
Submitted:
01 September 2025
Posted:
01 September 2025
You are already at the latest version
Abstract
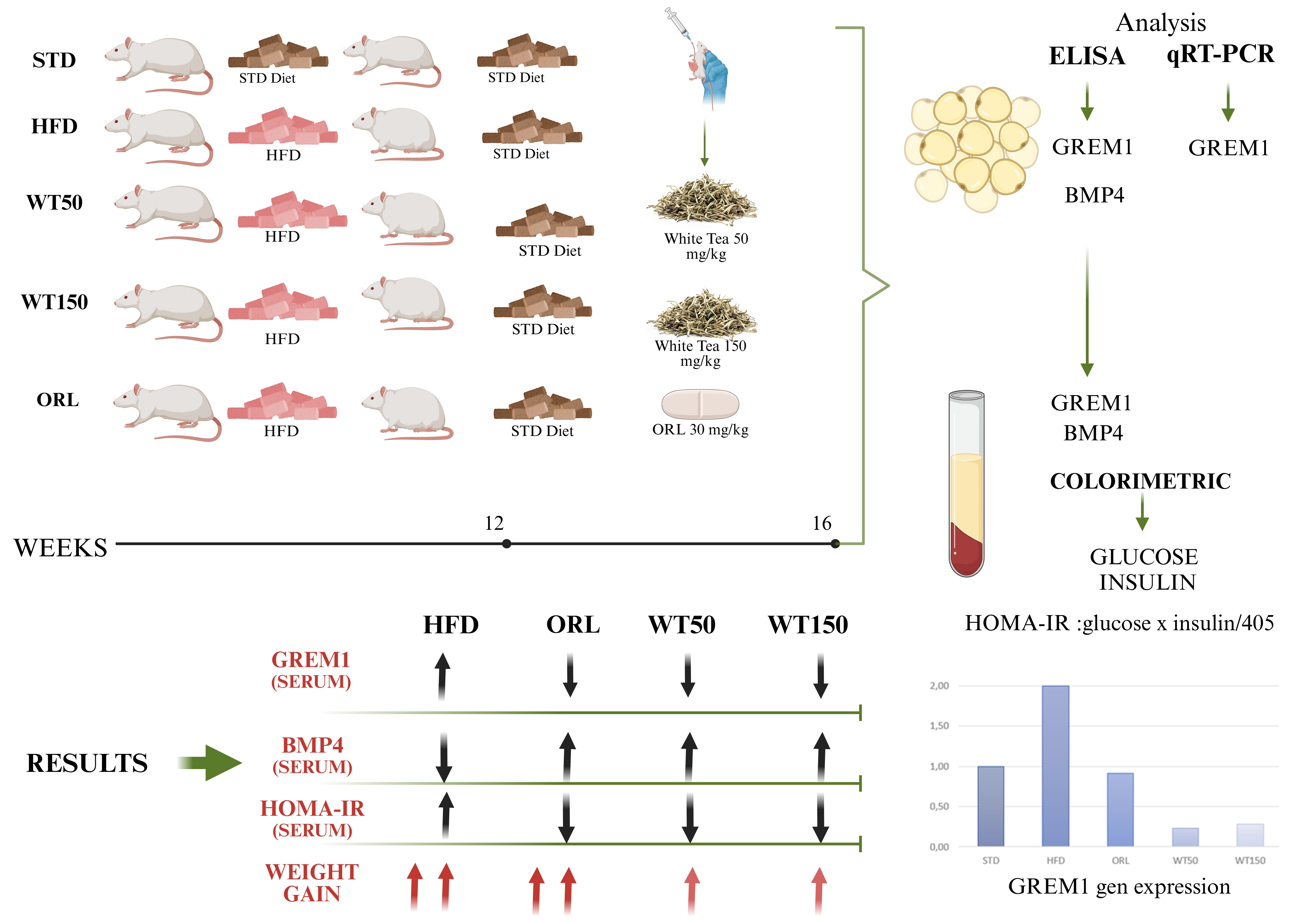
Background: Obesity is a major global health concern, being associated with insulin resistance and multiple metabolic disorders. Gremlin 1(GREM1), a bone morphogenetic protein(BMP) antagonist, is increasingly recognised as a key regulator of adipose tissue dysfunction and impaired thermogenesis in obesity. White tea has demonstrated antioxidant and anti-obesity properties in experimental models. The aim of this study was to evaluate the effects of white tea on metabolic parameters and GREM1 expression in rats made obese by a high-fat diet (HFD). Methods: A total of 40 male Sprague-Dawley rats were randomised into five groups: a standard diet group (STD); a high-fat diet group (HFD); an HFD + orlistat group (HFD + ORL); an HFD + 50 mg/kg white tea group (HFD + WT50); and an HFD + 150 mg/kg white tea group (HFD + WT150). Obesity was induced by feeding the rats a 45% high-fat diet for 3 weeks. Serum insulin, glucose and HOMA-IR levels were measured. Levels of GREM1 and BMP4 in serum and retroperitoneal adipose tissue were assessed. Results: White tea supplementation, significantly reduced weight gain and HOMA-IR compared to the HFD group. GREM1 mRNA expression in visceral adipose tissue decreased markedly in the WT50 and WT150 groups (p = 0.002 and p = 0.017, respectively). Serum GREM1 levels were significantly lower in the white tea-treated groups than in the HFD group (p = 0.011). Tissue BMP4 levels were only significantly reduced in the WT50 group (p = 0.005). There was a negative correlation between serum BMP4 levels and weight gain (rho = –0.440, p = 0.015). Conclusion: White tea has been shown to have a protective metabolic effect in obese rats, suppressing GREM1 expression and improving insulin sensitivity. White tea could therefore serve as both a dietary antioxidant and a molecular regulator of adipose tissue function in obesity management.
Keywords:
Gremlin1
; GREM1
; BMP4
; obesity
; diabetes mellitus
1. Introduction
Obesity is a growing global health problem. In recent decades, the global prevalence of obesity has increased dramatically, primarily due to increasingly sedentary lifestyles, urbanisation, and the overconsumption of high-calorie foods. Excessive accumulation of adipose tissue resulting from a chronic imbalance between energy intake and expenditure plays a central role in the development of metabolic syndrome and related disorders. It is now recognised as a major risk factor for various comorbidities, including cardiovascular diseases, insulin resistance, type 2 diabetes mellitus, obstructive sleep apnoea, osteoarthritis, certain cancers, and psychological disturbances such as depression and anxiety [1]. The economic burden of healthcare expenditure related to obesity continues to escalate worldwide, highlighting the urgent need for effective preventive and therapeutic strategies.
Adipose tissue is recognised as a highly dynamic and metabolically active organ. It is categorised into two types: white adipose tissue (WAT) and brown adipose tissue (BAT). These differ in origin, structure, function, and anatomical distribution. WAT is predominantly located in subcutaneous and visceral regions, serving as the main site for long-term energy storage in the form of triglycerides. It also secretes a variety of adipokines, cytokines and hormones that influence appetite, glucose metabolism and the secretion of inflammatory molecules. In contrast, brown adipose tissue (BAT) specialises in non-shivering thermogenesis, which is a heat-generating process driven by the expression of uncoupling protein 1 (UCP1) in the inner mitochondrial membrane. This tissue is particularly abundant in newborns but has also been identified in adult humans, primarily in the supraclavicular and perirenal regions [2]. The thermogenic capacity of BAT enables the dissipation of excess calories, making it metabolically favourable in the context of obesity and insulin resistance. Recently, a third population of thermogenic adipocytes has been identified: ‘beige’ or ‘brite’ cells. These cells can develop within white adipose tissue (WAT) depots in response to environmental stimuli such as cold exposure, exercise, and certain dietary components. This process, known as the ‘browning’ of white adipose tissue, reflects the plasticity of adipose tissue and its potential as a therapeutic target for metabolic diseases [3,4]. Therefore, identifying the molecular regulators that govern the balance between white and brown adipocyte phenotypes is crucial for developing novel anti-obesity strategies.
Tea is one of the most widely consumed beverages in the world. It is derived from the leaves of the Camellia sinensis plant, which belongs to the Theaceae family. Depending on the degree of fermentation and processing methods, tea can be broadly classified into several types, including green, black, oolong and white. White tea is considered the least processed form. It is produced by harvesting young buds and leaves at an early stage, which are then subjected to minimal withering and drying without rolling or oxidation. This gentle processing preserves its high content of bioactive compounds, particularly catechins and their derivatives, such as EGCG), epicatechin (EC) and epigallocatechin (EGC). These compounds are renowned for their potent antioxidant and anti-inflammatory properties [5]. White tea is primarily cultivated in regions with specific climatic conditions, such as China, India, Sri Lanka and Turkey. It is gaining increasing attention in the field of phytomedicine due to its rich polyphenolic profile. Studies have demonstrated that white tea exhibits a broad spectrum of biological activities, including anti-carcinogenic, cardioprotective, anti-obesity and anti-diabetic effects [6]. These health-promoting properties are largely attributed to white tea’s ability to modulate oxidative stress, improve lipid metabolism, enhance insulin sensitivity, and inhibit adipogenesis [7]. White tea retains higher levels of unoxidised catechins and flavonoids than green and black tea because it is minimally processed. This may result in superior protection against various metabolic and degenerative conditions. Studies have suggested that white tea can reduce lipid accumulation in adipocytes, downregulate pro-inflammatory cytokines, and enhance glucose uptake in insulin-resistant models. Furthermore, drinking white tea has been associated with improvements in endothelial function, lipid profile and body weight regulation [8]. Given these effects, white tea is considered a promising natural agent in the management and prevention of lifestyle-related disorders, including obesity and type 2 diabetes mellitus [9]. However, the precise molecular pathways underlying these beneficial metabolic effects are not fully understood and require further investigation.
Gremlin 1 (GREM1) is a highly conserved, secreted glycoprotein which functions as an antagonist of bone morphogenetic proteins (BMPs). It primarily targets BMP2, BMP4 and BMP7. By binding directly to these ligands, GREM1 prevents them from interacting with their receptors. This inhibits the SMAD1/5/8 signalling cascade downstream and suppresses the transcriptional activation of peroxisome proliferator-activated receptor gamma (PPARγ), a key regulator of adipocyte differentiation [10,11]. GREM1 thus plays a critical role in preserving the phenotype of white adipocytes while limiting the thermogenic browning process associated with improved metabolic flexibility and energy expenditure [12]. Among BMPs, BMP4 plays a key role in the differentiation of mesenchymal stem cells into brown adipocytes and the induction of uncoupling protein 1 (UCP1), a hallmark of mitochondrial thermogenesis [13]. Elevated GREM1 levels in obesity have been shown to impair this process, contributing to reduced energy dissipation and enhanced lipid accumulation [14]. In animal models fed a high-fat diet (HFD), GREM1 expression increases significantly in visceral adipose tissue, which is associated with impaired glucose tolerance and increased insulin resistance, even in the absence of significant weight gain [15,16]. Conversely, GREM1 inhibition, whether genetic or pharmacological, has been reported to restore insulin sensitivity, promote thermogenic gene expression, and reduce adipose tissue inflammation [17]. Further human studies support the metabolic role of GREM1. Circulating and tissue levels of GREM1 are elevated in individuals with obesity, type 2 diabetes mellitus (T2DM), and non-alcoholic fatty liver disease (NAFLD). GREM1 is also particularly enriched in visceral adipose tissue compared to subcutaneous tissue depots [15,18]. This distribution is strongly associated with increased HOMA-IR, fasting insulin and triglyceride levels, and is negatively correlated with adiponectin and insulin-stimulated glucose uptake [16]. Taken together, these findings highlight GREM1 as a contributor to metabolic dysfunction and a potential biomarker for obesity-related insulin resistance.
This study aimed to investigate the effects of white tea on obesity and insulin resistance in rats fed a high-fat diet. Specifically, we examined whether white tea could impact the Gremlin 1 (GREM1) and BMP4 signalling pathway, which is involved in regulating fat tissue function and energy balance. Two different doses of white tea were tested and compared with orlistat, a widely used anti-obesity medication. The goal was to gain a better understanding of how white tea might improve metabolic stress and inform the development of new obesity treatment strategies.
2. Materials and Methods
2.1. Experimental Animals and Study Groups
In the study, 40 male Sprague-Dawley rats weighing 150-200 grams and 6-8 weeks aged were first subjected to a 1 week acclimation period. After the adaptation period, the rats were randomly divided into five groups. The STD group acted as the control group and was fed only a chow diet. The HFD group was fed a high-fat diet. The ORL group was fed a high-fat diet and given orlistat once they had become obese. The WT50 group was fed a high-fat diet and given 50 mg/kg/day of white tea once they had become obese. The WT150 group received a high-fat diet and 150 mg/kg/day of white tea after becoming obese. All groups except the STD group were given ad libitum access to a high-fat diet to induce obesity. The composition of the high-fat diet (Arden Research & Experimental Co., Türkiye) provided 45% of total energy from fat (see Supplementary Table S1). The STD group received a chow diet (Bayramoğlu Feed, Erzurum, Türkiye) . After obesity was induced, the rats were given chow diet for three weeks. All animals were housed and fed in a sterile animal facility with a humidity of 55-60%, room temperature of 22±2°C, and a 12-hour light/dark cycle. Body temperature was maintained at 36-37°C with a rectal probe. Food and water were provided ad libitum. The rats’ weights were measured weekly, and a weight gain of at least 20% was accepted as a criterion for obesity [19]. During this period, they were administered 30 mg/kg orlistat and two different doses of white tea (50 mg/kg and 150 mg/kg) via oral gavage. At the end of the experiment, the rats were sacrificed after 12 hours of fasting. Anaesthesia was induced by intraperitoneal injection of 50 mg/kg ketamine hydrochloride (Ketalar, Pfizer Ltd., Istanbul, Turkey) and 10 mg/kg xylazine hydrochloride (Rompun, Bayer, Whippany, NJ). Blood samples were collected from the aorta and transferred to additive-free serum tubes. Retroperitoneal adipose tissue samples were collected and stored at -80°C for further study. All animals were treated according to the protocols for the care and use of laboratory animals under the guidelines of the National and International Research Council.
2.2. Preparation of White Tea Samples
White tea samples were obtained from the manufacturer, the General Directorate of Tea Enterprises in Rize, Turkey, and prepared by daily brewing. The leaves were harvested during the first flush only, in May, in line with ÇAYKUR’s standard practice. White tea is not harvested during later flushes. The preparation and dose selection of white tea were based on previous studies [20]. Before administration, the samples were cooled to room temperature and delivered to the subjects by oral gavage. Experimental groups received 1 mL/day and 5 mg/kg/day doses of the substances at the same time daily. Subjects in the other groups received 1 mL/day of water by gavage.
2.3. Preparation of Blood and Tissue Specimens
Serum samples were collected by centrifugation of blood collected from the subjects for 15 minutes at 1500xg at 2-8℃ after waiting for clotting. The collected sera were stored in a -80°C freezer until the day the samples were examined. For homogenization of the obtained retroperitoneal adipose tissue samples, 1 mL homogenization buffer was prepared with 20 mM sodium phosphate + 140 mM potassium chloride at pH 7.4. 1 mL of homogenization buffer was added to 0.1 g of tissue and homogenized [21]. After homogenization, centrifugation was performed at 800xg for 10 minutes at 2-8℃.
2.4. Analysis of Samples
Gremlin 1 (Cat: SG-21730; SinoGeneClon Biotech Co., Ltd., China), BMP 4 (Cat: SG-20681; SinoGeneClon Biotech Co., Ltd., China) and insulin (Cat: SG-20161; SinoGeneClon Biotech Co., Ltd., China) levels were analyzed by ELISA method. The HOMA-IR index was calculated using the formula glucose x insulin/405. Total RNA was isolated using a kit (High Pure RNA Isolation Kit, Roche, Germany) and its concentration was determined using µdrop plates on Thermo Multiskan Go (Thermo Fisher Scientific, USA). The High-Capacity cDNA Synthesis Kit was used to generating cDNA (Applied Biosystems, USA) from total RNA (1000 ng RNA/20 µl reaction). We performed qRT-PCR in a 96-well optical plate using LightCycler 480 Probes Master in a Roche Lightcycler 480 II, diluting the master mix with nuclease-free water. 30 ng of cDNA was used in each qRT-PCR reaction (20 µl) contained in 1 µl. The following probes were purchased from Thermo; Grem1 (Cat: 4448489) and GAPDH (Cat: 4448489). GAPDH gene expression was used as a reference for normalizing the results.
2.5. Statistical Analyses
Statistical analyses were performed using IBM SPSS Statistics, v23.0 (SPSS Inc., Chicago, USA) and Microsoft Office Excel. GREM1 expression was evaluated by fold change. Within-group distributions were reported as frequencies (n, %). The Kruskal-Wallis test with Bonferroni correction was used to assess differences between groups for continuous numerical variables. Spearman correlation analysis was performed to evaluate the relationship between data, and significant rho values were determined. Data are presented as median and minimum-maximum (min-max). p<0.05 was considered significant.
3. Results
3.1. HPLC Content and Total Polyphenol/Flavonoid Capacity of White Tea
The phenolic composition of white tea was characterized by Hüner Yiğit M et al. using HPLC-DAD analysis, and the quantitative results are provided in Table 1 [22]. Representative chromatograms of the standards are displayed in Supplementary Figure S1. Among the catechins, epigallocatechin (EGC) and epigallocatechin gallate (EGCG) were identified as the most abundant compounds in the white tea extract.
3.2. Weight gain and HOMA-IR index
A metabolic evaluation revealed significant differences between the groups in terms of both the weight gain and HOMA-IR index. In terms of weight gain, the WT50 and WT150 groups gained significantly less weight than the HFD group, with the lowest weight gain observed in the WT150 group (p = 0.001). The ORL group showed a wide range in weight gain (330 g (290–349)), comparable to the HFD group. The HFD group exhibited the highest HOMA-IR values (median: 12.5 (10.6–16.6)), indicating pronounced insulin resistance. By contrast, the ORL group showed significantly lower HOMA-IR levels (median: 9.6 (6.2–13)) (p = 0.007), as did the WT50 group (median: 9.35 (9.1–10.7)). The WT150 group had slightly lower values than the HFD group, but these did not reach statistical significance (median: 11.2 (9.9–12.4)) (Figure 1A,B; respectively)).
3.3. GREM1 Expression in Visceral Adipose Tissue
In the quantitative real-time PCR analysis, the expression of GREM1 in visceral adipose tissue was evaluated as a fold change relative to the non-obese control group. As illustrated in Figure 2, GREM1 expression was approximately twofold higher in the high-fat diet (HFD) group than in the control group. In contrast, expression levels in the WT50 and WT150 groups were significantly lower than in the HFD group (p = 0.002 and p = 0.017, respectively). While the ORL group did not show a statistically significant difference, its expression level was lower than that of the control group. Furthermore, the WT50 group exhibited significantly lower expression than the ORL group (p = 0.037).
GREM1 expression was evaluated by qRT-PCR and is presented as fold change relative to the STD group, which was normalized to 1.00. A 2.00-fold increase was observed in the HFD group. The ORL group showed a value close to baseline (0.92-fold). In contrast, the white tea–treated groups demonstrated marked reductions, with 0.23-fold in the WT50 group and 0.29-fold in the WT150 group.
3.4. Serum and Tissue Protein Levels of GREM1 and BMP-4.
Analysis of serum and tissue biomarkers revealed notable group-specific variations. Serum Gremlin-1 (sGREM1) concentrations were significantly higher in the HFD group (median: 8.6 ng/mL, range: 7.0–9.9) than in the treatment groups. Notably, the WT50 and WT150 groups exhibited markedly lower sGREM1 levels than the HFD group, with statistical significance (p = 0.011). The ORL also showed lower sGREM1 levels (7.4 ng/mL (6.8–8.8)), though this reduction was not statistically significant when compared with the HFD group (Figure 3A). There was no significant difference in serum BMP-4 (sBMP4) levels between groups (p = 0.611), with median values ranging from 426 pg/mL in the HFD group to 504 pg/mL in the WT50 group. This suggests that circulating BMP-4 concentrations were not substantially affected by the interventions (Figure 3B). Tissue levels of 1 (tGREM1) and BMP-4 (tBMP4) exhibited distinct profiles. Although tGREM1 concentrations were higher in the HFD (101.5 ng/g, 83.9–161.7) and ORL (108.4 ng/g, 60.3–127.0) groups than in the WT50 (87.3 ng/g, 85.4–106.8) and WT150 (98.0 ng/g, 50.9–116.5) groups, these differences were not statistically significant (p = 0.383) (Figure 3C). Notably, tissue BMP-4 levels were significantly lower only in the WT50 group (2.45 ng/g, 2.03–2.69) than in the HFD group (3.9 ng/g, 3.12–5.3), with p = 0.005, indicating a tissue-specific regulatory effect of low-dose white tea. No other significant differences in tBMP4 were observed between the remaining groups (Figure 3D).
3.5. Correlation Analysis of GREM1 and BMP4
Correlation analysis revealed a significant negative association between serum BMP-4 levels and weight gain (rho = –0.440, p = 0.015). A moderate positive correlation was observed between sGREM1 and tGREM1 levels (rho = 0.387, p = 0.035). Additionally, sGREM1 levels were positively correlated with tBMP4 levels (rho = 0.560, p = 0.001).
4. Discussion
The experimental data from our study confirm the role of GREM1 as a central regulator in the pathophysiology of obesity, as reported in the literature. The data also show that the metabolic improvement effects of white tea administration can be explained by changes in GREM1 levels. These findings suggest that white tea can modulate adipose tissue-specific molecular responses, with GREM1 potentially acting as a mediator or target structure in this process. Increased GREM1 gene expression in visceral adipose tissue, alongside the highest serum GREM1 levels observed in the HFD group, suggests that GREM1 is activated both locally and systemically in obesity-related pathophysiological processes. This corroborates the previously reported role of GREM1 in adipose tissue dysfunction and metabolic disorders [23]. The significant increase in insulin resistance in the same group suggests that this increase in GREM1 levels may be associated with metabolic disorders.
White tea intervention significantly reversed the disturbance to this molecular axis, particularly at low doses. Significant decreases in GREM1 gene expression and serum levels were observed in the white tea-treated group, alongside a notable reduction in weight gain and insulin resistance. These results imply that white tea could restrict adipocyte hypertrophy and enhance systemic insulin sensitivity by inhibiting GREM1 production. It has indeed been demonstrated in the literature that white tea catechins, particularly EGCG, modulate Wnt and BMP signalling pathways, reducing oxidative stress and inflammation [24,25]. In this context, it can be suggested that the metabolic improvement observed in our study was due to white tea suppressing GREM1-BMP4 antagonism. Given the multifaceted effects of GREM1 on regulating adipogenesis, angiogenesis, and immune response processes, the effects of white tea on this protein are not limited to lipid metabolism but also extend to the structural and functional components of the adipose tissue microenvironment. In particular, the antagonistic effect of GREM1 on BMP4 may inhibit the maturation of adipocyte progenitors and the development of healthy adipose tissue, which may in turn pathologise the expansion of adipose tissue and trigger adverse processes such as hypertrophy, hypoxia and inflammation [26]. Depletion of GREM1 by white tea may have broken this chain and promoted more balanced adipocyte differentiation and tissue remodelling. However, GREM1 is produced not only by adipocytes, but also by fibroblasts, endothelial cells, and immune cells in the stromal vascular fraction of adipose tissue [27,28]. Consequently, the white tea intervention may have exerted a far-reaching regulatory effect on all cellular components of adipose tissue, including not only fat cells. The effects of GREM1 on these cellular subunits are associated with tissue-level fibrosis, extracellular matrix (ECM) remodelling, and local immune responses. Suppressing this axis contributes to maintaining adipose tissue function.
Administering white tea at a dose of 50 mg/kg significantly reversed this pathological process. White tea administration suppressed GREM1 expression and serum levels, resulting in a significant reduction in weight gain and insulin resistance. Thus, white tea provided not only symptomatic improvement, but also played a direct regulatory role in the GREM1-BMP4 axis by targeting obesity-related molecular dysregulation. Previous studies have reported that white tea polyphenols, particularly the EGCG compound, have regulatory effects on the Wnt/BMP signalling pathways and reduce oxidative stress and inflammation. Our study is the first to demonstrate that these effects are exerted through GREM1 in a concrete model, making it possible to evaluate white tea not only for its antioxidant properties, but also as an active biological agent that can modulate gene and protein expression in adipose tissue. EGCG, one of the major catechins found in white tea, has been described as a potent biological agent capable of modulating multiple signalling pathways at the cellular level [29]. While the anti-adipogenic, anti-inflammatory and antioxidant effects of EGCG in obesity models are well documented, our study provides new evidence that these effects may be mediated through the GREM1-BMP4 axis. Previous studies have shown that EGCG suppresses the differentiation of preadipocytes by inhibiting the Wnt/β-catenin and MAPK/ERK pathways and reducing lipogenesis through AMPK activation. These pathways are also key regulatory elements affecting GREM1 and BMP4 expression [30]. While BMP4 acts as a critical morphogen for physiological adipogenesis, GREM1 is a protein that directly antagonises this pathway. Therefore, EGCG’s effect on enhancing BMP4 signalling or promoting GREM1 inhibition may play a decisive role in adipogenesis and adipose tissue plasticity. In our study, the significant decrease in BMP4 levels observed in the WT50 group following white tea administration suggests that this effect may be related not only to GREM1 suppression, but also to a rebalancing effect at the systemic level. Furthermore, EGCG has been reported to affect histone acetylation and DNA methylation at an epigenetic level, thereby permanently altering the expression profiles of adipogenesis-related genes [31]. These findings suggest that white tea may support long-term as well as temporary metabolic remodelling.
In conclusion, this study reaffirms the importance of GREM1 in the pathogenesis of obesity and provides some of the first evidence that bioactive agents of natural origin, such as white tea, could act as therapeutic tools in this context. Given that white tea and its EGCG have been shown to regulate adipogenesis, insulin sensitivity and inflammation via GREM1, these components could be incorporated into preventive and complementary treatment strategies in the future. These findings suggest that functional foods should be considered as both dietary supplements and biological agents that can modulate molecular targets. Further investigating the effects of GREM1 in different tissues, dose-dependent tea interventions and signalling mechanisms specific to this axis may enable the development of new generation bioactive intervention strategies for obesity in future studies.
This study has some limitations. Firstly, the use of an animal model limits the direct generalisation of the findings to humans. Firstly, using an animal model restricts the direct generalisation of the findings to humans. Secondly, the duration of white tea administration was kept short, and the long-term effects were not evaluated. Furthermore, the bioavailability of EGCG and other polyphenols in white tea was not measured, and important adipokines and inflammatory markers other than GREM1 were not analysed. The lack of histological evaluation of adipose tissue meant that the tissue could not reflect the molecular findings. Using only male rats also meant that sex-related effects were neglected. For these reasons, future studies should be more extensive and multifaceted in terms of both duration and scope.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
BŞ and MA* collaborated to conceptualize the study and develop the methodology. MA conducted the analysis, HK overseeing supervision and drafted the manuscript.
Funding
This study was supported by the Development Institute of Recep Tayyip Erdoğan University.
Ethics Approval and Consent to Participate
All animal experimental procedures were performed in accordance with relevant guidelines and regulations. The study was approved by Recep Tayyip Erdoğan University (RTEU) Animal Experiments Local Ethics Committee with decision number 2020/29. This study did not involve human participants; therefore, informed consent and consent for publication are not applicable.
Data Availability Statement
All data supporting the findings of this study are included in the article and its supplementary materials.
Acknowledgements
None.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Ghaben AL, Scherer PE. Adipogenesis and metabolic health. Nature reviews Molecular cell biology. 2019;20(4):242-58.
- Coelho M, Oliveira T, Fernandes R. State of the art paper Biochemistry of adipose tissue: an endocrine organ. Archives of medical science. 2013;9(2):191-200.
- Kahn CR, Wang G, Lee KY. Altered adipose tissue and adipocyte function in the pathogenesis of metabolic syndrome. The Journal of clinical investigation. 2019;129(10):3990-4000.
- Park A, Kim WK, Bae K-H. Distinction of white, beige and brown adipocytes derived from mesenchymal stem cells. World journal of stem cells. 2014;6(1):33.
- Vishnoi H, Bodla RB, Kant R, Bodla R. Green tea (Camellia sinensis) and its antioxidant property: A review. International Journal of Pharmaceutical Sciences and Research. 2018;9(5):1723-36.
- Sanlier N, Atik İ, Atik A. A minireview of effects of white tea consumption on diseases. Trends in Food Science & Technology. 2018;82:82-8.
- Xu R, Yang K, Li S, Dai M, Chen G. Effect of green tea consumption on blood lipids: a systematic review and meta-analysis of randomized controlled trials. Nutr J. 2020 May 20;19(1):48. PMID: 32434539; PMCID: PMC7240975. [CrossRef]
- Huner Yigit M, Atak M, Yigit E, Topal Suzan Z, Kivrak M, Uydu HA. White Tea Reduces Dyslipidemia, Inflammation, and Oxidative Stress in the Aortic Arch in a Model of Atherosclerosis Induced by Atherogenic Diet in ApoE Knockout Mice. Pharmaceuticals (Basel). 2024 Dec 17;17(12):1699. PMID: 39770544; PMCID: PMC11679696. [CrossRef]
- Abe SK, Inoue M. Green tea and cancer and cardiometabolic diseases: a review of the current epidemiological evidence. Eur J Clin Nutr. 2021 Jun;75(6):865-876. Epub 2020 Aug 20. PMID: 32820240; PMCID: PMC8189915. [CrossRef]
- Kišonaitė M, Wang X, Hyvönen M. Structure of Gremlin-1 and analysis of its interaction with BMP-2. Biochem J. 2016 Jun 1;473(11):1593-604. Epub 2016 Apr 1. PMID: 27036124; PMCID: PMC4888461. [CrossRef]
- Gustafson B, Hammarstedt A, Hedjazifar S, Hoffmann JM, Svensson PA, Grimsby J, Rondinone C, Smith U. BMP4 and BMP Antagonists Regulate Human White and Beige Adipogenesis. Diabetes. 2015 May;64(5):1670-81. Epub 2015 Jan 20. PMID: 25605802. [CrossRef]
- Qian SW, Tang Y, Li X, Liu Y, Zhang YY, Huang HY, Xue RD, Yu HY, Guo L, Gao HD, Liu Y, Sun X, Li YM, Jia WP, Tang QQ. BMP4-mediated brown fat-like changes in white adipose tissue alter glucose and energy homeostasis. Proc Natl Acad Sci U S A. 2013 Feb 26;110(9):E798-807. Epub 2013 Feb 6. PMID: 23388637; PMCID: PMC3587258. [CrossRef]
- Elsen M, Raschke S, Tennagels N, Schwahn U, Jelenik T, Roden M, Romacho T, Eckel J. BMP4 and BMP7 induce the white-to-brown transition of primary human adipose stem cells. Am J Physiol Cell Physiol. 2014 Mar 1;306(5):C431-40. Epub 2013 Nov 27. PMID: 24284793. [CrossRef]
- Hoffmann JM, Grünberg JR, Church C, Elias I, Palsdottir V, Jansson JO, Bosch F, Hammarstedt A, Hedjazifar S, Smith U. BMP4 Gene Therapy in Mature Mice Reduces BAT Activation but Protects from Obesity by Browning Subcutaneous Adipose Tissue. Cell Rep. 2017 Aug 1;20(5):1038-1049. PMID: 28768190. [CrossRef]
- Hammarstedt A, Gogg S, Hedjazifar S, Nerstedt A, Smith U. Impaired Adipogenesis and Dysfunctional Adipose Tissue in Human Hypertrophic Obesity. Physiol Rev. 2018 Oct 1;98(4):1911-1941. PMID: 30067159. [CrossRef]
- Hedjazifar S, Khatib Shahidi R, Hammarstedt A, Bonnet L, Church C, Boucher J, Blüher M, Smith U. The Novel Adipokine Gremlin 1 Antagonizes Insulin Action and Is Increased in Type 2 Diabetes and NAFLD/NASH. Diabetes. 2020 Mar;69(3):331-341. Epub 2019 Dec 27. PMID: 31882566. [CrossRef]
- Park A, Kim WK, Bae KH. Distinction of white, beige and brown adipocytes derived from mesenchymal stem cells. World J Stem Cells. 2014 Jan 26;6(1):33-42. PMID: 24567786; PMCID: PMC3927012. [CrossRef]
- Coelho M, Oliveira T, Fernandes R. Biochemistry of adipose tissue: an endocrine organ. Arch Med Sci. 2013 Apr 20;9(2):191-200. Epub 2013 Feb 10. PMID: 23671428; PMCID: PMC3648822. [CrossRef]
- Hariri N, Thibault L. High-fat diet-induced obesity in animal models. Nutrition research reviews. 2010;23(2):270-99.
- Yılmaz HK, Türker M, Kutlu EY, Mercantepe T, Pınarbaş E, Tümkaya L, Atak M. Investigation of the effects of white tea on liver fibrosis: An experimental animal model. Food Sci Nutr. 2024 Jan 30;12(4):2998-3006. PMID: 38628196; PMCID: PMC11016422. [CrossRef]
- Hariri N, Thibault L. High-fat diet-induced obesity in animal models. Nutrition research reviews. 2010;23(2):270-99.
- Huner Yigit, M. Huner Yigit, M., Atak, M., Yigit, E., Topal Suzan, Z., Kivrak, M., & Uydu, H. A. (2024). White Tea Reduces Dyslipidemia, Inflammation, and Oxidative Stress in the Aortic Arch in a Model of Atherosclerosis Induced by Atherogenic Diet in ApoE Knockout Mice. Pharmaceuticals, 17(12), 1699.
- Grillo E, Ravelli C, Colleluori G, D’Agostino F, Domenichini M, Giordano A, Mitola S. Role of gremlin-1 in the pathophysiology of the adipose tissues. Cytokine Growth Factor Rev. 2023 Feb;69:51-60. Epub 2022 Sep 14. PMID: 36155165. [CrossRef]
- Dashwood WM, Orner GA, Dashwood RH. Inhibition of beta-catenin/Tcf activity by white tea, green tea, and epigallocatechin-3-gallate (EGCG): minor contribution of H(2)O(2) at physiologically relevant EGCG concentrations. Biochem Biophys Res Commun. 2002 Aug 23;296(3):584-8. PMID: 12176021. [CrossRef]
- Teiten MH, Gaascht F, Dicato M, Diederich M. Targeting the wingless signaling pathway with natural compounds as chemopreventive or chemotherapeutic agents. Curr Pharm Biotechnol. 2012 Jan;13(1):245-54. PMID: 21466435. [CrossRef]
- Hoffmann JM, Grünberg JR, Church C, Elias I, Palsdottir V, Jansson JO, Bosch F, Hammarstedt A, Hedjazifar S, Smith U. BMP4 Gene Therapy in Mature Mice Reduces BAT Activation but Protects from Obesity by Browning Subcutaneous Adipose Tissue. Cell Rep. 2017 Aug 1;20(5):1038-1049. PMID: 28768190. [CrossRef]
- Ivet Elias, Sylvie Franckhauser, Tura Ferré, Laia Vilà, Sabrina Tafuro, Sergio Muñoz, Carles Roca, David Ramos, Anna Pujol, Efren Riu, Jesús Ruberte, Fatima Bosch; Adipose Tissue Overexpression of Vascular Endothelial Growth Factor Protects Against Diet-Induced Obesity and Insulin Resistance. Diabetes 1 July 2012; 61 (7): 1801–1813.
- Kapoor VN, Müller S, Keerthivasan S, Brown M, Chalouni C, Storm EE, Castiglioni A, Lane R, Nitschke M, Dominguez CX, Astarita JL, Krishnamurty AT, Carbone CB, Senbabaoglu Y, Wang AW, Wu X, Cremasco V, Roose-Girma M, Tam L, Doerr J, Chen MZ, Lee WP, Modrusan Z, Yang YA, Bourgon R, Sandoval W, Shaw AS, de Sauvage FJ, Mellman I, Moussion C, Turley SJ. Gremlin 1+ fibroblastic niche maintains dendritic cell homeostasis in lymphoid tissues. Nat Immunol. 2021 May;22(5):571-585. Epub 2021 Apr 26. PMID: 33903764. [CrossRef]
- Liu X, Zhou F, Wen M, Jiang S, Long P, Ke JP, Han Z, Zhu M, Zhou Y, Zhang L. LC-MS and GC-MS based metabolomics analysis revealed the impact of tea trichomes on the chemical and flavor characteristics of white tea. Food Res Int. 2024 Sep;191:114740. Epub 2024 Jul 8. PMID: 39059930. [CrossRef]
- Moqaddasi HR, Singh A, Mukherjee S, Rezai F, Gupta A, Srivastava S, Sridhar SB, Ahmad I, Dwivedi VD, Kumar S. Influencing hair regrowth with EGCG by targeting glycogen synthase kinase-3β activity: a molecular dynamics study. J Recept Signal Transduct Res. 2025 Apr;45(2):95-106. Epub 2025 Feb 18. PMID: 39964119. [CrossRef]
- Prasanth MI, Sivamaruthi BS, Cheong CSY, Verma K, Tencomnao T, Brimson JM, Prasansuklab A. Role of Epigenetic Modulation in Neurodegenerative Diseases: Implications of Phytochemical Interventions. Antioxidants (Basel). 2024 May 15;13(5):606. PMID: 38790711; PMCID: PMC11118909. [CrossRef]
Figure 1.
Weight gain (A) and HOMA-IR (B) in groups.
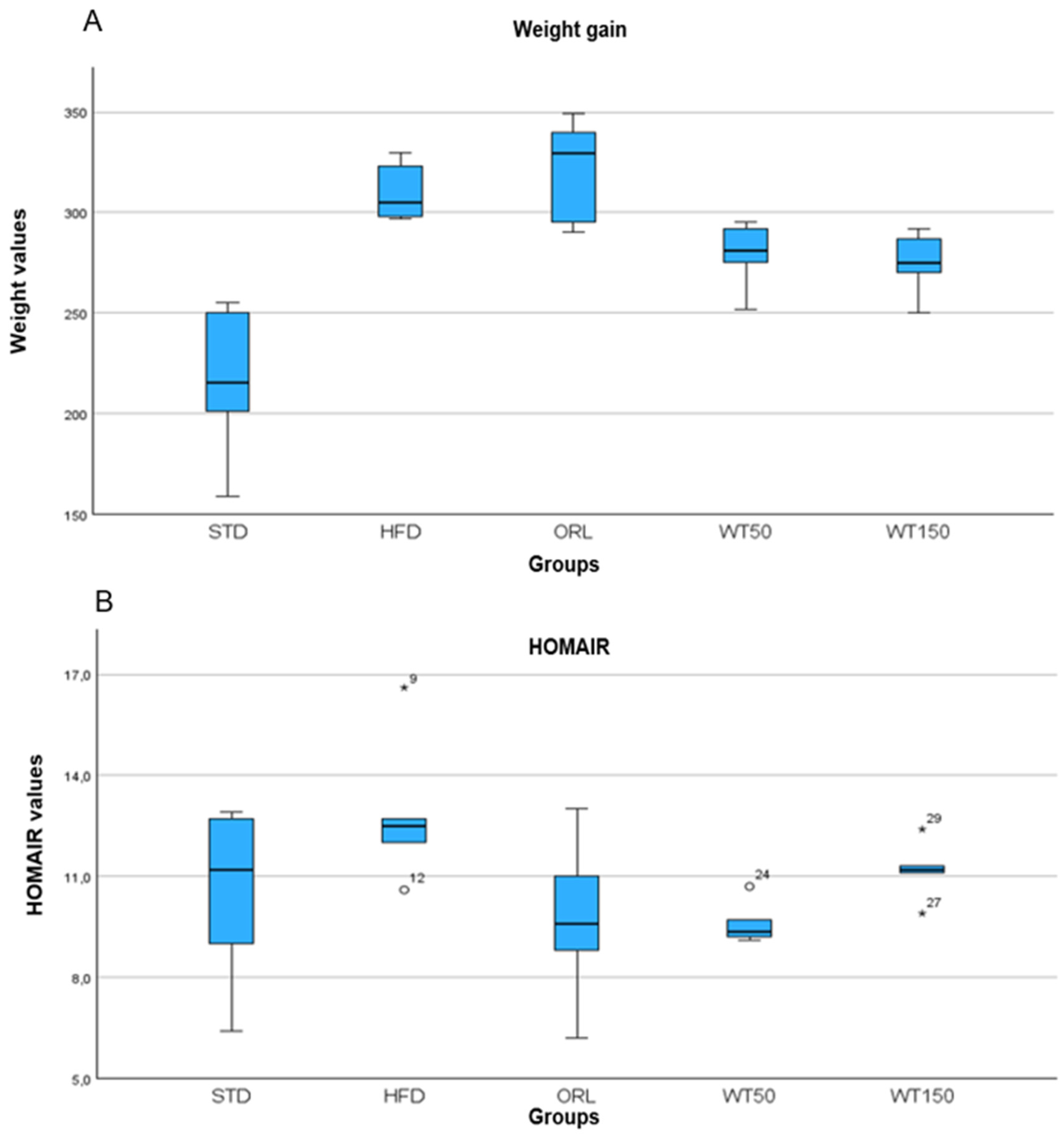
Figure 2.
GREM1 expression levels in visceral adipose tissue.
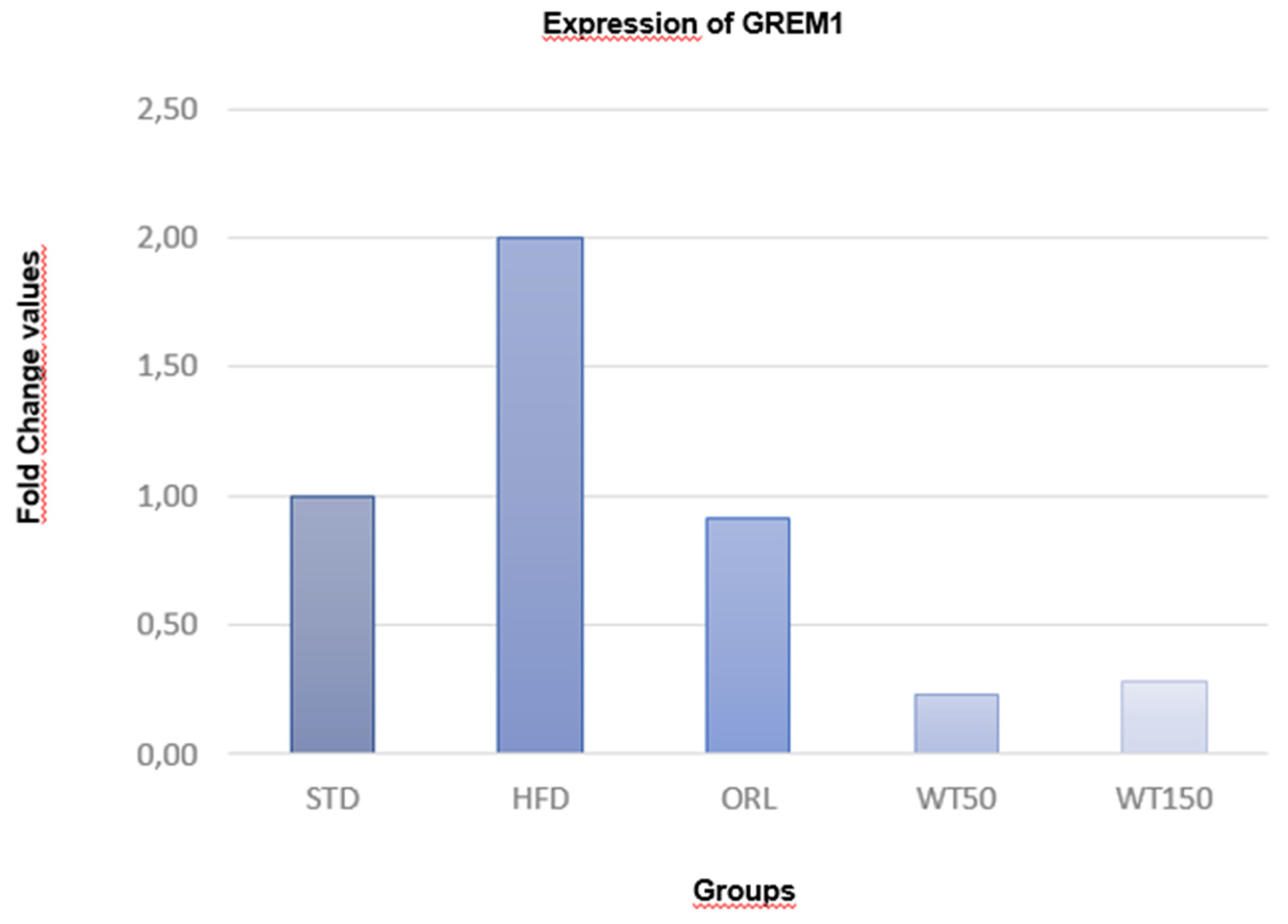
Figure 3.
Serum and tissue levels of GREM1 and BMP4 across experimental groups.Box plots show sGREM1, sBMP4, tGREM1 and tBMP4 concentrations in rats from the STD, HFD, ORL, and white tea–treated groups.
Figure 3.
Serum and tissue levels of GREM1 and BMP4 across experimental groups.Box plots show sGREM1, sBMP4, tGREM1 and tBMP4 concentrations in rats from the STD, HFD, ORL, and white tea–treated groups.
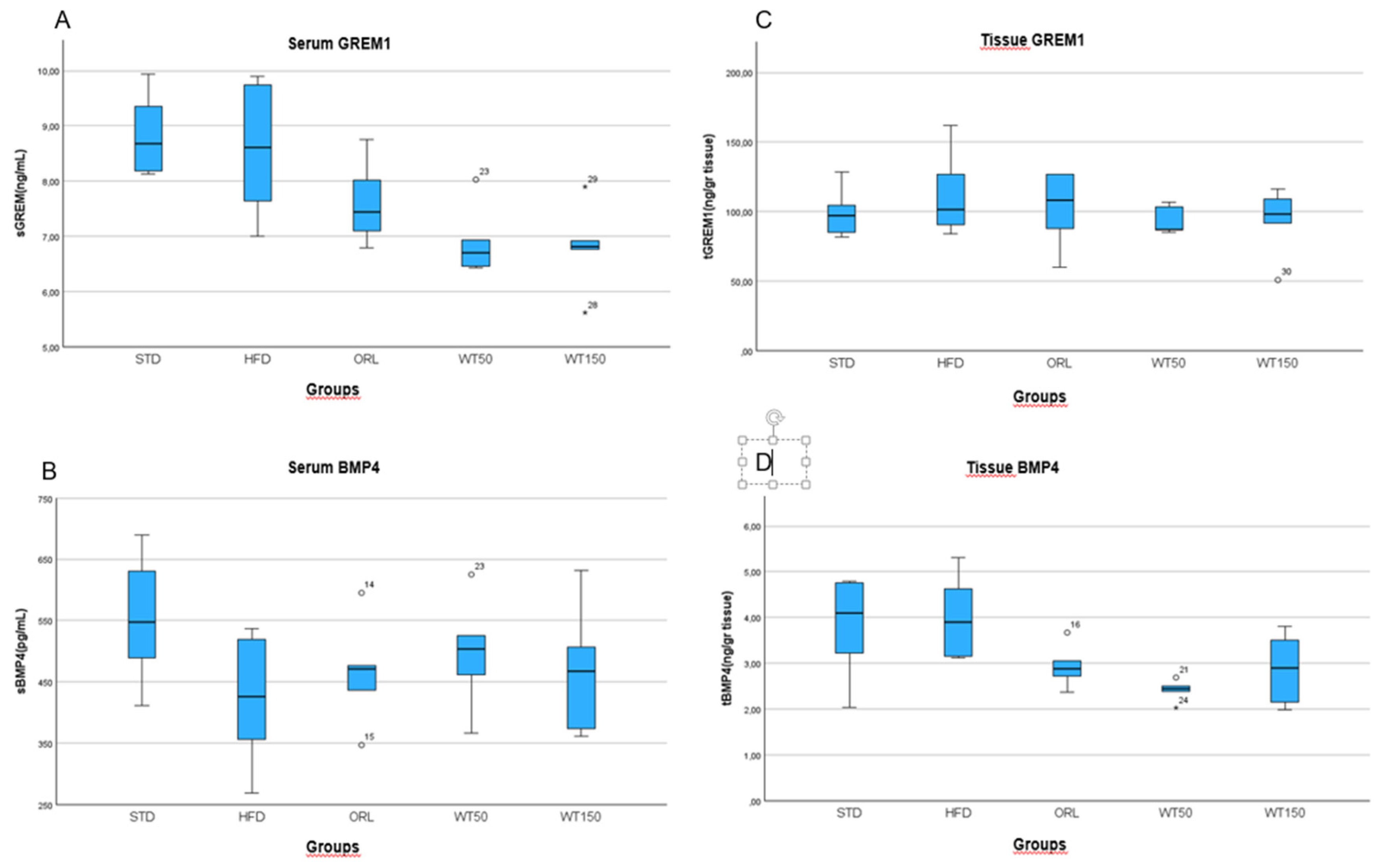

Table 1.
HPLC-DAD analysis of phenolic constituents of white tea.
| Number | Compound Name |
Retention Time (Min) |
Concentration (n = 2, µg/g) |
| 1 | Gallic acid | 3.202 | 5.55 ± 0.61 |
| 2 | Epigallocatechin | 4.499 | 261.4 ± 3.59 |
| 3 | Catechin | 5.488 | - |
| 4 | Caffeine | 7.750 | 101.51 ± 2.61 |
| 5 | Epigallocatechin 3-gallate | 8.678 | 119.8 ± 2.31 |
| 6 | Epicatechin | 9.800 | - |
| 7 | Epicatechin 3-gallate | 15.916 | 46.36 ± 1.70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



