1. Introduction
Ellagitannins from pomegranate, walnuts, and some berries are converted by intestinal microbial consortia into urolithins, a family of dibenzopyran-6-one metabolites. Among these, urolithin A (UA) is the best characterized and has been extensively reviewed for its metabolism, bioactivity, and associated gut microbiota [
1]. Interindividual differences in microbial composition lead to distinct urolithin metabotypes (UMA, UMB, and UM0), ranging from robust UA producers to nonproducers [
2]. A recent systematic review summarized bacteria implicated in ellagitannin transformation to urolithins and highlighted the biological and technical sources of variability that underlie these metabotypes [
3].
UA exhibits anti-inflammatory, antioxidant, and mitochondrial effects (including mitophagy induction) in experimental systems [
4,
5,
6]. At physiologically relevant concentrations, UA and its conjugates can attenuate pro-inflammatory signaling, including TNF-α-driven endothelial activation [
5] and broader inflammatory cascades [
6]. Importantly, urolithins and their conjugates have been reported to cross the blood-brain barrier (BBB) and modulate stimulus-dependent microglial activation [
7]. Reviews and preclinical studies position UA as a candidate postbiotic for brain aging and neurodegenerative trajectories, including Alzheimer’s disease models [
8,
9].
Cancer-associated gut dysbiosis is well documented in colorectal cancer (CRC), where reduced diversity and altered microbial metabolite profiles are consistently reported [
10,
11]. Although evidence is less mature in extraintestinal cancers, associations between gut dysbiosis and non-small-cell lung cancer (NSCLC) risk have been described and may influence treatment response [
12]. These observations support the concept that microbial metabolic capacity—not only taxonomic diversity—may modulate host inflammation and potentially tumor biology.
Melatonin is a pineal-derived hormone with pleiotropic immunomodulatory and oncostatic actions. Its major urinary metabolite, 6-sulfatoxymelatonin (6-SMT), is commonly used as a noninvasive surrogate of nocturnal melatonin secretion. Prospective studies have reported inverse associations between urinary 6-SMT and breast cancer risk in some cohorts [
16,
17,
18], and night shift work is associated with reduced 6-SMT and altered cortisol profiles [
19]. Urinary melatonin-sulfate has also been evaluated in prostate cancer [
20,
21] and prostate cancer risk studies [
22].
Despite converging evidence linking (i) microbiota-dependent ellagitannin-urolithin metabolism, (ii) cancer-associated dysbiosis, and (iii) circadian/melatonin biomarkers, these dimensions have rarely been assessed together in advanced solid tumors. We hypothesized that a composite signature involving selective dysbiosis, systemic and fecal UA depletion, and decreased 6-SMT would track with tumor aggressiveness. Here, we report a three-year pilot observational study comparing two advanced tumor cohorts and healthy controls, and we propose an integrated dysbiosis-UA-melatonin triad as a candidate composite biomarker and mechanistic framework.
2. Materials and Methods
2.1. Study Design and Participants
This pilot observational study was conducted over a three-year period. Adult Caucasian oncology patients were prospectively observed and categorized into two cohorts based on tumor aggressiveness and clinical status, and compared to a healthy control cohort.
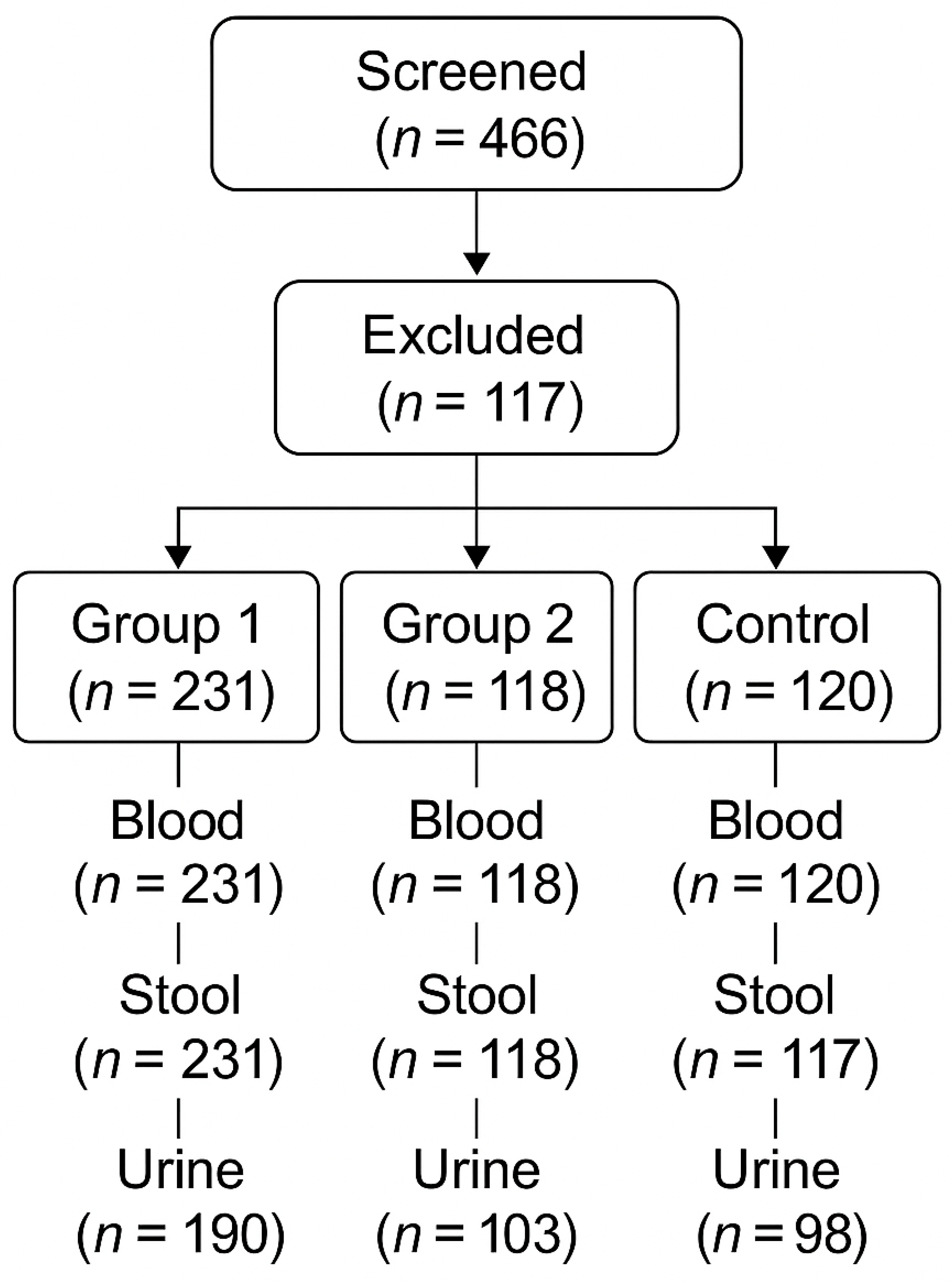
A total of 466 oncology patients were screened; 117 were excluded; and 349 were included and allocated to Group 1 (n = 231) or Group 2 (n = 118) (
Figure 1). Group 1 (high aggressiveness) included stage IV colorectal, hepatic, pancreatic, NSCLC, SCLC, TNBC, and glioblastoma cases (n = 231; 107 male, 124 female). Group 2 (lower aggressiveness) included stage IV ER/PR+++ HER2-negative breast cancer, WHO grade 2 astrocytoma, and stage IV carcinoid tumors (n = 118; 77 male, 41 female).
Healthy controls were recruited as an age- and sex-matched Caucasian cohort without known malignancy (n = 117; 75 male, 42 female). Additional control blood specimens were available for some analyses (
Figure 1).
Participant flow and specimen availability for blood, stool, and urine are summarized in
Figure 1.
2.2. Sample Collection and Processing
Blood, stool, and 24 h urine samples were collected according to a standardized protocol. Plasma was isolated from blood and stored under conditions appropriate for metabolite and cytokine analyses.
Stool specimens were processed for UA-related metabolite measurement and for culture-based microbiota assessment. Urine was collected over 24 h for quantification of 6-SMT.
2.3. Quantification of Urolithin A in Plasma and Stool
UA was quantified by high-performance liquid chromatography (HPLC) in plasma and stool extracts. The assay targeted UA and UA-related fecal metabolites; concentrations below the assay’s detection threshold were classified as undetectable.
All available plasma specimens were analyzed for UA. Stool UA analysis was performed for all available stool samples in each cohort (
Figure 1).
2.4. Culture-Based Microbiota Assessment
Stool cultures were performed using selective and differential media with particular attention to Streptococcus thermophilus, Enterococcus faecium, and Bacteroidetes, taxa hypothesized to relate to urolithin production and/or dysbiosis severity. Results were interpreted qualitatively (present/depleted/absent) in relation to the clinical grouping.
2.5. Inflammatory Cytokines and Urinary 6-SMT
Serum IL-6, TNF-α, IL-8, and IL-10 were measured using immunoassays performed according to the manufacturers’ instructions. Cytokine results were summarized as cohort-level distributions.
Twenty-four-hour urinary 6-SMT was measured as an index of melatonin output. All values were normalized to daily excretion (ng/day).
2.6. Statistical Analysis
Because this study was designed as a pilot observational investigation, analyses were primarily descriptive and focused on between-cohort contrasts. Continuous variables are reported using medians and interquartile ranges (IQRs) where available; categorical variables are reported as counts and percentages. Where comparisons were performed, nonparametric tests were used for continuous variables and χ² tests for categorical variables. A two-sided p-value < 0.05 was considered statistically significant.
3. Results
3.1. Study Design, Cohorts, and Specimen Availability
Oncology patients were screened (n = 466) and excluded (n = 117) according to protocol criteria, yielding 349 included patients allocated to Group 1 (n = 231) and Group 2 (n = 118). A healthy control cohort (n = 117) was enrolled in parallel.
Specimen availability for downstream analyses is summarized in
Figure 1. Blood and stool samples were available for all participants in Groups 1 and 2. For controls, blood was available for 120 individuals and stool for 117 individuals (
Figure 1). Twenty-four-hour urine samples were available for 190 participants in Group 1, 103 in Group 2, and 98 controls (
Figure 1).
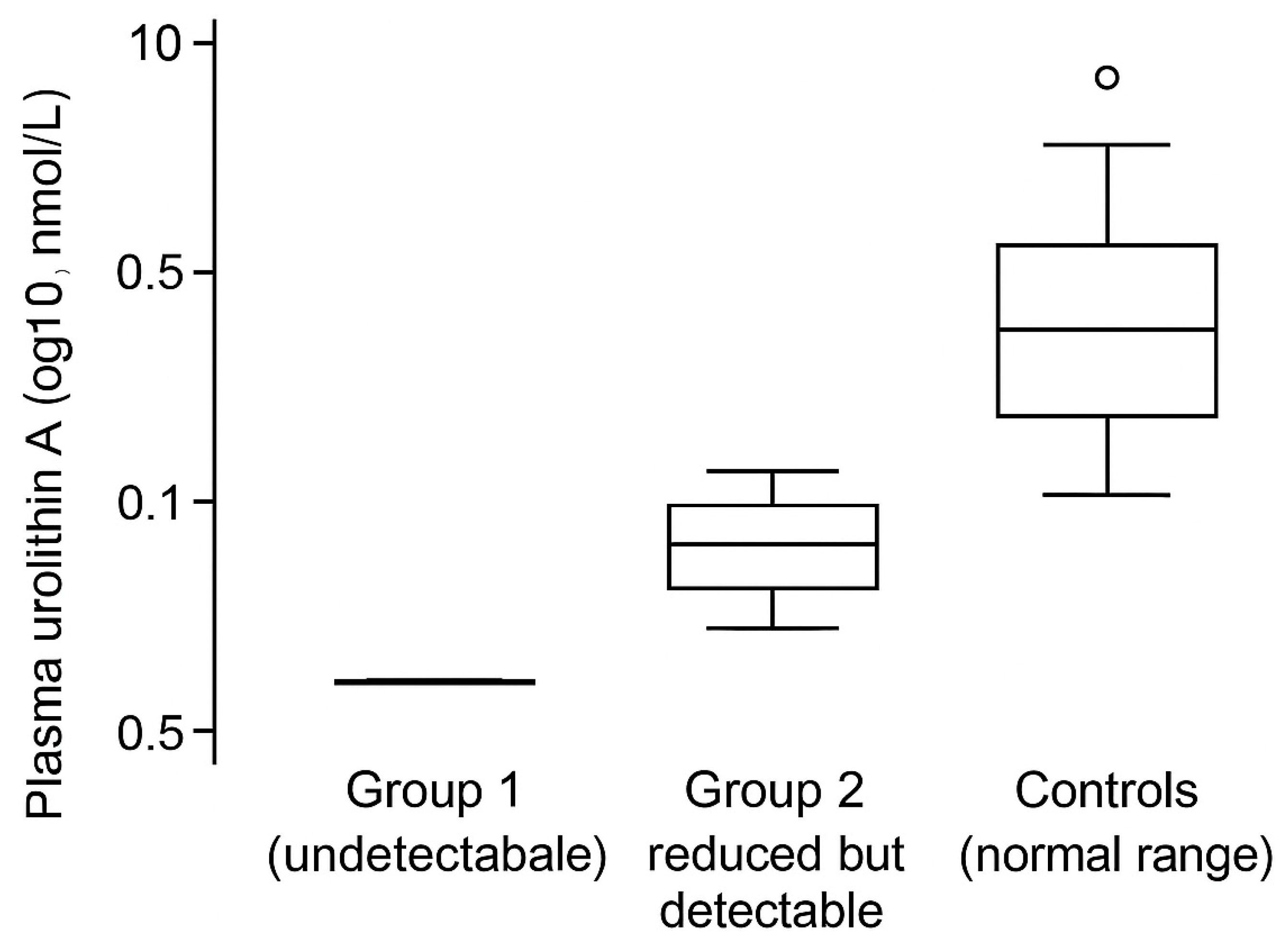
3.2. Plasma and Fecal Urolithin A Are Profoundly Depleted in the High-Aggressiveness Tumor Cohort
Plasma UA concentrations differed sharply across cohorts. In Group 1, UA was undetectable in plasma and fecal samples, consistent with a functional loss of urolithin-producing capacity. In Group 2, UA was reduced but remained detectable, whereas controls exhibited UA concentrations within the expected range (
Figure 2).
These group-level patterns suggest that advanced, high-mortality tumors may be accompanied by a collapse of the ellagitannin-to-urolithin metabolic axis, potentially reflecting both taxonomic dysbiosis and metabolic dysfunction (
Table 2).
3.3. Selective Dysbiosis of Putative Urolithin-Associated Taxa
Culture-based stool analysis showed marked depletion or absence of
Streptococcus thermophilus and
Enterococcus faecium in Group 1, along with a critical reduction in Bacteroidetes. Group 2 showed a moderate reduction in these taxa, while controls generally retained these groups (
Table 2).
While culture-based approaches provide limited taxonomic resolution, the reproducibility of the selective depletion pattern across participants supports the hypothesis that specific taxa (or consortia) may be linked to UA production status and overall dysbiosis severity.
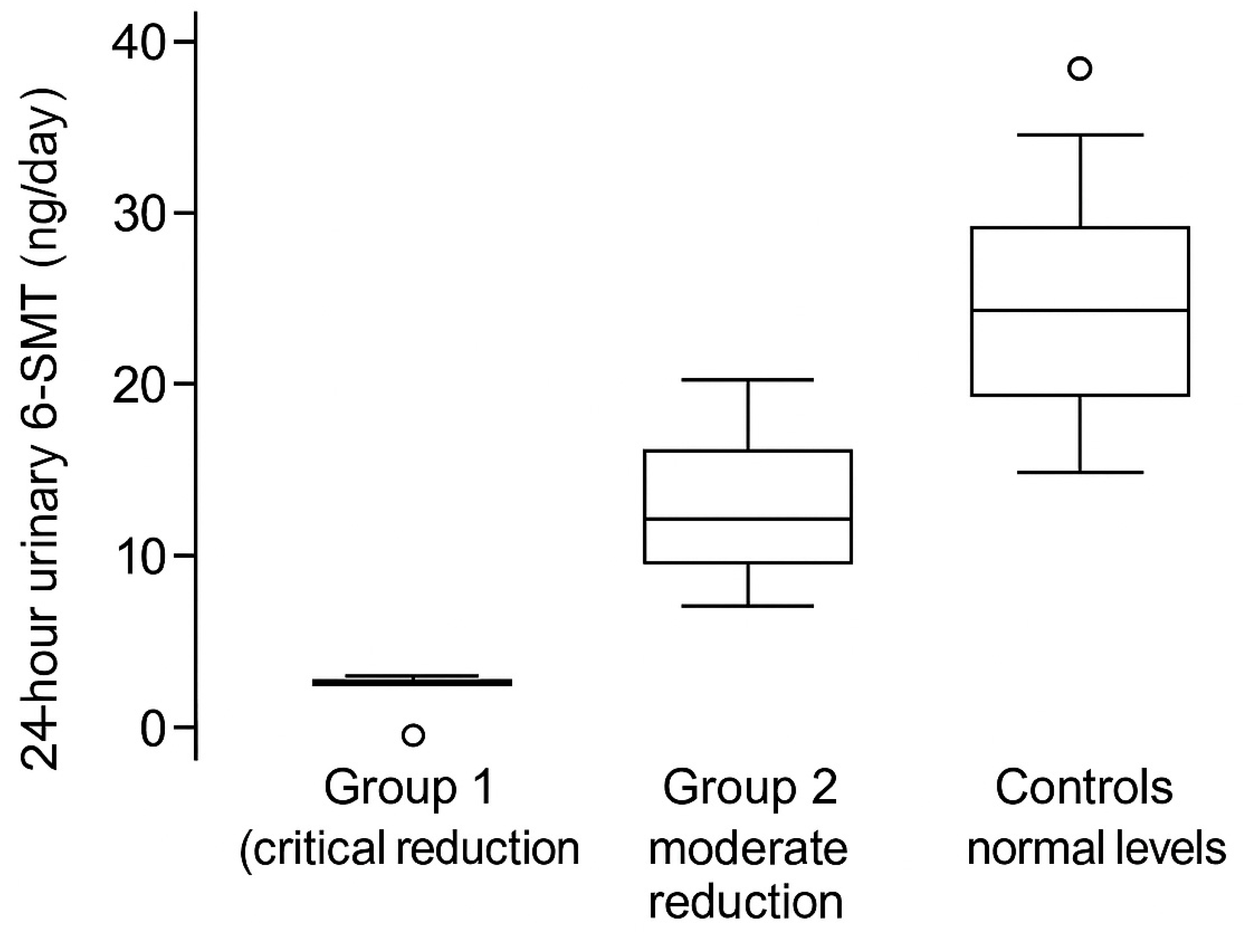
3.4. Urinary 6-SMT Is Reduced in Parallel with UA Depletion
Twenty-four-hour urinary 6-SMT was critically reduced in Group 1 and moderately reduced in Group 2, compared with controls (
Figure 3). This gradient suggests that diminished melatonin output may track with tumor aggressiveness and with the extent of UA depletion.
Notably, a subset of Group 1 exhibited near-zero 6-SMT excretion (
Figure 3), consistent with profoundly impaired nocturnal melatonin production or altered metabolism/excretion in the setting of systemic illness.
3.5. Pro-Inflammatory Cytokines Are Elevated with Preserved IL-10, Defining a Composite Triad
Group 1 displayed markedly elevated serum IL-6, TNF-α, and IL-8, whereas IL-10 remained within the normal range. Group 2 showed the same directionality with a smaller magnitude of elevation. Controls exhibited low levels of pro-inflammatory cytokines. Together, these findings indicate chronic, low-grade systemic inflammation that accompanies the dysbiosis-UA-melatonin pattern.
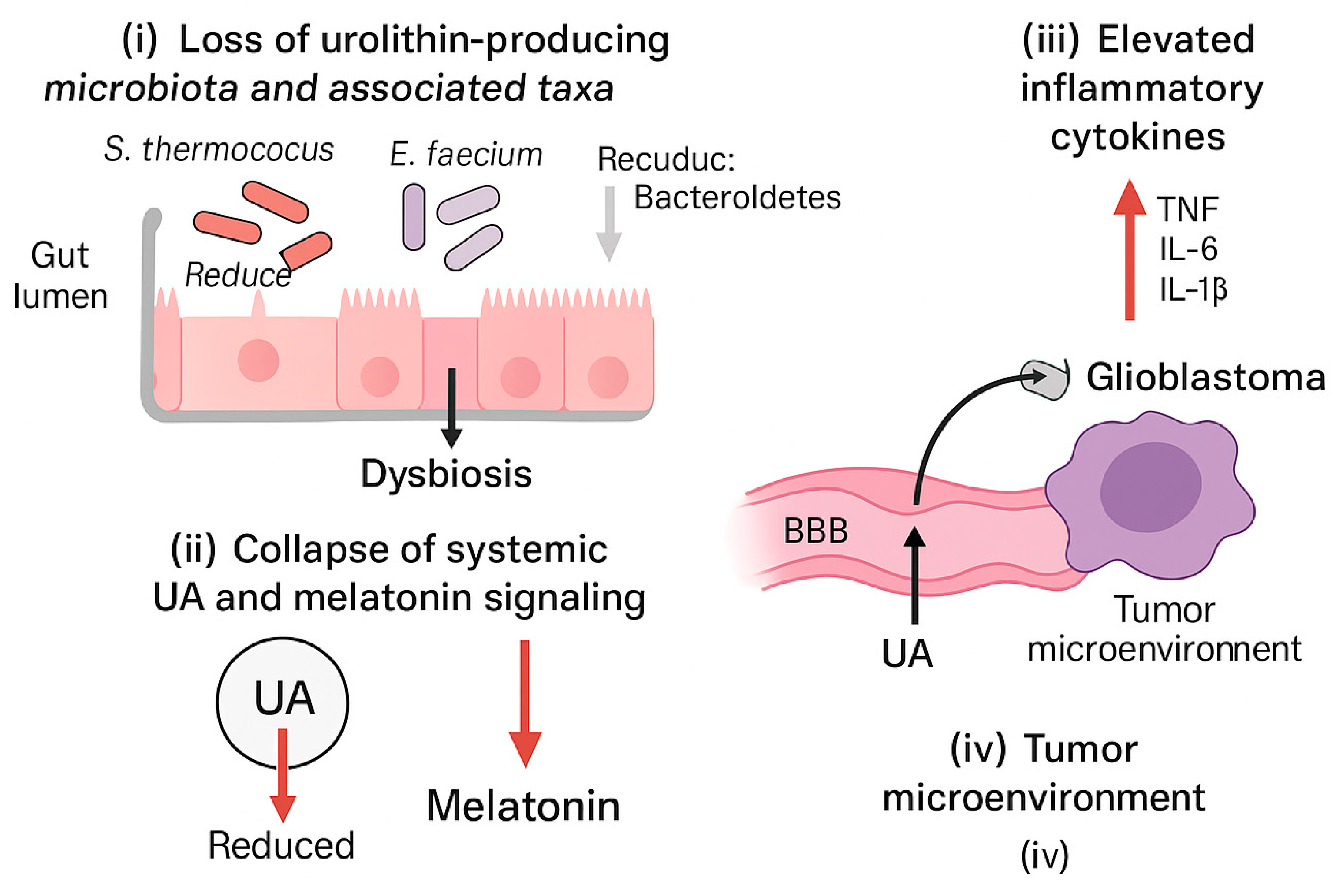
Based on the concordant patterns across compartments, we propose a composite dysbiosis-UA-melatonin triad characterized by (i) selective dysbiosis with depletion of putative urolithin-associated taxa, (ii) systemic and fecal UA depletion, and (iii) reduced 6-SMT coupled to elevated pro-inflammatory cytokines (
Figure 4;
Table 3).
Table 1.
Cohort definitions and baseline clinical characteristics (summary).
Table 1.
Cohort definitions and baseline clinical characteristics (summary).
| Cohort |
n |
Sex (M/F) |
Clinical definition / tumor types |
Notes |
| Group 1 (high aggressiveness) |
231 |
107/124 |
Stage IV colorectal, hepatic, pancreatic; NSCLC; SCLC; TNBC; glioblastoma |
UA undetectable; low 6-SMT |
| Group 2 (lower aggressiveness) |
118 |
77/41 |
Stage IV ER/PR+++ HER2-negative breast cancer; WHO grade 2 astrocytoma; stage IV carcinoid tumors |
UA reduced; 6-SMT reduced |
| Controls |
117 |
75/42 |
No known malignancy (healthy adults) |
UA detectable; 6-SMT normal |
Table 2.
Qualitative summary of key features across cohorts.
Table 2.
Qualitative summary of key features across cohorts.
| Parameter |
Group 1 |
Group 2 |
Controls |
| Plasma UA |
Undetectable |
Reduced but detectable |
Normal range |
| Fecal UA metabolites |
Undetectable |
Reduced |
Detectable |
| Streptococcus thermophilus |
Absent/depleted |
Reduced |
Present |
| Enterococcus faecium |
Absent/depleted |
Reduced |
Present |
| Bacteroidetes |
Critical reduction |
Moderate reduction |
Preserved |
| 24 h urinary 6-SMT |
Critical reduction |
Moderate reduction |
Normal levels |
| IL-6, TNF-α, IL-8 |
Markedly elevated |
Moderately elevated |
Low/normal |
| IL-10 |
Normal range |
Normal range |
Normal range |
Table 3.
Proposed dysbiosis-UA-melatonin triad categories for future validation.
Table 3.
Proposed dysbiosis-UA-melatonin triad categories for future validation.
| Category |
UA status |
6-SMT status |
Dysbiosis |
Cytokines |
Interpretation |
| A (triad-positive, high-risk) |
Undetectable |
Critical reduction |
Severe/selective depletion of taxa |
High IL-6/TNF-α/IL-8; IL-10 preserved |
Severe metabolic/inflammatory disruption |
| B (intermediate) |
Reduced but detectable |
Moderate reduction |
Moderate depletion of taxa |
Moderate elevation of IL-6/TNF-α/IL-8 |
Partial disruption; may track with lower aggressiveness |
| C (triad-negative) |
Normal range |
Normal levels |
No severe dysbiosis |
Normal cytokine profile |
Physiologic host–microbe axis preserved |
4. Discussion
4.1. Summary of Main Findings
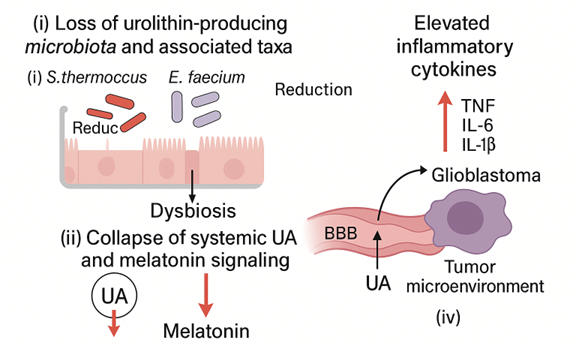
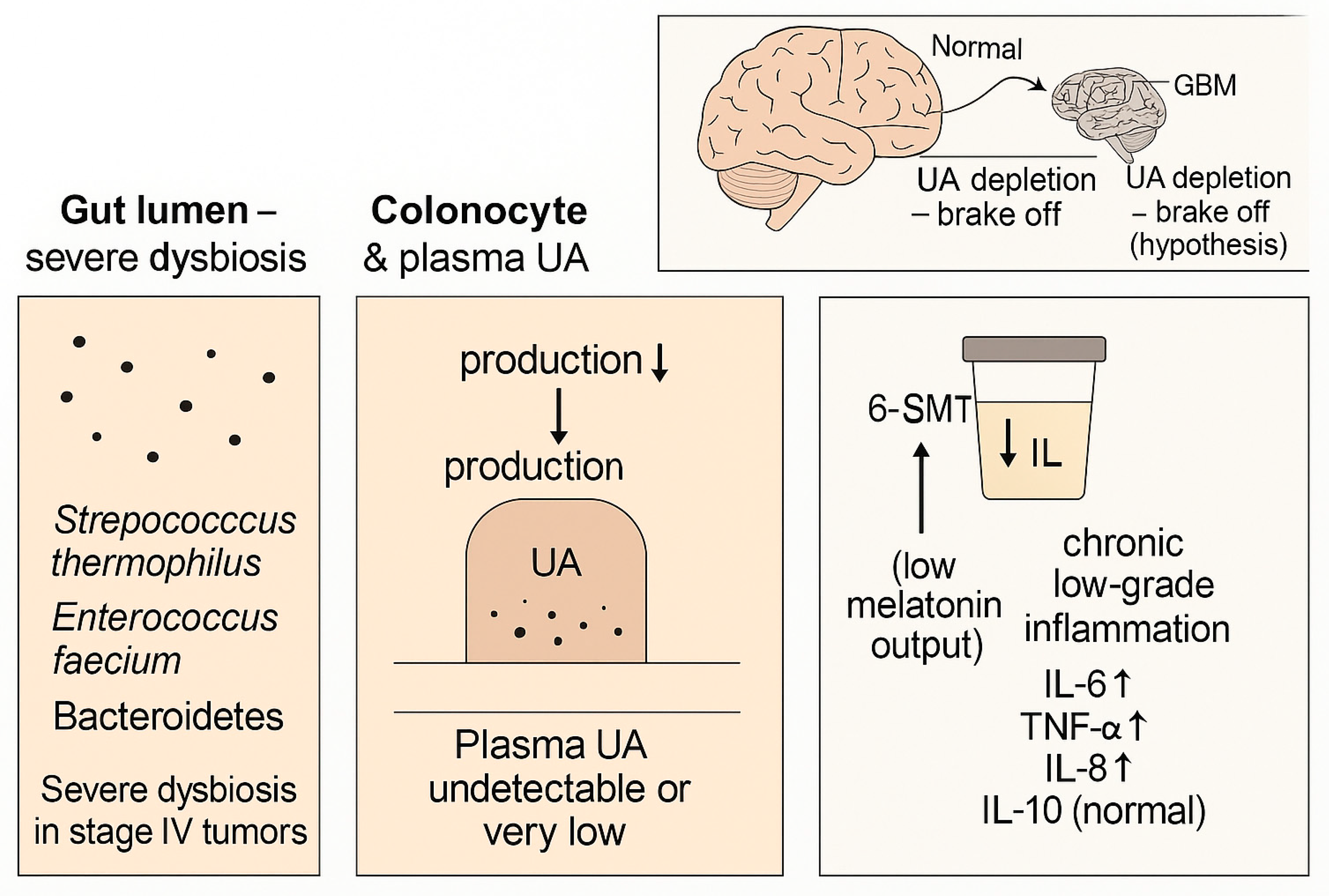
This pilot observational study suggests that advanced solid tumors with high mortality are associated with a reproducible multi-compartment pattern: (i) selective severe dysbiosis with depletion of key taxa, (ii) collapse of systemic and fecal UA availability, and (iii) reduced melatonin output (as 6-SMT) accompanied by elevated pro-inflammatory cytokines with preserved IL-10. In a lower-aggressiveness tumor cohort, the same pattern was present but attenuated, whereas controls exhibited measurable UA and normal 6-SMT.
We propose that this dysbiosis-UA-melatonin triad may serve as a composite biomarker framework for disease severity and as a hypothesis-generating model that integrates microbial metabolic capacity, systemic inflammation, and neuroendocrine status (
Figure 5).
4.2. Dysbiosis and Functional Loss of Urolithin Production
UA production depends on specific microbial consortia and is highly variable between individuals, giving rise to the recognized urolithin metabotypes [
1,
2,
3]. The observation that UA was undetectable in both plasma and stool in Group 1 suggests that, in at least a subset of advanced cancer patients, dysbiosis may extend beyond taxonomic shifts to a functional collapse of ellagitannin-to-urolithin biotransformation.
Because diet was not controlled and metagenomic profiling was not performed, we cannot distinguish whether UA depletion reflects loss of urolithin-producing taxa, suppression of relevant metabolic pathways, reduced substrate availability, or combined effects. Nonetheless, the concordance between culture-based depletion of selected taxa and the UA phenotype supports the plausibility of a metabolic bottleneck.
4.3. Linking UA Deficiency to Systemic Inflammation
UA has been reported to modulate inflammatory signaling and endothelial activation, including pathways driven by TNF-α [
5] and broader inflammatory cascades [
4,
6]. In our cohorts, UA depletion paralleled elevations in IL-6, TNF-α, and IL-8, consistent with a chronic inflammatory state. While causality cannot be inferred, one interpretation is that loss of a potentially anti-inflammatory postbiotic may contribute to a permissive cytokine milieu.
Alternatively, systemic illness, tumor burden, and therapy-related factors may drive both dysbiosis and inflammation, with UA depletion acting as a downstream marker of disrupted host–microbe metabolism. Longitudinal sampling will be required to determine temporality and directionality.
4.4. Potential Relevance to Glioblastoma and the BBB
A notable feature of our conceptual model is the potential intersection between UA availability and central nervous system (CNS) tumor biology. Urolithins and their conjugates have been reported to cross the BBB and to modulate microglial activation in experimental models [
7]. In parallel, UA has been discussed as a candidate postbiotic for brain aging and neurodegeneration, including Alzheimer’s disease models [
8,
9].
These observations motivate the hypothesis that systemic UA depletion could remove a protective ‘brake’ on neuroinflammatory or metabolic processes relevant to glioblastoma progression. This remains speculative and requires direct mechanistic testing in GBM models, including measurements of UA (and conjugates) in CNS compartments and evaluation of tumor-microenvironment interactions.
4.5. Melatonin Output as a Companion Signal in the Triad
Reduced urinary 6-SMT has been linked to cancer risk modulation in several epidemiologic settings, particularly in breast cancer cohorts [
16,
17,
18]. Circadian disruption and night shift work are associated with altered 6-SMT and cortisol profiles [
19]. Studies in prostate cancer further suggest that melatonin/6-SMT rhythms may relate to disease status [
20,
21], and urinary 6-SMT has been investigated in prostate cancer risk in a multiethnic cohort [
22].
In our cohorts, the reduction in 6-SMT tracked with tumor aggressiveness and with the UA phenotype. Whether this reflects altered pineal output, systemic inflammation, medication effects, sleep disruption, or changes in metabolism/excretion cannot be determined from this design. Nonetheless, incorporating a circadian/neuroendocrine biomarker alongside microbial-metabolic and inflammatory readouts may improve composite risk stratification.
4.6. Clinical Implications and Future Directions
If confirmed, the triad could inform screening or monitoring strategies in advanced solid tumors. Interventions might include dietary ellagitannin optimization, microbiota-directed therapies (e.g., prebiotics or carefully selected probiotics), direct UA supplementation, and/or circadian alignment strategies aimed at restoring melatonin rhythms.
Any microbiota-directed intervention should consider safety and strain specificity. For example, while certain
Enterococcus strains are used as probiotics and can produce bioactive exopolysaccharides [
13], functional properties of
Streptococcus thermophilus have been reviewed in food and industrial contexts [
14]. At the same time, concerns regarding enterococcal probiotic risk and the need for strain-level safety assessment have been highlighted [
15]. Future trials should prioritize well-characterized, nonpathogenic strains and incorporate metagenomic and metabolomic endpoints.
4.7. Limitations
This study has several limitations: (i) the observational design precludes causal inference; (ii) microbiota assessment relied on culture-based methods rather than sequencing; (iii) dietary intake of ellagitannins and use of antibiotics/probiotics were not quantitatively controlled; (iv) treatment exposures and other confounders were not modeled; and (v) detailed quantitative UA, cytokine, and 6-SMT distributions were not available for all participants in this report.
Accordingly, the findings should be interpreted as hypothesis-generating. Larger prospective cohorts with longitudinal sampling, comprehensive multi-omics profiling, and standardized dietary and circadian assessments are needed to validate the triad and clarify mechanisms.
5. Conclusions
In this pilot observational study, patients with highly aggressive advanced solid tumors exhibited a consistent triad comprising severe selective dysbiosis, collapse of systemic and fecal urolithin A availability, and reduced melatonin output (24 h urinary 6-SMT), accompanied by elevated pro-inflammatory cytokines with preserved IL-10. A lower-aggressiveness tumor cohort demonstrated an attenuated version of the same pattern, while controls showed measurable UA and normal 6-SMT.
These data motivate the dysbiosis-UA-melatonin triad as a candidate composite biomarker and a framework for mechanistic studies, including investigation of BBB-permeable postbiotics in glioblastoma biology. Prospective validation and intervention trials are warranted.
Author Contributions
All authors contributed equally to study design, patient management, data analysis, and manuscript preparation. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Institute for Personalized Medicine (Tbilisi, Georgia) and Foconsci Chemical Industry (Department of Biotechnology, China).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Institutional Review Board (IRB) under protocols #CN-2021-11 and #TX-UT-2021-08.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data are available from the corresponding author upon reasonable request.
Acknowledgments
The authors thank the patients, clinical teams, and laboratory staff for their dedication and support.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
UA, urolithin A; 6-SMT, 6-sulfatoxymelatonin; BBB, blood-brain barrier; CRC, colorectal cancer; GBM, glioblastoma; NSCLC, non-small-cell lung cancer; SCLC, small-cell lung cancer; TNBC, triple-negative breast cancer; ECOG, Eastern Cooperative Oncology Group; HPLC, high-performance liquid chromatography; IQR, interquartile range.
Appendix A. Figure Files for Submission
Table A1.
Figure numbering, captions, and file names (submission index).
Table A1.
Figure numbering, captions, and file names (submission index).
| Item |
Title (short) |
File name |
Notes |
| Graphical Abstract (optional) |
Overview schematic |
GraphicalAbstract.png |
Optional MDPI graphical abstract |
| Figure 1 |
Study design and participant flow |
Figure1.png |
Flow diagram |
| Figure 2 |
Plasma urolithin A across cohorts |
Figure2.png |
Box-and-whisker plot (log scale) |
| Figure 3 |
Urinary 6-SMT across cohorts |
Figure3.png |
Box-and-whisker plot (ng/day) |
| Figure 4 |
Compartmental schematic of triad |
Figure4.png |
Summary schematic/
hypothesis illustration |
| Figure 5 |
Conceptual model of dysbiosis-UA-melatonin triad |
Figure5.png |
Mechanistic/conceptual diagram |
References
- Garcia-Villalba, R.; Gimenez-Bastida, J.A.; Cortes-Martin, A.; Avila-Galvez, M.A.; Tomas-Barberan, F.A.; Selma, M.V.; Espin, J.C. Urolithins: A Comprehensive Update on Their Metabolism, Bioactivity, and Associated Gut Microbiota. Mol. Nutr. Food Res. 2022, 66, e2101019. [CrossRef]
- Selma, M.V.; Gonzalez-Sarrias, A.; Salas-Salvado, J.; Andres-Lacueva, C.; Alasalvar, C.; Orem, A.; Tomas-Barberan, F.A.; Espin, J.C. The Gut Microbiota Metabolism of Pomegranate or Walnut Ellagitannins Has a Major Impact on Their Bioavailability and Cardiometabolic Effects: Evidence from Human Intervention Studies. Clin. Nutr. 2018, 37, 897-905.
- Iglesias-Aguirre, C.E.; Cortes-Martin, A.; Avila-Galvez, M.A.; Garcia-Villalba, R.; Selma, M.V.; Espin, J.C. Gut Bacteria Involved in Ellagic Acid Metabolism to Yield Human Urolithin Metabotypes Revealed. J. Agric. Food Chem. 2023, 71, 4029-4035. [CrossRef]
- Piwowarski, J.P.; Kiss, A.K.; Granica, S.; Moeslinger, T. Urolithins, Gut Microbiota-Derived Metabolites of Ellagitannins, Inhibit LPS-Induced Inflammation in RAW 264.7 Murine Macrophages. Mol. Nutr. Food Res. 2015, 59, 2168-2177.
- Gimenez-Bastida, J.A.; Gonzalez-Sarrias, A.; Larrosa, M.; Tomas-Barberan, F.A.; Espin, J.C.; Garcia-Conesa, M.T. Ellagitannin Metabolites, Urolithin A Glucuronide and Its Aglycone Urolithin A, Ameliorate TNF-alpha-Induced Inflammation and Associated Molecular Markers in Human Aortic Endothelial Cells. Mol. Nutr. Food Res. 2012, 56, 784-796.
- Chen, X.; Zhang, S.; Zhang, D.; Li, H.; Wang, L. Urolithin B: A Review of Its Metabolism, Biological Activities, and Mechanisms. Front. Pharmacol. 2022, 13, 917266.
- Garay-Mayol, M.A.; Gimenez-Bastida, J.A.; Lopez-Canovas, A.; Poveda-Lopez, S.; Navarro-Orcajada, S.; Brito, J.B.; Alarcon-Fernandez, R.; Trujillo, M. Urolithins Cross the Blood-Brain Barrier and Reduce Stimulus-Dependent Microglial Activation. Food Funct. 2025, 16, 9006-9021.
- An, X.; Lu, S.; Wang, Y. Urolithins: A Prospective Alternative against Brain Aging. Nutrients 2023, 15, 3884.
- Hou, Q.; Chu, X.; Park, J.H. Urolithin A Improves Alzheimer’s Disease Cognition by Restoring Mitophagy and Lysosomal Function. Alzheimer’s Dement. 2024, 20, 4212-4233. [CrossRef]
- Fan, X.; Jin, Y.; Chen, G.; Ma, X.; Zhang, L. Gut Microbiota Dysbiosis Drives the Development of Colorectal Cancer. Digestion 2021, 102, 508-515. [CrossRef]
- Niekamp, J.H.; Kim, J.; Pankratz, N.; Zamani, N.; Zhao, M.; Lu, W.; Shukla, S.; Abbas, A.; Rhee, Y.Y.; Briski, D.; et al. Microbial Metabolite Dysbiosis and Colorectal Cancer. Gut Liver 2023, 17, 190-203.
- Wei, X.; Huang, Y.; Sun, Q.; Fu, L.; Lin, Z.; Wu, C. Impact of Gut Dysbiosis on the Risk of Non-Small-Cell Lung Cancer. Int. J. Environ. Res. Public Health 2022, 19, 15991.
- Tarique, M.; Ali, A.H.; Kizhakkayil, J.; Liu, S.Q.; Oz, F.; Dertli, E.; Kamal-Eldin, A.; Ayyash, M. Exopolysaccharides from Enterococcus faecium and Streptococcus thermophilus: Bioactivities, Gut Microbiome Effects, and Fermented Milk Rheology. Food Chem. X 2024, 21, 101073. [CrossRef]
- Huang, Y.-Y.; Lu, Y.-H.; Liu, X.-T.; Wu, W.-T.; Li, W.-Q.; Lai, S.-Q.; Aadil, R.M.; Rajoka, M.S.R.; Wang, L.-H.; Zeng, X.-A. Metabolic Properties, Functional Characteristics, and Practical Application of Streptococcus thermophilus. Food Rev. Int. 2024, 40, 792-813. [CrossRef]
- Im, E.J.; Lee, H.H.-Y.; Kim, M.; Kim, M.-K. Evaluation of Enterococcal Probiotic Usage and Review of Potential Health Benefits, Safety, and Risk of Antibiotic-Resistant Strain Emergence. Antibiotics 2023, 12, 1327.
- Schernhammer, E.S.; Berrino, F.; Krogh, V.; et al. Urinary 6-Sulfatoxymelatonin Levels and Risk of Breast Cancer in Postmenopausal Women. J. Natl. Cancer Inst. 2008, 100, 898-905. [CrossRef]
- Brown, S.B.; Arcaro, K.F.; Eliassen, A.H.; et al. Urinary Melatonin Concentration and the Risk of Breast Cancer in the Nurses’ Health Study II. Am. J. Epidemiol. 2015, 181, 155-162. [CrossRef]
- Wu, A.H.; Wang, R.; Koh, W.-P.; Stanczyk, F.Z.; Lee, H.-P.; Yu, M.C. Sleep Duration, Melatonin and Breast Cancer among Chinese Women in Singapore. Carcinogenesis 2008, 29, 1244-1248. [CrossRef]
- Mirick, D.K.; Bhatti, P.; Chen, C.; Nordt, F.; Stanczyk, F.Z.; Davis, S. Night Shift Work and Levels of 6-Sulfatoxymelatonin and Cortisol in Men. Cancer Epidemiol. Biomarkers Prev. 2013, 22, 1079-1087. [CrossRef]
- Bartsch, C.; Bartsch, H.; Schmidt, A.; Ilg, S.; Bichler, K.H.; Fluchter, S.H. Urinary Melatonin-Sulfate Excretion in Patients with Benign and Malignant Tumors of the Prostate. J. Urol. 1992, 148, 1165-1170.
- Tai, S.Y.; Huang, S.P.; Bao, B.Y.; Wu, M.T.; Wang, C.L.; Huang, C.Y. Circadian Variation of Melatonin and 6-Sulfatoxymelatonin in Patients with Prostate Cancer. Prostate 2016, 76, 544-556.
- Vaselkiv, J.B.; Cheng, I.; Chowdhury-Paulino, I.M.; et al. Urinary 6-Sulfatoxymelatonin Levels and Prostate Cancer Risk among Men in the Multiethnic Cohort. Cancer Epidemiol. Biomarkers Prev. 2022, 31, 688-691. [CrossRef]
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).