
Submitted:
01 December 2025
Posted:
02 December 2025
You are already at the latest version
Abstract
Salinity stress is an abiotic threat that affects crop growth, development, and productivity, placing a heavy burden on global agriculture. Therefore, researchers must thoroughly understand the various mechanisms by which crops tolerate salinity. This knowledge enables biotechnological strategies to enhance crop resilience and yield, helping to secure sustainable food supplies for the growing world population. Several organic osmolytes greatly influence how crop plants respond to salinity stress. Trehalose is one such osmolyte that has gained increasing attention because it effectively improves crop salinity response and tolerance by influencing physiological, biochemical, and signaling processes. Beyond trehalose itself, its metabolite trehalose-6-phosphate (T6P), along with key biosynthetic enzymes such as trehalose-6-phosphate synthase (TPS) and trehalose-6-phosphate phosphatase (TPP), is vital for helping crops adapt to salinity, especially when applied externally or via genetic modifications. Trehalose performs multiple roles: it acts as a cell hydration agent, an antioxidant, a gene regulator, a source of energy and carbon, a precursor for metabolic pathways, a signaling molecule, and a detoxifier of excess reactive oxygen species (ROS) in cells and organelles. This review carefully explores the structure of trehalose, its biosynthesis, protective molecular mechanisms, and the important functions of its metabolites in crop adaptation to salinity. Strategies focused on increasing trehalose levels and functions or boosting its metabolites are promising approaches for researchers aiming to enhance crop tolerance and yield in saline environments, which we highlight. We suggest that trehalose, its precursor, and the biosynthesis pathway play various roles under both normal and stressed conditions, potentially serving as biomarkers to assess crop tolerance and productivity under saline conditions. This review offers valuable insights into trehalose metabolism, paving the way for future engineering techniques to improve crop tolerance and yield in saline soils.
Keywords:
exogenous application
; genetic engineering
; gene modulator
; salinity stress
; signaling molecule
; trehalose
1. Introduction
As agriculture provides the food and raw materials needed for human nutrition and many economic industries, it represents a crucial part of the world economy. Unfortunately, biotic and abiotic stresses pose numerous hazards to the world’s agriculture, which is not in parallel with the escalating global population, placing a higher burden on food demands worldwide. Salinity stress is an abiotic constraint that restricts plant growth, development, distribution, and productivity. Approximately one billion hectares of global arable land have been severely impacted by salinity over the last decade, and it is expected to result in a 50% loss by 2050 [1]. Excess salts in the soil can adversely reduce plant water uptake, cellular turgor pressure, and expansion, specifically due to the osmotic stress of salinity. Long-term exposure to salinity results in the accumulation of toxic ions, leading to plant ion toxicity and nutrient imbalance, commonly referred to as ionic stress caused by salinity. In addition, salinity stress leads to increased ROS production, which damages membrane lipids, proteins, and nucleic acids [2,3]. These hazardous effects of salinity cause drastic modifications in plants’ morphological, physiological, biochemical, and molecular functions, which are less severe in salinity-tolerant crop plants/genotypes [4,5,6,7].
Salt-resistant crop plants handle salinity stress effects through various mechanisms: osmotic adjustment, ion homeostasis, antioxidant defense systems, and membrane and cell wall modification [6,8,9]. Figure 1 summarizes the harmful effects of salinity stress resulting in a reduction of crop yield. Mitigating the negative impacts of salinity stress and understanding how crop plants tolerate salinity are crucial for increasing agricultural productivity and food security. Tolerant crop plants exposed to salinity stress synthesize and accumulate large amounts of secondary metabolites to maintain a low intracellular osmotic potential, having signaling functions, cellular redox potential buffering, chaperones, a storage form of nitrogen and carbon, the induction of ROS detoxifying systems, and stabilizing cellular macromolecules [6,8,10,11,12]. It is worth mentioning that these metabolites mediate osmotic adjustment when their absolute concentrations are high. Still, they have singling and protective functions at low levels in response to stress conditions [13,14]. Additionally, several methods are used to reduce the damaging effects of high salinity: biotechnological approaches using advanced genetic engineering and non-genetic strategies that involve applying natural plant metabolites externally [10]. Recent genetic studies, along with metabolome and transcriptome analyses, have identified various candidate genes for salinity tolerance that should be used in genetic engineering efforts to improve crop resilience to saline environments [6,15,16]. As climate change is expected to worsen adverse conditions, using these metabolome biomarkers in agricultural research provides a promising method to develop crops with improved salinity stress tolerance. These crops can grow efficiently and produce high yields even under challenging conditions.
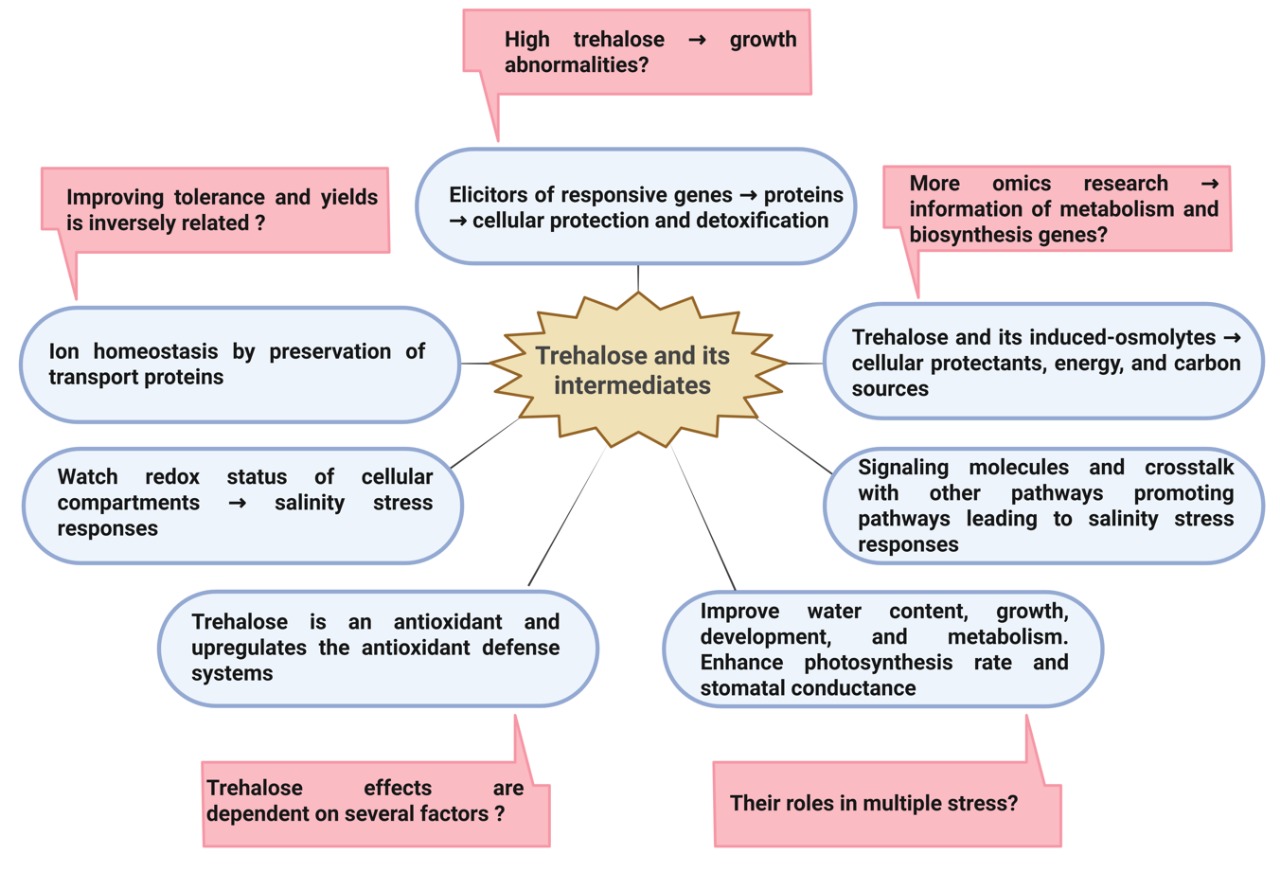
Evidence suggests that crop plants can produce trehalose, one of the frequently accumulated soluble sugars under salinity stress conditions, and was originally believed to serve as an osmoprotectant [17,18,19]. However, the presence of trehalose in plants at low levels (0.1-2 nmol g-1 FW) makes its role as an osmoprotectant unlikely [13]. A growing body of evidence indicates its implication in plant growth and development, numerous signaling cascades and metabolic pathways, ion homeostasis, as well as its vital role in stimulating stress tolerance [11,12,13,20,21,22,23,24,25]. Recently, the trehalose metabolic pathway has also been shown to play a crucial role as a regulator of carbon and nitrogen metabolism, as well as source-sink relationships, thereby largely influencing plant growth and development [24]. Additionally, under stressful conditions, trehalose has been shown to have an excellent capacity to protect cellular structures, outcompeting other sugars [26,27]. As such, trehalose exogenous application [12,13,25,27,28,29,30] and transformation of genes implicated in the trehalose metabolic pathway (trehalose-6-phosphate synthase, TPS; trehalose-6-phosphate phosphatase, TPP), resulting in elevated salinity tolerance and high-yielding crop plants [11,13,25,31,32]. Similarly, exogenous trehalose effectively reduces heat damage at high temperatures and improves the heat tolerance of rice varieties [33]. It achieves this by enhancing photosynthetic and carbohydrate metabolism, increasing antioxidant enzyme activity, raising osmoregulatory substance levels, and decreasing membrane lipid peroxidation in the flag leaves. Also, significant alterations in gene expression related to trehalose metabolism have been reported in response to a variety of abiotic stresses [13,24,34]. In addition to trehalose, evidence suggests trehalose-6-phosphate (T6P, a metabolite or precursor in the trehalose biosynthesis pathway) as a metabolic signal regulating sugar and energy homeostasis, and hence is a crucial regulatory component of plant metabolism and growth under stress [11,13,27,35]. We therefore believe that the external application of trehalose, engineering trehalose metabolism, and an intermediate and/or an enzyme of its biosynthetic pathway are potential biotechnological markers for alleviating salinity damage and enhancing crop tolerance and productivity. Furthermore, trehalose is also present in various organisms, including bacteria, yeast, fungi, insects, invertebrates, and plants, serving a wide range of functions [14,35,36,37]. In planktonic crustaceans, salinity-tolerant animals exhibited a positive correlation with trehalose concentrations, supporting the role of trehalose in salinity stress tolerance [38].
As for trehalose transport in plants, sugar transporters have been suggested to play essential roles in its transport and distribution throughout the plants [39,40]. These sugar transporters are typically categorized into two groups [39]: a) Those energized by electrochemical potential gradients created by proton pumps across the cellular membranes, i.e., secondary active transporters, and b) Sugar transporters that facilitate sugars to pass across membranes down the concentration gradients, i.e., facilitated diffusion. The purpose of this review is, therefore, to deliver updated evidence that trehalose is not only a sugar that acts as an innocent osmoprotectant osmolyte, and to answer the question of whether trehalose or its biosynthesis pathway intermediate (trehalose-6-phosphate) or enzyme (trehalose-6-phosphate synthase, TPS; trehalose-6-phosphate phosphatase, TPP) can be utilized as a biomarker to develop crop salinity-tolerant and yielding under saline stress. Reassessing the significance of trehalose or its derivatives accumulation in the homeostasis of crop plants that produce it and trehalose structure, biosynthesis, functional molecular mechanisms, engineering, and external application as efficient strategies to improve crop salinity and yield were discussed.
2. Trehalose Structure, Biosynthesis, and Protective Molecular Mechanisms
2.1. Structure and Properties
Trehalose is a natural soluble nonreducing disaccharide formed by a 1,1-glycosidic bond between two α-glucose units (Figure 2), giving it the name α-D-glucopyranosyl-(1→1)-α-D-glucopyranoside [14,19,41]. This bonding keeps trehalose in closed-ring form, stabilizes its structure at high temperatures, and is very resistant to acidic conditions and hydrolysis [14,42]. In addition, the glycosidic bond connecting the two glucose units has low energy (1 kcal/mol), resulting in a very stable structure relative to sucrose, which needs a high-energy bond (27 kcal/mol). Also, in the absence of trehalase, breaking trehalose into its two hexoses is not easy [19]. These superior trehalose physicochemical properties make it an exceptional sugar. Further, trehalose in aqueous solutions tends to form a concentration-dependent clustering and hydrogen bonds among them, thus forming clusters of various sizes [43]. Owing to trehalose’s ability to retain water, it is utilized in food, cosmetics, and as a drug. Among the three possible anomers of trehalose: α,β-1,1-, β,β-1,1-, and α,α1,1-, only α,α-trehalose (Figure 2) has been observed in living organisms [43].
2.2. Biosynthesis and Hydrolysis
It wasn’t until 1998 that genes encoding catalytically active trehalose 6-phosphate synthase (TPS, EC 2.4.1.15) and trehalose 6-phosphate phosphatase (TPP, EC 3.1.3.12) were identified in Arabidopsis, and the importance of trehalose metabolism led to its biosynthesis and heterologous expression of microbial TPS and TPP genes [18,42]. These genes are identified in all major plant taxa, indicating their capacity to synthesize trehalose and demonstrating its universal presence in the plant kingdom [13,14,24]. Five biological pathways support trehalose biosynthesis in various living groups. The most common pathway that plants use consists of two steps [13,41,43]: in the first step, the TPS enzyme catalyzes glucose transformation from uridine diphosphate-glucose (UDP-glucose) into glucose-6-phosphate (G6P), resulting in trehalose-6-phosphate (T6P) and UDP (Figure 3). In the second step, the TPP enzyme mediates the dephosphorylation of T6P into trehalose. TPP and T6P have been indicated to be located in the chloroplasts as well as other various cellular compartments, including vacuoles, cytosols, and nuclei [24,44,45]. This biosynthetic pathway is the most widely distributed in all organisms, where TPS and TPP are the major enzymes in trehalose metabolism, and also various isoforms of each are found in various plants [26,45]. For instance, 11 TPS genes and 10 TPP genes are identified in Arabidopsis [26], while in rice, 14 TPS and 13 TPP genes were identified [14]. The TPS genes are composed of class I and class II genes, where only AtTPS1 from the four genes encodes for an active synthase in Arabidopsis. On the other hand, it is worth mentioning that class II genes do not encode active enzymes [26], and hence, we believe future research should explore a possible role for their proteins under stress conditions. The two genes (TPS and TPP) and their homologs, coding trehalose, were identified in different plant species, and their expression in several crop species resulted in trehalose accumulation and enhanced abiotic stress tolerance [11,13,46,47]. However, several options should be considered to achieve more success by altering the trehalose metabolism of plants because overexpression of the trehalose biosynthetic genes leads to various growth aberrations [26,42,48]. That is, using the plant endogenous genes and the expression of TPS and TPP should be limited to certain levels, and at the same time, utilize stress-inducible promoters to regulate the introduced genes (42). Further, Eh et al. [13] presented evidence that these aberrant phenotypes are related to changes in T6P levels, and the co-expression of TPS and TPP is a successful method that results in no phenotypic abnormalities. The authors report that the absence of these aberrant phenotypes in response to the co-expression of microbial TPS and TPP genes is attributed to the contention that the fusion enzymes immediately convert T6P to trehalose without accumulating the intermediate T6P. Delorge et al. [26] also suggest modification of trehalose metabolism to improve stress tolerance without adverse effects through trehalase overexpression, which hydrolyzes trehalose into two glucose units (Figure 3). It is worth mentioning that salinity stress-related signaling molecules such as ABA and ROS regulate TPS and TPP genes and, hence, trehalose production [12,49,50]. ABA activates transcription factors like ABA-RESPONSIVE ELEMENT BINDING FACTOR 2 (ABF2) to bind to TPS/TPP promoters, which activate their expression, thus boosting trehalose production and enhancing plant tolerance to saline conditions (Figure 3). Trehalose accumulation triggers the production of ROS, creating a feedback loop that further enhances the ABA signaling pathway and strengthens ABA’s effects on processes like stomatal closure and other stress responses, which improves salinity tolerance [49,50]. For example, AtTPPD overexpression lines showed improved salinity tolerance due to hypersensitivity to redox changes in two cysteine residues of TPPD, which led to trehalose accumulation [51].
Trehalose can only be split into its monosaccharide residues under extreme hydrolytic agents or in the presence of the enzyme trehalase, which hydrolyzes trehalose into two glucose moieties [26]. It is proposed that the ubiquitous presence of the enzyme trehalase in plants may result in low amounts of trehalose frequently found in various plant species [26]. Trehalose breakdown can also occur by trehalose phosphorylase, producing glucose and glucose-1-phosphate [45]. Because of this controversy, it seems that trehalose itself may not be the effector but is more likely an intermediate and/or an enzyme of its biosynthetic pathway that plays a key role in crop growth, development, and stress tolerance [18,52].
2.3. Molecular Mechanisms of Trehalose/T6P’s Protective Role
Trehalose’s protective potential has been reported to be due to mechanisms that are a result of its physicochemical properties, particularly trehalose as a non-reducing sugar [14,46]. Due to its nonreducing nature, trehalose is less chemically reactive, making it compatible with cellular metabolism even at high concentrations. Its high levels do not harm cellular macromolecules but can help preserve them [46]. Three possible mechanisms have been suggested to explain how trehalose does not damage cellular structures: water replacement, glass formation, and chemical stability [19]. These trehalose protective mechanisms may all work together to stabilize the cellular macromolecules, and they are not mutually exclusive. All biological macromolecules are normally stabilized by a hydration layer around them, which is formed by hydrogen bonds between water and the cellular macromolecules. The water replacement theory suggests that trehalose preserves biological structures against dehydration, caused by different stresses, by trehalose replacement of the water molecules in the hydration layer, thereby helping to stabilize the biomolecules and inhibit their irreversible denaturation [19,25,46]. This feature of trehalose is due to the flexible nature of the α-(1→1) glycosidic bond that is greater than that of the glycosidic bond of other disaccharides; this allows trehalose to easily join the polar groups of the molecules and hence to have compatible functions with them [53]. Additionally, other disaccharides can displace water, but the trehalose hydration number exceeds that of the other disaccharides [19]. In this respect, Bezrukavnikov et al. [54] report that DNA bases have a higher affinity for trehalose than water, which promotes the melting of double-stranded DNA and stabilizes single-stranded nucleic acids. Secondly, trehalose can crystallize into a glass-like appearance under extreme dehydration; a particular property of trehalose that preserves cellular macromolecules from denaturation and recovers their functional activity upon rehydration [55]. Another function of this glassy formation is its ability to limit molecular motion and prevent protein aggregation and free radical diffusion under desiccation stress [42]. It thus seems that the key difference that makes trehalose surpass other disaccharides is its ability to form this glassy structure around molecules even in the absence of water, ensuring their stability even under harsh conditions [42,53,56]. It is worth mentioning that this glassy state has also been reported in low concentrations of trehalose, which was attributed to the transition from weak to strong correlated hydrogen bonds [57]. Thirdly, the chemical stability of trehalose is owing to the strong resistance of the α-(1→1) glycosidic bond to being cleaved by glucosidases and acid hydrolysis, and therefore the trehalose stability in solution at high temperatures and acidic conditions [56]. As mentioned earlier, this bonding of trehalose has low energy (1 kcal/mol) compared with another similar disaccharide, sucrose (27 kcal/mol). Because of these peculiar molecular features of trehalose, a focus of interest on its role in improving the performance of crop plants under various abiotic challenges has been reported [21,22,24,58,59,60,61,62]. These previous works report that trehalose efficiently stabilizes dehydrated enzymes, proteins, biological structures, and membrane lipids, which preserves their structures and hence protects them against the harsh impact of various abiotic stresses. Additionally, trehalose enhances crop salinity tolerance by mitigating the adverse effects of oxidative stress; trehalose acts through antioxidant defense system activation and/or functions as a ROS scavenger, thereby lowering ROS generation and their detrimental effects [25]. The signaling metabolite T6P, on the other hand, has the potential to relate reducing power with energy metabolism to fuel growth as detailed in a coming section.
As for the molecular mechanisms relating trehalose and T6P to salinity stress tolerance, T6P has been demonstrated to serve as a crucial signal linking carbohydrate metabolism to growth and stress responses [12,13,27,46]. That is, T6P is a central metabolic regulator that links carbohydrate availability to growth and stress responses. Also, Elevated T6P levels promote growth pathways, while lower T6P levels trigger the mobilization of carbon reserves for stress response. T6P can also regulate the expression of genes involved in stress tolerance and thus modulate plant resistance to salinity. On the other hand, trehalose has been reported to act as a compatible solute and a direct osmoprotectant to maintain water potential and prevent cellular damage, stabilizing cell membranes and proteins, and upregulating antioxidant genes [11,12,13,21,22,27,46]. In short, T6P acts as a sensor and signaling molecule, modulating cellular resources towards survival under salinity stress, while trehalose directly mitigates the damaging effects of salinity on cellular structures and functions.
3. Functions of Trehalose and Its Metabolites
Salinity stress seriously alters plants’ physiological, biochemical, and metabolic processes, resulting in drastic growth and yield loss. To overcome the stress damage, a large set of tolerance mechanisms operates in crop plants; some are constitutive, while others are expressed only when specific stress signals are perceived [41]. These events include changing signal transduction pathways, enhancing the downstream gene expression, ion homeostasis mediated by various transport systems, and accumulation of innocuous osmolytes in response to stress [8,42,46,63]. Among the compatible osmolytes, trehalose and T6P occupy an important place, have multiple functions in crop plants, and act as stress protectants and positive regulatory signals involved in adaptive responses and tolerance to stress conditions [11,13,20,21,22,64,65,66,67,68]. Accumulated trehalose, as well as its intermediate T6P, helps to overcome different adverse conditions by protecting biological molecules, inducing transcriptional activation of the stress-responsive genes, improving ion homeostasis, energy, and carbon reserve, signaling molecule, antioxidant, and promoting other antioxidants or inhibiting trehalase activity [11,19,34,41,42,46,47]. It is important to mention that once the stress condition is relieved, trehalose returns to normal levels, supportive of its important roles under stress conditions. However, the trehalose functions in plants remain controversial because transgenic plants overexpressing trehalose biosynthetic genes unexpectedly display a wide range of phenotypic abnormalities and stress symptoms [26,55,69,70]. The challenges caused by trehalose can be managed by altering the endogenous genes responsible for trehalose biosynthesis. It is essential to identify specific subcellular locations for these modifications and to appropriately direct them towards particular cell types, tissue categories, and stages of development. Such targeted interventions could significantly reduce the growth anomalies associated with trehalose. Eh et al. [13] recommend overexpression of microbial TPS and TPP fusion genes as a promising approach to exhibit no aberrant phenotypes. Moreover, evidence demonstrates that trehalose itself may not be the effector but more likely an intermediate (T6P) or an enzyme of its biosynthetic pathway (trehalose-6-phosphate synthase, TPS; trehalose-6-phosphate phosphatase, TPP) that plays a pivotal role in plants under stress conditions [18,46,47,52]. In this section, the different roles of trehalose or its metabolites in enhancing plant tolerance to salinity stress are discussed and evaluated. These mechanisms may all contribute to the mitigative functions of trehalose and do not exclude one another, i.e., they are not mutually exclusive.
3.1. Regulation of Crop Growth, Development, and Carbohydrate Metabolism
In response to saline conditions, plant content of trehalose, its metabolic intermediate, or an enzyme involved in its biosynthesis plays a key role in the regulation of growth, development, and metabolism [14,24,25,47,49,64,71]. One crucial role of trehalose reported by Yang et al. [69] is its benefit for vegetative and floral growth in salinity-stressed plants. In the work of Yang et al. [21,22,69] both vegetative and reproductive growth were largely improved by the trehalose supply, highlighting the possible role of trehalose in helping plants to antagonize salinity stress in part through the completion of the life cycle under saline situations. It is most likely that the role of trehalose during plant salinity response is critical for crop productivity because severe salinity stress will drastically interrupt plant floral growth, which eventually affects yield potential. Trehalose treatment also increased the growth parameters and yield of various plants, which declined under saline soil: quinoa plants [29], wheat cultivars [72], and Catharanthus roseus [73]. The improved growth and resistance to high salinity observed in the above works by trehalose were always associated with ionic regulation, osmotic adjustment, and enhanced antioxidant defense system. In the same trend, trehalose metabolism disruption was associated with severe alterations in embryogenesis, flower formation, organogenesis, growth, and senescence, highlighting trehalose and T6P’s essential roles in plant growth and development at all growth stages [24,74]. In addition, trehalose impacts sugar metabolism since the reduced level of trehalose through trehalase hydrolysis of trehalose causes a drastic decline in starch and sucrose contents [46], suggestive of trehalose’s role in carbon allocation and sugar metabolism. Also, T6P, which is an intermediate in trehalose biosynthesis, is crucial for embryonic and vegetative development of plants, and sucrose and starch metabolism [24,71]. The main role of T6P is to signal and regulate sucrose levels under normal and stressed conditions via acting as a negative feedback regulator [24,74], although how T6P senses and regulates sucrose levels remains unclear and needs further exploration. However, Blanford et al. [75] recently reported that T6P is a key for cellular sugar status and a strong inhibitor of Sucrose non-fermenting 1-related protein kinases 1 (SnRK1) through binding to SnRK1 catalytic subunit (KIN10), which disrupts the activation loop and precludes its phosphorylation and activation by GRIK1 under high sugar. Under these conditions, SnRK1 minimally phosphorylates its target proteins, switching from catabolism to anabolism [75]. In this respect, Eh et al. [13] indicate that when the T6P level is low, it enhances SnRK1 activity and cellular metabolism shifts toward the famine direction, strengthening stress responses. Conversely, when the T6P level is high, SnRK1 activity is suppressed, causing plant cell metabolism to shift towards a feast state, which in turn increases biomass and yield. Other studies showed that trehalose regulates embryo maturation, plant growth and development, flowering, carbohydrate and abscisic acid metabolism, as well as stress signaling [19,64]. In this respect, it is worth mentioning that the growth abnormalities found in the transformed plants are possibly due to the accumulation of T6P, confirming that T6P is a key mediator in plant development. Our proposal is supported by the finding that homozygous tps1 mutants exhibited embryo-lethal phenotypes in Arabidopsis. Similarly, when the TPS gene was rendered non-functional, aquatic invertebrate genotypes could not produce trehalose and suffered drastically decreased salinity tolerance, emphasizing the importance of this sugar and its metabolite in their survival [38]. As such, the AtTPS1 gene has been reported to be essential for root and shoot growth as well as for the transition to flowering [47]. Further, T6P has been reported to inhibit SnRK1, which is an important transcriptional regulator of metabolism, growth, and development in plants [24,35,59]. As detailed above, T6P seems to act as a potent switch for plant growth and catabolism under normal and stressful conditions [75]. Phan and Van Dijck [48] also report that the role of T6P as a signaling molecule, regulator of plant metabolism, growth, and development, becomes clear because trehalose-induced growth inhibition is related to T6P accumulation in Arabidopsis seedlings. High trehalose levels might lead to T6P dephosphorylation reduction, which increases T6P contents. In this work, the growth inhibition by T6P was restored by adding metabolizable sugars to the growth medium enriched with trehalose. This finding reveals that when the balance between T6P contents and sugar availability is impaired, growth is reduced, which can be restored by the sucrose addition to the medium, suggesting that T6P improves plant growth when the carbon supply is high. The results suggest that trehalose metabolism is important for plant development, and T6P adequate levels are essential for carbon utilization during normal growth. Based on this, it is obvious that the growth and reproduction of the salinity-stressed crop plants largely benefit from trehalose and/or its biosynthesis intermediates and enzymes.
Another example supporting the important role of T6P in regulating crop metabolism in response to salinity imposition is the sucrose: T6P ratio [24,59,76,77]. In plant cells, the sucrose: T6P ratio has been reported to affect various metabolic processes via the induction or repression of transcription factors in response to different stresses. For instance, elevated T6P levels repress the catalytic activity of SnRK1, a key transcriptional regulator affected by carbon and energy availability [19,78]. Consequently, T6P exhibits an adverse effect on genes that are upregulated by SnRK1 while simultaneously exerting a beneficial influence on genes that are downregulated by SnRK1 [77]. Furthermore, T6P appears to function as a negative feedback regulator, modulating sucrose concentrations through its interaction with SnRK1, which was previously discussed [75]. This regulatory loop between T6P and SnRK1 leads to a balance in growth when energy status is favorable or restrains metabolism when energy status is low. Henry et al. [76] further reported that the interaction between T6P and sucrose is a developmental stage-, tissue-, and cell-type-dependent process, and is regulated by environmental factors. Proposed mechanisms for the inhibition of SnRK1 by T6P are forwarded by Kerbler et al. [24]: T6P inhibits SnRK1 via interaction with unknown protein factors and indirectly by inhibiting the interaction of SnRK1 with SnRK1 activating kinases (SnAKs GRIKs). They also indicate that class II TPS proteins and non-catalytic TPS-like proteins similarly act as inhibitors of SnRK1. This suggestion, presented by Kerbler et al. [24], is currently further unmasked by the work of Blanford et al. [75]. It, therefore, appears that T6P and SnRK1 signaling are involved in stress response and recovery by regulating genes that utilize sucrose in growth and development and the synthesis of end products. The evidence indicates that the metabolism of trehalose, via its intermediates and associated enzymes, is crucial in regulating the distribution of carbon and the coordination between sources and sinks. Another transcription factor that regulates carbohydrate metabolism and is controlled by T6P is basic leucine zipper 11 (bZIP11) [76]. Therefore, it can be concluded that T6P indirectly modulates the developmental phase transitions, carbohydrate, and amino acid metabolisms since these processes are regulated by bZIPs. This contention is supported by the finding that when trehalose is supplied in high concentrations, the observed plant toxicity has been linked to T6P accumulation and bZIP11, as its overexpressing plants show insensitivity toward supplied trehalose [79]. It seems that a connection between T6P, SnRK1, or bZIP11 possibly explains the resulting toxicity of trehalose, and most likely impacts the responses and tolerance to stress. We recommend that in engineering interventions, T6P should be precisely modified and targeted to specific cell types and tissues to avoid unfavorable responses.
A further feature of trehalose’s impact on plant growth and development is provided by the fact that trehalose and ABA have synergistic effects on root growth and stomatal closure [49]. In this respect, ABA has been found to stimulate trehalose accumulation, and trehalose external supply significantly enhanced ABA biosynthesis in both the roots and leaves of Avicenna marina [50]. The expression level of TPPE, as one of the ten genes encoding TPPs and trehalose levels in Arabidopsis increased in response to ABA: the ABA--responsive transcription factor ABA responsive element-binding factor2 (ABF2) in the presence of ABA directly binds to the TPPE promoter and activates its expression. TPPE is, therefore, implicated in ABA--controlled stomatal movement and root elongation, since in the tppe mutant, stomatal closure was less sensitive to ABA than wild-type plants [24]. Similarly, Song et al. [50] have confirmed the direct binding of AmABF2 to the AmTPS9A promoter, activating its expression and activity in vitro and in vivo. Accordingly, the results uncover a new aspect of the ABA signaling pathway and provide a molecular basis for the trehalose role in crop responses to abiotic stresses. Moreover, it has recently been found that trehalose metabolic pathways are also important for normal seed germination [80]. For instance, hydrogen gas-enhanced seed germination in cucumber has been illustrated to be through enhancing trehalose biosynthetic enzyme activity and gene expression, decreasing starch level by promoting-related enzyme activities and their gene expression, thereby elevating the endogenous trehalose level [80]. It is, therefore, evident that hydrogen crosstalk with trehalose serves as a signaling molecule that plays an important part in seed germination. In conclusion, the regulation of plant growth, development, and sugar metabolism by T6P/trehalose is hence a noncontroversial fact, and their precise modulation is recommended for biotechnological interventions for crop resilience and yield potential improvement, which surely contributes to sustainable agriculture and global food security.
3.2. Trehalose/T6P Induces Other High Sugar Levels, Serving as an Energy and Carbon Source
Sugars have been demonstrated to provide carbon and energy for normal cellular metabolism and hence regulate plant metabolism, growth, and development under normal and stressed conditions [67]. In support, salinity-induced starch accumulation in tomato roots and leaves reduced the photosynthetic performance, chlorophyll content, and starch transport. At the same time, exogenous trehalose treatment overcomes these adverse effects and supplies the necessary sugars and energy for resilience processes [40]. Also, trehalose accumulation has been shown to influence sugar metabolism that is required for the osmoprotectant role under different environmental stresses [25,26,81,82]. Research by Sah et al. [64] similarly showed that nonreducing disaccharides (trehalose and sucrose) provided a soluble energy source as stable molecules, as well as acting as a protectant compound in crop plants in response to stress conditions. The previous authors reported that accumulated trehalose acts as a storage carbohydrate that possesses the unique feature of reversible water absorption capacity to maintain cell turgidity and protect biological molecules from stress-induced dehydration. In this respect, trehalose is superior to other sugars, conferring protection. In a study by Abdallah et al. [60], trehalose priming induced soluble sugar accumulation when rice cultivars were subjected to saline conditions, suggesting trehalose’s crucial role as a modulator of other carbohydrate molecules and sugar transport. Additionally, trehalose supply to salinity-stressed Arabidopsis resulted in improved shoot fresh weight and leaf water content relative to untreated plants, which was explained by the fact that trehalose induced the enzymes that catalyze the accumulation of storage carbohydrates in photosynthetic tissues [69]. Similarly, exogenous trehalose increases the activities of sucrose synthetic enzymes (sucrose phosphate synthase and sucrose synthase) and soluble sugars (trehalose, sucrose, glucose 6-phosphate, fructose 6-phosphate), which improves the energy status and membrane stability, and lowers chilling adversities in guava fruit stored in a cold environment [83]. Another recent work by Vichaiya et al. [84] illustrated that postharvest trehalose supply enhanced the expression of energy-producing related genes that encode (i.e., adenosine triphosphate synthase subunit β, adenosine diphosphate/adenosine triphosphate carrier) and stimulated the enzyme activities of nicotinamide adenine dinucleotide dehydrogenase, succinate dehydrogenase, and cytochrome c oxidase. These trehalose-inducible responses were associated with higher energy charges and reduced chilling damage in cold storage guava fruits. These findings suggest that trehalose supply resulted in a transient increase in sucrose level at early storage to act as sugar signaling to SnRK1 activation, which is required for maintaining energy homeostasis needed for chilling injury suppression. In a field experiment, a foliar spray of trehalose improved yield, yield components, and chemical composition of quinoa seeds under drought stress, which was attributed to trehalose serving as a storage carbohydrate and transport sugar [85]. Interestingly, Raza et al. [45] reported that T6P increased maize growth and development by improving carbon availability under cold stress. Also, as T6P has a dual function (a signal molecule and homeostatic regulator of sucrose levels), Kerbler et al. [24] indicated that T6P in source leaves controls the sucrose production to balance supply with demand for sucrose from growing sink organs. Further, in addition to T6P role as an important signaling metabolite, T6P regulates carbon assimilation and sugar status in plants and plays an essential role in plant development [74]. Based on that, trehalose/T6P has been demonstrated to indirectly have important roles as a carbon storage, energy source, and carbon transport molecule by triggering the synthesis of high levels of other sugars in response to stress conditions. However, Kerbler et al. [24] argued that a low level of trehalose, always found in higher plants, is inconsistent with its role as carbon storage. We therefore propose that other high levels of sugars regulated by trehalose may serve as carbon sources for energy, growth, and maintenance purposes in response to stress.
Further evidence concerning indirect trehalose’s role as an energy and carbon reserve is provided in this section. For example, one crucial role of trehalose in inducing plant-rhizobia interaction and thus improving abiotic stresses is its role in acting as a source of carbon and energy in different phases of the rhizobium-legume association [86]. Feng et al. [40] also demonstrated that an external supply of trehalose significantly upregulates the expression of fructokinase and phosphofructokinase in tomato plants under salinity stress, suggesting trehalose’s capacity to enable plants to accumulate sugars and maintain efficient respiration. The work also illustrated that, in addition to sugar’s role as the primary energy source for plant growth, it can also serve as a signal molecule under stress, as trehalose treatment was found to upregulate sugar signal transduction genes in tomato under salinity stress. Furthermore, the expression level of the monosaccharide transporter (MST3) gene and the sucrose transporter genes SUT1 and SUT4 were significantly upregulated under salinity and trehalose treatment [40], suggestive of trehalose’s ability to alter the source-sink transport pathway of sugars, thereby regulate the accumulation and distribution of other sugars to provide the required source of carbon and energy for tomato plants. It seems that tomato plants were unable to utilize carbon sources under salinity stress. Sadak [87] similarly revealed that trehalose foliar application enhanced growth parameters and accumulation of glucose, sucrose, trehalose, starch, and soluble sugars of salinity-stressed plants, indicative of the indirect impact of trehalose on carbohydrate metabolism to serve in turgor maintenance and as a reserve form of carbon. Under drought conditions, Bao et al. [68] reported that foliar spray of trehalose or T6P promoted starch, soluble sugar, and lignin accumulation in the petals, pointing to the role of T6P or trehalose as a positive regulatory signal participating in the accumulation of various sugars and enhancing the drought tolerance of rose plants. In summary, although trehalose (a soluble, non-reducing disaccharide) is present in low amounts in plants, it modulates sugar accumulation and distribution by affecting the activities of sugar transporters and simultaneously regulating sugar metabolism to adequately supply carbon and energy sources, thereby significantly improving plant salinity stress resistance. As trehalose is not always present in high enough levels in plants, it is obvious that trehalose’s role is mainly to modulate other sugar accumulation in sufficient amounts for the required energy and carbon sources under stress.
3.2.1. Induction of the Expression of Stress-Responsive Genes
Several reports indicate that trehalose-induced upregulation of stress-responsive genes improves crop resilience to stress conditions [14,24,46,55,88]. Products of these genes have been found to participate in tolerance traits in response to stress conditions. For example, Feng et al. [40] analyzed the metabolic changes in sugar and ABA in tomato seedlings treated with trehalose and salinity. They also determined the differential expression profiles of key genes involved in sugar metabolism. The authors report that the external supply of trehalose decreased the starch content and enhanced the soluble sugars by modulating the gene expression involved in the metabolism of starch and soluble sugars. The results also demonstrated that exogenous trehalose alters the sugar accumulation and distribution by upregulating sugar transporter genes and increasing the ABA content. This resulted in improving salinity tolerance by regulating the gene expression related to the synthesis and metabolism of ABA. In the same trend, trehalose added to 2-week-old liquid cultures containing Arabidopsis modified the transcript levels of transcription factors, cell wall composition, nitrogen metabolism, stress-responsive genes, defense-related genes, and fatty acid biosynthesis genes [88], suggesting that trehalose or its metabolite (trehalose6phosphate) regulated expression of genes (trehalose-6-phosphate synthase, TPS; trehalose-6-phosphate phosphatase, TPP) implicated in the mechanisms underpinning stress resilience. Another research by Bae et al. [89], where exogenous trehalose was applied to liquid cultures of Arabidopsis seedlings, found that trehalose rapidly altered disaccharide levels and induced detoxification and stress response proteins, highlighting trehalose’s role in modulating the stress-inducible genes that contribute to the observed important alterations. Additionally, Yang et al. [69] showed that exogenous trehalose not only increases the salinity tolerance of Arabidopsis but also possibly acts as a biological process inducer (i.e., ion homeostasis, ROS scavenging) involved in the salinity stress response, further evidence indicates trehalose’s capacity to trigger salinity-responsive genes regulating different processes that stimulate tolerance to saline imposition. Jiang et al. [90] also reported that when the IbTPS gene from sweet potato was transferred into tobacco, transgenic tobacco exhibited enhanced salinity tolerance, high trehalose, proline, and upregulation of stress-related genes (TPP, HSP70, DHN) compared with wild-type. The results indicate that enzymes of the trehalose biosynthesis pathway similarly contribute to enhanced salinity tolerance. Microarray analysis performed by Sah et al. [64] clearly indicates that both trehalose and T6P affect the expression levels of genes involved in abiotic stresses. Trehalose or T6P external application also induced drought-responsive genes under drought conditions; the response of genes was varied, suggesting functional differences between T6P and trehalose [68]. Phan and Van Dijck [48] similarly showed that trehalose-induced abiotic stress may be attributed to the upregulation of stress-responsive genes and transcription factors in various crops, rather than trehalose’s role as an osmoprotective molecule. Further, Kerbler et al. [24] reported that both T6P and TPP may act as signaling molecules, stress-inducible gene upregulators, water content elevation, ROS scavengers, stomatal conductance enhancers, and other processes that are involved in stress responses and resistance. Based on the above evidence, it is obvious that trehalose or its metabolites act as a modulator of stress-responsive genes that are implicated in the detoxification of stress’s harmful effects and enhancement of stress tolerance.
Further investigations support the fact that the improvement of trehalose-mediated abiotic stress tolerance is related to the activation of stress-inducible genes and transcription factors in addition to its role as a protectant molecule. For example, the reduced salinity harmful impact by exogenous trehalose application in rice was related to root integrity preservation, Na+ accumulation reduction, and activation of the genes responsible for osmotic adjustment [77]. Also, a study by Li et al. [91] revealed that in rice TPS overexpressing lines, the levels of trehalose and proline were highly elevated in the presence or absence of stress, and the expression of stress-related genes such as ELIP, HSP70, CRP, DHN6, LEA14A, and WS118 was enhanced up to twofold in transgenes comparing with wild-type plants. Recently, exogenous trehalose induced not only the antioxidant enzyme activities but also upregulated their related gene expression (SlCu/Zn-SOD, SlFe-SOD, SlMn-SOD, SlPOD, and SlCAT) in salinity-stressed tomato leaves [21], which greatly enhanced tomato plants to counteract the harmful effects of salinity. Similarly, foliar application of trehalose to rice cultivars enhanced antioxidant enzyme activities as well as their gene expression, resulting in decreased ROS levels under drought stress [34]. Another work by Hathout et al. [92] illustrated trehalose-induced inhibition of the enzyme trehalase resulted in the accumulation of tissue trehalose which was associated with starch and protein increase with the appearance of new protein patterns in two rice cultivars under saline soil, indicative of trehalose’s role as a modulator of genes involved in mechanisms that participate in rice salinity stress tolerance. Recent research by Vichaiya et al. [84] illustrated that postharvest trehalose supply upregulated the expression of energy-producing related genes, resulting in higher energy charges and lower chilling injury in cold storage guava. Further, Henry et al. [76] similarly reported that in maize, salinity upregulated the expression of several genes encoding enzymes and transporters involved in trehalose and sucrose metabolism. The research also revealed that the levels of many sugar metabolism intermediates, glycolysis, and tricarboxylic acid cycle were modified; a 2- to 4-fold increase in T6P levels in leaves, kernels, and cobs was observed at the silking stage. These findings suggest a general reconfiguration of gene expression and central metabolism in trehalose-treated maize under salinity stress. However, Figueroa and Lunn [42] argued that it is not clear from Henry and coworkers’ study whether the increase in T6P at the earliest developmental stage (i.e., silking) represents a primary response to salinity stress that promotes other gene expression and metabolic changes or if the altered T6P levels and T6P/sucrose ratios reflect a broader reconfiguration of metabolism in salinity-stressed plants. We believe that both responses might share the observed impacts because both effectors could induce such metabolic reconfiguration, and they are not mutually exclusive. Taken together, the previous findings indicate trehalose, its precursor, and/or enzymes of the synthesis pathway act as an upregulator of genes involved in salinity stress responses and tolerance in various crop plants. It is worth mentioning that although the upregulation of trehalose biosynthesis and exogenous trehalose supply showed protective, regulatory functions, and upregulation of stress-responsive gene expression in various crop plants under stress, the exact mode of action of trehalose or its metabolites in promoting such gene expression still needs further exploration and research.
3.2.2. Upregulation of Antioxidant Systems to Scavenge ROS
Salinity stress induces ROS production, which causes injury to macromolecules and membranes [2,3,9,14]. Therefore, the joint action of both antioxidant defense systems (i.e., enzymatic and non-enzymatic antioxidants) is vital to detoxify ROS and overcome salinity-induced oxidative stress in crop plants. Trehalose is one of the remarkable protectants due to its role that works directly to suppress the ROS or indirectly to activate other antioxidant systems and hence stabilizes the cellular membranes, proteins, and protein complexes under abiotic stresses [25,28,45,71,93]. However, when crops are overwhelmed by stresses and the perturbation becomes more dominant or evolves from various sources, an external application of trehalose or other relevant natural secondary metabolites (e.g., choline, β-carotene, anthocyanin) could be a promising strategy to combat the hazardous effects of stress. For instance, trehalose treatment efficiently elevates the endogenous trehalose concentration, which mainly induces the antioxidant defense system by enhancing the activities of antioxidant enzymes (superoxide dismutase, catalase, peroxidase) in two rice cultivars and quinoa plants, which improve plants’ capability to scavenge excessive ROS and overcome salinity stress effects [28,29,60]. In quinoa plants, in addition to activation of antioxidant enzymes, trehalose also stimulates nonenzymatic antioxidants (soluble sugars, trehalose, proline, free amino acids), clearly indicating trehalose’s capacity to upregulate both antioxidant defense systems, besides its direct effect as an antioxidant. Yang et al. [69] also demonstrated that exogenous trehalose decreased ROS, which minimizes the adverse effects of salinity stress: the results indicate that trehalose alleviated salinity-induced cellular ROS and the programmed cell death (PCD) process by enhancing the antioxidant enzyme activities and the level of compatible solutes serving as nonenzymatic antioxidants, which modulate plant salinity responses and promote tolerance to high salinity. Luo et al. [93] provided direct evidence that trehalose itself plays a key role in scavenging ROS in wheat under heat stress, indicative of trehalose’s role as an antioxidant. Additionally, overexpression of OsTPS1 enhanced rice tolerance to multiple abiotic stresses by increasing trehalose and proline levels as well as activating abiotic stress-related genes [91]. The accumulation of compatible osmolytes such as proline and glutamate was also found to remarkably improve the performance of salinity-stressed Catharanthus roseus plants when exogenous trehalose was applied [73]. It seems that elevated trehalose, proline, and glutamate act as nonenzymatic antioxidants that minimize oxidative damage induced under saline conditions. Recent investigation has positioned trehalose as a significant factor in enhancing the salinity tolerance of wheat: this was linked to its capacity to diminish ROS levels, increase the concentration of antioxidant compounds, and promote the synthesis of compatible osmolytes when exposed to NaCl stress [87]. The trehalose’s role in preventing salinity damage during stress was by acting as an antioxidant, triggering other antioxidants, and as a signaling molecule. Moreover, Feng et al. [40] and Islam et al. [25] showed that trehalose enhances the antioxidant enzyme activities and increases the proline level while reducing the MDA content, therefore, minimizing the adversities caused by salinity stress in tomato and Indian mustard plants. It is worth mentioning that exogenous trehalose enhanced antioxidant enzyme activities not during salinity stress but during the recovery period in rice seedlings; the effect was more pronounced in the salinity-sensitive cultivar [81]. This finding is suggestive of trehalose’s crucial role in eliminating oxidative stress in response to salinity imposition. The impact of trehalose during the recovery period is unclear, but its presence and effect may be extended after stress release in rice seedlings. Taken together, the results suggest that trehalose itself acts as an antioxidant, or by inducing the generation of nonenzymatic antioxidants and/or enhancing enzymatic antioxidant activities, enables plants to cope with salinity-induced ROS bursts and therefore contributes to the enhancement of tolerance to saline conditions. However, because of trehalose’s special chemical characteristics mentioned earlier, trehalose has been indicated to act as an antioxidant, though its underlying molecular mechanisms remain unclear and poorly understood.
Additional evidence supporting the role of trehalose or its derivatives in boosting the antioxidant defense system under stress is provided here. For example, researchers have reported that trehalose is a key regulator of antioxidant defense systems in plants exposed to various stress conditions [20,25,45,55,68,87]. Under drought stress, trehalose largely improves proline content and protects the biological molecules by enhancing the antioxidant defense system in flax plants [30]. Similarly, a foliar spray of trehalose or T6P elevated the production of secondary metabolites and soluble sugars that serve as nonenzymatic antioxidants under drought stress [68]. In a recent study by Wang et al. [94], trehalose-treated peaches had higher internal trehalose, sucrose, proline, and choline contents under chilling stress; these induced solutes most probably contribute to ROS scavenging and hence lower levels of chilling injury. Trehalose at 10 mM enhanced enzymatic antioxidant and glyoxalase activities, suggesting enhancement of ROS detoxification and reducing methylglyoxal in two maize genotypes under 150 mM NaCl stress [95]. Both effects protected maize genotypes against the harmful effects of salinity stress, which improved genotype resistance. Rohman and coworkers’ research also revealed that trehalose enhanced a reduction in lipid peroxidation, MDA, and ROS accumulation in response to salinity imposition, altogether pointing out trehalose’s capability to enhance maize salinity tolerance through acting as an antioxidant or regulating other antioxidants as well as glyoxalase systems. Moreover, Zulfiqar et al. [20] illustrated that trehalose treatment also elevated the internal content of trehalose, glycine betaine, and proline as well as the enzymatic antioxidant activities in sweet basil, a response that promotes drought tolerance. Also, increased antioxidant enzyme activities and their gene expression in leaves of rice cultivars in response to foliar trehalose application diminished ROS levels and reduced leaf electrolyte leakage under drought stress [34]. Recently, Islam et al. [25] reported that leaf-applied trehalose significantly increased Indian mustard salinity tolerance and yield by enhancing ion homeostasis, photosynthesis efficiency, antioxidant defense mechanisms, chlorophyll content, osmolyte accumulation, stomatal aperture, cell viability, and ROS scavenging under salinity stress. Based on the above sources, trehalose or its metabolites effectively modulate the cellular redox state, cell death, antioxidant defense system, and compatible osmolytes to control various crop performances and improve their resilience to salinity and other abiotic stresses. Therefore, we recommend trehalose as a cost-effective strategy for alleviating the salinity impacts on crop plants. However, further research is needed to explore how trehalose boosts antioxidant activities and the signaling crosstalk between this sugar and the antioxidant system under stress conditions.
3.3. Trehalose/T6P as a Signaling Molecule and Crosstalk with Sugars and Hormones
Despite being found very minutely in plants, it has many roles in plant cell metabolism that contribute to abiotic stress tolerance. For instance, Sah et al. [64] report that in most plants, trehalose serves as a signaling molecule rather than its direct involvement in abiotic stress alleviation. As such, in different living organisms, trehalose has been shown to act as a signaling molecule to regulate several metabolic events at different growth stages and shows crosstalk with other metabolites [13,40,43,55,64,68,96]. These reports indicate that trehalose metabolism is essential for normal plant growth and development, and its biosynthesis takes place via a phosphorylated intermediate T6P; the latter level alterations are comparable with sucrose concentration, which is the main product of photosynthesis and the major transportable sugar in plants [41,55]. That is, the intermediate T6P in the trehalose biosynthesis pathway senses the available sucrose and thus directly affects the response to harsh environmental situations. This is justified as T6P, trehalose, and/or one or more enzymes of their biosynthetic pathway are part of complex interaction networks concerted with hormone and sugar-induced signaling pathways, which may have a role at different developmental stages [55]. In addition, T6P is proposed to be implicated in the regulation of the SnRK1(SNF1/AMPK group of protein kinases) activity, which is a central modulator of energy and sugar homeostasis: T6P at low concentrations has been indicated to inhibit SnRK1 in vivo and in vitro [35]. For example, in Arabidopsis, T6P is a signaling molecule that regulates sucrose content and hence adjusts its level within the cell [55,74]. Also, the sugar-signaling metabolite T6P was increased in leaf, cob, and kernels at silking in response to salinity treatment, and showed to have a regulatory function in source and sink tissues [58,59,76]. Similarly, a study by Ibrahim and Abdellatif [97] supports trehalose’s role as a signaling molecule that induces wheat plants to enhance the synthesis of non-enzymatic antioxidants to scavenge ROS and thus minimize the damaging effects of water stress. The mechanism is that trehalose, as a signaling molecule, induces ROS production in plants under abiotic stresses, the latter sends a signal to promote enzymatic antioxidants to reduce ROS levels that are triggered by the stress. Additionally, drought stress inhibited the trehalose synthesis pathway in rose petals was alleviated by exogenously applied trehalose or T6P by promoting carbohydrate accumulation, secondary metabolites, and lignin in petals [68]. The results indicate that T6P or trehalose signaling is possibly related to these positive impacts and contributes to improved drought tolerance of rose plants, which maintains the quality of rose flowers under drought conditions. Therefore, the signaling function of trehalose and T6P, integrating development and metabolism about carbon supply, might be more essential than the other suggested protective or osmotic functions, although in some tissues and plants, the latter roles may be important and cannot be excluded [98].
From the studies carried out by Debast et al. [99] on potato, Henry et al. [76] on maize, Blanford et al. [75] on rose and the reviews by John et al. [41] and Onwe et al. [71], the following conclusions have been reached: a) TPS1 and/or T6P are major players in signaling pathways and gene modulation during seedling development and stress response, ii) trehalose as a stress response and signal plays a vital protective role during abiotic stress, and iii) crosstalk between trehalose and stress-responsive secondary metabolites that participate in plant stress tolerance is evident. For instance, transgenic potato plants with altered T6P contents were developed to find out whether T6P has a signaling role in the tubers. The work demonstrated that transgenic potato lines with elevated T6P contents exhibited decreased contents of starch and ATP, and stimulated respiration rate, indicative of high metabolic activity [99]. However, potato lines with lower T6P displayed soluble sugar accumulation, increased hexose phosphates and ATP, no change in starch content, and a strong reduction in tuber yield. The study also showed that the transgenic lines with target genes of SnRK1 that were involved in the promotion of cell proliferation and growth were downregulated, while those involved in inhibiting cell cycle progression were upregulated. The work of Debast et al. [99] also indicated that T6P accumulation in tubers drastically postponed sprouting, while those with reduced T6P sprouted earlier than the wild type; reduced ABA level was correlated with early sprouting of tubers. In agreement, Figueroa and Lunn [42] presented different lines of evidence that the link between growth, development, and carbon status to maintain sucrose levels within an optimal range is maintained. That is, in source leaves, T6P modifies sucrose levels by influencing sucrose synthesis, while in sink organs, T6P controls sucrose consumption. Moreover, the presence of interaction between sugars and plant hormones in metabolic pathways and stress response has also been suggested, which is supported by the result that the application of trehalose upregulated the gene expression responsible for the biosynthesis of ABA and downregulated metabolic genes, which eventually enhanced the ABA content in plants [40,96]. It is, therefore, clear from these findings that trehalose serves as a signaling molecule via controlling the ABA anabolic pathway, which results in ABA accumulation and enhances the salinity tolerance of tomato plants. The results provide greater insights into understanding the salinity tolerance mechanism of plants and have important significance for promoting salinity resilience and crop yield. A recent work by Wang et al. [94] also illustrated that trehalose and ABA have synergistic impacts on root growth and stomatal closure since ABA has been indicated to stimulate the expression of TPP and hence raise trehalose contents. Recently, Kerbler et al. [24] and Blanford et al. [75] similarly report T6P as an essential signaling molecule linking carbon metabolism to plant growth and development. The latter authors also indicated that TPPs play pivotal regulatory roles in regulating endogenous levels of T6P and trehalose, and T6P signaling, as well as in integrating environmental signals with plant metabolism. In conclusion, trehalose, its precursor T6P, and enzymes (TPP, TPS) of its biosynthesis pathway have a crucial role as signaling molecules, and they greatly function in the regulation of sugar metabolism as well as ABA levels, which strongly suggests crosstalk between them resulting in a link between stress, metabolism, and development. However, these metabolites’ roles in signaling, regulation of metabolism, hormone levels, and the development of plants during stress need further molecular elucidation and exploration.
Induction of Ion Homeostasis, Accumulation of Osmolytes, and Secondary Metabolites
The capacity of plants to retain ion homeostasis in response to high salinity is a pivotal trait in determining plant adaptation to saline soil [14,25]. Trehalose improves Catharanthus roseus salinity resistance due to trehalose’s ability to reduce the Na+/K+ ratio in salinity-stressed plants [73]. In this work, trehalose treatment increased sugar accumulation, net photosynthesis rate, transpiration rate, and stomatal conductance, suggesting that the trehalose-enhancement of sugars (i.e., trehalose, sucrose, fructose, glucose, soluble sugars, free amino acids, and alkaloids) allowed the salinity-stressed plants to maintain a status of water relations suitable for growth and photosynthesis under high saline situations. It can also be concluded from this research that trehalose efficiently induces osmolyte accumulation, which plays a part in the salinity tolerance mechanism. Further evidence supporting trehalose’s role in accelerating ion balance under saline conditions and hence enhancing crop resilience is that transgenes with increased trehalose content can maintain a greater selectivity for K+ over Na+ uptake in rice roots, which improved the rice salinity tolerance when grown under high salinity [58]. Another study by Garcia et al. [100] also illustrated that exogenous trehalose remarkably decreased Na+ accumulation triggered by salinity, suppressed salT (an osmotically regulated gene) expression, and enhanced growth in rice. Here, the proposed mechanism underpinning rice salinity tolerance by trehalose application is trehalose-induced Na+ exclusion from root cells by protecting membrane transport proteins in response to salinity stress. Furthermore, exogenous trehalose overcomes salinity-induced reduction in growth and reproduction of Arabidopsis under high salinity by ionic homeostasis maintenance, i.e., retained higher K+ and K+/Na+ ratios in the leaf and stem of the inflorescence [69]. In this investigation, it is also illustrated that trehalose was capable of restricting Na+ transport from leaves to the inflorescence stem. Phan and Van Dijck [48] similarly indicated that the mechanism by which trehalose can protect plants against stress is through its protection of proteins, membrane integrity, and ion pumps, which help to prevent Na+ uptake into chloroplasts. Taken together, trehalose has been found to regulate the K+/Na+ ratio and modify osmolyte metabolism and accumulation under saline conditions. However, the implicated molecular mechanisms in how trehalose regulates ionic balance under salinity imposition need further research and exploration.
In response to saline conditions, a strategy has emerged in plants to adjust the osmotic potential of the cell by overproduction of compatible solutes such as sugars, proline, and glycinebetaine [25,55]. Compatible solutes are important osmotic regulators as they can significantly lower the cell osmotic potential, providing the required water potential gradient for water absorption and hence turgidity retention, and they also stabilize cellular proteins and other cell components against the harmful effects of high salinity [59]. Further evidence indicating trehalose’s implication in accelerating osmolyte levels under saline conditions is provided in this section. For instance, exogenous trehalose increases the soluble sugar content in tomato seedlings by upregulating the expression of genes encoding enzymes responsible for their biosynthesis [40]. Additionally, trehalose also alters soluble sugar accumulation and distribution by upregulating the expression of genes related to sugar transporters [40]. In quinoa and rice plants, trehalose supply stimulates the production of compatible osmolytes such as proline, trehalose, total soluble sugars, and free amino acids under salinity stress [29,60], indicating trehalose’s capacity to upregulate osmolyte and secondary metabolite accumulation that are involved in osmoregulation and protective functions as well. Also, trehalose-induced compatible osmolytes and secondary metabolites production in maize [76], cucumber [80], Indian mustard [101,102], rice [31], and tomato [21] in response to high salinity has been documented. Externally applied trehalose promoted starch accumulation, soluble sugar, and lignin in the rose petals during drought stress, alleviating drought impacts and enhancing tolerance [68]. Collectively, these solutes participate in cell osmotic balance regulation, ROS scavenging as nonenzymatic antioxidants, as well as cellular structure integrity protection, and thus contribute to crop salinity stress alleviation and tolerance. It can also be inferred from these works that trehalose upregulates genes encoding enzymes that catalyze these solutes. However, the mechanism by which trehalose activates stress-responsive genes/proteins in response to saline conditions needs to be deciphered.
3.4. Trehalose as a Macromolecular Protector
An important strategy plants adopt to cope with various abiotic stresses is the biosynthesis and accumulation of excessive compatible solutes, which contribute to osmotic adjustment for maintaining continued water absorption. These osmolytes are called compatible osmolytes as they, even at high concentrations in the cytoplasm, are compatible with the cellular metabolism and function to balance the external lower water potential of the soil. Except in certain resurrection plants (e.g., Selaginella lepidophylla) and specific organs upon stress exposure, trehalose is hardly detectable in most plants, and therefore it is not considered a solute that contributes to osmotic adjustment under stress conditions [42,77]. It is most likely that other osmolytes take over this function, whereas trehalose serves as a cellular constituent protectant and signaling molecule under salinity imposition. Trehalose, hence, is reported to be a remarkably effective protectant due to its roles in stabilizing the cellular membranes, proteins, and protein complexes, and cell components under abiotic stresses [42,58,64,71]. Reportedly, these special functions of trehalose are related to its physicochemical properties because trehalose can protect protein and membrane structure deformation by replacing the hydrogen bonds of water molecules that form the water film surrounding these cellular structures as well as creating a gel phase in the cells exposed to dehydration, thereby protecting cells against drought and high salinity [42,52]. These protective properties of trehalose are reported to be superior to those of other sugars, such as sucrose, making it an ideal stress protectant [42,59,69]. In support, an in vitro experiment by Magazù et al. [103] illustrated the effectiveness of trehalose in protecting proteins from degradation and even outcompetes sucrose. In addition, lipid-based cells maintain their status of fluidity under stress conditions with a trehalose stabilizing role, preventing membrane leakage and fusion [19,56]. Abdallah et al. [60] also suggested that trehalose’s effective mitigating impact against salinity stress in two rice cultivars was likely due to its role in stabilizing antioxidant enzymes and proteins, as well as protecting other cellular structures, which suggests the protective function of trehalose macromolecules under stress. Additionally, interestingly, trehalose was one crucial alternative osmoprotectant to proline in rice cultivars exposed to NaCl stress [100]. In this investigation, while proline caused inhibitory effects, trehalose reduced Na+ accumulation, loss of chlorophyll, and enhanced growth under salinity. The protective role of trehalose is explained by its excellent capacity to stabilize lipid bilayer and enzyme functioning, thus preserving their integrity and role under salinity stress [100]. Thus, the study showed trehalose importance for rice more than proline under saline stress. Consistently, proline proposed functions in plants under salinity stress are argued by Mansour and Ali [104] as proline accumulation showed a negative correlation with salinity tolerance, and proline functions are not always confirmed under saline imposition in many plants.
Trehalose exogenous application also provides beneficial protection of the cellular macromolecules in plants facing salinity stress. For instance, trehalose via its direct interaction with cis double bonds of the cellular macromolecules has been indicated to protect unsaturated fatty acids against oxidative stress, preventing protein aggregation, and limiting the acetylation of lysine amino groups [105]. Another mechanism by which trehalose shows its capability to protect the cellular macromolecules and membranes is achieved through trehalose’s direct suppression of ROS damage effects, as trehalose itself is proposed as an antioxidant effector or indirectly activates the antioxidant defense systems that scavenge ROS [71,93]. The work of Luo et al. [93] demonstrated that trehalose plays a direct role in scavenging ROS (O2−, H2O2) in wheat under heat stress, which is possibly the case under saline conditions. Therefore, minimizing cellular ROS levels induced by trehalose protects membranes, proteins, and nucleic acids from oxidative damage triggered by saline imposition. Further, Fordil and Khan [72] reported trehalose alleviation of the harmful impacts of NaCl stress on ten Pakistani wheat varieties through trehalose’s capacity to act as an osmoprotective agent, reducing the damage caused by salinity. Trehalose’s protective function is also reported by Figueroa and Lunn [42] since trehalose accumulation in chloroplasts preserves their functionality under salinity stress, suggestive of trehalose stabilizing impact on the chloroplast architecture. Similarly, trehalose treatment increased the net photosynthesis rate and enhanced Catharanthus roseus salinity tolerance, which was partly attributed to trehalose’s protective effects on the photosynthetic apparatus [73]. Moreover, trehalose priming has been proposed to protect the macromolecular structures of maize plants against the destabilizing effect of salinity stress, which improves maize performance under high salinity [106]. Based on the above evidence, it is therefore obvious that trehalose effectively protects the cellular macromolecules against the hazardous effects of salinity imposition, thereby improving salinity resilience.
4. Genetic Engineering of Trehalose-Encoding Genes (TPS and TPP)
To develop salinity tolerance in crop plants, engineering a trehalose biosynthetic pathway is one of the most promising approaches because trehalose plays a chief role in regulating various tolerance mechanisms [31,40,64,90,107]. The development of transgenic lines with trehalose biosynthesis genes provides numerous possibilities for enhancing crop salinity resilience and thus may improve yield production. These transgenes also showed favorably altered nutritional metabolites of seeds and tubers. However, some transgenic plants overexpressing trehalose biosynthesis genes and over accumulating trehalose have been reported to be adversely affected [41,108]. Therefore, trehalose accumulation by biosynthesis gene overexpression should be precisely controlled to optimal subcellular levels to avoid such adverse effects on plants. In addition, trehalose naturally accumulates in a broad variation among plants and genotypes, indicating that trehalose’s optimal level of accumulation is species/genotype dependent. This also needs to be considered in transgenic approaches seeking to modify biosynthesis genes (TPS, TPP) to improve salinity tolerance. Several options are proposed to avoid such phenotypic alterations in response to engineering trehalose biosynthesis genes: a) a tight regulation between biosynthesis and degradation of T6P and/or trehalose at the cellular level should be balanced as an imbalance in the level of T6P and/or trehalose has been reported to result in these aberrant phenotypes [70], b) overexpression of trehalase is suggested to regulate trehalose metabolism and avoid trehalose overaccumulation [26], and c) regulation of the introduced genes by using stress-inducible promoters and limitation of TPS and TPP expression to certain subcellular levels [42]. These options should be considered in gene manipulation techniques to enhance trehalose production in crop plants without undesirable effects. The main results of various investigations that indicate the importance of trehalose biosynthesis gene transformation in modulating transgenic crop plant responses to saline conditions are reviewed and summarized in Table 1.
5. Trehalose Exogenous Application
The approach of trehalose exogenous application is based on the fact that when plants are overwhelmed by salinity stress and the perturbation exceeds the plant’s ability to deal with it or emerges from different sources in addition to the fact that trehalose is detected in very low level in plants, an exogenous supply of trehalose or T6P could be an efficient strategy to improve several crop plant performance under saline soil [26,28,29,40,47,68,72,123]. Under stressful saline conditions, exogenous trehalose could improve crop salinity tolerance, which most likely functions as a modulator of the biological processes implicated in crop salinity stress responses and adaptation. Trehalose external applications have been reported to have the advantages of being available, absorbable, transportable, cheap, and non-toxic [124]. A tabulated review of the responses of crop plants contrasting in salinity resistance to exogenously applied trehalose or T6P under saline conditions is given in Table 2. However, these trehalose-induced mitigative effects are not unique and similar but depend on various factors. Therefore, conclusions are drawn based on the results presented in Table 2 for consideration when trehalose is supplied externally. These include a) the trehalose impacts seem to rely on the concentration used, and they are species-specific. It is, therefore, recommended to justify trehalose concentration and other factors regulating its efficacy for each plant species/genotype, b) roots efficiently absorb trehalose which is easily transported to the shoot and leaves to trigger several adaptive mechanisms functioning as major defensive responses to saline stress, c) internal trehalose content depends on plant species/genotype, different providing approaches, tissue and cell type, and plant developmental stage at which trehalose applied. Therefore, these conditions should be considered when interpreting the results and drawing conclusions, d) T6P and trehalose foliar spray showed functional differences in rose plants under drought stress, as the response of gene expression to T6P or trehalose was different under stress. This also needs to be explored under saline imposition, e) trehalose toxicity to plants, when applied in high concentrations, depends on a link between T6P, SnRK1, bZIP11, sucrose, and starch, and f) The salinity’s drastic impacts on the expression of the trehalose pathway genes and T6P showed remarkable differences between source and sink tissues through T6P impact on SnRK1 activity. This impact occurs through differential effects on SnRK1 marker genes between source and sink tissues, possibly suggesting unrelated functions.
6. Conclusions and Perspectives
The current review presents various lines of evidence to raise awareness of the roles of trehalose, its precursor T6P, and biosynthetic enzymes. Figure 4 summarizes possible mechanisms by which trehalose and its intermediates promote crop plants’ salinity tolerance. Additionally, the engineering of trehalose biosynthetic genes and the exogenous application of trehalose or T6P are discussed to assist researchers in improving crop performance and yield potential under saline conditions. This includes a) As a signaling molecule, trehalose or T6P promotes various events and signaling pathways in response to salinity stress, b) Trehalose or T6P functions as a modulator of genes implicated in producing stress-responsive proteins that detoxify and protect crop species from the devastating impacts of salinity stress, c) Trehalose metabolism might watch the redox status of the cellular compartments to modulate diverse physiological processes that render salinity stress responses, d) Trehalose and most probably trehalose-induced accumulation of osmolytes act as an energy and carbon source as well as cellular structures protectant, e) Trehalose itself is an antioxidant (although needs elucidation) and upregulates other antioxidant defense system and f) Trehalose induces ion homeostasis, accumulation of osmolytes and secondary metabolites. In addition, exogenously applied trehalose, to compensate for the insufficient internal trehalose level, is a promising strategy to elevate internal trehalose levels and improve tolerance to saline soil in crop plants. Further, the genetic engineering of trehalose biosynthesis genes in crop plants can be a potential option to enhance their tolerance to saline conditions.
Future perspectives and research should focus on and include a) Evidence indicates that constitutive overexpression of the trehalose biosynthetic genes led to the growth abnormalities, we therefore should adopt the controlled upregulation of trehalose biosynthesis to develop transgenes with improved stress tolerance, but without phenotypic alterations, b) Although studies have efficiently manipulated trehalose content to improve crop tolerance and productivity, trehalose roles in tolerance to multiple stresses concerning plant growth, structure, and metabolism still need huge research and exploration, c) Further intensive -omics research would be an interesting approach for providing useful information concerning trehalose metabolism during developmental stages and under saline conditions, as well as for future trehalose genetic engineering of biosynthesis genes to promote salinity tolerance and yield in crop plants, d) Under natural saline habitats, trehalose accumulation, function, engineering, and pretreatment in crops are rare, and therefore, future research should focus on such evaluations to support our claims of the laboratory controlled results, e) Trehalose response to saline situations greatly depends upon species/genotype, salinity intensity and type, plant organ, trehalose exogenous concentration, and developmental stage at which trehalose is applied. Therefore, these prerequisites should be taken into consideration to reach a plausible conclusion before recommending trehalose for agricultural practices, f) In response to escalating world population and climate change pressures, our ultimate goal is to enhance crop productivity required for global food security, either under favorable or poor conditions alike. However, simultaneously stimulating both crop yield and tolerance is challenging, as the mechanisms determining stress tolerance and yield are typically inversely related. In this respect, we believe that the protocol by Paul et al. [129] to appropriately target T6P in certain cells, tissues, and developmental stages should be given serious consideration and adopted to enhance both productivity and stress tolerance. Therefore, we recommend that future research focus on appropriately modifying T6P levels to meet specific targets, aiming to simultaneously stimulate yield potential and stress tolerance. In this review, we lay the groundwork for future research to gain further insights into the possible molecular mechanisms behind trehalose and its intermediates’ functions in salinity tolerance, and to aid in their engineering for improving crop tolerance and yield in saline soils.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We appreciate the assistance of Nayera Mansour in preparing the manuscript figures.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- FAO (2015) Food and Agriculture Organization of the United Nations and ITPS, Intergovernmental Technical Panel on Soils Status of the world’s soil resources (SWSR), main report.
- Hasanuzzaman, M.; Raihan, R.H.; Masud, A.A.C.; Rahman, K.; Nowroz, F.; Rahman, M.; Nahar, K.; Fujita, M. Regulation of Reactive Oxygen Species and Antioxidant Defense in Plants under Salinity. Int. J. Mol. Sci. 2021, 22, 9326. [Google Scholar] [CrossRef]
- Kesawat, M.S.; Satheesh, N.; Kherawat, B.S.; Kumar, A.; Kim, H.-U.; Chung, S.-M.; Kumar, M. Regulation of Reactive Oxygen Species during Salt Stress in Plants and Their Crosstalk with Other Signaling Molecules—Current Perspectives and Future Directions. Plants 2023, 12, 864. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of Salinity Tolerance in Plants: Physiological, Biochemical, and Molecular Characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.A.; Sarkhosh, A.; Khan, N.; Balal, R.M.; Ali, S.; Rossi, L.; Gómez, C.; Mattson, N.; Nasim, W.; Garcia-Sanchez, F. Insights into the Physiological and Biochemical Impacts of Salt Stress on Plant Growth and Development. Agronomy 2020, 10, 938. [Google Scholar] [CrossRef]
- Atta, K.; Mondal, S.; Gorai, S.; Singh, A.P.; Kumari, A.; Ghosh, T.; Roy, A.; Hembram, S.; Gaikwad, D.J.; Mondal, S.; et al. Impacts of salinity stress on crop plants: improving salt tolerance through genetic and molecular dissection. Front. Plant Sci. 2023, 14, 1241736. [Google Scholar] [CrossRef] [PubMed]
- Harsha SG, Geetha GA, Manjugouda IP, Shilpashree VM, GuheA, Singh TH, Laxman RH, Satisha GC, Prathibha MD, Shivashankara KS (2025) Unraveling salinity stress tolerance: Contrasting morpho-physiological, biochemical, and ionic responses in diverse brinjal (Solanum melongena L.) genotypes. Plant Physiol Biochem 222:109666.
- Mansour MMF, Salama KHA (2019) Cellular mechanisms of plant salt tolerance. In: Giri B, Varma A (eds) Microorganisms in Saline Environment: Strategies and Functions, Springer, Switzerland, pp 169-210.
- Zhang H, Zhu J, Gong Z, Zhu J Abiotic stress responses in plants. Nature Rev Genet 2022, 23, 104–119. [CrossRef]
- Mansour, M.M.F. Anthocyanins: Biotechnological targets for enhancing crop tolerance to salinity stress. Sci. Hortic. 2023, 319. [Google Scholar] [CrossRef]
- Sarkar, A.K.; Sadhukhan, S. Imperative role of trehalose metabolism and trehalose-6-phosphate signaling on salt stress responses in plants. Physiol. Plant. 2022, 174, e13647. [Google Scholar] [CrossRef]
- Ibrahim, E.A. Review: Trehalose and its role in plant adaptation to salinity stress. Plant Sci. 2025, 357, 112533. [Google Scholar] [CrossRef]
- Eh, T.-J.; Jiang, Y.; Jiang, M.; Li, J.; Lei, P.; Ji, X.; Kim, H.-I.; Zhao, X.; Meng, F. The role of trehalose metabolism in plant stress tolerance. J. Adv. Res. 2024, 76, 57–72. [Google Scholar] [CrossRef]
- Han, Y.; Liang, A.; Xu, D.; Zhang, Y.; Shi, J.; Li, M.; Liu, T.; Qi, H. Versatile roles of trehalose in plant growth and development and responses to abiotic stress. Veg. Res. 2024, 4. [Google Scholar] [CrossRef]
- Goharrizi, K.; Karami, S.; Hamblin, M.; Momeni, M.; Basaki, T.; Dehnavi, M.; Nazari, M. Transcriptomic and proteomic profile approaches toward drought and salinity stresses. Biol. Plant. 2022, 66, 255–271. [Google Scholar] [CrossRef]
- Zhu, Y.; Ren, Y.; Liu, J.; Liang, W.; Zhang, Y.; Shen, F.; Ling, J.; Zhang, C. New Genes Identified as Modulating Salt Tolerance in Maize Seedlings Using the Combination of Transcriptome Analysis and BSA. Plants 2023, 12, 1331. [Google Scholar] [CrossRef] [PubMed]
- López, M.; Herrera-Cervera, J.A.; Lluch, C.; Tejera, N.A. Trehalose metabolism in root nodules of the model legume Lotus japonicus in response to salt stress. Physiol. Plant. 2006, 128, 701–709. [Google Scholar] [CrossRef]
- Grennan, A.K. The Role of Trehalose Biosynthesis in Plants. Plant Physiol. 2007, 144, 3–5. [Google Scholar] [CrossRef]
- Iturriaga, G.; Suárez, R.; Nova-Franco, B. Trehalose Metabolism: From Osmoprotection to Signaling. Int. J. Mol. Sci. 2009, 10, 3793–3810. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Chen, J.; Finnegan, P.M.; Younis, A.; Nafees, M.; Zorrig, W.; Ben Hamed, K. Application of Trehalose and Salicylic Acid Mitigates Drought Stress in Sweet Basil and Improves Plant Growth. Plants 2021, 10, 1078. [Google Scholar] [CrossRef]
- Yang, Y.; Yao, Y.; Li, J.; Zhang, J.; Zhang, X.; Hu, L.; Ding, D.; Bakpa, E.P.; Xie, J. Trehalose Alleviated Salt Stress in Tomato by Regulating ROS Metabolism, Photosynthesis, Osmolyte Synthesis, and Trehalose Metabolic Pathways. Front. Plant Sci. 2022, 13, 772948. [Google Scholar] [CrossRef]
- Yang, Y.; Xie, J.; Li, J.; Zhang, J.; Zhang, X.; Yao, Y.; Wang, C.; Niu, T.; Bakpa, E.P. Trehalose alleviates salt tolerance by improving photosynthetic performance and maintaining mineral ion homeostasis in tomato plants. Front. Plant Sci. 2022, 13, 974507. [Google Scholar] [CrossRef]
- Zhang, Z.; Sun, M.; Gao, Y.; Luo, Y. Exogenous trehalose differently improves photosynthetic carbon assimilation capacities in maize and wheat under heat stress. J. Plant Interactions 2022, 17, 361–370. [Google Scholar] [CrossRef]
- Kerbler, S.M.; Armijos-Jaramillo, V.; Lunn, J.E.; Vicente, R. The trehalose 6-phosphate phosphatase family in plants. Physiol. Plant. 2023, 175, e14096. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Mohammad, F.; Khan, M.N.; Corpas, F.J. Trehalose mitigates sodium chloride toxicity by improving ion homeostasis, membrane stability, and antioxidant defense system in Indian mustard. Plant Stress 2025, 15. [Google Scholar] [CrossRef]
- Delorge, I.; Janiak, M.; Carpentier, S.; Van Dijck, P. Fine tuning of trehalose biosynthesis and hydrolysis as novel tools for the generation of abiotic stress tolerant plants. Front. Plant Sci. 2014, 5, 147. [Google Scholar] [CrossRef] [PubMed]
- Mohsin, S.M.; Fardus, J.; Nagata, A.; Tamano, N.; Mitani, H.; Fujita, M. Comparative Study of Trehalose and Trehalose 6-Phosphate to Improve Antioxidant Defense Mechanisms in Wheat and Mustard Seedlings under Salt and Water Deficit Stresses. Stresses 2022, 2, 336–354. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Hossain, M.A.; Fujita, M. Trehalose pretreatment induces salt tolerance in rice (Oryza sativa L.) seedlings: oxidative damage and co-induction of antioxidant defense and glyoxalase systems. Protoplasma 2014, 252, 461–475. [Google Scholar] [CrossRef]
- Abdallah, M.M.-S.; El Sebai, T.N.; Ramadan, A.A.E.-M.; El-Bassiouny, H.M.S. Physiological and biochemical role of proline, trehalose, and compost on enhancing salinity tolerance of quinoa plant. Bull. Natl. Res. Cent. 2020, 44, 1–13. [Google Scholar] [CrossRef]
- Abid A, Shahbaz M Influence of exogenously applied trehalose on morphological and physiological traits of flax (Linum usitissimum) plant under water shortage conditions. Pak J Bot 2022, 59, 435–448.
- Joshi, R.; Sahoo, K.K.; Singh, A.K.; Anwar, K.; Pundir, P.; Gautam, R.K.; Krishnamurthy, S.L.; Sopory, S.K.; Pareek, A.; Singla-Pareek, S.L. Enhancing trehalose biosynthesis improves yield potential in marker-free transgenic rice under drought, saline, and sodic conditions. J. Exp. Bot. 2019, 71, 653–668. [Google Scholar] [CrossRef]
- Kumar, T.; Tiwari, N.; Bharadwaj, C.; Roorkiwal, M.; Reddy, S.P.P.; Patil, B.S.; Kumar, S.; Hamwieh, A.; Vinutha, T.; Bindra, S.; et al. A comprehensive analysis of Trehalose-6-phosphate synthase (TPS) gene for salinity tolerance in chickpea (Cicer arietinum L.). Sci. Rep. 2022, 12, 1–12. [Google Scholar] [CrossRef]
- Xu, P.; Ma, Y.; Wang, S.; Liu, M.; Zhu, T.; He, H.; Ke, J.; You, C.; Wu, L. Exogenous Trehalose can Reduce the Loss of Rice Yields Under High Temperature Stress at the Flowering Stage by Improving the Carbohydrate Metabolism and Photosynthesis Capacity of Flag Leaves. J. Plant Growth Regul. 2024, 44, 2892–2909. [Google Scholar] [CrossRef]
- Mohanan, A.; Kodigudla, A.; Raman, D.R.; Bakka, K.; Challabathula, D. Trehalose accumulation enhances drought tolerance by modulating photosynthesis and ROS-antioxidant balance in drought sensitive and tolerant rice cultivars. Physiol. Mol. Biol. Plants 2023, 29, 2035–2049. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.J.; Jhurreea, D.; Zhang, Y.; Primavesi, L.F.; Delatte, T.; Schluepmann, H.; Wingler, A. Up-regulation of biosynthetic processes associated with growth by trehalose 6-phosphate. Plant Signal. Behav. 2010, 5, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Maicas, S.; Guirao-Abad, J.P.; Argüelles, J.-C. Yeast trehalases: Two enzymes, one catalytic mission. Biochim. et Biophys. Acta (BBA) - Gen. Subj. 2016, 1860, 2249–2254. [Google Scholar] [CrossRef]
- Tang, B.; Wang, S.; Wang, S.-G.; Wang, H.-J.; Zhang, J.-Y.; Cui, S.-Y. Invertebrate Trehalose-6-Phosphate Synthase Gene: Genetic Architecture, Biochemistry, Physiological Function, and Potential Applications. Front. Physiol. 2018, 9, 30. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.L.; Nick, F.; Adhitama, N.; Fields, P.D.; Stillman, J.H.; Kato, Y.; Watanabe, H.; Ebert, D. Trehalose mediates salinity-stress tolerance in natural populations of a freshwater crustacean. Curr. Biol. 2024, 34, 4160–4169.e7. [Google Scholar] [CrossRef]
- Cha-um S, Rai V, Takabe T (2019) Proline, glycinebetaine, and trehalose uptake and inter-organ transport in plants under stress. In: Hossain MA, Kumar V, Burritt D, Fujita M, Makela PS (eds) Osmoprotectant-Mediated Abiotic Stress Tolerance in Plants, Springer Nature, Switzerland, pp 201-223.
- Feng, Y.; Chen, X.; He, Y.; Kou, X.; Xue, Z. Effects of Exogenous Trehalose on the Metabolism of Sugar and Abscisic Acid in Tomato Seedlings Under Salt Stress. Trans. Tianjin Univ. 2019, 25, 451–471. [Google Scholar] [CrossRef]
- John R, Raja V, Ahmad M, Majeed U, Ahmad S, Yaqoob U, Kaul T (2017) Trehalose: metabolism and role in stress signaling in plants. In: Sarwat M, Ahmad A, Abdin MZ, Ibrahim MM (eds) Stress Signaling in Plants: Genomics and Proteomics Perspective, Volume 2, Springer, Switzerland, pp 261-275.
- Figueroa, C.M.; Lunn, J.E. A Tale of Two Sugars: Trehalose 6-Phosphate and Sucrose. Plant Physiol. 2016, 172, 7–27. [Google Scholar] [CrossRef]
- Elbein, A.D.; Pan, Y.T.; Pastuszak, I.; Carroll, D. New insights on trehalose: A multifunctional molecule. Glycobiology 2003, 13, 17R–27R. [Google Scholar] [CrossRef]
- Martins, M.C.M.; Hejazi, M.; Fettke, J.; Steup, M.; Feil, R.; Krause, U.; Arrivault, S.; Vosloh, D.; Figueroa, C.M.; Ivakov, A.; et al. Feedback Inhibition of Starch Degradation in Arabidopsis Leaves Mediated by Trehalose 6-Phosphate. Plant Physiol. 2013, 163, 1142–1163. [Google Scholar] [CrossRef]
- Raza, A.; Bhardwaj, S.; Rahman, A.; García-Caparrós, P.; Habib, M.; Saeed, F.; Charagh, S.; Foyer, C.H.; Siddique, K.H.; Varshney, R.K. Trehalose: A sugar molecule involved in temperature stress management in plants. Crop. J. 2024, 12, 1–16. [Google Scholar] [CrossRef]
- Nawaz, M.; Hassan, M.U.; Chattha, M.U.; Mahmood, A.; Shah, A.N.; Hashem, M.; Alamri, S.; Batool, M.; Rasheed, A.; Thabit, M.A.; et al. Trehalose: a promising osmo-protectant against salinity stress—physiological and molecular mechanisms and future prospective. Mol. Biol. Rep. 2022, 49, 11255–11271. [Google Scholar] [CrossRef]
- Hassan, M.U.; Nawaz, M.; Shah, A.N.; Raza, A.; Barbanti, L.; Skalicky, M.; Hashem, M.; Brestic, M.; Pandey, S.; Alamri, S.; et al. Trehalose: A Key Player in Plant Growth Regulation and Tolerance to Abiotic Stresses. J. Plant Growth Regul. 2022, 42, 4935–4957. [Google Scholar] [CrossRef]
- Phan CHB, Van Dijck P (2019) Biosynthesis and degradation of trehalose and its potential to control plant growth, development, and (a)biotic stress tolerance. In: Hossain MA, Kumar V, Burritt D, Fujita M, Makela PS (eds) Osmoprotectant-Mediated Abiotic Stress Tolerance, Springer Nature, Switzerland, pp 175-199.
- Wang W, Chen Q, Xu S, Liu W, Zhu X, Song C (2020a) Trehalose--6--phosphate phosphatase E modulates ABA--controlled root growth and stomatal movement in Arabidopsis. J Integr Plant Biol 62, 1518–1534. [CrossRef] [PubMed]
- Song, L.; Xu, C.; Zhang, L.; Li, J.; Jiang, L.; Ma, D.; Guo, Z.; Wang, Q.; Wang, X.; Zheng, H. Trehalose along with ABA promotes the salt tolerance of Avicennia marina by regulating Na+ transport. Plant J. 2024, 119, 2349–2362. [Google Scholar] [CrossRef] [PubMed]
- Krasensky, J.; Broyart, C.; Rabanal, F.A.; Jonak, C. The Redox-Sensitive Chloroplast Trehalose-6-Phosphate Phosphatase AtTPPD Regulates Salt Stress Tolerance. Antioxidants Redox Signal. 2014, 21, 1289–1304. [Google Scholar] [CrossRef]
- Avonce, N.; Mendoza-Vargas, A.; Morett, E.; Iturriaga, G. Insights on the evolution of trehalose biosynthesis. BMC Evol. Biol. 2006, 6, 109–109. [Google Scholar] [CrossRef]
- Paul S, Paul S Trehalose induced modifications in the solvation pattern of N-methylacetamide. J Phys Chem B 2014, 118, 1052–1063. [CrossRef]
- Bezrukavnikov, S.; Mashaghi, A.; van Wijk, R.J.; Gu, C.; Yang, L.J.; Gao, Y.Q.; Tans, S.J. Trehalose facilitates DNA melting: a single-molecule optical tweezers study. Soft Matter 2014, 10, 7269–7277. [Google Scholar] [CrossRef]
- Kosar, F.; Akram, N.A.; Sadiq, M.; Al-Qurainy, F.; Ashraf, M. Trehalose: A Key Organic Osmolyte Effectively Involved in Plant Abiotic Stress Tolerance. J. Plant Growth Regul. 2018, 38, 606–618. [Google Scholar] [CrossRef]
- Crowe, J.H.; Crowe, L.M.; Chapman, D. Preservation of Membranes in Anhydrobiotic Organisms: The Role of Trehalose. Science 1984, 223, 701–703. [Google Scholar] [CrossRef]
- Olgenblum, G.I.; Sapir, L.; Harries, D. Properties of Aqueous Trehalose Mixtures: Glass Transition and Hydrogen Bonding. J. Chem. Theory Comput. 2020, 16, 1249–1262. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.K.; Kim, J.-K.; Owens, T.G.; Ranwala, A.P.; Choi, Y.D.; Kochian, L.V.; Wu, R.J. Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses. Proc. Natl. Acad. Sci. 2002, 99, 15898–15903. [Google Scholar] [CrossRef] [PubMed]
- Iordachescu M, Imai R (2011) Trehalose and abiotic stress in biological systems. In: Shanker A (ed) Abiotic Stress in Plants – Mechanisms and Adaptations, In Tech Open, Croatia, pp 215-234.
- Abdallah, M.-S.; Abdelgawad, Z.; El-Bassiouny, H. Alleviation of the adverse effects of salinity stress using trehalose in two rice varieties. South Afr. J. Bot. 2016, 103, 275–282. [Google Scholar] [CrossRef]
- Lu, Y.-P.; Zheng, P.-H.; Zhang, X.-X.; Wang, L.; Li, J.-T.; Zhang, Z.-L.; Xu, J.-R.; Cao, Y.-L.; Xian, J.-A.; Wang, A.-L.; et al. Effects of dietary trehalose on growth, trehalose content, non-specific immunity, gene expression and desiccation resistance of juvenile red claw crayfish (Cherax quadricarinatus). Fish Shellfish. Immunol. 2021, 119, 524–532. [Google Scholar] [CrossRef]
- Liu, J.; Wang, D.; Tong, R.; Ye, S.; Zhao, Y.; Wu, J.; Gan, Y. Engineered Rhizobia with Trehalose-Producing Genes Enhance Peanut Growth Under Salinity Stress. Agronomy 2025, 15, 974. [Google Scholar] [CrossRef]
- Younis, A.A.; Mansour, M.M.F. Hydrogen sulfide priming enhanced salinity tolerance in sunflower by modulating ion hemostasis, cellular redox balance, and gene expression. BMC Plant Biol. 2023, 23, 1–16. [Google Scholar] [CrossRef]
- Sah SK, Kaur G, Wani SH (2016) Metabolic engineering of compatible solute trehalose for abiotic stress tolerance in plants. In: Iqbal N, Nazar R, Khan NA (eds) Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies, Springer, India, pp 83-96.
- Roychoudhury A (2021) Trehalose in osmotic stress tolerance of plants. Bioingene PSJ 2:1-13.
- Zhang, Z.; Sun, M.; Gao, Y.; Luo, Y. Exogenous trehalose differently improves photosynthetic carbon assimilation capacities in maize and wheat under heat stress. J. Plant Interactions 2022, 17, 361–370. [Google Scholar] [CrossRef]
- Sharma P, Kapoor N, Dhiman S, Kour J, Singh A, Sharma A, Bhardwaj R (2023) Role of sugars in regulating physiological and molecular aspects of plants under abiotic stress. In: Sharma A, Pandey S, Bhardwaj R, Zheng B, Tripathi DK (eds) The Role of Growth Regulators and Phytohormones in Overcoming Environmental Stress, Academic Press, London, pp 355-374.
- Bao, M.; Xi, Y.; Wang, R.; Ma, Y.; Bai, M.; Wei, G.; Xu, Y.; Wang, J.; Feng, L. Trehalose signaling regulates metabolites associated with the quality of rose flowers under drought stress. Environ. Exp. Bot. 2024, 224. [Google Scholar] [CrossRef]
- Yang, L.; Zhao, X.; Zhu, H.; Paul, M.; Zu, Y.; Tang, Z. Exogenous trehalose largely alleviates ionic unbalance, ROS burst, and PCD occurrence induced by high salinity in Arabidopsis seedlings. Front. Plant Sci. 2014, 5, 570–570. [Google Scholar] [CrossRef]
- Bánfalvi Z )2019(Transgenic plants overexpressing trehalose biosynthetic genes and abiotic stress tolerance in plants. In: Hossain MA, Kumar V, Burritt D, Fujita M, Makela PS (eds) Osmoprotectant-Mediated Abiotic Stress Tolerance, Springer Nature, Switzerland, pp 225-239.
- Onwe, R.O.; Onwosi, C.O.; Ezugworie, F.N.; Ekwealor, C.C.; Okonkwo, C.C. Microbial trehalose boosts the ecological fitness of biocontrol agents, the viability of probiotics during long-term storage and plants tolerance to environmental-driven abiotic stress. Sci. Total. Environ. 2022, 806, 150432. [Google Scholar] [CrossRef]
- Fordil B, Khan N (2022) Trehalose treatment alleviates salt toxicity in wheat seedlings: a beneficial research attempt in agriculture sector of Pakistan. Hamdard Medicus 65:34-50.
- Chang, B.; Yang, L.; Cong, W.; Zu, Y.; Tang, Z. The improved resistance to high salinity induced by trehalose is associated with ionic regulation and osmotic adjustment in Catharanthus roseus. Plant Physiol. Biochem. 2014, 77, 140–148. [Google Scholar] [CrossRef]
- Ponnu, J.; Wahl, V.; Schmid, M. Trehalose-6-Phosphate: Connecting Plant Metabolism and Development. Front. Plant Sci. 2011, 2, 15488. [Google Scholar] [CrossRef] [PubMed]
- Blanford, J.; Zhai, Z.; Baer, M.D.; Guo, G.; Liu, H.; Liu, Q.; Raugei, S.; Shanklin, J. Molecular mechanism of trehalose 6-phosphate inhibition of the plant metabolic sensor kinase SnRK1. Sci. Adv. 2024, 10, eadn0895. [Google Scholar] [CrossRef] [PubMed]
- Henry, C.; Bledsoe, S.W.; Griffiths, C.A.; Kollman, A.; Paul, M.J.; Sakr, S.; Lagrimini, L.M. Differential Role for Trehalose Metabolism in Salt-Stressed Maize. Plant Physiol. 2015, 169, 1072–1089. [Google Scholar] [CrossRef] [PubMed]
- Kahraman M, Sevim G, Bor M (2019) The role of proline, glycinebetaine, and trehalose in stress-responsive gene expression. In: Hossain MA, Kumar V, Burritt D, Fujita M, Makela PS (eds) Osmoprotectant-Mediated Abiotic Stress Tolerance in Plants, Springer, Switzerland, pp 241-256.
- Nuccio, M.L.; Wu, J.; Mowers, R.; Zhou, H.-P.; Meghji, M.; Primavesi, L.F.; Paul, M.J.; Chen, X.; Gao, Y.; Haque, E.; et al. Expression of trehalose-6-phosphate phosphatase in maize ears improves yield in well-watered and drought conditions. Nat. Biotechnol. 2015, 33, 862–869. [Google Scholar] [CrossRef]
- Delatte, T.L.; Sedijani, P.; Kondou, Y.; Matsui, M.; de Jong, G.J.; Somsen, G.W.; Wiese-Klinkenberg, A.; Primavesi, L.F.; Paul, M.J.; Schluepmann, H. Growth Arrest by Trehalose-6-Phosphate: An Astonishing Case of Primary Metabolite Control over Growth by Way of the SnRK1 Signaling Pathway. Plant Physiol. 2011, 157, 160–174. [Google Scholar] [CrossRef]
- Huang, P.; Li, C.; Che, P.; Liu, H.; Zhao, Z.; Feng, L.; Liu, X.; Liao, W. Hydrogen Gas Enhanced Seed Germination by Increasing Trehalose Biosynthesis in Cucumber. J. Plant Growth Regul. 2022, 42, 3908–3922. [Google Scholar] [CrossRef]
- Nounjan, N.; Theerakulpisut, P. Effects of exogenous proline and trehalose on physiological responses in rice seedlings during salt-stress and after recovery. Plant, Soil Environ. 2012, 58, 309–315. [Google Scholar] [CrossRef]
- Redillas, M.C.F.R.; Park, S.-H.; Lee, J.W.; Kim, Y.S.; Jeong, J.S.; Jung, H.; Bang, S.W.; Hahn, T.-R.; Kim, J.-K. Accumulation of trehalose increases soluble sugar contents in rice plants conferring tolerance to drought and salt stress. Plant Biotechnol. Rep. 2011, 6, 89–96. [Google Scholar] [CrossRef]
- Vichaiya, T.; Faiyue, B.; Rotarayanont, S.; Uthaibutra, J.; Saengnil, K. Exogenous trehalose alleviates chilling injury of ‘Kim Ju’ guava by modulating soluble sugar and energy metabolisms. Sci. Hortic. 2022, 301. [Google Scholar] [CrossRef]
- Vichaiya, T.; Intarasit, S.; Umnajkitikorn, K.; Jangsutthivorawat, S.; Saengnil, K. Postharvest trehalose application alleviates chilling injuring of cold storage guava through upregulation of SnRK1 and energy charge. Sci. Hortic. 2023, 313. [Google Scholar] [CrossRef]
- Tarek A, Sadak M, Dawood MG (2017) Improving drought tolerance of quinoa plant by foliar treatment of trehalose. Agric Eng Int: CIGR Journal Special issue 245-25.
- Bharti A, Maheshwari HS, Chourasiya D, Prakash A, Sharma MP (2022) Role of trehalose in plant–rhizobia interaction and induced abiotic stress tolerance. In: Sigh HB, Vaishnav A (eds) New and Future Developments in Microbial Biotechnology and Bioengineering - Sustainable Agriculture: Revisiting Green Chemical, Elsevier, London, pp 245-263.
- Sadak, M.S. Physiological role of trehalose on enhancing salinity tolerance of wheat plant. Bull. Natl. Res. Cent. 2019, 43, 53. [Google Scholar] [CrossRef]
- Bae H, Herman E, Baily B, Sicher R Exogenous trehalose alters Arabidopsis transcripts involved in cell wall modification, abiotic stress, nitrogen metabolism, and plant defense. Physiol Plant 2005, 125, 114–126. [CrossRef]
- Bae, H.; Herman, E.; Sicher, R. Exogenous trehalose promotes non-structural carbohydrate accumulation and induces chemical detoxification and stress response proteins in Arabidopsis thaliana grown in liquid culture. Plant Sci. 2005, 168, 1293–1301. [Google Scholar] [CrossRef]
- Jiang, T.; Zhai, H.; Wang, F.-B.; Zhou, H.-N.; Si, Z.-Z.; He, S.-Z.; Liu, Q.-C. Cloning and Characterization of a Salt Tolerance-Associated Gene Encoding Trehalose-6-Phosphate Synthase in Sweetpotato. J. Integr. Agric. 2014, 13, 1651–1661. [Google Scholar] [CrossRef]
- Li, H.-W.; Zang, B.-S.; Deng, X.-W.; Wang, X.-P. Overexpression of the trehalose-6-phosphate synthase gene OsTPS1 enhances abiotic stress tolerance in rice. Planta 2011, 234, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Hathout TA, Samia M, Zinab A, Said E, Al Mokadem A (2015) Using validamycin A for salinity stress mitigation in two rice varieties differing in their salt tolerance level. Egypt J Bot 37:49-68.
- Luo, Y.; Li, W.-M.; Wang, W. Trehalose: Protector of antioxidant enzymes or reactive oxygen species scavenger under heat stress? Environ. Exp. Bot. 2008, 63, 378–384. [Google Scholar] [CrossRef]
- Wang, X.; Wei, Y.; Chen, Y.; Jiang, S.; Xu, F.; Wang, H.; Shao, X. NMR revealed that trehalose enhances sucrose accumulation and alleviates chilling injury in peach fruit. Sci. Hortic. 2022, 303. [Google Scholar] [CrossRef]
- Rohman, M.; Islam, R.; Monsur, M.B.; Amiruzzaman, M.; Fujita, M.; Hasanuzzaman, M. Trehalose Protects Maize Plants from Salt Stress and Phosphorus Deficiency. Plants 2019, 8, 568. [Google Scholar] [CrossRef]
- León, P.; Sheen, J. Sugar and hormone connections. Trends Plant Sci. 2003, 8, 110–116. [Google Scholar] [CrossRef]
- Ibrahim, H.A.; Abdellatif, Y.M. Effect of maltose and trehalose on growth, yield and some biochemical components of wheat plant under water stress. Ann. Agric. Sci. 2016, 61, 267–274. [Google Scholar] [CrossRef]
- Lunn, J.E.; Delorge, I.; Figueroa, C.M.; Van Dijck, P.; Stitt, M. Trehalose metabolism in plants. Plant J. 2014, 79, 544–567. [Google Scholar] [CrossRef]
- Debast, S.; Nunes-Nesi, A.; Hajirezaei, M.R.; Hofmann, J.; Sonnewald, U.; Fernie, A.R.; Börnke, F. Altering Trehalose-6-Phosphate Content in Transgenic Potato Tubers Affects Tuber Growth and Alters Responsiveness to Hormones during Sprouting. Plant Physiol. 2011, 156, 1754–1771. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.B.; Engler, J.; Iyer, S.; Gerats, T.; Van Montagu, M.; Caplan, A.B. Effects of Osmoprotectants upon NaCl Stress in Rice. Plant Physiol. 1997, 115, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Mohammad, F. Modulation of growth, photosynthetic efficiency, leaf biochemistry, cell viability and yield of Indian mustard by the application of trehalose. Sci. Hortic. 2021, 290. [Google Scholar] [CrossRef]
- Islam, S.; Mohammad, F.; Siddiqui, M.H.; Kalaji, H.M. Salicylic acid and trehalose attenuate salt toxicity in Brassica juncea L. by activating the stress defense mechanism. Environ. Pollut. 2023, 326, 121467. [Google Scholar] [CrossRef]
- Magazù, S.; Migliardo, F.; Benedetto, A.; La Torre, R.; Hennet, L. Bio-protective effects of homologous disaccharides on biological macromolecules. Eur. Biophys. J. 2011, 41, 361–367. [Google Scholar] [CrossRef]
- Mansour, M.M.F.; Ali, E.F. Evaluation of proline functions in saline conditions. Phytochemistry 2017, 140, 52–68. [Google Scholar] [CrossRef]
- Kuczyńska-Wiśnik, D.; Stojowska-Swędrzyńska, K.; Laskowska, E. Intracellular Protective Functions and Therapeutical Potential of Trehalose. Molecules 2024, 29, 2088. [Google Scholar] [CrossRef]
- Zeid IM Trehalose as osmoprotectant for maize under salinity-induced stress. Res J Agri Biol Sci 2009, 5, 613–622.
- Ye, N.; Wang, Y.; Yu, H.; Qin, Z.; Zhang, J.; Duan, M.; Liu, L. Abscisic Acid Enhances Trehalose Content via OsTPP3 to Improve Salt Tolerance in Rice Seedlings. Plants 2023, 12, 2665. [Google Scholar] [CrossRef]
- Zhang, S.-Z.; Yang, B.-P.; Feng, C.-L.; Tang, H.-L. Genetic Transformation of Tobacco with the Trehalose Synthase Gene from Grifola frondosa Fr. Enhances the Resistance to Drought and Salt in Tobacco. J. Integr. Plant Biol. 2005, 47, 579–587. [Google Scholar] [CrossRef]
- Abdelgawad ZA, Hathout TA, El-Khallal SM, Said EM, Al-Mokadem AZ (2014) Accumulation of trehalose mediates salt adaptation in rice seedlings. American-Eurasian J Agri Environ Sci 14:1450-1463.
- Jun, S.-S.; Yang, J.Y.; Choi, H.J.; Kim, N.-R.; Park, M.C.; Hong, Y.-N. Altered physiology in trehalose-producing transgenic tobacco plants: Enhanced tolerance to drought and salinity stresses. J. Plant Biol. 2005, 48, 456–466. [Google Scholar] [CrossRef]
- Wang, P.; Lei, X.; Lü, J.; Gao, C. Overexpression of the ThTPS gene enhanced salt and osmotic stress tolerance in Tamarix hispida. J. For. Res. 2020, 33, 299–308. [Google Scholar] [CrossRef]
- Cortina, C.; Culiáñez-Macià, F.A. Tomato abiotic stress enhanced tolerance by trehalose biosynthesis. Plant Sci. 2005, 169, 75–82. [Google Scholar] [CrossRef]
- Ge, L.-F.; Chao, D.-Y.; Shi, M.; Zhu, M.-Z.; Gao, J.-P.; Lin, H.-X. Overexpression of the trehalose-6-phosphate phosphatase gene OsTPP1 confers stress tolerance in rice and results in the activation of stress responsive genes. Planta 2008, 228, 191–201. [Google Scholar] [CrossRef]
- Park, S.H.; Jun, S.-S.; An, G.; Hong, Y.-N.; Park, M.C. A comparative study on the protective role of trehalose and LEA proteins against abiotic stresses in transgenic Chinese cabbage (Brassica campestris) overexpressingCaLEA orotsA. J. Plant Biol. 2003, 46, 277–286. [Google Scholar] [CrossRef]
- Almeida, A.M.; Villalobos, E.; Araújo, S.S.; Leyman, B.; Van Dijck, P.; Alfaro-Cardoso, L.; Fevereiro, P.S.; Torné, J.M.; Santos, D.M. Transformation of tobacco with an Arabidopsis thaliana gene involved in trehalose biosynthesis increases tolerance to several abiotic stresses. Euphytica 2005, 146, 165–176. [Google Scholar] [CrossRef]
- Miranda, J.A.; Avonce, N.; Suárez, R.; Thevelein, J.M.; Van Dijck, P.; Iturriaga, G. A bifunctional TPS–TPP enzyme from yeast confers tolerance to multiple and extreme abiotic-stress conditions in transgenic Arabidopsis. Planta 2007, 226, 1411–1421. [Google Scholar] [CrossRef]
- Zhao, F.; Li, Q.; Weng, M.; Wang, X.; Guo, B.; Wang, L.; Wang, W.; Duan, D.; Wang, B. Cloning of TPS gene from eelgrass species Zostera marina and its functional identification by genetic transformation in rice. Gene 2013, 531, 205–211. [Google Scholar] [CrossRef]
- Vishal B, Krishnamurthy P, Ramamoorthy R, Kumar PP OsTPS8 controls yield-related traits and confers salt stress tolerance in rice by enhancing suberin deposition. New Phytol 2019, 221, 1369–1386. [CrossRef] [PubMed]
- Islam, M.O.; Kato, H.; Shima, S.; Tezuka, D.; Matsui, H.; Imai, R. Functional identification of a rice trehalase gene involved in salt stress tolerance. Gene 2019, 685, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Jang, I.-C.; Oh, S.-J.; Seo, J.-S.; Choi, W.-B.; Song, S.I.; Kim, C.H.; Kim, Y.S.; Seo, H.-S.; Choi, Y.D.; Nahm, B.H.; et al. Expression of a Bifunctional Fusion of the Escherichia coli Genes for Trehalose-6-Phosphate Synthase and Trehalose-6-Phosphate Phosphatase in Transgenic Rice Plants Increases Trehalose Accumulation and Abiotic Stress Tolerance without Stunting Growth. Plant Physiol. 2003, 131, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Suárez, R.; Calderón, C.; Iturriaga, G. Enhanced Tolerance to Multiple Abiotic Stresses in Transgenic Alfalfa Accumulating Trehalose. Crop. Sci. 2009, 49, 1791–1799. [Google Scholar] [CrossRef]
- Shao, W.; Zhang, X.; Zhou, Z.; Ma, Y.; Chu, D.; Wang, L.; Yang, Y.; Du, L.; Du, Y.; Du, J.; et al. Genome- and transcriptome-wide identification of trehalose-6-phosphate phosphatases (TPP) gene family and their expression patterns under abiotic stress and exogenous trehalose in soybean. BMC Plant Biol. 2023, 23, 1–18. [Google Scholar] [CrossRef]
- AL-Zewany RGS, Al-Semmak QHA (2017) Effect of foliar-applied trehalose on wheat (Triticum aestivum L.) under salt stress condition. Karbala J Agri Sci 4:1-18.
- Lin, Y.; Zhang, J.; Gao, W.; Chen, Y.; Li, H.; Lawlor, D.W.; Paul, M.J.; Pan, W. Exogenous trehalose improves growth under limiting nitrogen through upregulation of nitrogen metabolism. BMC Plant Biol. 2017, 17, 247. [Google Scholar] [CrossRef]
- Mohamed HI, Akladious S, El-Beltagi H Mitigation the harmful effect of salt stress on physiological, biochemical and anatomical traits by foliar spray with trehalose on wheat cultivars. Fresenius Environ Bull 2018, 27, 7054–7065.
- Soares, F.A.; Na Chiangmai, P.; Duangkaew, P.; Pootaeng-On, Y. ; Nurhidayati Exogenous trehalose application in rice to mitigate saline stress at the tillering stage. Pesqui. Agropecu. Trop. 2023, 53, e75695. [Google Scholar] [CrossRef]
- Theerakulpisut P, Gunnula W Exogenous sorbitol and trehalose mitigated salt stress damage in salt-sensitive but not salt-tolerant rice seedlings. Asian J Crop Sci 2012, 4, 165–170. [CrossRef]
- Alhudhaibi, A.M.; Ibrahim, M.A.R.; Abd-Elaziz, S.M.S.; Farag, H.R.M.; Elsayed, S.M.; Ibrahim, H.A.; Hossain, A.S.; Alharbi, B.M.; Haouala, F.; Elkelish, A.; et al. Enhancing salt stress tolerance in wheat (Triticum aestivum) seedlings: insights from trehalose and mannitol. BMC Plant Biol. 2024, 24, 1–11. [Google Scholar] [CrossRef]
- Paul, M.J.; Gonzalez-Uriarte, A.; Griffiths, C.A.; Hassani-Pak, K. The Role of Trehalose 6-Phosphate in Crop Yield and Resilience. Plant Physiol. 2018, 177, 12–23. [Google Scholar] [CrossRef]
Figure 1.
Salinity stress harmful impacts (osmotic, ionic, oxidative stresses) result in crop growth, development, and yield reduction.
Figure 1.
Salinity stress harmful impacts (osmotic, ionic, oxidative stresses) result in crop growth, development, and yield reduction.
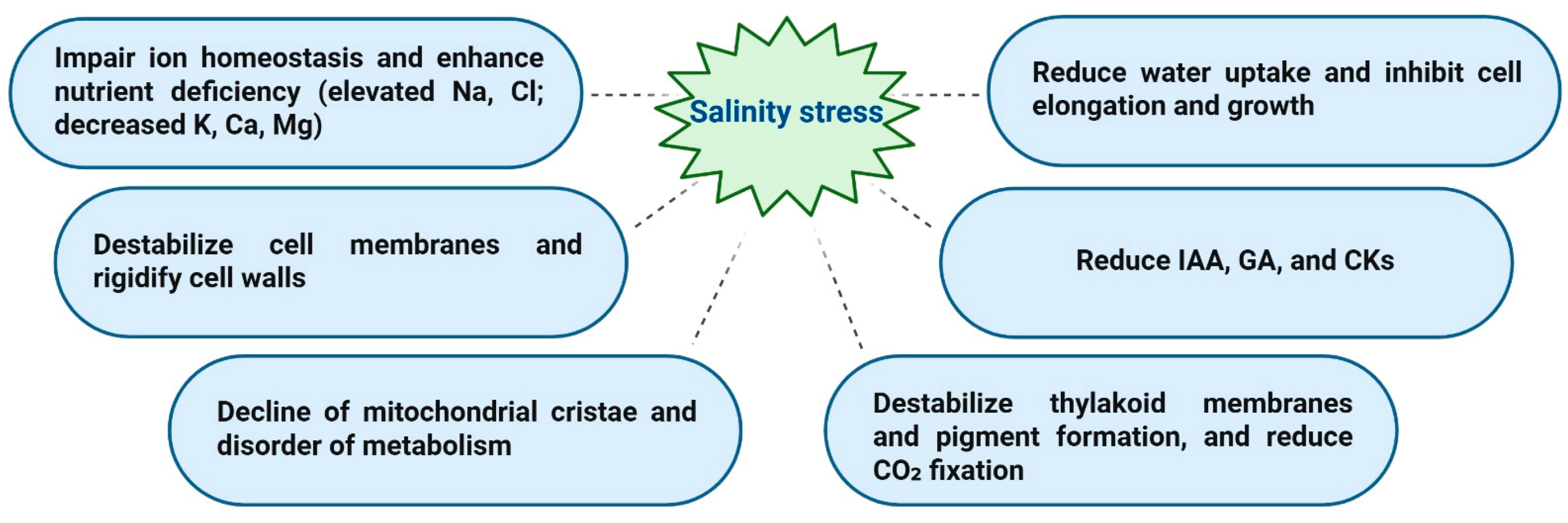
Figure 2.
Structure of trehalose: composed of two glucose units connected by 1,1 α-glycosidic bond.
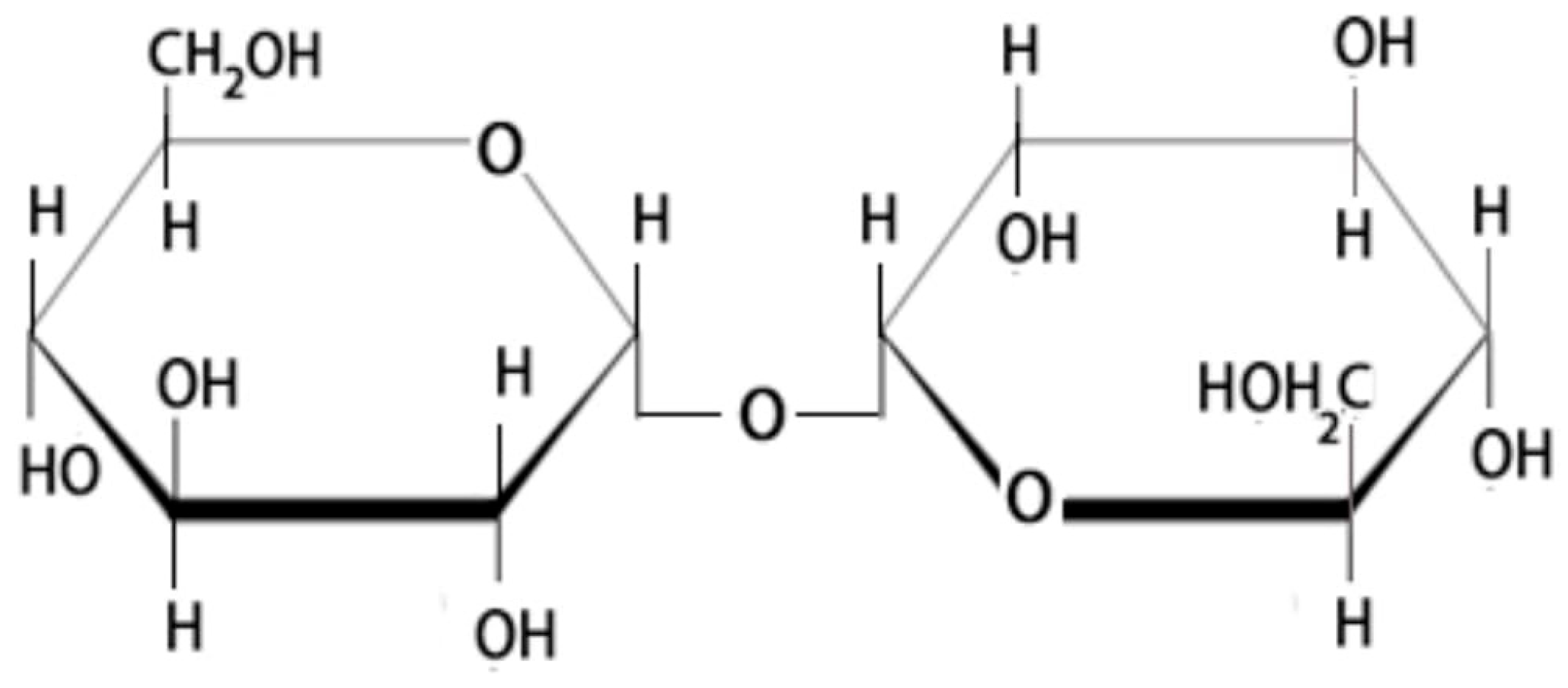
Figure 3.
Trehalose biosynthesis and hydrolysis pathway, and the implication of ABA and ROS as salinity signals contribute to salinity stress tolerance via trehalose biosynthesis enhancement. TPS, trehalose-6-phosphate synthase; TPP, trehalose-6-phosphate phosphatase; UDP, uridine diphosphate. ABA activates ABF2 that binds to TPS/TPP promoters and activates their expression, enhancing trehalose production and salinity tolerance. Trehalose elevates ROS formation that enhances the ABA signaling pathway and strengthens ABA’s effects on processes like stomatal closure and other stress responses, improving salinity tolerance.
Figure 3.
Trehalose biosynthesis and hydrolysis pathway, and the implication of ABA and ROS as salinity signals contribute to salinity stress tolerance via trehalose biosynthesis enhancement. TPS, trehalose-6-phosphate synthase; TPP, trehalose-6-phosphate phosphatase; UDP, uridine diphosphate. ABA activates ABF2 that binds to TPS/TPP promoters and activates their expression, enhancing trehalose production and salinity tolerance. Trehalose elevates ROS formation that enhances the ABA signaling pathway and strengthens ABA’s effects on processes like stomatal closure and other stress responses, improving salinity tolerance.
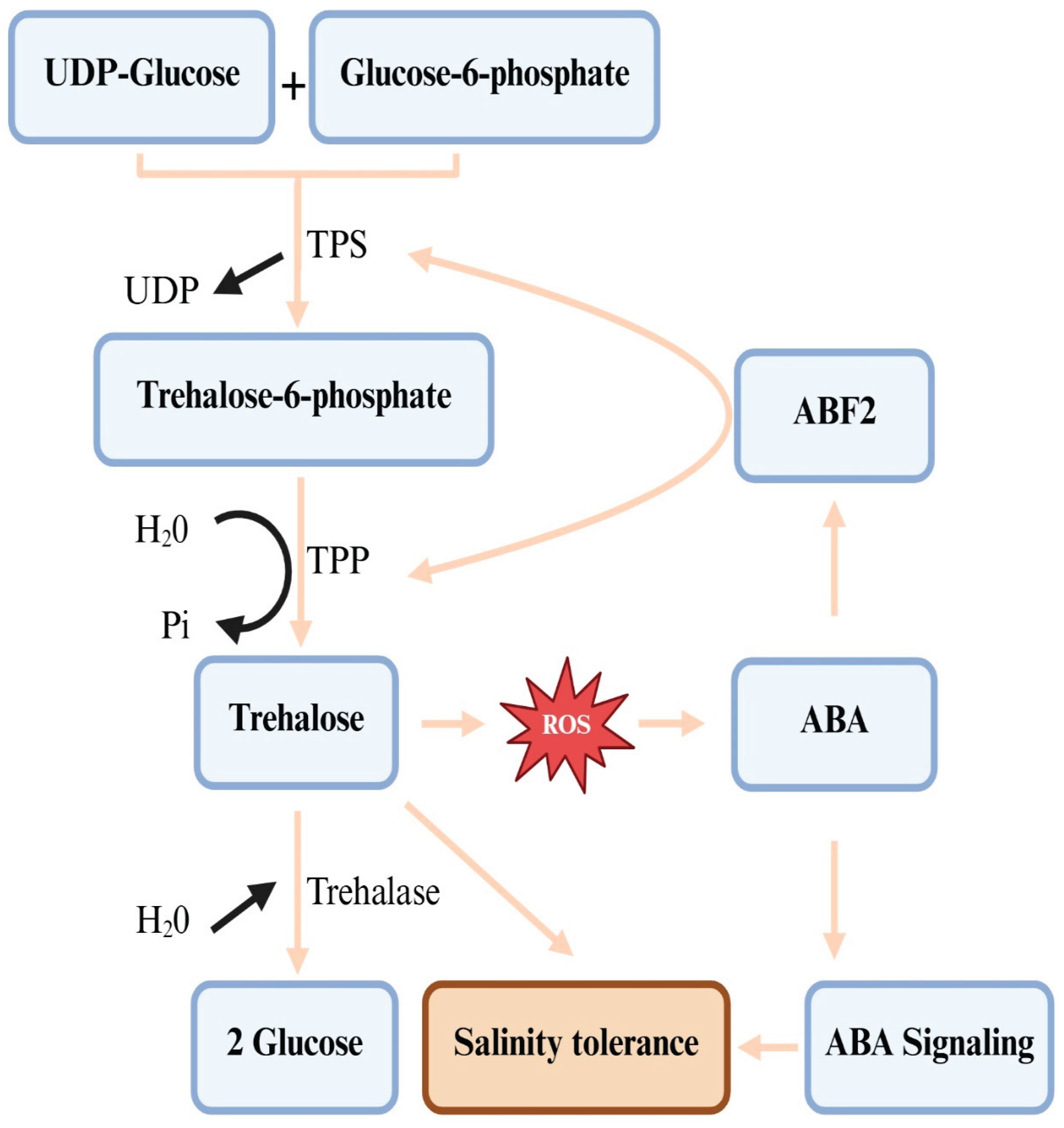
Figure 4.
Roles of trehalose and its intermediates in improving crop plant salinity resilience and yield. Future perspectives are proposed.
Figure 4.
Roles of trehalose and its intermediates in improving crop plant salinity resilience and yield. Future perspectives are proposed.
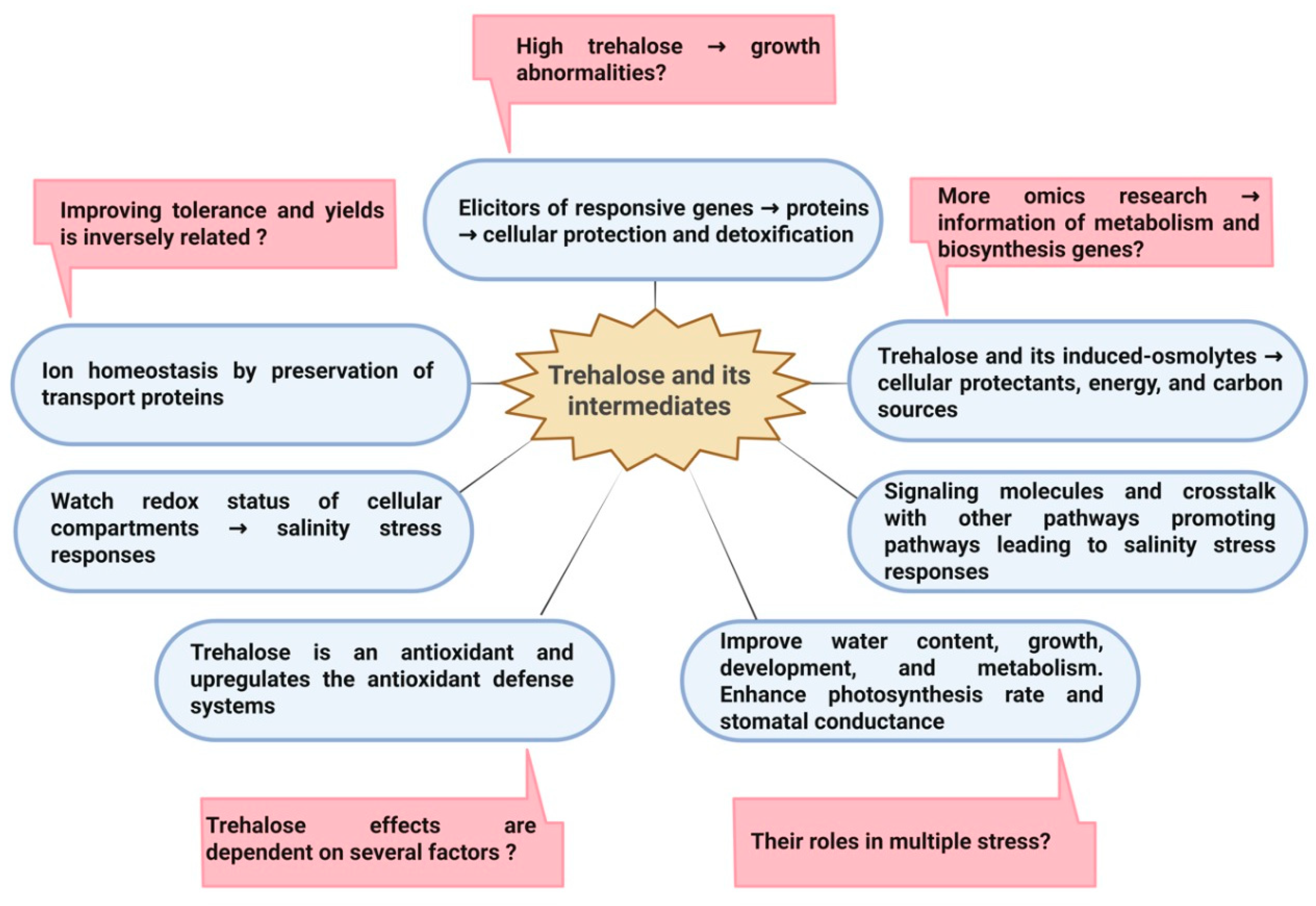

Table 1.
Transgenic crop species expressing trehalose biosynthesis genes that enhance tolerance to saline conditions.
Table 1.
Transgenic crop species expressing trehalose biosynthesis genes that enhance tolerance to saline conditions.
| Gene | Origin | Transgenic crop | Remarks | Reference |
|---|---|---|---|---|
| TPS | Oryza sativa | Oryza sativa | TPS gene was constitutively expressed in the leaves of two rice cultivars, contrasting in salinity tolerance, under stressed and non-stressed conditions, treated and untreated with validamycin A (a competitive inhibitor of trehalase enzyme). Higher transcript amounts were obtained in the presence of validamycin A. Salinity tolerant cultivar showed increased TPS transcript amounts relative to the sensitive one. Upregulation of TPS led to trehalose accumulation and increased gene expression that is responsible for producing osmoprotectants, because trehalose was too low to serve as an osmoprotectant. The study also revealed that the TPS expression pattern was consistent with the increase in TPS activity, confirming that the expression of the TPS gene was upregulated in response to salinity and validamycin treatments, and evidenced its important role in crop salinity tolerance. | [109] |
| TPS, TPP | Solanum Lycopersicon | Solanum Lycopersicon | Salinity upregulated the expression of TPS in tomato, and exogenous trehalose increased the endogenous trehalose, which caused a negative feedback regulation and suppressed the TPP expression. The study indicated that TPS, TPP, and trehalose exhibited differential expressions under salinity stress. The result suggests that the trehalose metabolic pathway could directly affect the salinity tolerance of crop plants. | [40] |
| otsA, otsB | Escherichia coli | Oryza sativa | Overexpression of these genes resulted in transgenic lines exhibiting sustained plant growth, increased trehalose, high soluble sugars, less oxidative damage, elevated capacity for photosynthesis, and ion homeostasis relative to wild type under salinity stress. The study argued against the primary role of trehalose as a compatible solute, as a low level of trehalose was obtained in transgenic rice plants. The work revealed that engineering crops for the overproduction of trehalose resulted in improved salinity tolerance and productivity. | [58] |
| AmTPS9A | Avicennia marina | Arabidopsis thaliana | AmTPS9A-overexpressing Arabidopsis showed increased salinity tolerance by upregulating the expression of genes encoding ion transporters that mediate Na+ efflux from the roots of transgenic Arabidopsis under NaCl treatments. | [50] |
| TPS (otsA) | Escherichia coli | Nicotiana tabacum | Transgenic plants produced small amounts of trehalose, maintained leaf turgidity and fresh weight, had more efficient seed germination, and showed a drastic delay in leaf withering and chlorosis under 250 mM NaCl. It appears that trehalose did not participate in osmotic adjustment but rather may have had a protective or signaling role. | [110] |
| TPS | Tamarix hispida | Tamarix hispida | ThTPS overexpression was induced by salinity treatment, which promoted the biosynthesis of trehalose, decreased the accumulation of O2− and H2O2, and thus improved the T. hispida salinity resistance, suggestive of enhancing the antioxidant defense system by trehalose under salinity. | [111] |
| TPSP | Escherichia coli | Oryza sativa | TPSP transgenic plants retained higher yield, RWC, chlorophyll content, trehalose content, K+/Na+ ratio, stomatal conductance, efficient photosynthesis, and rice seed nutritional levels compared with the wild type under salinity imposition. This finding confirms that trehalose acts as a signaling molecule that alters other metabolic switches, resulting in significant changes in the levels of other tolerance traits in transgenic plants, which support salinity tolerance under stress conditions. | [31] |
| TPS (TSase) | Grifola frondosa | Nicotiana tabaccum | Transgene tobacco showed increased trehalose, high water content, chlorophyll content, and antioxidant enzyme activities, enhanced salinity tolerance with obvious morphological changes but no growth inhibition. | [108] |
| AtTPPD | Arabidopsis thaliana | Arabidopsis thaliana | Plants overexpressing AtTPPD (its product enzyme AtTPPD is chloroplast-localized) produced high levels of starch and soluble sugars, which contributed to salinity tolerance. The result suggests that trehalose metabolism possibly regulates various biological processes by relaying the redox status of different cellular compartments. Also, under salinity stress, the transcript levels of the chloroplastic AtTPPD were highly increased, while tppd mutants were hypersensitive, indicative of the trehalose protective role of the thylakoid membranes when accumulated in the chloroplasts under salinity stress | [51] |
| IbTPS | Ipomoea batatas | Nicotiana tabaccum | Transgenic tobacco overexpressing the IbTPS gene resulted in significantly higher salinity tolerance, trehalose, and proline content than wild type under high salinity. Several stress tolerance-responsive genes were upregulated, suggesting that the IbTPS gene may enhance salinity tolerance by increasing the level of trehalose and proline and modulating the expression of stress tolerance-related genes. | [90] |
| TPS1 | Saccharomyces cerevisiae | Lycopersicon esculentum | TPS1 transgenic tomato plants displayed morphological changes such as thick shoots, rigid dark-green leaves, erect branches, and aberrant root development. The transgenic TPS1 tomato had leaves with high chlorophyll and starch contents. The study revealed that engineering tomato through trehalose biosynthesis showed increased salinity tolerance and yield in the presence and absence of stress. | [112] |
| TPS3 | Oryza sativa | Oryza sativa | ABA and salinity imposition upregulate OsTPP3 overexpression, which elevates trehalose content and improves salinity tolerance in rice seedlings. The work also revealed that the knockout of OsTPP3 reduced rice salinity tolerance associated with a decline in trehalose level. Trehalose application enhanced the salinity tolerance of the tpp3 mutant plant, indicating trehalose’s crucial role in improving crop salinity tolerance. | [107] |
| OsTPS1 | Oryza sativa | Oryza sativa | Overexpression of OsTPS1 improved the resilience of rice seedlings to salinity stress by elevating the content of trehalose and proline and upregulating stress-related genes under saline conditions relative to the wild type. | [91] |
| Ubi1: TPSP | Oryza sativa | Oryza sativa | Overexpression of Ubi1: TPSP induced a significant increase in endogenous trehalose and soluble sugars in transgenic rice and improved its salinity resistance to 150 mM NaCl. | [82] |
| OsTPP1 | Oryza sativa | Oryza sativa | OsTPP1 expression was upregulated under salinity, and its overexpression enhanced seed germination and trehalose levels, triggered stress-responsive genes, and upregulated the expression of OsTPS1, which contributed to rice salinity tolerance. The study indicates the potential use of OsTPP1 in the salinity stress engineering of crops of other crops. | [113] |
| otsA (TPS) | E. ctdi | Brassica campestris | Overexpression of OtsA in Chinese cabbage resulted in transgenic plants showing retained turgidity and photosynthesis rate, delayed necrosis, and remarkably improved salinity tolerance, but exhibited altered phenotypes, including stunted growth and aberrant root development relative to wildtype when subjected to 250 mM NaCl stress. | [114] |
| AtTPS1 | Arabidopsis thaliana | Nicotiana tabacum | Transgenic seeds germinated in different NaCl concentrations, scoring salinity tolerance compared with untransformed plants, confirming that AtTPS1 overexpressing lines stimulated crop tolerance to NaCl stress. | [115] |
| TPS1, TPS2 | Saccharomyces cerevisiae | Arabidopsis thaliana | Lines overexpressing the TPS1-TPS2 construct accumulated trehalose and exhibited a significant increase in salinity resilience with no morphological or growth alterations, while plants overexpressing the TPS1 alone exhibited modified growth, color, and shape. TPS1-TPS2 overexpressing lines were insensitive to glucose, confirming the proposed role of trehalose/T6P in modulating sugar sensing and carbohydrate metabolism. | [116] |
| ZmTPS | Zostera marina | Oryza sativa | Transgenic rice plants overexpressing ZmTPS showed increased endogenous trehalose and tolerance to 150 mM NaCl relative to untransformed control plants. Transgenic plants had no phenotypic aberrations. The work also illustrated that the transformed ZmTPS gene can be transmitted stably from the parent to the progeny in transgenic rice. | [117] |
| OsTPS8 | Oryza sativa | Oryza sativa | Constitutive overexpression of OsTPS8 enhanced soluble sugars and regulated the expression of genes involved in ABA signaling via SAPK9 regulation, which promoted suberin deposition in the root and reduced Na+ content in the shoot and root; this, in turn, improved salinity tolerance and grain yield in rice with no aberrant changes in plant growth and development under saline stress compared with the wild type. | [118] |
| OsTRE1 | Oryza sativa | Oryza sativa | Overexpressing the trehalase gene, OsTRE1, exhibited considerable increases in trehalase activity and remarkable declines in trehalose levels under 150 mM NaCl, with little change in the levels of other soluble sugars. Transgenic plants exhibited enhanced salinity resilience with no morphological alterations or growth defects, suggesting the involvement of OsTRE1 in salinity tolerance in rice. Trehalose accumulation appears not to be a prerequisite for better adaptation to salinity stress, but rather possibly the endogenous trehalose biosynthesis pathway. | [119] |
| otsA (Ubi1: TPSP) | Escherichia coli | Oryza sativa | Transgenic rice produced by the transformation of a gene encoding a bifunctional fusion (TPSP) of TPS and TPP of Escherichia coli elevated trehalose levels in the leaves and seeds, reduced the accumulation of potentially deleterious T6P, displayed no growth inhibition or visible morphological alterations, and enhanced tolerance to salinity stress. | [120] |
| TPS, TPP | Saccharomyces cerevisiae | Medicago sativa | When both genes (TPS, TPP) were fused and expressed, transgenic alfalfa plants displayed improved growth, biomass production, and a significant increase in salinity tolerance. TPS-TPP fusion protein is promising for crop salinity tolerance and enhanced yield under saline conditions. | [121] |
| otsA (TPS) | Mesorhizobium sp. CCBAU25338 | Arachis hypogaea | Overexpression of trehalose synthesis genes otsA in the peanut-nodulating rhizobium Mesorhizobium sp. CCBAU25338 enhanced the salinity stress tolerance and nitrogen-fixing capacity of the rhizobium strain, as well as increased endogenous trehalose, agronomic traits, and lowered oxidative damage in peanuts under salinity conditions. | [62] |
| GmTPP | Glycine max | Glycine max | The expression levels of GmTPPs were upregulated under saline-alkali stress in soybean and were tissue-specific (i.e., flowers, stems, roots, nodules, shoots, leaves, pods, and seeds), suggestive of triggering developmental processes and stress responses that confer stress resilience. | [122] |
Table 2.
Exogenous application of trehalose improves crop performance by triggering different adaptive mechanisms in response to saline conditions.
Table 2.
Exogenous application of trehalose improves crop performance by triggering different adaptive mechanisms in response to saline conditions.
| Crop species | Mode of application | Trehalose concentration (mM) | Response | Reference |
|---|---|---|---|---|
| Oryza sativa | Seed priming | 0, 25 | Alleviated the harmful effects of salinity stress, increased soluble sugars and proline contents contributing to osmotic adjustment, and elevated internal trehalose, stimulating the antioxidant enzyme activities in two rice varieties under salinity imposition. | [60] |
| Chenopodium quinoa Masr 1 | Foliar spray | 0, 2.5, 5 | Growth, total soluble sugars, proline, free amino acids, photosynthetic pigments, yield attributes, antioxidant enzyme activities, and seed nutritional value were improved in quinoa plants by trehalose foliar spray under salinity stress, more so when compost was added to the soil. | [29] |
| Triticum aestivum | Foliar spray | 0, 5, 10, 15 (mg L-1) | Trehalose foliar spray stimulated growth parameters, trehalose content, proline content, and antioxidant enzyme activities in response to salinity imposition; these effects were more pronounced with 15 mg L-1. | [123] |
| Avicennia marina | Added to the Hoagland solution | 0, 20 | Exogenous trehalose enhanced salt resilience by increasing Na+ efflux from the leaf salt gland and root, which reduces the Na+ content in the root and leaf. | [50] |
| Arabidopsis thaliana | Added to liquid cultures | 0, 30 | Levels of trehalose, sucrose, and starch were elevated in response to trehalose treatment. Two-dimensional gel electrophoresis identified nine altered proteins, four of which were responsible for detoxification or stress responses. This work indicates that the external supply of trehalose acted as a regulator of genes involved in responses to abiotic stresses. As exogenous trehalose altered transcript levels of several processes related to tolerance mechanisms, we suggest that trehalose, or metabolites derived from its biosynthesis pathway, are key modulators of gene expression in higher plants under stress. | [88,89] |
| Catharanthus roseus | Added to the nutrient solution | 0, 10, 30, 50 | Supplement of trehalose to the nutrient solution markedly alleviated the inhibitory effects of salinity on plant growth, relative water content, and photosynthetic rate by decreasing Na+, increasing K+, soluble sugars, free amino acids, and leaf gas exchange in leaves under 250 mM NaCl. The regulatory role of exogenous trehalose in stimulating salinity tolerance was optimal with 10 mM, while higher concentrations adversely affected plant growth. It appears that exogenous trehalose acted as a signal to induce the salinity-stressed plants to efficiently raise internal compatible solutes to regulate water loss and turgidity, leaf gas exchange, and ion homeostasis under salinity. | [73] |
| Solanum Lycopersicon | Added to the nutrient solution | 0, 1, 2, 3, 5, 10 | Exogenous trehalose increased growth characteristics, chlorophyll content, proline, internal trehalose, and antioxidant enzyme activities, while decreasing lipid peroxidation, which effectively enhanced tomato salinity tolerance in response to 200 mM NaCl stress. The study also showed that the starch content declined and soluble sugars elevated as trehalose modulated the gene expression of starch and soluble sugar metabolism, induced the upregulation of sugar transporter genes, and those related to the synthesis and metabolism of ABA. It is obvious that trehalose’s role in regulating the above processes significantly modulates sugar accumulation, content, and distribution, thereby improving plant stress resilience. Notably, 2 mM provided optimal responses, and higher concentrations showed decreased responses. | [40] |
| Triticum aestivum | Added to a growth medium | 0, 10, 50 | Trehalose supply significantly stimulated several growth parameters of ten wheat varieties under 150 mM NaCl stress. | [72] |
| Brassica juncea | Foliar spray | 0, 10 | Leaf-applied trehalose increased salinity tolerance and yield by enhancing ion homeostasis, photosynthesis efficiency, antioxidant defense mechanisms, chlorophyll content, osmolyte accumulation, stomatal aperture, cell viability, and ROS scavenging under salinity stress. | [25] |
| Solanum Lycopersicon | Foliar spray | 0, 5, 10, 25 | Trehalose application scavenges ROS by enhancing the activities of antioxidant enzymes and their related gene expression, improves growth, biomass production, proline, and glycine betaine content, increases K+ and K+/Na+ ratio, upregulates the expression of trehalose genes (SlTPS1, SlTPS5, SlTPS7, SlTPPJ, SlTPPH, and SlTRE), and the activity of enzymes involved in its metabolic pathway, which in turn altogether stimulated tomato salinity tolerance. Trehalose at 10 mM was the best mitigation concentration, while 25 mM caused leaf damage and adversely affected plant growth under salinity. | [21] |
| Solanum Lycopersicon | Foliar spray | 0, 10 | Trehalose foliar supply improves photosynthetic efficiency, increases the activity of Calvin cycle enzymes, upregulates the expression of their related genes, protects the photosynthetic electron transport system, affects the expressions of SlSOS1, SlNHX, SlHKT1.1, SlVHA, and SlHA-A, which retain ion homeostasis, induces stomatal opening, and alleviates salt-induced damage to the chloroplast membrane and structure under salinity. The study shows that trehalose renders salinity tolerance by upregulating the processes involved in mitigating salinity toxicity. | [22] |
| Zea mays | Added to the nutrient solution | 0, 10 | Trehalose application improved the performance of two maize genotypes in response to 150 mM NaCl by decreasing the Na+/K+ ratio, ROS, lipid peroxidation, methylglyoxal, and increasing leaf trehalose, salinity tolerance, indicative of regulating antioxidant and glyoxalase systems as well as ion homeostasis. | [95] |
| Triticum aestivum | Foliar spray | 0, 10, 50 | Trehalose enhanced wheat growth and accumulation of compatible solutes (i.e., glucose, sucrose, trehalose, phenolic compounds, total soluble sugars) but decreased lipid peroxidation, hydrogen peroxide, and lipoxygenase activity of salinity-stressed wheat plants. It seems that antioxidant compounds and compatible osmolytes play a crucial role in enhancing wheat performance under salinity by counteracting oxidative damage and, hence, protecting cellular structures. | [87] |
| Cucumis sativus | Added to the nutrient solution | 0, 0.05, 0.2, 0.4, 0.6, 0.8, 1% | Hydrogen-induced cucumber seed germination was done by enhancing enzyme activity and gene expression levels of trehalose metabolism-related genes, which in turn increased the endogenous trehalose content. Although the molecular mechanism and hydrogen crosstalk with other signaling molecules in inducing seed germination are not clear, this work provides new insights concerning the roles and interactions of hydrogen and trehalose during seed germination and confirms one of the fundamental trehalose roles. That is, trehalose as a key regulator of carbon metabolism, largely influences plant growth and development. | [80] |
| Brassica juncea | Foliar spray | 0, 10, 20, 30 | Under the nonsaline condition, trehalose application improved Indian mustard growth characteristics and yield by enhancing osmolyte accumulation and their enzymes’ activities, hence reducing the ROS content, while promoting the content of photosynthetic pigments, gas exchange, mineral acquisition, and root cell viability. These improvements were more pronounced by 10 mM, while other trehalose concentrations were either equally or less effective. | [101] |
| Brassica juncea | Foliar spray | 0, 10 | Foliar spraying of trehalose improved the enzymatic antioxidant activities, compatible solute content, water status, and membrane permeability while decreasing ROS production, lipid peroxidation, and Na+ levels under salinity stress. These effects resulted in improved Indian mustard growth, ion homeostasis, photosynthesis, yield, and salinity resilience. | [102] |
| Oryza sativa | Added to the nutrient solution | 0, 10 | Trehalose protects rice seedlings against NaCl stress by increasing the level of internal trehalose, which effectively reduces ROS accumulation, elevates nonenzymatic antioxidants, and co-activates the antioxidative and glyoxalase systems. | [28] |
| Arabidopsis thaliana | Added to the nutrient solution | 0, 0.5, 1, 5 | Trehalose improved Arabidopsis salinity tolerance by retaining K+ content and a high K+/Na+ ratio, decreasing Na+ level, enhancing internal soluble sugars, and the activities of antioxidant enzymes in plant tissues under NaCl imposition. Thus, it is evident that trehalose regulates plant redox state and ionic distribution under high salinity. Trehalose at 1 mM showed the best salinity tolerance responses. | [69] |
| Triticum aestivum | Plant spray | 0, 10 | Trehalose promoted the growth parameters, yield, leaf anatomical features, endogenous trehalose, amino acid, sugar, total carbohydrate, and total soluble protein contents in 4 wheat cultivars contrasting in their salinity tolerance under 200 mM NaCl. | [125] |
| Oryza sativa | Added to the nutrient solution | 0, 10 | Trehalose supplement to 200 mM NaCl-stressed rice cultivars, contrasting in salinity resilience, did not mitigate the growth reduction during stress, while during recovery plants previously supplied with trehalose displayed a significantly higher growth recovery compared with plants that received only NaCl treatment. This mitigative effect was due to reduced Na+/K+ ratio and H2O2 level, and enhanced ascorbate peroxidase activity. The impact was more pronounced in the salinity-sensitive cultivar. | [81] |
| Oryza sativa | Foliar spray at the tillering stage | 0, 50, 100, 150 | Trehalose enhanced RWC, chlorophyll, soluble sugars, reproductive tillers per plant, grains per panicle, 100-grain weight, percentage of filled grains per panicle, and grain yield per plant in the salt-tolerant variety relative to other varieties. | [126] |
| Oryza sativa | Added to a growth culture | 0, 1, 5, 10 | Application of low trehalose concentrations (up to 5 mM) significantly decreased Na+ accumulation, overcame the growth inhibition, and enhanced the expression of the salT gene, while higher concentrations (10 mM) preserved the root integrity, prevented the chlorophyll damage in leaf blades, and suppressed the expression of the NaCl-induced salT gene. Interestingly, trehalose did not prevent salt accumulation in plant cells but did reduce Na+ accumulation in laminae. It is clear that trehalose protects cellular structure under salinity, but the observed differences in response and between low and high trehalose concentrations might reflect different modes of action at various concentrations, as well as differences in the accumulation or catabolism of trehalose in different parts of the plant. | [100] |
| Rosa rugosa | Foliar spray | 50 μM T6P, 20 mM trehalose |
Exogenous trehalose or T6P improved the drought tolerance of rose plants by alleviating the injurious impact of drought stress, maintaining the rose flower quality, T6P and trehalose accumulation, adjusting the carbohydrate distribution, and altering the synthesis of secondary metabolites (geraniol, total flavonoids, and total anthocyanins). Foliar application of trehalose or T6P also promoted the contents of starch, soluble sugar, and lignin in the petals, pointing to the role of T6P or trehalose as a positive regulatory signal participating in enhancing the rose plant’s resilience to drought stress. | [68] |
| Oryza sativa | Added to a growth culture | 0, 5, 10 | Exogenously supplied trehalose stimulated the growth and alleviated the harsh effects of salinity stress by reducing H2O2 and lipid peroxidation in the sensitive cultivar. However, trehalose did not have any beneficial effect on the growth of the salinity-tolerant cultivar, indicative of trehalose’s protective roles in the salt-sensitive cultivar but not in the salt-tolerant one. Although the study did not determine the endogenous trehalose content, we assume that salinity tolerant cultivar might have sufficient trehalose as well as other tolerance traits, and thus exogenous trehalose showed no beneficial impact on this cultivar. | [127] |
| Oryza sativa | Foliar spray | 0, 0.5% | Trehalose improved the salinity resilience of rice seedlings by increasing the antioxidant enzyme activities under 100 mM NaCl treatment. The study also demonstrated that salinity-induced ABA escalated the expression of OsTPP3, resulting in elevated endogenous trehalose, which stimulated the rice salinity tolerance to NaCl stress. | [107] |
| Zea mays | Seed priming | 0, 10 | Trehalose alleviated the adverse impacts of high salinity on metabolic activity (Hill-reaction activity, photosynthetic pigments, and nucleic acids content), increased sugars, soluble proteins, and proline contents, leaf K+/Na+ ratio, the growth and salinity tolerance, while decreasing electrolyte leakage and lipid peroxidation of the root cells, and salinity expression detected by leaf protein banding patterns of salinity-stressed maize seedlings. It seems that trehalose upregulated various salinity stress-responsive genes that contribute to the tolerance mechanisms of maize seedlings under salinity stress. | [106] |
| Triticum aestivum | Seed presoaking | 0, 10 | Seed presoaking with trehalose improved the salinity tolerance of wheat seedlings by reducing lipid peroxidation, elevating osmotic adjustment osmolytes such as amino acids (especially proline), reducing sugars, and total soluble sugars, and improving the enzymatic and nonenzymatic antioxidants in wheat seedlings. This leads to maintaining the balance between pro-oxidants and antioxidants like phenols, flavonoids, proline, and soluble sugars. It is noteworthy that trehalose was more effective than mannitol. | [128] |
| Glycine max | Added to a growth culture | 10 μmol/L | Trehalose treatment upregulated the expression of TPP genes, and increased the carbohydrate and trehalose levels, while declining ROS content and thus mitigated the adverse impacts caused by saline-alkaline stress, especially in the saline-alkali-tolerant genotype. | [122] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



