Submitted:
21 November 2025
Posted:
24 November 2025
You are already at the latest version
Abstract
Obesity treatments increasingly target multiple pathways beyond appetite suppression. We evaluated KBN2202, a salicylate-derived small molecule, in a high-fat diet (60% kcal) mouse model using female and male C57BL/6J mice treated for 8 weeks with oral KBN2202 (20 mg/kg/day) or matched-volume vehicle (1% DMSO/PBS). Body weight and food intake were tracked weekly; serum hormones/cytokines, adipose tissue histology, and open-field behavior were assessed at study end. KBN2202 significantly reduced peri-ovarian gonadal white adipose tissue (gWAT) mass and adipocyte size in females without altering overall body weight. Circulating glucagon-like peptide-1 (GLP-1) increased, uncoupling protein 1 (UCP1) in gWAT showed a non-significant upward trend, and serum TNF-α was selectively decreased, while MCP-1 and IL-1β were unchanged. Despite higher apparent food intake, locomotor activity was unaltered and anxiety-like behavior was reduced. Male mice did not show comparable adipose effects. These findings indicate depot-specific, peripheral modulation of adipose remodeling, hormonal balance, and inflammatory tone by KBN2202, supporting its further investigation as an adipose-targeted metabolic modulator complementary to incretin-based therapies.
Keywords:
white adipose tissue
; obesity
; high-fat diet
; GLP-1
; TNF-α
1. Introduction
Obesity is defined as an abnormal or excessive accumulation of body fat and is regarded as a complex disease that leads to various metabolic and functional impairments beyond simple weight gain. The development of obesity is primarily driven by high-calorie diets, physical inactivity, imbalanced energy metabolism, and genetic factors [1]. In particular, high-fat and high-sugar diets cause a chronic energy surplus, which promotes adipocyte hyperplasia and hypertrophy, resulting in increased fat mass and inflammatory changes [2]. Visceral fat, such as gonadal white adipose tissue (gWAT), plays a central role in obesity-related metabolic dysfunction by secreting pro-inflammatory cytokines and contributing to insulin resistance [3].
Excessive fat accumulation contributes to a wide range of health problems. Direct consequences include joint pain, sleep apnea, and reduced physical mobility due to increased body weight [4]. Indirectly, obesity is strongly associated with a higher risk of chronic conditions such as type 2 diabetes, hypertension, dyslipidemia, cardiovascular disease, non-alcoholic fatty liver disease, and certain types of cancer [5,6]. Notably, the pathological expansion of adipocytes promotes immune cell infiltration and the secretion of pro-inflammatory cytokines such as TNF-α, IL-1β, MCP-1, and IL-6, leading to a state of low-grade chronic inflammation [7,8]. This inflammatory condition is recognized as a central mechanism that exacerbates the development and progression of obesity-related metabolic disorders [2,7,9].
In recent years, obesity treatment has evolved beyond simple weight loss to focus on metabolic improvement and long-term safety [10]. For example, semaglutide, a glucagon-like peptide-1 receptor agonist (GLP-1 RA), has demonstrated significant weight reduction by suppressing appetite and stimulating insulin secretion [11,12,13]. However, these agents still present several limitations. Reported issues include gastrointestinal side effects (e.g., nausea, vomiting, diarrhea, constipation), loss of muscle mass, poor tolerability, and weight regain after treatment discontinuation [11,14,15,16]. These challenges are largely attributed to their single-pathway mechanism of action and their limited capacity to modulate broader metabolic processes. Therefore, there is an increasing demand for multi-targeted therapeutic strategies that regulate both adipose tissue-specific pathways and systemic metabolic hormones more comprehensively.
Salicylic acid, a prototypical nonsteroidal anti-inflammatory drug (NSAID), has long been used to treat inflammatory conditions [17]. It is known to inhibit the NF-κB and IKKβ pathways [18,19], thereby suppressing the production of pro-inflammatory cytokines [20,21]. Beyond its anti-inflammatory actions, emerging evidence suggests that salicylic acid also exerts beneficial metabolic effects in obesity models, including improvements in hepatic steatosis, insulin resistance, and body weight regulation [21,22]. These mechanistic insights have led to increasing interest in salicylate-based derivatives as potential therapeutic agents for modulating obesity-related pathophysiology [23,24].
Based on these considerations, we hypothesized that KBN2202, a novel small-molecule compound structurally derived from salicylic acid, may serve as a multi-targeted therapeutic candidate capable of modulating adipose tissue expansion and metabolic hormone pathways. This hypothesis is supported by the dual role of salicylates in both anti-inflammatory signaling and energy balance regulation. In this context, we specifically focused on evaluating whether KBN2202 could overcome key limitations of current anti-obesity drugs by exerting broader adipose tissue- and hormone-centered mechanisms, rather than relying solely on appetite suppression or body weight reduction.
2. Results
2.1. Effects of KBN2202 on Body Weight and Dietary Intake
Eight weeks of high-fat diet (HFD) consumption resulted in a significant increase in body weight compared to the chow-fed controls in both female and male mice (Figure 1B-C; Supplementary Figure S1A,B). In female, KBN2202 administration showed a trend toward attenuated weight gain without reaching statistical significance, whereas in males, KBN2202 led to a marked increase in body weight compared with chow groups (p < 0.05). Analysis of dietary intake revealed that the HFD group consumed significantly less food than the chow group, while KBN2202-treated mice exhibited a marked increase in food intake compared with the HFD group in both sexes (Figure 1D; Supplementary Figure S1C). In female mice, individual in the KBN2202 group frequently displayed shredding and foraging behaviors, resulting in noticeable scattering of feed within the bedding. Although spilled chow was carefully collected and included in intake calculations, complete recovery was not always possible.
2.2. Suppression of gWAT Expansion and Adipocyte Hypertrophy by KBN2202
To evaluate the effects of KBN2202 on diet-induced obesity, the weights of adipose tissues and major organs were measured. In female mice, HFD feeding significantly increased the weights of gWAT and brown adipose tissue (BAT), indicating enhanced adipogenesis. KBN2202 treatment significantly suppressed gWAT weight gain by approximately 48% compared with the HFD group, whereas BAT weight remained unchanged (Figure 2A). To assess possible changes in muscle mass, the tibialis anterior (TA) muscle was analyzed, no significant differences in TA weight were observed among groups (Figure 2B). Although KBN2202 markedly reduced gWAT weight, total body weight remained unchanged, likely because modest increases in other organs (e.g., the liver, heart and kidney) offset the reduction in gWAT (Figure 2A-C, Figure 1B,C). In contrast, male mice did not exhibit a comparable response to KBN2202 (Supplementary Figure S1D–F). HFD feeding increased both gWAT and BAT weights relative to chow controls, but KBN2202 administration did not alter these parameters. To further investigate the anti-adipogenic effect of KBN2202 in females, adipocyte size in gWAT was quantified. After 8 weeks of HFD feeding, the average adipocyte area increased approximately three-fold (from ~1000 μm² to ~3800 μm²), indicating marked adipocyte hypertrophy (Figure 2D). KBN2202 treatment significantly reduced adipocyte size by nearly 50% compared to the HFD group.
2.3. Modulation of Thermogenic Markers and GLP-1 Levels by KBN2202
To evaluate the effects of KBN2202 on thermogenic and hormonal pathways, serum levels of GLP-1 and growth differentiation factor 15 (GDF15) and uncoupling protein 1 (UCP1) expression in gWAT were examined. Serum GLP-1 levels were significantly higher in the KBN2202 group than in the HFD group (p < 0.05) (Figure 3A). In contrast, serum GDF15 levels, which were markedly elevated in the HFD group, were significantly reduced following KBN2202 treatment, returning to values comparable to those of the chow group (p < 0.05) (Figure 3B). Immunohistochemical analysis showed that UCP1 expression was markedly decreased in the HFD group compared with the chow group (Figure 3C). KBN2202 tended to increase UCP1 immunoreactivity in gWAT compared with the HFD group, but this difference did not reach statistical significance (p = 0.07). (Figure 3D). Pearson correlation analysis revealed a positive correlation between UCP1 expression in gWAT and serum GLP-1 levels (Figure 3E), suggesting a potential link rather than a confirmed causal relationship. Conversely, a significant negative correlation was observed between the adipocyte area and UCP1 expression (Figure 3F), indicating that higher UCP1 levels may be associated with smaller adipocyte size.
2.4. Effects of KBN2202 on Systemic Inflammation in HFD-Fed Mice
To evaluate the potential anti-inflammatory effects of KBN2202 in an HFD-induced obesity model, serum concentrations of eight cytokines (MCP-1, IFN-γ, IL-1β, IL-4, IL-6, IL-10, IL-13, and TNF-α) were measured using a multiplex bead-based assay (Figure 4). Most cytokines were below the limit of blank (LOB) across all groups, indicating minimal systemic inflammatory activation under the current experimental conditions. Among detectable analytes, TNF-α showed a consistent reduction following KBN2202 treatment (p < 0.01; Figure 4A), whereas MCP-1 and IL-1β remained unchanged (Figure 4B,C). In the KBN2202 group, two samples were below the LOB and the remaining three produced identical plotted concentrations because median fluorescence intensity (MFI) signals on the Luminex® 100/200™ platform are displayed in 0.5-unit increments. The underlying fluorescence values, however, differed slightly (CV < 3%), confirming that these were independent biological replicates.
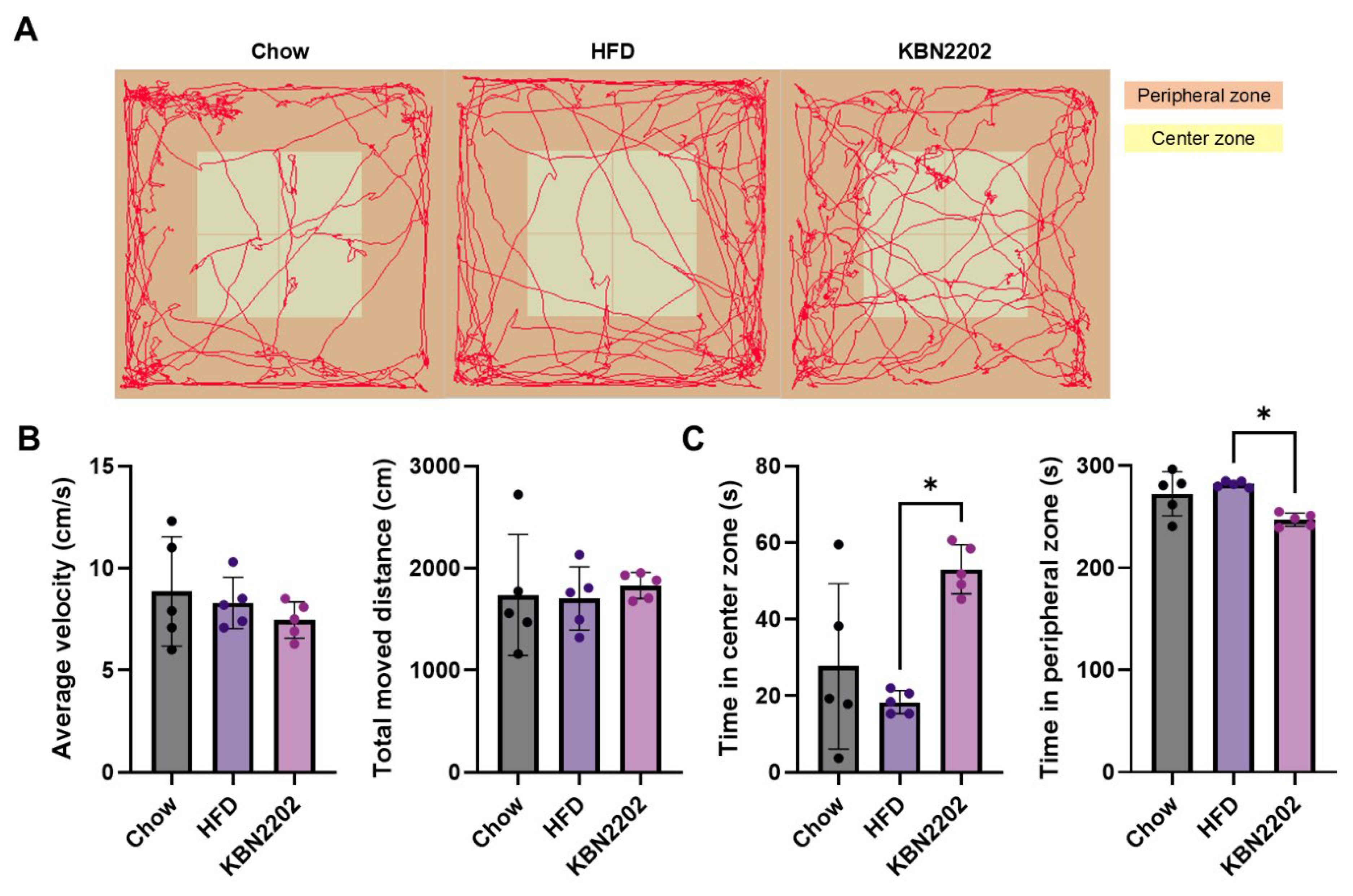
2.5. Effects of KBN2202 on Locomotor Activity and Anxiety-Like Behavior in the Open Field Test
To evaluate the effects of KBN2202 on locomotion and anxiety-related behavior, mice were subjected to the open field test (OFT) after eight weeks of HFD feeding. Representative locomotor tracks demonstrated no apparent differences in general movement patterns among groups (Figure 5A). Quantitative analysis confirmed that neither mean locomotor speed nor total distance traveled differed significantly across groups (Figure 5B), indicating no impact on baseline locomotor activity. However, KBN2202-treated mice spent significantly more time in the center zone and less time in the peripheral zone compared to the HFD group (p < 0.05; Figure 5C).
3. Discussion
Recent advances in obesity therapeutics have shifted toward multi-targeted approaches that address the limitations of current pharmacological agents, moving beyond weight loss strategies. For instance, although GLP-1 RAs are effective in reducing body weight, their efficacy appears to be reduced in patients with type 2 diabetes compared with non-diabetic individuals [25]. To overcome these limitations, combination therapies based on entero-pancreatic hormones are currently under development [26]. Additionally, to mitigate the adverse effect of muscle mass loss associated with GLP-1 RAs, preclinical studies are investigating the co-administration of GLP-1 RAs with bimagrumab, an antibody that blocks activin type II receptors [27]. These examples reflect a growing interest in combinatorial strategies that modulate multiple metabolic pathways simultaneously. Therefore, we investigated the anti-obesity effects of KBN2202, a novel small molecule structurally derived from salicylic acid, which possesses both anti-inflammatory [17,18,19] and metabolic regulatory properties [21,24,28] in a mouse model of HFD-induced obesity.
As shown in the male group, KBN2202 treatment did not attenuate body-weight gain or reduce adipose tissue mass, whereas female mice exhibited clear anti-adipogenic effects. These results indicate a sex-dependent difference in responsiveness to KBN2202. Because all experimental groups in the present study shared the same sex and hormonal background, comparisons between HFD and KBN2202-treated female mice minimized variability associated with sex hormones. Nevertheless, several studies have reported that hormonal and pharmacological pathways can interact to potentiate anti-obesity or anti-inflammatory actions [29,30]. The more pronounced efficacy of KBN2202 in females may be related to such cross-talk mechanisms. Estrogen signaling interacts with multiple metabolic pathways, including AMPK and NF-κB, which are also targeted by salicylate derivatives [18,31]. It is therefore possible that the anti-adipogenic and anti-inflammatory effects of KBN2202 in female mice are partly facilitated by estrogen-dependent modulation of these signaling networks.
HFD consumption induces body-weight gain through a chronic positive energy balance, leading to excessive caloric intake, increased adipogenesis, and reduced energy expenditure [32,33]. Mechanistically, this process promotes adipocyte hypertrophy and hyperplasia, particularly in visceral fat depots such as gWAT, which contribute to systemic inflammation and insulin resistance [34]. Moreover, HFD increases BAT mass as a compensatory response to elevated energy intake; however, thermogenic capacity in BAT and beige adipocytes are often impaired, limiting their ability to counteract obesity [35,36]. Current anti-obesity agents such as GLP-1 RA primarily reduce body weight by suppressing appetite through central mechanisms and secondarily improve glycemic control [11,37]. While appetite suppression plays a critical role in energy balance, direct regulation of adipose tissue expansion and function is also essential in combating obesity.
In this context, KBN2202 significantly suppressed HFD-induced gWAT expansion and adipocyte hypertrophy without affecting total food intake or locomotor activity. Although KBN2202 increased circulating GLP-1 levels and showed a non-significant trend toward higher UCP1 expression, these findings do not constitute direct evidence of enhanced thermogenesis. Nonetheless, targeting UCP1 activation remains an attractive approach to increase energy dissipation and promote metabolic flexibility [35,38,39]. Interestingly, several GLP-1 RAs, such as semaglutide, have been reported to induce mild browning of white adipose tissue and upregulate UCP1 expression, though this is generally considered a secondary consequence of appetite suppression [39]. In contrast, KBN2202’s effects on gWAT remodeling and hormonal regulation may indicate peripheral modulation of adipose tissue metabolism independent of central appetite pathways. The observed positive correlation between UCP1 and GLP-1 levels, together with the inverse association between UCP1 and adipocyte size, suggests a potential link between hormonal signaling and adipose metabolic capacity.
GDF15 is a stress-responsive cytokine belonging to the transforming growth factor-β (TGF-β) superfamily [40]. It is induced by various cellular stressors, including inflammation, hypoxia, mitochondrial dysfunction, and nutrient excess, and has been implicated in the regulation of energy balance and body weight [40,41]. Elevated circulating GDF15 levels are often observed in pathological states such as cancer, cardiovascular disease, and metabolic disorders, where it acts as an anorexigenic signal via its receptor GFRAL in the hindbrain to suppress appetite [40]. Based on these properties, recombinant GDF15 and GDF15 analogs have been explored as potential therapeutic agents for obesity, demonstrating robust weight loss effects in preclinical and early-phase clinical studies [42,43]. Importantly, several studies have reported that HFD feeding significantly increases circulating GDF15 levels in mice, suggesting its role as a metabolic stress marker in diet-induced obesity models [44,45]. In our study, serum GDF15 levels were significantly increased in HFD-fed mice, consistent with its role as a stress-induced cytokine in response to metabolic overload. In particular, KBN2202 treatment markedly reduced elevated GDF15 levels toward those seen in chow groups. Normalization of GDF15 may indicate mitigation of metabolic stress, though the mechanistic link with KBN2202 remains unclear.
Pro-inflammatory cytokines such as TNF-α, IL-1β, and MCP-1 are key mediators of adipose tissue remodeling, promoting lipogenesis, hypertrophy, and insulin resistance [46]. However, in the present study, serum levels of these cytokines were not significantly elevated in HFD-fed mice despite marked adipocyte hypertrophy and gWAT expansion. This cytokine pattern differs from that typically reported in long-term obesity models, where sustained HFD feeding induces systemic low-grade inflammation [47]. The discrepancy may reflect the relatively short duration of HFD exposure (8 weeks) or an early metabolic stage characterized by adipose remodeling rather than overt systemic inflammation [48]. Similar temporal dissociation between adiposity and circulating cytokine elevation has been observed in other short-term HFD models, in which local inflammatory activation precedes systemic cytokine release [49]. Another possible factor is assay sensitivity and biological variability, as low-grade inflammation in mild obesity can produce near-threshold cytokine levels that fluctuate around the lower limit of quantification [2]. Notably, KBN2202 treatment selectively reduced serum TNF-α concentrations even in the absence of systemic cytokine elevation. This selective modulation suggests that KBN2202 may influence intracellular signaling pathways involved in TNF-α regulation, potentially including the IKKβ/NF-κB axis, a known pharmacological target of salicylic acid derivatives [50]. By attenuating this pathway, KBN2202 could locally modulate inflammatory tone within adipose tissue and improve adipocyte function. TNF-α produced within adipose depots, rather than in circulation, is known to drive adipocyte dysfunction and insulin resistance [51]. Therefore, even under low systemic inflammatory conditions, partial suppression of TNF-α by KBN2202 may contribute to improved adipose homeostasis rather than broad anti-inflammatory effects.
The KBN2202 group showed an apparent increase in measured food intake, which may partially reflect behavioral factors such as feed shredding, foraging, or stress-relieving chewing rather than true hyperphagia. To further examine whether such behavioral alterations were associated with stress or locomotor changes, we conducted the open field test. This test not only allows assessment of anxiety-like behavior but also provides an index of locomotor activity, which can influence body-weight dynamics and energy expenditure. The results revealed that KBN2202-treated mice spent more time in the inner zone without changes in total distance traveled, indicating reduced anxiety-like behavior without altered general activity. These findings support the interpretation that the observed differences in food handling and body-weight outcomes were not due to hyperactivity or stress-induced behavioral artifacts, but instead reflect metabolic modulation by KBN2202. Although KBN2202 increased measured food intake and reduced anxiety-like behavior without altering locomotion, these findings do not establish causal links to metabolism. The amygdala–Neuropeptide Y (NPY) axis has been implicated in stress-related feeding and emotion regulation [52], but central pathways were not examined here. Thus, any central nervous system (CNS) interpretation remains speculative and beyond the scope of this study.
In conclusion, KBN2202 modulated adipose remodeling and selected metabolic parameters in an HFD-induced obesity model by suppressing adipose tissue expansion, modulating circulating levels of GLP-1, and showing a potential but unconfirmed effect on thermogenic gene expression and pro-inflammatory cytokines. Compared with existing therapies such as GLP-1 RAs, KBN2202 appears to provide both metabolic and anti-inflammatory benefits while potentially minimizing muscle-related adverse effects. Its dual regulatory actions on metabolic hormones and inflammatory pathways may offer a more balanced therapeutic profile, particularly for patients unresponsive to mono-targeted agents. However, several limitations should be considered. First, this study did not directly assess energy expenditure or insulin sensitivity, which are critical for fully understanding KBN2202’s metabolic effects. Second, the small sample size (n = 5 mice per group) may limit the statistical robustness of certain analyses, particularly those involving high variability such as UCP1 expression levels. Finally, the evaluation of muscle preservation was limited to TA muscle weight measurements without histological or functional assessments, which restricts definitive conclusions regarding potential muscle-sparing effects. Within these constraints, KBN2202 can be viewed as a peripheral metabolic modulator that acts on adipose tissue remodeling and inflammatory signaling, meriting further investigation alongside incretin-based therapeutic strategies.
4. Materials and Methods
4.1. Animal Model of High-Fat Diet-Induced Obesity
Four-week-old male and female C57BL/6J mice were purchased from DBL Co., Ltd. (Daejeon, South Korea) and housed under controlled conditions (21 ± 2°C, 12-hour light/dark cycle) with ad libitum access to food and water. After a one-week acclimation period, five-week-old male and female mice were randomly assigned to three groups (n=5 per group for each sex): normal chow (Chow), HFD, and HFD plus KBN2202 (KBN2202). All procedures were identical across sexes; sex-stratified analyses were prespecified. Female data are presented in main figures; male data in Supplementary Figure S1. The HFD provided 60% of total calories from fat, had an energy density of 5.24 kcal/g, and was obtained from Research Diets (D12492, Saeron Bio, Republic of Korea). Both male and female groups were maintained on their respective diets for eight weeks to induce obesity. The animal study was approved by the Institutional Animal Care and Use Committee of the Advanced Medical Bio Research Center (IACUC-2024-009) and conducted in accordance with institutional and national guidelines.
4.2. Drug Treatment
KBN2202 is a salicylic acid derivative (molecular formula C19H19NO4; molecular weight 325.35 g/mol) synthesized and provided by KAISER Bio Ltd., (Gyeongsan, Republic of Korea), and formulated as previously described [53]. Briefly, KBN2202 was initially dissolved in 1 mL of DMSO (200 mg/mL) and subsequently diluted with 99 mL of PBS to yield a final solution containing 1% DMSO and 2 mg/mL of the compound. This solution was administered orally at a dose of 20 mg/kg/day via oral gavage using a 20-gauge feeding needle. Mice in the Chow and HFD groups received daily oral gavage of vehicle (1% DMSO in PBS) at a matched volume. Drug administration began concurrently with the high-fat diet and continued for eight weeks (Figure 1A). This concurrent initiation was intended to evaluate the preventive efficacy of KBN2202 against diet-induced adipose expansion and inflammation, rather than to assess a therapeutic effect after obesity establishment.
4.3. Physiological Measurements and Tissue Collection
Body weight was recorded weekly throughout the experimental period, and food intake was measured daily. Food intake was measured with cage at the experimental unit. A fixed amount of chow was supplied every 24 hours, and the remaining food was collected and weighed the next day. Pellets dispersed into bedding were retrieved using a fine sieve and included in the remaining weight whenever possible. The total amount of consumed chow was then divided by the number of mice per cage to calculate average daily intake per mouse. After the final administration of HFD and KBN2202, behavioral assessment was performed using the OFT to evaluate locomotor activity and anxiety-like behavior. Two days after behavioral testing, mice were fasted for 6 h and subsequently anesthetized with Avertin (2,2,2-tribromoethanol, 250 mg/kg, intraperitoneally; T1420, Tokyo Chemical Industry Co., Ltd., Tokyo, Japan), followed by euthanasia via exsanguination. Blood samples were collected in Microtainer® SST tubes (365967; BD, Franklin Lakes, NJ, USA). Major organs—including the liver, heart, kidneys, gWAT, BAT, and TA muscle—were excised and weighed for further analysis. Serum was isolated by centrifugation at 3,000 rpm for 15 minutes at 25°C and stored at −80°C until further use.
4.4. Quantification of Serum Cytokines by ELISA and Multiplex Analysis
Serum levels of GLP-1 (BMS2194; Invitrogen, Waltham, MA, USA) and GDF15 (MGD150; R&D Systems, Minneapolis, MN, USA) were measured using a commercial ELISA kit according to the manufacturer’s instructions. All samples were analyzed in duplicate, and optical density was measured using a microplate reader (INNO-S, LTek, Sungnam, Korea). Serum cytokines, including MCP-1, IFN-γ, IL-1β, IL-4, IL-6, IL-10, IL-13, and TNF-α, were quantified using a multiplex bead-based assay (Mouse XL Cytokine Premixed Kit; R&D Systems, FCSTM20) on a Luminex® 100/200™ platform. Samples were diluted 1:2 in assay buffer and analyzed in duplicate according to the manufacturer’s instructions. MFI values were used for back-calculation of concentrations according to a 5-parameter logistic standard curve. On this platform, MFI signals are displayed in 0.5-unit increments, which can lead to discretized concentration outputs appearing as identical values when the true fluorescence intensities are very close. Each sample was analyzed in duplicate, and samples below the LOB were treated as non-detectable (0 pg/mL).
4.5. Histology and Immunohistochemistry
Tissue samples were fixed in 4% paraformaldehyde (PFA) for 24 h at 4 °C, embedded in paraffin, and sectioned at a thickness of 4 µm. Sections were deparaffinized, rehydrated, and stained using standard protocols for hematoxylin and eosin (H&E) staining. Adipocyte size was quantified from H&E-stained gWAT sections using ImageJ software (NIH, Bethesda, MD, USA). For each animal, three non-overlapping fields (200× magnification) were randomly selected from the central region of the tissue section. Cell boundaries were manually delineated using the “Freehand selection” tool, and adipocyte area (µm²) was measured after automated thresholding with consistent parameters across all samples. Approximately 150–200 adipocytes were analyzed per animal. Image analysis was performed by two independent observers blinded to group allocation. For immunohistochemical analysis of thermogenic activity, sections were deparaffinized, rehydrated, and subjected to antigen retrieval by heating in 10 mM sodium citrate buffer (pH 6.0) at 95 °C for 20 min, followed by cooling at room temperature. Endogenous peroxidase activity was quenched with 2% hydrogen peroxide for 10 min, and non-specific binding was blocked with 5% normal goat serum for 1 h at room temperature. Sections were then incubated overnight at 4 °C with a rabbit anti-UCP1 primary antibody (1:500, ab10983; Abcam, Cambridge, UK). Signal detection was performed using the ImmPRESS®-HRP Horse Anti-Rabbit IgG Polymer Reagent (MP-7401; Vector Laboratories, Newark, CA, USA) and a DAB substrate kit (SK-4103; Vector Laboratories, Newark, CA, USA). Inter-scapular brown adipose tissue (BAT) was used as a positive control for UCP1, and negative controls were prepared by omitting the primary antibody. Representative images were acquired using an OLYMPUS CKX3-HOUN bright-field microscope (Olympus Corporation, Tokyo, Japan). UCP1 expression levels were quantified by measuring mean DAB signal intensity in gWAT sections using ImageJ with consistent thresholding across samples.
4.6. Open Field Test (OFT)
Locomotor activity and anxiety-like behavior were assessed using the open field test in a dedicated sound-attenuated behavioral testing room. To minimize stress caused by external noise, continuous white noise (60 ± 5 dB) was played throughout the experiment. The open-field apparatus consisted of a square arena (40 × 40 × 40 cm) made of opaque white acrylic, placed under uniform illumination of approximately 150 lux provided by an overhead LED light source. Prior to testing, each mouse was acclimated to the behavioral testing room for 30 minutes to minimize novelty-induced stress. Mice were then individually placed in the center of the arena and allowed to freely explore for 10 minutes. A ceiling-mounted camera (Basler acA1300, Germany) positioned 100 cm above the arena was connected to an automated video-tracking system (EthoVision XT, version 11.5; Noldus Information Technology, Wageningen, Netherlands) for continuous tracking of the animal’s center of mass at a sampling rate of 15 frames per second. The center zone was defined as a 20 × 20 cm square in the middle of the arena. Between trials, the arena was thoroughly wiped with odourless wet wipes to remove residual scent cues without introducing stress-inducing odors. Behavioral parameters analyzed included total distance traveled (cm), mean velocity (cm/s), and time spent in the center and peripheral zones (s). To avoid early exploratory bias, behavioral data were analyzed from the final 5 minutes of the 10-minuate session. All data were analyzed offline by an investigator blinded to treatment allocation using the same tracking software.
4.7. Statistical Analysis
All data are presented as mean ± standard deviation (SD), and individual data points are shown in the figures. Statistical analyses were performed using GraphPad Prism 10 (GraphPad Software, San Diego, CA, USA). Data normality and variance homogeneity were verified using the Shapiro–Wilk and Brown–Forsythe tests, respectively. One-way analysis of variance (ANOVA) was used to compare groups, followed by Tukey’s multiple-comparison test when appropriate. Weekly body weight was analyzed using two-way repeated-measures ANOVA. A p < 0.05 was considered statistically significant. All analyses were conducted by investigators blinded to treatment allocation.
Supplementary Materials
The following supporting information can be downloaded at: https://doi.org/10.6084/m9.figshare.30662153, Supplementary Figure S1.
Author Contributions
Conceptualization, M.K., M.R.C. and S.-R.L.; methodology, M.K., J.-H.H. and S.H.C.; validation, M.K., J.-H.H., S.H.C., J.K., S.-Y.L., M.-G.S. and J.S.K.; investigation, M.K., J.-H.H., J.K., S.-Y.L., M.-G.S. and J.S.K.; data curation, M.K., J.-H.H., S.H.C., S.-Y.L., M.-G.S. and J.S.K.; writing—original draft, M.K. and J.-H.H.; writing—review and editing, M.R.C. and S.-R.L.; supervision, M.R.C. and S.-R.L.; project administration, M.R.C. and S.-R.L.; funding acquisition, S.-R.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Korea Bio-health Technology R&D Project (No. RS-2023-00267453) and the Korea Health Technology R&D Project (No. RS-2021-KH113822) through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare. It was also supported by the Bio & Medical Technology Development Program of the National Research Foundation of Korea (NRF), funded by the Ministry of Science and ICT (MSIT) (No. RS-2023-00223559), and by a separate grant from the NRF, also funded by MSIT (No. RS-2021-NR062031).
Institutional Review Board Statement
All animal experimental protocols in this study were approved by the Institutional Animal Care and Use Committee of the Advanced Medical Bio Research Center (approval No. IACUC-2024-009; Approval date: 10 Jun 2024) and reported in accordance with ARRIVE guidelines.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated and analyzed in this study are available from the corresponding author upon reasonable request.
Acknowledgments
During the preparation of this manuscript, the authors used OpenAI’s ChatGPT (GPT-4, accessed via ChatGPT Plus in July 2025) to assist with scientific phrasing and language refinement. All AI-generated content was carefully reviewed and edited by the authors, who take full responsibility for the final content of this publication.
Conflicts of Interest
KBN2202, the compound used in this study, was provided by KAISER Bio Ltd. The company had no involvement in the study design, data acquisition, analysis, or interpretation, nor in the decision to publish the results or the preparation of this manuscript. The authors declare no other competing interests.
Abbreviations
The following abbreviations are used in this manuscript:
| BAT | Brown adipose tissue |
| gWAT | Gonadal white adipose tissue |
| GDF15 | Growth differentiation factor 15 |
| GLP-1 | Glucagon-like peptide-1 |
| GLP-1RA | GLP-1 receptor agonist |
| HFD | High-fat diet |
| LOB | Limit of blank |
| MFI | Median fluorescence intensity |
| NSAID | Nonsteroidal anti-inflammatory drug |
| NPY | Neuropeptide Y |
| OFT | Open field test |
| TA | Tibialis anterior |
| UCP1 | Uncoupling protein 1 |
References
- Lin X, Li H. Obesity: Epidemiology, Pathophysiology, and Therapeutics. Front Endocrinol (Lausanne). 2021;12:706978. [CrossRef]
- Gregor MF, Hotamisligil GS. Inflammatory mechanisms in obesity. Annu Rev Immunol. 2011;29:415–45. [CrossRef]
- Bensussen A, Torres-Magallanes JA, Roces de Alvarez-Buylla E. Molecular tracking of insulin resistance and inflammation development on visceral adipose tissue. Front Immunol. 2023;14:1014778. [CrossRef]
- Bray GA, Kim KK, Wilding JPH, World Obesity F. Obesity: a chronic relapsing progressive disease process. A position statement of the World Obesity Federation. Obes Rev. 2017;18(7):715–23. [CrossRef]
- Guh DP, Zhang W, Bansback N, Amarsi Z, Birmingham CL, Anis AH. The incidence of co-morbidities related to obesity and overweight: a systematic review and meta-analysis. BMC Public Health. 2009;9:88. [CrossRef]
- Bluher M. Obesity: global epidemiology and pathogenesis. Nat Rev Endocrinol. 2019;15(5):288–98. [CrossRef]
- Saltiel AR, Olefsky JM. Inflammatory mechanisms linking obesity and metabolic disease. J Clin Invest. 2017;127(1):1–4. [CrossRef]
- Wellen KE, Hotamisligil GS. Obesity-induced inflammatory changes in adipose tissue. J Clin Invest. 2003;112(12):1785–8. [CrossRef]
- Hotamisligil GS. Inflammation and metabolic disorders. Nature. 2006;444(7121):860–7. [CrossRef]
- Muller TD, Bluher M, Tschop MH, DiMarchi RD. Anti-obesity drug discovery: advances and challenges. Nat Rev Drug Discov. 2022;21(3):201–23. [CrossRef]
- Wilding JPH, Batterham RL, Calanna S, Davies M, Van Gaal LF, Lingvay I, et al. Once-Weekly Semaglutide in Adults with Overweight or Obesity. N Engl J Med. 2021;384(11):989–1002. [CrossRef]
- Wilkinson L, Holst-Hansen T, Laursen PN, Rinnov AR, Batterham RL, Garvey WT. Effect of semaglutide 2.4 mg once weekly on 10-year type 2 diabetes risk in adults with overweight or obesity. Obesity (Silver Spring). 2023;31(9):2249–59. [CrossRef]
- Wadden TA, Bailey TS, Billings LK, Davies M, Frias JP, Koroleva A, et al. Effect of Subcutaneous Semaglutide vs Placebo as an Adjunct to Intensive Behavioral Therapy on Body Weight in Adults With Overweight or Obesity: The STEP 3 Randomized Clinical Trial. JAMA. 2021;325(14):1403–13. [CrossRef]
- Jensen SBK, Blond MB, Sandsdal RM, Olsen LM, Juhl CR, Lundgren JR, et al. Healthy weight loss maintenance with exercise, GLP-1 receptor agonist, or both combined followed by one year without treatment: a post-treatment analysis of a randomised placebo-controlled trial. EClinicalMedicine. 2024;69:102475. [CrossRef]
- Rubino D, Abrahamsson N, Davies M, Hesse D, Greenway FL, Jensen C, et al. Effect of Continued Weekly Subcutaneous Semaglutide vs Placebo on Weight Loss Maintenance in Adults With Overweight or Obesity: The STEP 4 Randomized Clinical Trial. JAMA. 2021;325(14):1414–25. [CrossRef]
- Klimova J, Mraz M, Kratochvilova H, Lacinova Z, Novak K, Michalsky D, et al. Gene Profile of Adipose Tissue of Patients with Pheochromocytoma/Paraganglioma. Biomedicines. 2022;10(3). [CrossRef]
- Vane JR, Botting RM. Mechanism of action of nonsteroidal anti-inflammatory drugs. Am J Med. 1998;104(3A):2S–8S; discussion 21S–22S. [CrossRef]
- Yin MJ, Yamamoto Y, Gaynor RB. The anti-inflammatory agents aspirin and salicylate inhibit the activity of I(kappa)B kinase-beta. Nature. 1998;396(6706):77–80. [CrossRef]
- Kopp E, Ghosh S. Inhibition of NF-kappa B by sodium salicylate and aspirin. Science. 1994;265(5174):956–9. [CrossRef]
- Hardie DG. AMPK: a key regulator of energy balance in the single cell and the whole organism. Int J Obes (Lond). 2008;32 Suppl 4:S7–12. [CrossRef]
- Hawley SA, Fullerton MD, Ross FA, Schertzer JD, Chevtzoff C, Walker KJ, et al. The ancient drug salicylate directly activates AMP-activated protein kinase. Science. 2012;336(6083):918–22. [CrossRef]
- Kim JK, Kim YJ, Fillmore JJ, Chen Y, Moore I, Lee J, et al. Prevention of fat-induced insulin resistance by salicylate. J Clin Invest. 2001;108(3):437–46. [CrossRef]
- Choi HE, Jeon EJ, Kim DY, Choi MJ, Yu H, Kim JI, et al. Sodium salicylate induces browning of white adipocytes via M2 macrophage polarization by HO-1 upregulation. Eur J Pharmacol. 2022;928:175085. [CrossRef]
- Smith BK, Ford RJ, Desjardins EM, Green AE, Hughes MC, Houde VP, et al. Salsalate (Salicylate) Uncouples Mitochondria, Improves Glucose Homeostasis, and Reduces Liver Lipids Independent of AMPK-beta1. Diabetes. 2016;65(11):3352–61. [CrossRef]
- Bergmann NC, Davies MJ, Lingvay I, Knop FK. Semaglutide for the treatment of overweight and obesity: A review. Diabetes Obes Metab. 2023;25(1):18–35. [CrossRef]
- Melson E, Ashraf U, Papamargaritis D, Davies MJ. What is the pipeline for future medications for obesity? Int J Obes (Lond). 2025;49(3):433–51. [CrossRef]
- Nunn E, Jaiswal N, Gavin M, Uehara K, Stefkovich M, Drareni K, et al. Antibody blockade of activin type II receptors preserves skeletal muscle mass and enhances fat loss during GLP-1 receptor agonism. Mol Metab. 2024;80:101880. [CrossRef]
- Deschamps D, Fisch C, Fromenty B, Berson A, Degott C, Pessayre D. Inhibition by salicylic acid of the activation and thus oxidation of long chain fatty acids. Possible role in the development of Reye's syndrome. J Pharmacol Exp Ther. 1991;259(2):894–904.
- Stubbins RE, Najjar K, Holcomb VB, Hong J, Nunez NP. Oestrogen alters adipocyte biology and protects female mice from adipocyte inflammation and insulin resistance. Diabetes Obes Metab. 2012;14(1):58–66. [CrossRef]
- Heine PA, Taylor JA, Iwamoto GA, Lubahn DB, Cooke PS. Increased adipose tissue in male and female estrogen receptor-alpha knockout mice. Proc Natl Acad Sci U S A. 2000;97(23):12729–34. [CrossRef]
- Lopez M, Tena-Sempere M. Estradiol effects on hypothalamic AMPK and BAT thermogenesis: A gateway for obesity treatment? Pharmacol Ther. 2017;178:109–22. [CrossRef]
- Winzell MS, Ahren B. The high-fat diet-fed mouse: a model for studying mechanisms and treatment of impaired glucose tolerance and type 2 diabetes. Diabetes. 2004;53 Suppl 3:S215–9. [CrossRef]
- Choi MS, Kim YJ, Kwon EY, Ryoo JY, Kim SR, Jung UJ. High-fat diet decreases energy expenditure and expression of genes controlling lipid metabolism, mitochondrial function and skeletal system development in the adipose tissue, along with increased expression of extracellular matrix remodelling- and inflammation-related genes. Br J Nutr. 2015;113(6):867–77. [CrossRef]
- An SM, Cho SH, Yoon JC. Adipose Tissue and Metabolic Health. Diabetes Metab J. 2023;47(5):595–611. [CrossRef]
- Ikeda K, Yamada T. UCP1 Dependent and Independent Thermogenesis in Brown and Beige Adipocytes. Front Endocrinol (Lausanne). 2020;11:498. [CrossRef]
- Kajimura S, Spiegelman BM, Seale P. Brown and Beige Fat: Physiological Roles beyond Heat Generation. Cell Metab. 2015;22(4):546–59. [CrossRef]
- Baggio LL, Drucker DJ. Glucagon-like peptide-1 receptors in the brain: controlling food intake and body weight. J Clin Invest. 2014;124(10):4223–6. [CrossRef]
- Fedorenko A, Lishko PV, Kirichok Y. Mechanism of fatty-acid-dependent UCP1 uncoupling in brown fat mitochondria. Cell. 2012;151(2):400–13. [CrossRef]
- Martins FF, Marinho TS, Cardoso LEM, Barbosa-da-Silva S, Souza-Mello V, Aguila MB, et al. Semaglutide (GLP-1 receptor agonist) stimulates browning on subcutaneous fat adipocytes and mitigates inflammation and endoplasmic reticulum stress in visceral fat adipocytes of obese mice. Cell Biochem Funct. 2022;40(8):903–13. [CrossRef]
- Li J, Hu X, Xie Z, Li J, Huang C, Huang Y. Overview of growth differentiation factor 15 (GDF15) in metabolic diseases. Biomed Pharmacother. 2024;176:116809. [CrossRef]
- Chang JY, Hong HJ, Kang SG, Kim JT, Zhang BY, Shong M. The Role of Growth Differentiation Factor 15 in Energy Metabolism. Diabetes Metab J. 2020;44(3):363–71. [CrossRef]
- Hale C, Veniant MM. Growth differentiation factor 15 as a potential therapeutic for treating obesity. Mol Metab. 2021;46:101117. [CrossRef]
- Benichou O, Coskun T, Gonciarz MD, Garhyan P, Adams AC, Du Y, et al. Discovery, development, and clinical proof of mechanism of LY3463251, a long-acting GDF15 receptor agonist. Cell Metab. 2023;35(2):274–86 e10. [CrossRef]
- Patel S, Alvarez-Guaita A, Melvin A, Rimmington D, Dattilo A, Miedzybrodzka EL, et al. GDF15 Provides an Endocrine Signal of Nutritional Stress in Mice and Humans. Cell Metab. 2019;29(3):707–18 e8. [CrossRef]
- L'Homme L, Sermikli BP, Haas JT, Fleury S, Quemener S, Guinot V, et al. Adipose tissue macrophage infiltration and hepatocyte stress increase GDF-15 throughout development of obesity to MASH. Nat Commun. 2024;15(1):7173. [CrossRef]
- Kawai T, Autieri MV, Scalia R. Adipose tissue inflammation and metabolic dysfunction in obesity. Am J Physiol Cell Physiol. 2021;320(3):C375–C91. [CrossRef]
- Kunz HE, Hart CR, Gries KJ, Parvizi M, Laurenti M, Dalla Man C, et al. Adipose tissue macrophage populations and inflammation are associated with systemic inflammation and insulin resistance in obesity. Am J Physiol Endocrinol Metab. 2021;321(1):E105–E21. [CrossRef]
- Kanda H, Tateya S, Tamori Y, Kotani K, Hiasa K, Kitazawa R, et al. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J Clin Invest. 2006;116(6):1494–505. [CrossRef]
- Rubin DA, Hackney AC. Inflammatory cytokines and metabolic risk factors during growth and maturation: influence of physical activity. Med Sport Sci. 2010;55:43–55. [CrossRef]
- Austin RL, Rune A, Bouzakri K, Zierath JR, Krook A. siRNA-mediated reduction of inhibitor of nuclear factor-kappaB kinase prevents tumor necrosis factor-alpha-induced insulin resistance in human skeletal muscle. Diabetes. 2008;57(8):2066–73. [CrossRef]
- Suganami T, Ogawa Y. Adipose tissue macrophages: their role in adipose tissue remodeling. J Leukoc Biol. 2010;88(1):33–9. [CrossRef]
- Ip CK, Zhang L, Farzi A, Qi Y, Clarke I, Reed F, et al. Amygdala NPY Circuits Promote the Development of Accelerated Obesity under Chronic Stress Conditions. Cell Metab. 2019;30(1):111–28 e6. [CrossRef]
- Lee S-Y, Kim JC, Choi MR, Song J, Kim M, Chang S-H, Kim JS, Park J-S and Lee S-R. KBN2202, a Salicylic Acid Derivative, Preserves Neuronal Architecture, Enhances Neurogenesis, Attenuates Amyloid and Inflammatory Pathology, and Restores Recognition Memory in 5xFAD Mice at an Advanced Stage of AD Pathophysiology. Int.J. Mol. Sci. 2025.;26(22). [CrossRef]
Figure 1.
Effects of KBN2202 on body weight and dietary intake in HFD-fed female mice (male data are presented in Supplementary Figure S1). (A) Schematic overview of the 8-week experimental design involving HFD feeding and daily oral administration of KBN2202 (20 mg/kg). (B) Weekly body weight measurements over the 8-week study period. HFD-fed mice showed significant increase in body weight compared to the chow group, while KBN2202-treated females showed a non-significant trend toward reduced weight gain. (C) Final body weight at 8 weeks. (D) Cumulative food intake calculated as the sum of daily consumption during the study. HFD-fed mice consumed significantly less food than chow-fed mice, whereas KBN2202-treated mice showed a significant increase in food intake compared to the HFD group. Data are presented as mean ± SD (n = 5 per group). Statistical significance was determined by one-way ANOVA followed by Tukey’s post hoc test (*p < 0.05, **p < 0.01, ****p < 0.0001).
Figure 1.
Effects of KBN2202 on body weight and dietary intake in HFD-fed female mice (male data are presented in Supplementary Figure S1). (A) Schematic overview of the 8-week experimental design involving HFD feeding and daily oral administration of KBN2202 (20 mg/kg). (B) Weekly body weight measurements over the 8-week study period. HFD-fed mice showed significant increase in body weight compared to the chow group, while KBN2202-treated females showed a non-significant trend toward reduced weight gain. (C) Final body weight at 8 weeks. (D) Cumulative food intake calculated as the sum of daily consumption during the study. HFD-fed mice consumed significantly less food than chow-fed mice, whereas KBN2202-treated mice showed a significant increase in food intake compared to the HFD group. Data are presented as mean ± SD (n = 5 per group). Statistical significance was determined by one-way ANOVA followed by Tukey’s post hoc test (*p < 0.05, **p < 0.01, ****p < 0.0001).
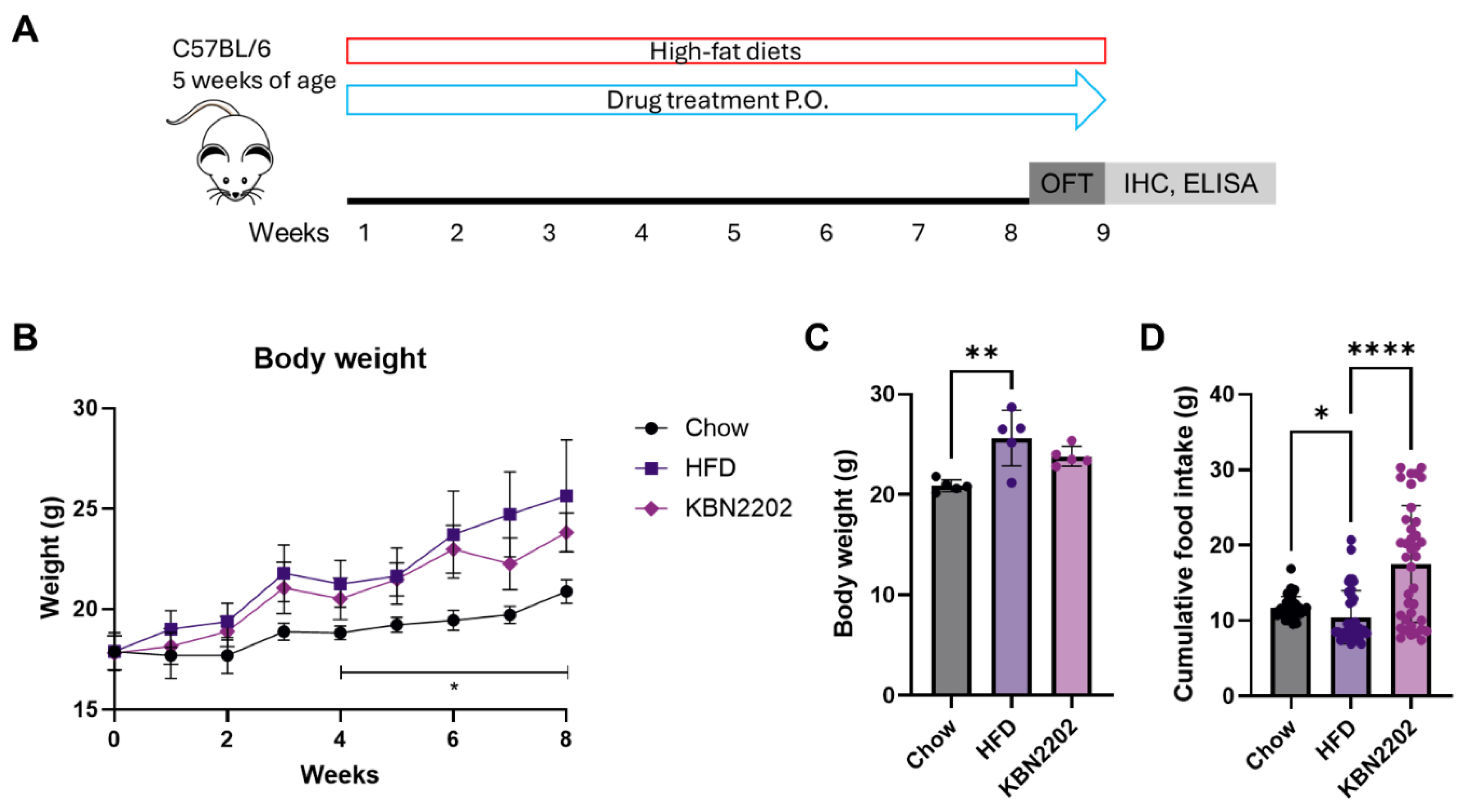
Figure 2.
Effects of KBN2202 on adipose tissue, organ weight, and adipocyte hypertrophy in females (A) Absolute weights of gWAT and BAT. KBN2202 significantly reduced gWAT weight compared to the HFD group, whereas BAT weight was unchanged. (B) Absolute weight of TA muscle (left and right), showing no significant differences among groups. (C) Absolute weights of liver, heart, and kidneys were comparable among groups. (D) Representative H&E-stained images of gWAT and quantification of adipocyte area. HFD group markedly increased adipocyte size, which was significantly reduced by KBN2202 treatment. Scale bar = 100 μm. Data shown are for female mice; corresponding male data are provided in Supplementary Figure S1. Data are presented as mean ± SD (n = 5 per group). Statistical significance was assessed by one-way ANOVA followed by Tukey’s post hoc test (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001).
Figure 2.
Effects of KBN2202 on adipose tissue, organ weight, and adipocyte hypertrophy in females (A) Absolute weights of gWAT and BAT. KBN2202 significantly reduced gWAT weight compared to the HFD group, whereas BAT weight was unchanged. (B) Absolute weight of TA muscle (left and right), showing no significant differences among groups. (C) Absolute weights of liver, heart, and kidneys were comparable among groups. (D) Representative H&E-stained images of gWAT and quantification of adipocyte area. HFD group markedly increased adipocyte size, which was significantly reduced by KBN2202 treatment. Scale bar = 100 μm. Data shown are for female mice; corresponding male data are provided in Supplementary Figure S1. Data are presented as mean ± SD (n = 5 per group). Statistical significance was assessed by one-way ANOVA followed by Tukey’s post hoc test (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001).
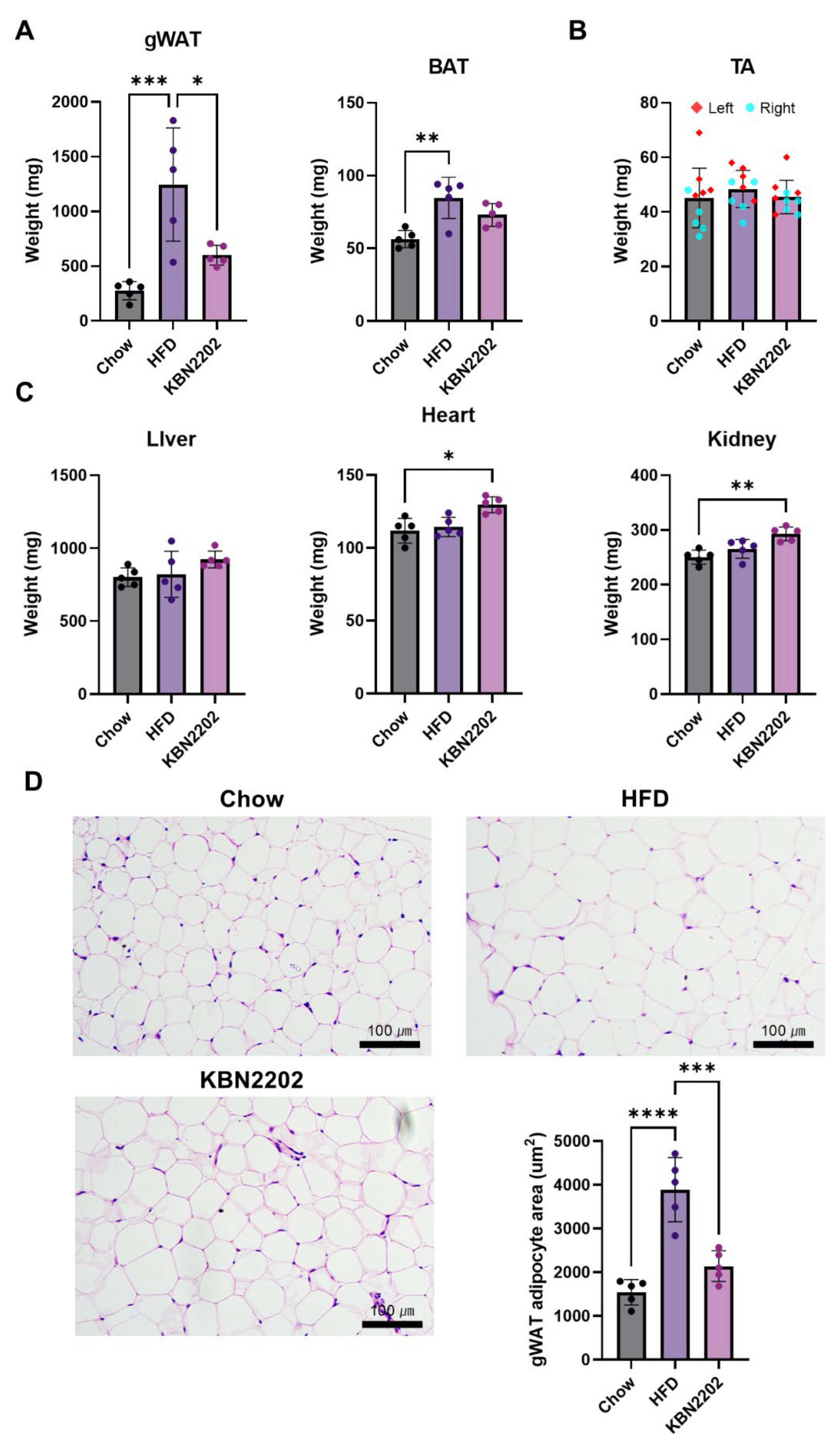
Figure 3.
Effects of KBN2202 on GLP-1, GDF15, and UCP1 expression and their correlation. (A) Serum GLP-1 levels were significantly elevated in the KBN2202-treated group compared with the HFD group (p < 0.05). (B) Serum GDF15 levels were markedly elevated in the HFD group and reduced in KBN2202-treated group. (C) Representative immunohistochemical staining of UCP1 in gWAT. Scale bar = 200 μm. (D) Quantification of UCP1 staining intensity in gWAT. KBN2202 increased UCP1 immunoreactivity by ~1.5-fold versus HFD, but this trend did not reach significance (p = 0.07). (E) Positive correlation between UCP1 intensity and serum GLP-1 levels (n = 15, r = 0.5242, p = 0.045). (F) Negative correlation between adipocyte area and UCP1 intensity (n = 15, r = –0.5599, p = 0.03). Data are shown as mean ± SD (n = 5 per group). Correlation was analyzed using pooled data from all groups (total n=15). Statistical significance was determined by one-way ANOVA followed by Tukey’s post hoc test.
Figure 3.
Effects of KBN2202 on GLP-1, GDF15, and UCP1 expression and their correlation. (A) Serum GLP-1 levels were significantly elevated in the KBN2202-treated group compared with the HFD group (p < 0.05). (B) Serum GDF15 levels were markedly elevated in the HFD group and reduced in KBN2202-treated group. (C) Representative immunohistochemical staining of UCP1 in gWAT. Scale bar = 200 μm. (D) Quantification of UCP1 staining intensity in gWAT. KBN2202 increased UCP1 immunoreactivity by ~1.5-fold versus HFD, but this trend did not reach significance (p = 0.07). (E) Positive correlation between UCP1 intensity and serum GLP-1 levels (n = 15, r = 0.5242, p = 0.045). (F) Negative correlation between adipocyte area and UCP1 intensity (n = 15, r = –0.5599, p = 0.03). Data are shown as mean ± SD (n = 5 per group). Correlation was analyzed using pooled data from all groups (total n=15). Statistical significance was determined by one-way ANOVA followed by Tukey’s post hoc test.
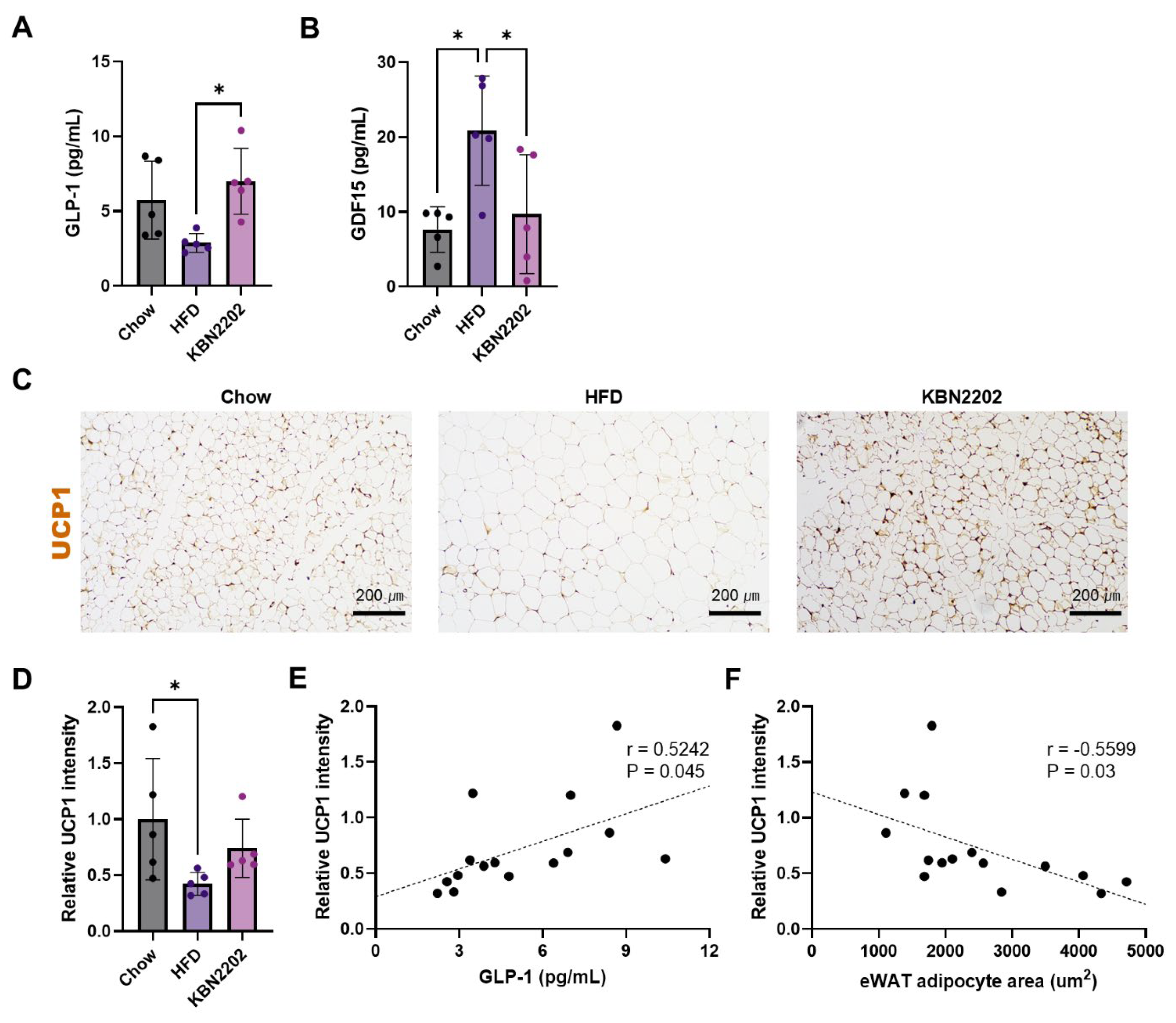
Figure 4.
Effects of KBN2202 on pro-inflammatory cytokines. Serum concentrations of (A) TNF-α, (B) MCP-1, and (C) IL-1β were quantified using a multiplex immunoassay. TNF-α levels were significantly reduced in the KBN2202 group compared with the HFD group (p < 0.01). MCP-1 and IL-1β levels did not differ significantly among groups. Data are presented as mean ± SD (n = 3–5 per group). Samples with concentrations below the assay’s limit of blank (LOB) were excluded from analysis. Statistical significance was determined by one-way ANOVA followed by Tukey’s post hoc test.
Figure 4.
Effects of KBN2202 on pro-inflammatory cytokines. Serum concentrations of (A) TNF-α, (B) MCP-1, and (C) IL-1β were quantified using a multiplex immunoassay. TNF-α levels were significantly reduced in the KBN2202 group compared with the HFD group (p < 0.01). MCP-1 and IL-1β levels did not differ significantly among groups. Data are presented as mean ± SD (n = 3–5 per group). Samples with concentrations below the assay’s limit of blank (LOB) were excluded from analysis. Statistical significance was determined by one-way ANOVA followed by Tukey’s post hoc test.
Figure 5.
Effects of KBN2202 on locomotor activity and anxiety-like behavior in the open field test. (A) Representative locomotor tracks during the final 5-minute of 10-minute test period. (B) Mean locomotor speed (cm/sec) and total distance traveled (cm). No significant differences were observed among groups in overall locomotor activity. (C) Time spent in the center and peripheral zone. KBN2202-treated mice spent significantly more time in the center zone and less time in the peripheral zone compared to the HFD group (p < 0.05) Data are presented as mean ± SD (n = 5 per group). Statistical significance was determined by one-way ANOVA followed by Tukey’s post hoc test.
Figure 5.
Effects of KBN2202 on locomotor activity and anxiety-like behavior in the open field test. (A) Representative locomotor tracks during the final 5-minute of 10-minute test period. (B) Mean locomotor speed (cm/sec) and total distance traveled (cm). No significant differences were observed among groups in overall locomotor activity. (C) Time spent in the center and peripheral zone. KBN2202-treated mice spent significantly more time in the center zone and less time in the peripheral zone compared to the HFD group (p < 0.05) Data are presented as mean ± SD (n = 5 per group). Statistical significance was determined by one-way ANOVA followed by Tukey’s post hoc test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



