
Submitted:
16 December 2025
Posted:
16 December 2025
You are already at the latest version
Abstract
Hybridization is considered an important process in plant evolution, especially in the origin domesticated plant taxa, with many crop species being the result of interspecific hybridization events. There are several unidentified lineages of Spondias in the Northeastern region of Brazil known only by vernacular names such as ‘cajaguela’, ‘umbu-cajá’, and ‘umbuguela’. These taxa are often regarded as being of hybrid origin, based on supposedly intermediate morphological features. However, the morphology-based hypotheses of hybrid origin and parentage of these Spondias taxa remains largely untested experimentally. We collected 355 accessions of Spondias including the putative hybrid taxa and both native and introduced species. We then reconstructed phylogenies of plastid and nuclear markers, and also haplotype networks in order to ascertain the genetic affinities between putative hybrids and other Spondias species. All taxa with intermediate morphology were confirmed as hybrids between their putative parental species. All hybrids involving S. purpurea (native to Mexico) appear to be F1 generation. The recently described Spondias bahiensis is shown to have originated from hybridization between S. tuberosa and S. venulosa. The other ‘umbu-cajá’ taxon found in Northeastern of Brazil is revealed to be the result of hybridization between S. mombin and S. tuberosa. Both the northern ‘umbu-cajá’ taxon and S. bahiensis appear to be well-established hybrid lineages and not early generation hybrids. Additionally, some introgression and backcrossing processes between S. bahiensis and one of the parents was also observed. Our findings confirm the hybrid origins of the domesticated Spondias taxa found in Northeastern Brazil.
Keywords:
1. Introduction
2. Results
3. Discussion
3.1. Hybrid Origins of Spondias Taxa Found in Northeastern Brazil
3.2. Mode and Time of Origin of the Spondias Hybrids from Northeastern Brazil
4. Materials and Methods
4.1. Taxon Sampling
4.2. DNA Extraction, Amplification and Sequencing
4.3. Sequence Edition, Alignment and Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stebbins, G.L. The role of hybridization in evolution. Proc. Amer. Phil. Soc. 1959, 103, 231–251. [Google Scholar]
- Raven, P.H. Systematics and plant population biology. Syst. Bot. 1976, 1, 284–316. [Google Scholar] [CrossRef]
- Rieseberg, L.H. The role of hybridization in evolution: Old wine in new skins. Am. J. Bot. 1995, 82, 944–953. [Google Scholar] [CrossRef]
- Ellstrand, N.C.; Whitkus, R.; Rieseberg, L.H. Distribution of spontaneous plant hybrids. Proc. Natl. Acad. Sci. USA 1996, 93, 5090–5093. [Google Scholar] [CrossRef]
- Arnold, M.L. Natural Hybridization and Evolution; Oxford University Press: New York, NY, USA, 1997. [Google Scholar]
- Levin, D.A. The Origin, Expansion, and Demise of Plant Species; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Rieseberg, L.H.; Raymond, O.; Rosenthal, D.M.; Lai, Z.; Livingstone, K.; Nakazato, T.; Durphy, T.L.; Schwarzbach, A.E.; Donovan, L.A.; Lexer, C. Major ecological transitions in wild sunflowers facilitated by hybridization. Science 2003, 301, 1211–1216. [Google Scholar] [CrossRef]
- Soltis, P.S.; Soltis, D.E. The role of hybridization in plant speciation. Annu. Rev. Plant Biol. 2009, 60, 561–588. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, Y. Evolution of polyploid Triticum wheats under cultivation: The role of domestication, natural hybridization and allopolyploid speciation in their diversification. Plant Cell Physiol. 2011, 52, 750–764. [Google Scholar] [CrossRef]
- Miller, A.J.; Gross, B.L. From forest to field: Perennial fruit crop domestication. Am. J. Bot. 2011, 98, 1389–1414. [Google Scholar] [CrossRef]
- Robinson, J.P.; Harris, S.A.; Juniper, B.E. Taxonomy of the genus Malus Mill. (Rosaceae) with emphasis on the cultivated apple, Malus domestica Borkh. Plant Syst. Evol. 2001, 226, 35–58. [Google Scholar] [CrossRef]
- Nikiforova, S.V.; Cavalieri, D.; Velasco, R.; Goremykin, V. Phylogenetic analysis of 47 chloroplast genomes clarifies the contribution of wild species to the domesticated apple maternal line. Mol. Biol. Evol. 2013, 30, 1751–1760. [Google Scholar] [CrossRef]
- Cornille, A.; Giraud, T.; Smulders, M.J.M.; Roldán-Ruiz, I.; Gladieux, P. The domestication and evolutionary ecology of apples. Trends Genet. 2014, 30, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Simmonds, N.W.; Shepherd, K. The taxonomy and origins of cultivated bananas. Bot. J. Linn. Soc. 1955, 55, 302–312. [Google Scholar] [CrossRef]
- Osoju, J.O.; Harrison, G.; Crouch, J.; Heslop-Harrison, J.S. Identification of the genomic constitution of Musa L. lines (bananas, plantains and hybrids) using molecular cytogenetics. Ann. Bot. 1997, 80, 787–793. [Google Scholar] [CrossRef]
- Perrier, X.; De Langhe, E.; Donohue, M.; Lentfer, C.; Vrydaghs, L.; Bakry, F.; Carreel, F.; Hippolyte, I.; Horry, J.-P.; Jenny, C.; et al. Multidisciplinary perspectives on banana (Musa spp.) domestication. Proc. Natl. Acad. Sci. USA 2011, 108, 11311–11318. [Google Scholar] [CrossRef]
- Cipriani, G.; Testolin, R.; Morgante, M. Paternal inheritance of plastids in interspecific hybrids of the genus Actinidia revealed by PCR-amplification of chloroplast DNA fragments. Mol. Gen. Genet. 1995, 247, 693–697. [Google Scholar] [CrossRef]
- Li, J.; Huang, H.; Sang, T. Molecular phylogeny and infrageneric classification of Actinidia (Actinidiaceae). Syst. Bot. 2002, 27, 408–415. [Google Scholar]
- Chat, J.; Jáuregui, B.; Petit, R.J.; Nadot, S. Reticulate evolution in kiwifruit (Actinidia, Actinidiaceae) identified by comparing their maternal and paternal phylogenies. Am. J. Bot. 2004, 91, 736–747. [Google Scholar] [CrossRef]
- NNicolosi, E.; Deng, Z.N.; Gentile, A.; La Malfa, S.; Continella, G.; Tribulato, E. Citrus phylogeny and genetic origin of important species as investigated by molecular markers. Theor. Appl. Genet. 2000, 100, 1155–1166. [Google Scholar] [CrossRef]
- Li, X.; Xie, R.; Lu, Z.; Zhou, Z. The origin of cultivated Citrus as inferred from Internal Transcribed Spacer and chloroplast DNA sequence and Amplified Fragment Length Polymorphism fingerprints. J. Am. Soc. Hortic. Sci. 2010, 135, 341–350. [Google Scholar] [CrossRef]
- Xu, Q.; Chen, L.-L.; Ruan, X.; Chen, D.; Zhu, A.; Chen, C.; Bertrand, D.; Jiao, W.-B.; Hao, B.-H.; Lyon, M.P.; et al. The draft genome of sweet orange (Citrus sinensis). Nat. Genet. 2013, 45, 59–66. [Google Scholar] [CrossRef]
- Wu, G.A.; Prochnik, S.; Jenkins, J.; Salse, J.; Hellsten, U.; Murat, F.; Perrier, X.; Ruiz, M.; Scalabrin, S.; et al. Sequencing of diverse mandarin, pummelo and orange genomes reveals complex history of admixture during Citrus domestication. Nat. Biotechnol. 2014, 32, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.D.; Daly, D.C.; Randrianasolo, A. The first report of Spondias native to Madagascar: Spondias tefyi, sp. nov. (Anacardiaceae). Brittonia 2012, 64, 263–267. [Google Scholar] [CrossRef]
- Miller, A.J.; Schaal, B.A. Domestication of a Mesoamerican cultivated fruit tree, Spondias purpurea. Proc. Natl. Acad. Sci. USA 2005, 102, 12801–12806. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.J. Characterization of a domesticated tree lineage (Spondias purpurea, Anacardiaceae) based on nuclear and chloroplast sequence data. J. Torrey Bot. Soc. 2008, 135, 463–474. [Google Scholar] [CrossRef]
- Urban, I. Plantae Haitienses et Domingenses novae vel rariores VII. a cl. E.L. Ekman 1924-1928 lectae. Ark. Bot. 1929, 22A, 1–115. [Google Scholar]
- Machado, M.C.; Carvalho, P.C.L.; van den Berg, C. Domestication, hybridization, speciation, and the origins of an economically important tree crop of Spondias (Anacardiaceae) from the Brazilian Caatinga dry forest. Neodiversity 2015, 8, 8–49. [Google Scholar] [CrossRef]
- Carvalho, P.C.L. Variabilidade morfológica, avaliação agronômica, filogenética e citogenética em Spondias (Anacardiaceae) no nordeste do Brasil. Ph.D. Thesis, Universidade Estadual de Feira de Santana, Feira de Santana, Brazil, 2006. [Google Scholar]
- Almeida, C.C.S.; Carvalho, P.C.L; Guerra, M. Karyotype differentiation among Spondias species and the putative hybrid Umbu-cajá (Anacardiaceae). Bot. J. Linn. Soc. 2007, 155, 541–547. [Google Scholar] [CrossRef]
- Souza, F.X. Spondias agroindustriais e os seus métodos de propagação. Doc. Embrapa-CNPAT 1998, 27, 1–28. [Google Scholar]
- Carvalho, P.C.L.; Soares-Filho, W.S.; Ritzinger, R.; Carvalho, J.A.B.S. Conservação de germoplasma de fruteiras tropicais com a participação do agricultor. Rev. Bras. Frutic. 2001, 23, 730–734. [Google Scholar] [CrossRef]
- Araújo, F.P.; Santos, C.A.F.; Oliveira, V.R. Fruticultura de sequeiro: Uma janela para o desenvolvimento sustentável. Instruções Técnicas da Embrapa Semiárido 2006, *73*, 1–6. Available online: http://www.infoteca.cnptia.embrapa.br/handle/doc/157633 (accessed on 9 December 2013).
- Ritzinger, R.; Soares-Filho, W.S.; Carvalho, P.C.L. Potencial da umbucajazeira para regiões semi-áridas da Bahia; Embrapa Mandioca e Fruticultura Tropical: Cruz das Almas, Brazil, 2006. [Google Scholar]
- Ritzinger, R.; Soares-Filho, W.S.; Carvalho, P.C.L. Evaluation of umbu-cajá germplasm in the state of Bahia, Brazil. Crop Breed. Appl. Biotechnol. 2008, 8, 181–186. [Google Scholar] [CrossRef]
- Ritzinger, R.; Soares-Filho, W.S.; Castellén, M.S. Coleção de Spondias sp. da Embrapa Mandioca e Fruticultura Tropical. In Spondias no Brasil: Umbu, Cajá e Espécies Afins; Lêderman, I.E., Lira-Júnior, J.S., Silva-Júnior, J.F., Eds.; IPA/UFPE: Recife, Brazil, 2008; pp. 86–90. [Google Scholar]
- Souza, F.X.; Costa, J.T.A. Produção de mudas das Spondias cajazeira, cajaraneira, cirigueleira, umbu-cajazeira e umbuzeiro. Doc. Embrapa Agroind. Trop. 2010, 133, 1–26. [Google Scholar]
- Soares-Filho, W.S. Prospecção e seleção de genótipos de umbu-cajazeira no estado da Bahia. In Botânica e Desenvolvimento Sustentável; Lucena, E.M.P., Amorim, A.V., Eds.; EdUECE: Fortaleza, Brazil, 2011; p. 371. [Google Scholar]
- Silva-Júnior, J.F.; Lêdo, A.S.; Silva, A.V.C.; Ramos, S.R.R. Recursos genéticos de fruteiras nativas e adaptadas do nordeste: Situação do germoplasma conservado ex situ na região. In Proceedings of the Congresso Brasileiro de Recursos Genéticos, 2, Belém, PA, Brazil, 2012; Sociedade Brasileira de Recursos Genéticos: Brasília, Brazil, 2012.
- Romano, M.R.; Soares-Filho, W.S.; Ritzinger, R.; Fonseca, N.; Machado, C.F. Aspectos técnicos introdutórios ao emprego de espôndias nativas do Nordeste brasileiro em Sistemas Agroflorestais. Comun. Téc. Embrapa Mandioca Frutic. 2013, 153, 1–6. [Google Scholar]
- Mason-Gamer, R.J.; Holsinger, K.E.; Jansen, R.K. Chloroplast DNA haplotype variation within and among populations of Coreopsis grandiflora (Asteraceae). Mol. Biol. Evol. 1995, 12, 371–381. [Google Scholar] [CrossRef]
- Soltis, D.E.; Kuzoff, R.K. Discordance between nuclear and chloroplast phylogenies in the Heuchera group (Saxifragaceae). Evolution 1995, 49, 727–742. [Google Scholar] [CrossRef] [PubMed]
- Kellogg, E.A.; Appels, R.; Mason-Gamer, R.J. When genes tell different stories: The diploid genera of Triticeae (Gramineae). Syst. Bot. 1996, 21, 321–347. [Google Scholar] [CrossRef]
- Rieseberg, L.H.; Whitton, J.; Linder, C.R. Molecular marker incongruence in plant hybrid zones and phylogenetic trees. Acta Bot. Neerl. 1996, 45, 243–262. [Google Scholar] [CrossRef]
- Sang, T.; Crawford, D.J.; Stuessy, T.F. Chloroplast DNA phylogeny, reticulate evolution, and biogeography of Paeonia (Paeoniaceae). Am. J. Bot. 1997, 84, 1120–1136. [Google Scholar] [CrossRef]
- Seelanan, T.; Schnabel, A.; Wendel, J.F. Congruence and consensus in the cotton tribe (Malvaceae). Syst. Bot. 1997, 22, 259–290. [Google Scholar] [CrossRef]
- Sang, T.; Zhong, Y. Testing hybridization hypotheses based on incongruent gene trees. Syst. Biol. 2000, 49, 422–434. [Google Scholar] [CrossRef]
- Ferguson, C.J.; Jansen, R.K. A chloroplast DNA phylogeny of eastern Phlox (Polemoniaceae): Implications of congruence and incongruence with the ITS phylogeny. Am. J. Bot. 2002, 89, 1324–1335. [Google Scholar] [CrossRef]
- Funk, D.J.; Omland, K.E. Species-level paraphyly and polyphyly: Frequency, causes, and consequences, with insights from animal mitochondrial DNA. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 397–423. [Google Scholar] [CrossRef]
- Linder, C.R.; Rieseberg, L.H. Reconstructing patterns of reticulate evolution in plants. Am. J. Bot. 2004, 91, 1700–1708. [Google Scholar] [CrossRef] [PubMed]
- Weeks, A.; Simpson, B.B. Molecular genetic evidence for interspecific hybridization among endemic Hispaniolan Bursera (Burseraceae). Am. J. Bot. 2004, 91, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Barber, J.C.; Finch, C.C.; Francisco-Ortega, J.; Santos-Guerra, A.; Jansen, R.K. Hybridization in Macaronesian Sideritis (Lamiaceae): Evidence from incongruence of multiple independent nuclear and chloroplast sequence datasets. Taxon 2007, 56, 74–88. [Google Scholar] [CrossRef]
- Soltis, D.E.; Mavrodiev, E.V.; Doyle, J.J.; Rauscher, J.; Soltis, P.S. ITS and ETS sequence data and phylogeny reconstruction in allopolyploids and hybrids. Syst. Bot. 2008, 33, 7–20. [Google Scholar] [CrossRef]
- Lo, E.Y.Y. Testing hybridization hypotheses and evaluating the evolutionary potential of hybrids in mangrove plant species. J. Evol. Biol. 2010, 23, 2249–2261. [Google Scholar] [CrossRef]
- Laureto, P.J.; Barkman, T.J. Nuclear and chloroplast DNA suggest a complex single origin for the threatened allopolyploid Solidago houghtonii (Asteraceae) involving reticulate evolution and introgression. Syst. Bot. 2011, 36, 209–226. [Google Scholar] [CrossRef]
- Cires, E.; Baltisberger, M.; Cuesta, C.; Vargas, P.; Fernández-Prieto, J.A. Allopolyploid origin of the Balkan endemic Ranunculus wettsteinii (Ranunculaceae) inferred from nuclear and plastid DNA sequences. Org. Divers. Evol. 2014, 14, 1–10. [Google Scholar] [CrossRef]
- Mogensen, H.L. The hows and whys of cytoplasmic inheritance in seed plants. Am. J. Bot. 1996, 83, 383–404. [Google Scholar]
- Zhang, Q.; Sodmergen. Why does biparental plastid inheritance revive in angiosperms? J. Plant Res. 2010, 123, 201–206. [Google Scholar] [CrossRef]
- Birky, C.W. Uniparental inheritance of organelle genes. Curr. Biol. 2008, 18, R692–R695. [Google Scholar] [CrossRef]
- Álvarez, I.; Wendel, J.F. Ribosomal ITS sequences and plant phylogenetic inference. Mol. Phylogenet. Evol. 2003, 29, 417–434. [Google Scholar] [CrossRef] [PubMed]
- Fuertes Aguilar, J.; Rosselló, J.A.; Nieto Feliner, G. Nuclear ribosomal DNA (nrDNA) concerted evolution in natural and artificial hybrids of Armeria (Plumbaginaceae). Mol. Ecol. 1999, 8, 1341–1346. [Google Scholar] [CrossRef] [PubMed]
- Franzke, A.; Mummenhoff, K. Recent hybrid speciation in Cardamine (Brassicaceae): Conversion of nuclear ribosomal ITS sequences in statu nascendi. Theor. Appl. Genet. 1999, 98, 831–834. [Google Scholar] [CrossRef]
- Miller, A.J.; Schaal, B.A. Domestication and the distribution of genetic variation in wild and cultivated populations of the Mesoamerican fruit tree Spondias purpurea L. (Anacardiaceae). Mol. Ecol. 2006, 15, 1467–1480. [Google Scholar] [CrossRef]
- Miller, A.J. Spondias. In Wild Crop Relatives: Genomic and Breeding Resources, Tropical and Subtropical Fruits; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 203–212. [Google Scholar] [CrossRef]
- Kiill, L.H.P.; Silva, T.A.; Araújo, F.P. Fenologia reprodutiva de espécies e híbridos do gênero Spondias L. (Anacardiaceae) em Petrolina, PE. Bol. Pesqui. Desenvolv. Embrapa Semiárido 2013, 110, 1–23. [Google Scholar]
- Ramos, M.C. Ecologia da polinização de taperebá (Spondias mombin L., Anacardiaceae) em área de floresta secundária no município de Santo Antônio do Tauá, Pará, Brasil. M.Sc. Thesis, Universidade Federal do Pará, Belém, Brazil, 2009. [Google Scholar]
- Alves, R.M.; Oliveira, C.C.; Fernandes, J.R.Q.; Oliveira, H.O. Estudo fenológico do taperebazeiro, Spondias mombin L., em sistema agroflorestal, no município de Santa Bárbara, Pará. In Anais do Congresso Brasileiro de Recursos Genéticos, 2, Belém, PA, Brazil, 2012; Sociedade Brasileira de Recursos Genéticos: Brasília, Brazil, 2012.
- Barbosa, D.C.A.; Alves, J.L.H.; Prazeres, S.M.; Paiva, A.M.A. Dados fenológicos de 10 espécies arbóreas de uma área de Caatinga (Alagoinha-PE). Acta Bot. Bras. 1989, 3, 109–118. [Google Scholar] [CrossRef]
- Nadia, T.L.; Machado, I.C.; Lopes, A.V. Polinização de Spondias tuberosa Arruda (Anacardiaceae) e análise da partilha de polinizadores com Ziziphus joazeiro Mart. (Rhamnaceae), espécies frutíferas e endêmicas da caatinga. Rev. Bras. Bot. 2007, 30, 89–100. [Google Scholar] [CrossRef]
- Lins-Neto, E.M.F.; Almeida, A.L.S.; Peroni, N.; Castro, C.C.; Albuquerque, U.P. Phenology of Spondias tuberosa Arruda (Anacardiaceae) under different landscape management regimes and a proposal for a rapid phenological diagnosis using local knowledge. J. Ethnobiol. Ethnomed. 2013, 9, 10. [Google Scholar] [CrossRef]
- Pinto, J.A. Diccionário de Botânica Brasileira ou Compêndio dos Vegetaes do Brasil, tanto Indígenas como Acclimatados; Typographia Perseverança: Rio de Janeiro, Brazil, 1873; p. 435. [Google Scholar]
- Engler, A. Anacardiaceae. In Flora Brasiliensis; Martius, C.F.P., Ed.; Fleischer: Munich, Germany, 1876; Volume 12, pp. 367–418. [Google Scholar]
- Braga, R. Plantas do Ceará. Rev. Inst. Ceará 1955, 69, 100–119. [Google Scholar]
- Bediaga, B. Joining pleasure and work in the making of science: The Jardim Botânico do Rio de Janeiro - 1808 to 1860. Hist. Cienc. Saude-Manguinhos 2007, 14, 1131–1157. [Google Scholar] [CrossRef]
- Pombo, N. As “riquezas do mundo” - Cobiça e ciência nos jardins botânicos de Caiena e Belém do Pará (1790-1803). Navigator 2010, 6, 52–60. [Google Scholar]
- Duss, R.P. Flore Phanérogamique des Antilles Françaises (Martinique et Guadeloupe); Protat Frères Imprimeurs: Mâcon, France, 1897; Volume 1, p. 256. [Google Scholar]
- Mazé, H. Contribution to the Flora of Guadeloupe.; The Government Printing: Basse-Terre, p. 191.
- Descourtilz, M.E. Flore Pittoresque et Médicale des Antilles; Pichard: Paris, France, 1827; Volume 5, p. 292. [Google Scholar]
- D’Orbigny, M.A. Voyage Pittoresque dans les Deux Amériques; Furne et Cie Libraires: Paris, France, 1841; p. 568. [Google Scholar]
- Peckolt, T. História das Plantas Alimentares e de Gozo do Brasil; H. Laemmert & C.: Rio de Janeiro, Brazil, 1871; Volume 5, p. 167. [Google Scholar]
- Barbosa Rodrigues, J. Hortus Fluminensis, ou, Breve Noticia sobre as Plantas Cultivadas no Jardim Botanico do Rio de Janeiro para Servir de Guia aos Visitantes; Leuzinger: Rio de Janeiro, Brazil, 1895; p. 388. [Google Scholar]
- Braga, R. Plantas do Nordeste, especialmente do Ceará, 2nd ed.; Imprensa Oficial: Fortaleza, Brazil, 1960; p. 540. [Google Scholar]
- Blog Visão Norte. A origem da seriguela na serra da Meruoca. 2014. Available online: http://visaonorte.blogspot.com.br/2013/01/a-origem-da-seriguela-na-serra-da.html (accessed on 3 March 2014).
- Carvalho, P.C.L.; Ritzinger, R.; Soares-Filho, W.S.; Lêdo, C.A.S. Características morfológicas, físicas e químicas de frutos de populações de umbu-cajazeira no estado da Bahia. Rev. Bras. Frutic. 2008, 30, 140–147. [Google Scholar] [CrossRef]
- Santos, L.A.; Dantas, A.C.V.L.; Fonseca, A.A.O.; Almeida, V.O.; Barroso, J.P. Caracterização química e físico-química de frutos de umbu-cajazeiras do semi-árido da Bahia por meio de análise multivariada. In Proceedings of the III Encontro da Rede de Recursos Genéticos Vegetais da Bahia, II Simpósio de Recursos Genéticos de plantas cultivadas no Nordeste Brasileiro, Vitória da Conquista, Brazil, 2008; UESB: Vitória da Conquista, Brazil, 2008.
- Santos, L.A.; Dantas, A.C.V.L.; Vilarinhos, A.D.; Fonseca, A.A.O.; Barroso, J.P. Classificação de frutos de umbu-cajazeira para consumo in natura e processado. In Proceedings of the Jornada Científica Embrapa Mandioca e Fruticultura Tropical, 3, Cruz das Almas, BA, Brazil, 2009; Embrapa Mandioca e Fruticultura Tropical: Cruz das Almas, Brazil, 2009.
- Santos, M.B. Conservação da polpa de umbu-cajá (Spondias spp.) por métodos combinados. M.Sc. Thesis, Universidade Federal do Recôncavo da Bahia, Cruz das Almas, Brazil, 2009. [Google Scholar]
- Santana, I.B.B.; Soares-Filho, W.S.; Ritzinger, R.; Amorim, E.P.; Costa, M.A.P.C.; Cruz, E.S.; Luquine, L.S. Análise conjunta de caracteres quantitativos e qualitativos em acessos de umbu-cajazeira. In Proceedings of the XXI Congresso Brasileiro de Fruticultura, Natal, RN, Brazil, 2010; Sociedade Brasileira de Fruticultura: Natal, Brazil, 2010.
- Santos, L.A. Caracterização de frutos e molecular de umbu-cajazeiras (Spondias sp.) no semiárido da Bahia. M.Sc. Thesis, Universidade Federal do Recôncavo da Bahia, Cruz das Almas, Brazil, 2010. [Google Scholar]
- Lima, M.S.S. Seleção e propagação de genótipos de umbu-cajazeira (Spondias sp.) da região semiárida da Bahia. M.Sc. Thesis, Universidade Federal do Recôncavo da Bahia, Cruz das Almas, Brazil, 2012. [Google Scholar]
- Santos, A.P.; Dantas, A.C.V.L.; Fonseca, A.A.O.; Lêdo, C.A.S.; Almeida, V.A.; Fonseca, M.D.S. Caracterização de frutos de umbu-cajazeiras das microrregiões de Santo Antônio de Jesus, Feira de Santana e Jequié, Bahia. Magistra 2012, 24, 271–279. [Google Scholar]
- Santana, I.B.B. Divergência genética entre acessos de umbu-cajazeira mediante análise multivariada utilizando marcadores morfoagronômicos e moleculares. M.Sc. Thesis, Universidade Federal do Recôncavo da Bahia, Cruz das Almas, Brazil, 2010. [Google Scholar]
- Santana, I.B.B.; Oliveira, E.J.; Soares-Filho, W.S.; Ritzinger, R.; Amorim, E.P.; Costa, M.A.P.C.; Moreira, R.F.C. Variabilidade genética entre acessos de umbu-cajazeira mediante análise de marcadores ISSR. Rev. Bras. Frutic. 2011, 33, 868–876. [Google Scholar] [CrossRef]
- Santana, I.B.B.; Soares-Filho, W.S.; Ritzinger, R.; Costa, M.A.P.C. Umbu-cajazeira: Boas perspectivas para o Semiárido baiano. Bahia Agric. 2011, 9, 101–105. [Google Scholar]
- Noronha, M.A.S.; Cardoso, E.A.; Dias, N.S. Características físico-químicas de frutos de umbu-cajá Spondias sp. proveniente dos Polos Baixo-Jaguaribe (CE) e Assu-Mossoró (RN). Rev. Bras. Prod. Agroind. 2000, 2, 91–96. [Google Scholar] [CrossRef]
- Lima, A.K.C.; Rezende, L.P.; Câmara, F.A.A.; Nunes, G.H.S. Propagação de cajarana (Spondias sp.) e Ciriguela (Spondias purpurea) por meio de estacas verdes enfolhadas nas condições climáticas de Mossoró-RN. Rev. Caatinga 2002, 15, 33–38. [Google Scholar]
- Silva-Júnior, J.F.; Bezerra, J.E.F.; Lêderman, I.E.; Alves, M.A.; Melo-Neto, M.L. Collecting, ex situ conservation and characterization of ‘cajá-umbu’ (Spondias mombin × Spondias tuberosa) germplasm in Pernambuco State, Brazil. Genet. Resour. Crop Evol. 2004, 51, 343–349. [Google Scholar] [CrossRef]
- Lira-Júnior, J.S. Caracterização molecular, físico-química de frutos e fenológica do banco de germoplasma de cajá-umbu na zona da mata de Pernambuco. M.Sc. Thesis, Universidade Federal Rural de Pernambuco, Recife, Brazil, 2005. [Google Scholar]
- Lira-Júnior, J.S.; Musser, R.S.; Melo, E.A.; Maciel, M.I.S.; Santos, V.F. Caracterização física e físico-química de frutos de cajá-umbu (Spondias spp.). Ciênc. Tecnol. Aliment. 2005, 25, 757–761. [Google Scholar] [CrossRef]
- Gondim et al. 2013; Gondim, P.J.S.; Silva, S.M.; Pereira, W.E.; Dantas, A.L.; Chaves-Neto, J.R.; Santos, L.F. Qualidade de frutos de genótipos de umbu-cajazeira (Spondias sp.). Rev. Bras. Eng. Agric. Ambient. 2013, 17, 1217–1221. [CrossRef]
- Silva, L.R.; Alves, R.S.; Aragão, F.A.S.; Silva, S.M.; Maia, L.K.; Nogueira, D.H. Qualidade de frutos de genótipos de umbu-cajazeiras (Spondias sp.) oriundos da microrregião de Iguatu, CE. Sci. Plena 2011, 7, 1–7. [Google Scholar]
- Vieira, A.R. Propagação assexuada e qualidade de frutos de genótipos de umbu-cajazeira da mesorregião centro-sul do Ceará. Ph.D. Thesis, Universidade Federal Rural do Semiárido, Mossoró, Brazil, 2013. [Google Scholar]
- Lira-Júnior, J.S.; Musser, R.S.; Lêderman, I.E.; Martins, L.S.S. Variabilidade entre genótipos de um banco de germoplasma de cajá-umbuzeiro (Spondias spp.) na Zona da Mata de Pernambuco. Rev. Bras. Ciênc. Agrár. 2008, 3, 116–120. [Google Scholar] [CrossRef]
- Lima, E.D.P.A.; Lima, C.A.A.; Aldrigue, M.L.; Gondim, P.J.S. Caracterização física e química dos frutos da umbu-cajazeira (Spondias spp) em cinco estádios de maturação, da polpa congelada e néctar. Rev. Bras. Frutic. 2002, 24, 338–343. [Google Scholar] [CrossRef]
- Soares-Filho, W.S.; Ritzinger, R. Pré-melhoramento genético de fruteiras nativas: Caso da umbu-cajazeira na Embrapa Mandioca e Fruticultura Tropical. In Curso Internacional de Pré-Melhoramento de Plantas; Lopes, M.A., Fávero, A.P., Ferreira, M.A.J., Faleiro, F.G., Eds.; Embrapa Recursos Genéticos e Biotecnologia: Brasília, Brazil, 2006. [Google Scholar]
- Fiaes, G.; Cardoso, R.L.; Viana, E.S. Aceitação sensorial de compota de umbu-cajá. In Proceedings of the Jornada Científica Embrapa Mandioca e Fruticultura Tropical, 3, Cruz das Almas, BA, Brazil, 2009; Embrapa Mandioca e Fruticultura Tropical: Cruz das Almas, Brazil, 2009.
- Bastos, L.P. Caracterização de frutos e propagação vegetativa de Spondias. M.Sc. Thesis, Universidade Federal do Recôncavo da Bahia, Cruz das Almas, Brazil, 2010. [Google Scholar]
- Soares-Filho, W.S. Prospecção e seleção de genótipos de umbu-cajazeira no estado da Bahia. In Botânica e Desenvolvimento Sustentável; Lucena, E.M.P., Amorim, A.V., Eds.; EdUECE: Fortaleza, Brazil, 2011; p. 371. [Google Scholar]
- Silva, G.A.; Brito, N.J.N.; Santos, E.C.G.; López, J.A.; Almeida, M.G. Gênero Spondias - aspectos botânicos, composição química e potencial farmacológico. Rev. Biol. Farm. 2014, 10, 8–22. [Google Scholar]
- Souza, F.X.; Souza, F.H.L.; Freitas, J.B.S. Caracterização morfológica de endocarpos de umbu-cajá. In Proceedings of the 48° Congresso Nacional de Botânica, Crato, CE, Brazil, 1997; Sociedade Botânica do Brasil: Crato, Brazil, 1997; p. 121. [Google Scholar]
- Behling, H.; Arz, H.W.; Pätzold, J.; Wefer, G. Late Quaternary vegetational and climate dynamics in northeastern Brazil, inferences from marine core GeoB 3104-1. Quat. Sci. Rev. 2000, 19, 981–994. [Google Scholar] [CrossRef]
- Pennington, R.T.; Prado, D.E.; Pendry, C.A. Neotropical seasonally dry forests and Quaternary vegetation changes. J. Biogeogr. 2000, 27, 261–273. [Google Scholar] [CrossRef]
- Auler, A.S.; Wang, X.; Edwards, R.L.; Cheng, H.; Cristalli, P.S.; Smart, P.L.; Richards, D.A. Palaeoenvironments in semi-arid northeastern Brazil inferred from high precision mass spectrometric speleothem and travertine ages and the dynamics of South American rainforests. Speleogenesis Evol. Karst Aquifers 2004, 2, 1–4. [Google Scholar]
- Auler, A.S.; Wang, X.; Edwards, R.L.; Cheng, H.; Cristalli, P.S.; Smart, P.L.; Richards, D.A. Quaternary ecological and geomorphic changes associated with rainfall events in presently semi-arid northeastern Brazil. J. Quat. Sci. 2004, 19, 693–701. [Google Scholar] [CrossRef]
- Shock, M.P. Holocene hunter-gatherer plant use and foraging choice - a test from Minas Gerais, Brazil. Ph.D. Dissertation, University of California, Santa Barbara, CA, USA, 2010. [Google Scholar]
- Shock, M.P.; Kipnis, R. Archaeobotanical insights into diet on the Brazilian savanna, 500-2000 BP. 2008. Available online: http://www.shockfamily.info/Myrtle/archaeobotanical.html (accessed on 19 April 2024).
- Shock, M.P.; Kipnis, R.; Bueno, L.; Silva, F.M. A chronology of the introduction of domesticated plants in central Brazil. Tipití 2013, 11, 52–59. [Google Scholar] [CrossRef]
- Lima, J.M.D. Arqueologia de Furna do Estrago, Brejo da Madre de Deus - PE. Clio (Série Arqueológica) 1985, 2, 97–111. [Google Scholar]
- Lima, J.M.D. Dois períodos de subsistência no Agreste pernambucano - 9000 e 2000 A.P. Clio (Série Arqueológica) 1991, 4, 57–61. [Google Scholar]
- Lima, M.G.S.M. Estudo arqueobotânico dos restos alimentares silvestres do sítio arqueológico Alcobaça, Buíque - PE. M.Sc. Thesis, Universidade Federal de Pernambuco, Recife, Brazil, 2010. [Google Scholar]
- Lima, J.M.D.; Schmitz, P.I.; Souza, S.M.F.M.; Beber, M.V. A Furna do Estrago no Brejo da Madre de Deus, PE. Pesquisas (Antropologia) 2012, 69, 5–159. [Google Scholar]
- Souza, G.S. Tratado Descriptivo do Brasil em 1587; Typographia de João Ignacio da Silva: Rio de Janeiro, Brazil, 1879; p. 882. [Google Scholar]
- Hughes, C.; Govindarajulu, R.; Robertson, A.; Filers, D.L.; Harris, S.A.; Bailey, C.D. Serendipitous backyard hybridization and the origin of crops. Proc. Natl. Acad. Sci. USA 2007, 104, 14389–14394. [Google Scholar] [CrossRef]
- Abbott, R.J. Plant invasions, interspecific hybridization and the evolution of new plant taxa. Trends Ecol. Evol. 1992, 7, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Ellstrand, N.C.; Prentice, H.C.; Hancock, J.F. Gene flow and introgression from domesticated plants into their wild relatives. Annu. Rev. Ecol. Syst. 1999, 30, 539–563. [Google Scholar] [CrossRef]
- Ellstrand, N.C.; Schierenbeck, K.A. Hybridization as a stimulus for the evolution of invasiveness in plants? Proc. Natl. Acad. Sci. USA 2000, 97, 7043–7050. [Google Scholar] [CrossRef]
- Arnold, M.L. Natural hybridization and the evolution of domesticated, pest and disease organisms. Mol. Ecol. 2004, 13, 997–1007. [Google Scholar] [CrossRef]
- Neiva, A.; Penna, B. Viagem científica pelo Norte da Bahia, sudoeste de Pernambuco, sul do Piauhí e de norte a sul de Goiaz. Mem. Inst. Oswaldo Cruz 1916, 8, 74–224. [Google Scholar] [CrossRef]
- Bondar, G. Moscas-das-frutas na Bahia. Bahia Rural 1949, 17, 26–27. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Barrera-Rojas, C.H.; van den Berg, C. Sand-mediated DNA extraction from orchids for genomic applications. BioRxiv 2025, 2025.10.08.681153. [Google Scholar] [CrossRef]
- Oxelman, B.; Lidén, M.; Berglund, D. Chloroplast rps16 intron phylogeny of the tribe Sileneae (Caryophyllaceae). Plant Syst. Evol. 1997, 206, 393–410. [Google Scholar] [CrossRef]
- Pell, S.K. Molecular systematics of the cashew family (Anacardiaceae). Ph.D. Dissertation, Louisiana State University, Baton Rouge, LA, USA, 2004. [Google Scholar]
- Baldwin, B.G.; Markos, S. Phylogenetic utility of the external transcribed spacer (ETS) of 18S-26S rDNA: Congruence of ETS and ITS trees of Calycadenia (Compositae). Mol. Phylogenet. Evol. 1998, 10, 449–463. [Google Scholar] [CrossRef] [PubMed]
- Samarakoon, T.; Wang, S.Y.; Alford, M.H. Enhancing PCR amplification of DNA from recalcitrant plant specimens using a trehalose-based additive. Appl. Plant Sci. 2013, 1, 1200236. [Google Scholar] [CrossRef]
- Paithankar, K.R.; Prasad, K.S. Precipitation of DNA by polyethylene glycol and ethanol. Nucleic Acids Res. 1991, 19, 1346. [Google Scholar] [CrossRef]
- Staden, R.; Beal, K.F.; Bonfield, J.K. The Staden Package, 1998. In Bioinformatics Methods and Protocols; Misener, S., Krawetz, S.A., Eds.; Humana Press: Totowa, NJ, USA, 2000; pp. 115–130. [Google Scholar]
- Galtier, N.; Gouy, M.; Gautier, C. SEAVIEW and PHYLO_WIN: Two graphic tools for sequence alignment and molecular phylogeny. Bioinformatics 1996, 12, 543–548. [Google Scholar] [CrossRef]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Doyle, J.J. Gene trees and species trees: Molecular systematics as one-character taxonomy. Syst. Bot. 1992, 17, 144–163. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP: Phylogenetic Analysis Using Parsimony (and Other Methods), version 4; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Ihaka, R.; Gentleman, R. R: A language for data analysis and graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar] [CrossRef]
- R Core Team 2014R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef]
- Inkscape 0.48.4. 2013. Available online: http://www.inkscape.org.
- Network 4.6.1.1. 2013. Available online: http://www.fluxus-engineering.com/ (accessed on 5 July 2013).
- Bandelt, H.-J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Mitchell, J.D.; Daly, D.C. A revision of Spondias L. (Anacardiaceae) in the Neotropics. Phytokeys 2015, 55, 1–92. [Google Scholar] [CrossRef]
- Kostermans, A.J.G.H. Kedondong, Ambarella, Amra. The Spondiadeae (Anacardiaceae) in Asia and the Pacific Area; Published by the Author: Bogor, Indonesia, 1991; p. 100. [Google Scholar]
- Campbell, C.W.; Sauls, J.W. Spondias in Florida. Fact Sheet HS-63; University of Florida: Gainesville, FL, USA, 1994. [Google Scholar]
- Duvall, C.S. On the origin of the tree Spondias mombin in Africa. J. Hist. Geogr. 2006, 32, 249–266. [Google Scholar] [CrossRef]
- Mitchell, J.D.; Daly, D.C. The “tortoise’s caja” - a new species of Spondias (Anacardiaceae) from southwestern Amazonia. Brittonia 1998, 50, 447–451. [Google Scholar] [CrossRef]
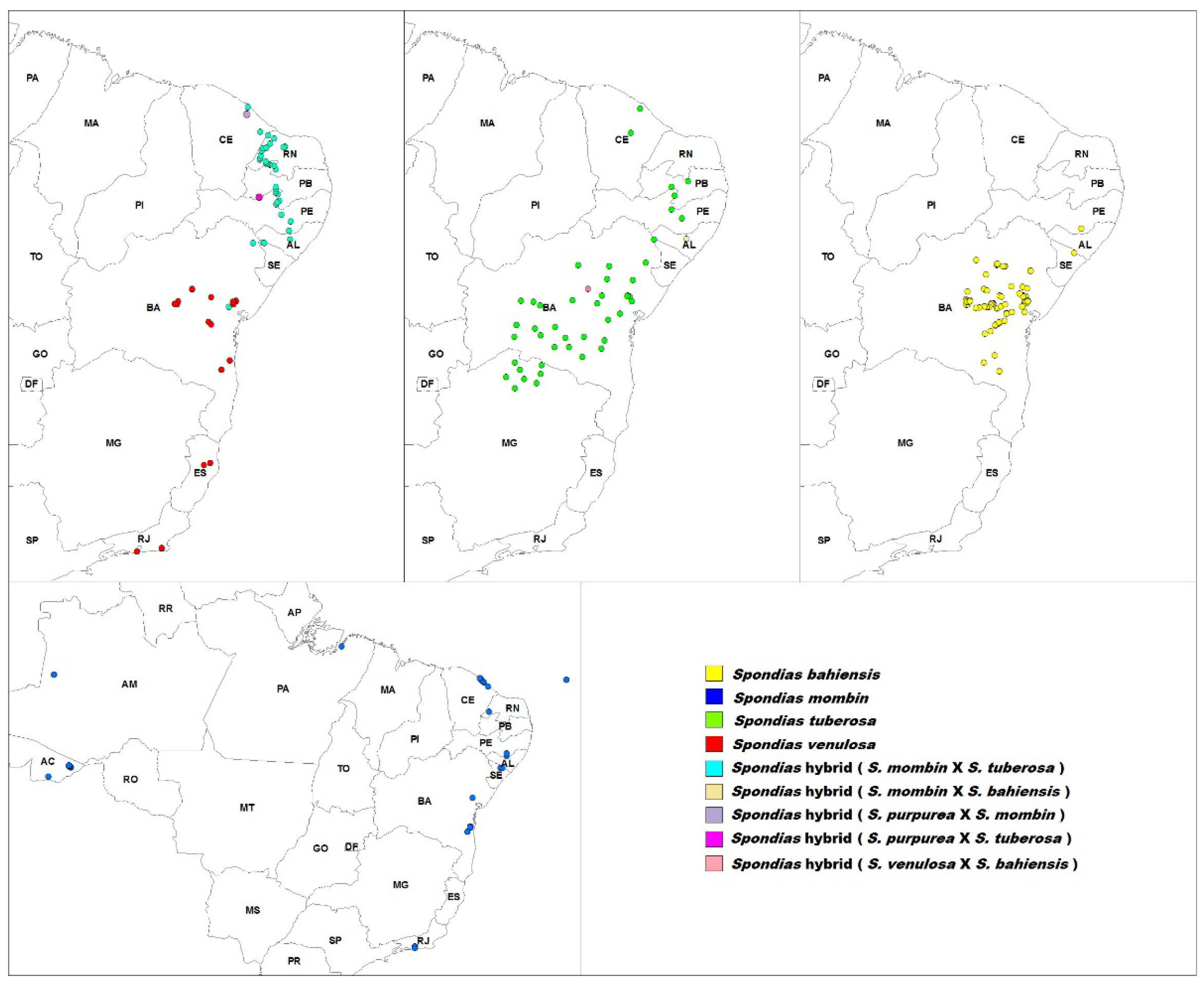
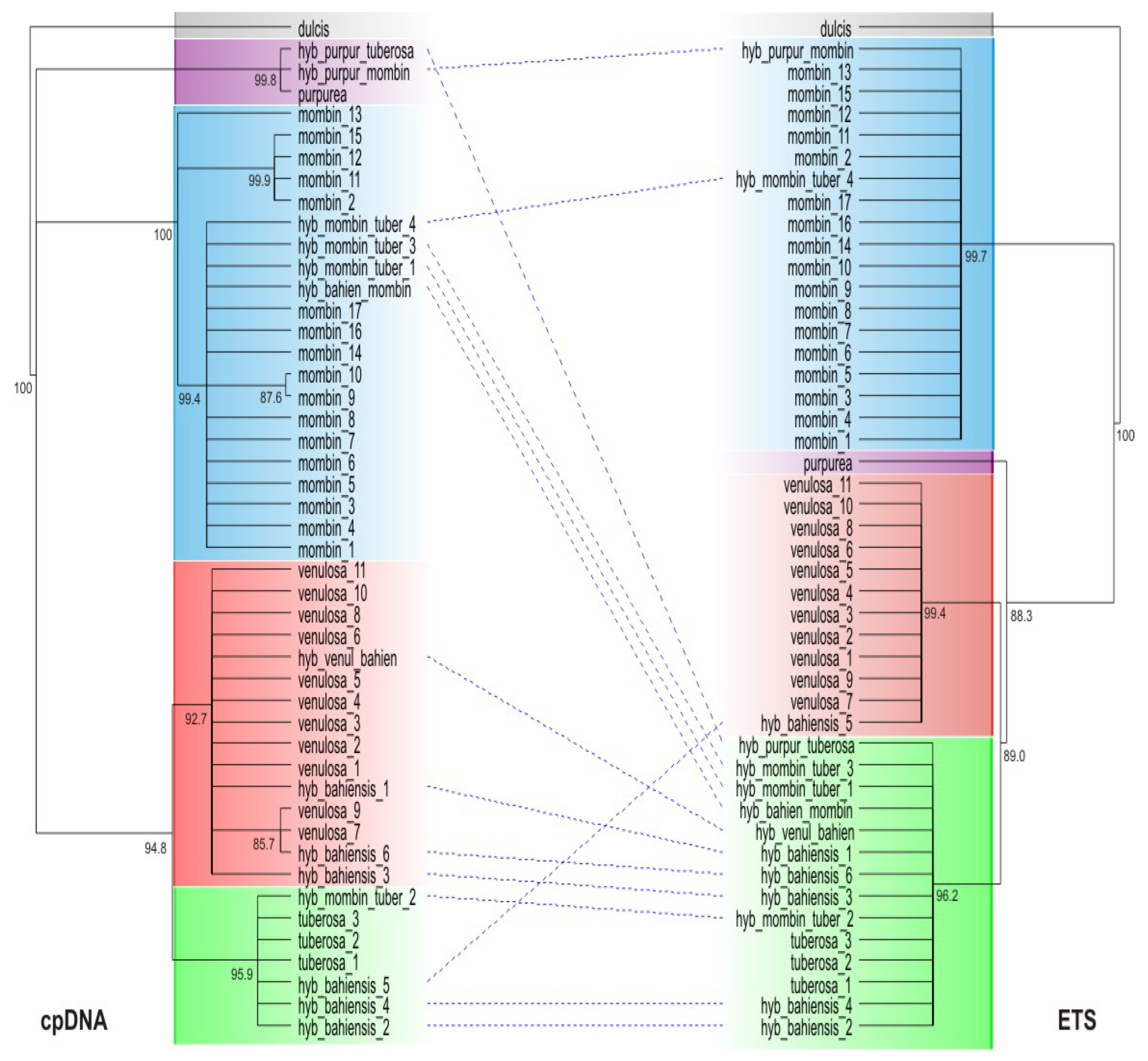
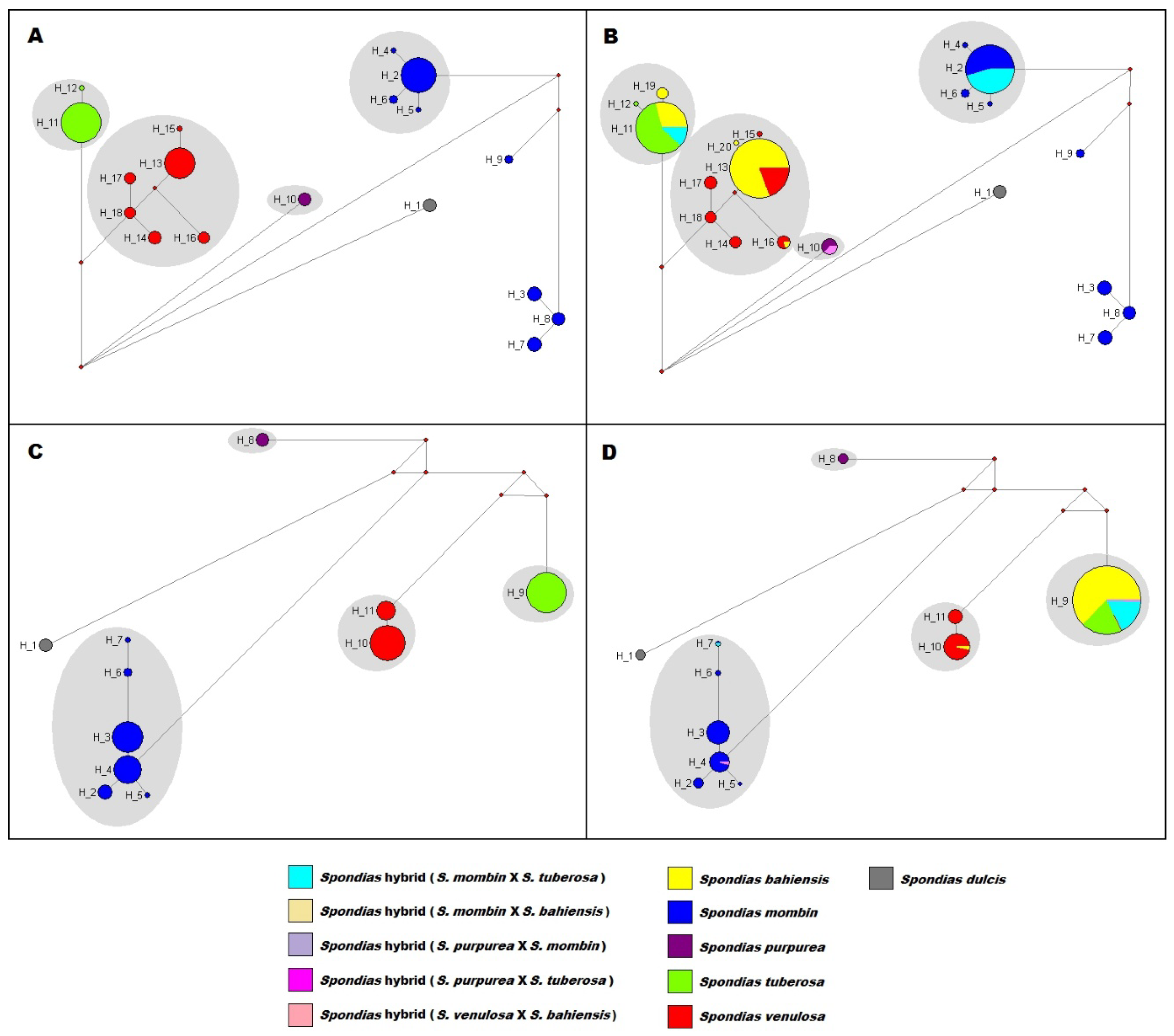
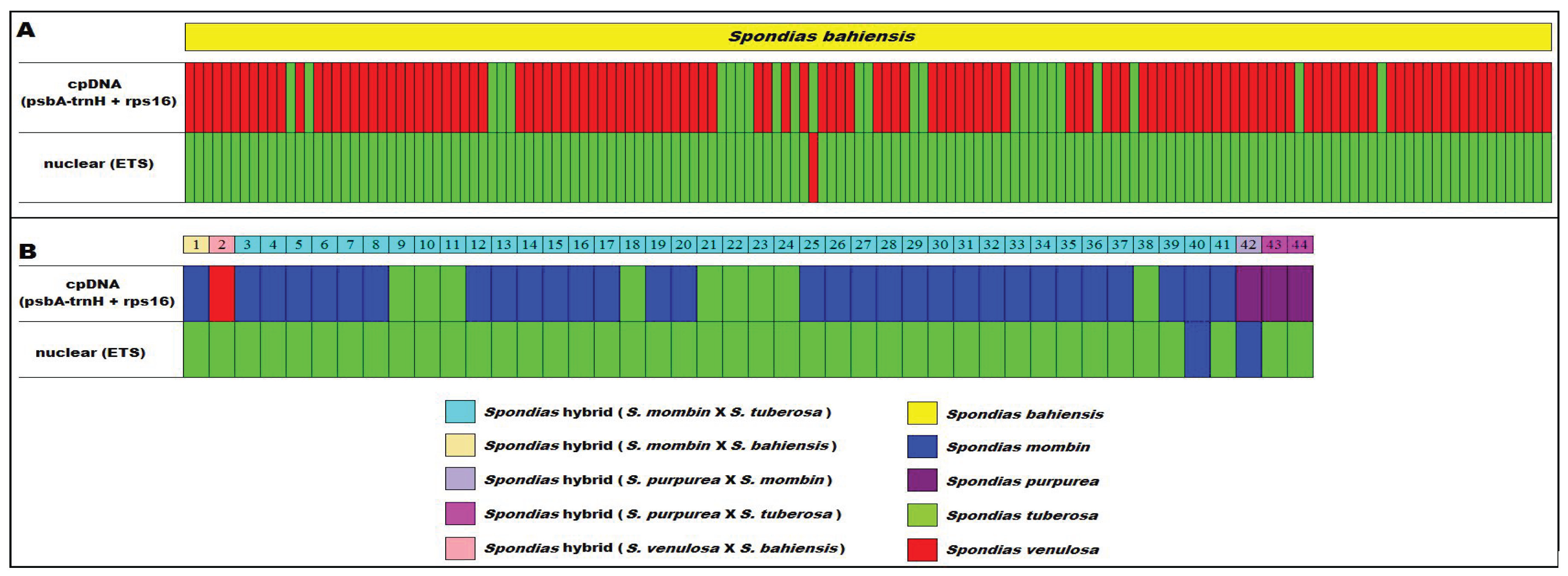

| mark | Species | Distribution |
|---|---|---|
| * | Spondias admirabilis J.D. Mitch. & Daly | Atlantic Forest of Rio de Janeiro, Brazil (Mitchell & Daly 2015) [149] |
| + | Spondias dulcis Parkinson | Originally from Society Islands (Kostermans 1991 [150]), Polynesia (Campbell & Sauls 1994 [151]), widely cultivated pantropically. |
| * | Spondias expeditionaria J.D. Mitch. & Daly | Atlantic Forest of Espírito Santo and Minas Gerais, Brazil (Mitchell & Daly 2015) [149] |
| Spondias globosa J.D.Mitch. & Daly | W Amazonia; outlier in Zulia, Venezuela (Mitchell & Daly 2015) [149] | |
| + | Spondias pinnata (L.f.) Kurtz | Originally from India to Myamar and Thailand (Kostermans 1991) [150], sporadically cultivated elsewhere, in Brazil found in a few collections. |
| + | Spondias purpurea L. | Originally from Mexico and Central America (Miller & Schaal 2005) [25], widely cultivated pantropically. |
| * | Spondias macrocarpa Engl. | Brazil, found in humid forests in southern Bahia, Espírito Santo, southeastern Minas Gerais and Rio de Janeiro states. |
| Spondias mombin L. | Widely distributed in the neotropics, naturalized in parts of Africa (Duvall 2006) [152], widely cultivated pantropically. | |
| Spondias testudinis J.D.Mitch. & Daly | Restricted to a small area in Bolivia (Pando), Brazil (Acre), and Peru (Huanuco, Ucayali) (Mitchell & Daly 1998) [153]. | |
| * | Spondias tuberosa Arruda | Brazil, found in most states of Northeastern Brazil and in the northern region of Minas Gerais state, growing in the Caatinga seasonally dry woodlands; sporadically cultivated elsewhere. |
| * | Spondias venulosa (Engl.) Engl. | Brazil, occurring in semi-deciduous forests in Bahia, Espírito Santo, Minas Gerais, and Rio de Janeiro states. |
| cpDNA dataset | ETS | |
|---|---|---|
| Total number of characters | 1590 | 371 |
| Number of constant characters | 1509 | 331 |
| Number of substitutions | 81 | 40 |
| Number of parsimony-uninformative characters | 22 | 13 |
| Number of parsimony-informative characters | 59 | 27 |
| Number of indels | 32 | 4 |
| Size range of indels | 1–96 | 2–3 |
| Number of trees retained in heuristic search | 11 | 2 |
| Tree length | 89 | 49 |
| Consistency index (CI) | 0.9438 | 0.8776 |
| CI excluding uninformative characters | 0.9254 | 0.8286 |
| Retention index (RI) | 0.9926 | 0.9855 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



