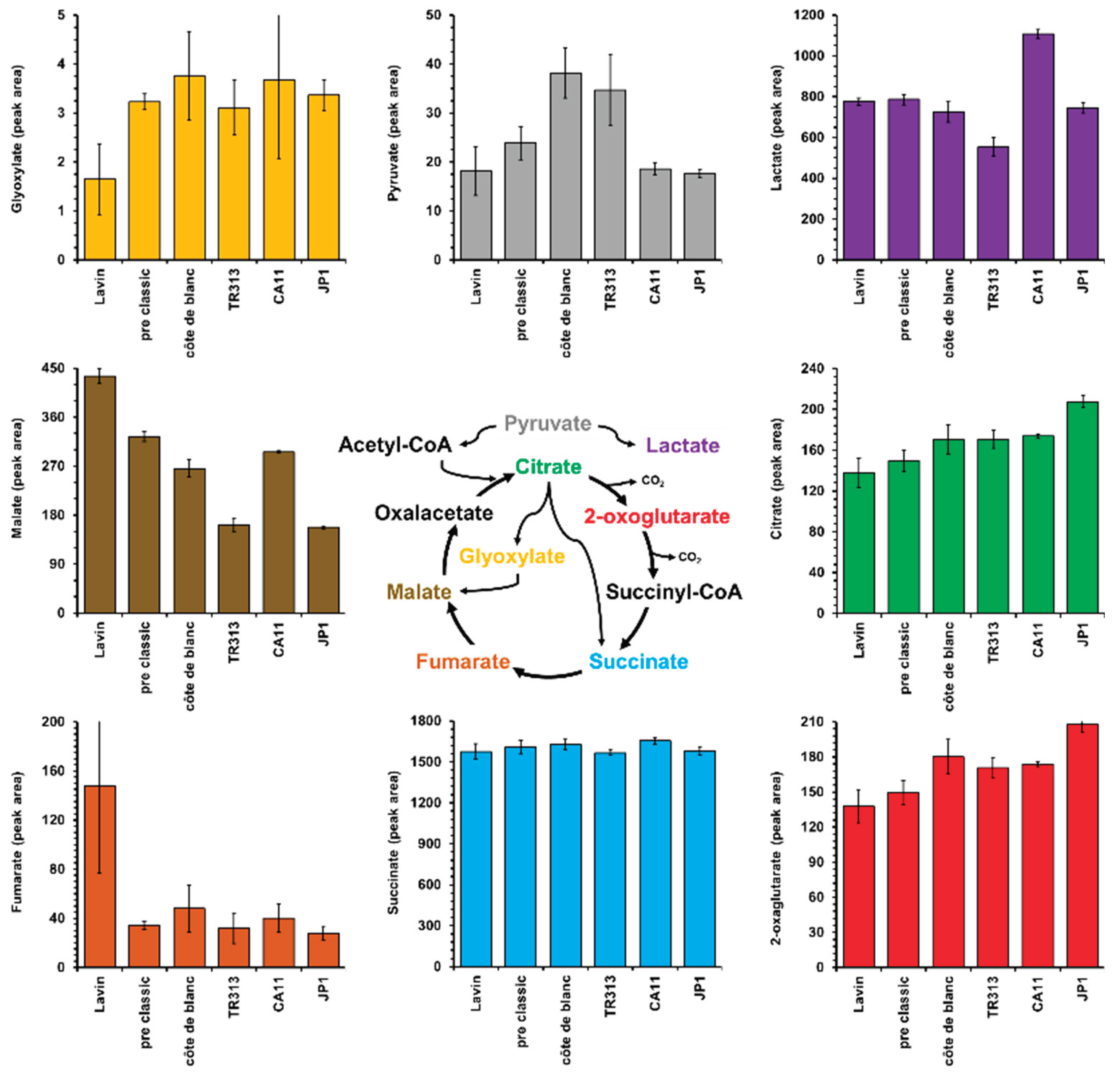
Figure 3.
Fermentative profile of Saccharomyces cerevisiae industrial strains Fermol Distiller JP1 (A) and Renascence TR313 (B) in wort of diluted bee-honey from the Pajeú semiarid hinterland, Pernambuco, Brazil. The consumption of fructose (grey circles), glucose (green circles) and sucrose (red circles), and the production of ethanol (blue circles) and glycerol (yellow circles) were measured up to 30 days of fermentation.
Figure 3.
Fermentative profile of Saccharomyces cerevisiae industrial strains Fermol Distiller JP1 (A) and Renascence TR313 (B) in wort of diluted bee-honey from the Pajeú semiarid hinterland, Pernambuco, Brazil. The consumption of fructose (grey circles), glucose (green circles) and sucrose (red circles), and the production of ethanol (blue circles) and glycerol (yellow circles) were measured up to 30 days of fermentation.
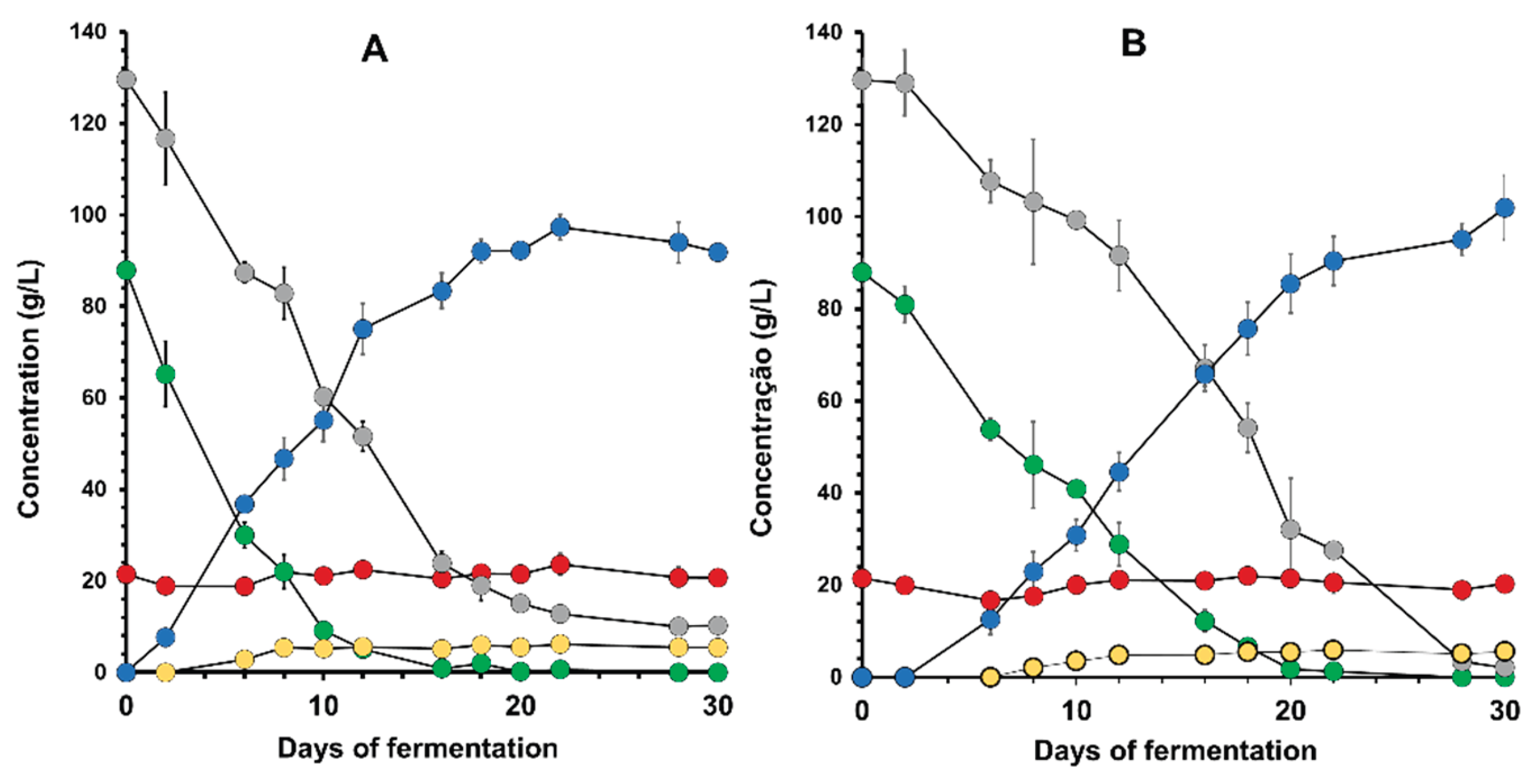
Figure 4.
Recalcitrant sugar present in the bee-honey from the Pajeú semiarid hinterland, Pern[26–28ambuco, Brazil, to the hydrolysis by yeast invertase. Mead containing sugar in the range of 20 g/L at the end of fermentation (closed circles) was mixed with the filtered supernatant of a 48h culture of Saccharomyces cerevisiae Fermol Distiller JP1 strain in YNB-sucrose synthetic medium. A 20 g/L sucrose solution (open circles) was used as a reference for enzyme activity.
Figure 4.
Recalcitrant sugar present in the bee-honey from the Pajeú semiarid hinterland, Pern[26–28ambuco, Brazil, to the hydrolysis by yeast invertase. Mead containing sugar in the range of 20 g/L at the end of fermentation (closed circles) was mixed with the filtered supernatant of a 48h culture of Saccharomyces cerevisiae Fermol Distiller JP1 strain in YNB-sucrose synthetic medium. A 20 g/L sucrose solution (open circles) was used as a reference for enzyme activity.
Figure 5.
Quantification of amino acids (A and B) and aliphatic organic acids (C and D) in the supernatant of honey-fermented must by industrial Saccharomyces cerevisiae strains JP1 (green columns) and TR313 (red columns). Abbreviations: Alanine (ALA), Aspartic acid (ASP), Glutamic acid (GLU), Glycine (GLY), Isoleucine (ISO), Leucine (LEU), Lysine (LYS), Phenylalanine (PHE), Threonine (THR), Tyrosine (TYR), Valine (VAL), Proline (PRO), 2-Hydroxybutyric acid (2-HBA), 2-Oxovaleric acid (2-OVA), 3-Methyl-2-oxovaleric acid (3-MOVA), 4-Aminobutyric acid (4-ABA), Adipic acid (AdA), Cabamic acid (CbA), Dehydroascorbic acid (DHAA), Glutaric acid (GtA), Glyceric acid (GyA) and 3-Aminobutyric acid (3-ABA).
Figure 5.
Quantification of amino acids (A and B) and aliphatic organic acids (C and D) in the supernatant of honey-fermented must by industrial Saccharomyces cerevisiae strains JP1 (green columns) and TR313 (red columns). Abbreviations: Alanine (ALA), Aspartic acid (ASP), Glutamic acid (GLU), Glycine (GLY), Isoleucine (ISO), Leucine (LEU), Lysine (LYS), Phenylalanine (PHE), Threonine (THR), Tyrosine (TYR), Valine (VAL), Proline (PRO), 2-Hydroxybutyric acid (2-HBA), 2-Oxovaleric acid (2-OVA), 3-Methyl-2-oxovaleric acid (3-MOVA), 4-Aminobutyric acid (4-ABA), Adipic acid (AdA), Cabamic acid (CbA), Dehydroascorbic acid (DHAA), Glutaric acid (GtA), Glyceric acid (GyA) and 3-Aminobutyric acid (3-ABA).
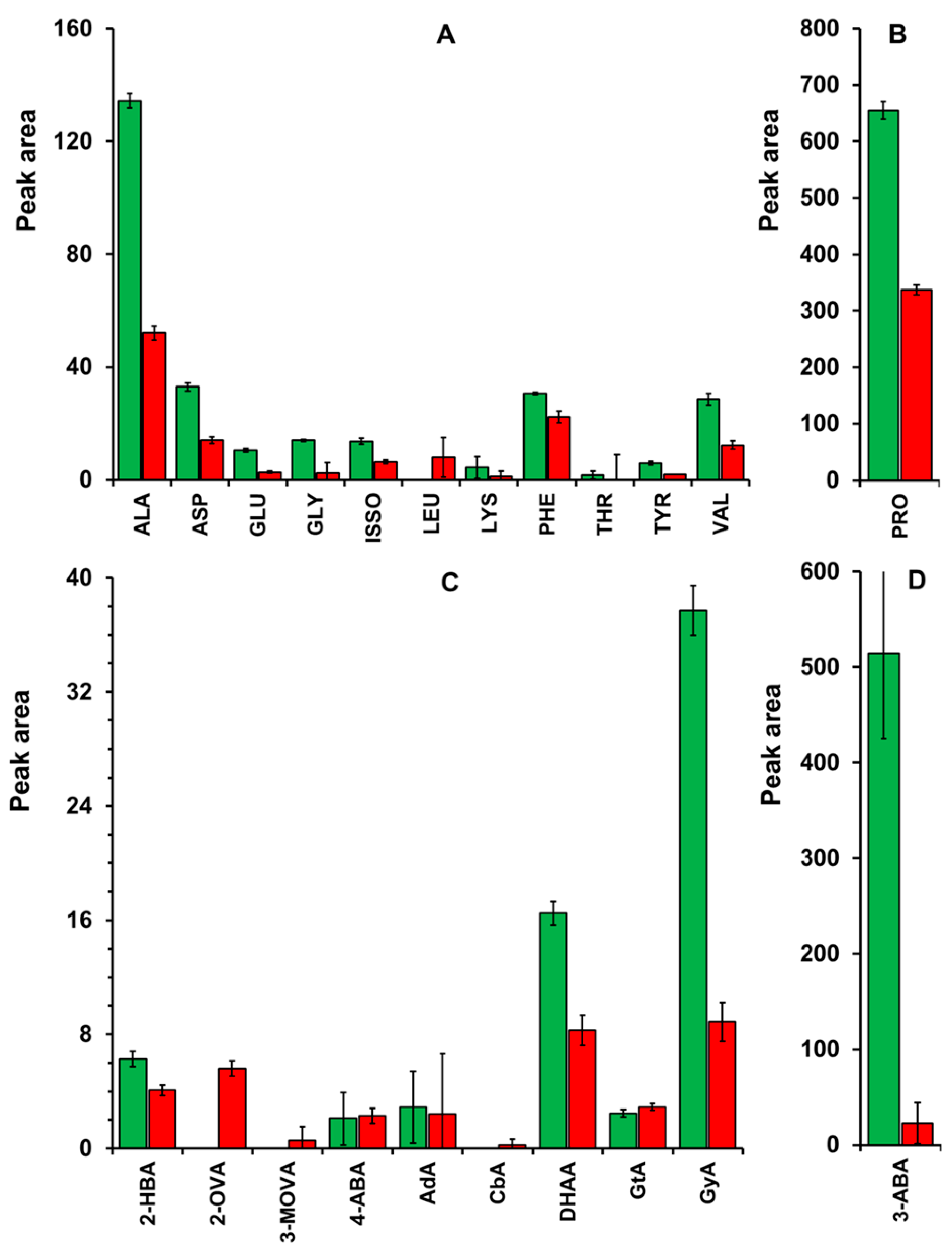
Figure 6.
Profile of volatile organic compounds produced by the Saccharomyces cerevisiae industrial strains Fermol Distiller JP1 (green columns) and Renascence TR313 (red columns) at the end of mead fermentation using diluted bee-honey from the Pajeú semiarid hinterland, Pernambuco, Brazil. Abbreviations: 2,3-Butanediol (2,3-BDO), 2-phenylethanol (2-PE), 4-Hydroxyphenylethanol (4-HPE), p-Anisic acid (ANAc), p-anisaldehyde (pANAd) and p-anisyl alcohol (pANOH).
Figure 6.
Profile of volatile organic compounds produced by the Saccharomyces cerevisiae industrial strains Fermol Distiller JP1 (green columns) and Renascence TR313 (red columns) at the end of mead fermentation using diluted bee-honey from the Pajeú semiarid hinterland, Pernambuco, Brazil. Abbreviations: 2,3-Butanediol (2,3-BDO), 2-phenylethanol (2-PE), 4-Hydroxyphenylethanol (4-HPE), p-Anisic acid (ANAc), p-anisaldehyde (pANAd) and p-anisyl alcohol (pANOH).
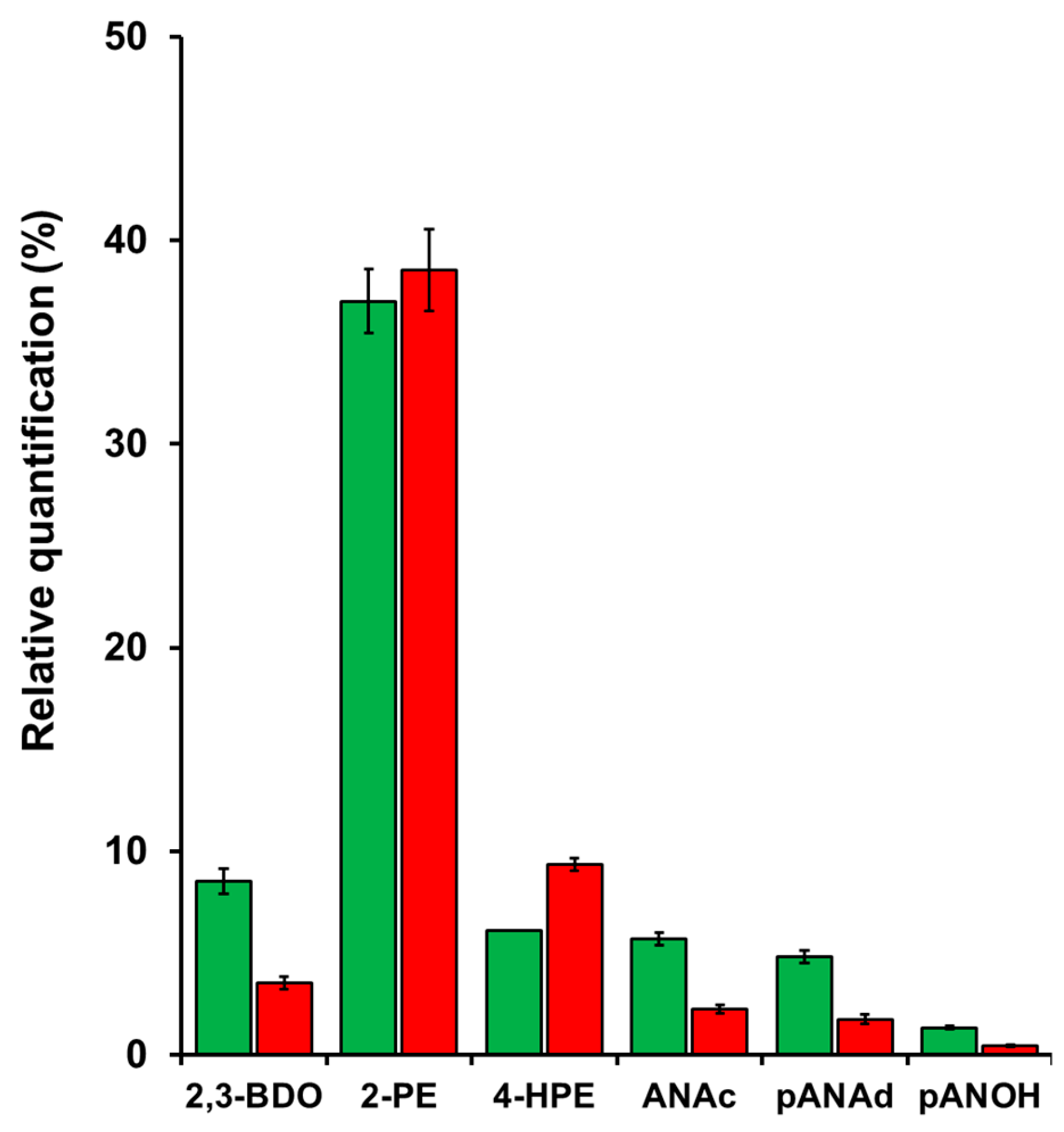
Figure 7.
Fermentative profile of Saccharomyces cerevisiae industrial strains Fermol Distiller JP1 in wort of diluted bee-honey from the Pajeú semiarid hinterland, Pernambuco, Brazil with supplemented must (A), high density of cells (B) and immobilized cells (C). The consumption of fructose (grey circles), glucose (green circles) and sucrose (red circles), and the production of ethanol (blue circles) and glycerol (yellow circles) were measured up to 30 days of fermentation.
Figure 7.
Fermentative profile of Saccharomyces cerevisiae industrial strains Fermol Distiller JP1 in wort of diluted bee-honey from the Pajeú semiarid hinterland, Pernambuco, Brazil with supplemented must (A), high density of cells (B) and immobilized cells (C). The consumption of fructose (grey circles), glucose (green circles) and sucrose (red circles), and the production of ethanol (blue circles) and glycerol (yellow circles) were measured up to 30 days of fermentation.
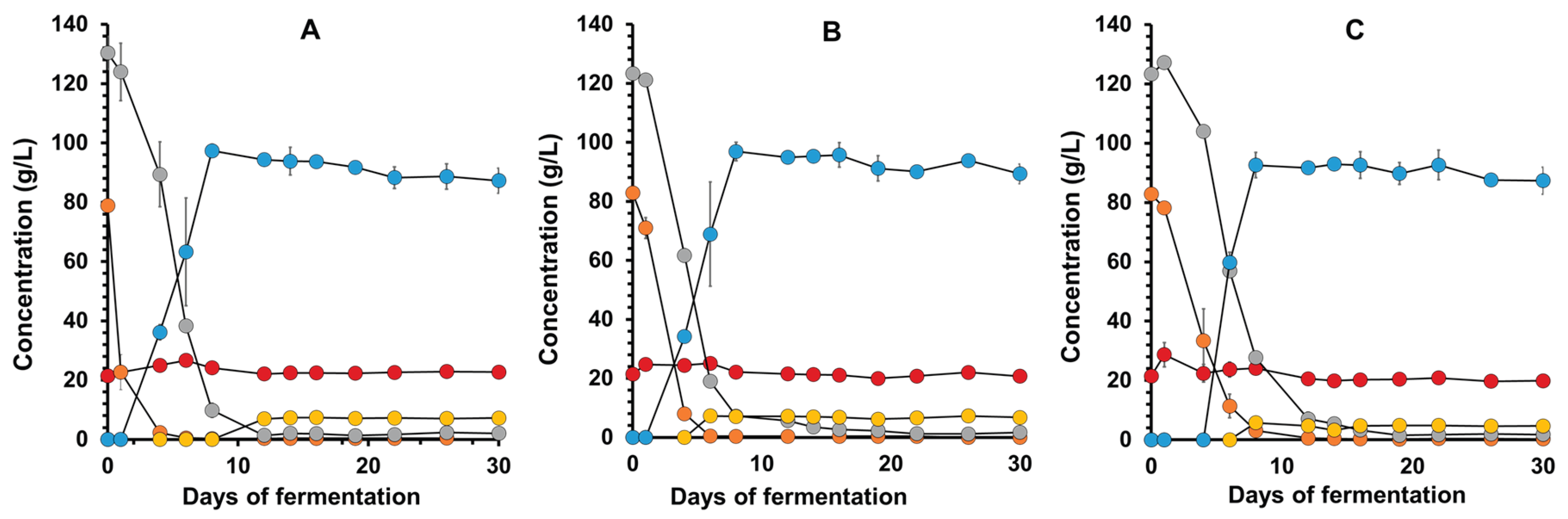

Table 1.
Industrial yeast strains used in this work.
| Code | Commercial name | Supplier | Product | Sensorial characteristics* |
|---|---|---|---|---|
| Sc01 | Saf-Instant | Lesaffre (France) | Bread | Neutral |
| Sc02 | CA-11 | LNF (Brazil) | Cachaça | Neutral |
| Sc03 | LalBrew Diamond | Lallemand (Austria) | German lager beer | Neutral |
| Sc04 | Red Star Premier | Lesaffre (France) | White wine and mead | Neutral, butter, bready |
| Sc05 | LalBrew Belle saison | Lallemand (Austria) | Belgium beer | Fruity and spicy |
| Sc06 | Renascence TR313 | Renaissance (Canada) | White wine | Passion fruits and berries |
| Sc07 | Lalvin k1v-1116 | Lallemand (Canada) | Red wine and mead | Floral aroma |
| Sc08 | Côte de blancs | Fermentis (Belgium) | White wine and mead | Fruity aroma |
| Sc09 | Premier classic | Fermentis (Belgium) | White wine and mead | Fruity aroma |
| Sc10 | Fermol Distiller JP1 | LNF (Brazil) | Cachaça and fuel-ethanol | Neutral |
* information provided by the manufacturer.
Table 2.
Physiological parameters of the fermentation of diluted bee-honey from Serra Talhada, in the Pajeú semiarid hinterland, Pernambuco, Brazil. Initial sugar of 220±11.5 g/L (oBrix = 12.6±1.6).
Table 2.
Physiological parameters of the fermentation of diluted bee-honey from Serra Talhada, in the Pajeú semiarid hinterland, Pernambuco, Brazil. Initial sugar of 220±11.5 g/L (oBrix = 12.6±1.6).
| Yeast strain | Code | Residual sugars (g/L) | Attenuation | Ethanol (%) |
|---|---|---|---|---|
| Saf-Instant | Sc01 | 44.6±0.7 | 80% | 7.91±0.18 |
| CA-11 | Sc02 | 49.2±2.7 | 78% | 7.91±0.11 |
| Lallemand Diamond | Sc03 | 22.7±1.4 | 90% | 8.79±0.07 |
| Red Star Premier | Sc04 | 49.4±1.2 | 78% | 7.95±0.32 |
| LalBrew Belle saison | Sc05 | 52.9±1.1 | 76% | 7.61±0.13 |
| Renascence TR313 | Sc06 | 47.0±2.0 | 79% | 7.61±0.13 |
| Lalvin k1v-1116 | Sc07 | 22.1±0.5 | 90% | 9.08±0.17 |
| Côte de blancs | Sc08 | 30.4±8.2 | 86% | 6.50±0.14 |
| Premier classic | Sc09 | 30.9±5.1 | 86% | 7.99±0.17 |
| Fermol distiller JP1 | Sc10 | 39.9±0.6 | 82% | 9.66±0,19 |
Table 3.
Physiological profile of bee honey fermentation of different origins in the semiarid area of Pernambuco, Brazil, by the industrial strain Saccharomyces cerevisiae JP1 for 8 days.
Table 3.
Physiological profile of bee honey fermentation of different origins in the semiarid area of Pernambuco, Brazil, by the industrial strain Saccharomyces cerevisiae JP1 for 8 days.
| Fermentation parameter | Honey origin | ||
|---|---|---|---|
| IB/M | ST/P | TF/P | |
| Initial sugar (g/L) | 274.9±38.1 | 284.6±8.4 | 281.5±23.3 |
| Residual sugar (g/L) | 78.5±4.8 | 70.0±1.5 | 55.1±3.1 |
| Consumed sugar (g/L) | 196.4±15.5 | 214.7±9.6 | 226.4±3.1 |
| Ethanol produced (g/L) | 79.7±16.2 | 88.3±3.3 | 86.1±4.7 |
| Glycerol produced (g/L) | 10.0±1.7 | 6.1±0.9 | 10.2±0.7 |
| Ethanol yield (g/g) | 0.41 | 0.41 | 0.38 |
| Glycerol yield (g/g) | 0.05 | 0.03 | 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



