
Submitted:
01 December 2025
Posted:
02 December 2025
You are already at the latest version
Abstract
The development of beekeeping in Ecuador has generated the need to strengthen the bee health program. Research on the main pathogens responsible for diseases like nosemosis, which can severely impact bee health, is of special interest. This study aims to detect and identify the Nosema apis and / or Nosema ceranae species affecting honeybee colonies located in the northern Sierra region of Ecuador through the amplification of RPB1 gene fragments by multiplex PCR and the phylogenetic analysis of N. ceranae based on the 16 S RNAr gene. Of the 164 honeybee samples collected from colonies located in the provinces of Carchi, Imbabura and Pichincha, N. apis was detected in 14.63% (24/164), and N. ceranae in 21.34% (35/164). Phylogenetic analysis showed that N. ceranae from Ecuador is closely related to the sequences from Argentina and Brazil. This study is the first in the country to report the presence of these two microsporidia species, being valuable for improving diagnostic capabilities.
Keywords:
Nosemosis
; Nosema
; N. ceranae
; N. apis
; RPB1gen and 16S RNAr gen
1. Introduction
Beekeeping activity in Ecuador has been growing steadily. In 2016, a total of 902 apiaries and 12188 colonies of domestic were registered, with most of them concentrated in the Sierra region (mountain area); where the provinces of Pichincha (22.79%), Imbabura (8.41%) and Carchi (7.99%) have the highest numbers of colonies [1]. Given the increase in this activity in the country, the Agencia de Regulación y Control Fito y Zoosanitario de Ecuador (AGROCALIDAD) aimed to get information regarding the health status of those colonies. They conducted a nationwide study of the main pathogens affecting honey bee colonies and reported the presence of Nosema sp. in 235 of the sampled apiaries. However, the species was not identified [1].
Microsporidia of the genus Nosema are intracellular parasites [2] with more than 150 described species [3] affecting both mammals [4] and insects [5] especially of the order Hymenoptera and Lepidoptera [6]. Nosemosis is a disease caused by the microsporidia Nosema apis [7] and/or Nosema ceranae [8,9,10] which has a worldwide distribution [11,12,13,14,15]. Nosema apis and Nosema ceranae have recently been reclassified as Vairomorpha [16] although its reclassification is still under discussion [17]. For this reason, several authors still retain the previous nomenclature of Nosema [18,19,20], while others have already adopted the new name Vairomorpha [21,22,23,24]. In this study, we will use the both name Nosema and Vairomorpha.
Within the Apidae family, the first report of infection was reported in 1996 by Nosema ceranae in Apis cerana [10]. As for Apis mellifera, the first report of N. ceranae as a new pathogen was reported in 2006 [8,25].
In the South American region, several countries such as Brazil [26], Argentina [27], Chile [28] or Uruguay [29] have reported the presence of these parasites in their colonies [30]. Within the Caribbean, Dominica Island reports the presence of both species [31], and to the north of the American continent the presence of this microsporidium was reported in Mexico [32], USA and Canada [33]. In addition, N. ceranae has been found in bumblebees of the genus Bombus [34,35] and in native stingless bees [36].
N. apis and N. ceranae can enter the honey bees by ingestion of spores with the food or by cleaning the alveoli and the inside of the hives [37,38,39,40]. The spores invade the cells of the intestinal wall of bees and multiply [41,42,43,44]. Traces of spores have also been found in other parts of the body, such as hypopharyngeal gland tissue, salivary glands, Malpighian tubules and fat body [45]. Infection by either or both Nosema species, can cause alterations in behavior [46], metabolism and nutrition [47], decreased bee life expectancy and mortality [48], as well as decreased honey production [49].
Reported prevalence levels of nosemosis varied depending on the country, the season and the species of the causal agent. For example, in Canada, prevalence of N. apis can range between 4-34% [50], in México 80-90% while N. ceranae was about 12-13% in Mexico [51], 58% in Saudi Arabia [52] or up to 66% in Romania [53]. Co-infections have also been recorded in hives from Turkey, the study carried out since 2009 to 2016, in different seasons, reports that the prevalence varies from 20-49% in samples [54].
Among the methods developed for Nosema diagnosis described in the OIE Terrestrial Manual [39] and in Standard methods for Nosema research [55] are microscopic observation of spores, fluorescence microscopy and Polymerase Chain Reaction (PCR), the latter being the most employed because under the light microscope it is difficult to differentiate morphologically these two Nosema species. Primers based on the 16S rRNA gene described by [56] are widely used [14,57,58]. However, authors such as Gisder and Genersch [59] have chosen to develop or use primers based on the large subunit of the RNA polymerase II gene (RPB1) to differentiate Nosema species [59,60,61], which is also used to observe population diversity [62] and genetic diversity [63] in N. apis and N. ceranae.
The aim of this study was to detect and differentiate the type of microsporidium that causes nosemosis in domestic colonies of honey bees in the northern highlands of Ecuador, using a multiplex PCR based on the RPB1 gene. In addition, we present the first molecular characterization of Nosema ceranae in apiaries in Ecuador based on the 16S rRNA gene.
2. Materials and Methods
2.1. Sample Collection
Based on the data obtained from the first beekeeping census carried out by AGROCALIDAD (2016) [1], the study area focused on the provinces of Pichincha and Imbabura given the greater concentration of apiaries (a) and hives (h) in the northern part of the Ecuadorian Sierra, and Carchi because it is the border province with Colombia.
Between the months of April to June 2017, samples of foragers honey bees were collected from the hive entrance (h=164), located in apiaries (a=29) in the three intervened provinces (Table 1). The location of the apiaries was recorded with the help of a Garmin 60Cs GPS and the EpiCollect 5 application.
2.2. Diagnostic Tests
For the diagnosis of Nosema sp. in honey bees, light microscopy and PCR laboratory tests were used. Each of the 164 samples was individually analyzed with both techniques. In addition, the florescence microscopy test was used on one of the samples diagnosed as co-infected by PCR to observe and compare the size of N. apis and N. ceranae spores.
2.3. Optical Microcopy Test and Determination of Spore Number
Abdomen from approximately 20 honey bees per colony were aseptically separated with forceps and scalpel, mixed with 1 mL of distilled water, macerated and placed in vials. An aliquot of the sample (10 µL) was placed on a Neubauer chamber and visualized with an optical microscope at 400X magnification.
The spore concentration was obtained by multiplying the average number of spores in the sample by the dilution factor and dividing by the product of the chamber area (mm) by the chamber depth (mm). The level of bee infestation was then classified according to the following scale: low (<1,000,000 spores/bee), medium (>1,000,000 <2,000,000 spores/bee) and high (more than 2,000,000) [64].
2.4. DNA Extraction of Nosema sp. in Honey Bees
The protocol used for DNA extraction was the described by Hamiduzzaman et al., (2010) [65] with modifications. The abdomens of 20 honey bees from each colony were placed in 2 mL vials. 500 µL of extraction buffer (0.03 M CTAB, 0.05 M Tris, 0.01 M EDTA, 1.1 M NaCl, pH 8.0), and 4 µL of Proteinase K (20 mg/ml) were added. Samples were triturated with a sterile pistil, vortexed, and incubated at 60 °C for 3 h with constant shaking, occasionally inverting the tubes during incubation. They were then centrifuged for 1 min at 14000 rpm, and the supernatant was transferred to a 1.5 mL vial. A double extraction with phenol-chloroform (1:1) was performed by adding 300 µL of this mixture, homogenizing the tubes by inversion and centrifuging them at 14000 rpm for 15 min; the supernatant was transferred to a new vial. Then 300 µL of chloroform was added and centrifuged at 10000 rpm for 5 min. 30 µL of 3 M sodium acetate and 600 µL of 95 % ethanol were added to the supernatant, mixed gently and stored at -20 °C overnight. The samples were centrifuged at 10000 rpm for 10 min and the ethanol was discarded. Subsequently, 1 mL of 75% ethanol (4 °C) was added and mixed briefly by vortexing. The pellet was then centrifuged for 3 min at 14000 rpm, the ethanol was discarded, and the pellet was allowed to dry. Finally, the DNA pellet was re-suspended in 100 µL of ultrapure distilled water free of nucleases, and the samples were incubated in a water bath at 65 °C for 10 min. Samples were incubated with RNAse at 37 °C for 10 min. The extracted DNA was stored at -20 °C until use.
2.5. Detection and Identification of Nosema sp. by Multiplex PCR
Obtained DNA was subjected to multiplex PCR, using two pairs of primers (Table 2), that amplify different regions of the RPB1 gene, depending on the Nosema species. The multiplex PCR assay consisted of adding 1X of PCR buffer, 0.5 µM of primers NosaRNAPol-F2 / NosaRNAPol-R2, for the detection of N. apis; 0.4 µM of primers NoscRNAPol-F2 / NoscRNAPol-R2 for N. ceranae; 1.75 mM MgCl2, 0.8 mM dNTP mix (0.2 mM/dNTP), 1.25 U/µL Taq polymerase enzyme (Invitrogen), 400 ng DNA, plus a volume of UltraPure ™ DNase/RNase-Free Distilled Water to complete 25 µL of reaction. Cycling conditions in the thermal cycler were 95 °C initial denaturation for 5 minutes; 40 one-minute cycles of denaturation steps at 94 °C, annealing primer 67 °C and extension at 72 °C, and a final extension cycle at 72 °C for 10 minutes. Additionally, a single PCR assay (Table 2) was performed on N. ceranae positive samples according to the results of the multiplex PCR. A 218 bp fragment of the 16S rRNA gene was amplified, following the protocol described by Higes et al. (2006) [8].
2.6. Sequencing, Molecular Characterization and Phylogenetic Analysis
After molecular detection of Nosema species by multiplex PCR, phylogenetic analysis of N. ceranae, was performed, as the species of greatest interest, based on the 16S rRNA gene primers (Tabla 2). Only those N. ceranae positive samples with a strong band intensity were chosen, and the products of the single PCR assay were sent for sequencing, in duplicate and in both directions by the Sanger method to Macrogen® Korea. Consensus sequences (n = 9) from Ecuador were compared with sequences of isolates available in GenBank.
A phylogenetic tree was constructed to determine the phylogenetic relationship between N. ceranae isolates from Ecuador and sequences belonging to Americas, Europe and Asia, as well as to observe the relationship between the sequences from this study and other sequences within the Nosema genus. The tree was constructed in MEGA 12 program using the consensus sequence from this study and the sequences available in GenBank. The maximum parsimony method, which uses the subtree-pruning-regrafting (SPR) algorithm), was employed for the analysis. This analysis involved 24 nucleotide sequences. There was a total of 222 positions in the final dataset. Trachipleistophora hominis was used as the outgroup.
2.7. Detection of Spores by Fluorescence Microscopy
Sample A27C2 was subjected to complementary analysis by fluorescence microscopy according to the modified procedure of Snow(2016) [66]. Smear samples of bee macerates were incubated for 2 h at room temperature in 3% glutaraldehyde (60 μL). To remove the fixative, two washes were performed, for 10 min, with 1 mL of PBS-T solution (PBS + 0.01 %Triton X-100). 500 μL of fluorescent FB28 (Calcofluor White M2R) was added. After overnight incubation at 4 °C in a humid chamber, the plates were washed twice with PBS-T solution. Next, as a co-stain, 200 μL of Hoesch DNA dye (1:2000) was added and incubated for 5 min at 4 °C in humid chamber. The plates were washed twice with PBS-T solution. Then, the plates were dried at room temperature in the dark, observation was performed on an Olympus IX53 microscope at 384.6 ms exposure in 20X lens.
3. Results
3.1. Detection of Nosema sp. Spores by Optical Microscopy
The oval shape of Nosema sp. spores was generally observed by light microscopy without identification of the species (Figure 1). The prevalence of Nosema sp. was 41.38 % (12/29) at the apiary level and 17.07 % (28/164) at the colony level.
The province of Pichincha (h = 63) had the highest number of positive samples (20/63), followed by the province of Carchi (5/33) and finally Imbabura (3/68) (Table 3). On the other hand, low (h = 9), medium (h = 2) and high (h = 17) levels of infestation or spore intensity were observed.
3.2. Identification of Nosema apis and Nosema ceranae by PCR
By multiplex PCR we detected Nosema sp. infection in 34.76% (59/164) of colonies and 86.21% (25/29) of apiaries. We determined the presence of N. apis and N. ceranae in the colonies of the three provinces with a prevalence of 14.63% (24/164) and 21.34% (35/164) respectively, finding also apiaries (a = 5) and colonies (h = 2) with double infections.
Among the three provinces, Pichincha had the highest prevalence of both N. apis (24.24 %) and N. ceranae (36.50 %) at the colony level. While in Imbabura and Carchi there was a higher prevalence of N. apis (14.70 % and 12.69 % respectively) than N. ceranae (13.23 % and 9.09 % respectively) in the colonies (Table 3). Table 3 gives details of the distribution of results (number, prevalence, and 95 % confidence intervals) for light microscopy and PCR tests, at the apiary, colony and provincial level.
3.3. Molecular Characterization and Phylogenetic Analysis of N. ceranae
BLAST analysis of the fragment sequence was 99.5-100 % identity with partial sequences of small subunit ribosomal RNA gene isolates. Nine sequences of N. ceranae (n = 9) were obtained from various sectors of the provinces of Imbabura (n = 3) and Pichincha (n = 6), accession numbers: PQ336918, PQ336919, PQ336920, PQ336921, PQ336922, PQ336923, PQ336924, PQ336925, PQ336926. Since all of them showed 100 % homology, only one sequence was used in the phylogenetic tree (PQ336918).
The phylogenetic analysis involved nine species of microsporidia belonging to this genus (Figure 2). We observed that the isolates from Ecuador are located within the same clade of N. ceranae, confirming that they belong to this species. Furthermore, the closest species are N. bombi and other Nosema species, leaving N. apis distantly related to N. ceranae.
Within of the N. ceranae clade (Figure 2), we don’t observe a grouped regionalization by continent. Thus, sequences from American continent are distributed among all N. ceranae subclades. Consequently, the isolate from Ecuador is placed at the same phylogenetic distance as isolates from Argentina and Brazil (South America), Saudi Arabia and Iran (Asia), and Spain and Lithuania (Europe).
3.4. Detection of Nosema sp. by Fluorescence Microscopy
Through fluorescence microscopy (Figure 3), the oval forms of Nosema sp. spores were visualized, being between 4 μm and 6 μm long, stained with the Hoesch dye which can stain the DNA of the cells and additionally it was possible to take the approximate measurements of them. The red staining of the spores indicates a positive result to the FB28 dye or calcofluor which is specific for chitin, polysaccharide component of the fungal cell wall. This specific staining allows us to confirm definitively that they are Nosema sp. microsporidia.
4. Discussion
This study revealed the presence of N. apis (14.63 %) and N. ceranae (21.34 %) in colonies from Ecuadorian territory. The prevalence of both microsporidia species results higher than reported by the regulatory agency (9 % / 235 apiaries sampled) in the 24 provinces of Ecuador [1]. Visualization of spores by light microscopy technique is relatively straightforward, experts may be able to discern between spores [67], with fluorescence microscopy it has been shown that calcofluor, specifically binds to chitin in the cell wall of mature spores [66]. Immature spores lack chitin, this method is useful for studies of mature spore localization within cells, but cannot differentiate between species [66].
Although the light microscopy technique is used as an initial screening for spore detection, diagnosis and differentiation of Nosema species has been carried out based on molecular methods such as PCR and its variants [68,69], because of its sensitivity and specificity [55,70], which is clearly observed in this research. Thus, it is proposed that this type of technique should be incorporated into regulatory agencies for greater accuracy in distribution and prevalence studies.
Although the presence of Nosema spp. has been previously reported in Ecuador [71,72], as a thesis of universities in the country, these studies were carried out using light microscopy, so the species of Nosema are not differentiated. Therefore, the present study is the first report of the distribution of N. apis and N. ceranae in the northern part of Ecuador with molecular characterization. Hence, both pathogens are present in the provinces studied. Furthermore, the prevalence of N. ceranae determined here is in addition to reports where higher prevalence than N. apis are expressed [45,49,51,73,74].
The detection of Nosema spp. in bees using calcofluor white and Hoechst staining proved highly effective, allowing for the identification of fluorescent spores measuring 4-6 µm. Calcofluor specifically stained the chitin in the spore wall, while Hoechst confirmed the presence of DNA in the nucleus, facilitating spore differentiation. These fluorescent stains, sensitive and rapid, outperform traditional methods such as phase-contrast microscopy, making them valuable for diagnosis and epidemiological monitoring [66,75].
The maximum parsimony using the 16S rRNA gene determined that there is no difference between geographically distant isolates within the South American continent. However, they are also related to sequences from Iran (Asia) and Lithuania (Europe). The finding agrees with those of [76] where they found identical haplotypes from samples originating from Spain, Slovenia and Kyrgyzstan. Molecular phylogeny using the small subunit revealed that N. apis and N. ceranae, pathogens of the same host, are highly divergent, and are not very close relatives within the genus Nosema [38,45,62].
The fact that N. apis and N. ceranae microsporidia are also present in hives in Ecuador contributes to a better understanding of the dynamics of diseases affecting the bee population. This will allow the implementation of specific preventive measures, adapted to the country's context, to control their spread.
5. Conclusions
This is the first report of N. ceranae in Ecuador. The prevalence found and the co-infections with N. apis, suggest a wide census of Nosema species throughout Ecuador. The detection of the two Nosema species in the apiaries and in the colonies shows the importance of contagion within the apiaries. Future studies using haplotype networks could determine if there are genetic variants infections between apiaries.
This study represents the first report of Nosema ceranae in Ecuador, highlighting its presence alongside Nosema apis and suggesting a broad distribution of Nosema species across the country. The observed co-infections underscore the potential for significant pathogen exchange within and between apiaries, emphasizing the need for improved biosecurity measures. The detection of both Nosema species within colonies also raises concerns about the potential cumulative effects on colony health and productivity.
Future research should prioritize molecular studies, such as haplotype network analyses, to explore the genetic diversity of Nosema populations and identify transmission pathways. Additionally, longitudinal studies on colony dynamics could clarify the impact of co-infections on bee health and survival. Implementing large-scale monitoring programs in Ecuador and globally will be essential to assess the prevalence and spread of Nosema species, enabling the development of targeted control strategies. These efforts could significantly contribute to safeguarding apiculture and ensuring pollination sustainability worldwide.
Author Contributions
Conceptualization, J.R. and C.S.; methodology, D.S., L.F., M.T. and C.CH.; software, D.S. and C.CH; validation, J.R., C.S., S.M., A.R., C.CH and K.A.; formal analysis, J.R and C.S.; investigation, J.R. and D.S.; resources, J.R.; data curation, D.S., J.R. and C.CH;. writing—original draft preparation, D.S.; writing—review and editing, J.R., C.S, S.M., M.T, C.CH and K.A.; visualization, J.R., C.S. and S.M.; supervision, J.R.; project administration, J.R.; funding acquisition, J.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
Universidad de las Fuerzas Armadas ESPE
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| GISAH | Grupo de Investigación en Sanidad Animal y Humana |
| RPB1 | RNA Polymerase II Subunit RPB1 |
| 16 S RNAr | 16S ribosomal RNA |
| PCR | Polymerase Chain Reaction |
| N. apis | Nosema apis |
| AGROCALIDAD | Agencia de Regulación y Control Fito y Zoosanitario (Ecuador) |
| N. ceranae | Nosema ceranae |
| OIE | World Organization for Animal Health |
| GPS | Global Positioning System |
| DNA | Deoxyribonucleic Acid |
| EDTA | Ethylenediaminetetraacetic Acid |
| RNAse | Ribonuclease |
| dNTP | Deoxyribonucleoside Triphosphate |
| RNA | Ribonucleic Acid |
| USA | United States of America |
References
- AGROCALIDAD Programa Nacional Sanitario Apícola. 2016, 60–60.
- Vávra, J.; Ronny Larsson, J.I. Structure of Microsporidia. In Microsporidia: Pathogens of Opportunity; Weiss, L.M., Becnel, J.J., Eds.; Wiley Blackwell, 2014; pp. 729–729.
- Sprague, V. Characterization and Composition of the Genus Nosema. In Selected Topics on the genus Nosema (Microsporida); Brooks, W.M., Ed.; 1978; Vol. 11, pp. I6–I6.
- Wasson, K.; Peper, R.L. Mammalian Microsporidiosis; Vet Pathol; 2000; Vol. 37, pp. 113–128;
- Becnel, J.J.; Andreadis, T.G. Microsporidia in Insects. In Microsporidia: Pathogens of Opportunity; Weiss, L.M., Becnel, J.J., Eds.; Wiley Blackwell, 2014; pp. 521–570 ISBN 978-1-118-39526-4.
- Wittner, M.; Weiss, L.M. The Microsporidia and Microsporidiosis; ASM press: Washington, DC, 1999; p. 553;
- Zander, E. Tierische Parasiten Als Krankheitserreger Bei Der Biene. Münch. Bienenztg. 1909, 196–204.
- Higes, M.; Martín, R.; Meana, A. Nosema ceranae, a New Microsporidian Parasite in Honeybees in Europe. J. Invertebr. Pathol. 2006, 92, 93–95. [Google Scholar] [CrossRef]
- Klee, J.; Besana, A.M.; Genersch, E.; Gisder, S.; Nanetti, A.; Tam, D.Q.; Chinh, T.X.; Puerta, F.; Ruz, J.M.; Kryger, P.; et al. Widespread Dispersal of the Microsporidian Nosema Ceranae, an Emergent Pathogen of the Western Honey Bee, Apis Mellifera. J. Invertebr. Pathol. 2007, 96, 1–10. [Google Scholar] [CrossRef]
- Fries, I.; Feng, F.; da Silva, A.; Slemenda, S.B.; Pieniazek, N.J. Nosema Ceranae n. sp. (Microspora, Nosematidae), Morphological and Molecular Characterization of a Microsporidian Parasite of the Asian Honey Bee Apis cerana (Hymenoptera, Apidae). Eur. J. Protistol. 1996, 32, 356–365. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee Declines Driven by Combined Stress from Parasites, Pesticides, and Lack of Flowers. Science 2015, 347. [Google Scholar] [CrossRef] [PubMed]
- Hristov, P.; Georgieva, A.A.; Radoslavov, G.; Sirakova, D.; Dzhebir, G.; Shumkova, R.; Neov, B.B.; Bouga, M.; Author, C.; Hristov, P.; et al. The First Report of the Prevalence of Nosema ceranae in Bulgaria. PeerJ 2018.
- Martín-Hernández, R.; Bartolomé, C.; Chejanovsky, N.; Le Conte, Y.; Dalmon, A.; Dussaubat, C.; García-Palencia, P.; Meana, A.; Pinto, M.A.; Soroker, V.; et al. Nosema ceranae in Apis mellifera: A 12 Years Postdetection Perspective. Environ. Microbiol. 2018, 20, 1302–1329. [Google Scholar] [CrossRef] [PubMed]
- Papini, R.; Mancianti, F.; Canovai, R.; Cosci, F.; Rocchigiani, G.; Benelli, G.; Canale, A. Prevalence of the Microsporidian Nosema ceranae in Honeybee (Apis mellifera) Apiaries in Central Italy. Saudi J. Biol. Sci. 2017, 24, 979–982. [Google Scholar] [CrossRef]
- Vavilova, V.Y.; Konopatskaia, I.; Luzyanin, S.L.; Woyciechowski, M.; Blinov, A.G. Parasites of the Genus Nosema, Crithidia and Lotmaria in the Honeybee and Bumblebee Populations: A Case Study in India. Vavilov J. Genet. Breed. 2017, 21, 943–951. [Google Scholar] [CrossRef]
- Tokarev, Y.S.; Huang, W.-F.; Solter, L.F.; Malysh, J.M.; Becnel, J.J.; Vossbrinck, C.R. A Formal Redefinition of the Genera Nosema and Vairimorpha (Microsporidia: Nosematidae) and Reassignment of Species Based on Molecular Phylogenetics. J. Invertebr. Pathol. 2020, 169, 107279–107279. [Google Scholar] [CrossRef]
- Bartolomé, C.; Higes, M.; Hernández, R.M.; Chen, Y.P.; Evans, J.D.; Huang, Q. The Recent Revision of the Genera Nosema and Vairimorpha (Microsporidia: Nosematidae) Was Flawed and Misleads the Bee Scientific Community. J. Invertebr. Pathol. 2024, 206, 108146–108146. [Google Scholar] [CrossRef]
- Benali, K.; Nora, C.-A.; Mohamed, C.; Hakim, T.; Khaoula, B.; Semir, G.; Suheil, B. First Molecular Detection and Geographical Distribution of Nosema apis & Nosema ceranae in Indigenous Honey Bees Reared in Algeria. Genet. Biodivers. J. 2023, 7, 141–150. [Google Scholar] [CrossRef]
- Lopes, A.R.; Martín-Hernández, R.; Higes, M.; Segura, S.K.; Henriques, D.; Pinto, M.A. First Detection of Nosema ceranae in Honey Bees (Apis mellifera L.) of the Macaronesian Archipelago of Madeira. J. Apic. Res. 2023, 62, 514–517. [Google Scholar] [CrossRef]
- Valizadeh, P.; Guzman-Novoa, E.; Goodwin, P.H. High Genetic Variability of Nosema ceranae Populations in Apis mellifera from East Asia Compared to Central Asia and the Americas. Biol. Invasions 2022, 24, 3133–3145. [Google Scholar] [CrossRef]
- Imani Baran, A.; Kalami, H.; Mazaheri, J.; Hamidian, G. Vairimorpha ceranae Was the Only Detected Microsporidian Species from Iranian Honey Bee Colonies: A Molecular and Phylogenetic Study. Parasitol. Res. 2022, 121, 355–366. [Google Scholar] [CrossRef]
- Pekagirbas, M.; Hacilarlioglu, S.; Kanlioglu, H.; Aydin, H.B.; Koc, B.; Bilgic, H.B.; Karagenc, T.; Bakirci, S. Dominancy of Vairimorpha ceranae: Microscopic and Molecular Detection in Aydın, West Turkey. Comp. Parasitol. 2025, 92. [Google Scholar] [CrossRef]
- Blot, N.; Clémencet, J.; Jourda, C.; Lefeuvre, P.; Warrit, N.; Esnault, O.; Delatte, H. Geographic Population Structure of the Honeybee Microsporidian Parasite Vairimorpha (Nosema) ceranae in the South West Indian Ocean. Sci. Rep. 2023, 13, 12122–12122. [Google Scholar] [CrossRef]
- Manjy, M.S.; Shaher, K.W. Molecular Detection of Vairimorpha ceranae and Determining the Incidence Rate of Honey Bee Hives and Workers of Apis mellifera L. IOP Conf. Ser. Earth Environ. Sci. 2025, 1449, 012054–012054. [Google Scholar] [CrossRef]
- Huang, W.-F.; Jiang, J.-H.; Chen, Y.-W.; Wang, C.-H. A Nosema ceranae Isolate from the Honeybee Apis mellifera. Apidologie 2007, 38, 30–37. [Google Scholar] [CrossRef]
- Teixeira, E.W.; dos Santos, L.G.; Sattler, A.; Message, D.M.; Alves, M.L.T.M.F.; Martins, M.F.; Grassi-Sella, M.L.; Francoy, T.M. Nosema ceranae Has Been Present in Brazil for More than Three Decades Infecting Africanized Honey Bees. J. Invertebr. Pathol. 2013, 114, 250–254. [Google Scholar] [CrossRef]
- Medici, S.K.; Sarlo, E.G.; Porrini, M.P.; Braunstein, M.; Eguaras, M.J. Genetic Variation and Widespread Dispersal of Nosema ceranae in Apis mellifera Apiaries from Argentina. Parasitol. Res. 2012, 110, 859–864. [Google Scholar] [CrossRef]
- Servicio Agrícola Ganadero (SAG) Chile Informe de Caso de Nosema ceranae En La Región Del Bío Bío, Octubre de 2009; 2010.
- Invernizzi, C.; Abud, C.; Tomasco, I.H.; Harriet, J.; Ramallo, G.; Campá, J.; Katz, H.; Gardiol, G.; Mendoza, Y. Presence of Nosema ceranae in Honeybees (Apis Mellifera) in Uruguay. J. Invertebr. Pathol. 2009, 101, 150–153. [Google Scholar] [CrossRef]
- Maggi, M.; Antúnez, K.; Ivernizzi, C.; Aldea, P.; Vargas, M.; Negri, P.; Brasero, C.; De Jong, D.; Message Dejair; Weinstein Teixeira, E. Honeybee Health in South America. Springer Verl. 2016, 835–854. [Google Scholar] [CrossRef]
- Rangel, J.; Gonzalez, A.; Stoner, M.; Hatter, A.; Traver, B.E. Genetic Diversity and Prevalence of Varroa destructor, Nosema apis, and N. ceranae in Managed Honey Bee (Apis Mellifera) Colonies in the Caribbean Island of Dominica, West Indies. J. Apic. Res. 2018, 57, 541–550. [Google Scholar] [CrossRef]
- Guerrero-Molina, C.; Correa-Benítez, A.; Hamiduzzaman, M.; Guzman-Novoa, E. Nosema ceranae Is an Old Resident of Honey Bee (Apis mellifera) Colonies in Mexico, Causing Infection Levels of One Million Spores per Bee or Higher during Summer and Fall. J. Invertebr. Pathol. 2016, 141, 38–40. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.R.; Shafer, A.B.A.; Rogers, R.E.L.; Shutler, D.; Stewart, D.T. First Detection of Nosema ceranae, a Microsporidian Parasite of European Honey Bees (Apis mellifera), in Canada and Central USA. J. Invertebr. Pathol. 2008, 97, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Plischuk, S.; Martín-Hernández, R.; Prieto, L.; Lucía, M.; Botías, C.; Meana, A.; Abrahamovich, A.H.; Lange, C.; Higes, M. South American Native Bumblebees (Hymenoptera: Apidae) Infected by Nosema ceranae (Microsporidia), an Emerging Pathogen of Honeybees (Apis mellifera). Environ. Microbiol. Rep. 2009, 1, 131–135. [Google Scholar] [CrossRef]
- Arbulo, N.; Antúnez, K.; Salvarrey, S.; Santos, E.; Branchiccela, B.; Martín-Hernández, R.; Higes, M.; Invernizzi, C. High Prevalence and Infection Levels of Nosema ceranae in Bumblebees Bombus atratus and Bombus bellicosus from Uruguay. J. Invertebr. Pathol. 2015, 130, 165–168. [Google Scholar] [CrossRef]
- Porrini, M.P.; Porrini, L.P.; Garrido, P.M.; de Melo e Silva Neto, C.; Porrini, D.P.; Muller, F.; Nuñez, L.A.; Alvarez, L.; Iriarte, P.F.; Eguaras, M.J. Nosema ceranae in South American Native Stingless Bees and Social Wasp. Microb. Ecol. 2017, 74, 761–764. [Google Scholar] [CrossRef]
- Fries, I. Nosema apis—a Parasite in the Honey Bee Colony. Bee World 1993, 74, 5–19. [Google Scholar] [CrossRef]
- Fries, I. Nosema ceranae in European Honey Bees (Apis mellifera). J. Invertebr. Pathol. 2010, 103, S73–S79. [Google Scholar] [CrossRef]
- OIE Nosemosis de Las Abejas Melíferas. In Manual Terrestres de la OIE; 2018.
- Smith, M.L. The Honey Bee Parasite Nosema ceranae: Transmissible via Food Exchange? PLoS ONE 2012, 7, e43319–e43319. [Google Scholar] [CrossRef]
- Bailey, L. The Infection of the Ventriculus of the Adult Honeybee by Nosema apis (Zander). Parasitology 1955, 45, 86–94. [Google Scholar] [CrossRef]
- Dussaubat, C.; Brunet, J.-L.; Higes, M.; Colbourne, J.K.; Lopez, J.; Choi, J.-H.; Martín-Hernández, R.; Botías, C.; Cousin, M.; McDonnell, C.; et al. Gut Pathology and Responses to the Microsporidium Nosema ceranae in the Honey Bee Apis mellifera. PloS One 2012, 7, e37017–e37017. [Google Scholar] [CrossRef]
- Huang, W.-F.; Solter, L.F.; Yau, P.M.; Imai, B.S. Nosema ceranae Escapes Fumagillin Control in Honey Bees. PLoS Pathog. 2013, 9, e1003185–e1003185. [Google Scholar] [CrossRef]
- Higes, M.; García-Palencia, P.; Urbieta, A.; Nanetti, A.; Martín-Hernández, R. Nosema apis and Nosema ceranae Tissue Tropism in Worker Honey Bees (Apis mellifera). Vet. Pathol. 2020, 57, 132–138. [Google Scholar] [CrossRef]
- Chen, Y.P.; Evans, J.D.; Murphy, C.; Gutell, R.; Zuker, M.; Gundensen-Rindal, D.; Pettis, J.S. Morphological, Molecular, and Phylogenetic Characterization of Nosema ceranae, a Microsporidian Parasite Isolated from the European Honey Bee, Apis mellifera. J. Eukaryot. Microbiol. 2009, 56, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Gage, S.L.; Kramer, C.; Calle, S.; Carroll, M.; Heien, M.; DeGrandi-Hoffman, G. Nosema ceranae Parasitism Impacts Olfactory Learning and Memory and Neurochemistry in Honey Bees (Apis mellifera). J. Exp. Biol. 2018, 221. [Google Scholar] [CrossRef] [PubMed]
- Holt, H.L.; Aronstein, K.A.; Grozinger, C.M. Chronic Parasitization by Nosema Microsporidia Causes Global Expression Changes in Core Nutritional, Metabolic and Behavioral Pathways in Honey Bee Workers (Apis mellifera). BMC Genomics 2013, 14, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.R.; Shutler, D.; Burgher-MacLellan, K.L.; Rogers, R.E.L. Infra-Population and -Community Dynamics of the Parasites Nosema apis and Nosema ceranae, and Consequences for Honey Bee (Apis mellifera) Hosts. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Botías, C.; Martin-Hernández, R.; Barrios, L.; Meana, A.; Higes, M. Nosema spp. Infection and Its Negative Effects on Honey Bees (Apis mellifera iberiensis) at the Colony Level. Vet. Res. 2013, 44, 25–25. [Google Scholar] [CrossRef]
- Emsen, B.; Guzman-Novoa, E.; Hamiduzzaman, M.M.; Eccles, L.; Lacey, B.; Ruiz-P??rez, R.A.; Nasr, M. Higher Prevalence and Levels of Nosema ceranae than Nosema apis Infections in Canadian Honey Bee Colonies. Parasitol. Res. 2016, 115, 175–181. [Google Scholar] [CrossRef]
- González, S.A.C.; Valencia, G.L.; Cabrera, C.O.; Gómez Gómez, S.D.; Torres, K.M.; Blandón, K.O.E.; Guerrero Velázquez, J.G.; Paz, L.E.S.; Trasviña Muñoz, E.; Monge Navarro, F.J. Prevalence and Geographical Distribution of Nosema apis and Nosema ceranae in Apiaries of Northwest Mexico Using a Duplex Real-Time PCR with Melting-Curve Analysis. J. Apic. Res. 2019, 59, 195–203. [Google Scholar] [CrossRef]
- Ansari, M.J.; Al-Ghamdi, A.; Nuru, A.; Khan, K.A.; Alattal, Y. Geographical Distribution and Molecular Detection of Nosema ceranae from Indigenous Honey Bees of Saudi Arabia. Saudi J. Biol. Sci. 2017, 24, 983–991. [Google Scholar] [CrossRef]
- Mederle, N.; Lobo, M.; Morariu, S.; Morariu, F.; Darabus, G.; Mederle, O.A.; Matos, O. Microscopic and Molecular Detection of Nosema ceranae in Honeybee Apis mellifera L. from Romania:: Status on Pathogen Worldwide Distribution. Rev. Chim. 2018, 69, 3761–3772. [Google Scholar] [CrossRef]
- Özkırım, A.; Schiesser, A.; Keskin, N. Dynamics of Nosema apis and Nosema ceranae Co-Infection Seasonally in Honey Bee (Apis Mellifera L.) Colonies. J. Apic. Sci. 2019, 63, 41–48. [Google Scholar] [CrossRef]
- Fries, I.; Chauzat, M.-P.; Chen, Y.-P.; Doublet, V.; Genersch, E.; Gisder, S.; Higes, M.; McMahon, D.P.; Martín-Hernández, R.; Natsopoulou, M.; et al. Standard Methods for Nosema Research. J. Apic. Res. 2013, 52, 1–28. [Google Scholar] [CrossRef]
- Martín-Hernández, R.; Meana, A.; Prieto, L.; Salvador, A.M.; Garrido-Bailón, E.; Higes, M. Outcome of Colonization of Apis Mellifera by Nosema ceranae. Appl. Environ. Microbiol. 2007, 73, 6331–6338. [Google Scholar] [CrossRef]
- Aroee, F.; Azizi, H.; Shiran, B.; Pirali Kheirabadi, K. Molecular Identification of Nosema Species in Provinces of Fars, Chaharmahal and Bakhtiari and Isfahan (Southwestern Iran). Asian Pac. J. Trop. Biomed. 2017, 7, 10–13. [Google Scholar] [CrossRef]
- Botias, C.; Martin-Hernandez, R.; Garrido-Bailon, E.; Anderson, D.; Higes, M. Nosema ceranae Isolate NOS034 16S Ribosomal RNA Gene, Partial Sequence - Nucleotide - NCBI. 2009.
- Gisder, S.; Genersch, E. Molecular Differentiation of Nosema apis and Nosema ceranae Based on Species-Specific Sequence Differences in a Protein Coding Gene. J. Invertebr. Pathol. 2013, 113, 1–6. [Google Scholar] [CrossRef]
- Bacela-Spychalska, K.; Wattier, R.; Teixeira, M.; Cordaux, R.; Quiles, A.; Grabowski, M.; Wroblewski, P.; Ovcharenko, M.; Grabner, D.; Weber, D.; et al. Widespread Infection, Diversification and Old Host Associations of Nosema Microsporidia in European Freshwater Gammarids (Amphipoda). PLOS Pathog. 2023, 19, e1011560–e1011560. [Google Scholar] [CrossRef]
- Ironside, J.E. Multiple Losses of Sex within a Single Genus of Microsporidia. BMC Evol. Biol. 2007, 7, 48–48. [Google Scholar] [CrossRef]
- Maside, X.; Gómez-Moracho, T.; Jara, L.; Martín-Hernández, R.; De la Rúa, P.; Higes, M.; Bartolomé, C. Population Genetics of Nosema apis and Nosema ceranae: One Host (Apis mellifera) and Two Different Histories. PloS One 2015, 10, e0145609–e0145609. [Google Scholar] [CrossRef]
- Gómez-Moracho, T.; Maside, X.; Martín-Hernández, R.; Higes, M.; Bartolomé, C. High Levels of Genetic Diversity in Nosema ceranae within Apis mellifera Colonies. Parasitology 2014, 141, 475–481. [Google Scholar] [CrossRef]
- Yucel, B.; Dogaroglu, M. The Impact of Nosema apis Z. Infestation of Honey Bee (Apis mellifera L.) Colonies after Using Different Treatment Methods and Their Effects on the Population Levels of Workers and Honey Production on Consecutive Years. Pak. J. Biol. Sci. 2005, 8, 1142–1145. [Google Scholar] [CrossRef]
- Hamiduzzaman, M.M.; Guzman-Novoa, E.; Goodwin, P.H. A Multiplex PCR Assay to Diagnose and Quantify Nosema infections in Honey Bees (Apis mellifera). J. Invertebr. Pathol. 2010, 105, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Snow, J.W. A Fluorescent Method for Visualization of Nosema Infection in Whole-Mount Honey Bee Tissues. J. Invertebr. Pathol. 2016, 135, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Milbrath, M.O.; Van Tran, T.; Huang, W.-F.; Solter, L.F.; Tarpy, D.R.; Lawrence, F.; Huang, Z.Y. Comparative Virulence and Competition between Nosema apis and Nosema ceranae in Honey Bees (Apis mellifera). J. Invertebr. Pathol. 2015, 125, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, L.; Beaman, L.; Holloway, B.; Rinderer, T.E. External and Internal Detection of Nosema ceranae on Honey Bees Using Real-Time PCR. J. Invertebr. Pathol. 2012, 109, 323–325. [Google Scholar] [CrossRef]
- Szalanski, A.L.; Tripodi, A.D.; Trammel, C.E. Molecular Detection of Nosema apis and N. ceranae from Southwestern and South Central USA Feral Africanized and European Honey Bees, Apis mellifera (Hymenoptera: Apidae). Fla. Entomol. 2014, 97, 585–589. [Google Scholar] [CrossRef]
- Rivière, M.-P.; Ribière, M.; Chauzat, M.-P. Recent Molecular Biology Methods for Foulbrood and Nosemosis Diagnosis. Rev. Sci. Tech. OIE 2013, 32, 885–892. [Google Scholar] [CrossRef]
- Amoguimba, E. Determinación de La Prevalencia y Georreferenciación de Varoosis y Nosemosis En Colmenares de Apis mellifera En Tres Provincias Del Ecuador En El Año 2015 (Chimborazo, Tungurahua y Bolivar). 2015, 95–95.
- Vivas, J.L. Prevalencia de Nosema (Nosema spp.) En Colmenares de La Región Norte y Centro Norte Del Ecuador. 2015, 81–81.
- Gajger, I.T.; Vugrek, O.; Grilec, D.; Petrinec, Z. Prevalence and Distribution of Nosema ceranae in Croatian Honeybee Colonies. Vet. Med. (Praha) 2010, 55, 457–462. [Google Scholar] [CrossRef]
- Higes, M.; Martín-Hernández, R.; Meana, A. Nosema ceranae in Europe: An Emergent Type C Nosemosis. Apidologie 2010, 41, 375–392. [Google Scholar] [CrossRef]
- Martín-Hernández, R.; Meana, A.; García-Palencia, P.; Marín, P.; Botías, C.; Garrido-Bailón, E.; Barrios, L.; Higes, M. Effect of Temperature on the Biotic Potential of Honeybee Microsporidia. Appl. Environ. Microbiol. 2009, 75, 2554–2557. [Google Scholar] [CrossRef]
- Sagastume, S.; del Águila, C.; Martín-Hernández, R.; Higes, M.; Henriques-Gil, N. Polymorphism and Recombination for rDNA in the Putatively Asexual Microsporidian Nosema ceranae, a Pathogen of Honeybees. Environ. Microbiol. 2011, 13, 84–95. [Google Scholar] [CrossRef]
Figure 1.
Spores (black circles) of Nosema sp. by optical microscopic examination (400X).
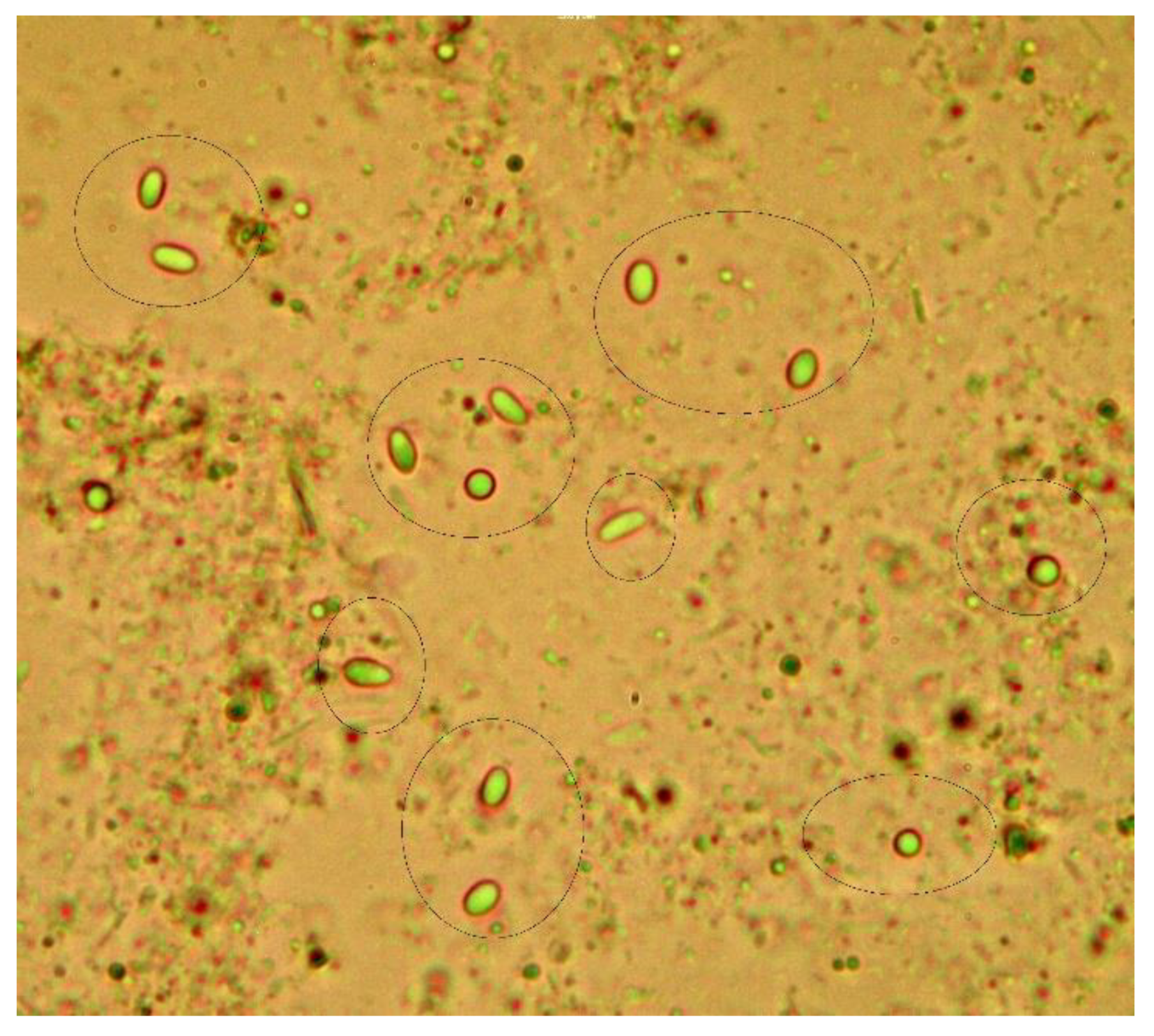
Figure 2.
Phylogenetic tree of microsporidia Nosema based on the sequences of the small subunit rRNA and constructed by Maximum Parsimony analysis. Blue: N. ceranae from Americas; gold: other Nosema species; green: N. apis; black: N. ceranae from continents other than Americas; red: N. ceranae from Ecuador. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches. The consistency index is (0.838), the retention index is (0.883), and the composite index is 0.783 (0.740) for all sites and parsimony-informative sites (in parentheses).
Figure 2.
Phylogenetic tree of microsporidia Nosema based on the sequences of the small subunit rRNA and constructed by Maximum Parsimony analysis. Blue: N. ceranae from Americas; gold: other Nosema species; green: N. apis; black: N. ceranae from continents other than Americas; red: N. ceranae from Ecuador. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches. The consistency index is (0.838), the retention index is (0.883), and the composite index is 0.783 (0.740) for all sites and parsimony-informative sites (in parentheses).
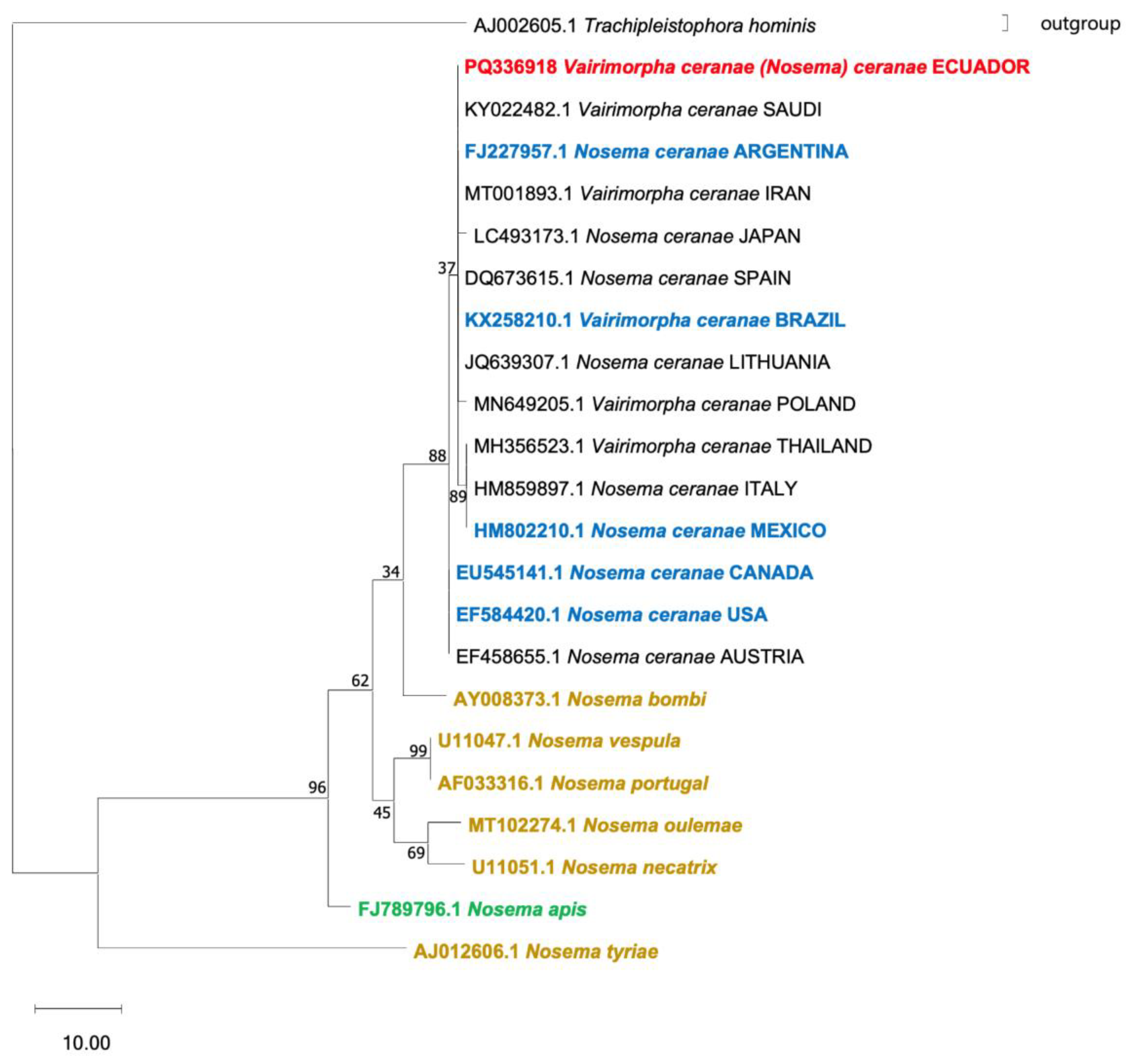
Figure 3.
Spores of Nosema sp. observed by Olympus IX53 fluorescence microscopy. Nosema’s spores stained with A) Hoesch dye and B) FB28 red dye at 40X.
Figure 3.
Spores of Nosema sp. observed by Olympus IX53 fluorescence microscopy. Nosema’s spores stained with A) Hoesch dye and B) FB28 red dye at 40X.
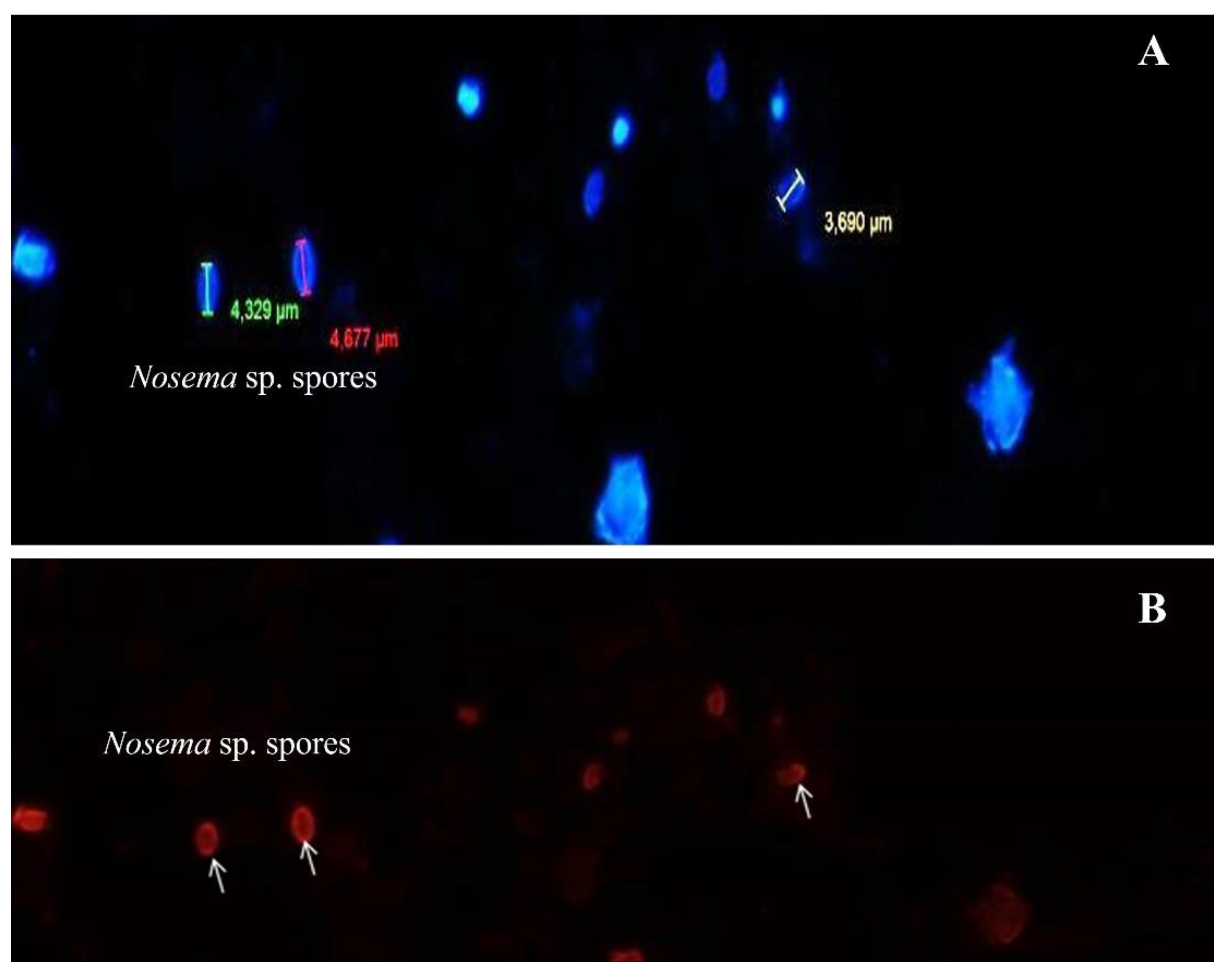

Table 1.
Distribution of existing and sampled apiaries and beehives in the three provinces surveyed.
Table 1.
Distribution of existing and sampled apiaries and beehives in the three provinces surveyed.
| Province | Number and percentage of apiaries of the national totala | Number and percentage of apiaries Sampling | Number and percentage of beehives of the national totalb | Number and percentage of beehives Sampling |
| Carchi | 40 (4,43%) | 4 (10%) | 974 (7,99%) | 33 (3,39%) |
| Imbabura | 74 (8,20%) | 13 (17,58%) | 1025 (8,41%) | 68 (6,64%) |
| Pichincha | 108 (11,97%) | 13 (12,04) | 2778 (22,79) | 63 (2,28%) |
| Total | 222/902a (24,61%) | 30 (13,51%) | 4777/12188 b | 164 (3,43%) |
a Apiaries at national level; b Number of hives nationwide.
Table 2.
Primers used for the detection and phylogenetic analysis of N. apis and N. ceranae.
| Primer name | Sequence (5’-3’) | Species | Fragment size |
| NosaRNAPol-F2* NosaRNAPol-R2 |
AGCAAGAGACGTTTCTGGTACCTCA CCTTCACGACCACCCATGGCA |
Nosemaapis | 297 bp |
| NoscRNAPol-F2* NoscRNAPol-R2 |
TGGGTTCCCTAAACCTGGTGGTTT TCACATGACCTGGTGCTCCTTCT |
Nosemaceranae | 662 bp |
| 218MITOC-FOR** 218MITOC-REV |
CGGCGACGATGTGATATGAAAATATTAA CCCGGTCATTCTCAAACAAAAAACCG |
Nosemaceranae | 218-219 bp |
Table 3.
Frequency of Nosema sp. by microscopy and multiplex PCR in the analysis of apiaries and hives and by each province.
Table 3.
Frequency of Nosema sp. by microscopy and multiplex PCR in the analysis of apiaries and hives and by each province.
| Province | Number apiaries | Microscopy | PCR | Total hives | Microscopy | PCR | ||||
| Number – prevalence in % (95% CI) Nosema sp. |
Number - prevalence in % (95% CI) N. apis |
Number – prevalence in % (95% CI) N. ceranae |
Number -prevalence in% (95% CI) Co-infection |
Number – prevalence in % (95% CI) Nosema sp. |
Number – prevalence in % (95% CI) N. apis |
Number – prevalence in % (95% CI) N. ceranae |
Number – prevalence in % (95% CI) Co-infection |
|||
| Carchi | 4 | 2 - 50% (6.76 – 93.24) |
3 - 75% (19.41 – 99.37) |
1 - 25% (0.63 – 80.59) |
1 - 25% (0.63 – 80.59) |
33 | 6 - 18.18% (6.98 – 35.46) |
6 – 18.18% (6.98 – 35.46) |
3 - 9.09% (1.92 – 24.33) |
1 - 3.03% (0.08 – 15.76) |
| Imbabura | 13 | 3 - 23.08% (5.04 -53.81) |
5 – 38.46% (13.86 – 68.42) |
7 – .15% (19.22 -74.87) |
0-* | 68 | 3 - 4.41% (0.92 – 12.36) |
10 – 14.71% (7.282- 25.39) |
9 - 13.24% (6.33 – 23.64) |
0-* |
| Pichincha | 12 | 7 - 58.33% (27.67 - 84.83) |
4 - 33.33% (9.92 – 65.11) |
8 - 66.67% (34.89 – 90.08) |
2 – 8.33% (0.21 – 38.48) |
63 | 19 - 30.16% (19.23 – 53.02) |
8 - 12.70% (5.65 - 23.50) |
23 - 36.51% (24.73 – 49.6) |
1 - 1.59% (0.04 – 8.53) |
| TOTAL | 29 | 12 – 41.38% (23.52 – 61.06) |
12 – 41.38% (25.52 – 61.06) |
15 – 51.72% (32.53 – 70.55) |
2 – 6.90% (0.85 – 22.77) |
164 | 28 - 17.07% (11.65 – 23.72) |
24 - 14.63% (9.61 – 20.99) |
35 - 21.34% (15.34 – 28.41) |
2 - 1.22% (0.15 4.34) |
*: IC does not apply.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



