Submitted:
13 December 2025
Posted:
16 December 2025
You are already at the latest version
Abstract
To optimize juvenile barramundi (Lates calcarifer) feeding strategies, this study compared cannibalism (CB), formulated feed (FF), and mixed feed (MIX: formulated + biological feed) on growth, physiology, and immune-related gene expression. 36-day-old juveniles (initial body weight) were randomized into 3 groups (1 Lates calcarifer was placed in each tank, with 15 replicates in each group.) for a 20-day trial. Growth performance: MIX group showed significantly higher weight gain rate (862.31 ± 346.66) and specific growth rate (93.2 ± 42.48) than FF and CB groups (P < 0.05); CB group outperformed FF (P< 0.05). Physiology: MIX had the highest alkaline phosphatase (AKP) and pyruvate kinase (PK) activities (P < 0.05), and significantly elevated alanine aminotransferase (ALT) activity, lipid peroxide (LPO) content, but the lowest catalase (CAT) activity (P < 0.05). Gene expression: CB group had the highest lysosomal protease (cts1a) and glycolytic gene (eno3) levels (P < 0.05); FF group showed higher heat-shock protein 90(hsp90) and pro-inflammatory cytokine (IL1β) expression (P < 0.05). FF exhibited the highest SOD activity and IL1β levels (P < 0.05), indicating strong antioxidant. Conclusion: MIX promotes growth but risks liver damage/oxidative stress; CB serves as emergency nutrition but requires management to avoid; FF exhibits significant antioxidant advantages despite poor growth performance. "Mixed feeding + immune enhancers" is recommended for industrial seedling production to balance growth and health.
Keywords:
barramundi
; feeding strategy
; cannibalism
; growth performance
; physiological and biochemical
; immune genes
; industrial seedling production
1. Introduction
Cannibalism is a strategy for fish to cope with harsh environments or promote the rapid growth of small individuals [1]. In nature, this strategy can effectively preserve the survival of the population under conditions of hunger and lack of food, and it is easier to prey on conspecifics within the feeding group under high-competition conditions in the wild [2]. At the same time, the occurrence of cannibalism also indicates that the digestive tract of the cannibal has fully developed, enabling it to withstand the struggle of the prey before death and to digest and absorb the complex nutritional components of the whole small fish [3]. However, under artificial breeding conditions, where sufficient food can be provided and food and nutrition are no longer limiting factors for growth, cannibalism still occurs, especially after obvious size differences appear [4]. Why does cannibalism still occur when there is an abundant supply of food? Under the conditions of providing sufficient artificial food and an abundant supply of prey, what differences will occur in the growth rate, digestive enzyme activity, immune enzyme activity, and the expression levels of genes related to digestive enzymes and immune enzymes of the predators?
The Asian seabass (Lates calcarifer) is an important economic fish with advantages such as fast growth and strong adaptability, and it is widely farmed in regions such as Asia and Australia [5,6]. However, during the fry rearing stage, there is a widespread and serious phenomenon of cannibalism among Asian seabass larvae and juveniles, that is, larger individuals prey on smaller conspecifics, resulting in a significant decrease in survival rate and seriously affecting the farming efficiency [7]. Cannibalism is one of the key bottlenecks in the fry rearing of carnivorous fish, and its occurrence is affected by multiple factors, including food type, size difference, hunger stress, and stocking density [8,9]. Currently, research on the cannibalistic behavior of Asian seabass mainly focuses on farming management measures (such as graded rearing) and feeding strategies, while it is still unclear how cannibalism affects the growth, digestive physiology, and immune status of Asian seabass.
Studies have shown that there are significant differences in nutritional components, palatability, and digestive utilization between artificial food and live food (such as small fry) [10,11]. For example, when juvenile Giant Salamander (Andrias davidianus) are fed artificial food, the activities of intestinal digestive enzymes (such as trypsin) and the expression levels of genes related to lipid metabolism (such as Fatty Acid Synthase, Carnitine Palmitoyltransferase I) are significantly lower than those in the live - food group, which may lead to insufficient nutrient intake, exacerbate growth dispersion, and further induce cannibalism [10]. In addition, changes in the immune status (such as a decrease in lysozyme activity or an up-regulation of inflammatory factor expression) may further exacerbate cannibalistic behavior [12], but relevant research is still limited.
This study focuses on the growth performance of juvenile barramundi (Lates calcarifer) under different feeding strategies, the changes in the activities of digestion - and immunity - related enzymes, and the regulatory differences in the expression of key genes. The aim is to reveal the mechanism by which cannibalistic behavior affects the physiological state and metabolic function of predators. Despite the sufficient supply of feed under artificial breeding conditions, cannibalism among conspecifics still occurs frequently, suggesting that this behavior may not only be driven by hunger but is also closely related to nutrient acquisition efficiency, energy metabolism optimization, and physiological state regulation. Therefore, this study clearly proposes three specific objectives:
(1) Compare the growth rates and weight changes of barramundi in the FF group, CB group, and MIX group to evaluate whether cannibalism brings growth advantages.
(2) Measure the differences in the activities of digestive enzymes (such as proteases and lipases), metabolic enzymes (such as ALT, LDH, AKP, etc.), and antioxidant enzymes (such as SOD, POD, MDA, etc.) in the liver and whole - body tissues to analyze the effects of different nutrient sources on the body’s metabolic load and oxidative stress level.
(3) Analyze the expression levels of immunity - related genes (such as myD88, IFNγ1, IL1β) and stress - response genes (hsp90) through qRT - PCR to explore whether long - term consumption of live prey or mixed feed affects the individual’s immune status and disease - resistance potential. The significance of this study lies in the fact that it is the first to systematically evaluate the comprehensive impact of cannibalistic behavior on the physiological health of barramundi under non - hunger stress conditions. It breaks through the traditional cognitive limitation of simply regarding cannibalism as a “negative loss” and helps to re - understand the ecological and evolutionary significance of this behavior from the perspective of “predator benefits”. The research results can provide a theoretical basis for optimizing the fry - rearing strategies of barramundi. For example, by adjusting the feed structure and intervening in early feeding behavior, the growth dispersion and cannibalism rate within the population can be reduced, thereby improving the breeding survival rate and production efficiency. At the same time, this study also provides a technical path and theoretical reference for the research on the cannibalism regulation mechanism of other carnivorous fish.
2. Materials and Methods
2.1. Experimental Fish and Design
The juvenile barramundi (Lates calcarifer) used in this experiment were obtained from the Hainan Base of the South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences. The fertilized eggs were self-bred, and the juvenile fish were obtained after hatching and nursing. At the beginning of the experiment, the fish were 36 days old and had been successfully domesticated to eat artificial feed. The juvenile fish were randomly divided into three groups: the CB group is cannibalism group, only feed by smaller juvenile barramundi; the FF group with only formulated feed, and the MIX group with a mixture of smaller juvenile barramundi and formulated feed. All the formulated feeds were commercial compound feeds (Haitong compound feed for juvenile fish, Santong Bio-engineering (Weifang) Co., Ltd.), and the feed particle size was selected according to the mouth diameter of the barramundi. The experiment was carried out in 36L tanks. The water quality conditions of each treatment were consistent with those in the domestication stage. 1 experimental fish was placed in each tank, 15 tanks for each treatment. The experiment is divided into 3 treatments in total. The entire experimental period was 20 days. All the water tanks are connected to the same recirculating aquaculture system.
In the CB and MIX treatments, 15 live preys were fed to each tank every 24 hours. The preys were smaller barramundi, and the body length of the preys was 40%-50% of that of the predators. In addition, 10,000 juvenile fish of similar size were selected in advance as potential preys and temporarily kept in a floating bucket. After measuring the wet body weight to estimate the biomass of the prey (OPB, g), the preys were released into the tank respectively for the predators. During the experiment, the number of preys released in each experimental bucket (OPN) was recorded daily, and the body weight of the predators was measured. The FF and MIX treatments were fed with enough feed three times a day (6:00, 14:00, 22:00). The aquaculture water was maintained under the following conditions: salinity 32‰, dissolved oxygen > 7.50 mg/L, pH 7.93 ± 0.12, ammonia nitrogen < 0.10 mg/L, and nitrite nitrogen < 0.05 mg/L. After 20 days of the experiment, 5 fish were randomly selected, over - anesthetized with MS - 222, and the whole - fish tissues were quickly collected for enzyme activity determination and RNA extraction. All samples were quickly frozen in liquid nitrogen and transferred to - 80 °C for subsequent analysis.
2.2. Research and Analysis of Oxidation Capacity and Metabolic Enzyme Activity
Approximately 0.1 g of tissue was collected from each juvenile barramundi and placed in a 2-mL centrifuge tube. Then, 9 times the volume of 0.86% normal saline solution was added. The tissues were homogenized on ice using a Prima PB100 handheld homogenizer (Gloucester, England). The homogenate was centrifuged at 3500 ×g for 10 min, and the supernatant was collected and stored at - 80 °C for analysis of antioxidant capacity and metabolic enzyme activity.
The antioxidant indicators in the tissues mainly included superoxide dismutase (SOD), lipid peroxide (LPO), peroxidase (POD), malondialdehyde (MDA), glutathione reductase (GR), and peroxidase (POD). These antioxidant indicators were measured using commercial detection kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) according to the kit instructions. The protein content in the homogenate was measured using a BCA kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
The metabolic enzyme activity indicators mainly included alanine aminotransferase (ALT), lactate dehydrogenase (LDH), alkaline phosphatase (AKP), and acid phosphatase (ACP). These metabolic enzyme activity indicators were measured using commercial detection kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) according to the kit instructions. The protein content in the homogenate was measured by the BCA method (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
2.3. Sampling and Determination
The experimental period was 20 days. After the experiment, the predators were anesthetized and weighed. After weighing, the whole fish was ground to measure enzyme activity and genes.
The whole-fish homogenate of barramundi under different feeding strategies was used for RNA extraction. Total RNA was extracted using the Thermo Scientific GeneJET RNA Purification Kit (Waltham, MA, USA). According to the manufacturer’s instructions, the integrity, concentration, and purity of RNA were detected using agarose gel electrophoresis, Nanodrop2000 (Thermo Fish Scientific Waltham, MA, USA), and 2100 Bioanalyser (Agilent Technologies, Waldbronn, Germany). Only high - quality RNA (OD260/280 = 1.8 - 2.2, OD260/230 ≥ 2.0, 28S:18S ≥ 1.0, Total RNA > 10 μg) could be used for subsequent experiments. 2 μl of total RNA was pipetted, and the first - strand cDNA was synthesized by reverse transcription according to the instructions of the EasyScript All - in - One First - Strand cDNA Synthesis SuperMix for qPCR (One - Step gDNA Removal) kit, which was used as a template for subsequent gene expression determination [13].
In the quantification of immune gene expression at each time point, the gene expression level of the CB group was used as a normalization index for gene relative quantification. Five immune and metabolic single genes were selected for verification by quantitative real-time PCR (qRT-PCR) using a real-time qPCR analysis system (Analytik Jena GmbH, Germany) and SYBR Green (Tiangen Biotech Co., Ltd., China). Specific primers were designed using Primer Premier 5 software (Table 1). The total reaction mixture was 20 μl, including 10 μl of 2×RealUniversal PreMix, 0.6 μl of each primer (10 μM), and 2 μl of diluted cDNA. First, it was denatured at 95 °C for 15 minutes, followed by 40 cycles of amplification (95 °C, 10 s; 58 °C, 20 s; 72 °C, 30 s). At the end of each qRT-PCR cycle, a melting curve analysis of the primers was performed to ensure that only specific products were obtained and no primer dimers were formed. Each experiment included a no-template control to verify that the PCR reaction mixture was free of contamination. The relative mRNA expression level of the target gene was determined by the 2 - ΔΔCt method, with the β-actin gene as the internal reference gene and the CG group at 0 h for normalization. The reaction efficiency was 90 - 110%, and the Pearson correlation coefficient (R2) > 0.97.
2.4. Statistical Analysis
The data were expressed as mean (X) ± standard deviation (SD). One-way ANOVA was used to analyze the data using SPSS 19.0 software, and Duncan’s multiple comparison method was used to analyze the significant differences between groups, with a significance level of P < 0.05.
3. Results
3.1. Growth Performance
As can be seen from Table 2, both the average weight gain rate and specific growth rate showed the order of MIX > CB > FF. That is, the MIX group had the highest growth and the fastest growth rate. Followed by the CB group, the FF group had the least weight gain and the slowest growth rate. However, there was no significant difference in the weight gain rate and specific growth rate between the MIX group, the FF group and the CB group (P > 0.05). Feeding solely on artificial feed (FF) resulted in the poorest growth performance. Although cannibalism among conspecifics (CB) was not significantly better than the FF group, it showed higher growth potential.
3.2. Effects of Different Baits on the Physiological and Biochemical Indicators of Lates calcarifer
As can be seen from Figure 1a, the ALT levels in the FF group and the CB group were significantly lower (P < 0.05) than those in the MIX group. The LDH activity was stable in the three groups of experiments (Figure 1b), with no significant differences between groups and small differences within groups, suggesting that none of the three feeding methods would cause significant differences in general cell damage to Lates calcarifer. The ACP activity in the CB group was significantly lower than that in the other two groups (Figure 1c).
As shown in Figure 1d, the SOD activity in the CB group was
the lowest, significantly lower (P < 0.05) than that in the FF group and the MIX group, and it was stable within the group. The SOD activities in the FF group and the MIX group were relatively high, with no significant difference between the two groups, and at the same time, there were relatively large differences within the groups. The CAT activity in the MIX group was much lower than that in the FF and CB groups (Figure 1e), and the difference within the MIX group was small. The CAT activity values in the FF group and the CB group were relatively high, with no significant difference between the groups, and the CAT activity values within the groups were unstable. It is worth noting that although the MIX group showed the optimal growth performance, its SOD and CAT activities were relatively low, suggesting the possible existence of antioxidant stress; this was confirmed in its subsequent immune gene regulation - the expression of the pro - inflammatory factor IFNγ1 in the MIX group increased, which might be a compensatory response of the body to oxidative stress.
The POD activity values showed the order of MIX group > FF group > CB group (Figure 1f), and the differences between the groups were significant (P < 0.05). The small error values indicated that the activity values within the groups were stable. The GR activity values showed the order of FF group > MIX group > CB group (Figure 1g), and the differences between MIX group and FF group or between MIX group and CB group were not significant (P > 0.05). The small error values indicated that the activity values within the groups were stable, but the GR activity value in the FF group was significantly higher (P < 0.05) than that in the CB group.
The MDA content was in the order of FF group > MIX group > CB group (Figure 1h). There was no significant difference between adjacent groups (P > 0.05). The small error values in the MIX group and CB group indicated stable content within the groups. However, the MDA content in the FF group was significantly higher (P < 0.05) than that in the CB group. However, the difference between groups was not significant, suggesting that the overall effect was small. As shown in the Figure 1i, the LPO content was in the order of MIX group > FF group > CB group. There was significant difference between adjacent groups (P < 0.05). The small error values in these grous indicated stable content within the groups.
As shown in Figure 1j, the AKP activity value was in the order of MIX group > FF group > CB group, and there were significant differences between groups (P < 0.05), with small fluctuations within each group. The PK activity value was in the order of MIX group > CB group > FF group (Figure 1k), and there were significant differences between groups (P < 0.05), with small fluctuations within each group.
3.3. Regulation of Gene Expression: Differences in Immunity, Metabolism, and Stress Response
To evaluate the effects of diet types on the innate immune capacity of juvenile barramundi, this study detected the gene expression levels of myD88, a key adapter molecule in the Toll-like receptor signaling pathway, and C3, a core component of the complement system. The results showed that diet types had significant effects on the expression of both immune genes (p < 0.05). As shown in Figure 2a and Figure 2b, the expression patterns of myD88 and C3 were the highest in the mixed-feeding group (MIX), and their expression levels were significantly higher than those in the formulated feed group (FF) and the cannibalism group (CB). Moreover, the expression levels in the FF group were significantly higher than those in the CB group (P < 0.05).
Under different diet feeding conditions, there were significant differences in the expression of immune-related genes in juvenile barramundi (Figure 2c, Figure 2d and Figure 2e). The expression level of the pro-inflammatory factor IFNγ1 was relatively high in the FF and MIX groups (Figure 2c), maintaining above 0.6 U/gprot, while it decreased significantly in the CB group, only about 0.3 U/gprot. The expression of pfnd6 (Figure 2d), which is related to the cytoskeleton and immune cell migration, was the highest in the FF group (about 1.0 U/gprot), decreased significantly in the MIX group, and was the lowest in the CB group (less than 0.2 U/gprot). This result suggests that formulated feed may have a stronger promoting effect on the motility of immune cells in juvenile fish, while feeding solely on small fish carcasses is not conducive to the expression of this gene. In terms of the expression of the lysosomal protease-related gene cts1a (Figure 2e), the CB group showed the highest level (about 1.2 U/gprot), significantly higher than the FF and MIX groups.
Under different diet feeding conditions, there were significant differences in the expression levels of metabolism - and stress-related genes in juvenile barramundi (Figure 2f, 2g, and 2h). The expression of hsp90 (Figure 2f), which is related to the molecular chaperone function, was maintained at relatively high levels in the FF and MIX groups (about 0.95 U/gprot and 0.8 U/gprot, respectively), while it decreased significantly in the CB group, only about 0.4 U/gprot. This suggests that formulated feed and mixed feed contribute to enhancing the molecular chaperone protection of juvenile fish under stress. The expression trend of the glycolysis-related gene eno3 (Figure 2g) was the opposite: it was the highest in the CB group, reaching about 1.7 U/gprot, significantly higher than the other two groups(p<0.05), followed by the MIX group (about 1.1 U/gprot), and the lowest in the FF group, only about 0.8 U/gprot. There was no significant difference between the MIX group and the FF group(p>0.05). The expression of ppl (Figure 2h) was the highest in the FF group, reaching about 1.05 U/gprot, decreased significantly to about 0.5 U/gprot in the MIX group, and was the lowest in the CB group, only about 0.2 U/gprot.
Under different diet feeding conditions, there were significant differences in the expression levels of the immune-related genes tegt and IL1β in juvenile barramundi (Figure 2i and 2j). Among them, the expression level of tegt (Figure 2i) was the highest in the FF group (about 0.85 U/gprot), significantly higher than that in the MIX group (about 0.2 U/gprot) and the CB group (about 0.15 U/gprot). In terms of the expression of the immune-regulatory gene IL1β(Figure 2j), the levels in the FF and CB groups were relatively high (about 1.2 and 1.0 U/gprot, respectively), and no significantly different between this two groups, while the MIX group had the lowest level, only about 0.6 U/gprot, MIX group significantly lower than the other two groups(p<0.05). This shows that feeding with formulated feed or small fish carcasses alone is conducive to the high expression of IL1β, while the expression decreased significantly under the mixed-feed condition.
4. Discussion
This study compared the effects of artificial compound feed (FF), mixed feed (MIX), and cannibalism of conspecifics (CB) on the growth performance, physiological and biochemical indicators, and gene expression of juvenile barramundi (Lates calcarifer). It revealed the “growth-health” trade-off relationship under different feeding strategies. The core findings and discussions are as follows:
4.1. Growth Advantages and Potential Risks of Mixed Feeding (MIX)
The weight gain rate (862.31 ± 346.66) and specific growth rate (93.2 ± 42.48) of the MIX group were significantly higher than those of the FF group and the CB group (P < 0.05), indicating that the mixed feeding of “artificial feed + biological feed” can promote the growth of juvenile barramundi. This is consistent with the general conclusion in the larval rearing of marine fish that mixed feeding is required during the transition from biological feed to artificial feed to improve feeding efficiency [14].
Potential mechanism: The LPO content was in the order of MIX group > FF group > CB group. There was significant difference between adjacent groups (P < 0.05), suggesting that the intensity of the lipid peroxidation reaction caused by the three types of feed was significantly different. LPO is an intermediate product in the process of lipid peroxidation, indicating that the lipid peroxidation reaction in the MIX group was actively occurring and was significantly higher than that in the FF group and CB group. It is shown that feeding artificial feed leads to an increase in fish body peroxides, and it is necessary to increase LOP to resist the negative effects [15]. The MIX group had the highest activities of AKP (substance metabolism) and PK (glycolysis) (P < 0.05), suggesting that the mixed feed may accelerate energy conversion and growth by optimizing nutritional balance (such as essential amino acids and unsaturated fatty acids) and digestion utilization rate. This is consistent with the view proposed by Safari, Omid, et al. (2014) that mixed feed can improve the metabolic enzyme activities of marine fish through nutritional complementarity [16].
Although the MIX group had the best growth performance, there were the following health risks:
Liver damage risk: The ALT activity increased significantly (P < 0.05), indicating that feeding on artificial compound feed alone or feeding on conspecifics would not cause damage to the liver of Lates calcarifer, while mixed feeding would cause liver damage [17], which may be related to the nutritional imbalance of biological feed (such as brine shrimp) or the accumulation of anti-nutritional factors in artificial feed. This is consistent with the result reported by Chatzifotis, Stavros et al.(2011). that long-term mixed feeding leads to an increased metabolic load on the liver of sea bass [18].
Weak antioxidant defense: The CAT activity was the lowest (P < 0.05), and the LPO content was significantly higher than that of the FF group (P < 0.05), indicating that it faced higher oxidative stress. This may be related to the mechanism proposed by Lushchak(2006) [19] Michael J Considine et al. (2021) that rapid growth leads to the accumulation of reactive oxygen species. When the growth rate exceeds a certain threshold, the antioxidant system of fish will be temporarily inhibited.
Imbalance of immune homeostasis: The pro-inflammatory genes such as myD88 and C3 were highly expressed, while the anti-inflammatory factor IL1β decreased significantly (P < 0.05), suggesting that “rapid growth may come at the cost of immune regulation”. This is consistent with the theory of growth-immunity resource allocation trade-off in teleost fish [20].
4.2. Adaptive Characteristics of Cannibalism of Conspecifics (CB)
Although the weight gain rate (542.16 ± 61.64) and specific growth rate (62.61 ± 38.27) of the CB group were lower than those of the MIX group, they were significantly higher than those of the FF group (P < 0.05), indicating that juvenile barramundi can obtain nutrients and maintain a moderate growth rate through cannibalism of conspecifics. This is consistent with the carnivorous habit of barramundi in the natural environment. However, its cannibalistic behavior may be affected by factors such as stocking density and individual size difference, and should be avoided through graded culture in actual production [21].
Physiological and genetic adaptations:The ACP activity in the CB group was significantly lower than that in the other two groups, indicating that in this experiment, feeding on artificial compound feed by Lates calcarifer accelerated the lysis or apoptosis rate of red blood cells [22]. The PK activity value was in the order of MIX group > CB group > FF group (Figure 1k), and there were significant differences between groups (P < 0.05), with small fluctuations within each group. The efficiency of the glycolytic pathway and energy production in the CB group was the lowest, which may be related to the composition of the fish body [23]. The expression level of the pro-inflammatory factor IFNγ1 was relatively high in the FF and MIX groups (Figure 2c), maintaining above 0.6 U/gprot, while it decreased significantly in the CB group, only about 0.3 U/gprot. This indicates that cannibalism can more effectively activate the inflammatory immune response of juvenile fish [12]. In terms of the expression of the lysosomal protease-related gene cts1a (Figure 2e), the CB group showed the highest level (about 1.2 U/gprot), significantly higher than the FF and MIX groups. This trend indicates that the diet of small fish carcasses may induce stronger lysosomal activity and antigen-processing ability, while the formulated feed and mixed feed groups showed weaker effects [24]. The expression trend of the glycolysis-related gene eno3 (Figure 2g) was the opposite: it was the highest in the CB group, reaching about 1.7 U/gprot, significantly higher than the other two groups(p<0.05), followed by the MIX group (about 1.1 U/gprot), and the lowest in the FF group, only about 0.8 U/gprot. There was no significant difference between the MIX group and the FF group(p>0.05). This indicates that the diet mainly composed of small fish carcasses may more strongly induce the expression of genes related to energy metabolism [25].
Enhanced lysosomal function: The expression level of cts1a (lysosomal protease) in the CB group was the highest (about 1.2 U/gprot), suggesting that cannibalism of conspecifics may reduce the risk of pathogen infection by improving antigen processing and clearance ability. This is consistent with the phenomenon found by Corcos, D. (2015) in carnivorous fish that cannibalistic behavior is accompanied by the upregulation of immune-related genes [26].
Efficient energy metabolism: The expression level of eno3 (glycolysis gene) in the CB group was significantly higher than that of the other groups (P < 0.05), indicating that it may quickly meet the energy demand through a high glycolysis rate. This is consistent with the metabolic adaptation of carnivorous fish to high-protein feed [27,28].
4.3. Limitations and Optimization Directions of Artificial Feed (FF)
Growth short-board: The weight gain rate and specific growth rate of the FF group were the lowest (P < 0.05), which may be due to the poor palatability of artificial feed, single nutritional components (such as lack of essential fatty acids like DHA and EPA), or the inhibition of digestion and absorption by anti - nutritional factors (such as phytic acid). This is consistent with the conclusion of Le Pape et al. (2015) and Melaku et al. (2024) in the study of artificial feed for marine fish that single feed is likely to cause slow growth of juvenile fish [20,29].
Potential advantages: The MDA content in the FF group was significantly higher (P < 0.05) than that in the CB group. This indicates that the proportion of artificial feed in the diet affects the production of MDA, and the higher the content of artificial feed in the diet, the higher the amount of MDA produced [30]. The expression of ppl (Figure 2h) was the highest in the FF group, reaching about 1.05 U/gprot, decreased significantly to about 0.5 U/gprot in the MIX group, and was the lowest in the CB group, only about 0.2 U/gprot. This result indicates that formulated feed can more effectively induce the expression of ppl [31]. Among them, the expression level of tegt (Figure 2i) was the highest in the FF group (about 0.85 U/gprot), significantly higher than that in the MIX group (about 0.2 U/gprot) and the CB group (about 0.15 U/gprot). This result indicates that formulated feed can significantly promote the expression of tegt, while the diet of small fish carcasses or mixed feed showed lower expression [32]. The FF group had higher expression levels of SOD activity (antioxidant), hsp90 (stress protection), and IL1β (anti-inflammatory), indicating its advantages in immune homeostasis and stress adaptation. This suggests that artificial feed can further improve growth performance by adding immune enhancers (such as probiotics and β-glucan) or nutritional fortifiers (such as selenium and vitamin E) while retaining its health advantages [33,34].
5. Conclusions
Mixed feeding (MIX): It is the best feeding strategy for the growth performance of juvenile barramundi. However, its “rapid growth” is accompanied by risks of liver damage, decreased antioxidant capacity, and imbalance of immune homeostasis. Nutritional regulation (such as adding liver-protecting agents and antioxidants) is needed to reduce the health costs.
Cannibalism of conspecifics (CB): It can be used as an emergency nutritional source under natural conditions. Its high lysosomal activity and glycolysis efficiency provide a physiological basis for the carnivorous adaptation of barramundi. However, environmental optimization (such as reasonable stocking density and sufficient feed) is needed in aquaculture to reduce cannibalistic behavior.
Artificial feed (FF): When used alone, it has a poor growth effect. However, its characteristic of high expression of antioxidant enzymes suggests that in the future, the coordinated improvement of “growth-health” can be achieved through formula optimization (such as adding bioactive substances).
Application suggestions: In the industrial larval rearing of barramundi, it is recommended to adopt the strategy of “mainly mixed feeding and supplemented with artificial feed”, and add antioxidants (such as vitamin C) and immune enhancers specifically to balance the growth rate and aquaculture benefits.
Author Contributions
Conceptualization, Junhua Huang and Shengjie Zhou; Methodology, Yun Wei and Yuzhuo You; Software, Songyuan Liu and Yun Wei; Validation, Zhengyi Fu and Yuzhuo You; Formal Analysis, Yun Wei and Songyuan Liu; Investigation, Junhua Huang and Jing Hu; Resources, Zemin Bai and Zhenmin Bao; Data Curation, Yun Wei and Junhua Huang; Writing—Original Draft, Yun Wei and Shengjie Zhou.; Writing—Review and Editing, Zhengyi Fu., and Zhenmin Bao; Visualization, Jing Hu and Zemin Bai; Supervision, Zhenmin Bao; Project Administration, Zemin Bai and Jing Hu; Funding Acquisition, Zemin Bai. All authors have read and approved the final version of the manuscript.
Funding
This work was supported by the Hainan Province Natural Science Foundation enterprise talent project (grant number 324QY579), Central Fund for Guiding Local Science and Technology Development (guike ZY22096005, guike AD21238026). Central Public-interest Scientific Institution Basal Research Fund, CAFS(2025XT05)
Institutional Review Board Statement
The animal study was reviewed and approved by the Animal Care and Use Committee of the South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences. The ethical code is 324QY579, which was approved in April 2024.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data supporting the results of this study can be obtained from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Pereira, L.S.; Agostinho, A.A.; Winemiller, K.O. Revisiting cannibalism in fishes. Reviews in fish biology and fisheries 2017, 27, 499-513. [CrossRef]
- Ward, A.J.; Webster, M.M.; Hart, P.J. Intraspecific food competition in fishes. Fish and Fisheries 2006, 7, 231-261. [CrossRef]
- Fox, L.R. Cannibalism in natural populations. Annual review of ecology and systematics 1975, 87-106.
- Hecht, T.; Pienaar, A.G. A review of cannibalism and its implications in fish larviculture. Journal of the World Aquaculture Society 1993, 24, 246-261. [CrossRef]
- Yue, G.H. Asian seabass (Lates calcarifer): a versatile species for marine and freshwater aquaculture. Reviews in Aquaculture 2025, 17, e70054. [CrossRef]
- Huang, J.; Fu, Z.; Yu, W.; Bai, Z.; Ma, Z. Toxic Effects of Carbaryl Exposure on Juvenile Asian Seabass (Lates calcarifer). Journal of Xenobiotics 2024, 14, 923-938. [CrossRef]
- Islam, M.A.; Bosu, A.; Hasan, M.M.; Yasmin, F.; Khan, A.B.S.; Akhter, M.; Ullah, M.R.; Karim, E.; Rashid, M.H.; Mahmud, Y. Culture technique of seabass, Lates calcarifer in Asia: A review. International Journal of Science and Technology Research Archive 2023, 4, 6-17. [CrossRef]
- Kubitza, F.; Lovshin, L.L. Formulated diets, feeding strategies, and cannibalism control during intensive culture of juvenile carnivorous fishes. Reviews in Fisheries Science 1999, 7, 1-22. [CrossRef]
- Kalaiselvan, P.; Ranjan, A.; Nazir, M.I.; Suresh, E. Exploring ontogenic development and larval rearing of striped murrel (Channa striatus). Aquaculture International 2024, 32, 9711-9754. [CrossRef]
- Fan, H.; Huang, P.; Zhu, J.; Li, J.; Jian, S.; Hou, J.; Wang, Z.; Li, L.; Zhao, D. Effects of Artificial Compound Feed for Live Fish Feed on Growth Performance, Nutrient Composition, Digestive Performance, and Gut Microbiota of Giant Salamander (Andrias davidianus). Fishes 2024, 10, 11. [CrossRef]
- Khan, N.; Qureshi, N.A.; Nasir, M.; Rasool, F.; Iqbal, K.J. Effect of artificial diet and culture systems on sensory quality of fried fish flesh of Indian major carps. Pakistan J. Zool 2011, 43, 1177-1182.
- Murray, R.L.; Tah, S.; Koprivnikar, J.; Rowe, L.; McCauley, S.J. Exposure to potentially cannibalistic conspecifics induces an increased immune response. Ecological Entomology 2020, 45, 355-363. [CrossRef]
- Huang, J.; Fu, Z.; Liu, X.; Ma, Z. Splenic tissue injury and physiological response mechanisms in juvenile yellowfin tuna (Thunnus albacares) under acute cold stress. Developmental & Comparative Immunology 2025, 105421. [CrossRef]
- Xue, S.; Ding, J.; Li, J.; Jiang, Z.; Fang, J.; Zhao, F.; Mao, Y. Effects of live, artificial and mixed feeds on the growth and energy budget of Penaeus vannamei. Aquaculture Reports 2021, 19, 100634. [CrossRef]
- Wang, J.; Xu, H.; Zuo, R.; Mai, K.; Xu, W.; Ai, Q. Effects of oxidised dietary fish oil and high-dose vitamin E supplementation on growth performance, feed utilisation and antioxidant defence enzyme activities of juvenile large yellow croaker (Larmichthys crocea). British Journal of Nutrition 2016, 115, 1531-1538. [CrossRef]
- Safari, O.; Shahsavani, D.; Paolucci, M.; Atash, M.M.S. Single or combined effects of fructo-and mannan oligosaccharide supplements on the growth performance, nutrient digestibility, immune responses and stress resistance of juvenile narrow clawed crayfish, Astacus leptodactylus leptodactylus Eschscholtz, 1823. Aquaculture 2014, 432, 192-203. [CrossRef]
- Li, Y.; Shi, J.; Shi, S.; Liu, Z.; Li, Z.; Li, J. Effect of live, frozen and artificial feeds on digestive enzymes, aminotransferase, histology of liver and intestine in mandarin fish hybrid (Siniperca chuatsi♀× Siniperca scherzeri♂). 2015.
- Chatzifotis, S.; Papadaki, M.; Despoti, S.; Roufidou, C.; Antonopoulou, E. Effect of starvation and re-feeding on reproductive indices, body weight, plasma metabolites and oxidative enzymes of sea bass (Dicentrarchus labrax). Aquaculture 2011, 316, 53-59. [CrossRef]
- Lushchak, V.I.; Bagnyukova, T.V. Effects of different environmental oxygen levels on free radical processes in fish. Comparative Biochemistry and Physiology Part B Biochemistry and Molecular Biology 2006, 144, 283-289. [CrossRef]
- Melaku, S.; Geremew, A.; Getahun, A.; Mengestou, S.; Belay, A. Challenges and prospects of using live feed substitutes for larval fish. Fisheries and Aquatic Sciences 2024, 27, 475-487. [CrossRef]
- Romano, N.; Zeng, C. Cannibalism of decapod crustaceans and implications for their aquaculture: a review of its prevalence, influencing factors, and mitigating methods. Reviews in Fisheries Science & Aquaculture 2017, 25, 42-69. [CrossRef]
- Lim, K.C.; Yusoff, F.M.; Shariff, M.; Kamarudin, M.S.; Nagao, N. Dietary supplementation of astaxanthin enhances hemato-biochemistry and innate immunity of Asian seabass, Lates calcarifer (Bloch, 1790). Aquaculture 2019, 512, 734339. [CrossRef]
- Vazquez, A.; Liu, J.; Zhou, Y.; Oltvai, Z.N. Catabolic efficiency of aerobic glycolysis: the Warburg effect revisited. BMC systems biology 2010, 4, 58. [CrossRef]
- Johnstone, C.; Chaves-Pozo, E. Antigen presentation and autophagy in teleost adaptive immunity. International journal of molecular sciences 2022, 23, 4899. [CrossRef]
- Gong, Y.; Chen, W.; Han, D.; Zhu, X.; Yang, Y.; Jin, J.; Liu, H.; Xie, S. Effects of food restriction on growth, body composition and gene expression related in regulation of lipid metabolism and food intake in grass carp. Aquaculture 2017, 469, 28-35. [CrossRef]
- Corcos, D. Food–Nonfood Discrimination in Ancestral Vertebrates: Gamete Cannibalism and the Origin of the Adaptive Immune System. Scandinavian Journal of Immunology 2015, 82, 409-417. [CrossRef]
- Sánchez-Muros, M.a.J.; Garcı́a-Rejón, L.; Garcı́a-Salguero, L.; Lupiáñez, J.A. Long-term nutritional effects on the primary liver and kidney metabolism in rainbow trout. Adaptive response to starvation and a high-protein, carbohydrate-free diet on glutamate dehydrogenase and alanine aminotransferase kinetics. The international journal of biochemistry & cell biology 1998, 30, 55-63. [CrossRef]
- Clemente-Suárez, V.J.; Redondo-Flórez, L.; Beltrán-Velasco, A.I.; Yáñez-Sepúlveda, R.; Rubio-Zarapuz, A.; Martín-Rodríguez, A.; Navarro-Jimenez, E.; Tornero-Aguilera, J.F. Human Digestive Physiology and Evolutionary Diet: A Metabolomic Perspective on Carnivorous and Scavenger Adaptations. Metabolites 2025, 15, 453. [CrossRef]
- Le Pape, O.; Bonhommeau, S. The food limitation hypothesis for juvenile marine fish. Fish and Fisheries 2015, 16, 373-398. [CrossRef]
- Zhang, Z.; Yuan, X.; Wu, H.; Gao, J.; Wu, J.; Xiong, Z.; Feng, Z.; Xie, M.; Li, S.; Xie, Z. The effect of short-term artificial feed domestication on the expression of oxidative-stress-related genes and antioxidant capacity in the liver and gill tissues of Mandarin fish (Siniperca chuatsi). Genes 2024, 15, 487. [CrossRef]
- Boczonadi, V.; Määttä, A. Functional analysis of periplakin and envoplakin, cytoskeletal linkers, and cornified envelope precursor proteins. Methods in enzymology 2016, 569, 309-329. [CrossRef]
- Fan, Y.; Wang, Z.; Ren, C.; Ma, T.; Deng, K.; Feng, X.; Li, F.; Wang, F.; Zhang, Y. Effect of dietary energy restriction and subsequent compensatory feeding on testicular transcriptome in developing rams. Theriogenology 2018, 119, 198-207. [CrossRef]
- Dawood, M.A.; Koshio, S.; Esteban, M.Á. Beneficial roles of feed additives as immunostimulants in aquaculture: a review. Reviews in Aquaculture 2018, 10, 950-974. [CrossRef]
- Hu, X.; Ma, W.; Zhang, D.; Tian, Z.; Yang, Y.; Huang, Y.; Hong, Y. Application of natural antioxidants as feed additives in aquaculture: A review. Biology 2025, 14, 87. [CrossRef]
Figure 1.
Effects of different baits on the physiological and biochemical indicators of Lates calcarifer. Note: Different letters indicate significant differences (p < 0.05). The same letters represent no significant difference (p > 0.05).
Figure 1.
Effects of different baits on the physiological and biochemical indicators of Lates calcarifer. Note: Different letters indicate significant differences (p < 0.05). The same letters represent no significant difference (p > 0.05).
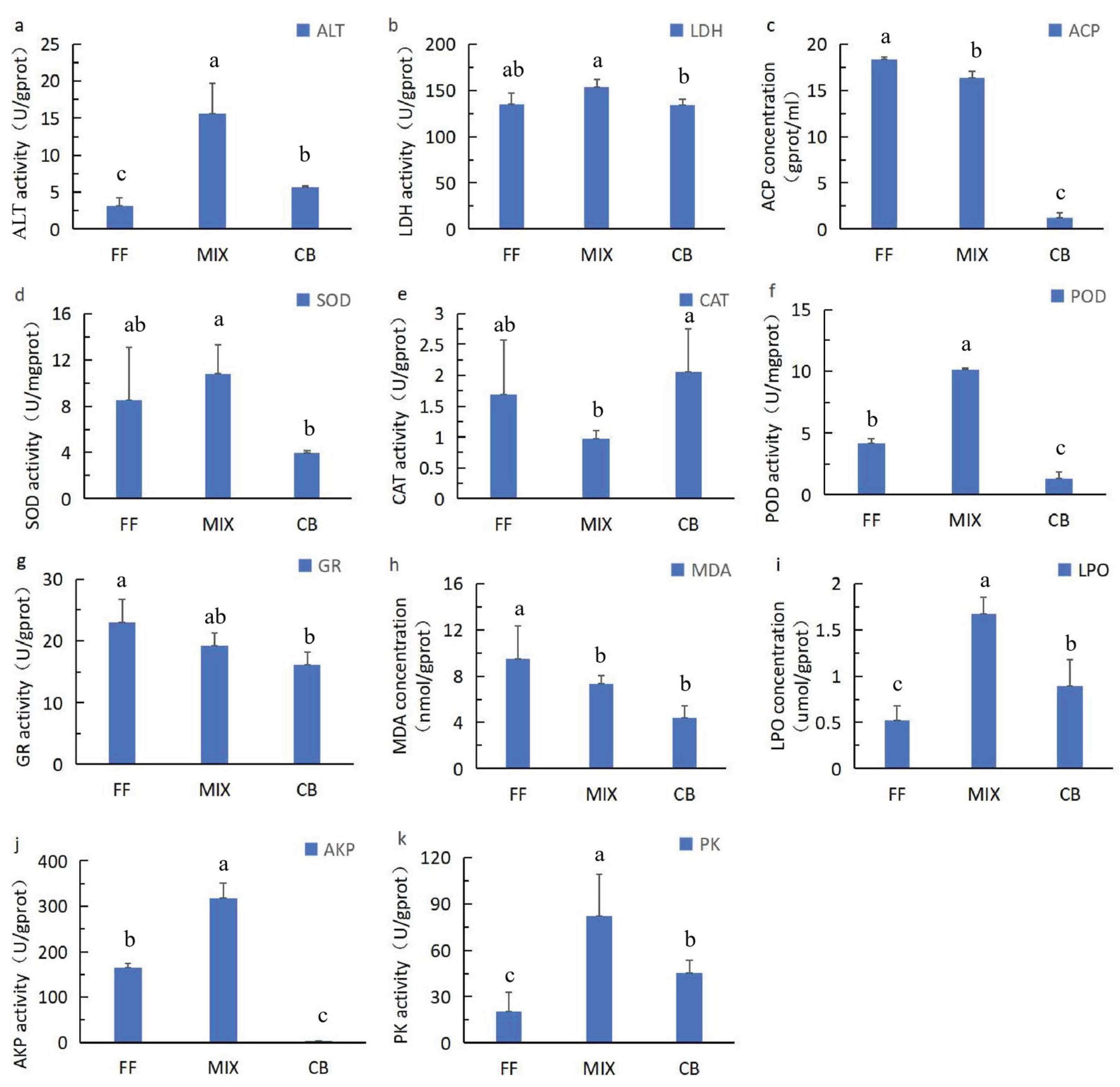
Figure 2.
Effects of different baits on the relative gene expression levels of Lates calcarifer. Note: Different letters indicate significant differences (p < 0.05). The same letters represent no significant difference (p > 0.05).
Figure 2.
Effects of different baits on the relative gene expression levels of Lates calcarifer. Note: Different letters indicate significant differences (p < 0.05). The same letters represent no significant difference (p > 0.05).
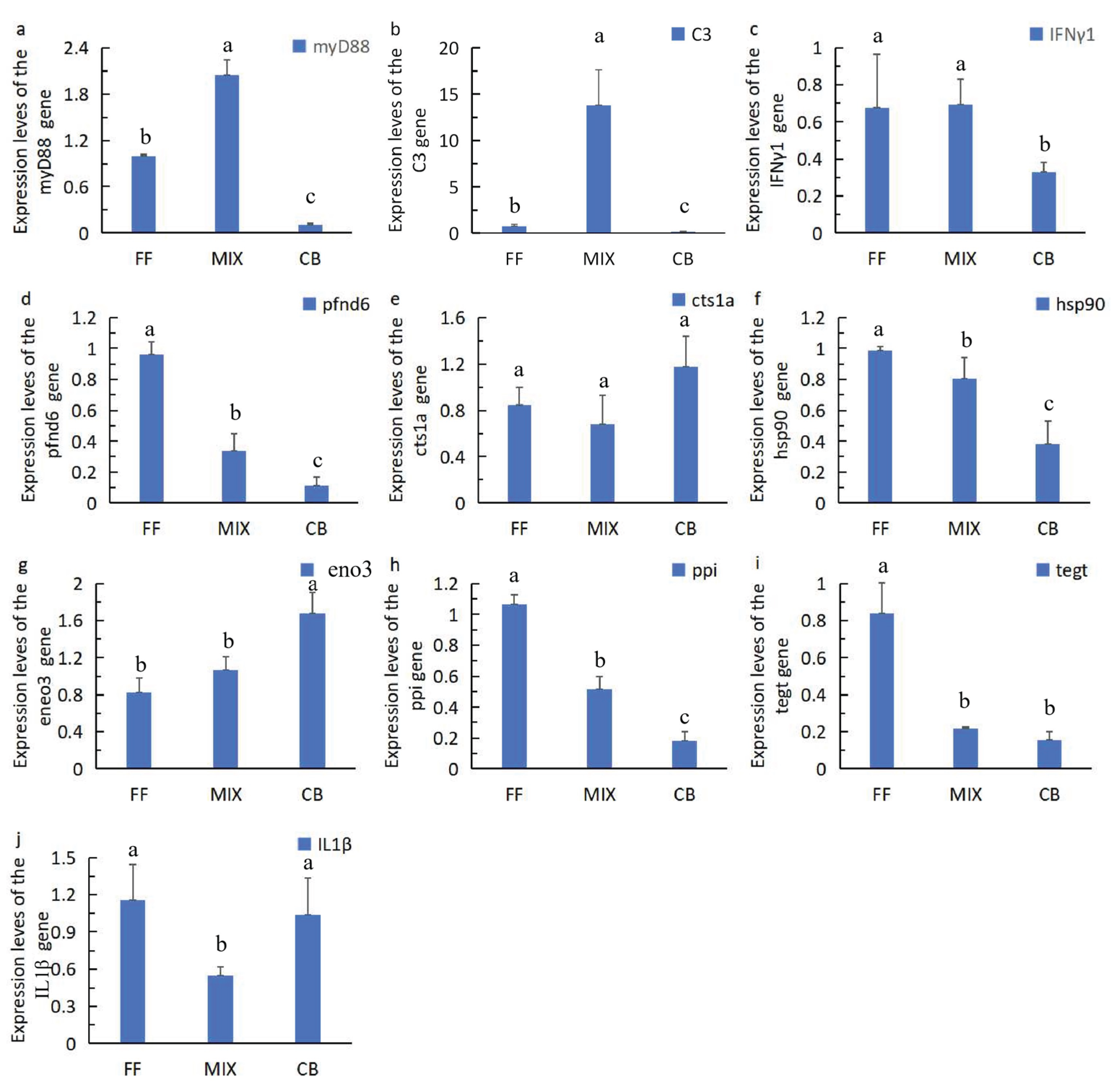

Table 1.
Sequences of specific primers used for qRT-PCR analysis.
| Gene Abbreviation |
Primer Sequence (5’–3’) | Amplicon Size (bp) | Accession NO | |
|---|---|---|---|---|
| Immune-related genes | C3 | F: AAATGCTGCCATCGTTCC R: CCAGTGACCTTCAGACCAAA |
175 | XM_018679796 |
| myD88 | F: TCCAACTTGCCTCCTAAT R: AAGCGTGATACAGAAACC |
111 | XM_018673801.2 | |
| IFNγ1 | F: TACCAGGAGCAGGACAAGC R: TCGTCAGGCAGCGAACTT |
134 | NM_001360734 | |
| pfnd6 | F: TCCCGTAAGGACCAACAG R: GCACCCAGATAGAGGAAGG |
173 | JQ247193.1 | |
| cts1a | F: CACATCCGTCCGTCTTCT R: GCTCAGCGACACCAGTTT |
120 | EU143238.1 | |
| Metabolism genes | hsp90 | F: ACGATGATGAGCAGTATGCC R: CAAACAGGGTGATGGGGTA |
210 |
XM018661637 |
| eno3 | F: GCATAGCAAGAGACAGTGC R: GGAGGTGTAGATGTATTGTC |
100 | XM_018666223.2 | |
| ppl | F: ATTCCCCAGGTGATGAGTC R: GCGAATGTGTCGTGTGTCT |
104 | XM_018697911.2 | |
| Stress Response genes |
β-actin | F: AACCAAACGCCCAACAACT R: ATAACTGAAGCCATGCCAATG |
112 | XM_018667666 |
| IL1β | F: GACCCTACACCCAGCAAC R: GACGTCCAATGGGCTTTCT |
174 | XM_018669006.1 | |
| tegt | F: ATTCCCCAGGTGATGAGTC R: GCGAATGTGTCGTGTGTCT |
111 | XM_018686203.2 | |
Note: C3(Complement component 3), myD88(Myeloid differentiation primary response 88), IFNγ1(Interferon gamma 1), pfnd6(Profilin2 domain family member 6), cts1a(Cathepsin A), hsp90(Heat Shock Protein 90), eno3(enolase 3),ppl(periplakin), IL1β(Interleukin 1 beta), β-actin(beta-actin), tegt(testis enhanced gene transcript).
Table 2.
Effects of different feeding strategies on the growth performance of juvenile Lates calcarifer.
Table 2.
Effects of different feeding strategies on the growth performance of juvenile Lates calcarifer.
| Initial Weight(g) | Weight Gain Rate(%) | Specific Growth Rate(%/d-1) | |
|---|---|---|---|
| FF | 0.83±0.16a | 496.68±342.92a | 41.89±56.19a |
| MIX | 0.98±0.53a | 862.31±346.66a | 93.2±42.48a |
| CB | 10.3±0.53a | 542.16±61.64a | 62.61±38.27a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



