
Submitted:
21 November 2025
Posted:
24 November 2025
You are already at the latest version
Abstract
Zinc homeostasis is fundamental to metabolic health, orchestrated by the coordinated actions of two major zinc transporter families: ZIP (Zrt- and Irt-like Proteins) and ZnT (Zinc Transporters). ZIP-transporters Facilitates zinc influx into the cytosol from the extracellular space or from the lumen of intracellular organelles, whereas ZnT-transporters control zinc efflux from the cytosol to the extracellular space or facilitate its sequestration into intracellular vesicles and organelles, concurrently harboring the meticulous intracellular zinc homeostasis. This equilibrium is essential for all critical functions like cellular response, metabolic control, or immune pathway alteration. Disruption of this homeostasis is a driver of different pathological alterations like metabolic inflammation, a chronic low-grade inflammatory state underlying obesity, type 2 diabetes, and nonalcoholic fatty liver disease. Recent studies revealed that ZIP and ZnT transporters dynamically regulate metabolic and inflammatory cues, with their tissue-specific expression varying by tissue and acclimating to different physiological and pathological conditions. Recent advanced research in molecular and genetic understanding has helped to deepen our knowledge on the interplay of activity between ZIP and ZnT transporters and their crosstalk in metabolic tissues, underscoring the potential therapeutic prospect for restoring zinc balance and ameliorating metabolic inflammation. This review provides a comprehensive overview that covers the function, regulation, and the interactive crosstalk of ZIP and ZnT zinc transporters in metabolic tissues, and its pathological condition.
Keywords:
Zinc transporters
; ZIP family
; ZnT family
; metabolic inflammation
; obesity
; diabetes
; immune response
; zinc homeostasis
1. Introduction
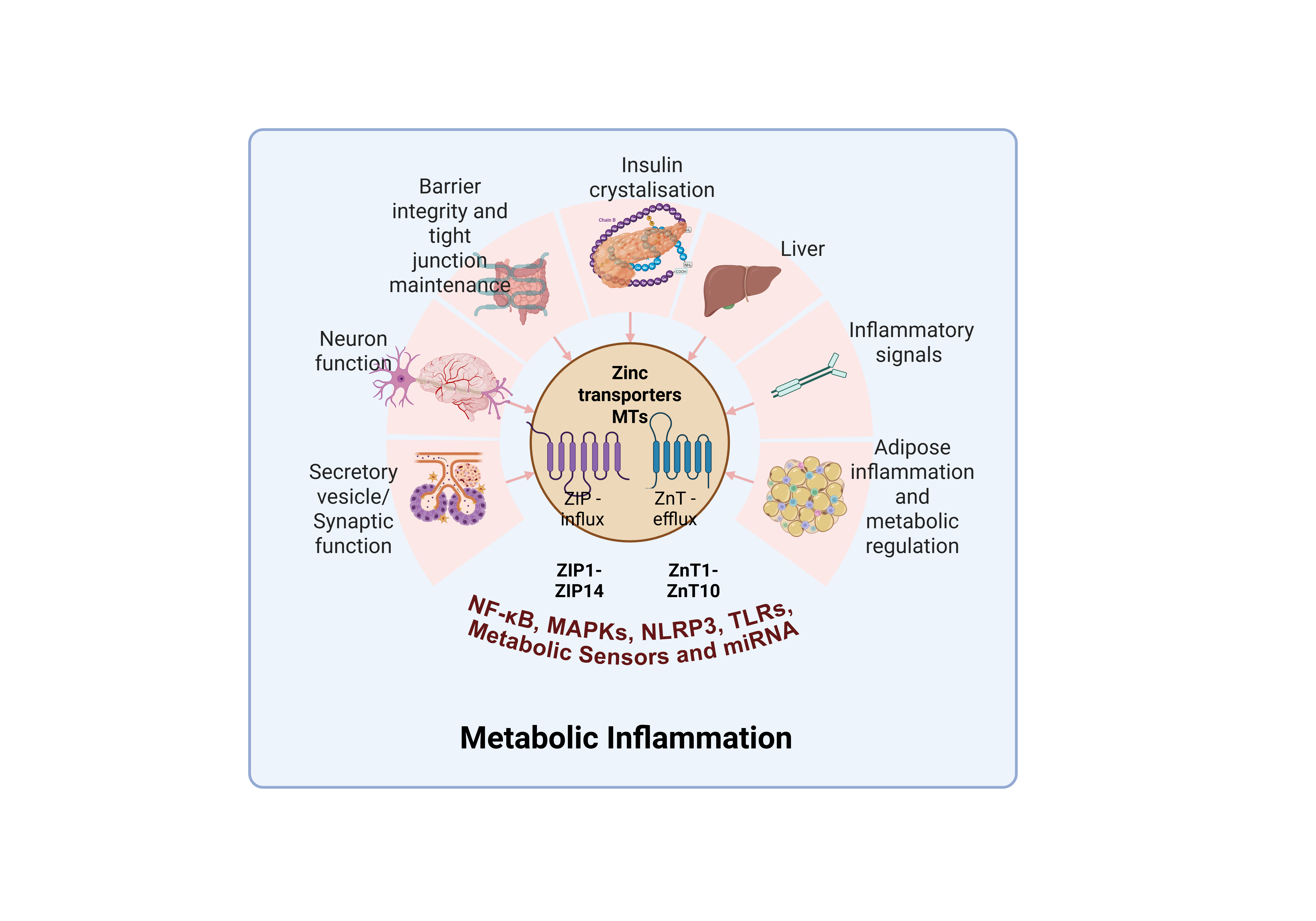
Zinc is an essential trace element intricately interwoven into the foundation of human physiology. It protects cellular homeostasis, controls gene transcription, supports metabolic reactions, and intervenes intercellular communication and many more. Computational researches indicate that approximately one-tenth of the human proteome possesses zinc-binding capability, while over three percent of human genes encode zinc finger–containing proteins, underscoring the profound functional significance of zinc in regulating diverse biological systems [1]. A tightly organized orchestration of membrane-bound transport mechanisms which controls zinc's intracellular equilibrium, essential for its biological activity. Two primary families of zinc transporters are chiefly responsible for maintaining zinc homeostasis. The SLC30 family (ZnT), which facilitates zinc efflux from the cytoplasm to the extracellular space or into intracellular compartments. On the other hand, the SLC39 family (ZIP), which mediates zinc influx into the cytoplasm from either extracellular sources or intracellular organelles [1]. Complementing these transporters, another indispensable protein, metallothioneins (MTs) a cysteine-rich, zinc-binding proteins that function as intracellular zinc reservoirs and also bind to excess free zinc ions to avoid cytotoxicity [2].
Research have identified an intricate association between zinc equilibrium and metabolic inflammation, a phenomenon also known as metaflammation which is a chronic, low-grade inflammatory state arising from metabolic disorders rather than classical infectious triggers. Metaflammation is a defining feature of several prevalent metabolic diseases, including type 2 diabetes (T2D), nonalcoholic fatty liver disease (NAFLD), obesity and cardiovascular disease. Persistent metabolic stress, especially developed from dietary habits which are rich in saturated fats and refined carbohydrates, initiates inflammatory cascades in different metabolic organs. These insults result in the secretion of pro-inflammatory cytokines like interleukin-1β (IL-1β) and tumor necrosis factor-alpha (TNF-α), which subsequently interfere with insulin signaling, impair glucose metabolism, and promote metabolic dysfunction [3].
Emerging research evidence indicates that zinc transporter dysfunction plays a crucial role in the pathophysiology of metaflammation. To maintain the intracellular zinc pool, several zinc transporters exhibit tissue-specific critical roles that significantly contribute to immune and metabolic regulation. In light of this, ZIP14 is increased in hepatocytes during inflammation and affects insulin sensitivity. In pancreatic cells, ZnT8 plays the crucial function for insulin formation, secretion and its malfunction leads to decreased insulin production or β-cell stress [4,5]. Meanwhile, ZIP4 & 8 and ZnT2 are involved in immune cell activation and gut epithelial barrier function, respectively both of which are pivotal for maintaining systemic inflammatory balance [6,7].
Importantly, besides maintaining the zinc flux zinc transporters are active participants in immune and metabolic signaling pathways. Transcriptional regulation of zinc transporter genes involves key inflammatory transcription factors such as nuclear factor-kappa B (NF-κB) and metal-regulatory transcription factor 1 (MTF-1) [8]. Furthermore, the innate immune system components including Toll-like receptors (TLRs) and the NLRP3 (NOD-like receptor family, pyrin domain-containing 3) inflammasome are known to modulate the expression of various transporters in response to metabolic stressors. Recent insights have further identified zinc as a second messenger in intracellular signaling and get influenced by this stimuli [9].
In this review, we endeavored to consolidate existing information about the unique and interrelated roles of ZIP and ZnT transporters in metabolic inflammation. We tried to overview their molecular processes, tissue-specific functions, and connections with immune-metabolic signaling cascades. We also accentuated their potential as therapeutic targets in metabolic disorders and inflammation by elucidating the zinc-dependent regulatory webs that govern these functions. Overall, we wanted to highlight the translational prospect of zinc signaling ordinance in reforming metabolic health and mitigating chronic inflammation.
2. Key Machinery for Zinc Homeostasis
Maintaining tightly regulated cellular and tissue zinc concentrations is essential for the function of hundreds of enzymes, transcription factors, and signaling molecules. Zinc homeostasis requires the coordinated action of membrane-bound transporter families, metallothioneins, and organelle-specific trafficking mechanisms enabling dynamic control over import, export, and intracellular zinc distribution.
2.1. ZnT and ZIP Families: the Core Zinc Transporters
The ZIP (Zrt/Irt-like protein) transporter family (1-14 members) is essential for preserving inter-cellular zinc status by facilitating zinc transfer from the extracellular milieu to the cytoplasm or from intracellular organelle reserves. The transport system is crucial for maintaining zinc homeostasis within the cells or whole system, particularly in zinc deficiency or cellular stress to ensure adequate zinc supply for supporting the variety of biochemical processes that are essential for cellular function [10].
On the other hand, the ZnT (zinc transporter) transporter family (ZnT1-ZnT10) is responsible for facilitating the efflux of zinc from the cytoplasm, either into the extracellular space or into intracellular organelles such as the Golgi apparatus, vesicles, and mitochondria. These transporters are essential for maintaining cellular zinc homeostasis by excess zinc induced cytotoxicity and ensuring proper compartmentalization of zinc pools, critical for specific signaling pathways within the cell [11].
The "yin-yang" regulation mechanism of these two families maintains cellular zinc homeostasis in opposition. ZIP transporters deliver cytosolic zinc for cellular processes, whereas ZnT transporters export excess zinc to ensure optimal storage for optimal cellular performance [12].
2.2. Metallothionein: A Crucial Intracellular Zinc Buffer
Metallothioneins (MTs), a group of small, cysteine-rich proteins, function as the crucial intracellular zinc buffers and dynamic reservoir, which help in tightly regulating the concentration of free/labile zinc ions [Zn2+] in the picomolar (pM) to low nanomolar (nM) range [13]. This dynamic binding allows MTs to buffer both steady-state and transient levels of free zinc ions (which are typically in the picomolar range), thereby controlling zinc availability for a wide range of biological processes, including enzyme activity, gene expression, boosting immunity and fighting against oxidative stress via zinc-finger transcription factors. Considering the complex role of metallothioneins, especially the partly zinc-depleted variant, MT2, it functions not only as a zinc buffer but also dynamically regulates cytosolic zinc concentrations by both donating and accepting zinc ions. to maintain zinc pool, releasing it when required for enzymatic or structural activities inside the cell [13,14]. Moreover, MTs are essential for eliminating surplus zinc and so evading the cytotoxic consequences of zinc aggregation. Of note, MTs are involved in redox regulation, as their zinc-thiolate clusters are highly sensitive to oxidative and reductive changes, allowing them to modulate cellular redox status and mitigate oxidative stress as well as cellular defense [6,15]. The MTF-1 meticulously controls MTs synthesis. MTF-1 recognizes an increase in free zinc and activates MTs to control the surplus zinc [16].
The synchronized and antagonistic function of ZIP and ZnT transporters, in conjunction with zinc-binding protein- MTs, is crucial for maintaining the cytosolic free zinc content within a confined, low range (pM to low nM values). By shuttling zinc across designated membranes, with precise orchestrated mechanism they maintain the intracellular zinc gradients essential for finest cellular signaling and general physiological function.
2.3. Key Cellular Zinc Compartments and Organelle-Specific Zinc Trafficking.
Zinc has a markedly heterogeneous distribution across different cellular compartments, each of which maintains specific zinc concentrations targeted for certain enzymatic, structural, or signaling roles. The complementary functions of ZnT and ZIP transporters coordinate zinc flow across organellar membranes to dynamically modulate subcellular zinc distribution. Precise regulation of the zinc pool is critical for basic cell survival, redox equilibrium, and intracellular signaling.
Endoplasmic Reticulum (ER): In the endoplasmic reticulum, zinc transporters such as ZnT5 and ZnT7 play a critical role in facilitating zinc influx into the ER lumen where this transported zinc is essential for the proper folding and structural maturation of nascent proteins, including several crucial zinc-requiring ectoenzymes like Phosphatidylinositol-glycan anchor biosynthesis (PIG) proteins, Sphingomyelin phosphodiesterase 1 (SMPD1), Chaperones etc [17,18]. Zinc present in the ER promotes the activity of certain folding enzymes and chaperones, and it is crucial for managing unfolded protein response (UPR) pathways for cellular stress adaptation, hence aiding cellular responses to metabolic or oxidative stressors [19].
Golgi Apparatus: Zinc is crucial in the Golgi apparatus as a cofactor for specific enzymes involved in protein post-translational modification and transport. ERp44, an early secretory pathway chaperone, binds Zn2+ to regulate client binding and release for protein flow and homeostasis. In return for protons, ZnT4, ZnT5/ZnT6, and ZnT7 membrane transporter complexes import Zn2+ into the Golgi lumen. Amagai research group in their study showed that systematic ZnT-knockdowns exhibited ZnT4, ZnT5/ZnT6, and ZnT7 control labile Zn2+ content in the distal, middle, and proximal Golgi, respectively, consistent with their local ZnT-mediated Zn2+ fluxes regulate ERp44 localization, trafficking, and client-retrieval activity, as shown by time-course imaging of cells undergoing synchronized secretory protein traffic and functional experiments [20]. Additionally, key zinc-dependent processes and enzymes include Golgi alpha-mannosidase II (GMII or MAN2A1/MAN2A2), Beta-4-galactosyltransferase, Alkaline phosphatases (ALPs) which mostly depends on ZnT4-7 to maintain zinc homeostasis within the Golgi lumen [21,22,23]. Zinc supports these enzymatic activities related to glycosylation, protein packaging, and vesicle trafficking. Disruption in these transporters particularly ZnT4 mutations has been shown to impair secretory pathway efficiency and compromise protein maturation, potentially leading to broader cellular dysfunction in secretory tissues.
Mitochondria: Mitochondrial zinc homeostasis, albeit less well-characterized, is increasingly acknowledged as vital regulatory axis. Recent research emphasizes the importance of precisely regulated mitochondrial zinc homeostasis as both deficiency and excess can affect redox homeostasis, increase oxidative stress, and cause metabolic inflammation and mitochondrial dysfunction by targeting specific electron transport chain (ETC) complexes [24]. Several ongoing research have shown interest to define the involvement of transporters in mitochondrial zinc transportation, highlighting a promising novel approach for targeted therapies in different mitochondria-dependent pathological condition, including metabolic disorders and cancer.
Vesicles and Insulin Granules: Vesicles accumulate high concentrations of zinc within their lumen through specific transporters of the ZnT (SLC30) family, such as ZnT2, ZnT3 and ZnT8, which pump zinc from the cytosol into secretory and synaptic vesicles. As per cryo-EM and immunolocalization studiesZnT transporters are required for vesicular zinc storage and controlled zinc secretion in neuronal, endocrine, and epithelial cells [25]. ZnT2 (SLC30A2) is a vesicular zinc transporter that directs zinc into lysosomes and secretory vesicles, especially in mammary epithelial cells, enabling zinc secretion into breast milk [26]. The ZnT2 mutation degrades this process, which inturn leads to transient newborn zinc deficit (TNZD) in breastfed babies due to low milk zinc levels [27]. ZnT8 transports zinc into insulin granules in pancreatic β-cells, facilitating insulin crystallization and further maturation and secretion. Furthermore, zinc-mediated crystallization is crucial for maintaining proper glucose homeostasis in the body, and dysfunction of this transporter contributes to β-cell stress and T2D [28].
Lysosomes and Endosomes: Zinc trafficking within lysosomes and endosomes is systematically regulated by the ZnT and ZIP family transporters, the TRPML1 ion channel, and lysosomal exocytosis. ZnT2 and ZnT4 localize to the lysosomal membrane and mediate zinc import into the lumen for storage or detoxification, as documented by recent experimental knockdown (TRPML1KD) and colocalization studies. TRPML1 (MCOLN1) serves as a key channel for zinc efflux from lysosomal stores to the cytoplasm, with loss-of-function (LOF) causing pathological zinc accumulation and lysosomal enlargement, a phenotype seen in mucolipidosis type IV, a rare genetic disorder. The function of these transporters is modulated by lysosomal acidification (via V-ATPase) and transcription factor, MTF-1, which upregulate the expression of ZnT2 and ZnT4 and its in response to excess zinc [29]. ZIP transporters like ZIP8 and ZIP13 can also regulate lysosomal-cytosolic zinc exchange and immune signaling [30]. Proper zinc balance in these organelles is crucial for protein degradation, autophagy, immune activation, and receptor recycling, and dysregulation is implicated in immune disorders, metabolic disease, and lysosomal storage pathologies.
This intricate and compartmentalized zinc trafficking system ensures that each organelle maintains optimal zinc concentrations for its specialized functions. Alterations in the zinc concentration within the organelles, either due to mutations in transporter genes, inflammation or metabolite disturbances, can compromise the integrity and proper function of respective organelles. This may leads to disruption in fundamental operations including autophagy, protein folding, and immune signaling, which ultimately direct to cellular dysfunction and tailored pathological condition. Thus, precise regulation of zinc distribution within organelles is a critical mechanism underpinning cellular physiology and pathology, going beyond basic maintenance to serve as a foundation for organelle and cell health.

Table 1.
Zinc Homeostasis Components.
| Organelle | Transporters Involved | Function & Physiological Significance | Key Experimental Study Details & Insights | Reference |
|---|---|---|---|---|
| Cytosol | ZIP family (influx), ZnT family (efflux into organelles/extracellular space) | Maintains low free Zn²⁺ (picomolar) to support signaling and prevent toxicity; main hub for zinc-sensitive enzymes and kinases. | ZIP1/ZIP3 found in intracellular organelles (HEK293, mouse), localize dynamically based on zinc status .Cytosolic zinc wave observed in mast cells, dependent on ZIP/ZnT activity. | [31] |
| Secretory Vesicles(e.g., synaptic, insulin granules) | ZnT3, ZnT8, ZnT2, ZnT4 | Accumulate high vesicular zinc (millimolar range); ZnT3 loads synaptic vesicles (neurons); ZnT8 loads insulin granules (pancreatic β cells); ZnT2 regulates glandular vesicles; ZnT4 traffics vesicles in secretory tissues. | ZnT8 knockout and variant studies confirm granule-specific insulin packaging/diabetes risk. ZnT3 is involved in heterogeneous synaptic vesicle assembly and neurotransmission. ZnT2 is critical for zinc vesicle formation and stress protection. | [32,33,34,35,36,37,38] |
| Golgi Apparatus & ER | ZnT5, ZnT6, ZnT7 (influx into lumen); ZIP7, ZIP13 (efflux into cytosol) | Zinc is required for folding and activation of secreted/membrane proteins, e.g., tissue-nonspecific alkaline phosphatase (TNAP), ERp44; ZIP exports zinc in "zinc wave" for cytosolic signaling; ZnT5/6/7 localized to Golgi & ER, essential for ALP activation. | ZnT5 variant B localizes to ER, colocalizes with ZIP7, forming zinc efflux pathway; ZIP7 essential for cytosolic zinc signaling. ZnT7 localizes to proximal Golgi and regulates ERp44-dependent homeostasis. ZnT5/6/7 activate tissue-nonspecific alkaline phosphatase in two-step mechanism. | [17,20,21,39,40,41] |
| Mitochondria | ZnT2 (suggested), ZnT9 (SLC30A9), ZIP family (potential roles) | Mitochondrial zinc pools regulate oxidative metabolism, apoptosis, mitophagy; zinc influx/efflux affects mitochondrial stress resilience and cytochrome c release. | SLC30A9 (ZnT9) loss causes zinc mishandling and mitochondrial overload in HeLa cells, shown by live dye tracking and ERC coevolution analysis. Zn-induced mitochondrial swelling triggers mPTP opening, mediates apoptosis. | [24,42,43,44,45] |
| Lysosome-Related Organelles (LROs) | CDF-2 (ZnT family), ZIPT-2.3 (ZIP family, C. elegans), ZnT4 | Dynamic zinc storage and release; maintain organellar zinc pools for protein degradation/homeostasis, critical in stress adaptation. | In C. elegans, CDF-2 stores zinc in LROs during excess, ZIPT-2.3 releases zinc during deficiency; co-regulation and colocalization confirmed by super-resolution microscopy and transgenics. Morphologic changes reflect zinc status, transporter levels. | [46] |
| Nucleus | Metallothioneins (MTs), potential transporters | Zinc primarily bound to transcription factors (zinc fingers), essential for gene expression/DNA replication; labile pool controls TF binding/dynamics. | Single-molecule microscopy reveals that zinc availability modulates DNA binding of zinc finger TFs (MTF-1, CTCF, GR) in live mammalian cells; zinc depletion shortens TF dwell time .MTs buffer nuclear zinc and protect against oxidative injury . | [47,48] |
3. Functional Links Between Zinc and Metaflammation
Metaflammation is classified as a persistent, low-grade systemic low-grade inflammation that originates in metabolic tissues in response to factors like overnutrition and sedentary lifestyles rather than infections by microorganisms. This condition is distinct from acute inflammation, which is transitory, subsides upon rejuvenation. Metaflammation, which affects metabolically active tissues such as adipose tissue, liver, pancreas, and skeletal muscle, stays constant nevertheless is less intense [49].
The hallmark of metaflammation is the sustained production of pro-inflammatory cytokines, such as IL-1β, TNF-α, and interleukin-6 (IL-6).Metaflammation is intricately associated with metabolic overload, a condition resulting from uncontrolled intake of food components which triggers a complex and crucial crosstalk between the systemic immune and metabolic pathways. The core mechanism involves specific immune cells that reside within metabolic tissues, most notably tissue-resident macrophages and other components of the innate immune system [50]. These interactions can lead to insulin resistance, glucose intolerance, and disturbances in lipid metabolism, thereby contributing to the development of chronic health conditions such as type 2 diabetes (T2D), nonalcoholic fatty liver disease (NAFLD), atherosclerosis, and hypertension associated with obesity. The key molecular mechanisms underlying metaflammation include persistent signaling through pattern recognition receptors (PRRs), notably TLRs, activation of the NLRP3 inflammasome, mitochondrial dysfunction, ER stress etc [49].
The choice of dietary intake plays a major impact in establishing and aggravating metaflammation. With great example, Western diets high in saturated fatty acids (SFAs) and processed carbs fuel metabolic organ inflammatory signals [51,52,53]. Lipotoxicity, ER stress, and NLRP3 inflammasome activation arise from saturated fatty acid stimulation of TLR4. This cascade increases IL-1β production and promotes pro-inflammatory macrophage polarization in adipose tissue [54,55]. Importantly, overconsuming glucose and fructose facilitates the production of advanced glycation end-products (AGEs), which worsen liver inflammation and lipid metabolism via the receptor for AGEs (RAGE) signaling pathway [56,57,58]. The chronic state of overnutrition also drives the production of ROS due to mitochondrial overload and the activity of NADPH oxidase [59,60]. This redox can displace zinc from specific zinc-binding proteins like cysteine thiols, MTs, which can further interfare in signalings dependant to cytosolic zinc status [61]. Thus, metabolic tissues can convert into immunological-active organs, addressing the close relationship between food signaling and immune responses in metaflammation.
3.1. The Influence of Zinc Transporter Imbalance in Metaflammation
Zinc plays a dual role in modulating metaflammation. An imbalance in zinc equilibrium, either through deficiency or mislocalization within the cell’s compartments, disrupts immune homeostasis and increases both oxidative stress and inflammatory signaling pathways [62,63]. Importantly, zinc deprived condition influences in cytokine production, affecting in a pro-inflammatory profile activation, primarily featured with increased levels of TNF-α, IL-1β, and IL-6, coupled with a decline in the anti-inflammatory cytokine such as IL-10 [64]. This imbalance fosters the polarization of macrophages towards the M1 phenotype, impairs T-cell growth, and limits the efficacy of regulatory T-cells, consequently intensifying insulin resistance [65,66].
The expression of zinc transporters is tightly regulated by inflammatory signals. For instance, in liver, ZIP14 is upregulated by IL-6. This upregulation leads to enhanced zinc accumulation in the liver and subsequent changes in how zinc is distributed throughout the body, ultimately affecting insulin sensitivity [67]. Dysfunctional ZnT8 can impair the crystallization and secretion of insulin in pancreatic β-cells, while ZIP8 influences NF-κB signaling in immune cells. Disruptions in the function or expression of specific zinc transporters are indeed linked to impaired metabolic function and sustained inflammatory responses involving several mechanisms. Specific levels of zinc are also critical in modulating inflammatory responses within the system, primarily by inhibiting the NF-κB signaling pathway [68,69]. This inhibition occurs through the blockage of IκB kinase activity, a key enzyme responsible for the phosphorylation and subsequent degradation of IκB proteins. Under normal, zinc-sufficient conditions, NF-κB is held in an inactive state in the cytoplasm by IκB proteins. In response to inflammatory stimuli (like infection or cytokines), IκB is phosphorylated by the IκB kinase (IKK) complex, ubiquitinated, and degraded by the proteasome. This degradation allows NF-κB to translocate into the nucleus and activate pro-inflammatory gene expression [69]. In contrast, zinc deficiency has been shown to have detrimental effects on inflammation regulation. A lack of adequate zinc levels leads to the enhanced activation of the NLRP3 inflammasome, a multi-protein complex that plays a pivotal role in the innate immune response which results in increased secretion of IL-1β, a potent pro-inflammatory cytokine that drives inflammatory processes [70].
The increased activity of the NLRP3 inflammasome and the resultant overproduction of IL-1β lead to a condition of metaflammation in zinc deficiency. Thus, zinc deprivation results in intensification of inflammatory pathways and promotes the advancement of metabolic illnesses highlighting the need of sustaining optimal zinc levels for immune system health and inflammatory management.
3.2. Zinc Transporters and microRNA-Mediated Regulation of Metaflammation
Zinc transporters controll the biogenesis and expression patterns of numerous microRNAs (miRNAs) involved in immune and metabolic processes, which are also regulated by precisely controlled intracellular zinc concentrations. Zinc-modulated miRNAs serve as key post-transcriptional influencer inchronic inflammation by regulating cytokine synthesis, immune cell differentiation, and metabolic adaptability. Recent studies demonstrated that transporters such as ZIP14 modulate the expression of zinc-sensitive miRNAs such as miR-675 and others within intestinal and hepatic tissues [71]. In enterocytes ZIP14 deficiency has been exhibited to impact intracellular zinc balance, resulting in modified miRNA profiles which in turn influence the transcriptional regulation of inflammatory pathways, including NF-κB and STAT3 signaling, thereby fostering a pro-inflammatory condition and undermining gut barrier integrity. Moreover, aberrant zinc homeostasis resulting from transporter dysfunction in different metabolic tissue has been shown to regulate the expression of miRNAs by “zinc–miRNA axis” linked to insulin resistance and lipid metabolism, further contributing to metaflammation [72]. Another study has shown that zinc deficiency upregulate miR-21 and promotes inflammation in adipose tissue by targeting the tumor suppressor PDCD4 whereas, zinc repletion can normalize miR-21 levels and attenuate these inflammatory effects [72]. Similarly, miR-34a is increased in states of zinc deficiency and obesity, exacerbating insulin resistance and driving pro-inflammatory M1 macrophage polarization through suppression of SIRT1 and Klf4, as confirmed in both murine and human adipose tissue models.
In obesity and NAFLD, miR-122 influences systemic cholesterol and lipid metabolism by regulating key enzyme expressions and signaling pathways. Studies show that miR-122 generally acts to promote hepatic lipogenesis and cholesterol synthesis in a healthy liver, and its dysregulation in disease states has complex and sometimes conflicting effects. Different studies evidenced a complex, dose-dependent relationship between zinc status and miRNA expression, specifically involving miR-122 and miR-144-3p in metabolic tissues [73]. Experimental alteration in zinc status have been shown to influence miR-122 expression and associated lipid profiles. Alternatively, excessive zinc exposure upregulated miR-144-3p, which has been exhibited to impairs the Nrf2 antioxidant defense pathway and exacerbates oxidative stress and insulin resistance, as demonstrated in in vitro hepatocyte and adipocyte models. Evidence together supports the hypothesis that zinc transporter-mediated miRNA regulation may constitute a basic molecular axis connecting zinc homeostasis to the regulation of inflammatory, metabolic, and immunological signaling pathways.
Table 2.
Zinc-Related Impacts of Pathogenic Triggers on Inflammatory Outcomes.
| Pathogenic Trigger | Zinc-Related Impact | Inflammatory Outcome / Mechanism | Mechanistic/Clinical Notes | |
|---|---|---|---|---|
| SFAs, ROS, glucose overload | Disrupt zinc transporter expression: ZIP14 ↑ (in hepatocytes, adipocytes), ZnT8 ↓ (in pancreatic β-cells) | NLRP3 inflammasome activation, increased IL-1β/IL-18 release, chronic metabolic inflammation | ZIP14 upregulation in response to TLR4 activation and IL-6 drives hepatic/adipose zinc accumulation and insulin resistance. ZnT8 downregulation impairs insulin granule formation and β-cell function. High glucose and ROS amplify IL-1β/IL-18 via NLRP3 . | [54,67,68] |
| Zinc deficiency | Reduced Treg cell numbers, increased NF-κB activation; impaired metallothionein buffering | Chronic low-grade inflammation, heightened NLRP3 activation, increased cytokine output | Zn deficiency leads to lysosomal stress, ROS generation, and NLRP3 inflammasome activation/secretion of IL-1β .Zinc supplementation inhibits NLRP3 and supports immune balance. | [59,70,74] |
| Zinc transporter dysfunction | Alters zinc distribution in pancreas (ZnT8), liver/adipose (ZIP14), gut (ZnT2/ZIP8) | Insulin resistance, gut barrier leakiness, cytokine imbalance | Genetic or acquired dysfunction in ZnT8/ZIP14 impairs insulin packaging/secretion and hepatic/adipose zinc homeostasis. ZnT2/ZIP8 regulate intestinal barrier integrity; dysfunction increases permeability and systemic inflammation. | [67,68,75,76] |
| Oxidative stress | Displaces zinc from protein binding sites, impairs antioxidant function | Amplifies ROS, triggers NLRP3 activation, further immune cell recruitment and cytokine release | Oxidative stress displaces zinc, activates stress kinases, and amplifies proinflammatory signaling. Zinc repletion reduces ROS and NLRP3 activity, supporting antioxidant defenses. | [59,77] |
4. Tissue-Specific Roles of Zinc Transporters
Zinc transporters exhibit specific, non-redundant roles depending on the tissue and the subcellular compartment where they are expressed, allowing for fine-tuned control of zinc homeostasis for particular physiological functions. The following discussion delves into the evidence highlighting the functions of key zinc transporters in specific organs, particularly their implications for metaflammation and metabolic disorders.
Liver- The Role of ZIP and ZnT Transporters in inflammation and insulin resistance: The liver serves as a pivotal regulator of metabolism and acts as a primary site for the redistribution of zinc during inflammatory responses. Among the several zinc transporters, ZIP14 is a pivotal zinc importer, with its expression markedly increased in hepatocytes during both acute and chronic inflammation. This induction is mostly initiated by pro-inflammatory cytokines, including IL-6 and TNF-α [78]. The overexpression of ZIP14 enhances the inflow of zinc into hepatocytes, which is closely associated with the acute phase response and marked by a fast decrease in blood zinc levels while hepatic zinc reserves simultaneously increase. ZIP14 located in the cytosol and the mitochondrial of hepatocytes, where it conjunctively work with ZIP8 to maintain zinc balance within mitochondria [79]. Controlling pathways like glycolysis, gluconeogenesis, glycogenesis, and glycogenolysis helps liver cells conserve energy and adapt to insulin and glucose fluctuations. ZIP14 regulates insulin signaling pathways via affecting PTP1B and c-Met phosphorylation [78]. The ZIP14KO animals demonstrated hepatic insulin resistance, glucose intolerance, and lipid metabolic issues. These mice also exhibited increased body fat and altered expression of glucose and lipid metabolism genes such GLUT2, SREBP-1c, and FASN [80]. ZIP4/14 and ZnT9 depletion or dysregulation impairs the liver's response to inflammatory stimuli and regeneration signals, increasing the likelihood of NAFLD and metabolic syndrome [81]. ZnT transporters export zinc from hepatocytes or store it intracellularly to lower cytosolic zinc levels. ZnT8 is an interesting exception to the steady expression of most ZnT transporters during inflammation. Studies have shown that ZnT8 deficiency in ZnT8 KO mice can protect from the liver injury by acetaminophen (APAP)-induced oxidative stress and damage [82]. This protection is linked to an increase in hepatic zinc and MT levels, which enhances the liver's antioxidant defense system, and it is also associated with reduced liver cell death and inflammation. This protective strategy involves compensatory ZIP4 and ZIP14 overexpression, which increases hepatic zinc absorption, antioxidant protein production, and hepatocyte proliferation [81,82]. Overall, ZIP14 and ZnT8 work in concert to control the metabolic pathways and response to inflammation in the liver for proper maintenance of zinc homeostasis. Understanding these mechanisms is essential for elucidating the pathways that contribute to metabolic diseases and developing targeted therapeutic strategies.
Pancreas- ZIP and ZnT Transporters in Insulin Granule Biogenesis and Inflammation: Zinc, is indispensable for insulin production, storage, and controlled secretion in pancreatic β-cells. The ZIP and ZnT, each transporter performs distinct, non-overlapping roles necessary for insulin granule production and cellular responses to metabolic and inflammatory stressors. ZnT8 is the main zinc exporter in insulin-producing β-cells membranes where it transports cytosolic zinc into the lumen for insulin crystallization [83]. This transport is essential for insulin crystallization, which allows dense storage, proteolytic degradation protection, and structural stability. ZnT8-KO mice showed significant reduction in crystalline insulin granules. Moreover in ZnT8−/− knockout mice, β-cells form immature, less dense granules packed with water-soluble insulin instead of solid zinc-insulin crystals [84]. These mice can still process and secrete basal insulin, but they struggle to maintain glucose homeostasis under metabolic stress, such as a high-fat diet. During high metabolic demand, crystalline insulin produced from ZnT8 is essential for reserve and packaging. Human genome-wide association studies (GWAS) have identified risk variants in the SLC30A8 gene, which encodes the ZnT8, that are associated with a higher risk for T2D [85,86]. This gene is strongly linked to T2D pathogenesis, with specific common polymorphisms and rare LOF mutations impacting the risk of developing the disease. The finding underscores ZnT8's role in T2D, suggesting it as a potential therapeutic target. In addition to ZnT8, islet cells have ZnT5 and ZnT7 in their ER and Golgi apparatus [87]. These transporters collaborate to give zinc for proinsulin folding, hexamerization, and maturation before granulation. Proper insulin production and storage granule integrity orchestrated by the functioning of these transporters. The ZIP family of zinc importers helps maintain β-cell zinc homeostasis, especially under high insulin demand. Glucose stimulation upregulates transporters such ZIP6, ZIP7, ZIP8, and ZIP14 in mouse and human islets, increasing zinc absorption to replace granule reserves and fulfill cell biosynthetic needs. Recent study highlighted that ZIP14 acts as a negative regulator of glucose-stimulated insulin secretion (GSIS) by regulating the zinc trafficking and proper compartmentalization of intracellular zinc, particularly involving the ER [88]. This signifies the dynamic interplay between ZIP and ZnT transporters in regulating the zinc trafficking during insulin production, storage, and secretion. On the other hand, inflammatory states considering exposure to cytokines may drastically alter the expression and function of zinc transporters in β-cells during inflammatory conditions [89]. This modification may cause β-cell malfunction and death, which is frequently evinced in metabolic disorders like T2D. Pro-inflammatory stimuli can disrupt the expression of ZnT8 and ZIP transporters, thereby impairing zinc-dependent processes and linking immunometabolic stress to β-cell failure. In overview, zinc transporters particularly ZnT8 and the various members of the ZIP family are indispensable for orchestrating the biogenesis, structural integrity, and functionality of insulin granules in pancreatic β-cells. Dysregulation of these transporters results in decreased insulin processing, secretion, and β-cell resilience under inflammatory and metabolic stress. Understanding zinc homeostasis in pancreas function is crucial since such abnormalities are the key cause for diabetes and other metabolic diseases.
Adipose Tissue: As well-known, ZIP and ZnT transporters serve as crucial regulators of zinc homeostasis in the adipose tissue also, and their dysregulation profoundly affects adipose inflammation, insulin resistance, and overall metabolic health in this tissue. Within this complex tissue, zinc transporters particularly those belonging to the ZIP and ZnT families are essential regulators of immune responses, metabolic functions, inflammation, and insulin sensitivity across various adipose depots. Among these transporters, most important candidate is ZIP14 due to its significant increased expression in the white adipose tissue (WAT) during systemic inflammation and metabolic stress [90]. This upregulation is especially prominent in adipocytes in the conditions like lipopolysaccharide-induced endotoxemia, where inflammatory signals concomitantly stimulate a rise in ZIP14 expression. ZIP14 overexpression facilitate a surge in zinc influx into cells, effectively redistributing zinc in response to pro-inflammatory cytokines: IL-6 and TNF-α. Studies have revealed striking consequences of ZIP14 deficiency in the ZIP14KO murine model. ZIP14 deficient mice exhibited significant adipocyte hypertrophy, increased leptin production, and raised concentrations of pro-inflammatory cytokines such as IL-6, TNF-α, and IL-1β [91]. Additionally, ZIP14 deletion disrupts normal adipocyte differentiation and altered fat mass to lean mass ratio, resulting in a phenotype marked by metabolic endotoxemia, excessive adipose tissue growth, and systemic insulin resistance [92]. This diseased condition closely resembles the characteristics of human metabolic syndrome and T2D, highlighting the significance of ZIP14 in metabolic health. Moreover, in mice, the deficiency of ZIP14 leads to extensive systemic dysfunctions beyond adipocyte impacts, particularly chronic, low-grade systemic inflammation (metabolic endotoxemia), substantial manganese (Mn) accumulation in the brain resulting in neurotoxicity and motor impairments, as well as disrupted glucose and iron metabolism [93,94].
Another study showed that ZIP14 inhibition resulted in reduced bioavailability of cytosolic zinc, leading to dysregulation of pivotal inflammatory pathways, including the JAK/STAT and NF-κB signaling pathways [95]. The other members such as ZIP13, ZIP8, and ZnT7, have also been shown to play significant roles in modulating adipose tissue functionality and inflammatory responses [96]. In study ZIP13-null mice displayed enhanced thermogenesis and improved insulin sensitivity, suggesting a complex interplay between zinc transport and adipose tissue browning [97]. On the other hand, apart from zinc efflux, the other family i.e. ZnT transporters, significantly influences on adipokine secretion, fat metabolism, and local immune responses. In chronic inflammatory conditions, such as obesity, studies has documented association with altered expression and functionality of both ZIP and ZnT transporters, perpetuating a vicious cycle of adipose inflammation and metabolic dysfunction. Overall, the ZIP and ZnT transporter families' delicate balance of zinc influx and efflux determines adipose tissue's immunological and metabolic characteristics. The findings on ZIP disruption and inflammation, abnormal fat accumulation, and insulin resistance highlights the mechanistic importance of zinc homeostasis in adipose health and illness, making zinc transporters interesting targets for obesity-related metabolic diseases. The investigation into these transporters may lead to new treatments for metabolic illnesses associated to malfunctioning adipose tissue.
Gut Barrier: Firstly, it is well documented that zinc homeostasis is a critical player to maintain the gut health and its permeability. inc is essential for maintaining the integrity of the intestinal mucosal barrier, and protecting against pathogens and toxins [98]. A complex network of all the zinc transporters, intricately regulated to maintain the intestinal barrier and regulates immunological responses. Gut epithelium and immunological cells need these transporters to precisely absorb and distribute zinc. Proper zinc homeostasis maintains tight junctions, the mucosal layer, antimicrobial peptide production, and resident and recruited immune cell activity. ZIP4, the ZIP family's major zinc transporter, is located at enterocytes' apical membranes [6,99]. Of note, studies have shown that ZIP4 mutation result in acrodermatitis enteropathica, a condition that highlights the transporter’s critical role in zinc homeostasis and overall gut health [100]. Other ZIP transporter family member, including ZIP2, ZIP6, ZIP7, and ZIP10, are also transcriptionally regulated in response to dietary zinc levels and activation of the aryl hydrocarbon receptor (AHR), which collectively boosts zinc import and fortifies intestinal barrier function [101]. The zinc influx is critical player for maintaining the integrity of tight junction proteins including claudins and occludins which is again mediated by specific zinc transporters, primarily ZIP4 for dietary absorption and ZIP14 for systemic zinc uptake into enterocytes [102]. In ZIP14 knockout (ZIP14KO) mice, the observed increase in intestinal permeability, local inflammatory responses, and reduction in major histocompatibility complex class II (MHCII) gene expression are a consequence of intracellular zinc deficiency in intestinal epithelial cells (IECs) and the resulting epigenetic modifications and altered signaling pathways [103]. At the same time, Loss of the ZIP14 transporter in Zip14ΔIEC experimental mice notably impacted intestinal integrity by disrupting chromatin accessibility and altering gene expression, which compromised the gut barrier and immune response through mechanisms involving zinc-dependent histone deacetylases (HDACs) and key transcription factors like CIITA. Furthermore, ZIP14 has been shown to influence gut microbiome composition, important for epithelial zinc homeostasis, microbial diversity, and immune functionality [92]. ZnT2 is predominantly found in the secretory granules of Paneth and goblet cells, where it plays a critical role for the secretion of antimicrobial peptides and mucins [104]. In the ZnT2-null mouse model, the Paneth cells, exhibited impaired function due to zinc-deficient secretory granules, leading to a compromised chemical barrier. Though ZnT1 and other ZnT members have been studied less extensively in the context of the intestine, they are believed to play a protective role by preventing cytosolic zinc overload and modulating zinc signaling during episodes of intestinal stress.
On the other hand, zinc is essential for tight junction protein production and maintenance, and deficits due to transporter failure or diet may cause intestinal permeability, or "leaky gut." A weakened mucosal layer, decreased antimicrobial peptide synthesis, and dysregulated innate and adaptive immune responses are linked with this disease. In past decades, the research showed the significance of Zinc acting as a signaling cofactor via GPR39, activating PKCζ to improve epithelial barrier function [105]. Zinc supplementation and plant-derived AHR ligands increase zinc importer transcription, notably ZIP4, which has been shown to upregulate tight junction and mucin genes, strengthening barrier defense mechanisms [101]. Moreover, zinc transporter dysfunction is shown to weaken gut physical and chemical barriers and intestinal immune cell antigen presentation and cytokine transmission. This disruption may enhance microbial product translocation into systemic circulation, causing inflammation and metabolic abnormalities which may further leads to different pathological conditions such as metaflammation, IBD, obesity, and T2D [6,7,75,106]. Thus, ZIP and ZnT transporters are essential for zinc-dependent gut barrier function, immunological homeostasis, and microbiota composition. Their malfunction might significantly impact on gut integrity and immunological competency, exposing people to chronic inflammatory diseases and health concerns which underscore for better understanding of transporters' complex functions in zinc homeostasis and metabolic illness.
Table 3.
Tissue-Specific Zinc Transporters, Functions, and Pathological Implications with Supporting Studies.
Table 3.
Tissue-Specific Zinc Transporters, Functions, and Pathological Implications with Supporting Studies.
| Tissue | Zinc Transporters | Functions | Pathological Implications & Supporting Studies | References |
|---|---|---|---|---|
| Liver | ZIP14, ZIP8, ZIP1, ZIP10, ZnT1, ZnT5, ZnT6 | ZIPs mediate hepatic zinc uptake (ZIP14, ZIP8), respond to inflammation/acute phase (ZIP14); ZIP1/ZIP10 support vesicular influx; ZnT1 exports zinc. | Dysregulated ZIP14 leads to hepatic inflammation, NAFLD, and insulin resistance; ZnT1 alterations impact systemic zinc homeostasis. ZIP1 transits between plasma membrane and intracellular vesicles based on zinc levels. | [79,81,107,108,109] |
| Pancreas | ZnT8, ZnT5, ZnT7, ZIP6, ZIP7, ZIP8, ZIP1 | ZnT8 loads zinc into insulin granules for packaging/maturation (T2D linkage); ZnT5/7 regulate zinc in ER/Golgi for hormone biosynthesis; ZIPs maintain cytosolic/organelle zinc homeostasis; ZIP1 & ZIP3 is detected in mouse/pig pancreatic tissue. | ZnT8 dysfunction causes β-cell failure, impaired insulin secretion, and diabetes risk; ZIP6/7/8 impairment affects proinsulin processing and stress responses in beta cells. ZIP1 can localize to vesicular structures in transfected cells. | [37,87,110] |
| Adipose Tissue | ZIP14, ZIP13, ZIP8, ZIP1, ZnT7, ZnT5 | ZIP14 mediates zinc influx during inflammation, impacts immune signaling; ZIP13 regulates secretory pathway and adipocyte differentiation/BMP/TGF-β signaling (knockout leads to vesicular zinc build-up in fibroblasts); ZnT7 influences fat metabolism. | ZIP13 dysfunction leads to adipose inflammation and altered fat mass. ZIP14 upregulation is linked with metabolic syndrome and obesity-associated inflammation. ZnT7 influences insulin sensitivity and adiposity. ZIP1 and ZIP8 involved in adipocyte zinc homeostasis and cytokine response. | [91,96,97,111,112] |
| Gut (Intestine) | ZIP4, ZIP8, ZIP1, ZIP10, ZnT1, ZnT2, ZnT4 | ZIP4 mediates dietary zinc absorption on apical surface (ZIP4 mutations: acrodermatitis enteropathica); ZIP8 maintains immune cell zinc levels; ZIP1/ZIP10 contribute to epithelial zinc balance; ZnT1 exports into circulation (basolateral), ZnT2 supports zinc granule secretion in Paneth/goblet cells. | ZIP4 essential for intestinal health and systemic zinc; ZnT1 expressed highly in gut, supporting serum zinc levels. ZnT4 contributes to vesicle trafficking in enterocytes. ZIP8, ZIP1, ZIP10 regulate intestinal immunity and barrier integrity. | [99,100,101,104,113,114] |
| Kidney | ZIP8, ZIP1, ZnT3, ZIP10, ZnT1, ZnT4, ZnT8 | ZIPs/ZnTs support renal zinc reabsorption, homeostasis, and excretion; ZnT1/2 mRNA unique in kidney; ZnT4 and ZnT6 traffic in vesicular compartment. | Zinc imbalance impairs kidney function, ZnT3/ZIP8/ZIP1 changes affect nephropathy risk; transporter regulation controls acute-phase systemic zinc redistribution. | [60,115,116,117,118] |
| Brain | ZIP3, ZIP8, ZIP1, ZIP6, ZIP7, ZnT1, ZnT3, ZnT4, ZnT6 | ZnT3 loads zinc into synaptic vesicles for neurotransmission; ZIP3/8/1/6 manage neuronal zinc influx and homeostasis; ZIP7 located constitutively in Golgi/ER in neurons and glia; ZnT4, ZnT6 detected in neural vesicles. | ZnT3 critical for synaptic plasticity, alteration linked to neurodegeneration; ZIP7 antibody stains Golgi/ER regions in diverse cell types, including neurons and glia. Transporter imbalance may affect cognition and neuroinflammation. | [119,120,121] |
5. Zinc-Modulated Signaling Pathways
Zinc is known as a dynamic signal transducer, also serve as a second messenger by regulating signaling cascades, affecting protein activities, and enabling cellular communication between external stimuli and intracellular responses. Recent studies have shown zinc plays a role with rapid intracellular zinc release concomitant with calcium signaling, to influence cellular responses [122]. Intracellular zinc signaling is divided into two categories. One is the relatively swift, stimulus-driven “zinc waves,” and the other involves more gradual, transcription-mediated changes depending on the expression of zinc transporters [1]. Zinc waves occur within minutes following receptor activation such as the FcɛRI in mast cells requiring both calcium influx and the activation of the MEK signaling cascade. This phenomenon leads to a transient increase in free cytosolic zinc levels which directly modulates the critical protein phosphatases and kinases activity. Notably, zinc inhibits the activity of phosphatases, thus prolonging the activation of mitogen-activated protein kinases (MAPK) including ERK, JNK, and p38. Beyond these rapid signaling events, zinc also influences long-term cellular responses by affecting expression and function level of different transcription factors, most notably the MTF-1. Conversely, MTF-1 senses cytosolic zinc fluctuations and subsequently regulates the expression of MTs and Zinc transporters, involved with zinc homeostasis [16]. By activating these genes, MTF-1 helps maintain intracellular zinc balance and cellular adaptation to stress or excess metal exposure.
5.1. Zinc’s Crosstalk with Canonical Pathways: NF-κB, MAPKs, NLRP3, TLRs,
Zinc plays a crucial role in suppressing the nuclear translocation of NF-κB by stabilizing its inhibitor, IκB, and directly inhibiting the activity of upstream kinases [123]. Conversely, when zinc levels are depleted, there is an enhancement in NF-κB-driven inflammatory gene expression, which can activate NLRP3 inflammasomes, a key component in metaflammation and immune responses [54]. The interplay between zinc waves and extracellular zinc influx activates key signaling pathways such as ERK and Akt. This happens through the phosphatases inhibition and direct stimulation of Ras and MEK, resulting to the subsequent phosphorylation and activation of CREB and Fosl1 [124,125]. Depleted zinc levels also activate stress-responsive kinases like JNK and p38, which are integral to the cellular response to inflammatory stimuli and stress [126]. Adequate intracellular zinc levels serve to restrict the activation of the NLRP3 inflammasome, thereby mitigating the release of the pro-inflammatory cytokine IL-1β [127]. In contrast, zinc chelation or deficiency removes this inhibitory effect, resulting in an exaggerated inflammatory response.
Crosstalk Between Zinc, TLRs, and Metabolic Sensors: Zinc is essential for the immune system, playing a dual role as a cofactor and a signaling molecule (secondary messenger) that connects innate immunity and metabolic regulation. As a cofactor, it regulates thousands of proteins involved in basic cell functions like DNA replication and cell division. Interestingly, zinc ions also can function as a signaling molecule, modulating intracellular signaling pathways in immune cells and serving as a second messenger in response to stimuli. This dual function allows zinc to markedly affect cellular responses to diverse danger signals, especially via its interaction with TLR and metabolic sensor pathways. On the other hand, TLRs are crucial pattern recognition receptors (PRRs) that serve as a first line of defense in the immune system by detecting molecules from microbes (like LPS) and also from the body (damage-associated molecular patterns or DAMPs) [128]. Recent research shows that TLRs can also be activated by metabolic stresses, such as saturated fatty acids, and that this activation is linked to chronic inflammation in metabolic diseases like obesity. Zinc's involvement in TLR signaling can be categorized into several critical functions which has been discussed briefly here. After being stimulated by LPS or palmitate, it directs rapid activation of NF-κB, with subsequent enhancement of pro-inflammatory cytokines [129]. Though this initial response is tightly regulated by negative feedback mechanisms to prevent excessive inflammation and potential tissue damage. As a feedback mechanism this activation also triggers an upregulation of zinc importers, particularly ZIP14, facilitating the sequestration of zinc in the liver, a commonly observed response during acute inflammation [93]. Wherein, adequate zinc level is critical for negative regulation to curb excessive inflammation driven by TLRs.
In addition, zinc pilots MyD88-dependent pathways, to decrease pro-inflammatory cytokine production by direct inhibition of IKK, preventing the activation and nuclear translocation of the NF-κB. By blocking NF-κB, zinc restricts the downstream inflammatory response [130]. Zinc does indeed influence the functions of precise regulatory proteins like A-20 and the other zinc-finger protein, which again underscores its crucial role in preserving the critical equilibrium or the fine line between immune activation and its tolerance [131]. Research from past decades consistently highlighting importance of zinc as a trace element essential for the immune system, often modulating immune responses by serving as a critical cofactor for various enzymes and regulatory molecules.
Transporter Interactions Zinc transporters, particularly ZIP8 and ZIP14, play a significant role in modulating pro-inflammatory signaling. They help restrict NF-κB activity and influence the activity of protein tyrosine phosphatases, thus finely tuning the cellular responses to TLR activation [69,89,93]. Zinc and Metabolic Sensors Beyond its role in immune signaling, zinc is also integral to the regulation of various metabolic sensors that align energy and nutrient status with immune function.
Zinc is known to modulate AMPK activity, which serves as a master regulator of cellular energy homeostasis and fatty acid oxidation. As it is known that AMPK integrates metabolic stress signals to maintain cellular energy equilibrium and has anti-inflammatory characteristics. Moreover, zinc may affect mTOR signaling pathways, which are vital for controlling autophagy, immune cell differentiation, and anabolic activities necessary for cell growth and metabolism [132]. Of note, zinc has been shown as a critical regulator in experimental tissues for activating insulin receptors and the ensuing IRS-1/PI3K/Akt signaling pathway, facilitating effective glucose absorption and metabolism [133]. The zinc transporter ZnT8 is particularly important for insulin crystallization and secretion in β-cells, while zinc in peripheral tissues plays a role in modulating insulin sensitivity and downstream metabolic pathways. As mentioned earlier, in pathological conditions such as obesity and type 2 diabetes, metabolic stressors like SFA and ROS trigger chronic, low-grade inflammation via activation of TLRs and inflammasomes, which in turn leads to altered expression of zinc transporters like increased ZIP14 and decreased ZnT8 [93]. Disruption of intracellular zinc levels initiates a malicious cycle between metabolic and inflammatory signaling pathways by impairing the body's innate anti-inflammatory and insulin-sensitizing mechanisms, which in turn further dysregulate zinc equilibrium. Thus, extensive experimental and clinical research underscores zinc's critical role as an integrator of immune and metabolic signaling pathways. As shown, zinc modulates TLR responses and metabolic sensors, hence influencing cytokine production, metabolic adaptability, and the course of many illnesses. Consequently, focusing on zinc homeostasis and the related transporter pathways offers a viable therapeutic strategy for treating inflammatory and metabolic disorders.
Table 4.
Zinc-Modulated Signaling Pathways, Roles, and Physiological Impacts (with Literature Support).
Table 4.
Zinc-Modulated Signaling Pathways, Roles, and Physiological Impacts (with Literature Support).
| Signaling Pathway | Zinc’s Role | Physiological Impact | Study Details | References |
|---|---|---|---|---|
| Zinc Waves | Acts as a second messenger; rapid release from ER/perinuclear stores after receptor stimulation (FcεRI, TLR, cAMP/PKA) | Modulates protein tyrosine phosphatase activity, prolongs MAPK activation, amplifies/controls cytokine (IL-6, TNF-α) production | Zinc waves occur within minutes after FcεRI crosslinking, dependent on Ca²⁺ and MEK signals. Inhibits phosphatases and sustains MAPKs; first described in mast cells. | [1] |
| NF-κB | Inhibits IκB kinase (IKK), stabilizes IκB, directly and indirectly restricts NF-κB nuclear translocation | Suppresses pro-inflammatory gene expression (e.g., TNF-α, IL-1β); zinc deficiency or transporter dysfunction relieves this suppression | Zinc wave enhances cytokine gene induction via prolonged MAPK and potentially NF-κB activation after FcεRI stimulation .Zinc essentially gates the amplitude/duration of the NF-κB response . | [1,9,54] |
| MTF-1 | Direct zinc sensor; zinc binding activates metal response elements, upregulating metallothioneins and select ZnT genes | Promotes cellular defense against oxidative stress; increases zinc buffering capacity; adapts transporter profile to stress | Zinc exposure or cytosolic elevation leads to MTF-1 nuclear translocation and oxidative stress protection, well-documented in immune and liver cells . | [16] |
| MAPKs (ERK, JNK, p38) | Zinc waves/influx modulate phosphorylation, inhibiting protein phosphatases, sustaining MAPK signaling | Controls cell proliferation, inflammation, cytokine output, and survival/differentiation signals | Zinc ionophores mimic zinc wave by prolonging MAPK activation, increasing late-phase IL-6/TNF-α expression in mast cells . | [1] |
| NLRP3 Inflammasome | Zinc deficiency or oxidation-driven displacement of zinc from proteins activates NLRP3 inflammasome, increases IL-1β | Promotes metaflammation, insulin resistance, and chronic inflammatory disease | Zinc supplementation inhibits NLRP3 activation; deficiency/oxidative stress enhances it. Linked to response in macrophages, adipose tissue . | [54] |
| TLRs | Zinc suppresses MyD88 and canonical NF-κB pathway activation in TLR4/2 signaling, modulates inflammatory threshold | Prevents excessive cytokine release on microbial/metabolic stimulation; restricts prolonged inflammation | TLR activation results in rapid transporter regulation and a decrease in free zinc as an early signal for dendritic cell activation. ZIP14 and ZIP8 up/downregulation tightly couple TLR activity to zinc homeostasis. | [93,130] |
| Insulin Signaling | Zinc enhances Akt activation, supports phosphorylation cascade; ZnT8 ensures proper insulin packaging/release | Promotes glucose uptake, insulin secretion, and β-cell function; deficiency linked to impaired glycemic control | ZnT8 mutations disrupt insulin granule biogenesis and secretion, increasing T2D risk. Zinc signaling also influences IRS-1/PI3K/Akt sensitivity in target tissues. | [133,134] |
6. Therapeutic Implications and Translational Potential of Zinc Transporters
Recent advancements have established zinc transporters as promising therapeutic targets for metabolic disorders like T2D, NAFLD, and obesity. The therapeutic landscape has been traversed via principal intervention strategies which includes employing promising clinically approved small-molecule modulators to affect zinc transporter activity, the implementation of nutritional zinc supplementation, and the direct targeting of zinc transporter proteins as viable drug candidates in clinical applications.
Small Molecule Modulators of Zinc Transporters: The development of small compounds that may regulate the activity or expression of zinc transporters has considerable promise for reestablishing zinc homeostasis and improving cellular signaling pathways affected in several disease conditions. For instance, though currently, there are no specific, approved small-molecule drugs that directly target the zinc transporter ZIP5 for clinical use in T2D, NAFLD, or obesity. On the basis of existing research on genetic studies and experimental models, which suggest that pharmacological inhibition of SLC39A5 could be a viable therapeutic strategy in the future. This effect has been observed in human subjects harboring LOF mutations as well as in genetically modified mouse models, even when subjected to high-fat, high-fructose diets [135]. Furthermore, targeting the transporter ZIP13 has shown promise in modulating adipocyte browning, which could be beneficial in the context of obesity and T2D management [97,136]. ZnT8 in pancreatic beta cells for insulin synthesis, storage, and release, positioning those as a key target for diabetes therapies. Strategy-based therapies for insulin resistance, glucose intolerance, and dysregulated lipid metabolism may include zinc transporter augmentation or inhibition [137]. For targeted therapeutics for metabolic illnesses such as T2D, NAFLD, and obesity. Research shows that addressing zinc imbalances in metabolic organs including the pancreas, liver, adipose tissue, and muscle may restore cellular function.
Nutritional interventions involving zinc supplementation have been linked to favorable outcomes in glycemic control and overall metabolic health, as evidenced by various animal studies and clinical trials [74,138,139]. Supplementation study from different research has been shown to has the ability to improve insulin sensitivity in individuals, decrease fasting glucose levels, lower triglyceride concentrations, and alleviate systemic inflammation, especially in those with zinc insufficiency [59,74]. In populations such as obese mice and humans suffering from metabolic syndrome or T2D, zinc supplementation has been associated with improved insulin response and beneficial alterations in metabolic profiles. Personalized zinc therapy is an emerging field, and while routine clinical practice does not yet widely offer specific zinc dosing based on an individual's full genetic profile, certain genetic tests and specialized medical consultation are available. To pursue personalized zinc therapy based on genetic variations, you would typically need to undergo genetic testing ordered by a medical professional or a specialized nutritionist, followed by consultation with a dietitian or genetic counselor to tailor a plan. This field is a developing area of nutritional science, and the clinical application of specific genetic variants for general zinc supplementation recommendations is still evolving.
Zinc Transporters as Druggable Targets in T2D, NAFLD, and Obesity: Several zinc transporters exhibit significant translational potential for drug development. Research indicates that genetic ablation or pharmacological inhibition of ZIP5 transporter can lower the risk of diabetes and provide protection against hepatic metabolic dysfunction, supported by clinical evidence from large population studies and direct animal experimentation [140]. Variants of ZnT8 transporter, whether gain- or loss-of-function, have been associated with altered susceptibility to T2D, primarily through their impact on insulin storage and release. Current investigations are exploring modulation of ZnT8 as a therapeutic strategy for diabetes management [134,141]. On the other hand, ZIP13 and ZIP14 transporters have been documented as essential controller of adipogenesis, insulin sensitivity, and hepatic glucose metabolism as discussed earlier. Their modification may target concerns such as insulin resistance, adipose tissue inflammation, and steatosis with considerable implications for the treatment of obesity and NAFLD. Though there is no currently FDA-approved drugs that specifically target ZIP13 or ZIP14 for the treatment of metabolic disorders. However, research in animal models has identified potential therapeutic strategies involving the modulation of zinc signaling pathways associated with these transporters. For example bortezomib, used in genetic disorder (Spondylocheirodysplastic Ehlers-Danlos syndrome, SCD-EDS) caused by mutations in the ZIP13 gene helped to restore normal ZIP13 protein levels and intracellular zinc homeostasis in cellular models which can be a promising repurposed drug option for common metabolic diseases [142]. Therefore, targeting the zinc signaling cascade might reveal a potential restorative approach. As from the above discussion we know that zinc transporters not only orchestrate systemic zinc disbandment but also engage in cellular signaling mechanisms that control metabolic operations and inflammatory responses. Continued advancements in our understanding of the structure, regulation, and function of these transporters will accelerate the development of innovative small molecules and biologics, paving the way for new therapeutic options in the management of diabetes, fatty liver disease, and obesity.
6.1. Pharmaceutical Development and Contemporary Case Analyses:
In the realm of drug development, significant advancements have emerged, particularly with regards to targeting of different zinc transporters considering different pathological conditions. These efforts are mainly focused on two primary strategies: the direct modulation of transporter activity and the chemical inhibition of excessive zinc trafficking. Recent study, published in April 2025, used gene-edited human embryonic stem cell-derived β cells (SC-β cells) and human primary islet models to specifically examine the role of zinc and demonstrated that an increased influx of zinc causes ER stress, eventually resulting in β cell death, a mechanism of considerable importance in diabetes pathogenesis [143]. Another study by Batta et al. aligns closely with current efforts to pharmacologically modulate zinc homeostasis by identifying small-molecule ligands that selectively target members of the ZIP and ZnT transporter families [144]. Using in silico docking, Batta and a different research group screened multiple phytochemical and drug-like compounds against modeled structures of ZIP4, ZIP8, ZIP10, and ZnT8, identifying several candidates with notable predicted binding affinity most prominently quercetin, kaempferol, genistein, resveratrol, and curcumin [145]. The recognized compounds had promising potential and interactions with the transporter's active or regulatory regions, suggesting possible altering influences on zinc translocation. This study seeks to realize the ultimate goal of developing isoform-specific modulators for zinc transporters. It founded the first computational framework to facilitate the selection of potent compounds, thereby encouraging further biochemical confirmation and therapeutic outcome.
On the other hand, utilizing a sophisticated drug-screening platform centered on isogenic SC-β cells, researchers made a groundbreaking discovery that low-doses of anisomycin (at a concentration of 25 nM), which is conventionally recognized as a protein synthesis inhibitor at higher doses, showed selective inhibition of the over-transportation of zinc [143]. Remarkably, treatment with a low-dose (25 nM) anisomycin was found to protect human pancreatic beta cells from stress-induced cell death and prevent the onset of T2D in mice fed a high-fat diet. This low-dose anisomycin not only safeguarded human β cells from stress-induced cell death but also demonstrated efficacy in vivo. Rui et al showed that when administered to primary human islets and diabetic mice, it successfully prevented the onset of type 2 diabetes induced by high-fat diet. This unique pharmaceutical technique targeting zinc transporters is a novel therapeutic approach to maintaining β-cell function in diabetic population [143]. A significant milestone in the field of translational zinc biology has been achieved with the identification of LOF mutations in the zinc transporter ZIP5. Comprehensive large-scale exome and cohort studies across many ethnic groups have shown that people with uncommon LOF mutations in ZIP5 have significantly increased blood zinc levels and a much decreased risk of developing T2D [146]. Functional assays corroborated that these mutations lead to a complete loss of transporter function, resulting in systemic zinc elevation. Similar to human genetic findings, ZIP5/Slc39a5-null mice had elevated tissue zinc levels, decreased fasting glucose, improved hepatic insulin sensitivity, and reduced liver inflammation and fibrosis, even with congenital or diet-induced obesity [146]. These strong in vivo data suggest that ZIP5 may be a viable metabolic illness treatment target, especially for individuals who do not benefit from zinc supplementation alone. Clinical investigations of zinc supplementation in metabolic syndrome, obesity, and diabetes patients have consistently shown improvements in insulin resistance and metabolic biomarkers. However, recent approaches are unwinding to prioritize the modulation of intracellular zinc trafficking.
To circumvent the limits of "one-size-fits-all" supplementing techniques, small compounds or transporter proteins are used to address the molecular causes of metabolic disease dysregulations. Due to both gain- and loss-of-function variants of ZnT8 have been shown to affect β cell function and insulin granule biogenesis it drawn attention in diabetes. This duality in genetic variation justifies the exploration of both pharmacological enhancement and inhibition as viable strategies, tailored according to individual patient genotypes.
Recent advancements in zinc-centric pharmacological research have highlighted zinc transporters as potential therapeutic targets for inflammatory disorders, such as rheumatoid arthritis (RA). Case studies in experimental arthritis models, namely collagen-induced arthritis (CIA) mice and ex vivo synovial fibroblast systems, show that the modulation of zinc flux may influence critical inflammatory pathways [147,148]. ZIP8 has been shown to be highly elevated in arthritic synovium and has been proposed as a druggable target due to its zinc-dependent stimulation of MMP-9 and MMP-13, which is directly responsible for cartilage degradation [149]. Small-molecule ZIP8 inhibitors and siRNA-mediated suppression methods have shown effectiveness in mitigating joint inflammation and matrix deterioration in CIA models [150]. Moreover, targeting the ZIP14 transporter, which moves zinc in response to cytokines in tissues like the liver and joints or other synovial tissue, can reduce systemic inflammation and the acute-phase response [67,151,152]. Compounds that regulate ZnT1 and ZnT5 activity are being studied for their ability to restore cytosolic zinc buffering capacity and inhibit NF-κB–mediated cytokine production [153]. These case studies together highlight the potential of zinc-transporter-focused therapies, establishing them as promising candidates for the development of next-generation anti-inflammatory drugs.
Table 5.
Zinc Transporter Proteins as Therapeutic Drug Targets: Clinical Trials and Translational Studies.
Table 5.
Zinc Transporter Proteins as Therapeutic Drug Targets: Clinical Trials and Translational Studies.
| Zinc Transporter / Target | Mechanism / Rationale | Disease / Condition | Drug/Intervention Type | Study Details | references |
|---|---|---|---|---|---|
| ZnT8 | Zinc transport into insulin granules (β-cell specific); impacts insulin maturation and secretion | Type 2 Diabetes (T2D) | Targeted modulator / Precision therapy (in development) | GWAS and rare variant studies: loss-of-function alleles reduce T2D risk .Ongoing drug development focused on enhancing or mimicking protective variants. ZnT8-KO mouse studies confirm islet-specific function. | [154,155] |
| ZIP5 | Regulates glucose sensing & insulin secretion in β-cells; impacts gut/pancreas zinc handling | Diabetes, Metabolic Diseases | Small molecule/Genetic modulation (preclinical) | Mouse knockout protects against glucose dysregulation and pancreatic zinc toxicity. SLC39A5 variants studied in large cohorts; shown to modulate serum zinc and glucose homeostasis in humans and animals. | [146] |
| ZIP8 | Modulates zinc uptake in gut/liver/adipose; influences innate immunity, metabolism, Crohn's disease risk | Crohn's Disease, Gut/Liver Inflammation | Genetic and pharmacological modulation (early translational phase) | Functional variant linked with Crohn's disease and microbiome composition. Modifiers of ZIP8 studied in immune/inflammatory disease animal models. | [156,157] |
| ZIP10 | Controls B-cell receptor signaling, humoral immunity, anti-apoptotic signaling | Hematologic malignancy, Immunodeficiency | Genetic targeting / Therapeutic antibodies (preclinical) | ZIP10 critical for B cell survival; mouse genetic studies. ZIP10 inhibitors/enhancers under investigation for immune modulation; drug development in early preclinical phase. | [158] |
| ZIP13 | Regulates vascular and cardiac/skin function; upregulation linked to fibrosis and inflammation | Cardiovascular disease, Fibrosis | Small molecule inhibitor / antisense RNA (in development) | Mouse ZIP13 downregulation reduces ischemia/reperfusion injury via CaMKII regulation. Pharmacologic inhibition as a therapeutic strategy is under study. | [159] |
| ZIP4 | Dietary zinc absorption / homeostasis; overexpression in cancer | Pancreatic & GI cancers, Acrodermatitis Enteropathica | Antibody drugs / Antisense oligonucleotides | Anti-ZIP4 therapies in preclinical cancer studies. Genetic therapies for acrodermatitis enteropathica under development. | [99,160] |
| ZnT1 | Exports zinc from cells; affects systemic and tissue zinc levels | Zinc deficiency/excess, intestinal disorders | Dietary/Pharmacological / Translational biomarker | Plays a role in dietary and supplemental zinc absorption; involved in biomarker trial (NCT01062347) . | [161,162] |
| SLC transporters (class) | General therapeutic target class: several subtypes including SLC30A, SLC39A individually druggable | T2D, metabolic, cancer, inflammation | Small molecule/Monoclonal antibodies/combo therapy | SLC30A8 and related SLCs identified as most promising for metabolic indications from human genetic & animal studies; some SLCs targeted in marketed and experimental cancer/metabolic drugs. | [163,164,165] |
7. Future Perspectives and Open Questions
Zinc's role as a dynamic immunometabolic regulator has changed our knowledge of chronic illness development and opened new treatment avenues. However, zinc signaling's molecular details, tissue-specific control, and translational promise for precision metabolic therapy are still little understood. One major challenge lies in the context-specific nature of zinc transporter regulation. To achieve this, single-cell mapping of transporter expression in physiological and pathological states and tissue-specific delivery strategies like nanoparticle carriers or gene promoters are needed to precisely modulate transporter function. To achieve therapeutic selectivity, ligand-specific modulators that utilize transporter isoform structural differences might be designed. We also need biomarkers that can track intracellular zinc dynamics and transporter function in real time. Circulating miRNAs (e.g., miR-34a in NAFLD), metallothionein levels as proxies for cytosolic zinc buffering and redox balance, and zinc-sensitive contrast agents or genetically encoded biosensors for noninvasive PET or MRI zinc flux imaging seem promising. Genetic variants in SLC30 and SLC39 families may predict zinc-targeted therapy response, allowing patient classification and customized intervention. T2D and NAFLD are complicated, hence zinc-based monotherapies may not be enough. Instead, integrative combination techniques can work better. Zinc modulation may boost the metabolic advantages of GLP-1 receptor agonists, synergize with anti-inflammatory medicines like IL-1β inhibitors or NLRP3 antagonists, or improve gut-immune interaction with microbiome-directed therapy. Co-targeting AMPK, mTOR, and zinc homeostasis may improve cellular energy balance and immune-metabolic integration. To maximize its therapeutic potential, future translational research should focus on multi-axis techniques and incorporate zinc signaling into metabolic and inflammatory control.
Conclusion:
Both preclinical and human genetic studies have underscored the critical role zinc homeostasis in metabolic health, showing that alterations in transporter expression, genetic mutations, or disease-induced mis-localization can all lead to the pathogenesis of T2D, NFLD, obesity, and/or related metabolic disturbances. Breakthrough studies in imaging and biosensor advances have made zinc a second messenger and revealed its signaling mechanisms. Zinc transporter mutations, protect metabolic illness models, proving that targeting transporter activity when zinc supplementation is inadequate is therapeutic. Though, precise regulation of zinc flux in disease-relevant tissues is a critical emerging area in biomedical research, as future therapeutic success is highly dependent on addressing tissue specificity and context-driven treatment methods. Altogether, modulation of zinc transporter function—either by small molecules, gene targeting, or precision nutritional strategies—holds significant translational potential to restore immunometabolic balance and reduce the burden of chronic metabolic diseases. Integrating zinc biology into clinical practice for metabolic inflammation is an area of active research facing severe challenges and opportunities in biomarker development, drug delivery, and combinatorial treatment paradigms. Continued multidisciplinary efforts are required to translate current findings into next-generation therapies.
References
- Chen, B.; Yu, P.; Chan, W.N.; Xie, F.; Zhang, Y.; Liang, L.; Leung, K.T.; Lo, K.W.; Yu, J.; Tse, G.M.K.; et al. Cellular Zinc Metabolism and Zinc Signaling: From Biological Functions to Diseases and Therapeutic Targets. Signal Transduct. Target. Ther. 2024, 9, 6. [CrossRef]
- Kimura, T.; Kambe, T. The Functions of Metallothionein and ZIP and ZnT Transporters: An Overview and Perspective. Int. J. Mol. Sci. 2016, 17, 336. [CrossRef]
- Charles-Messance, H.; Mitchelson, K.A.J.; Castro, E.D.M.; Sheedy, F.J.; Roche, H.M. Regulating Metabolic Inflammation by Nutritional Modulation. J. Allergy Clin. Immunol. 2020, 146, 706–720. [CrossRef]
- Aydemir, T.B.; Troche, C.; Kim, M.-H.; Cousins, R.J. Hepatic ZIP14-Mediated Zinc Transport Contributes to Endosomal Insulin Receptor Trafficking and Glucose Metabolism*. J. Biol. Chem. 2016, 291, 23939–23951. [CrossRef]
- Barman, S.; Srinivasan, K. Diabetes and Zinc Dyshomeostasis: Can Zinc Supplementation Mitigate Diabetic Complications? Crit. Rev. Food Sci. Nutr. 2022, 62, 1046–1061. [CrossRef]
- Barman, S.; Pradeep, S.R.; Srinivasan, K. Zinc Supplementation Mitigates Its Dyshomeostasis in Experimental Diabetic Rats by Regulating the Expression of Zinc Transporters and Metallothionein†. Metallomics 2017, 9, 1765–1777. [CrossRef]
- Barman, S.; Srinivasan, K. Ameliorative Effect of Zinc Supplementation on Compromised Small Intestinal Health in Streptozotocin-Induced Diabetic Rats. Chem. Biol. Interact. 2019, 307, 37–50. [CrossRef]
- Fan, Y.-G.; Wu, T.-Y.; Zhao, L.-X.; Jia, R.-J.; Ren, H.; Hou, W.-J.; Wang, Z.-Y. From Zinc Homeostasis to Disease Progression: Unveiling the Neurodegenerative Puzzle. Pharmacol. Res. 2024, 199, 107039. [CrossRef]
- Paik, S.; Kim, J.K.; Silwal, P.; Sasakawa, C.; Jo, E.-K. An Update on the Regulatory Mechanisms of NLRP3 Inflammasome Activation. Cell. Mol. Immunol. 2021, 18, 1141–1160. [CrossRef]
- Jeong, J.; Eide, D.J. The SLC39 Family of Zinc Transporters. Mol. Aspects Med. 2013, 34, 612–619. [CrossRef]
- Kambe, T.; Taylor, K.M.; Fu, D. Zinc Transporters and Their Functional Integration in Mammalian Cells. J. Biol. Chem. 2021, 296, 100320. [CrossRef]
- Roca-Umbert, A.; Garcia-Calleja, J.; Vogel-González, M.; Fierro-Villegas, A.; Ill-Raga, G.; Herrera-Fernández, V.; Bosnjak, A.; Muntané, G.; Gutiérrez, E.; Campelo, F.; et al. Human Genetic Adaptation Related to Cellular Zinc Homeostasis. PLOS Genet. 2023, 19, e1010950. [CrossRef]
- Krężel, A.; Maret, W. The Bioinorganic Chemistry of Mammalian Metallothioneins. Chem. Rev. 2021, 121, 14594–14648. [CrossRef]
- Bafaro, E.; Liu, Y.; Xu, Y.; Dempski, R.E. The Emerging Role of Zinc Transporters in Cellular Homeostasis and Cancer. Signal Transduct. Target. Ther. 2017, 2, 17029. [CrossRef]
- Oteiza, P.I. Zinc and the Modulation of Redox Homeostasis. Free Radic. Biol. Med. 2012, 53, 1748–1759. [CrossRef]
- Laity, J.H.; Andrews, G.K. Understanding the Mechanisms of Zinc-Sensing by Metal-Response Element Binding Transcription Factor-1 (MTF-1). Arch. Biochem. Biophys. 2007, 463, 201–210. [CrossRef]
- Watanabe, S.; Amagai, Y.; Sannino, S.; Tempio, T.; Anelli, T.; Harayama, M.; Masui, S.; Sorrentino, I.; Yamada, M.; Sitia, R.; et al. Zinc Regulates ERp44-Dependent Protein Quality Control in the Early Secretory Pathway. Nat. Commun. 2019, 10, 603. [CrossRef]
- Wagatsuma, T.; Shimotsuma, K.; Sogo, A.; Sato, R.; Kubo, N.; Ueda, S.; Uchida, Y.; Kinoshita, M.; Kambe, T. Zinc Transport via ZNT5-6 and ZNT7 Is Critical for Cell Surface Glycosylphosphatidylinositol-Anchored Protein Expression. J. Biol. Chem. 2022, 298, 102011. [CrossRef]
- Kim, M.-H.; Aydemir, T.B.; Kim, J.; Cousins, R.J. Hepatic ZIP14-Mediated Zinc Transport Is Required for Adaptation to Endoplasmic Reticulum Stress. Proc. Natl. Acad. Sci. U. S. A. 2017, 114, E5805–E5814. [CrossRef]
- Amagai, Y.; Yamada, M.; Kowada, T.; Watanabe, T.; Du, Y.; Liu, R.; Naramoto, S.; Watanabe, S.; Kyozuka, J.; Anelli, T.; et al. Zinc Homeostasis Governed by Golgi-Resident ZnT Family Members Regulates ERp44-Mediated Proteostasis at the ER-Golgi Interface. Nat. Commun. 2023, 14, 2683. [CrossRef]
- Suzuki, T.; Ishihara, K.; Migaki, H.; Ishihara, K.; Nagao, M.; Yamaguchi-Iwai, Y.; Kambe, T. Two Different Zinc Transport Complexes of Cation Diffusion Facilitator Proteins Localized in the Secretory Pathway Operate to Activate Alkaline Phosphatases in Vertebrate Cells*. J. Biol. Chem. 2005, 280, 30956–30962. [CrossRef]
- Yuasa, H.; Morino, N.; Wagatsuma, T.; Munekane, M.; Ueda, S.; Matsunaga, M.; Uchida, Y.; Katayama, T.; Katoh, T.; Kambe, T. ZNT5-6 and ZNT7 Play an Integral Role in Protein N-Glycosylation by Supplying Zn2+ to Golgi α-Mannosidase II. J. Biol. Chem. 2024, 300, 107378. [CrossRef]
- McCormick, N.H.; Kelleher, S.L. ZnT4 Provides Zinc to Zinc-Dependent Proteins in the Trans-Golgi Network Critical for Cell Function and Zn Export in Mammary Epithelial Cells. Am. J. Physiol.-Cell Physiol. 2012, 303, C291–C297. [CrossRef]
- Liu, H.Y.; Gale, J.R.; Reynolds, I.J.; Weiss, J.H.; Aizenman, E. The Multifaceted Roles of Zinc in Neuronal Mitochondrial Dysfunction. Biomedicines 2021, 9, 489. [CrossRef]
- Ishida, H.; Yo, R.; Zhang, Z.; Shimizu, T.; Ohto, U. Cryo-EM Structures of the Zinc Transporters ZnT3 and ZnT4 Provide Insights into Their Transport Mechanisms. FEBS Lett. 2025, 599, 41–52. [CrossRef]
- Rivera, O.C.; Geddes, D.T.; Barber-Zucker, S.; Zarivach, R.; Gagnon, A.; Soybel, D.I.; Kelleher, S.L. A Common Genetic Variant in Zinc Transporter ZnT2 (Thr288Ser) Is Present in Women with Low Milk Volume and Alters Lysosome Function and Cell Energetics. Am. J. Physiol.-Cell Physiol. 2020, 318, C1166–C1177. [CrossRef]
- Chowanadisai, W.; Lönnerdal, B.; Kelleher, S.L. Identification of a Mutation in SLC30A2 (ZnT-2) in Women with Low Milk Zinc Concentration That Results in Transient Neonatal Zinc Deficiency*. J. Biol. Chem. 2006, 281, 39699–39707. [CrossRef]
- Davidson, H.W.; Wenzlau, J.M.; O’Brien, R.M. Zinc Transporter 8 (ZnT8) and β Cell Function. Trends Endocrinol. Metab. 2014, 25, 415–424. [CrossRef]
- Kukic, I.; Lee, J.K.; Coblentz, J.; Kelleher, S.L.; Kiselyov, K. Zinc-Dependent Lysosomal Enlargement in TRPML1-Deficient Cells Involves MTF-1 Transcription Factor and ZnT4 (Slc30a4) Transporter. Biochem. J. 2013, 451, 155–163. [CrossRef]
- Aydemir, T.B.; Liuzzi, J.P.; McClellan, S.; Cousins, R.J. Zinc Transporter ZIP8 (SLC39A8) and Zinc Influence IFN-γ Expression in Activated Human T Cells. J. Leukoc. Biol. 2009, 86, 337–348. [CrossRef]
- Wang, F.; Dufner-Beattie, J.; Kim, B.-E.; Petris, M.J.; Andrews, G.; Eide, D.J. Zinc-Stimulated Endocytosis Controls Activity of the Mouse ZIP1 and ZIP3 Zinc Uptake Transporters. J. Biol. Chem. 2004, 279, 24631–24639. [CrossRef]
- Xian, Y.; Zhou, M.; Hu, Y.; Liu, J.; Zhu, W.; Wang, Y. Free Zinc Determines the Formability of the Vesicular Dense Core in Diabetic Beta Cells. Cell Insight 2022, 1, 100020. [CrossRef]
- McAllister, B.B.; Dyck, R.H. Zinc Transporter 3 (ZnT3) and Vesicular Zinc in Central Nervous System Function. Neurosci. Biobehav. Rev. 2017, 80, 329–350. [CrossRef]
- Davidson, H.W.; Wenzlau, J.M.; O’Brien, R.M. ZINC TRANSPORTER 8 (ZNT8) AND BETA CELL FUNCTION. Trends Endocrinol. Metab. TEM 2014, 25, 415–424. [CrossRef]
- Lee, S.; Rivera, O.C.; Kelleher, S.L. Zinc Transporter 2 Interacts with Vacuolar ATPase and Is Required for Polarization, Vesicle Acidification, and Secretion in Mammary Epithelial Cells. J. Biol. Chem. 2017, 292, 21598–21613. [CrossRef]
- Kambe, T.; Matsunaga, M.; Takeda, T. Understanding the Contribution of Zinc Transporters in the Function of the Early Secretory Pathway. Int. J. Mol. Sci. 2017, 18, 2179. [CrossRef]
- Wijesekara, N.; Dai, F.F.; Hardy, A.B.; Giglou, P.R.; Bhattacharjee, A.; Koshkin, V.; Chimienti, F.; Gaisano, H.Y.; Rutter, G.A.; Wheeler, M.B. Beta Cell Specific ZnT8 Deletion in Mice Causes Marked Defects in Insulin Processing, Crystallization and Secretion. Diabetologia 2010, 53, 1656–1668. [CrossRef]
- Palmiter, R.D.; Cole, T.B.; Quaife, C.J.; Findley, S.D. ZnT-3, a Putative Transporter of Zinc into Synaptic Vesicles. Proc. Natl. Acad. Sci. U. S. A. 1996, 93, 14934–14939. [CrossRef]
- Fukunaka, A.; Kurokawa, Y.; Teranishi, F.; Sekler, I.; Oda, K.; Ackland, M.L.; Faundez, V.; Hiromura, M.; Masuda, S.; Nagao, M.; et al. Tissue Nonspecific Alkaline Phosphatase Is Activated via a Two-Step Mechanism by Zinc Transport Complexes in the Early Secretory Pathway. J. Biol. Chem. 2011, 286, 16363–16373. [CrossRef]
- Bafaro, E.; Liu, Y.; Xu, Y.; Dempski, R.E. The Emerging Role of Zinc Transporters in Cellular Homeostasis and Cancer. Signal Transduct. Target. Ther. 2017, 2, 17029. [CrossRef]
- Thornton, J.K.; Taylor, K.M.; Ford, D.; Valentine, R.A. Differential Subcellular Localization of the Splice Variants of the Zinc Transporter ZnT5 Is Dictated by the Different C-Terminal Regions. PLOS ONE 2011, 6, e23878. [CrossRef]
- Bai, M.; Cui, Y.; Sang, Z.; Gao, S.; Zhao, H.; Mei, X. Zinc Ions Regulate Mitochondrial Quality Control in Neurons under Oxidative Stress and Reduce PANoptosis in Spinal Cord Injury Models via the Lgals3-Bax Pathway. Free Radic. Biol. Med. 2024, 221, 169–180. [CrossRef]
- Dabravolski, S.A.; Sadykhov, N.K.; Kartuesov, A.G.; Borisov, E.E.; Sukhorukov, V.N.; Orekhov, A.N. Interplay between Zn2+ Homeostasis and Mitochondrial Functions in Cardiovascular Diseases and Heart Ageing. Int. J. Mol. Sci. 2022, 23, 6890. [CrossRef]
- Deng, H.; Qiao, X.; Xie, T.; Fu, W.; Li, H.; Zhao, Y.; Guo, M.; Feng, Y.; Chen, L.; Zhao, Y.; et al. SLC-30A9 Is Required for Zn2+ Homeostasis, Zn2+ Mobilization, and Mitochondrial Health. Proc. Natl. Acad. Sci. 2021, 118, e2023909118. [CrossRef]
- Jiang, D.; Sullivan, P.G.; Sensi, S.L.; Steward, O.; Weiss, J.H. Zn2+ Induces Permeability Transition Pore Opening and Release of Pro-Apoptotic Peptides from Neuronal Mitochondria *. J. Biol. Chem. 2001, 276, 47524–47529. [CrossRef]
- Mendoza, A.D.; Dietrich, N.; Tan, C.-H.; Herrera, D.; Kasiah, J.; Payne, Z.; Cubillas, C.; Schneider, D.L.; Kornfeld, K. Lysosome-Related Organelles Contain an Expansion Compartment That Mediates Delivery of Zinc Transporters to Promote Homeostasis. Proc. Natl. Acad. Sci. 2024, 121, e2307143121. [CrossRef]
- Kimura, T.; Kambe, T. The Functions of Metallothionein and ZIP and ZnT Transporters: An Overview and Perspective. Int. J. Mol. Sci. 2016, 17, 336. [CrossRef]
- Lichtlen, P.; Schaffner, W. Putting Its Fingers on Stressful Situations: The Heavy Metal-regulatory Transcription Factor MTF-1. BioEssays 2001, 23, 1010–1017. [CrossRef]
- Charles-Messance, H.; Mitchelson, K.A.J.; De Marco Castro, E.; Sheedy, F.J.; Roche, H.M. Regulating Metabolic Inflammation by Nutritional Modulation. J. Allergy Clin. Immunol. 2020, 146, 706–720. [CrossRef]
- Hotamisligil, G.S. Foundations of Immunometabolism and Implications for Metabolic Health and Disease. Immunity 2017, 47, 406–420. [CrossRef]
- Clemente-Suárez, V.J.; Beltrán-Velasco, A.I.; Redondo-Flórez, L.; Martín-Rodríguez, A.; Tornero-Aguilera, J.F. Global Impacts of Western Diet and Its Effects on Metabolism and Health: A Narrative Review. Nutrients 2023, 15, 2749. [CrossRef]
- Diet Review: Anti-Inflammatory Diet • The Nutrition Source 2021.
- Milanski, M.; Degasperi, G.; Coope, A.; Morari, J.; Denis, R.; Cintra, D.E.; Tsukumo, D.M.L.; Anhe, G.; Amaral, M.E.; Takahashi, H.K.; et al. Saturated Fatty Acids Produce an Inflammatory Response Predominantly through the Activation of TLR4 Signaling in Hypothalamus: Implications for the Pathogenesis of Obesity. J. Neurosci. 2009, 29, 359–370. [CrossRef]
- Finucane, O.M.; Lyons, C.L.; Murphy, A.M.; Reynolds, C.M.; Klinger, R.; Healy, N.P.; Cooke, A.A.; Coll, R.C.; McAllan, L.; Nilaweera, K.N.; et al. Monounsaturated Fatty Acid–Enriched High-Fat Diets Impede Adipose NLRP3 Inflammasome–Mediated IL-1β Secretion and Insulin Resistance Despite Obesity. Diabetes 2015, 64, 2116–2128. [CrossRef]
- Li, X.; Ren, Y.; Chang, K.; Wu, W.; Griffiths, H.R.; Lu, S.; Gao, D. Adipose Tissue Macrophages as Potential Targets for Obesity and Metabolic Diseases. Front. Immunol. 2023, 14, 1153915. [CrossRef]
- Barman, S.; Srinivasan, K. Zinc Supplementation Ameliorates Diabetic Cataract Through Modulation of Crystallin Proteins and Polyol Pathway in Experimental Rats. Biol. Trace Elem. Res. 2019, 187, 212–223. [CrossRef]
- Izgilov, R.; Kislev, N.; Omari, E.; Benayahu, D. Advanced Glycation End-Products Accelerate Amyloid Deposits in Adipocyte’s Lipid Droplets. Cell Death Dis. 2024, 15, 846. [CrossRef]
- Asadipooya, K.; Lankarani, K.B.; Raj, R.; Kalantarhormozi, M. RAGE Is a Potential Cause of Onset and Progression of Nonalcoholic Fatty Liver Disease. Int. J. Endocrinol. 2019, 2019, 2151302. [CrossRef]
- Barman, S.; Srinivasan, K. Attenuation of Oxidative Stress and Cardioprotective Effects of Zinc Supplementation in Experimental Diabetic Rats. Br. J. Nutr. 2017, 117, 335–350. [CrossRef]
- Barman, S.; Pradeep, S.R.; Srinivasan, K. Zinc Supplementation Alleviates the Progression of Diabetic Nephropathy by Inhibiting the Overexpression of Oxidative-Stress-Mediated Molecular Markers in Streptozotocin-Induced Experimental Rats. J. Nutr. Biochem. 2018, 54, 113–129. [CrossRef]
- Hotamisligil, G.S. Inflammation, Metaflammation and Immunometabolic Disorders. Nature 2017, 542, 177–185. [CrossRef]
- Marreiro, D. do N.; Cruz, K.J.C.; Morais, J.B.S.; Beserra, J.B.; Severo, J.S.; de Oliveira, A.R.S. Zinc and Oxidative Stress: Current Mechanisms. Antioxidants 2017, 6, 24. [CrossRef]
- Wessels, I.; Maywald, M.; Rink, L. Zinc as a Gatekeeper of Immune Function. Nutrients 2017, 9, 1286. [CrossRef]
- Wessels, I.; Haase, H.; Engelhardt, G.; Rink, L.; Uciechowski, P. Zinc Deficiency Induces Production of the Proinflammatory Cytokines IL-1β and TNFα in Promyeloid Cells via Epigenetic and Redox-Dependent Mechanisms. J. Nutr. Biochem. 2013, 24, 289–297. [CrossRef]
- Dierichs, L.; Kloubert, V.; Rink, L. Cellular Zinc Homeostasis Modulates Polarization of THP-1-Derived Macrophages. Eur. J. Nutr. 2018, 57, 2161–2169. [CrossRef]
- Kulik, L.; Maywald, M.; Kloubert, V.; Wessels, I.; Rink, L. Zinc Deficiency Drives Th17 Polarization and Promotes Loss of Treg Cell Function. J. Nutr. Biochem. 2019, 63, 11–18. [CrossRef]
- Liuzzi, J.P.; Lichten, L.A.; Rivera, S.; Blanchard, R.K.; Aydemir, T.B.; Knutson, M.D.; Ganz, T.; Cousins, R.J. Interleukin-6 Regulates the Zinc Transporter Zip14 in Liver and Contributes to the Hypozincemia of the Acute-Phase Response. Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 6843–6848. [CrossRef]
- Merriman, C.; Fu, D. Down-Regulation of the Islet-Specific Zinc Transporter-8 (ZnT8) Protects Human Insulinoma Cells against Inflammatory Stress. J. Biol. Chem. 2019, 294, 16992–17006. [CrossRef]
- Liu, M.-J.; Bao, S.; Gálvez-Peralta, M.; Pyle, C.J.; Rudawsky, A.C.; Pavlovicz, R.E.; Killilea, D.W.; Li, C.; Nebert, D.W.; Wewers, M.D.; et al. ZIP8 Regulates Host Defense through Zinc-Mediated Inhibition of NF-κB. Cell Rep. 2013, 3, 386–400. [CrossRef]
- Summersgill, H.; England, H.; Lopez-Castejon, G.; Lawrence, C.B.; Luheshi, N.M.; Pahle, J.; Mendes, P.; Brough, D. Zinc Depletion Regulates the Processing and Secretion of IL-1β. Cell Death Dis. 2014, 5, e1040. [CrossRef]
- Jimenez-Rondan, F.R.; Ruggiero, C.H.; Cousins, R.J. Long Noncoding RNA, MicroRNA, Zn Transporter Zip14 (Slc39a14) and Inflammation in Mice. Nutrients 2022, 14, 5114. [CrossRef]
- Taslim, N.A.; Graciela, A.M.; Harbuwono, D.S.; Syauki, A.Y.; Anthony, A.N.; Ashari, N.; Aman, A.M.; Tjandrawinata, R.R.; Hardinsyah, H.; Bukhari, A.; et al. Zinc as a Modulator of miRNA Signaling in Obesity. Nutrients 2025, 17, 3375. [CrossRef]
- Ye, Z.; Cheng, M.; Fan, L.; Ma, J.; Zhang, Y.; Gu, P.; Xie, Y.; You, X.; Zhou, M.; Wang, B.; et al. Plasma microRNA Expression Profiles Associated with Zinc Exposure and Type 2 Diabetes Mellitus: Exploring Potential Role of miR-144-3p in Zinc-Induced Insulin Resistance. Environ. Int. 2023, 172, 107807. [CrossRef]
- Barman, S.; Srinivasan, K. Zinc Supplementation Alleviates Hyperglycemia and Associated Metabolic Abnormalities in Streptozotocin-Induced Diabetic Rats. Can. J. Physiol. Pharmacol. 2016, 94, 1356–1365. [CrossRef]
- Mitchell, S.B.; Aydemir, T.B. Roles of Zinc and Zinc Transporters in Development, Progression, and Treatment of Inflammatory Bowel Disease (IBD). Front. Nutr. 2025, 12, 1649658. [CrossRef]
- Wan, Y.; Zhang, B. The Impact of Zinc and Zinc Homeostasis on the Intestinal Mucosal Barrier and Intestinal Diseases. Biomolecules 2022, 12, 900. [CrossRef]
- Fan, Y.; Zhang, X.; Yang, L.; Wang, J.; Hu, Y.; Bian, A.; Liu, J.; Ma, J. Zinc Inhibits High Glucose-Induced NLRP3 Inflammasome Activation in Human Peritoneal Mesothelial Cells. Mol. Med. Rep. 2017, 16, 5195–5202. [CrossRef]
- Aydemir, T.B.; Cousins, R.J. The Multiple Faces of the Metal Transporter ZIP14 (SLC39A14). J. Nutr. 2018, 148, 174–184. [CrossRef]
- Winslow, J.W.W.; Limesand, K.H.; Zhao, N. The Functions of ZIP8, ZIP14, and ZnT10 in the Regulation of Systemic Manganese Homeostasis. Int. J. Mol. Sci. 2020, 21, 3304. [CrossRef]
- Aydemir, T.B.; Chang, S.-M.; Guthrie, G.J.; Maki, A.B.; Ryu, M.-S.; Karabiyik, A.; Cousins, R.J. Zinc Transporter ZIP14 Functions in Hepatic Zinc, Iron and Glucose Homeostasis during the Innate Immune Response (Endotoxemia). PLOS ONE 2012, 7, e48679. [CrossRef]
- Gartmann, L.; Wex, T.; Grüngreiff, K.; Reinhold, D.; Kalinski, T.; Malfertheiner, P.; Schütte, K. Expression of Zinc Transporters ZIP4, ZIP14 and ZnT9 in Hepatic Carcinogenesis—An Immunohistochemical Study. J. Trace Elem. Med. Biol. 2018, 49, 35–42. [CrossRef]
- Su, W.; Feng, M.; Liu, Y.; Cao, R.; Liu, Y.; Tang, J.; Pan, K.; Lan, R.; Mao, Z. ZnT8 Deficiency Protects From APAP-Induced Acute Liver Injury by Reducing Oxidative Stress Through Upregulating Hepatic Zinc and Metallothioneins. Front. Pharmacol. 2021, 12. [CrossRef]
- Germanos, M.; Gao, A.; Taper, M.; Yau, B.; Kebede, M.A. Inside the Insulin Secretory Granule. Metabolites 2021, 11, 515. [CrossRef]
- Lemaire, K.; Ravier, M.A.; Schraenen, A.; Creemers, J.W.M.; Van de Plas, R.; Granvik, M.; Van Lommel, L.; Waelkens, E.; Chimienti, F.; Rutter, G.A.; et al. Insulin Crystallization Depends on Zinc Transporter ZnT8 Expression, but Is Not Required for Normal Glucose Homeostasis in Mice. Proc. Natl. Acad. Sci. 2009, 106, 14872–14877. [CrossRef]
- da Silva Xavier, G.; Bellomo, E.A.; McGinty, J.A.; French, P.M.; Rutter, G.A. Animal Models of GWAS-Identified Type 2 Diabetes Genes. J. Diabetes Res. 2013, 2013, 906590. [CrossRef]
- Sladek, R.; Rocheleau, G.; Rung, J.; Dina, C.; Shen, L.; Serre, D.; Boutin, P.; Vincent, D.; Belisle, A.; Hadjadj, S.; et al. A Genome-Wide Association Study Identifies Novel Risk Loci for Type 2 Diabetes. Nature 2007, 445, 881–885. [CrossRef]
- Cai, Y.; Kirschke, C.P.; Huang, L. SLC30A Family Expression in the Pancreatic Islets of Humans and Mice: Cellular Localization in the β-Cells. J. Mol. Histol. 2018, 49, 133–145. [CrossRef]
- Liu, Y.; Batchuluun, B.; Ho, L.; Zhu, D.; Prentice, K.J.; Bhattacharjee, A.; Zhang, M.; Pourasgari, F.; Hardy, A.B.; Taylor, K.M.; et al. Characterization of Zinc Influx Transporters (ZIPs) in Pancreatic β Cells. J. Biol. Chem. 2015, 290, 18757–18769. [CrossRef]
- Egefjord, L.; Jensen, J.L.; Bang-Berthelsen, C.H.; Petersen, A.B.; Smidt, K.; Schmitz, O.; Karlsen, A.E.; Pociot, F.; Chimienti, F.; Rungby, J.; et al. Zinc Transporter Gene Expression Is Regulated by Pro-Inflammatory Cytokines: A Potential Role for Zinc Transporters in Beta-Cell Apoptosis? BMC Endocr. Disord. 2009, 9, 7. [CrossRef]
- Troche, C.; Beker Aydemir, T.; Cousins, R.J. Zinc Transporter Slc39a14 Regulates Inflammatory Signaling Associated with Hypertrophic Adiposity. Am. J. Physiol. - Endocrinol. Metab. 2016, 310, E258–E268. [CrossRef]
- Aydemir, T.B.; Kim, M.-H.; Kim, J.; Colon-Perez, L.M.; Banan, G.; Mareci, T.H.; Febo, M.; Cousins, R.J. Metal Transporter Zip14 (Slc39a14) Deletion in Mice Increases Manganese Deposition and Produces Neurotoxic Signatures and Diminished Motor Activity. J. Neurosci. 2017, 37, 5996–6006. [CrossRef]
- Mitchell, S.B.; Thorn, T.L.; Lee, M.-T.; Kim, Y.; Comrie, J.M.C.; Bai, Z.S.; Johnson, E.L.; Aydemir, T.B. Metal Transporter SLC39A14/ZIP14 Modulates Regulation between the Gut Microbiome and Host Metabolism. Am. J. Physiol. - Gastrointest. Liver Physiol. 2023, 325, G593–G607. [CrossRef]
- Aydemir, T.B.; Chang, S.-M.; Guthrie, G.J.; Maki, A.B.; Ryu, M.-S.; Karabiyik, A.; Cousins, R.J. Zinc Transporter ZIP14 Functions in Hepatic Zinc, Iron and Glucose Homeostasis during the Innate Immune Response (Endotoxemia). PLOS ONE 2012, 7, e48679. [CrossRef]
- McCabe, S.; Limesand, K.; Zhao, N. Recent Progress toward Understanding the Role of ZIP14 in Regulating Systemic Manganese Homeostasis. Comput. Struct. Biotechnol. J. 2023, 21, 2332–2338. [CrossRef]
- Troche, C.; Beker Aydemir, T.; Cousins, R.J. Zinc Transporter Slc39a14 Regulates Inflammatory Signaling Associated with Hypertrophic Adiposity. Am. J. Physiol. - Endocrinol. Metab. 2016, 310, E258–E268. [CrossRef]
- Tepaamorndech, S.; Kirschke, C.P.; Pedersen, T.L.; Keyes, W.R.; Newman, J.W.; Huang, L. Zinc Transporter 7 Deficiency Affects Lipid Synthesis in Adipocytes by Inhibiting Insulin-Dependent Akt Activation and Glucose Uptake. FEBS J. 2016, 283, 378–394. [CrossRef]
- Fukunaka, A.; Fukada, T.; Bhin, J.; Suzuki, L.; Tsuzuki, T.; Takamine, Y.; Bin, B.-H.; Yoshihara, T.; Ichinoseki-Sekine, N.; Naito, H.; et al. Zinc Transporter ZIP13 Suppresses Beige Adipocyte Biogenesis and Energy Expenditure by Regulating C/EBP-β Expression. PLoS Genet. 2017, 13, e1006950. [CrossRef]
- Barman, S.; Srinivasan, K. Enhanced Intestinal Absorption of Micronutrients in Streptozotocin-Induced Diabetic Rats Maintained on Zinc Supplementation. J. Trace Elem. Med. Biol. 2018, 50, 182–187. [CrossRef]
- Andrews, G.K. Regulation and Function of Zip4, the Acrodermatitis Enteropathica Gene. Biochem. Soc. Trans. 2008, 36, 1242–1246. [CrossRef]
- Kuliyev, E.; Zhang, C.; Sui, D.; Hu, J. Zinc Transporter Mutations Linked to Acrodermatitis Enteropathica Disrupt Function and Cause Mistrafficking. J. Biol. Chem. 2021, 296, 100269. [CrossRef]
- Hu, X.; Xiao, W.; Lei, Y.; Green, A.; Lee, X.; Maradana, M.R.; Gao, Y.; Xie, X.; Wang, R.; Chennell, G.; et al. Aryl Hydrocarbon Receptor Utilises Cellular Zinc Signals to Maintain the Gut Epithelial Barrier 2022, 2022.11.03.515052.
- Wan, Y.; Zhang, B. The Impact of Zinc and Zinc Homeostasis on the Intestinal Mucosal Barrier and Intestinal Diseases. Biomolecules 2022, 12, 900. [CrossRef]
- Jimenez-Rondan, F.R.; Ruggiero, C.H.; Riva, A.; Yu, F.; Stafford, L.S.; Cross, T.R.; Larkin, J.; Cousins, R.J. Deletion of Metal Transporter Zip14 Reduces Major Histocompatibility Complex II Expression in Murine Small Intestinal Epithelial Cells. Proc. Natl. Acad. Sci. 2025, 122, e2422321121. [CrossRef]
- Podany, A.B.; Wright, J.; Lamendella, R.; Soybel, D.I.; Kelleher, S.L. ZnT2-Mediated Zinc Import Into Paneth Cell Granules Is Necessary for Coordinated Secretion and Paneth Cell Function in Mice. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 369–383. [CrossRef]
- Shao, Y.-X.; Lei, Z.; Wolf, P.G.; Gao, Y.; Guo, Y.-M.; Zhang, B.-K. Zinc Supplementation, via GPR39, Upregulates PKCζ to Protect Intestinal Barrier Integrity in Caco-2 Cells Challenged by Salmonella Enterica Serovar Typhimurium. J. Nutr. 2017, 147, 1282–1289. [CrossRef]
- Franco, C.; Canzoniero, L.M.T. Zinc Homeostasis and Redox Alterations in Obesity. Front. Endocrinol. 2024, 14. [CrossRef]
- Lichten, L.A.; Ryu, M.-S.; Guo, L.; Embury, J.; Cousins, R.J. MTF-1-Mediated Repression of the Zinc Transporter Zip10 Is Alleviated by Zinc Restriction. PloS One 2011, 6, e21526. [CrossRef]
- Jou, M.-Y.; Hall, A.G.; Philipps, A.F.; Kelleher, S.L.; Lönnerdal, B. Tissue-Specific Alterations in Zinc Transporter Expression in Intestine and Liver Reflect a Threshold for Homeostatic Compensation during Dietary Zinc Deficiency in Weanling Rats. J. Nutr. 2009, 139, 835–841. [CrossRef]
- Ziatdinova, M.M.; Мунирoвна, З.М.; Valova, Y.V.; В, В.Я.; Mukhammadiyeva, G.F.; Ф, М.Г.; Fazlieva, A.S.; С, Ф.А.; Karimov, D.D.; Д, К.Д.; et al. Analysis of MT1 and ZIP1 Gene Expression in the Liver of Rats with Chronic Poisoning with Cadmium Chloride. Hyg. Sanit. 2021, 100, 1298–1302. [CrossRef]
- Dufner-Beattie, J.; Huang, Z.L.; Geiser, J.; Xu, W.; Andrews, G.K. Mouse ZIP1 and ZIP3 Genes Together Are Essential for Adaptation to Dietary Zinc Deficiency during Pregnancy. Genes. N. Y. N 2000 2006, 44, 239–251. [CrossRef]
- Fukada, T.; Civic, N.; Furuichi, T.; Shimoda, S.; Mishima, K.; Higashiyama, H.; Idaira, Y.; Asada, Y.; Kitamura, H.; Yamasaki, S.; et al. The Zinc Transporter SLC39A13/ZIP13 Is Required for Connective Tissue Development; Its Involvement in BMP/TGF-β Signaling Pathways. PLoS ONE 2008, 3, e3642. [CrossRef]
- Bahadoran, Z.; Ghafouri-Taleghani, F.; Todorčević, M. Zinc and Adipose Organ Dysfunction: Molecular Insights into Obesity and Metabolic Disorders. Curr. Nutr. Rep. 2025, 14, 117. [CrossRef]
- Samuelson, D.R.; Haq, S.; Knoell, D.L. Divalent Metal Uptake and the Role of ZIP8 in Host Defense Against Pathogens. Front. Cell Dev. Biol. 2022, 10. [CrossRef]
- McMahon, R.J.; Cousins, R.J. Regulation of the Zinc Transporter ZnT-1 by Dietary Zinc. Proc. Natl. Acad. Sci. U. S. A. 1998, 95, 4841–4846. [CrossRef]
- Cai, Z.; Wu, X.; Wang, T.; Song, Z.; Ni, P.; Zhong, M.; Su, Y.; Xie, E.; Sun, S.; Lin, Y.; et al. SLC39A8-Mediated Zinc Dyshomeostasis Potentiates Kidney Disease. Proc. Natl. Acad. Sci. U. S. A. 2025, 122, e2426352122. [CrossRef]
- Chen, W.; Lu, H.; Ying, Y.; Li, H.; Shen, H.; Cai, J. Zinc and Chronic Kidney Disease: A Review. J. Nutr. Sci. Vitaminol. (Tokyo) 2024, 70, 98–105. [CrossRef]
- Chi, Y.; Zhang, X.; Liang, D.; Wang, Y.; Cai, X.; Dong, J.; Li, L.; Chi, Z. ZnT8 Exerts Anti-Apoptosis of Kidney Tubular Epithelial Cell in Diabetic Kidney Disease Through TNFAIP3-NF-κB Signal Pathways. Biol. Trace Elem. Res. 2023, 201, 2442–2457. [CrossRef]
- Liuzzi, J.P.; Blanchard, R.K.; Cousins, R.J. Differential Regulation of Zinc Transporter 1, 2, and 4 mRNA Expression by Dietary Zinc in Rats. J. Nutr. 2001, 131, 46–52. [CrossRef]
- McAllister, B.B.; Dyck, R.H. Zinc Transporter 3 (ZnT3) and Vesicular Zinc in Central Nervous System Function. Neurosci. Biobehav. Rev. 2017, 80, 329–350. [CrossRef]
- Grubman, A.; Lidgerwood, G.E.; Duncan, C.; Bica, L.; Tan, J.-L.; Parker, S.J.; Caragounis, A.; Meyerowitz, J.; Volitakis, I.; Moujalled, D.; et al. Deregulation of Subcellular Biometal Homeostasis through Loss of the Metal Transporter, Zip7, in a Childhood Neurodegenerative Disorder. Acta Neuropathol. Commun. 2014, 2, 25. [CrossRef]
- Sabouri, S.; Rostamirad, M.; Dempski, R.E. Unlocking the Brain’s Zinc Code: Implications for Cognitive Function and Disease. Front. Biophys. 2024, 2. [CrossRef]
- Alshawaf, A.J.; Alnassar, S.A.; Al-Mohanna, F.A. The Interplay of Intracellular Calcium and Zinc Ions in Response to Electric Field Stimulation in Primary Rat Cortical Neurons in Vitro. Front. Cell. Neurosci. 2023, 17. [CrossRef]
- Jarosz, M.; Olbert, M.; Wyszogrodzka, G.; Młyniec, K.; Librowski, T. Antioxidant and Anti-Inflammatory Effects of Zinc. Zinc-Dependent NF-κB Signaling. Inflammopharmacology 2017, 25, 11–24. [CrossRef]
- Chen, B.; Yu, P.; Chan, W.N.; Xie, F.; Zhang, Y.; Liang, L.; Leung, K.T.; Lo, K.W.; Yu, J.; Tse, G.M.K.; et al. Cellular Zinc Metabolism and Zinc Signaling: From Biological Functions to Diseases and Therapeutic Targets. Signal Transduct. Target. Ther. 2024, 9, 6. [CrossRef]
- Anson, K.J.; Corbet, G.A.; Palmer, A.E. Zn2+ Influx Activates ERK and Akt Signaling Pathways. Proc. Natl. Acad. Sci. U. S. A. 2021, 118, e2015786118. [CrossRef]
- Zhu, B.; Yang, C.; Liu, D.; Zhi, Q.; Hua, Z.-C. Zinc Depletion Induces JNK/P38 Phosphorylation and Suppresses Akt/mTOR Expression in Acute Promyelocytic NB4 Cells. J. Trace Elem. Med. Biol. 2023, 79, 127264. [CrossRef]
- Summersgill, H.; England, H.; Lopez-Castejon, G.; Lawrence, C.B.; Luheshi, N.M.; Pahle, J.; Mendes, P.; Brough, D. Zinc Depletion Regulates the Processing and Secretion of IL-1β. Cell Death Dis. 2014, 5, e1040–e1040. [CrossRef]
- Maywald, M.; Wessels, I.; Rink, L. Zinc Signals and Immunity. Int. J. Mol. Sci. 2017, 18, 2222. [CrossRef]
- Fessler, M.B.; Rudel, L.L.; Brown, M. Toll-like Receptor Signaling Links Dietary Fatty Acids to the Metabolic Syndrome. Curr. Opin. Lipidol. 2009, 20, 379–385. [CrossRef]
- Kim, B.; Lee, W.-W. Regulatory Role of Zinc in Immune Cell Signaling. Mol. Cells 2021, 44, 335–341. [CrossRef]
- Prasad, A.S.; Bao, B. Molecular Mechanisms of Zinc as a Pro-Antioxidant Mediator: Clinical Therapeutic Implications. Antioxidants 2019, 8, 164. [CrossRef]
- Kim, Y.-H.; Eom, J.-W.; Koh, J.-Y. Mechanism of Zinc Excitotoxicity: A Focus on AMPK. Front. Neurosci. 2020, 14. [CrossRef]
- Wong, V.V.T.; Nissom, P.M.; Sim, S.-L.; Yeo, J.H.M.; Chuah, S.-H.; Yap, M.G.S. Zinc as an Insulin Replacement in Hybridoma Cultures. Biotechnol. Bioeng. 2006, 93, 553–563. [CrossRef]
- Huang, Q.; Du, J.; Merriman, C.; Gong, Z. Genetic, Functional, and Immunological Study of ZnT8 in Diabetes. Int. J. Endocrinol. 2019, 2019, 1524905. [CrossRef]
- Chim, S.M.; Howell, K.; Dronzek, J.; Wu, W.; Hout, C.V.; Ferreira, M.A.R.; Ye, B.; Li, A.; Brydges, S.; Arunachalam, V.; et al. Genetic Inactivation of Zinc Transporter SLC39A5 Improves Liver Function and Hyperglycemia in Obesogenic Settings. eLife 2023, 12. [CrossRef]
- Fukunaka, A.; Fujitani, Y. Role of Zinc Homeostasis in the Pathogenesis of Diabetes and Obesity. Int. J. Mol. Sci. 2018, 19. [CrossRef]
- Horton, T.M.; Allegretti, P.A.; Lee, S.; Moeller, H.P.; Smith, M.; Annes, J.P. Zinc-Chelating Small Molecules Preferentially Accumulate and Function within Pancreatic β Cells. Cell Chem. Biol. 2019, 26, 213-222.e6. [CrossRef]
- Ranasinghe, P.; Wathurapatha, W.S.; Galappatthy, P.; Katulanda, P.; Jayawardena, R.; Constantine, G.R. Zinc Supplementation in Prediabetes: A Randomized Double-Blind Placebo-Controlled Clinical Trial. J. Diabetes 2018, 10, 386–397. [CrossRef]
- Ranasinghe, P.; Jayawardena, R.; Pigera, A.; Katulanda, P.; Constantine, G.R.; Galappaththy, P. Zinc Supplementation in Pre-Diabetes: Study Protocol for a Randomized Controlled Trial. Trials 2013, 14, 52. [CrossRef]
- Chim, S.M.; Howell, K.; Dronzek, J.; Wu, W.; Hout, C.V.; Ferreira, M.A.R.; Ye, B.; Li, A.; Brydges, S.; Arunachalam, V.; et al. Genetic Inactivation of Zinc Transporter SLC39A5 Improves Liver Function and Hyperglycemia in Obesogenic Settings 2021, 2021.12.08.21267440.
- Davidson, H.W.; Wenzlau, J.M.; O’Brien, R.M. ZINC TRANSPORTER 8 (ZNT8) AND BETA CELL FUNCTION. Trends Endocrinol. Metab. TEM 2014, 25, 415–424. [CrossRef]
- Bin, B.-H.; Hojyo, S.; Ryong Lee, T.; Fukada, T. Spondylocheirodysplastic Ehlers-Danlos Syndrome (SCD-EDS) and the Mutant Zinc Transporter ZIP13. Rare Dis. 2014, 2, e974982. [CrossRef]
- Hu, R.; Ma, Q.; Kong, Y.; Wang, Z.; Xu, M.; Chen, X.; Su, Y.; Xiao, T.; He, Q.; Wang, X.; et al. A Compound Screen Based on Isogenic hESC-Derived β Cell Reveals an Inhibitor Targeting ZnT8-Mediated Zinc Transportation to Protect Pancreatic β Cell from Stress-Induced Cell Death. Adv. Sci. 2025, 12, 2413161. [CrossRef]
- Batta, I.; Sharma, G. Molecular Docking Simulation to Predict Inhibitors Against Zinc Transporters 2024.
- Jangid, H.; Shah, N.H.; Dar, M.A.; Wani, A.K. Identification of Potent Phytochemical Inhibitors against Mobilized Colistin Resistance: Molecular Docking, MD Simulations, ADMET, and Toxicity Predictions. Silico Res. Biomed. 2025, 1, 100013. [CrossRef]
- Chim, S.M.; Howell, K.; Dronzek, J.; Wu, W.; Hout, C.V.; Ferreira, M.A.R.; Ye, B.; Li, A.; Brydges, S.; Arunachalam, V.; et al. Genetic Inactivation of Zinc Transporter SLC39A5 Improves Liver Function and Hyperglycemia in Obesogenic Settings. eLife 2023, 12. [CrossRef]
- Kim, B.; Kim, H.Y.; Yoon, B.R.; Yeo, J.; In Jung, J.; Yu, K.-S.; Kim, H.C.; Yoo, S.-J.; Park, J.K.; Kang, S.W.; et al. Cytoplasmic Zinc Promotes IL-1β Production by Monocytes and Macrophages through mTORC1-Induced Glycolysis in Rheumatoid Arthritis. Sci. Signal. 2022, 15, eabi7400. [CrossRef]
- Kang, J.-A.; Kwak, J.-S.; Park, S.-H.; Sim, K.-Y.; Kim, S.K.; Shin, Y.; Jung, I.J.; Yang, J.-I.; Chun, J.-S.; Park, S.-G. ZIP8 Exacerbates Collagen-Induced Arthritis by Increasing Pathogenic T Cell Responses. Exp. Mol. Med. 2021, 53, 560–571. [CrossRef]
- Kim, J.-H.; Jeon, J.; Shin, M.; Won, Y.; Lee, M.; Kwak, J.-S.; Lee, G.; Rhee, J.; Ryu, J.-H.; Chun, C.-H.; et al. Regulation of the Catabolic Cascade in Osteoarthritis by the Zinc-ZIP8-MTF1 Axis. Cell 2014, 156, 730–743. [CrossRef]
- Kang, J.-A.; Kwak, J.-S.; Park, S.-H.; Sim, K.-Y.; Kim, S.K.; Shin, Y.; Jung, I.J.; Yang, J.-I.; Chun, J.-S.; Park, S.-G. ZIP8 Exacerbates Collagen-Induced Arthritis by Increasing Pathogenic T Cell Responses. Exp. Mol. Med. 2021, 53, 560–571. [CrossRef]
- Troche, C.; Beker Aydemir, T.; Cousins, R.J. Zinc Transporter Slc39a14 Regulates Inflammatory Signaling Associated with Hypertrophic Adiposity. Am. J. Physiol.-Endocrinol. Metab. 2016, 310, E258–E268. [CrossRef]
- Gálvez-Peralta, M.; Wang, Z.; Bao, S.; Knoell, D.L.; Nebert, D.W. Tissue-Specific Induction of Mouse ZIP8 and ZIP14 Divalent Cation/Bicarbonate Symporters by, and Cytokine Response to, Inflammatory Signals. Int. J. Toxicol. 2014, 33, 246–258. [CrossRef]
- Briassoulis, G.; Briassoulis, P.; Ilia, S.; Miliaraki, M.; Briassouli, E. The Anti-Oxidative, Anti-Inflammatory, Anti-Apoptotic, and Anti-Necroptotic Role of Zinc in COVID-19 and Sepsis. Antioxidants 2023, 12, 1942. [CrossRef]
- Barman, N.; Haque, M.A.; Ridwan, M.; Ghosh, D.; Islam, A.B.M.M.K. Loss-of-Function Variant of SLC30A8 Rs13266634 (C > T) Protects against Type 2 Diabetes by Stabilizing ZnT8: Insights from Epidemiological and Computational Analyses. J. Genet. Eng. Biotechnol. 2025, 23, 100565. [CrossRef]
- Thomsen, S.K.; Gloyn, A.L. Human Genetics as a Model for Target Validation: Finding New Therapies for Diabetes. Diabetologia 2017, 60, 960–970. [CrossRef]
- Yang, J.C.; Zhao, M.; Chernikova, D.; Arias-Jayo, N.; Zhou, Y.; Situ, J.; Gutta, A.; Chang, C.; Liang, F.; Lagishetty, V.; et al. ZIP8 A391T Crohn’s Disease-Linked Risk Variant Induces Colonic Metal Ion Dyshomeostasis, Microbiome Compositional Shifts, and Inflammation. Dig. Dis. Sci. 2024, 69, 3760–3772. [CrossRef]
- Hall, S.C.; Smith, D.R.; Dyavar, S.R.; Wyatt, T.A.; Samuelson, D.R.; Bailey, K.L.; Knoell, D.L. Critical Role of Zinc Transporter (Zip8) in Myeloid Innate Immune Cell Function and the Host Response against Bacterial Pneumonia. J. Immunol. Baltim. Md 1950 2021, 207, 1357–1370. [CrossRef]
- Miyai, T.; Hojyo, S.; Ikawa, T.; Kawamura, M.; Irié, T.; Ogura, H.; Hijikata, A.; Bin, B.-H.; Yasuda, T.; Kitamura, H.; et al. Zinc Transporter SLC39A10/ZIP10 Facilitates Antiapoptotic Signaling during Early B-Cell Development. Proc. Natl. Acad. Sci. U. S. A. 2014, 111, 11780–11785. [CrossRef]
- Wang, J.; Cheng, X.; Zhao, H.; Yang, Q.; Xu, Z. Downregulation of the Zinc Transporter SLC39A13 (ZIP13) Is Responsible for the Activation of CaMKII at Reperfusion and Leads to Myocardial Ischemia/Reperfusion Injury in Mouse Hearts. J. Mol. Cell. Cardiol. 2021, 152, 69–79. [CrossRef]
- Weaver, B.; Zhang, Y.; Hiscox, S.; Guo, G.; Apte, U.; Taylor, K.; Sheline, C.; Wang, L.; Andrews, G. Zip4 (Slc39a4) Expression Is Activated in Hepatocellular Carcinomas and Functions to Repress Apoptosis, Enhance Cell Cycle and Increase Migration. PloS One 2010, 5. [CrossRef]
- Sun, S.; Xie, E.; Xu, S.; Ji, S.; Wang, S.; Shen, J.; Wang, R.; Shen, X.; Su, Y.; Song, Z.; et al. The Intestinal Transporter SLC30A1 Plays a Critical Role in Regulating Systemic Zinc Homeostasis. Adv. Sci. 2024, 11, 2406421. [CrossRef]
- Trial | NCT01062347 Available online: https://cdek.pharmacy.purdue.edu/trial/NCT01062347/ (accessed on 19 November 2025).
- Hara, T.; Yoshigai, E.; Ohashi, T.; Fukada, T. Zinc Transporters as Potential Therapeutic Targets: An Updated Review. J. Pharmacol. Sci. 2022, 148, 221–228. [CrossRef]
- Liu, L.; Yang, J.; Wang, C. Analysis of the Prognostic Significance of Solute Carrier (SLC) Family 39 Genes in Breast Cancer. Biosci. Rep. 2020, 40, BSR20200764. [CrossRef]
- Liu, Y.; Wei, L.; Zhu, Z.; Ren, S.; Jiang, H.; Huang, Y.; Sun, X.; Sui, X.; Jin, L.; Sun, X. Zinc Transporters Serve as Prognostic Predictors and Their Expression Correlates with Immune Cell Infiltration in Specific Cancer: A Pan-Cancer Analysis. J. Cancer 2024, 15, 939–954. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



