
Submitted:
28 November 2025
Posted:
01 December 2025
You are already at the latest version
Abstract
The success of invasive species relies heavily on the production, dispersal and genetic composition of propagules. For range expanding species, breeding strategy and level of reproductive investment will strongly in-fluence their capacity to establish and invade new areas. A hermaphroditic lifestyle provides the advantage of increasing the number of seed bearing individuals within a population while a dioecious habit may enable more rapid adaptation to new environments, improve resource use efficiency, fecundity and dispersal. Pittosporum undulatum, a tree native to coastal areas of southeastern Australia, has many characteristics of an invasive species within and beyond its native range. A previous study detected a male bias within invasive populations, with a high proportion of fruit deriving from female-only trees, leading to recommendations the removal of ‘matriarch’ trees as a simple management technique. We expanded that study and investigated different breeding systems populations of P. undulatum by assessing tree density, gender, resource availability and fruit load of individuals in 14 populations sited along the spectrum from native to invasive populations. All populations were comprised of either females or hermaphrodites. No male-only trees observed within the study. More females produced more fruit than hermaphrodites, especially in native site. This could not be attributed to environmental differences between sites. These data support the current management practices of targeting the removal of females as a simple method for containing invasions given the benefits of reducing the workload and spreading limited management resource. Our work highlights the value in understanding the breeding strategy employed by focal invasive species as a means of developing improved and more targeted control methods.
Keywords:
breeding strategy
; gynodioecious
; Pittosporum undulatum
; resource allocation
; fruit load
; forest management
; invasive plants
; flowering
; flowers
1. Introduction
The success of invasive species relies heavily on the production, dispersal and genetic composition of propagules [1]. Relative to their native range, invasive species commonly show increases in fecundity [2] and self-compatibility [3]. Thus, a focus on the mating systems and reproductive allocations of invasive species may improve our understanding of the processes that promote the establishment and expansion of invasive populations [1,2,4] and potentially guide control strategies[1,5,6,7].
Baker[8] considered the role of self-compatibility in the colonisation of islands. Due to the haphazard nature of long distance dispersal of propagules to islands, the likelihood of establishing within close proximity to a potential mate is thought to be low. Thus, hermaphroditic species with a self-compatible breeding system should predominate among island colonisers, as they have the capacity for uniparental reproduction through self-pollination[9]. The same concept can be extended to an invasion front, where individuals may be establishing in relative isolation[10]. Baker’s Law, as this idea is now referred to, implies that in invasive plants self-compatible hermaphrodite reproductive systems might predominate, especially at the margins of an invasive range.
Any initial advantage that hermaphrodites may have in founding of new populations need not persist as a population grows[11]. Metapopulation dynamics of an invasion may mean that female plants or hermaphroditic plants that have female-biased sexual allocation are favoured during the initial phase of population expansion, as females will tend to contribute disproportionately to early population growth[12,13]. Only later in consolidated populations with higher population density and female availability would the selective advantage of males and male allocation increase. Consistent with this, high seed fecundity is also a notable features of individuals in invasive populations[2,14]. We might also expect invasive populations to shift their breeding systems toward gynodioecy or toward a greater proportion of female plants in already gynodioecious species[15].
In large, well established invasive populations, novel resource environments may alter the sex expression of plants or skew flowering sex ratios[16]. Individual plants in diclinous species can often change sex expression according to their size or resource status[17,18], and individuals of monoecious species are known to shift their floral sex ratios in response to resource availability[19]. In these cases, female expression was almost universally favoured by greater moisture, soil nutrients, or sunlight. Some degree of segregation of male and females along resource gradients has been noted in dioecious species, with males tending to predominate at the drier, poorer end of the gradient and females at more fertile[20,21]. If invasive ranges offer a more favourable resource environment, for example through greater resource availability[22], we might expect to find a female bias in sex expression or sex occurrence among plants within those invasive populations. Alternatively, if permissive environments allow small plants to survive, and small size favours male expression, invasive populations may show male-biased sex expression of sex ratios[16].
Suites of morphological and life history traits that promote dispersal are likely to be exaggerated at an invasion front because the best dispersers are most likely to arrive at the front, mate with other recent arrivals, and, to the extent that traits are heritable, pass them to the next generation that itself may extend the front [23,24] Phillips, 2010 #57}[25]. This idea, which has been called the ‘Olympic Village effect’ in the context of movement adaptations for animal dispersal[26], suggests that, in invasive plants, traits that affect dispersal and establishment (e.g. of seeds and fruits) might differ between the native and introduced ranges. Such processes seem to account for lower wing loadings (thus, slower descent and more horizontal dispersal) of the winged seeds in more recently derived populations of Pinus contorta following its post-glacial range expansion in North America[27]. For species with animal-dispersed fruits, we might expect selection for dispersal ability to favour greater fruit load and higher probability of fruiting[2,28]. Large seed size, in contrast, is thought to diminish dispersal ability but may enhance competitive ability and stress tolerance during seedling establishment[29,30].
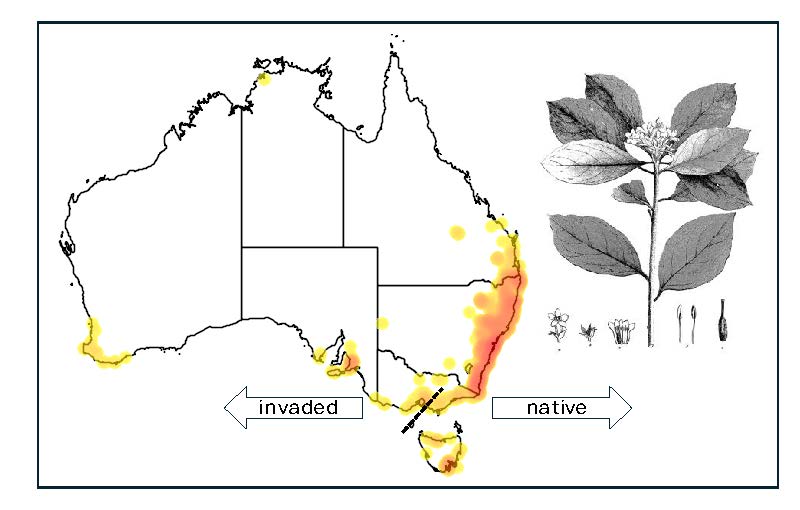
Pittosporum undulatum Vent. (Sweet Pittosporum) a long-lived woody invader with many characteristics of an invasive species within and beyond its native range of coastal south-eastern Australia[31,32,33,34] (Figure 1). Within Australia, sites dominated by P. undulatum have reduced biodiversity and species richness of both plants and birds[33,35,36], resulting calls for its control, despite its ‘native’ status. ‘Native species’ in Australia are typically defined as ones that were present at the time of European invasion, with those that are first recorded after this date regarded as ‘naturalised’[37,38]. Those whose ranges are still expanding are typically referred to as ‘invasive’. While this simplistic concept ignores the introduction and widespread translocation of plants by First Peoples prior to this date[39,40], we believe the terminology is appropriate for P. undulatum, as there is clear change in distribution since Europeans arrived [35]. It is also invasive in New Zealand, Portugal, Jamaica, Hawaii, and is an emergent weed in South Africa [31,41,42,43,44].
Previous studies indicated that individual P. undulatum trees can be male, female or hermaphroditic, but how this relates to its environment or invasive status remains unknown[41,45,46]. Here we measured the frequency of sexual types in P. undulatum sampled from populations growing across a spectrum from ‘native’ to ‘non-native’ locations. We also assessed the probability of fruit set, fruit load and seed size, traits that may affect the ability to reach and establish in novel sites. We hypothesise that P. undulatum from invasive populations will: (1) have a higher proportion of females than native populations; (2) produce relatively more fruit; and (3) have a greater number of seeds and/or smaller seeds, relative to native populations. In addition, in order to determine whether seed production was correlated with resource availability, we measured the concentration of total nitrogen , total carbon and carbon isotope discrimination (δC13) in leaves of trees at each site
2. Materials and Methods
2.1. Study Species: Pittosporum undulatum Vent. (Sweet Pittosporum)
P. undulatum is found in a range of habitats but is most commonly found in the temperate rainforests[31]. Horticultural propagation followed by altered fire regimes and the introduction of new avian vectors such as European blackbirds (Turdus merula L.) and have all contributed to the spread of this species across mainland south-eastern Australia [31,32,33,35,47,48,49]. P. undulatum is known to establish quickly after disturbance, although it can also become invasive at undisturbed locations[46,48,50]. Once established, mature trees can reach heights of 8-30 m. Individuals form dense canopies, shading out the undergrowth and reducing structural diversity, floristic composition and the integrity of ecological systems[31,33,51]. Original theories of it being allelopathic are now largely discredited[31,34].
2.2. Site Description
We investigated variations in tree density, sex, resource availability and fruit load for native and invasive populations of P. undulatum. Seven populations within temperature Eucalyptus forests of East Gippsland in southeastern Victoria, Australia, were selected to represent native populations (Table 1). A further seven populations across peri-urban areas of Melbourne, in south-eastern Australia, were selected to represent invasive populations (Table 1).
2.3. Sex Determination and Resource Analysis
Sex expression of individual P. undulatum plants was determined during Spring (September–October 2016; Table 1). At each site a 20 m x 20 m quadrat was established and mature trees growing within the quadrat were examined for the presence or absence of male and female floral structures and labelled. Within each quadrat, five leaves from five randomly selected individuals of each sex were sampled for nutrient analysis, specifically leaf nitrogen, carbon13 and chlorophyll content. Leaves were selected from approximately the third stem of a branch. Leaves were dried in an oven for 48 hours at 60 degrees before being ground in a homogenizing tissue mill. Total elemental nitrogen, carbon and δC13 were measured on finely ground freeze dried leaf samples using a LECO CNS2000 analyser (Environmental Analysis Laboratory, Southern Cross University, NSW, Australia).
2.4. Fruit Load and Seed Mass Determination
The original sample plots were re-examined six months later (between 1 Februry-20 March 2017). True fruit loads were assessed for the same individual plants as above using a ranking system from 0–11 where a rank of 0 equated to no capsules observed; 1 equated up to 50 capsules, 2 equated to 50-100 and so on up to 11 which equated to 500-550 capsules (the maximum observed). Capsules were collected and the seeds removed from their fruit casing and cleaned of mucilage using tissue paper. All seeds from each fruit were weighed together and the mean seed weight calculated.
2.5. Statistical Analysis
All analysis was conducted using the R statistical program[52]. Variation in mature plant density, proportion of females within a populations across native and invasive populations, and differences in seed number and weight across populations were analysed through unpaired t tests. Differences between native and invasive sites were compared using equal variance t tests. Differences between each reproductive type in the proportion of fruiting trees were examined through a generalised linear model. Comparison of the mean fruit load rank between reproductive types across populations were also examined via a generalised linear model. Linear modelling was also used to investigate variation in the proportion of individuals producing fruit, and the mean rank of fruit production across sex and origin. Data was arcsine square root and cube root transformed respectively prior to analysis to meet the conditions of normality. Non-linear modelling was used to analyse the influence of nutrient availability to fruit production. Statistical tables are included in the Supplementary Information.
3. Results
3.1. Reproductive Traits
All populations were comprised of either females or hermaphrodites, with no male trees observed within the study. Native and invasive sites did not differ significantly in the mean density in the proportion of females (P=0.217; Table 2).
No significant difference was detected in the proportion of fruiting individuals between populations of different origins either (P= 0.469; Figure 2; Supplementary Table S2). There were, however, differences at a population level in the presence of capsules with significantly more fruit set from female, compared to hermaphrodite flowers (Figure 2A; Supplementary Table S3). Females in native sites were also more likely to fruit comparative to females in invasive populations (P<0.01; Supplementary Table S3). When examining the quantity of fruit produced by each tree, female individuals appeared more likely to produce higher quantities of fruit (Figure 2B; Supplementary Table S4). The mean mass of seeds produced in invasive populations was approximately twice that of seeds from native populations (Table 2; Supplementary Table S5).
3.2. Resource Availability: Tree Density and Leaf N, C and ∂C13
Native and invasive sites did not differ significantly in the mean density of P. undulatum trees (P=0.718; Table 2; Supplementary Table S4). Little variation was observed in ∂C13 and total C concentration among populations regardless of origin. A weak correlation was detected between leaf nitrogen and fruit production (slope estimate= 5.540, adjusted r2 = 0.23) (Figure 3A, 3B, 3C and Figure 4).
4. Discussion
Investigating how the breeding system and sexual expression of invasive species differs between native and novel environments may improve our understanding of pest species and their management[26,53,54,55,56]. Furthermore, contrasting the performance of species within and beyond their native ranges can be use useful in testing ecological theory[4,57,58,59]. This study explored the breeding strategy and reproductive ecology of a range expanding invasive tree and considered how these factors varied between native and invasive populations. We expected to find a high level of female sex expression among plants in invasive populations, because higher numbers of females are likely to enhance population expansion at the invasion front through seed production [12]. We also anticipated a higher investment in reproductive traits for invasive populations, as these traits are likely to both extend the dispersal capacity of seeds and improve their chance of survival to maturity within novel environments. However, populations present a mosaic of both sex ratio and female traits, and our expectations could be met in different ways.
4.1. The Proportion of Female and Hermaphrodite Trees Was Similar Across All Sites
The most striking observation in this study was the lack of male trees within any population regardless of its native or invasive origin. Populations were comprised of either females or hermaphroditic individuals. This is surprising as male-bias is more common in trees and in plants that depend on biotic seed dispersal[60]. Importantly, the lack of male trees is in stark contrast to previous studies exploring P. undulatum populations within southeastern Victoria, that instead showed a male bias within invasive population[45,46]. A hermaphroditic bias has been observed within an invasive population of P. undulatum in Jamaica[41]. Of the 60 trees sampled in their study, Goodland and Healey [41] found 78.4% of individuals to be hermaphroditic, with the remainder female. Previous work in Victoria has suggested females make up between 30%-40% of invasive P. undulatum populations, with the remainder being male [46], whilst Mullet [45]found approximately 9% of male flowering plants went on to produce fruit. These distinct results suggest that the proportion of males, females and hermaphrodites making up P. undulatum populations may be highly variable and certainly less consistent then previously proposed.
Theory suggests hermaphroditism should be more common in populations with younger age cohorts and in lower densities [11,15]. This theory could imply that as invasive populations of our study are relatively younger (approximately 10-30 years) they should be less dense and therefore support higher proportions of hermaphrodites relative to older aged native populations. Instead, our sample populations showed an approximate even proportion of female and hermaphroditic individuals, and though variable, a similar density of individuals across populations, in both cases regardless of their native or invasive origin. Our prediction of greater female representation in invasive populations has therefore not been met. One possibility is that despite the younger age of invasive P. undulatum populations, the equivalent density of trees at native and invasive sites means that the selective pressure for a higher proportion of females may not be as strong for the established invasive populations of this study, comparative to a population at the very early stages of invasion/range expansion, where tree density is lower.
4.2. Fruit Production Was Higher in Female Trees in the Native Range
Female trees were far more likely to produce fruit than hermaphrodites, with significantly higher fruit loads, suggesting that hermaphrodites are predominately filling the role of males as pollen producers. Baker’s law postulates the selective advantage that hermaphrodites may have due to their capacity to self-pollinate in environments where mates are sparse[8]. The strong persistence of females within all populations together with observations of hermaphrodites generally fulfilling the male role within P. undulatum populations suggest there is no strong pressure for self-fertilisation. Given the high stand density that P. undulatum populations can reach along with the consistent and ongoing introduction of P. undulatum to invasion sites[31,33,47], mate proximity may not be an issue, which would therefore reduce selective pressure for self-fertilisation. In this instance, enhanced reproductive and growth traits may present a stronger selective advantage, improving the capacity to establish, develop and reproduce within varied and disturbed environments.
Contrary to expectations, females from native populations were more likely to fruit relative to those from the invasive range. Higher fruit set in females in native populations, may indicate that individuals within invasive populations might not be reaching their full fruiting capacity. A variety of factors, such as the level of disturbance, reduced pollination services or presence of facilitator species could contribute to this[61,62]. Regardless, the result may be of concern for land managers, as the ongoing naturalisation of invasive populations may potentially reduce this fruiting constraint and therefore the expansion of invasive populations.
4.3. Seed Size Was Greater in the Native Range
Seeds produced in the invasive range were significantly larger than in the native range. Seed mass reflects maternal investment, with larger mass representing a potential establishment advantage through greater stored resources. Findings of greater seed mass within invasive populations throughout the literature is mixed, with data supporting [63,64,65] and failing to support [66,67] the hypothesis. Large seed typically do not disperse as far as smaller seeds. This may explain, at least in part, the nucleation-style pattern of invasion seen in P. undulatum[35]. Heavier seeds may be advantageous to populations expanding into new sites as a larger seed mass may improve the capacity for a plant to withstand the unpredictable environments found at novel and disturbed sites[68,69]. Because humans have assisted the introduction of P. undulatum to novel environments, there may not have been the strong selection on dispersal traits that we expected based on studies of other invasive plants. However, traits that enhance seedling establishment and thus population growth may be favoured in young populations[12,13]. Moreover, the high levels of specialised metabolites may enhance tolerance to abiotic and biotic stresses[34]. Avian seed dispersal may be important to P. undulatum dispersal into new sites, which may further emphasise the benefits of enhanced establishment traits over dispersal characteristics[35,44].
4.4. Influence of Resources on Seed and Fruit Production
Production of fleshy fruits is resource intensive, thus females within dioecious populations are likely to incur higher reproductive costs[70]. However, pollen production may also require high nitrogen resources which may in part explain why both females and hermaphrodites had similar levels of nitrogen[71]. Female trees with higher leaf nitrogen concentrations tended to have a higher fruit load. The scaling exponent of 5.54 is much higher than unity, and indicates the dramatic benefits increased nitrogen availability may have on fruit production. Exponent values of this magnitude have been found in other plant species[72]. The scaling factor of nitrogen availability on fruit production has implications on management of the species, as populations downstream from agricultural areas may be more prone to range expansion. Population density was similar in all sites and cannot be considered a factor in affecting flowering, sex or fruit production.
5. Conclusions: Management and Control of P. undulatum
Our work highlights the value in understanding the breeding strategy employed by a focal invasive species as a means of developing improved and more targeted control methods. Current management practises focus on removing females from populations where the range is expanding, with a focus on trees that have large fruit loads, as these are the individuals that promote further colonising spread via seed dispersal[46]. This action has the benefit of reducing the workload and spreading limited management resources. Though our work has observed evidence of hermaphroditic individuals producing fruit, a general trend for significantly greater fruit production and seed size in females would support their targeted removal, as proposed by Gleadow and Walker [46]. However, this policy assumes sex ratios are consistent across all populations, and that the breeding system displayed by individuals remains fixed, regardless of fluctuations in resources, time and stochastic factors. Anecdotal reports suggest sexual labiality in P. undulatum may present a factor in the species control, with the removal of all females from a population in one season, followed by the production of fruit by trees previously considered “male”. Further study of this matter is advocated as a means of improving management efforts.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Supplementary Table S1. t test comparing differences in total tree density, the proportion of females and proportion of fruiting individuals among native and invasive populations of Pittosporum undulatum sampled across Victoria; Supplementary Table S2. Generalized Linear model comparing variation in the proportion of fruit producing individuals between female and hermaphroditic trees from native and invasive populations. Supplementary Table S3. Linear model comparing variation in ranked fruit production between female and hermaphrodite trees from native and invasive populations.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used: Conceptualization, B.O’L., R.G., M.B. and S.V..; methodology, B.O’L., M.B. and S.V.; software, B.O’L.; validation, B.O’L. and M.B..; formal analysis, B.O’L.; investigation, B.O’L.; resources, R.G.; data curation, B.O’L.; writing—original draft preparation, B.O’L.; writing—review and editing, R.G.., MB. and S.V.; visualization, B.O’L., R.G.; supervision, R.G., M.B. and S.V.; project administration, R.G.; funding acquisition, R.G.. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
This research was funded in park through the Parks Victoria Research Partners Program. O’Leary was supported through an Australian Government Research Training Program (RTP).
Data Availability Statement
Data is contained within the article or supplementary material.
Acknowledgments
This research was supported by an Australian Government Research Training Program (RTP) Scholarship. The authors acknowledge Parks Victoria, Melbourne Water, Yarra Ranges Council and Mornington Peninsula Council for their support and site access. The authors also thank all field volunteers and StopPitt for assistance and guidance.The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Barrett, S.C.H., R.I. Colautti, and C.G. Eckert, Plant reproductive systems and evolution during biological invasion. Molecular Ecology, 2008. 17(1): p. 373-383. [CrossRef]
- Burns, J.H., et al., Greater sexual reproduction contributes to differences in demography of invasive plants and their noninvasive relatives. Ecology, 2013. 94(5): p. 995-1004. [CrossRef]
- Petanidou, T., et al., Self-compatibility and plant invasiveness: Comparing species in native and invasive ranges. Perspectives in Plant Ecology Evolution and Systematics, 2012. 14(1): p. 3-12. [CrossRef]
- Carvalho, C., et al., Multivariate selection mediated by aridity predicts divergence of drought-resistant traits along natural aridity gradients of an invasive weed. New Phytologist, 2022. 234(3): p. 1088-1100. [CrossRef]
- Lambrinos, J.G., How interactions between ecology and evolution influence contemporary invasion dynamics. Ecology, 2004. 85(8): p. 2061-2070. [CrossRef]
- Barrett, S.C.H., Why reproductive systems matter for the invasion biology of plants, in Fifty Years of Invasion Ecology: The Legacy of Charles Elton. 2011. p. 195-210.
- Albert, A., et al., Strategies for a successful plant invasion: the reproduction of Phragmites australis in north-eastern North America. Journal of Ecology, 2015. 103(6): p. 1529-1537. [CrossRef]
- Baker, H.G., Self compatibility and establishment after long distance dispersal. Evolution, 1955. 9(3): p. 347-349. [CrossRef]
- Grossenbacher, D.L., et al., Self-compatibility is over-represented on islands. New Phytologist, 2017. 215(1): p. 469-478.
- Pannell, J.R., et al., The scope of Baker’s law. New Phytologist, 2015. 208(3): p. 656-667. [CrossRef]
- Eppley, S.M. and J.R. Pannell, Density-dependent self-fertilization and male versus hermaphrodite siring success in an androdioecious plant. Evolution, 2007. 61(10): p. 2349-2359. [CrossRef]
- Pannell, J., The maintenance of gynodioecy and androdioecy in a metapopulation. Evolution, 1997. 51(1): p. 10-20.
- Eppley, S.M. and J.R. Pannell, Sexual systems and measures of occupancy and abundance in an annual plant: Testing the metapopulation model. American Naturalist, 2007. 169(1): p. 20-28. [CrossRef]
- Bossdorf, O., et al., Phenotypic and genetic differentiation between native and introduced plant populations. Oecologia, 2005. 144(1): p. 1-11. [CrossRef]
- Pannell, J.R., et al., Gender variation and transitions between sexual systems in Mercurialis annua (Euphorbiaceae). International Journal of Plant Sciences, 2008. 169(1): p. 129-139. [CrossRef]
- Abe, T., et al., Sex ratio variation of Bischofia javanica Bl. (Euphorbiaceae) between native habitat, Okinawa (Ryukyu Islands), and invaded hab itat, Ogasawara (Bonin Islands). International Journal of Plant Sciences, 2002. 163(6): p. 1011-1016. [CrossRef]
- Freeman, D.C., K.T. Harper, and E.L. Charnov, Sex change in plants - old and new observations and new hypotheses. Oecologia, 1980. 47(2): p. 222-232. [CrossRef]
- Delph, L.F. and D.E. Wolf, Evolutionary consequences of gender plasticity in genetically dimorphic breeding systems. New Phytologist, 2005. 166(1): p. 119-128.
- Freeman, D.C., et al., Influence of environment on the floral sex-ratio of monoecious plants. Evolution, 1981. 35(1): p. 194-197.
- Freeman, D.C., L.G. Klikoff, and K.T. Harper, Differential resource utilization by sexes of dioecious plants. Science, 1976. 193(4253): p. 597-599. [CrossRef]
- Bierzychudek, P. and V. Eckhart, Spatial segregation of the sexes of dioecious plants. American Naturalist, 1988. 132(1): p. 34-43. [CrossRef]
- Davis, M.A., J.P. Grime, and K. Thompson, Fluctuating resources in plant communities: a general theory of invasibility. Journal of Ecology, 2000. 88(3): p. 528-534. [CrossRef]
- Travis, J.M.J. and C. Dytham, Dispersal evolution during invasions. Evolutionary Ecology Research, 2002. 4(8): p. 1119-1129.
- Simmons, A.D. and C.D. Thomas, Changes in dispersal during species’ range expansions. American Naturalist, 2004. 164(3): p. 378-395. [CrossRef]
- Tabassum, S. and M.R. Leishman, Have your cake and eat it too: greater dispersal ability and faster germination towards range edges of an invasive plant species in eastern Australia. Biological Invasions, 2018. 20(5): p. 1199-1210. [CrossRef]
- Phillips, B.L., G.P. Brown, and R. Shine, Life-history evolution in range-shifting populations. Ecology, 2010. 91(6): p. 1617-1627.
- Cwynar, L.C. and G.M. Macdonald, Geographical variation of lodgepole pine in relation to population history. American Naturalist, 1987. 129(3): p. 463-469. [CrossRef]
- Naniwadekar, R., C. Mishra, and A. Datta, Fruit resource tracking by hornbill species at multiple scales in a tropical forest in India. Journal of Tropical Ecology, 2015. 31: p. 477-490. [CrossRef]
- Westoby, M., M. Leishman, and J. Lord, Comparative ecology of seed size and dispersal. Philosophical Transactions of the Royal Society B-Biological Sciences, 1996. 351(1345): p. 1309-1317. [CrossRef]
- Turnbull, L.A., M. Rees, and M.J. Crawley, Seed mass and the competition/colonization trade-off: a sowing experiment. Journal of Ecology, 1999. 87(5): p. 899-912. [CrossRef]
- Gleadow, R.M. and D.H. Ashton, Invasion by Pittosporum undulatum of the forests of central Victoria. I Invasion patterns and plant morphology. Australian Journal of Botany, 1981. 29(6): p. 705-720. [CrossRef]
- Gleadow, R.M., K.S. Rowan, and D.H. Ashton, Invasion by Pittosporum undulatum of the forests of central Victoria. IV Shade tolerance. Australian Journal of Botany, 1983. 31(2): p. 151-160. [CrossRef]
- O’Leary, B., et al., Integrating the Passenger-Driver hypothesis and plant community functional traits to the restoration of lands degraded by invasive trees. Forest Ecology and Management, 2018. 408: p. 112-120. [CrossRef]
- Pasquini, D., et al., Is the invasiveness of Pittosporum undulatum in eucalypt forests explained by the wide-ranging effects of its secondary metabolites? Forests, 2023. 14(1): p. 23. [CrossRef]
- Gleadow, R.M., Invasion by Pittosporum undulatum of the forests of central Victoria. II Dispersal, germination and establishment. Australian Journal of Botany, 1982. 30(2): p. 185-198. [CrossRef]
- O’Leary, B.A., et al., Bird community recovery following removal of an invasive tree. Ecological Solutions and Evidence, 2021. 2(2). [CrossRef]
- Gallagher, R.V. and M.R. Leishman, Invasive plants and invaded ecosystems in Australia: implications for biodiversity, in Austral Ark: The State of Wildlife in Australia and New Zealand, A. Stow, N. Maclean, and G.I. Holwell, Editors. 2014, Cambridge University Press. p. 105-133.
- Fensham, R.J. and B. Laffineur, Defining the native and naturalised flora for the Australian continent. Australian Journal of Botany, 2019. 67((1)): p. 55-69. [CrossRef]
- Silcock, J.L., Aboriginal translocations: The intentional propagation and dispersal of plants in Aboriginal Australia. Journal of Ethnobiology, 2025. 38(3): p. 390-405. [CrossRef]
- McNiven, I.J. and L. Russell, Innovation: Knowledge and Ingenuity. First Knowledges Innovation, ed. M. Neale. 2023, Australia: Thames and Hudson. 256.
- Goodland, T. and J.R. Healey, The invasion of Jamaican montane rainforests by the Australian tree Pittosporum undulatum. 1996, School of Agricultural and Forest Sciences, University of Wales: Bangor, UK. p. 55.
- Hortal, J., et al., Assessing the areas under risk of invasion within islands through potential distribution modelling: The case of Pittosporum undulatum in Sao Miguel, Azores. Journal for Nature Conservation, 2010. 18(4): p. 247-257. [CrossRef]
- Lourenço, P., et al., Distribution, habitat and biomass of Pittosporum undulatum, the most important woody plant invader in the Azores Archipelago. Forest Ecology and Management, 2011. 262(2): p. 178-187. [CrossRef]
- Mokotjomela, T.M., C.F. Musil, and K.J. Esler, Frugivorous birds visit fruits of emerging alien shrub species more frequently than those of native shrub species in the South African Mediterranean climate region. South African Journal of Botany, 2013. 86: p. 73-78. [CrossRef]
- Mullett, T.L. Ecological aspects of Sweet Pittosporum (Pittosporum undulatum Vent.): Implications for control and management. in Eleventh Australian Weeds Conference 1996.
- Gleadow, R.M. and J. Walker, The invasion of Pittosporum undulatum in the Dandenong Ranges, Victoria: Realising predictions about rates and impact. Plant Protection Quarterly, 2014. 29: p. 103–109.
- Gleadow, R.M. and K.S. Rowan, Invasion by Pittosporum undulatum of the forests of central Victoria. III Effects of temperature and light on growth and drought resistance. Australian Journal of Botany, 1982. 30(3): p. 347-357. [CrossRef]
- Rose, S. and P.G. Fairweather, Changes in floristic composition of urban bushland invaded by Pittosporum undulatum in northern Sydney, Australia. Australian Journal of Botany, 1997. 45(1): p. 123-149. [CrossRef]
- Gleadow, R.M. and I. Narayan, Temperature thresholds for germination and survival of Pittosporum undulatum: implications for management by fire. Acta Oecologica, 2007. 31(2): p. 151-157. [CrossRef]
- Bellingham, P.J., et al., Endemic trees in a tropical biodiversity hotspot imperilled by an invasive tree. Biological Conservation, 2018. 217: p. 47-53. [CrossRef]
- Mullett, T.L., Effects of the native environmental weed Pittosporum undulatum Vent. (Sweet Pittosporum) on plant biodiversity. Plant Protection Quarterly, 2001. 16: p. 117–121.
- R Core Team, A language and environment for statistical computing. 2017, R Foundation for Statistical Computing: Vienna, Austria.
- Blossey, B. and R. Notzold, Evolution of increased competitive ability in invasive nonindigenous plants - a hypothesis. Journal of Ecology, 1995. 83(5): p. 887-889. [CrossRef]
- Keane, R.M. and M.J. Crawley, Exotic plant invasions and the enemy release hypothesis. Trends in Ecology & Evolution, 2002. 17(4): p. 164-170.
- Hejda, M., et al., Invasion success of alien plants: do habitat affinities in the native distribution range matter? Global Ecology and Biogeography, 2009. 18(3): p. 372-382. [CrossRef]
- van Etten, M.L., et al., Not all weeds are created equal: A database approach uncovers differences in the sexual system of native and introduced weeds. Ecology and Evolution, 2017. 7(8): p. 2636-2642.
- Hierro, J.L., J.L. Maron, and R.M. Callaway, A biogeographical approach to plant invasions: the importance of studying exotics in their introduced and native range. Journal of Ecology, 2005. 93(1): p. 5-15. [CrossRef]
- Hierro, J.L., et al., Disturbance facilitates invasion: The effects are stronger abroad than at home. American Naturalist, 2006. 168(2): p. 144-156. [CrossRef]
- Sax, D.F., et al., Ecological and evolutionary insights from species invasions. Trends in Ecology & Evolution, 2007. 22(9): p. 465-471. [CrossRef]
- Field, D.L., M. Pickup, and S.C.H. Barrett, Comparative analyses of sex-ratio variation in dioecious flowering plants. Evolution, 2013. 67(3): p. 661-672. [CrossRef]
- Nuñez, M.A., T.R. Horton, and D. Simberloff, Lack of belowground mutualisms hinders Pinaceae invasions. Ecology, 2009. 90(9): p. 2352-2359. [CrossRef]
- Eckert, C.G., et al., Plant mating systems in a changing world. Trends in Ecology & Evolution, 2010. 25(1): p. 35-43. [CrossRef]
- Crawley, M.J., P.H. Harvey, and A. Purvis, Comparative ecology of the native and alien floras of the British Isles. Philosophical Transactions of the Royal Society B-Biological Sciences, 1996. 351(1345): p. 1251-1259. [CrossRef]
- Daws, M.I., et al., Do invasive species have bigger seeds? Evidence from intra- and inter-specific comparisons. South African Journal of Botany, 2007. 73(1): p. 138-143. [CrossRef]
- Correia, M., et al., Evidence for enemy release and increased seed production and size for two invasive Australian acacias. Journal of Ecology, 2016. 104(5): p. 1391-1399. [CrossRef]
- Rejmfinek, M., A theory of seed plant invasiveness: The first sketch. Biological Conservation, 1996. 78(1-2): p. 171–181.
- Mason, R.A.B., et al., Reproductive output of invasive versus native plants. Global Ecology and Biogeography, 2008. 17(5): p. 633-640. [CrossRef]
- Muller-Landau, H.C., The tolerance-fecundity trade-off and the maintenance of diversity in seed size. Proceedings of the National Academy of Sciences of the United States of America, 2010. 107(9): p. 4242-4247.
- van Kleunen, M., W. Dawson, and N. Maurel, Characteristics of successful alien plants. Molecular Ecology, 2015. 24(9): p. 1954-1968. [CrossRef]
- Sinclair, J.P., J. Emlen, and D.C. Freeman, Biased sex ratios in plants: Theory and trends. Botanical Review, 2012. 78(1): p. 63-86. [CrossRef]
- Harris, M.S. and J.R. Pannell, Roots, shoots and reproduction: sexual dimorphism in size and costs of reproductive allocation in an annual herb. Proceedings of the Royal Society B-Biological Sciences, 2008. 275(1651): p. 2595-2602. [CrossRef]
- Hubbell, S.P., Seed predation and the coexistence of tree species in tropical forests. OIKOS, 1980. 35(2): p. 214–229. [CrossRef]
Figure 1.
A) Map of Australia showing current distribution of Pittosporum undulatum (Atlas of Living Australia http://www.ala.org.au). The dotted line indicates the native distribution (Gleadow and Ashton 1981). State borders are defined by the solid lines; (B) Location of the 14 sampling sites across Victoria that span native and expanded ranges: Morwell, Lakes Entrance, Lake Tyres, Marlo and Mallacoota are all considered native populations. Remaining locations represent invasive populations.
Figure 1.
A) Map of Australia showing current distribution of Pittosporum undulatum (Atlas of Living Australia http://www.ala.org.au). The dotted line indicates the native distribution (Gleadow and Ashton 1981). State borders are defined by the solid lines; (B) Location of the 14 sampling sites across Victoria that span native and expanded ranges: Morwell, Lakes Entrance, Lake Tyres, Marlo and Mallacoota are all considered native populations. Remaining locations represent invasive populations.
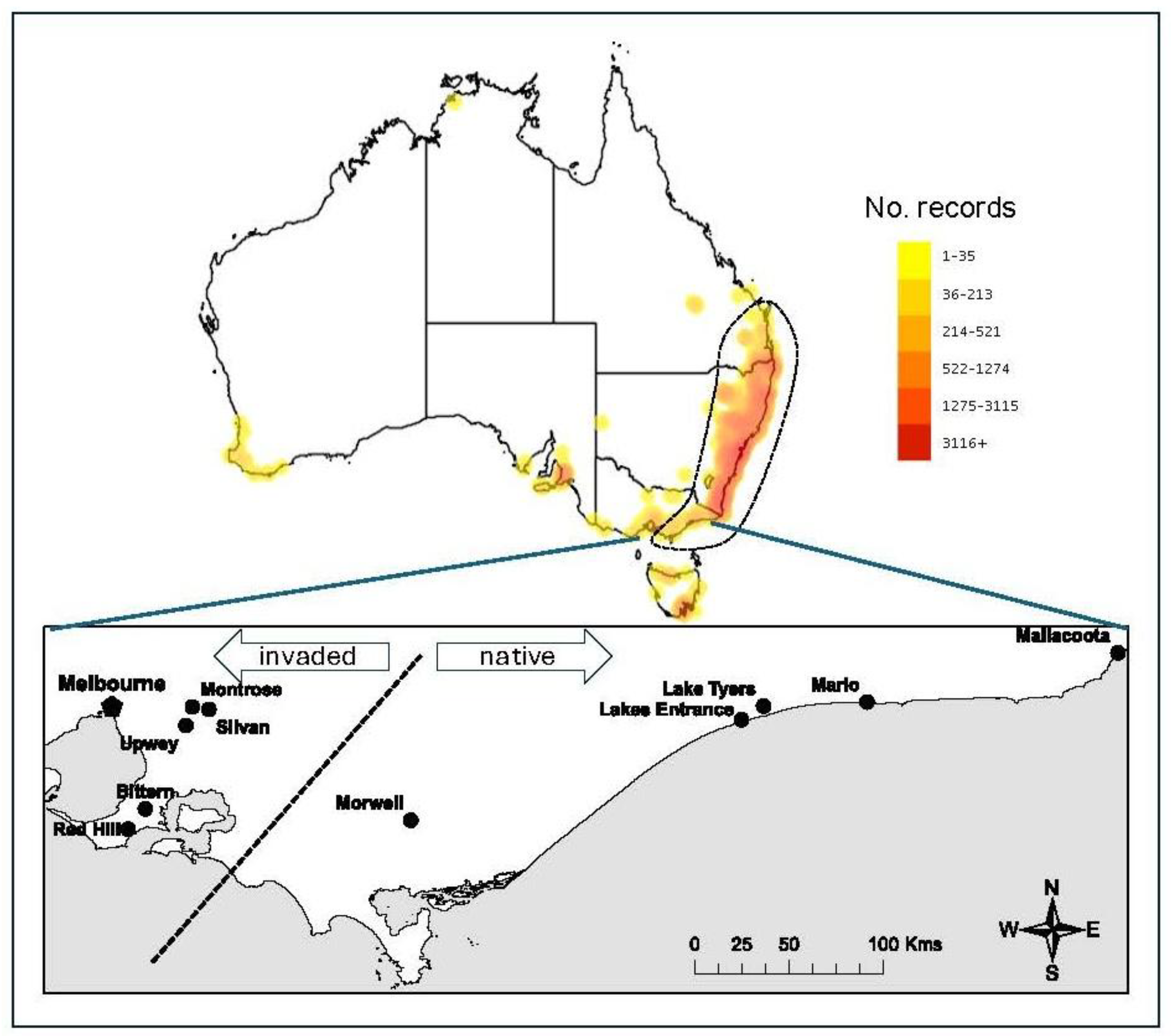
Figure 2.
A) Proportion of female and hermaphrodite individuals fruiting in native and invasive populations . (B) Variation in the mean fruit rank (an index of numbers of fruit) for individual females and hermaphrodites within native and invasive populations. Closed circles represent trees from native populations and open circles those from invaded populations. For full statistical analysis see Supplementary Tables S2 and S3.
Figure 2.
A) Proportion of female and hermaphrodite individuals fruiting in native and invasive populations . (B) Variation in the mean fruit rank (an index of numbers of fruit) for individual females and hermaphrodites within native and invasive populations. Closed circles represent trees from native populations and open circles those from invaded populations. For full statistical analysis see Supplementary Tables S2 and S3.
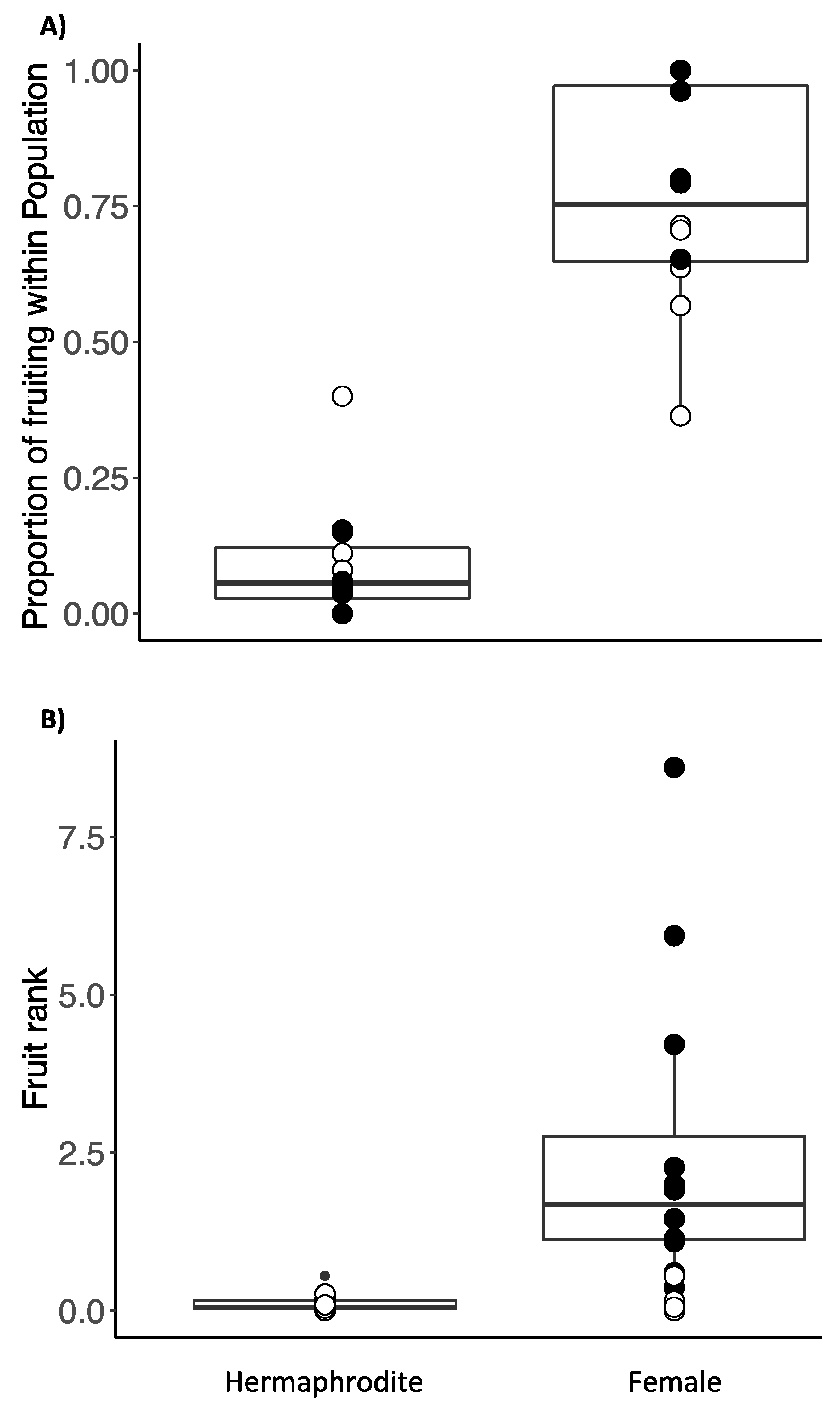
Figure 3.
Mean site fruit rank relative to leaf chemical traits. (A) dC13 ‰ (B), Total Nitrogen (% dw); (C) Total Carbon (% dw). Females are represented by closed circles and hermaphrodites by open circles. No significant correlations were detected overall between traits and fruit rank.
Figure 3.
Mean site fruit rank relative to leaf chemical traits. (A) dC13 ‰ (B), Total Nitrogen (% dw); (C) Total Carbon (% dw). Females are represented by closed circles and hermaphrodites by open circles. No significant correlations were detected overall between traits and fruit rank.
Figure 4.
Leaf nitrogen concentration for P. undulatum trees collected from 14 different populations. Bars represent the mean of 5 replicates ± 1SE. Grey bars = female, white bars = Hermaphrodite. Statistical analysis is in Supplementary Table S5.
Figure 4.
Leaf nitrogen concentration for P. undulatum trees collected from 14 different populations. Bars represent the mean of 5 replicates ± 1SE. Grey bars = female, white bars = Hermaphrodite. Statistical analysis is in Supplementary Table S5.
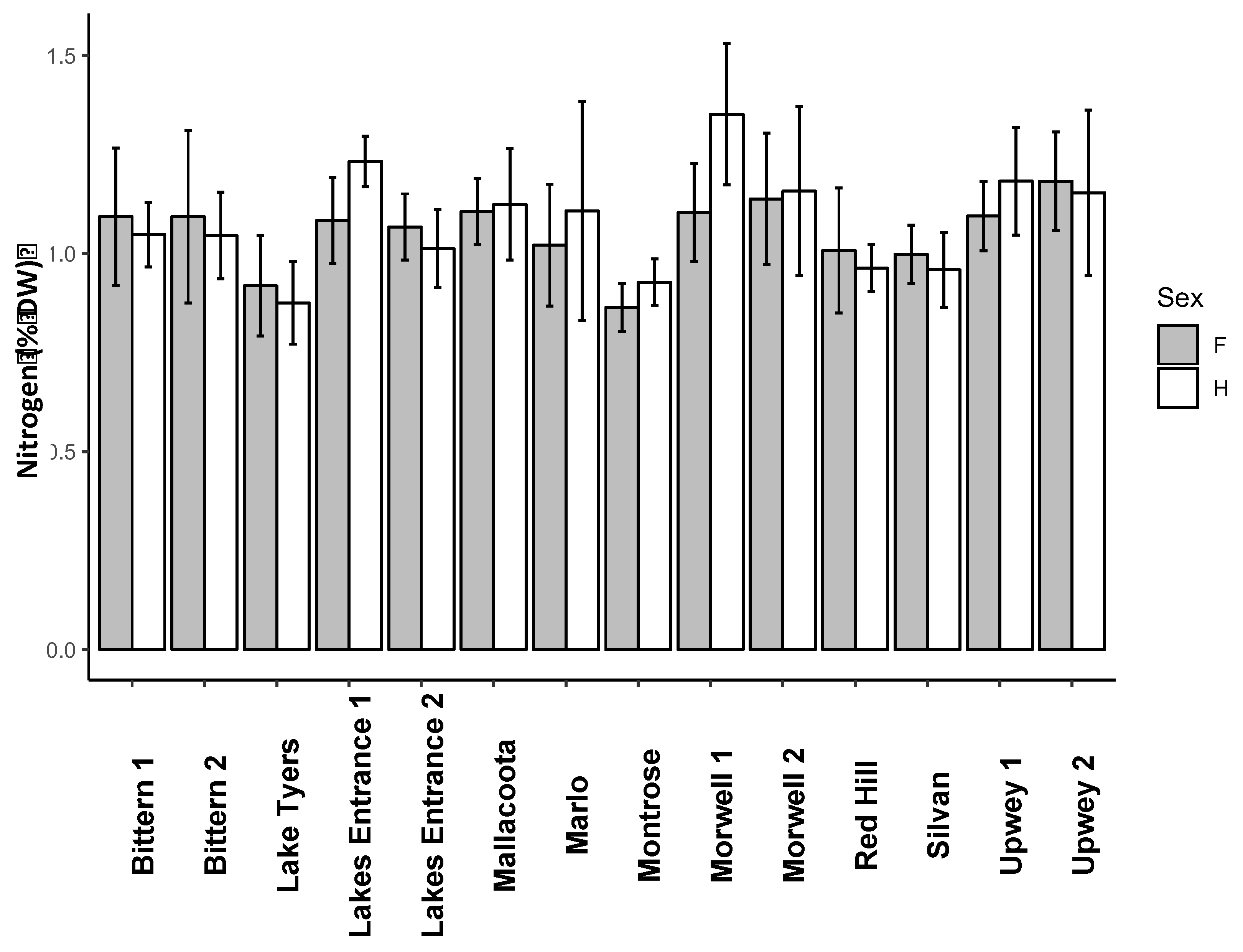

Table 1.
Location and elevation of 14 sites sampled across Victoria, Australia. Seven populations are considered to be within the ‘native’ range and seven where P. undulatum was only more recently recorded (see Figure 1).
Table 1.
Location and elevation of 14 sites sampled across Victoria, Australia. Seven populations are considered to be within the ‘native’ range and seven where P. undulatum was only more recently recorded (see Figure 1).
| Site | Origin | Location | Elevation (m) |
| Morwell National Park 1 | Native | Lat:-38.36 Lon:146.40 |
184 |
| Morwell National Park 2 | Native | Lat:-38.36 Lon:146.40 |
184 |
| Lakes Entrance 1 | Native | Lat: -37.88 Lon: 147.96 |
40 |
| Lakes Entrance 2 | Native | Lat: -37.88 Lon: 147.96 |
40 |
| Lake Tyers State park | Native | Lat:-37.76 Lon:148.07 |
89 |
| Marlo | Native | Lat -37.79 Lon: 148.55 |
22 |
| Mallacoota | Native | Lat: -37.56 Lon:149.76 |
19 |
| Red Hill | Invaded | Lat:-38.39 Lon:145.02 |
131 |
| Bittern 1 | Invaded | Lat: -38.30 Lon: 145.12 |
81 |
| Bittern 2 | Invaded | Lat: -38.30 Lon: 145.12 |
81 |
| Upwey 1 | Invaded | Lat: -37.90 Lon:145.31 |
291 |
| Upwey 2 | Invaded | Lat: -37.90 Lon:145.31 |
291 |
| Montrose | Invaded | Lat:-37.84 Lon:145.33 |
222 |
| Silvan | Invaded | Lat:-37.83 Lon:145.42 |
293 |
Table 2.
Number of trees per 20 m x 20 m plot (tree density), number of females per plot, mean number of seeds per fruit capsule (± 1 SD) and mean seed mass (g dw) (± 1SD) across all sampled populations. Fruit availability limited sampling for the sites at Silvan and Bittern. Differences between native and invaded sites are not significantly different (P <0.05, Supplementary Table S1).
Table 2.
Number of trees per 20 m x 20 m plot (tree density), number of females per plot, mean number of seeds per fruit capsule (± 1 SD) and mean seed mass (g dw) (± 1SD) across all sampled populations. Fruit availability limited sampling for the sites at Silvan and Bittern. Differences between native and invaded sites are not significantly different (P <0.05, Supplementary Table S1).
| Trees | Females | Seed number | Seed weight (g) | |
| Native populations | ||||
| Morwell 1 | 30 | 10 | 32.0 ± 1.8 | 0.0023 ±0.0003 |
| Morwell 2 | 8 | 5 | 28.0 ± 3.6 | 0.002 ±0.0001 |
| Lakes Entrance 1 | 103 | 46 | 33.8 ± 1.3 | 0.0036 ± 0.0003 |
| Lakes Entrance 2 | 34 | 16 | 27.1 ± 3.6 | 0.0038 ± 0.0001 |
| Lake Tyers | 22 | 8 | 25.0 ± 1.4 | 0.0029 ± 0.0026 |
| Marlo | 62 | 34 | 27.3 ± 4.9 | 0.0040 ± 0.0014 |
| Mallacoota |
143 | 71 | 28.7 ± 5.7 | 0.0027 ± 0.0006 |
| Invasive populations (rural) | ||||
| Red Hill | 159 | 72 | 27.6 ± 4.0 | 0.0082 ± 0.0026 |
| Bittern 1 | 51 | 15 | N/A | |
| Bittern 2 | 89 | 44 | N/A | |
| Upwey 1 | 30 | 14 | 26.8 ± 4.2 | 0.0071 ± 0.0017 |
| Upwey 2 | 29 | 12 | 28.7 ± 5.4 | 0.0066 ± 0.0016 |
| Montrose | 84 | 31 | 27.9 ± 4.7 | 0.0028 ± 0.0009 |
| Silvan | 27 | 10 | N/A | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



