
Submitted:
10 February 2025
Posted:
11 February 2025
You are already at the latest version
Abstract
The globose scale (GS) Sphaerolecanium prunastri (Boyer de Fonscolombe) (Hemiptera: Coccidae) is a serious pest affecting plants within the Rosaceae, notably wild apricot, Armeniaca vulgaris (Lamarck). Following its initial detection in 2019, more than 80% of valleys with wild apricots have become affected in the Ili River basin of Tianshan Mountains in Xinjiang, China. This study assessed GS population dynamics post invasion and its effects on growth and reproductive traits of wild apricot trees from 2019 to 2024. Nymph densities decreased over the years but remained high, with densities on shoots per 20 cm of 986 (1st instar nymph) and 120 (2nd instar nymph) in 2024, respectively. Damage has declined, with high damage rankings decreasing from 24% to 11% of wild apricot trees. However, mortality of trees was higher (25%) in infested than non-infested areas (13%). Interestingly, GS feeding stimulated the growth of spring shoots but significantly reduced the reproductive capacity of wild apricot. Heavily infested trees exhibited increased shoot length (2-3 times), decreased fruit yield (20-fold), lower flowering percentage (8-fold), and reduced flower bud density (2-fold) compared to non-infested trees. Overall, despite a decrease in damage severity, wild apricot forests remain threatened by GS. Implementing Integrated Pest Management (IPM) strategies is essential for effective GS management and recovery of wild apricot forests.
Keywords:
fruit set
; pest density
; plant compensation
; plant-insect interactions
; wild apricot forest
1. Introduction
Scale insects are common and destructive pests that frequently infest horticultural, fruit and ornamental plants [1]. Their small size makes them difficult to detect and identify, facilitating their rapid spread to new areas [2,3]. Some scale species can cause significant yield losses and economic damage, and even profound ecological impacts in natural systems. For example, the invasion of tortoise pine scale (Toumeyella parvicornis) in Turks and Caicos Islands has resulted in the infestation of more than 90% of the pine forested area, and severe damage to several Pinus species, seriously disturbing the balance of the forest ecosystem [4]. This species has also invaded Europe [5], causing higher mortality of stone pines (Pinus pinea L.) in a Coastal Pinewood in Southern Italy [6].
The globose scale (GS) Sphaerolecanium prunastri (Boyer de Fonscolombe) (Hemiptera: Coccoidea), a soft scale insect, is an invasive species [7]. Native to central and southern Europe it is now widespread across the Mediterranean, North America, the Middle East, Central and East Asia [8]. In Turkey, GS is considered a key pest of Prunus spp. [9], where it severely damages plum, peach and apricot trees [10,11,12], reaching 100% infestation in some orchards [9,13]. In 2019, an outbreak of GS was detected in wild apricot (Armeniaca vulgaris) forests in Xinjiang, China [7], particularly in the Ili Valley of Tianshan Mountains which has 1440 ha of patchily distributed remnant wild apricot forest [14]. The GS infestation has led to a decline in the physiological health of wild apricot populations, causing dieback, stunted growth and increased mortality of trees. This plant species, the ancestor of the cultivated apricot, is a tertiary relict plant and an important genetic resource for further breeding [15]. It is estimated that more than 80% of wild apricot trees in this valley have been infested with the pest [16], posing a serious threat to these trees and the resilience of the wild apricot forest ecosystem.
GS has one generation a year and overwinters as 2nd-instar nymphs on wild apricot trees [7]. The 2nd-instar nymph goes through the longest life history stage. At the beginning of July 2nd-instar nymph feed on phloem of young branches and then overwinter from October to late March of the following year. After overwintering, it pupates from late March to early April [7]. The overwintering 2nd-instar nymph stage coincides with bud development and flowering of wild apricot. The long-term damage stress seriously affects the flowering and germination of wild apricots [17]. Adults appear in late May and early June, when they increase in feeding secreting a large amount of honeydew [7]. Eggs hatch in early to mid-June and after a short period of crawling, 1st-instar nymph settle on young branches (less than 3 years old) and feed on the phloem, developing into 2nd-instar nymph in July [7]. Infested wild apricot are seriously damaged by female adults and 1st-instar nymph in early summer which coincides with the most vigorous growth and fruit-setting period [18].
The sap-sucking behavior of scale insects causes a cascade of direct effects, including branch dieback and secondary damage such as sooty mold [19] which reduces photosynthetic activity and stunts the growth of infested branches [15]. To mitigate the impact of GS on wild apricot, various management strategies have been implemented. The introduction of ladybird beetles (Harmonia axyridis) as a biological control agent had limited success, with field efficacy below 35% [20]. Although a variety of natural parasitic enemies have been identified within wild apricot forests [21,22], their impact on GS populations is low [23]. Since 2022, a large-scale aerial spraying program has been initiated to suppress GS populations. Nevertheless, its control efficacy has been challenging due to the mountainous topography, adverse weather, and GS biological traits such as a heavy protective waxy covering. As a result, GS populations continue to persist at damaging levels and pose a major risk to germplasm of wild apricots.
A rational assessment of pest damage can provide the foundation for decisions on management, especially in targeting areas to spray, selecting appropriate methods and determining the optimal timing of controls [24]. There were some basic assessments for wild apricots damage including GS densities and damage caused during the initial GS outbreaks [16], but there remains a gap between the invasion, rapid spread and the practical requirements for management GS, which has a more extensive and complex impact on wild apricots over time. This includes changes in the damage caused by GS and the cumulative effects on the growth and reproductive capacity of wild apricot. Such assessments are needed to monitor both pest population dynamics and tree heath simultaneously over a long period.
In this study we clarify the status of the GS population and the extent of the damage it has caused since the initial outbreak. In this study we: (1) survey population changes of GS by estimating pest density over 6 years following its first detection; (2) assess the change of GS damage to wild apricot trees over this period; and (3) evaluate the cumulative effects of GS damage on tree growth and reproduction, including spring shoot growth, fruit yield, and flower bud density.
2. Materials and Methods
2.1. Experimental Design
2.1.1. Survey Site
This study was carried out from 2019 to 2024 (excluding 2023) in wild apricot forests located in Gongliu, Xinyuan and Huocheng Counties, China at latitude 43°14’02”N to 44°42’88”N and longitude 83°03’87”E to 83°48’54”E; 1026.33-1376.6 m above sea level.
Four areas in Gongliu County and two areas in Xinyuan County, comprising 90% of the distribution of remnant forest where wild apricots are the dominant species, were selected as experimental sites. Another area, Huocheng County, was the control site as wild apricot was not yet infested by GS. At each survey location, we established replicate circular sample plots with a radius of 20 meters for a total 12, 4, and 3 plots in Gongliu, Xinyuan, and Huocheng Counties, respectively. Replicate plots in each site were spaced 1-3 kilometers apart (Table 1)
2.1.2. Density Survey of Sphaerolecanium prunastri
GS density surveys were carried out from 2019 to 2024, except for 2023. Surveys of nymph density began in early April, after the overwintering phase of the 2nd instar nymph. The density of 1st instar nymphs was estimated in early to mid-June. In each area, we randomly sampled 15 trees to assess nymph density. A 3-years old branch (1.5-3 cm in diameter) was cut from the east, south, west, and north directions of each tree at a height of 4-6 m. Branchlets measuring 20 cm in length were used to count 1st and 2nd instar nymphs because GS were concentrated on these. All densities reported in the results are per 20cm.
2.1.3. Damage Ranking of Wild Apricot
To assess the damage due to GS infestation, we surveyed the damage ranking of trees and the number of dead individuals. Tree damage was categorized as: 0=non-infested, 1=least damaged (1-10%), 2=low damage (11-31%), 3=medium (31-50%), 4=high (51-70%) and 5= very high (71-100%) damage based on the percentage of branches damaged by GS in each tree. This monitoring was conducted in early to mid-June between 2021 and 2024. We assessed the mortality of wild apricot in each plot to evaluate the cumulative effect of GS on tree health in 2021 and 2024. The dead individuals were identified based on the presence of old GS damage and the absence of new branches and green leaves.
2.1.4. The growth of Wild Apricots
The growth of spring shoots can indirectly reflect the potential growth and vigor of trees within a year. We observed the length of spring shoots (fresh shoots of the current year) on branches across trees with different damage rankings from 2021 to 2023. In early to mid-June, when wild apricot tree growth is at the most vigorous stage, 15 trees were randomly chosen from non-infested, low and high damage ranking and labelled. The non-infest trees were chosen from all plots in Huocheng County, which were free from pest scale as controls. There were no trees in this category in infested counties. The low and high damage trees were selected from the plots in Gongliu and Xinyuan Counties. Following the method used for surveying the densities of 1st- instar nymph, we cut four 3-year-old branches (less than 3 year growth), and measured the length of five spring shoots on each branch using a tape measure. A total of 20 spring shoots were measured per tree.
2.1.5. Assessment of Flower Buds and Fruit Yield
In 2020, we assessed the density of flower buds at three survey sites in Gongliu County. Then, from 2022 to 2024, we systematically assessed fruit yield within all plots and assessed flowering in 2023.
Assessment of flower buds was carried out in plots located in Xiaomohuer, Jinqikesai and Kuolesai Valleys (see Table 1). Five wild apricot trees were randomly selected from low, medium and high damage rankings in each plot. In late August and early September, one 3-years old branch was cut in the east, south, west, and north directions of each tree. The length of the branch was measured and the number of flower buds counted [25,26]. Finally, the density of flower buds per meter was calculated.
Assessment of fruit yield and flowering: In late June to early July, during the ripening of wild apricot fruit, we estimated the fruit yield for all non-infested trees and those with low and high levels of damage by counting the number of fruits, as weighing all fruit was impractical. Non-infested trees were sampled from three plots in Huocheng County, while low and high damaged trees were selected from all plots in Gongliu and Xinyuan Counties. To establish a weight estimation criterion, we weighed 50 fruits (approximately 500g) in each plot, which allowed us to estimate the yield for each plot. Furthermore, in early to mid-April 2023, when wild apricots were in full bloom, we assessed the flowering rate of low, medium, and high damage trees across all plots in Gongliu County. The flowering rate was calculated as the percentage of blooms relative to the entire canopy.
2.2. Data Analysis
One-way analysis of variance (ANOVA) was used to analysis the differences in GS density, flower bud density, and percentage flowering across damage rankings (SPSS Institute Inc 20). Years and damage ranking were fixed factors, and other observed variables were dependent variables. Data normality and homogeneity of variance were checked by Shapiro-Wilk tests (Shapiro & Wilk 1965). Two-way analysis of variance (ANOVA) was used to analyze the differences in spring shoot length and fruit yield across years and damage rankings. Years and damage ranking were fixed factors, survey site was a random factor and other observed variables were dependent variables. The means were separated using Tukey’s post-hoc test. Linear correlation (Origin 2021) was used to analyze the correlation between average damage ranking and fruit yields.
3. Results
3.1. Population Dynamics of GS Nymph Over Years
There was a significant difference among years for the density of 1st and 2nd instar nymphs (1st: F(4,34)=13.63, p<0.001; 2nd: F(3,32)=9.89, p<0.001) (Figure 1). Both 1st and 2nd instar nymph densities exhibited a similar trend, rapidly increasing in 2020, peaking in 2021 (1st: 1944 ± 325; 2nd: 454 ± 127). First-instar nymph density was low in 2019 at the start of the outbreak (38 ± 7) compared to other years. No significant differences were observed among 2020, 2021, 2022, and 2024. Second-instar nymph density was significantly lower in 2020 (137 ± 21) and 2024 (120 ± 19) compared to other years. Population densities of 2nd instar nymph declined to levels similar to 2020. However, 1st instar nymph densities remained over 1000 scale pest per 20cm.
3.2. Damage Ranking of Wild Apricot
There were significant differences in damage among years (Figure 2a). Between 2021 and 2023, while high and medium damage ranking increased to 29.6% and 27.6%, respectively, high damage decreased to 11.2% in 2024. Correspondingly, low damage ranking increased from 56.2% to 67.3%, which indicated that the damage of wild apricot tress has been gradually declining. However, mortality of wild apricot in damaged areas was higher than non-infested areas. In 2021 and 2024, the percentage of dead trees in damage areas reached 27.6% and 26.7% while 13.0% and 12.4% mortality of trees occurred in non-infested area n these years respectively (Figure 2b).
3.3. Length of Spring Shoots Among Damage Rankings
GS infestation had a positive stimulating effect on the growth of spring shoots, but the effect was attenuated with years. The length of spring shoots was significantly related to year and damage ranking (F(4,861)=8.48, p<0.001) (Figure 3). There was a positive correlation between the extent of damage and stem length. Non-infested trees exhibited stem lengths of 6.1 to 7.9 cm, while those with low and high damage had stem lengths of 7.9 to 18.3 cm and 15.1 to 20.7 cm, respectively. The length of spring shoots was 2-3 times longer compared with undamaged one.
The shoot length response of infested trees gradually attenuated with years. The shoot length of trees with low and high damage rankings were significantly lower in 2022 and 2023 than 2021. The length with low damage in descending order with year were 2021 (18.3 ± 0.4cm)>2022 (13.3±10.8cm)>2023 (7.9±0.4cm). High damaged trees had stem lengths of 15.5 cm in 2023 and 15.1cm in 2022, significantly shorter than the length of 20.7cm in 2021 (Figure 3). However, the shoot growth of non-infested trees did not change significantly among three years indicating GS has a cumulative damage effect on shoot growth. The length of the shoots gradually decreases with years (Figure 3).
3.4. Flower Buds and Flowering
GS negatively affected reproductive capacity affecting both flower buds and flower abundance per tree. The density of flower buds and flowering percentage per tree were different among three damage rankings (flower bud: F(2,478)=31.83, p<0.001; flowering percentage per tree: F(2,429)=331.59, p<0.001). With the increasing damage, flower bud density and flowering percentage per tree decreased. The flower bud density of low damage trees (88.5 ± 4.7 per tree) was about 2 times higher than high damage trees (46.2 ± 3.1). Flowering percentage of low damage (59.8 ± 2.0 per tree) was more 8 times higher than high damage trees (7.5 ± 0.9 per tree) (Figure 4).
3.5. Fruit Yields Loss
The fruit yield differed among three damage rankings and years (F(4,1241)=30.58, p<0.001) (Figure 5). Average fruit yield decreased with increasing damage ranking and years except in 2024 which experienced extreme freezing conditions in spring. In 2022 and 2023, average fruit yield among three damage rankings in descending order: non-infest (in 2022: 8.4±0.9 kg; in 2023: 2.2±0.4 kg) > low damage (2022: 2.0±0.58kg; 2023: 0.9±0.2 kg)> high damage (2022: 0.3±0.1kg; 2023: 0.1±0.1 kg). In 2024, fruit yield of non-infested trees was less than 0.1 kg per tree and were not significantly different from uninfected trees. Fruit yield was reduced by more than 20 times in damaged trees compared to non-infested trees without considering the effect of freezing conditions on fruit yield in 2024. There was a significant negative relationship between average fruit yield and damage ranking scores (Figure 6), which indicated GS has a decreasing effect on the fruiting of wild apricots.
4. Discussion
The invasion and subsequent outbreak of S. prunastri has had a profound detrimental impact on wild apricot forests, disrupting both individual tree growth and potentially the ecology of this unique system. Our observations revealed the changes in the population of first and second instar nymphs over the course of the invasion. A rapid increase in nymph density was observed from 2019 to 2021, culminating in a peak, followed by a gradual decline from 2022 to 2024. Concurrently, a gradual decrease in damage levels within the wild apricot forests was observed (Figure 2). Our findings indicate a counterintuitive response in severely damaged wild apricot trees. While experiencing increased spring shoot lengths, these trees exhibited significantly lower fruit yields, flowering percentages, and flower bud densities. This suggests that cumulative damage inflicted by S. prunastri stimulates compensatory branch growth while simultaneously suppressing reproductive capacity. It is crucial to note that the stimulatory effect of S. prunastri on branch growth appeared to diminish over time, as evidenced by the gradual decrease in spring shoot length across the years (Figure 3), whereas un-infested trees showed no response over time.
4.1. GS Population Development and Damage on the Wild Apricot Forest
GS females can establish a large population size in a short time because of high fecundity with 400-800 eggs per female [9,23]. The population of GS in our study areas developed rapidly in the two years following the initial detection of invasive populations in 2019. The densities of 1st instar nymph increased dramatically to more than 1,000 per 20 cm branch in 2020 and 2021, which far exceeded the densities of less than 100 at the time of the initial population (Figure 1). Serious damage occurred to infested trees in this period (Figure 2a). In wild apricot forests, the density of GS varies amongst regions. Wang et al.[16] showed that 1st instar nymph density decreased from over 1,000 to less than 500 in some areas from 2019 to 2020, while its densities increased 100 times in others. From 2022 to 2024, the density and damage of GS began to decrease gradually. The decline is likely the result of large-scale aerial spraying and pruning management in 2022 and 2023. However, 1st-instar nymph density in 2024 was still 26 times higher than in 2019. The cumulative damage from high GS density seems to accelerate the death of wild apricot. Compared to less than 13% mortality in un-infested areas, tree mortality nearly doubled to 25% in GS-infest areas. GS direct physiological effects likely played a decisive role in the death of wild apricot. Severe ductal embolism occurred in xylem of wild apricot showing decline, which induced hydraulic dysfunction and reduced the synthesis and storage of non-structural carbohydrate [27]. Severe carbohydrate-water imbalance does not seem to defend against high-density GS. We expect the high density of GS may continue to accelerate the death of wild apricots.
4.2. The Effect of GS on the Growth of Wild Apricots
Infested wild apricots had longer spring shoot lengths compared to non-infested trees (Figure 3). Trees with higher damage ranking had spring shoots, that were approximately 2-3 times longer than the non-infested trees. This a compensatory growth response to GS damage on wild apricot. Compensatory growth, such as increase in leaf area, new shoots and root-crown ratio, is a positive response of plants to insect damage by altering resource allocation, physiology, and/or phenology [28,29,30]. Trees can be affected by increased sink demand from sap-sucking pests, inducing nutrients to move toward the damaged site [31]. The relocation of nutrients driven by GS feeding accumulated in fresh branches and led increased shoot growth of infested branches (Figure 3). However, the health of trees gradually declined with years because of the accumulated damage and less source of carbohydrate or “carbon starvation” [27]. This means that there are insufficient nutrients allocating to the growth of branch and the growth of the spring shoot gradually decreased with years.
4.3. The Effect of GS on the Reproductive Capacity of Wild Apricots
Sap-feeding insects can significantly impair the reproductive capacity of woody plants [31,32]. This occurs primarily due to competition for vital nutrients between the sucking pests and the plant's reproductive organs, such as buds, flowers and fruits [31,33,34]. Previous research has demonstrated a significant reduction in fruit size of wild apricots infested with GS[15]. Our findings confirm these observations, revealing a significant decline in flower bud density, flowering percentage, and fruit yield in wild apricots with increasing levels of GS infestation. Furthermore, the detrimental impact of GS on wild apricot fruit production is exacerbated by the development of sooty mold, a secondary consequence of the infestation. Sooty mold has been shown to reduce stomatal conductance and photosynthetic capacity, ultimately leading to a decrease in both the quantity and quality of fruit yield[9]. It is crucial to note that the flowering and fruit set of wild apricots are inherently susceptible to spring frosts, further compounding the challenges faced by these plants. In 2022, wild apricot yields reached 8 kg per plant in un-infested areas but was less than 1 kg per tree due to freezing conditions in 2023 and 2024 (Figure 5). The reduced yields due to GS and freezing will decrease the seed bank in soil, reducing recruitment of seedlings and ultimately forest regeneration and forest resilience [35].
4.4. Integrated Management Approach to GS
The implementation of integrated pest management (IPM) strategies can effectively reduce GS populations and help recovery wild apricot forest health[15] Large-scale aerial spraying programs have been successfully employed in wild apricot forests, resulting in significant reductions in GS densities across most areas [16]. Additionally, the pruning of infested branches has proven to be an effective approach for controlling GS populations and mitigating damage levels in wild apricot trees[36], as was the case with Agrilus mali in wild apple[37]. In the long term, biological control strategies should be prioritized as a sustainable and practical approach. This is supported by successful examples in Turkey, where parasitism rates from eight hymenopterous species for GS exceeded 70% in cultivated orchards [13]. Furthermore, a comprehensive suite of nursery planting options to replace dead trees should be integrated with other management tactics to enhance the resilience of wild apricot forests and boost their ecological defenses against GS. For example, the postponement of spring grazing and reduction of grazing frequency can contribute to the survival of wild apricot seedlings and increase the stability of wild apricot tree populations.
5. Conclusions
Our study demonstrated that the population dynamics of GS, along with the damage ranking of wild apricot, have been gradually decreasing over the years. However, wild apricot still suffers damage because of the continuing high densities of GS. More than 25% of wild apricots died in the GS-infested areas. Furthermore, the cumulative damage inflicted by GS on wild apricot significantly stimulates the growth of new shoots, but adversely affects the reproduction capacity including fruit yield, flowering percentage, and flower bud density. This cumulative damage effect becomes particularly pronounced as the damage ranking increases, raising concerns about the destiny of wild apricot. Moving forward, it is essential to strengthen the monitoring and integrated pest management (IPM) strategies against GS to reduce its density below the damage threshold level firstly. Additionally, these efforts should be integrated with other conservation and restoration strategies to enhance the resilience of wild apricot against GS, by enhancing recruitment of young trees in various landscapes in Tianshan Mountains.
Author Contributions
Z.G. G. and Z.Z.L. conceived and designed the experiments; P Z., Y.F. L. and C.H. L. performed the experiments; P.Z. analyzed the data; P.Z. and Z.Z.L. wrote the manuscript main text. Z.K.D., B.R., M.P.Z. and Z.Z.L. revised the manuscript. All authors approved the manuscript for submission.
Funding
This research was funded by the Tianshan Talents Program of Xinjiang Uygur Autonomous Region, grant number 2023TSYCCX0029 and Central finance forestry and grass science and technology promotion demonstration project, grant number Xin [2024]TG02.
Data Availability Statement
The original contributions presented in the study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
The author is grateful to Yuxuan Li and Sen Wang of Shihezi University, Ming Li of Xinjiang Agricultural University, and Zhao Tang of Tarim University for their help in field work, and Mr. Zhijun Cui and Huailiang Ma of Gongliu County Forestry and Grassland Bureau for their technical help.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Miller, D.; Miller, G.; Hodges, G.S.; Davidson, J.A. Introduced scale insects (Hemiptera: Coccoidea) of the United States and their impact on U.S. agriculture. Proc. Entomol. Soc. Washington, 2004, 107, pp.123–158. [Google Scholar]
- Morales, M.; Denno, B.; Miller, D.; Miller, G.; Ben-Dov, Y.; Hardy, N. ScaleNet: a literature-based model of scale insect biology and systematics. Database (Oxford). 2016, bav118, 1–5. [Google Scholar]
- Amouroux, P.; Crochard, D.; Germain, J.F.; Correa, M.; Ampuero, J.; Groussier, G.; Kreiter, P.; Malausa, T.; Zaviezo, T. Genetic diversity of armored scales (Hemiptera: Diaspididae) and soft scales (Hemiptera: Coccidae) in Chile. Sci Rep. 2017, 7, 2014. [Google Scholar] [CrossRef]
- Malumphy, C.; Hamilton, M.; Manco, B.N.; Green, P.; Sanchez, M.; Corcoran, M.; Salamanca, E. Toumeyella parvicornis (Hemiptera: Coccidae), causing severe decline of Pinus caribaea var. Bahamensis in the Turks and Caicos Islands. Fla. Entomol 2012, 95, 113–119. [Google Scholar] [CrossRef]
- Di Sora, N.; Contarini, M.; Rossini, L.; Turco, S.; Brugneti, F.; Metaliaj, R.; Vejsiu, I.; Peri, L.; Speranza, S. First report of Toumeyella parvicornis (Cockerell) (Hemiptera: Coccidae) in Albania and its potential spread in the coastal area of the Balkans. Bull. OEPP 2024, 54, 160–165. [Google Scholar] [CrossRef]
- Nicoletti, R.; De Masi, L.; Migliozzi, A.; Calandrelli, M.M. Analysis of dieback in a coastal pinewood in Campania, Southern Italy, through High-Resolution Remote Sensing. Plants 2024, 13, 182. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, Z.; Linghu, W.; Gao, G. Sphaerolecanium prunastri (Hemiptera: Coccoidea: Coccidae), a new pest in wild fruit forests, Xinjiang. Forest Research 2021, 34, 152–158. [Google Scholar]
- Çiftçi, Ü.; Bolu, H. First records of Coccomorpha (Hemiptera) species in Diyarbakır, Turkey. J Entomol Sci 2021, 56, 235–245. [Google Scholar] [CrossRef]
- Karaca, I.; Japoshvili, G.; Demirozer, O. The chalcid parasitoid complex (Hymenoptera: Chalcidoidea) associated with the globose scale (Sphaerolecanium prunastri Fonscolombe) (Hemiptera: Coccoidea) in Isparta Province, Turkey and some east European countries. J Plant Dis Prot 2003, 110, 505–511. [Google Scholar] [CrossRef]
- Gülmez, M.; Ülgentürk, S.; Ulusoy, M. Soft scale (Hemiptera: Coccomorpha: Coccidae) species on fruit orchards of Diyarbakır and Elazığ provinces in TürkiyeDiyarbakır ve Elazığ illeri meyve bahçelerindeki Koşnil (Hemiptera:Coccomorpha: Coccidae) türleri. Turk. j. entomol 2023, 47, 199–213. [Google Scholar] [CrossRef]
- Özgen, i.; Bolu, H. Determination of Sphaerolecanium prunastri (Boyer de Fonscolombe, 1834) (Hemiptera: Coccidae) plum scale, the distribution, infestations and natural enemies in Malatya province in Turkey. Turk. j. entomol 2009, 33, 83–91. [Google Scholar]
- Bulak, Y.; Yildirim, E. Evaluation of hosts and distribution of Hyalopterus pruni (Geoffroy, 1762) and Sphaerolecanium prunastri (Boyer de Fonscolombe, 1834), which are pests of fruit trees in Iğdır. International conference on food, agriculture and animal sciences, Sivas,Turkey, 27-29 April 2023.
- Japoshvili, G.; Ay, R.; Karaca, I.; Gabroshvili, N.; Barjadze, S.; Chaladze, G. Studies on the parasitoid complex attacking the globose scale Sphaerolecanium prunastri (Fonscolombe) (Hemiptera: Coccoidea) on prunus species in Turkey. J. Kans. Entomol. Soc 2009, 81, 339–344. [Google Scholar] [CrossRef]
- Li, L.; Hai, Y.; Mohammat, A.; Tang, Z.; Fang, J. Community structure and conservation of wild fruit forests in the IIi Valley, Xinjiang. Arid Zone Res 2011, 28, 60–66. [Google Scholar] [CrossRef]
- Linghu, W.; Lu, Z.; Wang, Y.; Gao, G. The effects of globose scale (Sphaerolecanium prunastri) infestation on the growth of wild apricot (Prunus armeniaca) trees. Forests 2023, 14, 2032. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, Z.; Linghu, W.; Wang, Q.; Gao, G. Occurrence and harm of Sphaerolecanium prunastri in wild fruit forests in Xinjiang. Xinjiang Agric. Sci. 2022, 59, 1741–1747. [Google Scholar]
- Diao, Y. Primary research on generative biological characteristics of wild apricot(Armeniaca vulgaris Lam.) in Xinjiang. Master, Xinjiang Agriculture University, Xinjiang, 2009.
- You, L.; Chen, G.; Liu, L.; Jie, G.; Liao, K. Dynamics and correlation analysis of growth and development of root system and above -ground part of Armeniaca vulgaris Lam. Xinyuan wild fruit forest. J. Xinjiang Agric. Univ. 2019, 42, 9–14. [Google Scholar]
- Durovic, G.; Ülgentürk, S. Description of plum scale immatures stages (Sphaerolecanium prunastri (Boyer de Fonscolombe) (Hemiptera: Coccidae). YYU J AGR SCI 2021, 31, 982–994. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, Y.; Deng, B.; Ahmad, M.; Zalucki, M.P.; Gao, G.; Lu, Z. Harmonia axyridis (Boyer de Fonscolombe) (Hemiptera: Coccoidea) as a potential biological control agent of the invasive soft scale, Sphaerolecanium prunastri (Boyer de Fonscolombe) (Hemiptera: Coccoidea) in native wild apricot forests. Egypt J Biol Pest Control 2024, 34, 28. [Google Scholar] [CrossRef]
- Zirao, L.; Huang, L.; Zhao, W.; Yao, Y. A new species of the genus Coccophagus (Hymenoptera: Aphelinidae) associated with Sphaerolecanium prunastri (Hemiptera: Coccoidea) from the Tianshan Mountains, Xinjiang. Entomotaxonomia 2022, 44, 228–239. [Google Scholar]
- Cao, Z.; Zhang, Z.; Kang, N.; ZHao, Q.; Hu, H. Research of parasitoids of Sphaerolecanium prunastri fonscolombe in the western Tianshan wild fruit forest. Xinjiang Agric. Sci. 2024, 61, 971–983. [Google Scholar]
- Wang, Y. Biology of Sphaerolecanium prunastri and its control by dominant natural enemy in the wild fruit forest in Yili Master, Xinjiang Agriculture University, Xinjiang, 2021.
- Park, Y.S.; Chung, Y.J. Hazard rating of pine trees from a forest insect pest using artificial neural networks. For. Ecol. Manag 2006, 222, 222–233. [Google Scholar] [CrossRef]
- Polat, A.A.; Çalışkan, O. Fruit set and yield of apricot cultivars under subtropical climate conditions of Hatay, Turkey. Agric. Sci. Technol 2014, 16, 863–872. [Google Scholar]
- Tarantino, A.; Lops, F.; Disciglio, G.; Lopriore, G. Effects of plant biostimulants on fruit set, growth, yield and fruit quality attributes of ‘Orange rubis’ apricot (Prunus armeniaca L.) cultivar in two consecutive years. Sci. Hortic 2018, 239, 26–34. [Google Scholar] [CrossRef]
- Ouyang, Y.; Gong, X.; Duan, C.; Zhang, C.; Chenyang, M.; Han, P.; Zhang, Y.; Hao, G. Water-and carbon-related physiological mechanisms underlying the decline of wild apricot trees in Ili, Xinjiang, China. Chin. J. Plant Ecol 2024, 48, 1192–1201. [Google Scholar]
- Tiffin, P. Mechanisms of tolerance to herbivore damage:what do we know? Evol. Ecol 2000, 14, 523–536. [Google Scholar] [CrossRef]
- Hawkes, C.V.; Sullivan, J.J. The impact of herbivory on plants in different resource conditions: a meta-analysis. Ecology 2001, 82, 2045–2058. [Google Scholar] [CrossRef]
- Stevens, M.T.; Kruger, E.L.; Lindroth, R.L. Variation in tolerance to herbivory is mediated by differences in biomass allocation in aspen. Funct Ecol 2008, 22, 40–47. [Google Scholar] [CrossRef]
- Zvereva, E.L.; Lanta, V.; Kozlov, M.V. Effects of sap-feeding insect herbivores on growth and reproduction of woody plants: a meta-analysis of experimental studies. Oecologia 2010, 163, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Dilley, B.; Davies, D.; Glass, T.; Bond, A.; Ryan, P. Severe impact of introduced scale insects on Island Trees threatens endemic finches at the Tristan da Cunha archipelago. Biol. Conserv 2020, 251, 108761. [Google Scholar] [CrossRef]
- Larson, K.C.; Whitham, T.G. Competition between gall aphids and natural plant sinks: plant architecture affects resistance to galling. Oecologia 1997, 109, 575–582. [Google Scholar] [CrossRef]
- Gonda-King, L.; Gómez, S.; Martin, J.L.; Orians, C.M.; Preisser, E.L. Tree responses to an invasive sap-feeding insect. Plant Ecol 2014, 215, 297–304. [Google Scholar] [CrossRef]
- Shi, D.; Guo, C.; Jiang, N.; Tang, Y.; Zhen, F.; Wang, J.; Liao, K.; Liu, L. Characteristics and spatial distribution pattern of natural regeneration young plants of Prunus armeniaca in Xinjiang, China. Chin. J. Plant Ecol 2023, 47, 515–529. [Google Scholar] [CrossRef]
- Linghu, W.; Lu, Z.; Wang, Y.; Wang, Q.; Gao, G. Effects of pruning time and intensity on the population density of Sphaerolecanium prunastri (Boyer de Fonscolombe) and growth of Armeniaca vulgaris Lamarck. Northern Horticulture 2021, 34–41. [Google Scholar]
- Zhang, P.; Li, Y.; Wang, T.; Zhang, X.; Yanlong, Z.; Xu, H.; Jashenko, R.; Dong, Z.; Zalucki, M.; Lu, Z. Pruning can recover the health of wild apple forests attacked by the wood borer Agrilus mali in central Eurasia. Entomol Gen 2024, 44, 545–552. [Google Scholar] [CrossRef]
Figure 1.
The density dynamics of 1st and 2nd instar nymph from 2019 to 2024 in wild apricot forests (no data in 2023). Capital letters represent the difference of 2nd instar nymph among years; lowercase letters represent the difference of 1st instar nymph among years. ND: no data.
Figure 1.
The density dynamics of 1st and 2nd instar nymph from 2019 to 2024 in wild apricot forests (no data in 2023). Capital letters represent the difference of 2nd instar nymph among years; lowercase letters represent the difference of 1st instar nymph among years. ND: no data.
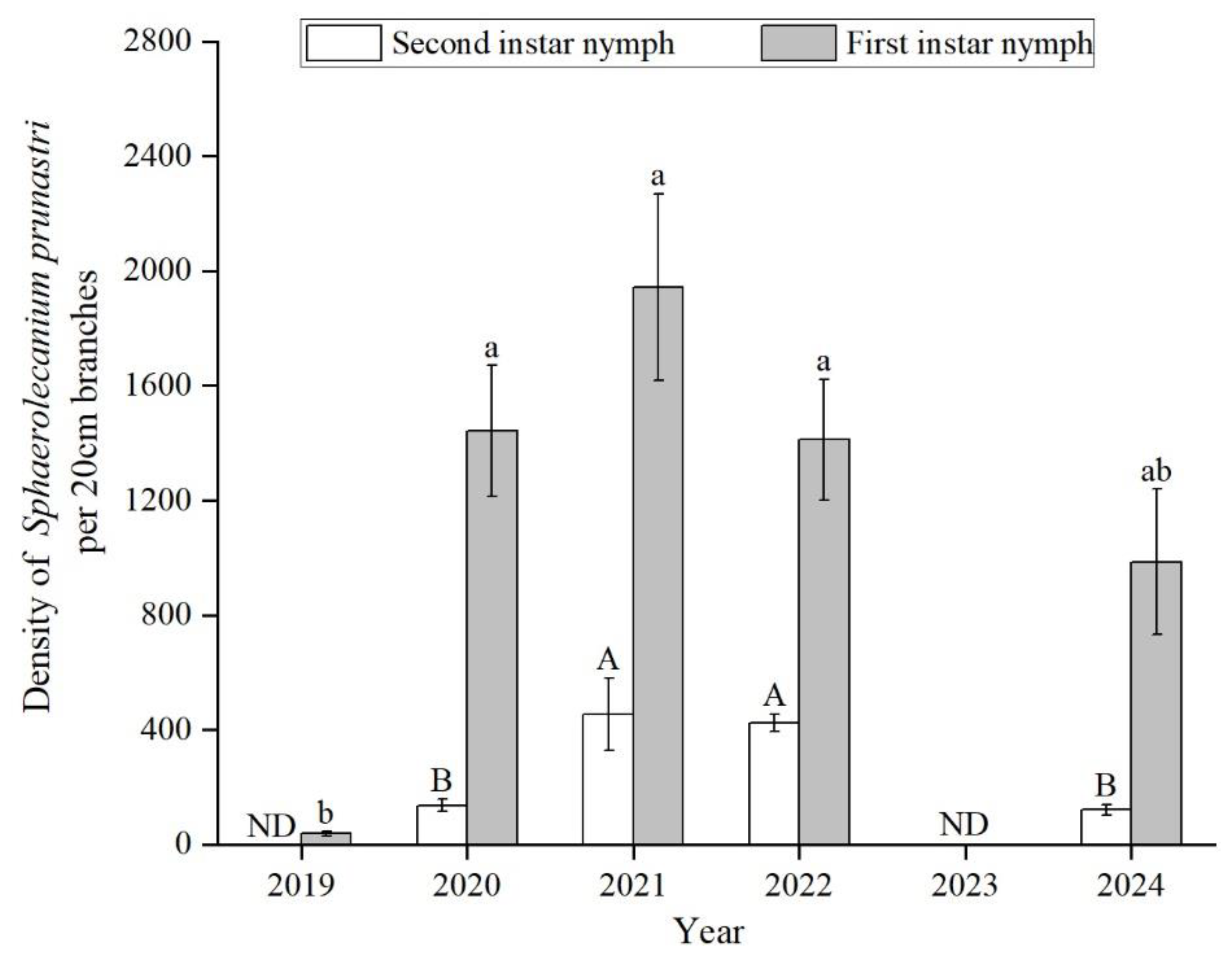
Figure 2.
Proportion of trees in different damage ranking (a) and proportion of dead and live wild apricot trees over time (b).
Figure 2.
Proportion of trees in different damage ranking (a) and proportion of dead and live wild apricot trees over time (b).
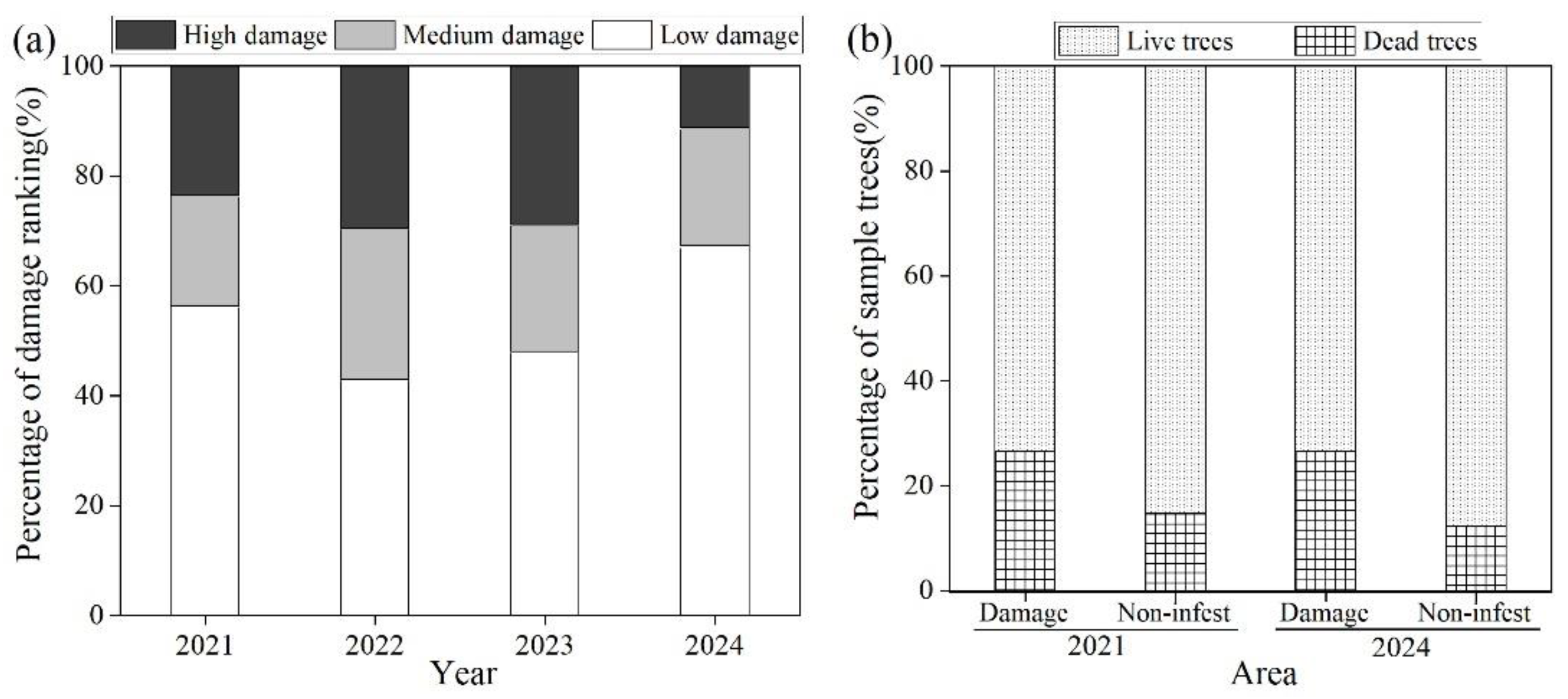
Figure 3.
The length of spring shoots among three damage rankings. Capital letters represent the difference among years; lowercase letters represent the difference of the spring shoot length among three damage rankings.
Figure 3.
The length of spring shoots among three damage rankings. Capital letters represent the difference among years; lowercase letters represent the difference of the spring shoot length among three damage rankings.
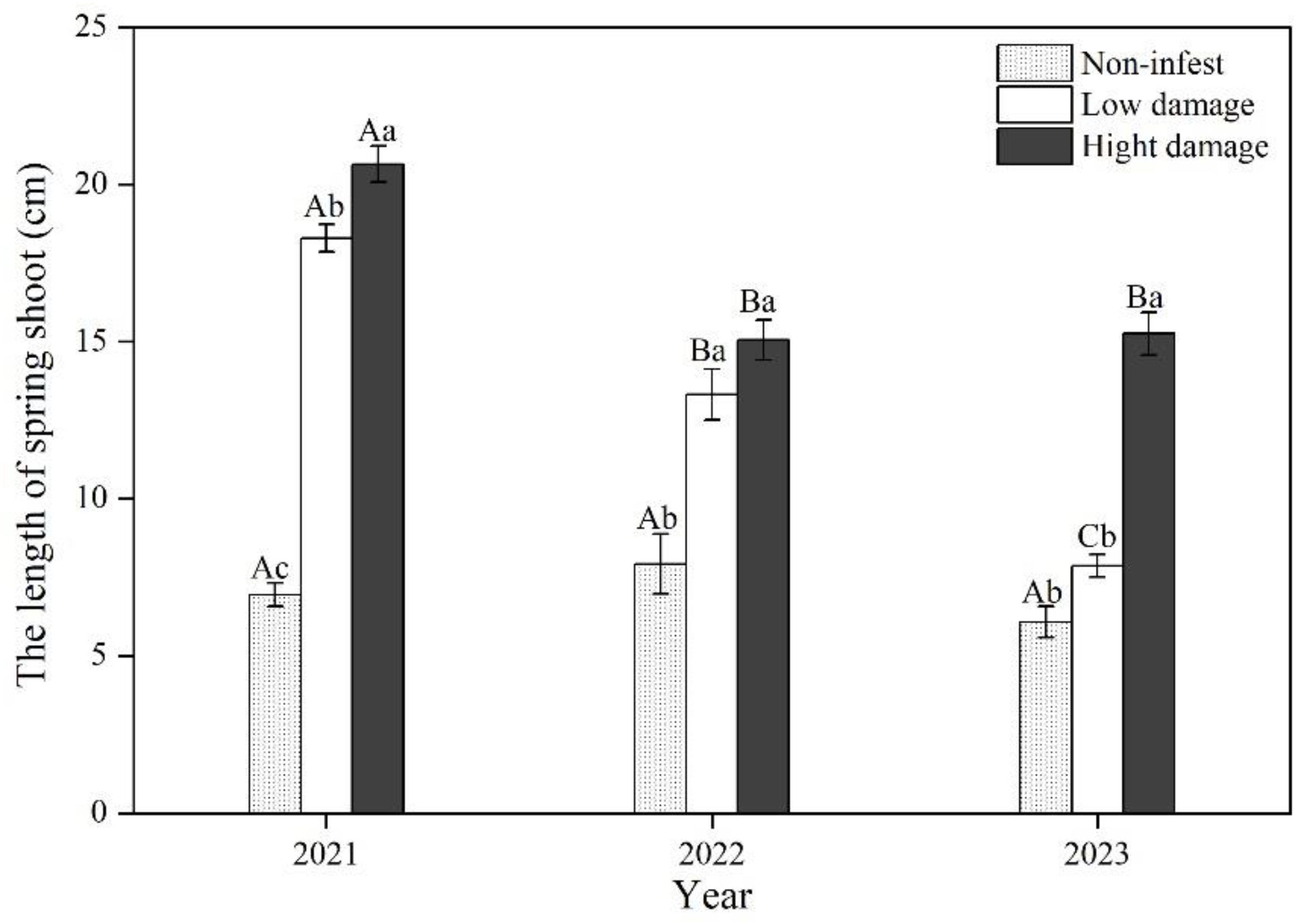
Figure 4.
The density of flower bud (a) and percentage of flowering per wild apricot tree (b) associated with three damage rankings.
Figure 4.
The density of flower bud (a) and percentage of flowering per wild apricot tree (b) associated with three damage rankings.
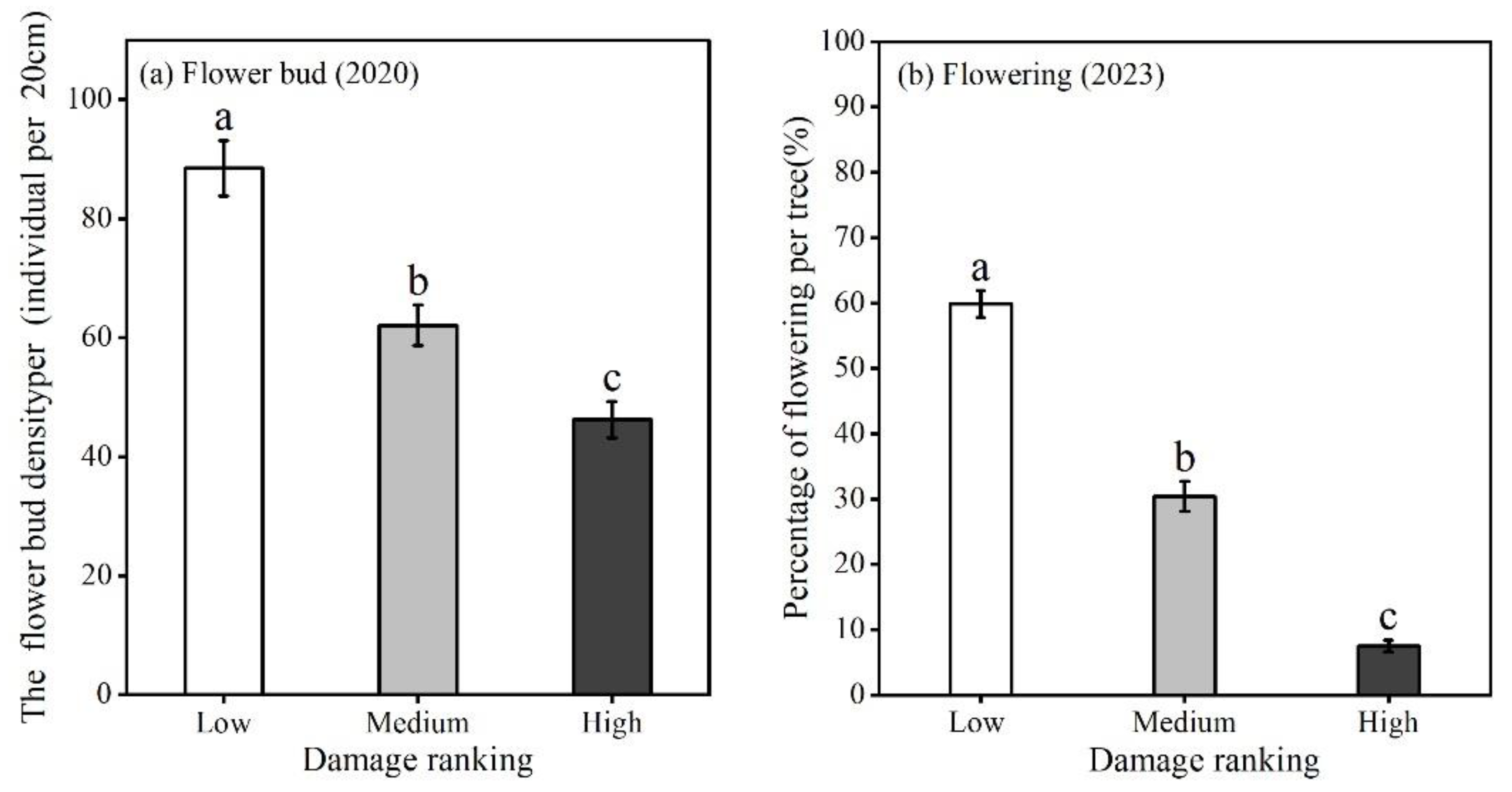
Figure 5.
The fruit yield of wild apricot among three damage rankings. Capital letters represent the difference of fruit yield among years; lowercase letters represent the difference of fruit yield among three damage rankings.
Figure 5.
The fruit yield of wild apricot among three damage rankings. Capital letters represent the difference of fruit yield among years; lowercase letters represent the difference of fruit yield among three damage rankings.
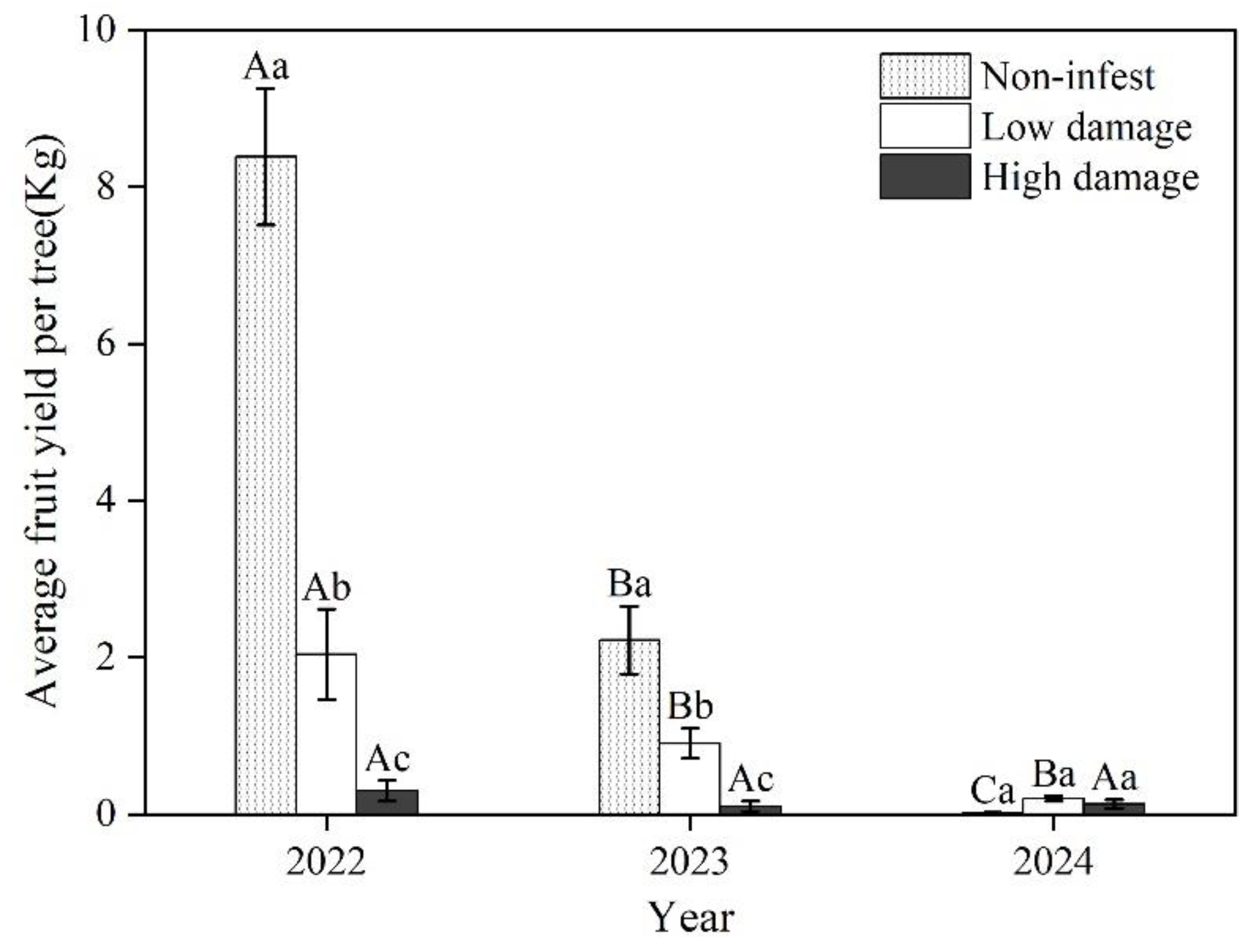
Figure 6.
Average fruit yield per survey site relative to damage ranking score of wild apricot in 2022(a) and 2023(b). The line shows the linear regression.
Figure 6.
Average fruit yield per survey site relative to damage ranking score of wild apricot in 2022(a) and 2023(b). The line shows the linear regression.
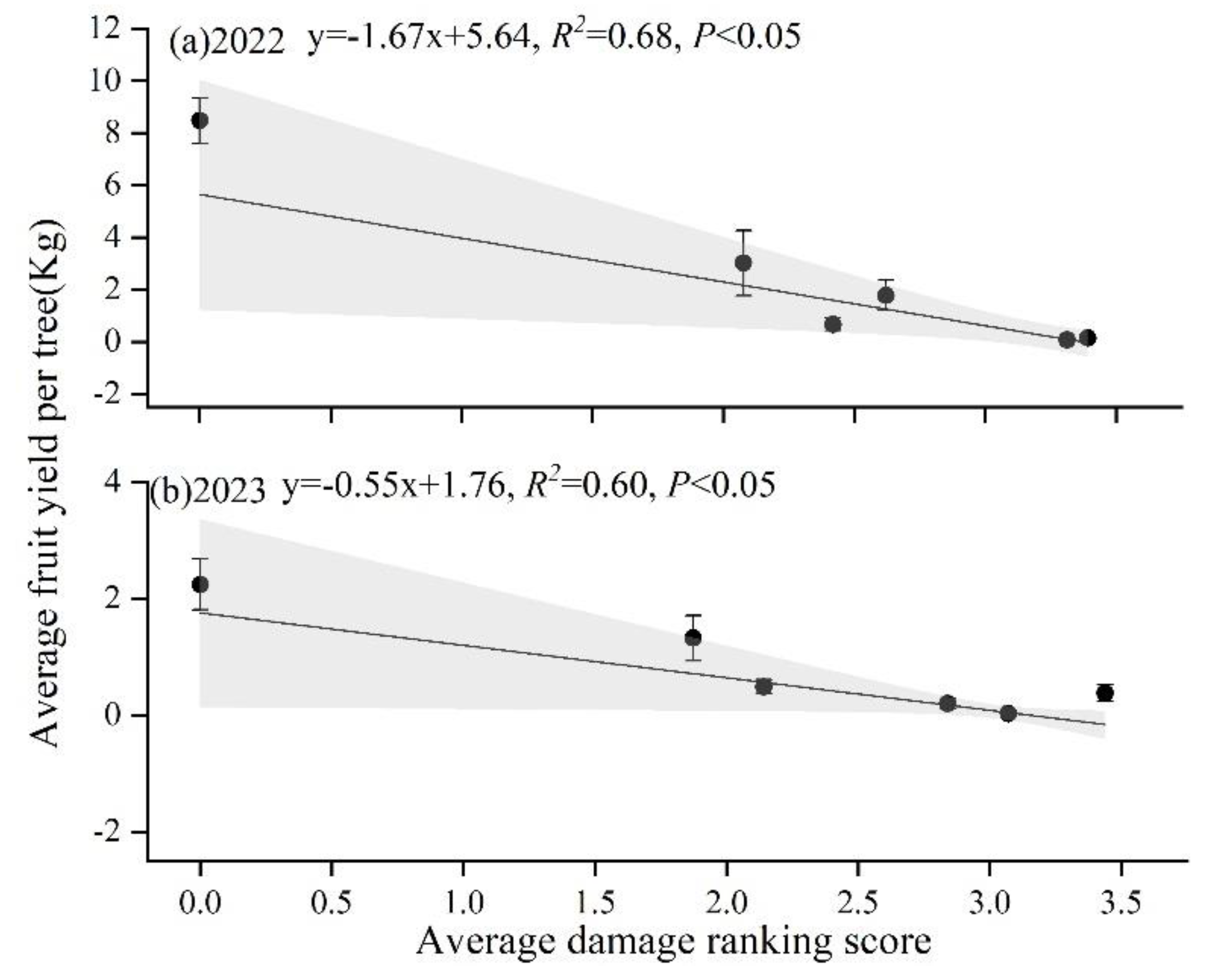

Table 1.
Location of sample plots in remnant wild apricot forests in Ili River basin, Tianshan mountains. Tree density was assessed in 2021.
Table 1.
Location of sample plots in remnant wild apricot forests in Ili River basin, Tianshan mountains. Tree density was assessed in 2021.
| County | Survey site | Plot number | Latitude (N) | Longitude € | Altitude (m) | Height (m) | DBH (cm) | Wild apricot trees density (number /ha) |
|---|---|---|---|---|---|---|---|---|
| Gong liu | Kuolesai | 1 | 43.234 | 82.811 | 1248.10 | 7.93±0.35 | 21.81±1.48 | 224 |
| 2 | 43.259 | 82.857 | 1343.23 | 7.28±0.40 | 13.57 ±1.68 | 248 | ||
| 3 | 43.248 | 82.697 | 1190.32 | 8.28±0.24 | 23.78±1.73 | 264 | ||
| Jinqikesai | 1 | 43.248 | 82.857 | 1321.14 | 7.79±0.37 | 22.2±2.52 | 288 | |
| 2 | 43.246 | 82.857 | 1343.23 | 7.94±0.43 | 21.96±1.5 | 240 | ||
| 3 | 43.248 | 82.858 | 1319.34 | 7.25±0.44 | 15.57±1.56 | 312 | ||
| Keersenbulake | 1 | 43.363 | 82.120 | 1252.33 | 6.30±0.41 | 18.26±1.36 | 264 | |
| 2 | 43.349 | 82.298 | 1237.30 | 8.31±0.51 | 17.50±1.71 | 232 | ||
| 3 | 43.349 | 82.299 | 1211.59 | 8.05±0.70 | 15.92±0.98 | 240 | ||
| Xiaomohuer | 1 | 43.178 | 82.733 | 1376.60 | 8.02±0.26 | 27.99±0.88 | 176 | |
| 2 | 43.141 | 82.442 | 1245.00 | 9.49±0.45 | 22.25±1.29 | 264 | ||
| 3 | 43.177 | 82.733 | 1356.96 | 6.33±0.57 | 27.52±1.35 | 152 | ||
| Xin yuan | Tuergeng | 1 | 43.561 | 83.482 | 1026.33 | 6.05±0.56 | 19.98±2.02 | 360 |
| 2 | 43.560 | 83.401 | 1110.72 | 6.24±0.61 | 26.6±3.37 | 288 | ||
| 3 | 43.539 | 83.485 | 1055.23 | 7.54±0.95 | 19.92±2.90 | 264 | ||
| Talede | 4 | 43.338 | 83.039 | 1165.06 | 6.77±0.51 | 20.94±1.37 | 240 | |
| Huo cheng | Xiaoxigou | 1 | 44.380 | 80.818 | 1084.52 | 5.72±0.47 | 27.17±1.74 | 208 |
| 2 | 44.429 | 80.832 | 1160.95 | 5.62±0.22 | 25.37±1.70 | 216 | ||
| 3 | 44.394 | 80.814 | 1051.36 | 6.39±0.22 | 18.57±1.25 | 232 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



