
Submitted:
28 November 2025
Posted:
02 December 2025
You are already at the latest version
Abstract
The meta-analysis examines the effects of eCO₂ on the growth, yield, and nutritional composition of two widely consumed leafy vegetables: kale (Brassica oleracea) and spinach (Spinacia oleracea). Following the Collaboration for Environmental Evidence (CEE) guidelines, we systematically reviewed studies that reported on the impacts of eCO₂ on these crops and conducted a meta-analysis to quantify the overall as well as sub-group responses via moderator analyses. A random-effects model was used to calculate effect sizes (Hedges' g), and confidence intervals (CI) were set at 95%. Our results reveal that eCO₂ significantly increased the biomass of spinach (g = 1.21, p < 0.01, CI [0.88, 1.54]) and kale (g = 0.97, p < 0.05, CI [0.65, 1.29]). However, the analysis also detected a significant decrease in protein content in both crops under eCO₂ conditions (spinach: g = -0.76, p = 0.03, CI [-1.10, -0.42]; kale: g = -0.61, p = 0.04, CI [-0.95, -0.27]). Additionally, calcium and magnesium concentrations declined in kale (g = -0.55, p = 0.05, CI [-0.89, -0.21]), and spinach showed a stronger reduction in nutrient content overall. The variability in response across different CO2 concentrations and exposure times further underscores the complexity of eCO2 effects. These findings suggest that while eCO₂ may enhance crop yields, it also leads to a dilution of essential nutrients, raising concerns about the trade-offs between productivity and nutritional quality. The study concludes that targeted breeding programmes, strategic precision agriculture, sustainable agricultural practices and policy interventions will be critical in mitigating the negative nutritional effects of eCO₂ to ensure food security in a changing climate.
Keywords:
elevated carbon dioxide
; kale
; spinach
; crop nutrients
; nutrition
; climate change
; food security
; eCO2
1. Introduction
Climate change is exerting unprecedented pressure on global agricultural systems, and one of the most significant drivers of this change is the increase in atmospheric CO₂ [1]. As CO₂ levels continue to rise, understanding its impact on crop production and nutritional quality is critical for ensuring food security, sustainable diet and human health [2]. Numerous studies have documented elevated CO₂ (eCO₂) effects on a range of crops, mostly cereals and legumes, revealing its potential to enhance photosynthesis, growth, and yield [3]. Most recently, there has been eCO₂ dilution effects documented, where essential micronutrients are concentrated in lower quantities per unit of biomass (compromising the nutritional value of crops), are being explored [4]. However, while key staple crops have been extensively studied, leafy vegetables, particularly kale (Brassica oleracea) and spinach (Spinacia oleracea), have received comparatively very little attention [5]. This oversight is concerning, given the essential role that leafy greens play in global nutrition and food security. This meta-analysis aims to synthesise existing research on the effects of eCO₂ on spinach and kale, two commonly consumed leafy greens, and to elucidate how variations in CO₂ concentration and specific crop types influence overall nutritional quality.
Kale is recognised for its high concentrations of vitamins A, C, and K, making it a key contributor to daily nutritional requirements, particularly for maintaining vision, skin health, and blood coagulation [6]. Additionally, kale is rich in glucosinolates, compounds known for their potential cancer-preventive properties, as well as iron, fibre and calcium, which are critical for bone health [7]. Similarly, spinach is another leafy green of high nutritional value, particularly noted for its rich content of vitamins A, C, and K, as well as essential minerals such as iron, magnesium, and folate [8]. Spinach plays a crucial role in combating iron-deficiency anaemia due to its non-heme iron content, and its high levels of antioxidants contribute to cellular protection and immune function [9].
Both leafy greens have received increasing attention for their nutritional benefits, and their consumption has risen globally. Kale consumption has increased, particularly in North America and Europe, where it is consumed both raw and cooked, contributing to traditional and modern diets [10]. Spinach is widely consumed as a staple vegetable in regions such as South Asia, West Africa, and the Mediterranean, where it is often integrated into a variety of dishes [11]. Their consumption makes them a significant part of both plant-based and mixed diets, with an estimated 24 million tons produced annually worldwide [12]. In the United States alone, spinach cultivation has more than doubled in the last 40 years, reaching an average of 0.6 kg per capita annually [13]. Kale, now often labelled a "superfood" due to its dense nutrient profile, has experienced increasing growth in cultivation, with demand increasing in many Western countries, particularly due to its high antioxidant content. According to the nutrient value reference in the UK, for example, spinach contains 2.8 mg of iron, 379 mg of potassium, and 53 mg of calcium per 100g, while kale provides 120 mg of vitamin C, 92 µg of vitamin K, and 120 mg of calcium per 100g [14]. Thus, reductions in these nutrient levels due to eCO₂-induced nutrient dilution could have significant public health implications, especially among populations that rely heavily on these greens for their micronutrient intake.
Despite the well-documented nutritional interest of kale and spinach, the effects of eCO₂ on their constituents have been less studied compared to staple crops. Preliminary research suggests that eCO₂ may differentially impact various nutritional components in leafy greens, potentially increasing carbohydrate content but decreasing protein and mineral concentrations [15]. Most long-term studies, such as those conducted in Free-Air CO₂ Enrichment (FACE) experiments, have provided valuable insights into these dynamics by simulating future CO₂ conditions in real-world settings; however, very few have investigated leafy vegetable crops [16, 17]. Though these studies have shown that initial benefits in growth and yield may be observed in crops, sustained exposure to eCO₂ often reveals more complex interactions, which include the potential for nutrient dilution and altered phytochemical profiles [4]. Additionally, the response of crops to eCO₂ is not uniform and can be influenced by several factors, including the duration of exposure, the concentration of CO₂, and the specific crop variety [18].
This meta-analysis provides a comprehensive assessment of how eCO₂ affects the nutritional quality of spinach and kale. By examining factors such as CO₂ concentration, crop type, and exposure duration, this study aims to determine the main causes of variability in response and their implications for crop quality. The findings will contribute to a better understanding of how future CO₂ levels might impact these widely consumed leafy greens and to inform strategies for mitigating potential adverse effects on human nutrition and food security.
Understanding the nuances of eCO₂ impacts on crop quality is crucial for developing adaptive strategies in agriculture. Given the projected increase in atmospheric CO₂ and its potential effects on crop nutrition, this meta-analysis provides essential insights that can guide future research and policy-making to ensure the sustainability of food systems and the nutritional adequacy of crops under changing climatic conditions.
2. Materials and Methods
Our meta-analysis adhered to a systematic approach, following the standards established by the Collaboration for Environmental Evidence and Standards for Evidence Synthesis [19]. Our construction was based on a systematic map (Supplement Material, pp 1-3) which provided the basis for our research questions and study selection criteria, detailing the effects of eCO₂ on various crops, including kale and spinach. Our main research question was: “What published evidence exists for the effects of eCO2 on the nutritional components of kale and spinach crops?” Our analysis expanded on this question by incorporating secondary questions related to differences in effects by crop type, CO₂ concentration, duration of exposure, species type, and nutritional outcomes.
2.1. PECO Components and Criteria
Population (P): We included studies explicitly focusing on kale (Brassica oleracea and all common names) and spinach (Spinacia oleracea and all common names), including various cultivars. Studies on other crops or combined datasets that did not provide disaggregated results for kale and spinach were excluded.
Exposure (E): We selected studies that investigated the impact of CO₂ levels ranging from 650 ppm to over 3000 ppm. Studies were excluded if they combined CO₂ exposure with other factors (e.g., drought or heat) without isolating CO₂ effects.
Comparator (C): Only studies comparing CO₂-exposed crops to controls grown under ambient CO₂ conditions (approximately 400-450 ppm) were included. Studies lacking a proper control or baseline comparator were excluded.
Outcome (O): We focused on changes in nutritional constituents namely, carbohydrates, minerals, vitamins, nitrogenous compounds, yields, and photosynthetic parameters. Studies were required to report these outcomes for kale and spinach without aggregation with other crops.
2.2 Literature Search, Study Selection, and Data Extraction
The literature search was systematically conducted over a number of databases, including Scopus, ScienceDirect, Web of Science Core Collection, Google Scholar, CAB Abstracts, and PubMed, with searches concluding on March 12, 2024. The search strategy aimed to capture all relevant peer-reviewed articles examining eCO₂ effects on kale and spinach across all years (i.e. all published studies till date were synthesised in this analysis). This search process resulted in the discovery of 872 studies. References were managed using Zotero, which facilitated the removal of duplicates and the organisation of the literature. After removing 138 duplicates, 734 unique articles were filtered for PECO eligibility using the SysRev review tool where only titles and abstracts were assessed. Relevant articles (N=42) were then subjected to a full-text review based on predefined PECO criteria (full details see Supplementary Table 1) as well as assessment of statistical rigour and completeness of data. Following several unsuccessful email requests, seven full texts could not be assessed. In addition, 29 studies were excluded after full-text review for not reporting the required outcomes (e.g., incomplete data, combined data on effects of eCO2 with other environmental stressors; lacking necessary statistical details; and relevant data could not be obtained from authors directly). Overall, 13 studies were retained, providing 346 effect sizes (full flow diagram: Supplementary Figure 1). The review was done in tandem by 2 reviewers (JUE and JOO), and any disputes were passed to a 3rd reviewer (SR). Data extraction was performed from text, tables, and figures of the selected studies, and MetaDigitize software was utilised to obtain data from graphs. At the same time, summary statistics were calculated from studies providing raw data via Excel. An outlier analysis was conducted, leading to the inclusion of only 339 effect sizes in the final meta-analysis (see supplementary R Markdown file). Information on potential moderators such as species type, CO₂ exposure duration, and outcome categories was also extracted and coded.
2.3. Effect Size and Variance Calculations
Hedges' g [20] was used to calculate effect sizes., a measure that accounts for small sample sizes via the formula:
Where M₁ and M₂ represent the means of the CO₂-exposed and control groups, respectively, while SD_pooled is the pooled standard deviation.
The pooled standard deviation is calculated using:
with s1 and s2 representing the standard deviations and n1 and n2 the sample sizes. The Hedges’ correction factor (J) was applied as follows:
The corrected effect size is:
The variance of each effect size was calculated to estimate precision, with the standard error of g given by:
More details of the calculations are found in Supplementary file p. 6.
2.4. Statistical Analysis
Statistical analyses were carried out using R (version 4.3.3) with the {metafor} package (version 3.8.1) [21]. An overall meta-analytic model was constructed to estimate the general impact of eCO₂ on nutritional quality across kale and spinach (Figure 3A). This model employed a random-effects approach to account for variability among studies, assessed through the Q-test and I² statistic [22].
Single-moderator analyses were performed to explore the influence of factors such as crop type, CO₂ exposure levels, exposure duration, and species type on effect sizes. Multi-moderator models were developed to examine interactions between these factors and account for residual heterogeneity. Due to potential dependencies in effect sizes (e.g., multiple outcomes per study), multilevel mixed-effects models were used, incorporating random effects for study identifiers.
Outlier analysis and sensitivity checks ensured the robustness of findings (Supplementary Figures 3&4). Publication bias was assessed using funnel plots (Supplementary Figures 5-9 and 14), and no substantial asymmetry was observed. Data visualisation was conducted using the {orchaRd} package, providing explicit graphical representations of effect sizes and confidence intervals [23]. Full methodological details, including data extraction and R scripts, are available in Supplementary Files 1, 2, and 3.
3. Results
3.1. Bibliography summary
Overall, after the analyses of outliers, 339 effect sizes generated from 13 articles were retained. Most effect sizes at the time the studies were conducted (and effects evaluated) showed changes in yield and biomass (39%); a smaller percentage examined modifications in nitrogenous compounds (18%), with only a few being connected with mineral contents (14%), photosynthetic components (10%), phytochemicals (7%), vitamin contents (6%) or carbohydrate contents (5%) (Figure 1A).
Of the effect sizes calculated, 170 were from crops exposed to eCO2 for 28 days with the remainder split between 14 days (56), 35 days (35), 26 days (14), 20 days (11), 40 days (11), 60 days (11), 80 days (11), 25 days (8), 29 days (7), 56 days (6), 16 days (2), 30 days (2) and 43 days (2) respectively (Figure 1B). The number of research articles published yearly did not appear to increase significantly from 1997 to 2024, as shown in Figure 1C. Furthermore, the geographic distribution of the research (Figure 2) shows that most were conducted in the United States of America. The CO2 levels crops were exposed to during the experiments in the USA were around 720 - 60,000ppm; Korea 700 - 1,600ppm; China 700 - 800ppm; India 650ppm; Italy 800ppm; Japan 800ppm; and Canada 1,000ppm.
Seventy-nine per cent of the effect sizes originated from studies investigating the effect of eCO2 on kale, while the remaining twenty-one per cen, investigated spinach crops. The effect sizes associated with kale species were from Brassica oleracea cv. alboglabra Bailey (41%) followed by Palmifolia DC (20%), Virdis (20%), Sijicutiao (9%), acephala Winterbor F1 (5%), stem marrow kale (2%), Toscano (0.3%), Winterbor (0.3%) and unspecified (2%) cultivars respectively. Most of the effect sizes were associated with spinach species were from unspecified cultivars (45%) with the remainder split between Spinacea oleracea cv. Gigante invernale (15%), Huangjia (15%), Wase Crone (10%), Melody (1%), Harmony (1%), Bloomsdale LS (1%) and Ipomoea aquatica cv. Forssk (11%) (see Supplementary Material Figure 2).
3.2. Overall and combined effect
The meta-analytic approach evaluating the combined effect of eCO2 unveiled a moderate increase in yield, biomass and nutritional components of kale and spinach crops exposed to atmospheric CO2 metrics ranging from 650 to 60,000ppm compared with crops cultivated at ambient levels of 350-400ppm (Hedges’ g = 1.04; CI 0.40, 1.69; P = 0.0043) (Figure 3 and Supplementary Table 2). Sizeable residual heterogeneity both between (Ibetween2 > 33%) and within (Iwithin2 > 54%) studies was apparent. The model was incrementally expanded to incorporate single moderators to identify important sources of response variability and evaluate the impacts of eCO2 across the types of crops, outcome category, specific constituents measured, period of exposure time and CO2 levels. Testing the combined interactive effect of eCO 2 presence relative to crop types, group and CO2 levels revealed that a portion of the heterogeneity seen in the overall model was caused by the difference in crop type, level of CO2 and category of outcome comparator (QM31,308=3.7943, P<0.001) (See Figure 4 and Supplementary Table 3-9).
Testing sub-group responses showed that eCO2 caused a slightly greater increase in overall yield and nutritional parameters of spinach crops (Hedges’ g=1.06, CI= 0.05, 2.07) compared to kale crops (Hedges’ g=1.03, CI=0.17, 1.90) (Omnibus test of moderators: QM2,337=5.703, P=0.0037) (Figure 4A). Across both crops types combined, only the following outcome category showed statistically significant difference - yields (Hedges’ g=1.15, CI=0.41, 1.89), carbohydrates (Hedges’ g=1.34, CI=0.22, 2.46), nitrogenous compounds (Hedges’ g=1.21, CI=0.38, 2.04) and photosynthetic parameters (Hedges’ g=1.18, CI=0.28, 2.07) with increases. While the changes observed in vitamins (Hedges’ g=0.42, CI=-0.63,1.48), mineral contents (Hedges’ g=0.30, CI=-0.61, 1.21) and other phytochemicals (Hedges’ g=0.47, CI=-0.57, 1.51) were not significant (Omnibus test of moderators detected significant differences (QM7,337=4.0457, P=0.0003)) (Figure 4C). For the differences across CO2 levels, the model showed that as eCO2 level increased, there was a combined increased on yields and all nutritional parameters in both kale and spinach (Omnibus Test of Moderators (QM(df1 = 4, df2 = 335) = 3.3138, p-val = 0.0111) (Figure 4B). There was no significant changes or difference between the exposure time across all groups (Figure 4D).
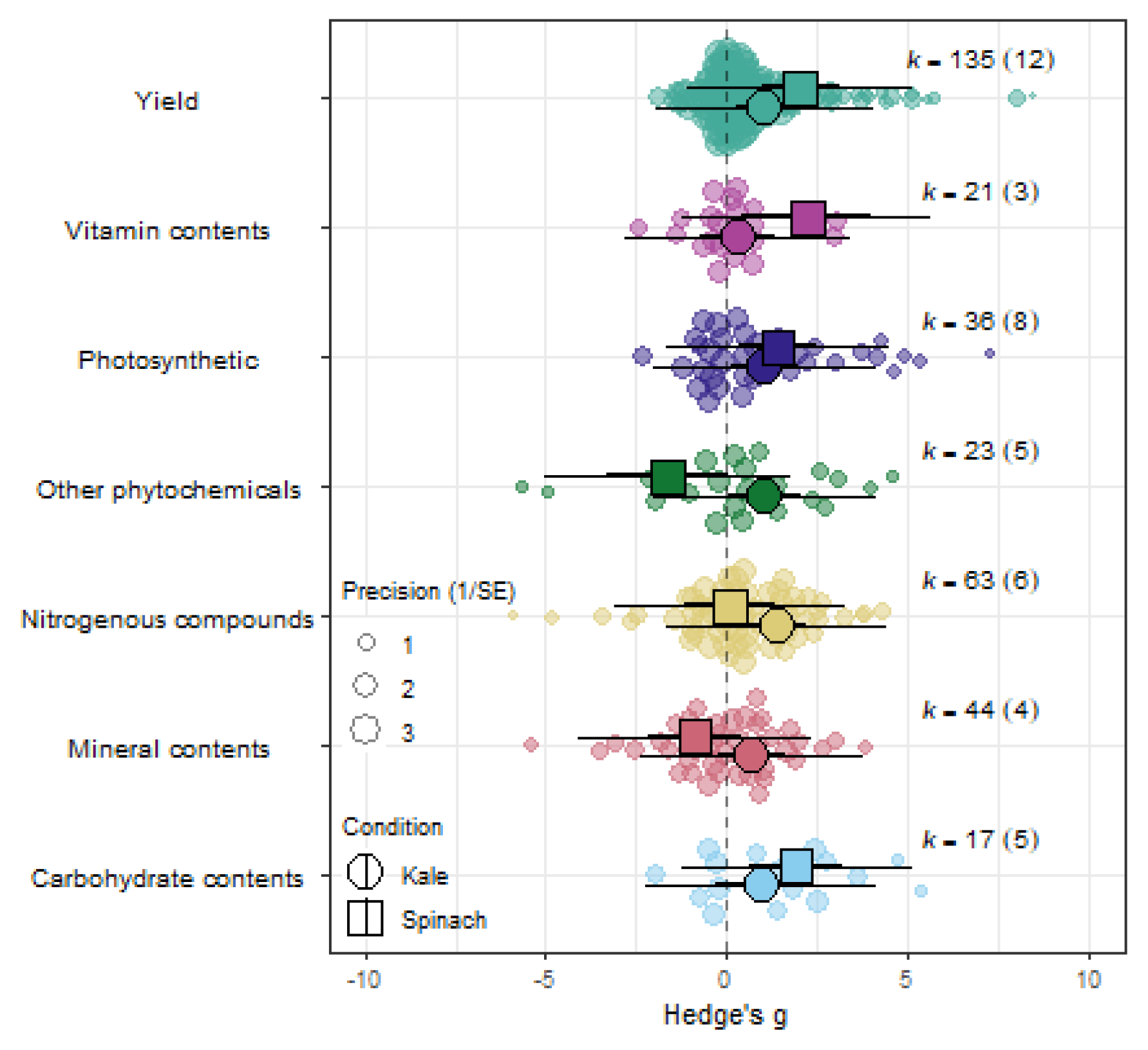
When isolating the effect of eCO2 on kale crops only across the different outcome categories, there was significant increase in photosynthetic parameters, yields, nitrogenous compounds and other phytochemicals only. Similarly, spinach crops showed increase in photosynthetic parameters, yields, vitamin and carbohydrates contents. All other constituents showed no significant changes whether higher or lower (QM6,263=1.6922, P=0.1231) (see Figure. 5 and Supplementary Table 15).
Figure 3.
(a) The overall meta-analytic model demonstrates that eCO2 causes moderate combined increase of biomass and nutritional constituents of both kale and spinach crops (Hedges' g value of 1.04, 95% CI [0.40, 1.69], t12 = 3.5125; P = 0.0043). In all orchard plots, individual effect sizes from studies are shown by the coloured bubbles in each figure, the estimated mean Hedges' g values are represented by the circular dots, the 95% confidence intervals are represented by the strong error bars, and the 95% prediction interval is represented by the thin error bars. Each group's impact size is represented by k, and the number of studies from which each effect size was derived is shown in brackets. (b) A caterpillar plot was also used to visualise the overall meta-analytic model. At the bottom, shown in red with a black 95% confidence interval bar, is the calculated mean Hedges' g value. Each effect size is represented by a yellow dot with green 95% CI bars, and they are sorted by magnitude, drawing from 13 distinct publications and 339 effect sizes.
Figure 3.
(a) The overall meta-analytic model demonstrates that eCO2 causes moderate combined increase of biomass and nutritional constituents of both kale and spinach crops (Hedges' g value of 1.04, 95% CI [0.40, 1.69], t12 = 3.5125; P = 0.0043). In all orchard plots, individual effect sizes from studies are shown by the coloured bubbles in each figure, the estimated mean Hedges' g values are represented by the circular dots, the 95% confidence intervals are represented by the strong error bars, and the 95% prediction interval is represented by the thin error bars. Each group's impact size is represented by k, and the number of studies from which each effect size was derived is shown in brackets. (b) A caterpillar plot was also used to visualise the overall meta-analytic model. At the bottom, shown in red with a black 95% confidence interval bar, is the calculated mean Hedges' g value. Each effect size is represented by a yellow dot with green 95% CI bars, and they are sorted by magnitude, drawing from 13 distinct publications and 339 effect sizes.
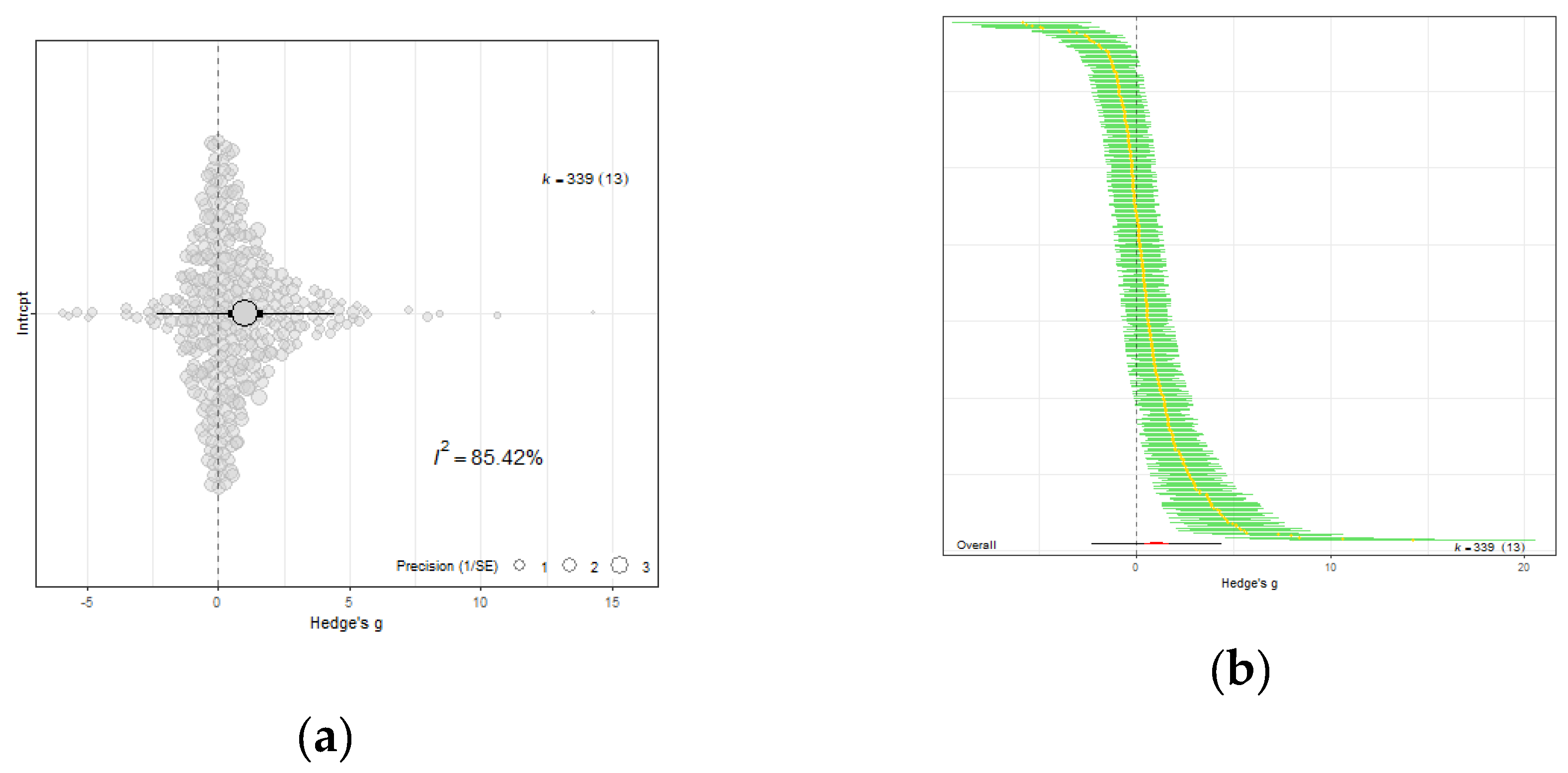
Figure 4.
(a) Using crop type as moderator, the model shows that eCO2 causes more overall increase of biomass and nutritional constituents in exposed spinach crops (Hedges’ g=1.06, CI= [0.05, 2.07]) compared to kale (Hedges’ g=1.03, CI=[0.17, 1.90]) ( t337=2.6321, P=0.0233*); (b) Using CO2 levels as moderator, the model show that the higher the CO2 level, the greater the combined increase of biomass and nutritional contents in both spinach and kale crops - 650-800ppm level (Hedges’ g=0.95, CI=0.21, 1.69, t335=2.9153, P=0.0172*), 900-1000ppm (Hedges’ g=1.06, CI=0.16, 1.96, t335=3.2617, P=0.7292), 1300-1900ppm (Hedges’ g=1.19, CI=0.35, 2.04, t335=3.7860, P=0.30846) and 3000ppm> (Hedges’ g=1.48, CI=0.50, 2.46, t335=4.3399, P=0.1552); (c) Using the outcome category as moderator, the model show that eCO2 causes increase in only yields (Hedges’ g=1.15, CI=0.41, 1.89, t332=2.4368, P=0.6264), carbohydrates (Hedges’ g=1.34, CI=0.22, 2.46, t332=2.9541, P=0.0265*), nitrogenous compounds (Hedges’ g=1.21, CI=0.38, 2.04, t332=2.6259, P=0.7657) and photosynthetic components (Hedges’ g=1.18, CI=0.28, 2.07, t332=2.5654, P=0.7200). The changes in mineral (Hedges’ g=0.30, CI=-0.61, 1.21, t332=0.4407, P=0.0135*), vitamins (Hedges’ g=0.42, CI=-0.63,1.48, t332=0.9707, P=0.0516) and other phytochemicals (Hedges’ g=0.47, CI=-0.57, 1.51, t332=1.0996, P=0.0690) were not significant; (d) Bubble plot assessing whether the number of days plants were exposed to affect the yield and nutritional response to the effects of eCO2 (expressed as Hedges g), points coloured blue are spinach species while red are kale (t1,13=0.2363, P=0.9397).
Figure 4.
(a) Using crop type as moderator, the model shows that eCO2 causes more overall increase of biomass and nutritional constituents in exposed spinach crops (Hedges’ g=1.06, CI= [0.05, 2.07]) compared to kale (Hedges’ g=1.03, CI=[0.17, 1.90]) ( t337=2.6321, P=0.0233*); (b) Using CO2 levels as moderator, the model show that the higher the CO2 level, the greater the combined increase of biomass and nutritional contents in both spinach and kale crops - 650-800ppm level (Hedges’ g=0.95, CI=0.21, 1.69, t335=2.9153, P=0.0172*), 900-1000ppm (Hedges’ g=1.06, CI=0.16, 1.96, t335=3.2617, P=0.7292), 1300-1900ppm (Hedges’ g=1.19, CI=0.35, 2.04, t335=3.7860, P=0.30846) and 3000ppm> (Hedges’ g=1.48, CI=0.50, 2.46, t335=4.3399, P=0.1552); (c) Using the outcome category as moderator, the model show that eCO2 causes increase in only yields (Hedges’ g=1.15, CI=0.41, 1.89, t332=2.4368, P=0.6264), carbohydrates (Hedges’ g=1.34, CI=0.22, 2.46, t332=2.9541, P=0.0265*), nitrogenous compounds (Hedges’ g=1.21, CI=0.38, 2.04, t332=2.6259, P=0.7657) and photosynthetic components (Hedges’ g=1.18, CI=0.28, 2.07, t332=2.5654, P=0.7200). The changes in mineral (Hedges’ g=0.30, CI=-0.61, 1.21, t332=0.4407, P=0.0135*), vitamins (Hedges’ g=0.42, CI=-0.63,1.48, t332=0.9707, P=0.0516) and other phytochemicals (Hedges’ g=0.47, CI=-0.57, 1.51, t332=1.0996, P=0.0690) were not significant; (d) Bubble plot assessing whether the number of days plants were exposed to affect the yield and nutritional response to the effects of eCO2 (expressed as Hedges g), points coloured blue are spinach species while red are kale (t1,13=0.2363, P=0.9397).
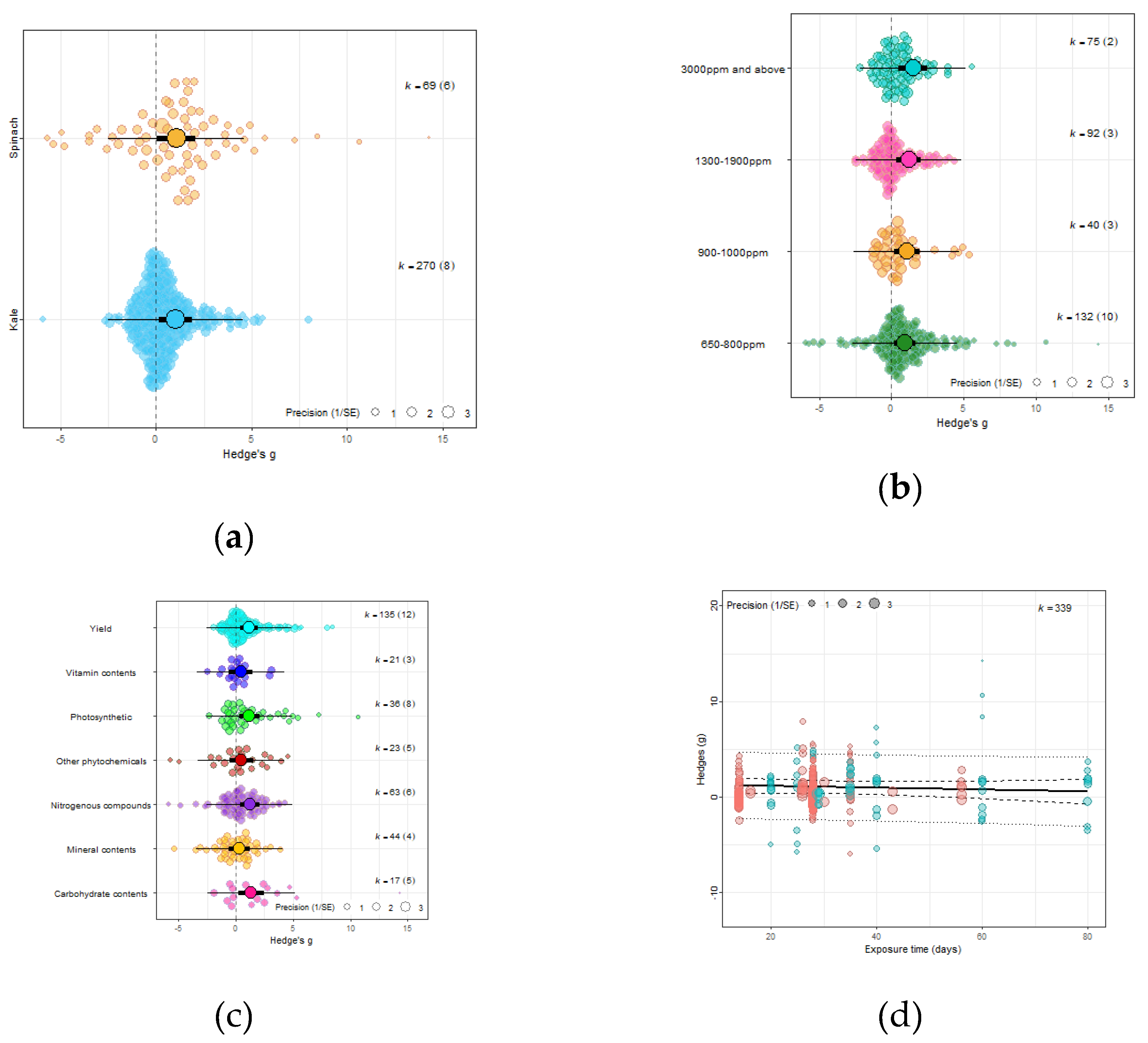
Figure 5.
Orchard plot showing the combined effect of eCO2 on all crops with a complex model of crop type and outcome category as moderators.
Figure 5.
Orchard plot showing the combined effect of eCO2 on all crops with a complex model of crop type and outcome category as moderators.
3.3. Effect on individual constituents
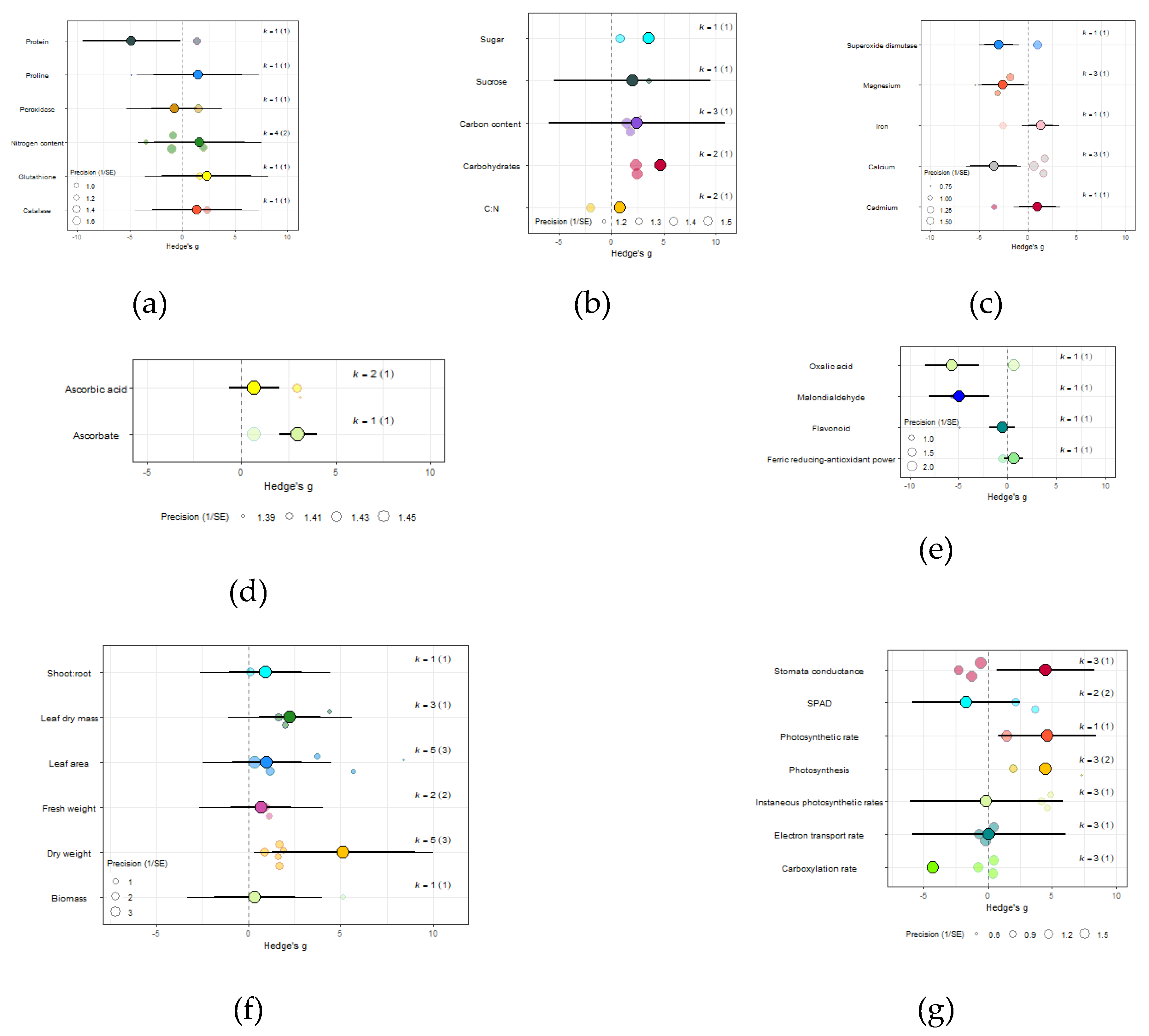
The impact of eCO2 varied according to the type of specific constituents measured. Within yields category of kale crops, only fresh weight (Hedges’ g=2.15, CI=1.52,2.79), leaf area (Hedges’ g=1.00, CI=0.36,1.65), plant height (Hedges’ g=0.95, CI=0.17,1.77), root surface area (Hedges’ g=7.22, CI=4.89,9.56) and root-shoot ratio (Hedges’ g=1.98, CI=0.60,3.35) showed increases to eCO2 (Figure 6G) While eCO2 increased only dry weight (Hedges’ g=5.15, CI=1.28,9.02) and leaf dry mass (Hedges’ g=2.24, CI=0.60,3.90) in spinach crops (Figure 7F) (QM14=121.5006, P<0.0001).
In the carbohydrate category, eCO2 caused an increase within kale crops only in the carbon-nitrogen ratio (Hedges’ g=4.72, CI=1.76,7.68) and unclassified carbohydrate contents (Hedges’ g=5.31, CI=2.04,8.58) (Figure 6A and 6B) (QM3=20.3902, P=0.0001). All other specific measured constituents did not show any significant changes (increase or decrease).
For protein and nitrogenous compounds, kale showed increases in 4-methoxyglucobrassin (Hedges’ g=0.80, CI=0.08,1.52), glucobrassin (Hedges’ g=1.26, CI=0.54,1.99) and progoitrin (Hedges’ g=1.12, CI=0.40,1.84) but decrease in nitrogen-sulfur ratio (Hedges’ g=-2.68, CI=-5.33,-0.02) and unclassified nitrogen contents (Hedges’ g=-5.90, CI=-9.89,-1.91) (Figure 6F). While spinach showed decrease in unclassified proteins (Hedges’ g=-4.85, CI=-9.52,-0.17) (Figure 7A) (QM9=46.8107, P<0.0001).
Sulfur contents under the mineral category was significantly decreased in kale crops (Hedges’ g=1.27, CI=0.18,2.37) (Figure 6E). While for spinach crops showed decrease in calcium (Hedges’ g=-3.50, CI=-5.90,-1.10), magnesium (Hedges’ g=-2.56, CI=-4.71,-0.40), superoxide dismutase (Hedges’ g=-2.95, Ci=-4.40,-1.50) but increase in iron (Hedges’ g=1.32, CI=0.06,2.57) (Figure 7C) (QM7=15.4177, P=0.0310). There were no significant changes for all other specific measure constituent.
Similarly, eCO2 caused an increase in chlorophyll values (Hedges’ g=3.89, CI=1.97,5.80) of kale crops only (Figure 6C). While spinach showed increase in photosynthetic rates (Hedges’ g=4.62, CI=0.80,8.44) and stomata conductance (Hedges’ g=4.51, CI=0.70,8.31) but decrease in carboxylation rate (Hedges’ g=-4.29, CI=-8.37,-0.21) (Figure 7G) (QM2=16.5623, P=0.0003). Kale crops showed decrease in vitamin B (Hedges’ g=-0.65, CI=-1.23,-0.08) (Figure 6B) while spinach showed increase in ascorbate (Hedges’ g=3.02, CI=2.03,4.01) (Figure 7D) (QM1=4.9116, P=0.0267).
For other phytochemical constituents, spinach crops showed only decreases and they were in malondialdehyde (Hedges’ g=-4.94, Ci=-8.01,-1.86) and oxalic acid (Hedges’ g=-5.70, CI=-8.44,-2.96) (Figure 7E). While kale showed decrease in aliphatic contents (Hedges’ g=-1.01, Ci=-1.91,-0.10) but increase in crude fat (Hedges’ g=4.52, Ci=1.37,7.68), glucoalyssin (Hedges’ g=1.64, Ci=0.67,2.61), glucoerucin (Hedges’ g=2.53, Ci=0.19,4.87) and glucoraphanin (Hedges’ g=2.72, CI=0.32,5.12) (Figure 6D) (QM9=38.0996, P<0.0001). Other specific measured constituents did not show any significant changes (also see Supplementary Table 14).
4. Discussion
4.1. Summary of the Combined Effect of Elevated CO2 on Spinach and Kale
The overall findings of this meta-analysis revealed a significant combined moderate increase of yield, biomass and nutritional contents in spinach and kale grown under eCO2 conditions compared to ambient equivalents, with a Hedges' g value of 1.04 (P=0.0043). This aligns with prior studies demonstrating the stimulating effect of eCO2 on plant growth and yield across the same crop species [24]. The meta-analysis also identified significant variability (heterogeneity) whereas suggesting that the response to eCO2 is not uniform and varies across different crops, cultivars, specific CO2 level utilised and constituents measured. This is consistent with previous research indicating that while eCO2 generally enhances photosynthetic activity and biomass production, the degree of this enhancement can vary widely depending on crop type, cultivar, and other environmental factors [18]. Notably, spinach exhibited a stronger response to eCO2 compared to kale, suggesting that species-specific physiological traits may influence the degree of responsiveness to elevated CO2 [25]. This finding aligns with studies on leafy vegetables that show variability in response to eCO2, where spinach generally shows more pronounced growth increases due to its higher photosynthetic efficiency and lower nitrogen use efficiency [26].
The impact of cultivar types on the response to eCO2 was not evident in this meta-analysis, though Brassica oleracea cv. Palmifolia DC and Brassica oleracea cv. Viridis were highlighted to behave differently from the rest, and each variety still had a different estimate mean value and confidence interval. Variation within cultivars can be associated with differences in genetic makeup, which affect physiological traits such as stomatal conductance, nutrient uptake efficiency, and carbon allocation [27]. Such findings have been reported in other crops, where the response to eCO2 varies significantly among different cultivars. For instance, in wheat, cultivars with higher intrinsic water-use efficiency and nitrogen-use efficiency tend to show greater yield responses under eCO2 conditions [28, 29]. This suggests that breeding and selection of cultivars with specific traits could be helpful for optimising crop performance in future CO2-enriched environments [30].
The meta-analysis found no significant difference in response to eCO2 across different exposure times, suggesting that the duration of exposure may not be a key factor in determining the overall impact of eCO2 on spinach and kale. This finding contrasts with some earlier studies that indicated a time-dependent response, where initial growth stimulation under eCO2 could taper off over time due to nutrient limitations or acclimation effects [15]. However, the lack of a significant time effect in this analysis could be due to the relatively short duration of most studies included, which may not capture long-term acclimation processes. Statistics from long-term experiments that fully understand the temporal dynamics of eCO2 effects would only be applicable in perennial crops or multi-year cultivation systems, unfortunately [17].
The response of crops to increasing CO2 levels revealed a strong positive correlation, with higher CO2 concentrations (beyond 800 ppm) leading to greater effect size estimates. This finding contrasts with previous studies that suggested an optimal response at intermediate CO2 levels (600–800 ppm) before plateauing [3; 31]. Instead, our results indicate that as CO2 levels rise, crops continue to exhibit enhanced photosynthetic activity, biomass accumulation, and yield improvements without clear saturation effects within the studied range. This pattern aligns with recent research suggesting that certain C3 crops, including leafy vegetables, may sustain productivity gains under elevated CO2 well beyond previously assumed thresholds [32]. The observed response in this meta-analysis emphasises how crucial it is to consider specific CO2 concentrations in future projections of crop performance under climate change scenarios.
4.2. Effect on Nutritional Components and Implications for Global Health
(i) Yields and Biomass
Elevated CO2 was found to significantly increase yields and biomass in spinach and kale, particularly in parameters such as root surface area, plant height, fresh weight, and leaf dry mass. This aligns with numerous studies that have documented similar increases in biomass and yield under eCO2 across various crop species [33]. The enhancement in biomass is primarily driven by increased photosynthetic rates and reduced photorespiration, which are direct responses to higher atmospheric CO2 [32]. However, the magnitude of yield increases can be variable depending on other factors such as nutrient availability, water supply, and cultivar characteristics [34]. In particular, the increase in biomass does not always translate into proportional increases in yield quality, as changes in nutrient content can occur simultaneously [35].
The impact of eCO2 on crop quality also has broader implications for global food security. As crop yields increase under eCO2, the dilution effect, where higher biomass dilutes the concentration of nutrients, could lead to a paradox where more food is produced but with lower nutritional value. This could exacerbate micro-deficiency problem, where people have enough calories to eat but lack essential nutrients, leading to malnutrition [36]. In addition, the variability in response among different crops suggests that some regions may be more adversely affected than others, depending on their primary food sources. This could lead to greater disparities in nutritional outcomes, with populations in certain areas experiencing more severe nutrient deficiencies.
(ii) Carbohydrate/carbon contents
The analysis showed that eCO2 led to a significant increase in the carbon-nitrogen ratio and unclassified carbohydrate contents in kale only, whereas no significant changes were observed in other measured constituents in both kale and spinach. The demonstrated increase is consistent with the general understanding that eCO2 stimulates carbohydrate synthesis through enhanced photosynthetic carbon fixation [37]. And increased carbohydrate accumulation under eCO2 has been reported in various crops, including wheat, rice, and maise, often leading to higher starch and sugar content [30]. However, this increase in carbohydrates can be accompanied by a reduction in protein concentration, leading to a dilution effect that could impact the nutritional quality of crops [38]. This trade-off between carbohydrate enrichment and protein reduction is a critical consideration for the nutritional implications of future crops grown under elevated CO2 conditions. Moreso, while increased carbohydrate accumulation has been well-documented in staple crops such as wheat, rice, and maize [30], our results suggest that leafy vegetables may exhibit a more selective response, with only certain carbohydrate fractions being affected.
Higher sugar contents, which may first seem to be advantageous for producing biomass in plants, have concerning implications for nutrition. Studies have indicated that increased crop sugar levels, especially in frequently consumed food crops, may be a factor in the rising rates of obesity and associated metabolic diseases like diabetes in developed nations [39, 40]. In the United States, for example, the growth in carbohydrate content under eCO2 conditions may cause people to consume identical amounts of food but with considerably more sugar [41]. According to the CDC, more than 42% of adult Americans are obese, and more than 11% of people have diabetes [39]. The obesity pandemic may be fuelled by this "hidden-sugar" effect, which can lead to an increase in calorie intake without a commensurate gain in satiety.
(iii) Proteins and Nitrogenous Compounds
The meta-analysis found that nitrogenous compounds, particularly nitrogen-sulfur ration and other unclassified nitrogen contents, showed a significant decrease under eCO2, which is indicative of a potential decline in protein content. This is in line with numerous studies showing that eCO2 often leads to reduced nitrogen concentration in plant tissues, likely due to a combination of factors such as dilution by increased carbohydrate content and reduced nitrogen uptake efficiency [42]. The reduction in protein content under eCO2 has been a consistent finding across various crop species, including cereals and legumes, which raises concerns about the potential impact on human nutrition, especially in regions where plant-based diets are predominant [43, 44]. The mechanisms underlying this reduction are complex and may involve changes in root-to-shoot nitrogen allocation, altered nitrogen assimilation pathways, and interactions with other environmental stressors such as drought or nutrient limitation [45].
The decline in protein content is particularly concerning given that billions of people worldwide rely on plant-based sources for their protein intake, especially in developing countries where animal protein is less accessible [44, 46]. A reduction in protein content could exacerbate issues of malnutrition, leading to increased rates of stunting, impaired cognitive development, and weakened immune systems among vulnerable populations [2]. Moreover, the decline in protein quality, indicated by a reduction in essential amino acids, could further impact dietary adequacy. Studies have shown that eCO2 can lead to reductions in lysine and other essential amino acids in staple crops like wheat, rice, and soybeans, which are crucial for maintaining muscle mass, enzyme function, and overall metabolic health [31].
(iv) Minerals
Mineral contents, including calcium, magnesium, and sulfur, showed varied responses to eCO2 in spinach and kale. While sulfur contents in kale showed a moderate increase, other minerals, such as calcium and magnesium, exhibited significant decrease in spinach. This differential response is consistent with previous research indicating that eCO2 can alter the nutrient composition of crops, often leading to reductions in essential minerals [47]. The decline in mineral content under eCO2 is thought to be related to the dilution effect, where the increased biomass production leads to lower concentrations of minerals per unit of plant tissue [35]. Additionally, changes in soil chemistry and nutrient uptake dynamics under eCO2 could contribute to these effects [35].
The observed decline in essential minerals such as calcium and magnesium in kale and spinach grown under eCO2 has severe implications for global nutrition as they exacerbate micronutrient deficiencies [36]. Reduction of minerals in crop produce contributes to the global problem of “hidden hunger” (a characteristic of micronutrient deficits), which the World Health Organization has identified as a significant worldwide health concern, especially in low- and middle-income nations [48]. Over two billion people worldwide suffer from hidden hunger, especially in low-income areas [6]. According to the most recent global assessment conducted by WHO, over 30% of people worldwide lack sufficient iron, and roughly 17% need zinc [48]. Micronutrients are vital for bone health, immune function, and metabolic processes, and deficiency has serious negative effects on health, including compromised immune systems, decreased cognitive development, and increased susceptibility to infections. The reduction in calcium and iron content due to eCO2 has been associated with a rise in the occurrences of anaemia, osteoporosis, and other related health problems [5]. Given that crop nutritional quality is declining under eCO2, these numbers are projected to increase.
(v) Vitamins
Vitamin responses to eCO2 were variable, with vitamin B showing a significant decrease and ascorbate increase in this meta-analysis. The impact of eCO2 on vitamin content has received less research than other nutrients, but existing research suggests that the effects can be highly specific to the type of vitamin and crop species [49]. For instance, studies on rice and wheat have reported both increases and decreases in various vitamins under eCO2, with factors such as cultivar, soil conditions, and interactions with other environmental variables playing critical roles [24]. The findings in spinach and kale suggest that while some vitamins may benefit from eCO2, others could decline, potentially affecting the overall nutritional quality of these crops. Additionally, any decline in vitamin contents could have implications on global nutrition because kale and spinach are recognised for their high concentrations of vitamins A, C, and K, making them key contributors to daily nutritional requirements, particularly for maintaining vision, skin health, and blood coagulation [6].
(vi) Other Phytochemicals
Phytochemicals such as glucosinolates showed a significant decrease under eCO2, with compounds like glucoalyssin, glucoerucin, and glucoraphanin exhibiting a decline in kale. In contrast, kale also showed an increase in aliphatic glucosinolate content, while spinach exhibited a significant decrease in malondialdehyde and oxalic acid. This is in line with findings from prior research that have revealed changes in specific phytochemicals under eCO2, possibly as a response to altered carbon allocation and secondary metabolite synthesis pathways [46].
The response of phytochemicals to eCO2 is intricate and subject to change based on the specific compound and crop species. For instance, some studies have reported increased concentrations of flavonoids and phenolic compounds under eCO2, while others have found reductions [2, 8; 45]. These changes in phytochemical content can have significant implications for the health benefits of crops, as many phytochemicals are known for their antioxidant, anti-inflammatory, and anti-carcinogenic properties [8]. The increase in aliphatic glucosinolates in kale, for example, could enhance its cancer-preventive properties, while the decline in glucoalyssin, glucoerucin, and glucoraphanin may reduce its overall health-promoting potential.
Phytochemicals play a crucial role in the prevention of long-term illnesses such as cancer, diabetes, and cardiovascular disease. The findings of this meta-analysis indicate that eCO2 can alter the concentration of these compounds in crops, with some increasing while others decrease. This variability could affect the protective health benefits that these crops offer [6]. For instance, while the increase in aliphatic glucosinolates in kale may enhance its anti-cancer properties, the decline in key glucosinolates, crude fat, malondialdehyde, and oxalic acid suggests that eCO2 induces complex biochemical trade-offs that may influence the nutritional and functional properties of both kale and spinach [8].
(vii) Photosynthetic Parameters
The meta-analysis indicated that eCO2 significantly enhanced several photosynthetic characteristics, including stomatal conductance, photosynthetic rate, and chlorophyll concentration in kale and spinach. These findings are consistent with the well-documented effect of eCO2 in increasing the photosynthetic capacity of plants by enhancing the carboxylation efficiency of RuBisCO (the enzyme ribulose-1,5-bisphosphate carboxylase/oxygenase) and reducing photorespiration [50]. The increase in photosynthetic activity under eCO2 often leads to higher biomass production, as observed in this study. However, the relationship between photosynthetic enhancement and yield is not always straightforward, as factors such as nutrient availability, water use efficiency, and the balance between source and sink capacities can modulate the overall growth response [17]. In spinach, the enhanced photosynthesis was reflected in increased biomass, but in kale, the response was more variable, suggesting that other physiological or environmental factors may be limiting the full realisation of the CO2 fertilisation effect.
The long-term implications of eCO2 on global health are profound, particularly as they relate to human nutrition and food security. As the meta-analysis has shown, while eCO2 can lead to increased crop yields, the quality of these crops—specifically their nutritional content—can be adversely affected. This poses significant challenges for global health, especially in regions where plant-based diets are predominant, and nutrient deficiencies are already a concern.
4.3. Implications for Future Sustainability and Agriculture
Long-term FACE experiments in the USA, such as those conducted on soybeans, wheat, and rice, have provided critical insights into the sustained impact of eCO2 on crops. These studies have demonstrated that while initial increases in photosynthesis and growth are common, these benefits can diminish over time due to nutrient limitations, particularly nitrogen [26]. For example, the SoyFACE project in Illinois has shown that while soybean yields increase under eCO2, the protein content decreases, posing challenges for both human nutrition and livestock feed [5]. The long-term nature of these studies also reveals the potential for acclimation effects, where plants may initially respond positively to eCO2, but over time, the benefits plateau or even decline. This acclimation is often linked to limitations in other resources, such as water and nutrients, highlighting the need for integrated approaches that consider the full range of environmental factors affecting crop growth [51].
Given the findings of both this meta-analysis and long-term FACE studies, there is a clear need for sustainable agricultural practices that can mitigate the negative impacts of eCO2 on crop nutrition. Soil nutrient management will be crucial in this regard. Practices such as precision agriculture, which optimises nutrient application based on real-time monitoring of crop needs, could help maintain or even improve the nutritional quality of crops under eCO2 conditions [18]. In addition, biofortification particularly, breeding crops to increase their nutrient content, could play a key role in addressing the reductions in essential minerals and vitamins observed under eCO2. This approach has already shown success in increasing the iron and zinc content of staple crops like rice and wheat and could be extended to other crops affected by eCO2 [47].
Diversified cropping systems, which involve growing a variety of crops rather than relying on monocultures, could also enhance resilience to the impacts of eCO2. By incorporating crops that are less sensitive to eCO2-induced nutritional changes, farmers can reduce the risk of nutrient deficiencies in their produce [52]. Moreover, integrating traditional and indigenous crop varieties, which may have unique adaptations to local environmental conditions, could provide additional resilience against the challenges posed by climate change and eCO2 [26]. The challenges posed by eCO2 for agriculture are multifaceted, involving not only the direct effects on crop yields and nutritional quality but also broader implications for agricultural sustainability, food systems, and global food security.
4.4. Limitations, Research Gaps, and Future Directions
This meta-analysis presents several limitations that must be considered in interpreting its findings, as well as important research gaps that need to be addressed to fully understand the implications of eCO₂ on agricultural productivity and nutrition. First, the scope of available data for spinach and kale under eCO₂ conditions remains limited, with most studies being conducted in controlled environments within temperate climates. While these studies offer valuable insights, the lack of data from diverse geographical regions restricts the ability to generalise the findings globally. Factors such as soil quality, water availability, and local climate can significantly influence how crops respond to elevated CO₂, limiting the applicability of current results. Future research should prioritise gathering data from regions outside the temperate zone and consider varying environmental stressors such as drought, heat, and nutrient availability to comprehensively understand global agricultural systems under eCO₂.
Second, a major limitation of this meta-analysis is the variability in experimental designs across the studies included. Differences in soil types, nutrient availability, water supply, growth conditions, the measurement of crop quality parameters (e.g., biomass, nutrient concentration) and experimental duration can all influence the outcomes of eCO2 experiments, leading to heterogeneity in the results [38]. This variability complicates the task of drawing definitive conclusions about the effects of eCO2 on crop nutrition. Although efforts were made to account for key variables, differences in how crop quality parameters, such as biomass and nutrient concentrations, were measured may contribute to heterogeneity in the results. This variability underscores the importance of standardising experimental protocols in future studies or employing meta-regression techniques to account for differences in experimental conditions [51]. In addition, it should be noted that while a sizeable number of effect sizes were available, the overall meta-analysis is based on outcomes from 13 primary studies only. While outcomes most certainly indicate overall converging trends in both spinach and kale crops in response to eCO2, more research and better accessibility to data is required.
A key challenge is the relatively short growing periods for these annual and biennial crops, meaning that studies are inherently limited to short-term effects of eCO₂. This limits our understanding of how repeated seasonal exposures might alter crop physiology or nutritional quality over time. However, given the life cycle of kale and spinach, conducting truly long-term studies remains a practical challenge.
Additionally, there is a risk of publication bias in this field, with studies showing significant effects of eCO₂ on yield and nutrient composition more likely to be published. Null or inconclusive results may be underreported, which could skew the overall interpretation of the data. Although this meta-analysis employed funnel plots to assess for such bias, it cannot entirely eliminate the possibility that underrepresented studies may impact the synthesis of the results.
More analysis needs to be done to quantify the net effect of the interaction between eCO₂ and other environmental stressors, such as water availability and heat, on leafy vegetables. For example, water stress has been shown to exacerbate the negative effects of eCO₂ on nitrogen assimilation in plants, leading to further declines in protein content [43]. Investigating these combined stressors is essential for understanding how crops will respond to future climate scenarios. Furthermore, the broader nutritional and health implications of eCO₂-induced changes in crop quality, such as reductions in essential nutrients and phytochemicals, remain underexplored. Epidemiological studies linking these changes to public health outcomes, particularly in vulnerable populations, could offer valuable insights into the long-term consequences of eCO₂ on human nutrition [45].
The findings raise concerns about the dilution of essential nutrients, particularly proteins and minerals, in crops grown under eCO2 conditions. The increase in carbohydrate content, coupled with a decline in nitrogenous compounds and key minerals like calcium and magnesium, underscores the potential for a trade-off between yield and nutritional quality. This phenomenon, often referred to as the "dilution effect," poses significant challenges for global health, particularly in regions where plant-based diets are predominant. The reduction in essential nutrients could exacerbate existing issues of malnutrition, leading to adverse health outcomes such as stunting, cognitive impairments, and weakened immune systems, particularly in vulnerable populations.
In terms of practical applications, the findings of this meta-analysis suggest several avenues for mitigating the negative impacts of eCO2 on crop nutrition. One potential strategy is the use of soil supplements, such as nitrogen fertilisers, to counteract the dilution of protein content. However, the environmental sustainability of such interventions must be carefully considered, especially in the context of global efforts to mitigate agricultural greenhouse gas emissions and enhance soil health [52, 53]. Precision agriculture techniques, which optimize nutrient application based on real-time monitoring, could help maintain crop nutritional quality. Additionally, exploring the potential of biofortification and genetic modification including the use of CRISPR technology to enhance the nutrient content of crops under eCO2 conditions could offer another promising approach. Such technologies could be used to develop crop varieties that are not only more resilient to eCO2 but also capable of maintaining or even increasing their nutritional quality [47]. Diversified cropping systems that incorporate a variety of crops with different sensitivities to eCO2 could reduce the risk of nutrient deficiencies and enhance agricultural resilience.
5. Conclusions
This meta-analysis highlights both opportunities and challenges posed by eCO₂ for agricultural productivity and global nutrition. While eCO₂ enhances the biomass and yield of spinach and kale, there is significant variability in crop responses, with spinach showing stronger benefits compared to kale. The findings suggest that targeted breeding programs to enhance physiological traits like photosynthetic and nutrient-use efficiency could maximise these benefits. However, the concurrent reduction in essential nutrients, particularly proteins, vitamins and minerals, due to eCO₂ presents a major concern. This "dilution effect" poses risks for global nutrition, particularly in regions where plant-based diets are dominant and could exacerbate existing malnutrition (including obesity and micro-deficiencies) as well as related health issues. The diminishing benefits of eCO₂ over time, as shown by long-term studies, further emphasise the importance of integrated nutrient management. To mitigate these negative impacts, strategies such as precision agriculture, biofortification, and genetic modification should be prioritised. Developing resilient crop varieties and adopting diversified cropping systems will be essential for balancing productivity with nutritional quality. Addressing these challenges is crucial for maintaining global food security in an era of rising atmospheric CO₂ levels.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
“Conceptualization, J.E. and R.S.; methodology, J.E. and S.Z.; software, S.Z.; validation, R.S., R.W. and K.L.; formal analysis, J.E. and J.O; investigation, J.E. and J.O; resources, S.Z.; data curation, J.E.; writing—original draft preparation, J.E.; writing—review and editing, J.E., S.Z., R.S., R.W., F.P., A.F. and K.L.; visualization, J.E.; supervision, R.S., R.W., F.P., A.F. and K.L.; project administration, R.S.; funding acquisition, R.S. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
“This research was funded by LIVERPOOL JOHN MOORES UNIVERSITY VC STUDENTSHIPS FACULTY FUNDING, title: Climate change alters the nutritional value of crops. 2024-2028”
Data Availability Statement
“All data generated or analysed during this study are included in this published article and its supplementary information file.”
Conflicts of Interest
“The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
Abbreviations
The following abbreviations are used in this manuscript:
| CO2 | Atmospheric carbon dioxide |
| eCO2 | Elevated carbon dioxide |
| RuBisCO | The enzyme ribulose-1,5-bisphosphate carboxylase/oxygenase |
| FACE | Free-air CO2 enrichment |
| ppm | Parts per million |
| CEE | Collaboration for Environmental Evidence guidelines |
| PECO | Population, Exposure, Comparator and Outcome eligibility criteria |
References
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Available online: https://www.ipcc.ch/report/ar6/wg1/ (assessed on 16th March 2025).
- Wheeler, R.M.; Spencer, L.E.; Bhuiyan, R.H.; Mickens, M.A.; Bunchek, J.M.; van Santen, E.; Massa, G.D.; Romeyn, M.W. Effects of elevated and super-elevated carbon dioxide on salad crops for space. Journal of Plant Interactions 2023, 19. [Google Scholar] [CrossRef]
- Bunce, J.A. Responses of soybeans and wheat to elevated CO2 in free-air and open top chamber systems. Field Crops Research 2016, 186, 78–85. [Google Scholar] [CrossRef]
- Dias, J.S. Nutritional quality and health benefits of vegetables: A review. Food and Nutrition Sciences 2012, 3, 1354–1374. [Google Scholar] [CrossRef]
- Myers, S.S.; Zanobetti, A.; Kloog, I. , Huybers, P.; Leakey, A.D.B.; Bloom, A.J.; Carlisle, E.; Dietterich, L.H.; Fitzgerald, G.; Hasegawa, T.; Holbrook, N.M.; Nelson, R.L.; Ottman, M.J.; Raboy, V.; Sakai, H.; Sartor, K.A.; Schwartz, J.; Seneweera, S.; Tausz, M.; Usui, Y. Increasing CO2 threatens human nutrition. Nature 2014, 510, 139–142. [Google Scholar] [CrossRef]
- Black, R.E.; Victora, C.G.; Walker, S.P.; Bhutta, Z.A.; Christian, P.; de Onis, M.; Ezzati, M.; Grantham-McGregor, S.; Katz, J.; Martorell, R.; Uauy, R. Maternal and child undernutrition and overweight in low-income and middle-income countries. The Lancet 2013, 382, 427–451. [Google Scholar] [CrossRef] [PubMed]
- Megan, W.; What are the health benefits of kale? Med. News Today 2020. Available online: https://www.medicalnewstoday.com/articles/270435.php (assessed on 8th November 2024).
- Wang, X.; Li, D.; Song, X. Elevated CO2 mitigates the effects of cadmium stress on vegetable growth and antioxidant systems. Plant, Soil and Environment 2023, 69, 202–209. [Google Scholar] [CrossRef]
- Ramezanifar, H.; Yazdanpanah, N.; Golkar Hamzee Yazd, H.; Tavousi, M.; Mahmoodabadi, M. Spinach growth regulation due to interactive salinity, water, and nitrogen stresses. J. Plant Growth Regul. 2022, 41, 1654–1671. [Google Scholar] [CrossRef]
- Šamec, D.; Urlić, B.; Salopek-Sondi, B. Kale (Brassica oleracea var. acephala) as a superfood: review of the scientific evidence behind. Crit. Rev. Food Sci. Nutr. 2018, 20, 1–37. [Google Scholar] [CrossRef]
- Tian, H.; Lu, C.; Pan, S.; Yang, J.; Miao, R.; Ren, W.; Yu, Q.; Fu, B.; Jin, F.-F.; Lu, Y.; Melillo, J.; Ouyang, Z.; Palm, C.; Reilly, J. Optimizing resource use efficiencies in the food–energy–water nexus for sustainable agriculture: from conceptual model to decision support system. Curr. Opin. Environ. Sustain. 2018, 33, 104–113. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAOSTAT Statistical Database: Production of Spinach 2023. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on day month year).
- United States Department of Agriculture. FoodData Central: Kale, raw 2020. Available online: https://fdc.nal.usda.gov (assessed on 20th September 2024).
- Public Health England. Composition of Foods Integrated Dataset (CoFID). London: Public Health England 2022. Available online: https://www.gov.uk/government/publications/composition-of-foods-integrated-dataset-cofid (assessed on 20th September 2024).
- Taub, D.R.; Miller, B.; Allen, H. Effects of elevated CO2 on the protein concentration of food crops: A meta-analysis. Global Change Biology 2008, 14, 565–575. [Google Scholar] [CrossRef]
- Shanker, A.K.; Gunnapaneni, D.; Bhanu, D.; Vanaja, M.; Lakshmi, N.J.; Yadav, S.K.; Prabhakar, M.; Singh, V.K. Elevated CO2 and Water Stress in Combination in Plants: Brothers in Arms or Partners in Crime? Biology 2022, 11, 1330. [Google Scholar] [CrossRef]
- Leakey, A.D.B.; Ainsworth, E.A.; Bernacchi, C.J.; Rogers, A.; Long, S.P.; Ort, D.R. Elevated CO2 effects on plant carbon, nitrogen, and water relations: Six important lessons from FACE. Journal of Experimental Botany 2009, 60, 2859–2876. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, A.; Reddy, K.R.; Walne, C.H.; Barickman, T.C.; Brazel, S.; Chastain, D.; Gao, W. Individual and Interactive Effects of Multiple Abiotic Stress Treatments on Early-Season Growth and Development of Two Brassica Species. Agriculture 2022, 12, 453. [Google Scholar] [CrossRef]
- Collaboration for Environmental Evidence. Guidelines and Standards for Evidence synthesis in Environmental Management 2022. Version 5.1 (Pullin, A.S.; Frampton, G.K.; Livoreil, B.; Petrokofsky, G. Eds). Available online: www.environmentalevidence.org/information-for-authors. (accessed on 18 February 2024).
- Newson, R.B.; Formulas for estimating and pooling Hedges’ g parameters in a meta-analysis 2020. Roger Newson Resources, 1-6. Available online: https://www.rogernewsonresources.org.uk/miscdocs/metahedgesg1.pdf (accessed on 18 February 2024).
- RStudio Team. RStudio: Integrated Development for R 2020. RStudio, PBC, Boston, MA URL. Available online: http://www.rstudio.com/ (accessed on 18 February 2024).
- Viechtbauer, W. Conducting meta-analyses in R with the metafor package. Journal of Statistical Software 2010, 36, 1–48. [Google Scholar] [CrossRef]
- Nakagawa, S.; Lagisz, M.; O'Dea, R.E.; Pottier, P.; Rutkowska, J.; Senior, A.M.; Yang, Y.; Noble, D.W.A. orchaRd 2.0: An R package for visualising meta-analyses with orchard plots. Methods in Ecology and Evolution 2023. [Google Scholar] [CrossRef]
- Li, F.; Guo, D.; Gao, X.; Zhao, X. Water Deficit Modulates the CO2 Fertilisation Effect on Plant Gas Exchange and Leaf-Level Water Use Efficiency: A Meta-Analysis. Frontiers in plant science 2021, 12, 775477. [Google Scholar] [CrossRef] [PubMed]
- Proietti, S.; Moscatello, S.; Giacomelli, G.A.; Battistelli, A. Influence of the interaction between light intensity and CO2 concentration on productivity and quality of spinach (Spinacia oleracea L.) grown in fully controlled environment. Advances in Space Research 2013, 52, 1193–1200. [Google Scholar] [CrossRef]
- Ritchie, H.; Reay, D.; Higgins, P. The impact of global dietary guidelines on climate change. Global Environmental Change 2018, 49, 46–55. [Google Scholar] [CrossRef]
- Rogers, A.; Ainsworth, E.A.; Leakey, A.D.B. Will elevated carbon dioxide concentration amplify the benefits of nitrogen fixation in legumes? Plant Physiology 2009, 151, 1009–1016. [Google Scholar] [CrossRef]
- Ekele, J.U.; Webster, R.; Perez de Heredia, F.; Lane, K.E.; Fadel, A.; Symonds, R.C. Current impacts of elevated CO2 on crop nutritional quality: a review using wheat as a case study. Stress Biol. 2025, 5, 34. [Google Scholar] [CrossRef]
- Fitzgerald, G.J.; Tausz, M.; O'Leary, G.; Mollah, M.R.; Tausz-Posch, S.; Seneweera, S.; Norton, R.M.; Fitzgerald, G.J. Elevated atmospheric CO2 can dramatically increase wheat yields in semi-arid environments and buffer against heat waves. Global Change Biology 2016, 22, 2269–2284. [Google Scholar] [CrossRef]
- Ziska, L.H. Rising Carbon Dioxide and Global Nutrition: Evidence and Action Needed. Plants (Basel) 2022, 11, 1000. [Google Scholar] [CrossRef] [PubMed]
- Nomura, K.; Yasutake, D.; Kaneko, T.; Takada, A.; Okayasu, T.; Ozaki, Y.; Mori, M.; Kitano, M. Long-term compound interest effect of CO2 enrichment on the carbon balance and growth of a leafy vegetable canopy. Scientia Horticulturae 2021, 283, 110060. [Google Scholar] [CrossRef]
- Jain, V.; Pal, M.; Raj, A.; Khetarpal, S. Photosynthesis and nutrient composition of spinach and fenugreek grown under elevated carbon dioxide concentration. Biol Plant 2007, 51, 559–562. [Google Scholar] [CrossRef]
- Jin, C.W.; Du, S.T.; Zhang, Y.S.; Tang, C.; Lin, X.Y. Atmospheric nitric oxide stimulates plant growth and improves the quality of spinach (Spinacia oleracea). Annals of Applied Biology 2009, 155, 113–120. [Google Scholar] [CrossRef]
- Reich, P.B.; Hobbie; S. E., Lee, T.; Ellsworth, D.S.; West, J.B.; Tilman, D.; Knops, J.M.; Naeem, S.; Trost, J. Nitrogen limitation constrains sustainability of ecosystem response to CO2. Nature 2006, 440, 922–925. [Google Scholar] [CrossRef]
- Loladze, I. Hidden shift of the ionome of plants exposed to elevated CO2 depletes minerals at the base of human nutrition. eLife 2014, 3, e02245. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.R.; Myers, S.S. Impact of anthropogenic CO2 emissions on global human nutrition. Nature Climate Change 2018, 8, 834–839. [Google Scholar] [CrossRef]
- La, G.X.; Fang, P.; Teng, Y.B.; Li, Y.J.; Lin, X.Y. Effect of CO2 enrichment on the glucosinolate contents under different nitrogen levels in bolting stem of Chinese kale (Brassica alboglabra L.). J. Zhejiang Univ. Sci. B. 2009, 10, 454–64. [Google Scholar] [CrossRef]
- Erwin, J.; Gesick, E. Photosynthetic responses of swiss chard, kale, and spinach cultivars to irradiance and carbon dioxide concentration. HortScience 2017, 52, 706–712. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. National Diabetes Statistics Report 2022. Available online: https:://www.cdc.gov/. (accessed on 10 September 2024).
- Hruby, A.; Hu, F.B. The epidemiology of obesity: a big picture. Pharmacoeconomics 2015, 33, 673–689. [Google Scholar] [CrossRef]
- Parker, E.D.; Lin, J.; Mahoney, T.; Ume, N.; Yang, G.; Gabbay, R.A.; ElSayed, N.A.; Bannuru, R.R. Economic Costs of Diabetes in the U.S. in 2022. Diabetes care 2024, 47, 26–43. [Google Scholar] [CrossRef]
- Bloom, A.J.; Burger, M.; Asensio, J.S.R.; Cousins, A.B. Carbon dioxide enrichment inhibits nitrate assimilation in wheat and Arabidopsis. Science 2010, 328, 899–903. [Google Scholar] [CrossRef]
- Sehgal, A.; Reddy, K.R.; Walne, C.H.; Barickman, T.C.; Brazel, S.; Chastain, D.; Gao, W. Climate Stressors on Growth, Yield, and Functional Biochemistry of two Brassica Species, Kale and Mustard. Life 12, 1546. [CrossRef] [PubMed]
- World Health Organization. Healthy diets. (WHO fact sheet series 2020). Available online: https://www.who.int/news-room/fact-sheets/detail/healthy-diet (accessed on 10 May 2024).
- Chowdhury, M.; Kiraga, S.; Islam, M.N.; Ali, M.; Reza, M.N.; Lee, W.H.; Chung, S.O. Effects of Temperature, Relative Humidity, and Carbon Dioxide Concentration on Growth and Glucosinolate Content of Kale Grown in a Plant Factory. Foods (Basel, Switzerland) 2021, 10, 1524. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef]
- Zhu, C.; Naqvi, S.; Gomez-Galera, S.; Pelacho, A.M.; Capell, T.; Christou, P. Transgenic strategies for the nutritional enhancement of plants. Trends in Plant Science 2018, 12, 548–555. [Google Scholar] [CrossRef]
- World Health Organization. Micronutrient deficiencies – malnutrition 2024. Available online: https://www.who.int/news-room/fact-sheets/detail/malnutrition (accessed on 16 May 2024).
- Tisserat, B.; Herman, C.; Silman, R.; Bothast, R.J. Using Ultra-high Carbon Dioxide Levels Enhances Plantlet Growth In Vitro. HortTechnology 1997, 7, 282–289. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant, Cell & Environment 2007, 30, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Haworth, M.; Hoshika, Y.; Killi, D. Has the Impact of Rising CO2 on Plants been Exaggerated by Meta-Analysis of Free Air CO2 Enrichment Studies? Frontiers in plant science 2016, 7, 1153. [Google Scholar] [CrossRef]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proceedings of the National Academy of Sciences (21 November 2011) 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
- Reekie, G.E. , MacDougall, G., Wong, I. & Hicklenton, P.R. Effect of sink size on growth response to elevated atmospheric CO2 within the genus Brassica. Canadian Journal of Botany 1998, 76, 829–835. [Google Scholar] [CrossRef]
Figure 1.
(a) Effect size distribution per classified outcome category; (b) Effect size distribution per period of CO2 exposure (time); (c) Periodic pattern of the release of articles.
Figure 1.
(a) Effect size distribution per classified outcome category; (b) Effect size distribution per period of CO2 exposure (time); (c) Periodic pattern of the release of articles.
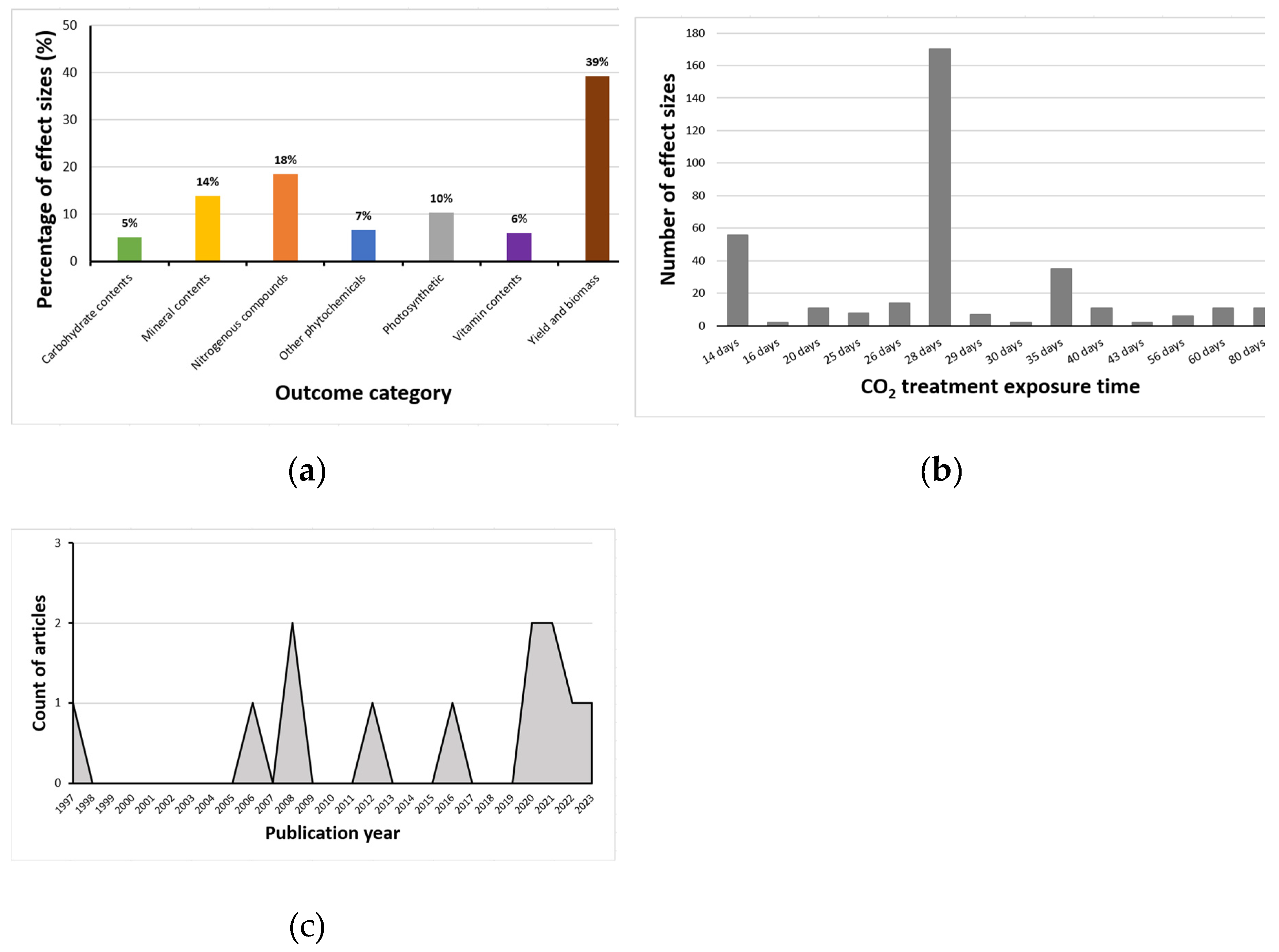
Figure 2.
Effect sizes' geographical dispersion
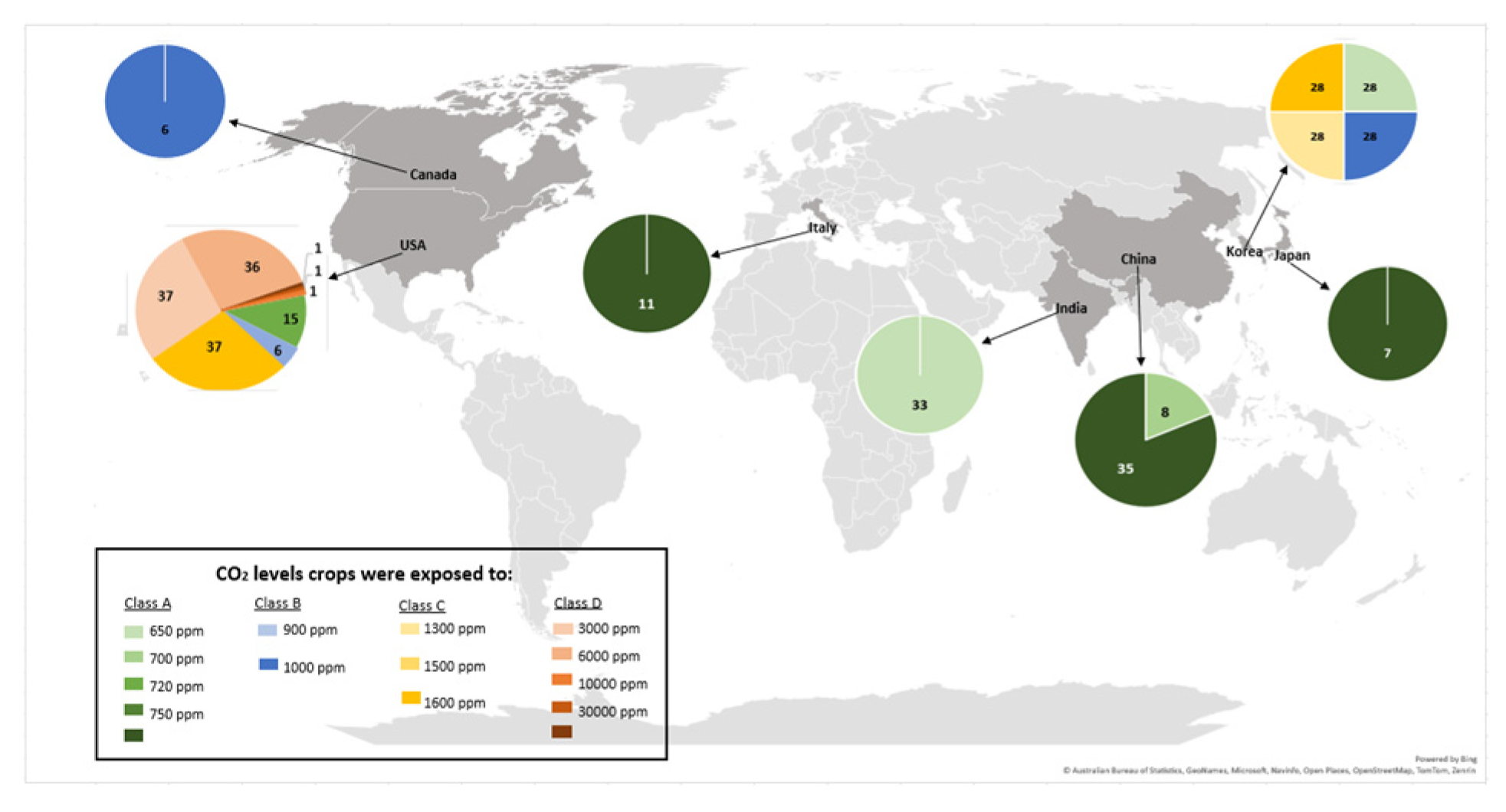
Figure 6.
For kale crops only (a) Effect of eCO2 on carbohydrate contents with constituent type as a moderator. (b) Effect of eCO2 on vitamin contents only. (c) Effect of eCO2 on photosynthetic components contents. (d) Effect of eCO2 on non-classed phytochemical contents. (e) Effect of eCO2 on mineral contents. (f) Effect of eCO2 on nitrogenous compounds. (g) Effect of eCO2 on yield and biomass components.
Figure 6.
For kale crops only (a) Effect of eCO2 on carbohydrate contents with constituent type as a moderator. (b) Effect of eCO2 on vitamin contents only. (c) Effect of eCO2 on photosynthetic components contents. (d) Effect of eCO2 on non-classed phytochemical contents. (e) Effect of eCO2 on mineral contents. (f) Effect of eCO2 on nitrogenous compounds. (g) Effect of eCO2 on yield and biomass components.
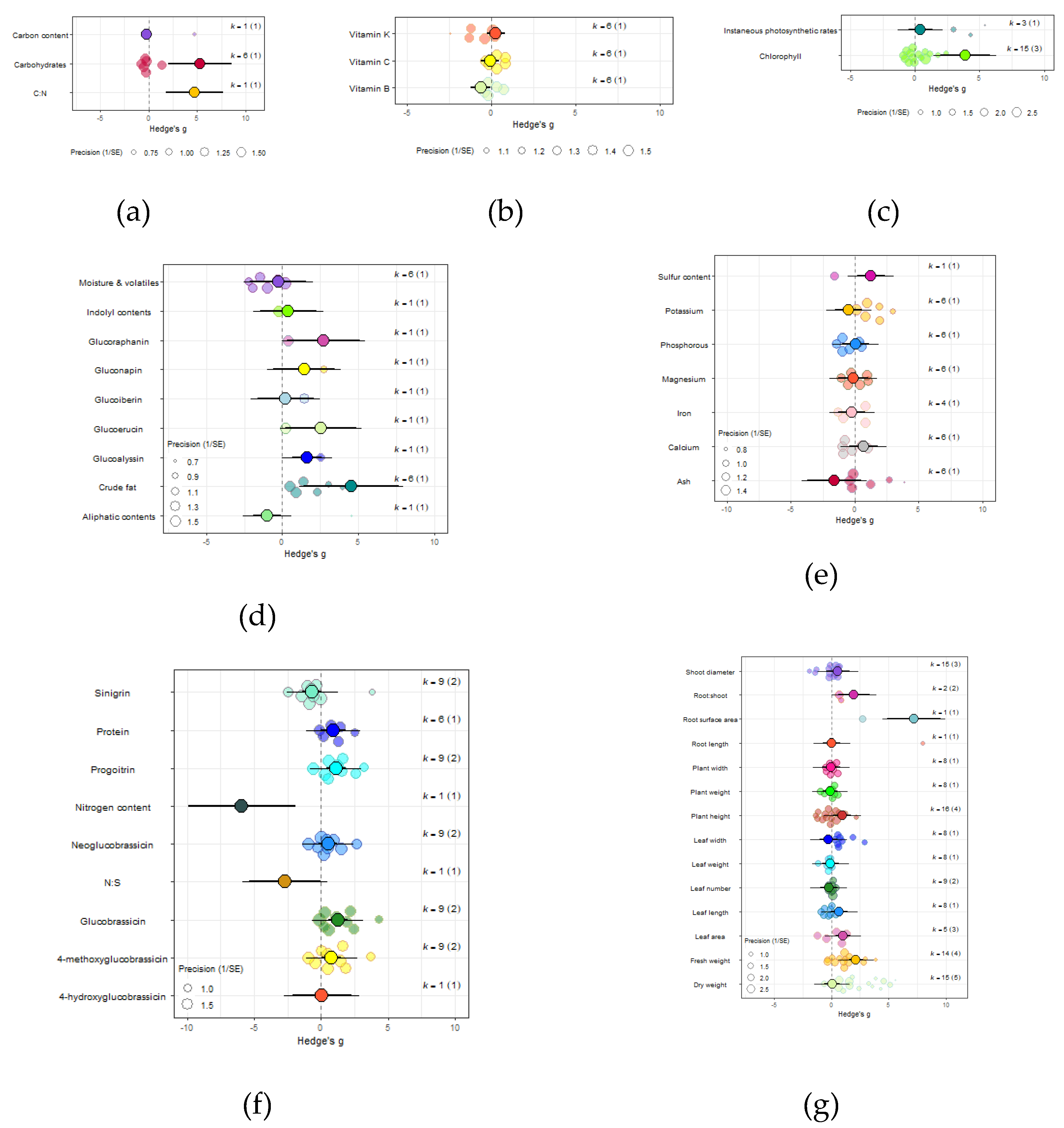
Figure 7.
For spinach crops only (a) Effect of eCO2 on nitrogenous compounds with constituent type as a moderator. (b) Effect of eCO2 on carbohydrate contents. (c) Effect of eCO2 on mineral contents. (d) Effect of eCO2 on vitamin contents. (e) Effect of eCO2 on non-classed phytochemical contents. (f) Effect of eCO2 on yield components. (g) Effect of eCO2 on photosynthetic components.
Figure 7.
For spinach crops only (a) Effect of eCO2 on nitrogenous compounds with constituent type as a moderator. (b) Effect of eCO2 on carbohydrate contents. (c) Effect of eCO2 on mineral contents. (d) Effect of eCO2 on vitamin contents. (e) Effect of eCO2 on non-classed phytochemical contents. (f) Effect of eCO2 on yield components. (g) Effect of eCO2 on photosynthetic components.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



