Submitted:
19 November 2025
Posted:
19 November 2025
You are already at the latest version
Abstract
This review elucidates the foundational principles of nicotinamide adenine dinucleotide (NAD+) homeostasis in humans, emphasizing its depletion during aging and in age-associated disorders. Subsequently, the discussion extends to NAD+ precursors and their prospective therapeutic applications, with insights derived from research utilizing zebrafish as a disease model. This information sheds light on the growing interest in NAD and its metabolism in the medical field, while also sparking curiosity among researchers focused on fish studies. The review further explores the role of nicotinamide in fish, encompassing core NAD+ metabolism, its participation in oxidative stress, environmental challenges, and the mitigation of pollutant-induced toxicity. Additionally, the implications of NAD+ in fish neurobiology, immune regulation, host-pathogen interactions, skin, eggs, and post-mortem muscle were considered. Dietary modulation of NAD+ pathways to enhance growth, immunity, and product quality in aquaculture has also been highlighted. This review highlights the significance of NAD+ metabolism in fish biology, covering cellular energy production, physiological processes, and environmental adaptation, and proposes targeting NAD+-related pathways as a strategy for aquaculture and fish health management.
Keywords:
NAD+
; metabolism
; NAD+ boosters
; sirtuins
; homeostasis
; fish
; physiology
1. Introduction
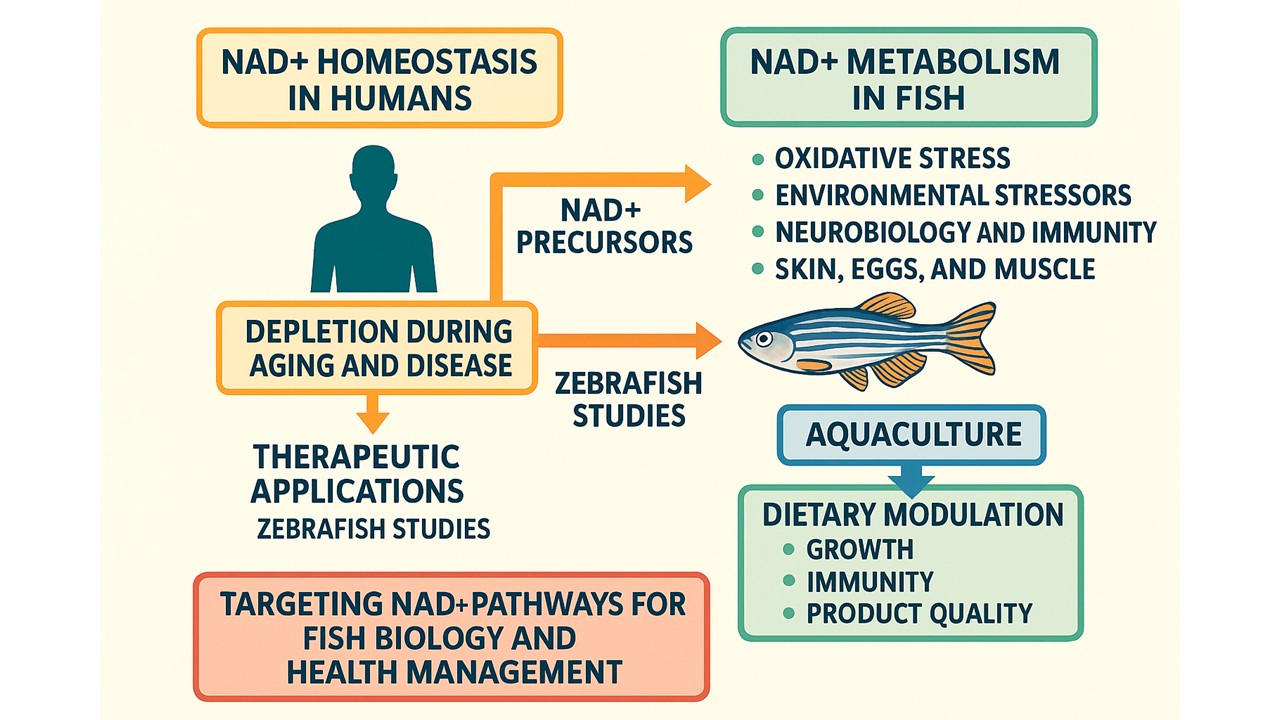
Nicotinamide adenine dinucleotide (NAD) was initially identified as a cofactor in yeast fermentation in 1906, originally termed 'cozymase.' Its structure, consisting of adenine, phosphate, and a reducing sugar, was elucidated in the 1930s, and its function as a hydride transfer agent was clarified in 1936. NAD+ research was conducted using the three Nobel Prizes. Interest in NAD+ significantly increased in the early 2000s following its identification as a co-substrate for sirtuins (SIRTs), which are essential for regulating longevity and metabolism. NAD+ and NADH are indispensable for electron exchange reactions, particularly those mediated by oxidoreductases, which involve hydride transfer. NAD+ functions as an electron acceptor, whereas NADH serves as an electron donor and plays a vital role in catabolic pathways, such as glycolysis, fatty acid β-oxidation, and the tricarboxylic acid cycle. Currently, there is renewed scientific interest in NAD+, owing to its recently discovered role in regulating metabolism and longevity in humans [1]. This article presents a narrative review delineating the fundamental concepts of NAD+ homeostasis in humans. It subsequently examines evidence implicating NAD+ depletion during the aging process and various age-related disorders. This body of knowledge has prompted investigations into NAD+ precursors, their potential therapeutic value, and the effects of NAD+ in disease models using zebrafish. Finally, this review discusses the current state of research concerning studies conducted on fish, elucidating the relationships between NAD+ and related molecules and their most significant functions in these animals. The ultimate objective was to clarify the key concepts, assess different NAD+ boosters in aquafeeds and their bioavailability, conduct comparative analyses to estimate potential requirements for each fish species, and explore optimal outcomes for the aquaculture industry.
2. Overview of NAD+ Biology And Its Balance in Human Health and Disease
The subcellular distribution of NAD+ and its biosynthetic enzymes vary across the cellular compartments. The nucleo-cytosolic NAD+ pool is considered to be interchangeable between cytosolic and nuclear pools, with similar concentrations in both [2]. The mitochondrial NAD+ pool, traditionally thought to be separated from the nucleo-cytosolic pool, may involve an unidentified mammalian mitochondrial NAD+ transporter [3]. Nicotinamide mononucleotide adenylyl transferase (NMNAT) is a non-histone chromatin-associated protein with distinct properties. Studies have shown that NMNAT is distributed in the nucleus with specific binding affinities different from those of histones, suggesting its role in chromatin structure and nuclear processes, such as DNA repair [4]. Now it is known that different NMNAT isoforms exhibit distinct subcellular localizations: NMNAT1 is in the nucleus, NMNAT2 in the cytosol and Golgi, and NMNAT3 in mitochondria [5]. Consequently, variations in the subcellular distribution of NAD+ across tissues with distinct metabolic functions and requirements may be substantial. The allocation of NAD+ and its biosynthetic components within cellular compartments facilitates the regulation of NAD+-dependent processes in various cellular regions and tissues [1].
NAD can be synthesized via de novo and salvage pathways. De novo NAD+ synthesis occurs in the cytosol, where all enzymes are localized [2]. De novo synthesis begins with dietary tryptophan (Trp), whereas salvage pathways use vitamin B3 molecules such as nicotinic acid (NA), nicotinamide (NAM), and nicotinamide riboside (NR) from the diet for NAD production in tissues. Both pathways could benefit aquaculture because fish diets can be supplemented to enhance NAD synthesis; however, these studies are still in their infancy. A recent review showed that the nicotinamide phosphoribosyltransferase (NAMPT)-driven NAD+ salvage pathway supports muscle health by maintaining mitochondrial function; reducing oxidative stress and inflammation; and promoting autophagy, muscle stem cell function, and neuromuscular junction integrity in aging and diseases [6]. These factors demonstrate the importance of NAD+ metabolism regulation through salvage pathway activation in combating metabolic, mitochondrial, neurotoxic, and muscle aging dysfunctions [6].
The main signaling pathways that consume NAD+ include SIRTs, poly (ADP-ribose) polymerases (PARPs), and cyclic ADP-ribose synthases (cADPRSs). SIRTs are conserved NAD+-dependent deacetylases; therefore, their functions are intrinsically linked to cellular metabolism [7]. Localization varies across cellular compartments, potentially enabling compartment-specific regulation of NAD+ pools [8]. PARPs consume NAD+ and are involved in DNA repair and other cellular functions, whereas cADPRSs, specifically, CD38 are examples of enzymes that cleave NAD+ to generate secondary messengers involved in calcium signaling. These enzymes share the common property of irreversibly cleaving NAD+ into NAM and ADP-ribose moieties. They act as metabolic sensors and significantly influence organ metabolism, function, and aging [9].
Energy status influences NAD+ homeostasis in cells and organisms. Conditions characterized by limited energy availability, such as caloric restriction, fasting, and physical exercise, have been shown to elevate NAD+ levels. In contrast, excessive energy consumption, particularly through diets high in fat or a combination of high fat and sucrose, results in NAD+ depletion within metabolic organs [10]. Circadian rhythms affect NAD+ levels and exhibit diurnal fluctuations in the liver. The circadian clock regulates NAD+ biosynthetic enzymes (e.g., NAMPT), whereas NAD+ consumers such as PARP-1, SIRT1, SIRT6, and SIRT3 regulate the circadian clock [11]. Mechanistic synthesis of the NAD+-sirtuin axis in circadian rhythm and metabolic regulation elucidates how NAD+ levels oscillate in a circadian manner through the rhythmic expression of biosynthetic enzymes, such as NAMPT, whereas sirtuins, such as SIRT1 and SIRT6, modulate clock components and circadian-controlled metabolic genes. Therapeutic tactics, including chronopharmacology, NAD⁺ boosters, and SIRTs’ modulators to restore circadian synchronization and improve age-related metabolic and neurodegenerative pathologies via the NAD⁺–sirtuin–clock network have been proposed [12].
Strategies to enhance NAD+ synthesis include supplementation with NAD+ precursors (e.g., NAM, NA, NR, and NMN), stimulation of NAD+ synthesis enzymes (such as NAMPT), and activation of NAD(P)H-quinone oxidoreductase 1 (NQO1) [13,14]. To prevent NAD+ depletion, it is also possible to inhibit enzymes that consume NAD+ (e.g., SIRTs, PARP-1, and CD38) [15]. Additionally, the regulation of metabolic pathways is crucial, as it can divert metabolites from NAD+ production, and their inhibition may result in elevated NAD+ levels. For instance, enzymes, such as nicotinamide N-methyltransferase (NNMT), facilitate the methylation of nicotinamide (NAM) by utilizing S-adenosyl methionine (SAM) as a methyl donor, resulting in the production of 1-methylnicotinamide (MNAM) and S-adenosylhomocysteine [16]. Understanding these factors is crucial for formulating strategies to sustain NAD+ levels, which may be beneficial in age-related and metabolic diseases.
2.1. Reduction in NAD+ Levels Is Associated with Aging and Numerous Age-Related Diseases.
NAD+ depletion is associated with aging and age-related disorders that affect physiological systems. Prior research in mammals has demonstrated that DNA damage resulting from aberrant nutritional status intensifies cellular NAD+ consumption. Consequently, a reduction in NAD+ levels leads to oxidative stress and contributes to pathological processes underlying metabolic diseases [17]. NAD+ depletion is linked to neurodegenerative disorders (including Alzheimer's disease, Parkinson's disease, amyotrophic lateral sclerosis, and prion diseases), all of which are characterized by protein misfolding and proteotoxic stress [18]. In addition, alcoholic and non-alcoholic fatty liver diseases are also associated with decreased NAD+ levels, as well as different cardiovascular diseases (including cardiac ischemia, cardiomyopathies, and cardiac hypertrophy) [19] and muscular disorders (e.g., muscular dystrophies, mitochondrial myopathies, and age-related sarcopenia). As a result, enhanced NAD+ levels have shown promise in preserving muscle function in animal models under these conditions [20] and kidney disorders (e.g., acute kidney injury, chronic kidney disease, or diabetic nephropathy), characterized by impaired mitochondrial function and diminished SIRT signaling [21]. In addition, maintenance of hepatic NAD+ content has shown protective effects against hepatic lipid accumulation and liver damage in various animal models [22]. Furthermore, a wide spectrum of metabolic disorders, such as obesity and type 2 diabetes, are associated with altered NAD+ homeostasis in various tissues. The restoration of NAD+ levels, which has been shown to enhance mitochondrial function and confer protection in animal models, represents a promising therapeutic avenue for these conditions.
2.2. Potential Therapeutic Value of NAD+ Precursors
Owing to their significant bioactivity, directly supplying animals with exogenous NAD+ is challenging [17,23]. As mentioned in this review, vitamin B3, also known as NA and NAM, and derivatives like NR and NMN are NAD+ precursor vitamins [24,25]. The administration of NAD+ donors can elevate NAD+ concentrations within cells, thereby ameliorating metabolic dysfunction. Research indicates that these NAD+ precursors exert distinct physiological effects because of their unique characteristics and effectiveness in enhancing NAD+ levels in mammalian cells [17,26]. For instance, oral administration of NR enhances hepatic NAD+ levels in mice more effectively than NA or NAM [27]. A recent review has evaluated NMN and NR as NAD⁺-boosting precursors. Both compounds increase NAD⁺ levels and are beneficial for aging- and metabolism-related health. Preclinical findings suggest that NR may be more efficient at increasing NAD⁺ levels, since NMN requires extracellular conversion to NR before cellular uptake. Animal studies have shown that NMN has superior effects in specific contexts, suggesting tissue-specific advantages [28]. These findings indicate the potential advantages of NAD+ enhancement in addressing age-related and metabolic disorders in humans. Although preclinical research has shown promise, clinical evidence is still in its early stages of development. Most human studies conducted to date have been short-term, spanning weeks to months. There is a notable lack of data regarding long-term NAD+ supplementation in humans. Consequently, further research involving long-term clinical trials with larger cohorts is necessary to fully comprehend the therapeutic potential of NAD+ [1,29]. Studies have shown that NAD+ precursors such as NR are well tolerated by humans over short periods [30]. Research indicates that the effects of NAD+ supplementation vary, with individual differences influenced by factors such as age, health status, and metabolic conditions. Therefore, caution must be exercised in this context [1]. Rigorous clinical trials are essential for assessing the effects and risks associated with NAD+ supplementation in humans. Further research is required to identify potential adverse effects and confirm the safety of long-term supplementation. These considerations underscore the importance of clinical trials in mitigating the risks associated with the unregulated use of NAD+ boosters.
Promising results from NAD+ supplementation suggest its potential application in oncology and anti-aging therapies. A promising treatment strategy for radioprotection, with potential applications in oncology, highlights the effectiveness of administering NR alongside other components without diminishing the efficacy of radiotherapy in tumor xenograft models. Combinatorial treatment with polyphenols, pterostilbene, and silibinin, along with NR and Toll-like receptor 2/6 (TLR2/6) ligand (FSL-1), provides radioprotective effects in mice exposed to lethal γ-radiation. While polyphenols alone ensured short-term survival (30 days), only the complete combination conferred long-term protection, with 90% survival at one-year post-irradiation. The protective mechanisms involve nuclear factor-erythroid 2 related factor 2 (Nrf2)-mediated antioxidant responses, DNA repair through PARP1, suppression of nuclear factor kappa-light-chain-enhancer of activated B cell (NF-κB) inflammation, mitochondrial stabilization via peroxisome proliferator-activated receptor gamma coactivator-1 alpha (PGC-1α)/SIRT1/SIRT3, and accelerated hematopoietic recovery [31]. However, the geroscience hypothesis posits that addressing the core elements of aging could prevent age-related diseases and prolong healthy life. Research has explored interventions including senolytics, NAD+ enhancers, and metformin. NAD+ enhancement using NMN and NR precursors increases the health span of model organisms, although human results vary [32]. Clinical trials in older adults and obese individuals have demonstrated its safety and modest improvements in insulin sensitivity and aerobic capacity. Although NMN and NR are promising NAD⁺ precursors, NR shows high bioavailability [33]. Current research has indicated that these agents have the potential to improve health. Nevertheless, further studies are required to conduct clinical comparisons and ascertain optimal dosages, benefits, and safety profiles.
3. Zebrafish (Danio rerio) Models for Exploring NAD+-Related Pathways in Humans
Given the conservation of metabolic pathways across animal species, research on lower animals provides insights into human metabolic diseases [34]. Studies have shown similar pathophysiological pathways in metabolic diseases in mammalian and fish models [35,36,37]. In addition, fish are reliable and cost-effective experimental alternatives to mammals [38,39]. Therefore, the function of NAD+ in disease models has been extensively investigated using zebrafish as a model organism. This review chronologically presents available studies and their principal findings, showing the progress of research in this field.
Nrk2b, a nicotinamide riboside kinase in zebrafish muscle, is essential for muscle morphogenesis by regulating NAD⁺-dependent cell–matrix adhesion at the myotendinous junction (MTJ). Nrk2b-mediated NAD⁺ biosynthesis enables laminin polymerization. In Nrk2b-deficient embryos, muscle fibers extend beyond the somite boundaries. NAD⁺ rescues MTJ morphology in Nrk2b morphants but not in laminin mutants. Both Nrk2b and laminin control paxillin localization to adhesion complexes. Paxillin overexpression restores MTJ integrity in Nrk2b-deficient embryos, revealing an essential Nrk2b → NAD⁺ → laminin adhesion → paxillin localization pathway for muscle boundary formation [40]. Increasing NAD⁺ levels via supplementation or Nrk2b-mediated biosynthesis improves muscular dystrophy in zebrafish lacking laminin-binding complexes. Boosting NAD⁺ restores the basement membrane organization through laminin polymerization, thereby enhancing locomotor performance. Paxillin overexpression partially rescued muscle structure but not motility. Integrin α6 has been identified as a noncanonical laminin receptor, revealing an Nrk2b → NAD⁺ → laminin assembly → integrin α6/paxillin pathway that enhances muscle–extracellular matrix resilience, suggesting a therapeutic tool for muscular dystrophy [41]. This research team also showed that NAD⁺ supplementation improved muscular dystrophy pathology in a zebrafish model of Duchenne muscular dystrophy (DMD) by restoring NAD⁺ homeostasis and activating repair mechanisms. This study linked muscle damage to NAD⁺ depletion, which worsened mitochondrial dysfunction and impaired SIRT1-mediated stress responses. Nicotinamide administration enhanced SIRT1 activation, improved mitochondrial function, and stabilized muscle membranes. These findings identified NAD⁺ biosynthesis as a therapeutic target for DMD, highlighting its role in muscle regeneration through metabolic-epigenetic crosstalk [42].
Numerous studies have focused on mitigating the adverse effects associated with various diseases through restoration of NAD+ levels using different methodologies. Resveratrol reduces hepatic lipid accumulation and improves lipid profiles by upregulating fatty acid oxidation genes and suppressing lipogenic markers. SIRT1 activation enhances mitochondrial function and insulin sensitivity, demonstrating that the NAD+–sirtuin pathway plays a role in lipid regulation. This study demonstrated that resveratrol effectively counteracts diet-induced dysregulation of lipid metabolism in zebrafish by activating NAD+-dependent SIRT1 signaling [43]. Chronic ethanol exposure induces hepatic steatosis and inflammation in zebrafish, mirroring alcoholic liver disease in mammals. Ethanol-treated zebrafish showed lipid accumulation, oxidative stress, and increased levels of pro-inflammatory markers, with evidence of liver injury. Metabolic and inflammatory dysregulation aligns with ethanol-induced suppression of NAD+-dependent SIRT1 activity, which regulates lipid metabolism and inflammation. These findings establish zebrafish as a model for alcoholic liver disease research, demonstrating conserved mechanisms by which ethanol disrupts NAD+ homeostasis, leading to metabolic-inflammatory effects, and enabling the investigation of NAD+-targeting therapies [44]. In another study, adult zebrafish exposed to 0.5% ethanol for 24-72 hours showed elevated serum alanine aminotransferase (ALT) levels at 48 h and 72 h, with increased hepatic expression of proinflammatory and lipogenic genes, indicating liver injury. Drug trials showed that nicotinamide riboside TES1025, (an inhibitor of amino-β-carboxymuconate-ε-semialdehyde decarboxylase, which is a key enzyme in the tryptophan-kynurenine-NAD+ metabolic pathway that elevates NAD+) and riboflavin suppressed ethanol-induced ALT elevation and inflammatory gene expression. The authors reproduced an ethanol-induced hepatic injury model, revealing mechanisms such as oxidative stress via reactive oxygen species (ROS) and lipid peroxidation, NF-κB-mediated inflammation, and steatosis. Ethanol metabolism impairs antioxidant defenses. The validity of this model was confirmed when N-acetylcysteine and silymarin attenuated injury by suppressing ROS/NF-κB and restoring AMP-activated protein kinase (AMPK)-mediated lipid metabolism. This validates adult zebrafish as a platform for evaluating hepatoprotective agents against ethanol toxicity, showing that NAD⁺ boosting and antioxidant supplementation can mitigate hepatic damage in vivo [45].
NAMPT, the rate-limiting enzyme in NAD⁺ salvage, drives PARP1 hyperactivation and parthanato cell death, thereby promoting inflammation in zebrafish and in human skin models. Inhibition of NAMPT, PARP1, and NADPH oxidases and overexpression of PAR-glycohydrolase reduced oxidative stress, DNA damage, and cell death. NAD⁺ supplementation reversed these protective effects, confirming that NAMPT-NAD+ is associated with the inflammatory pathology. This study found elevated NAMPT and PARP1 activities with apoptosis-inducing factor mitochondria associated 1 (AIM1) nuclear translocation in psoriatic skin, highlighting this pathway as a therapeutic target [46].
Another interesting study demonstrated that propionate induces intestinal oxidative stress in zebrafish through the NAD+-dependent SIRT3-regulated propionylation of mitochondrial superoxide dismutase (SOD2). Propionate exposure causes metabolic dysregulation through impaired NAD+/SIRT3 signaling, leading to SOD2 hyperpropionylation and inactivation. This mechanism reveals how fatty acids can induce oxidative stress despite their beneficial effects on gut health [47]. Further research is required to elucidate the established connections between microbial metabolites, post-translational modifications, and redox homeostasis in vertebrates. In addition, a dietary formulation of NMN with astaxanthin (a potent antioxidant) and blood orange extract (NOA) has been studied in aging zebrafish. NOA supplement was converted to NAD+ in vivo with superior bioavailability. NOA-treated fish showed reduced age-related decline, enhanced activity, improved sleep, and better skin health, with increased ATP synthesis. These findings show that NOA offers enhanced NAD+ augmentation and anti-aging benefits in zebrafish [48].
Exposure of zebrafish larvae to decabromodiphenyl ethane (DBDPE) induces insulin resistance, lipid accumulation, and neurotoxicity through acetylcholinesterase inhibition and apoptosis by disrupting mitochondrial function through reduced oxidative phosphorylation, respiratory activities, membrane potential, and ATP production. These negative effects were reversed by NR [49]. Aspartame exposure causes developmental defects in zebrafish embryos by disrupting NAD+-dependent SIRT1/FOXO3a (Forkhead Box O3a) signaling in neurons. Aspartame reduces NAD+ levels, impairing SIRT1-mediated FOXO3a deacetylation and leading to oxidative stress and neuronal apoptosis. These findings revealed the role of the NAD+/SIRT1/FOXO3a pathway in neurodevelopment and demonstrated how artificial sweeteners affect metabolic pathways during development. These results further suggest that maintaining NAD+ homeostasis may mitigate aspartame-induced neurotoxicity, thereby affecting vertebrate development [50]. Recently, a zebrafish model was established for Congenital NAD⁺ deficiency disorders, showing developmental anomalies similar to human congenital NAD deficiency disorder (CNDD) and vertebral-anal-cardiac-tracheoesophageal fistula-renal-limb (VACTERL) association syndrome. 2 amino1,3,4 thiadiazole (ATDA)-induced neural tube defects, craniofacial malformations, and cardiac abnormalities were rescued by NAM supplementation, confirming the role of NAD⁺ depletion. This demonstrates the utility of zebrafish as a model to study NAD⁺-dependent malformations and investigate CNDD and VACTERL phenotypes in vivo [51].
Perfluorooctanoic acid (PFOA), widely used to manufacture products such as Gore-Tex and Teflon, induces premature ovarian insufficiency in adult female zebrafish, reducing spawning, eggs, and gonadal index, while increasing embryonic mortality. PFOA suppresses NAD⁺ biosynthesis and impairs mitochondrial integrity in oocytes, resulting in disorganized mitochondria and a decreased membrane potential. NMN supplementation alleviates reproductive defects by restoring NAD+ levels. These findings suggest that NMN is a potential therapeutic agent for chemical disorders caused by zebrafish gonads [52]. A recent study on a phosphoribosylpyrophosphate 1 (PRPS1) variant in X-linked hearing loss showed a connection to NAD+ homeostasis. PRPS1 catalyzes purine nucleotide synthesis, generating the PRPP required for purine synthesis and NAD+ salvage pathway. The Ser115Gly variant impairs GTP/ATP production in zebrafish, potentially disrupting NAD+ levels by reducing GTP availability for NAD+ synthesis and compromising NAD+-dependent processes through ATP depletion [53].
Studies have also focused on emulating diseases affecting the nervous system. NR significantly prolonged survival and improved motor function in a zebrafish model of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced Parkinson's disease. Transcriptomic and metabolomic analyses revealed that NR modulates glucose metabolism by downregulating gluconeogenic enzymes and upregulating glycolytic enzymes, while attenuating endoplasmic reticulum stress. These effects underpin the observed neurobehavioral improvements. The study showed that NR exerts neuroprotective effects in the MPTP zebrafish model by enhancing energy metabolism and mitigating endoplasmic reticulum (ER) stress, providing mechanistic insights into potential therapeutic approaches for Parkinson's [54]. Zebrafish models of TANK-binding kinase 1 (TBK1, a serine/threonine kinase that is constitutively expressed and involved in various cellular processes, including innate immunity and autophagy) deficiency showed that TBK1 loss causes motor neuron degeneration and impaired swimming. Metabolomics revealed that TBK1 disruption leads to NAM pathway dysregulation and NAD⁺ depletion, with NR supplementation rescuing the motor function. Proteomic analysis showed elevated inflammatory markers and necroptosis, the inhibition of which improved survival [55].
These studies underscore the usefulness of zebrafish as a model for proposed diseases, as well as the fact that gaining a deeper understanding of NAD+ metabolism in vertebrates in general, and in species of interest in particular, can help us better understand the most common pathologies affecting these organisms. Moreover, it would be interesting to learn how to increase NAD+ reserves, which seem to be consistently depleted or exhausted during these pathological processes.
4. Nicotinamide and Related Metabolites in Fish
This section provides an in-depth analysis of NAD+ metabolism and its complex roles in fish physiology, health, and adaptation to environmental change. These findings highlight the diverse functions of NAD+ and emphasize the crucial importance of its related pathways in various biological processes in fish (Figure 1). The results are systematically organized for clarity, following a chronological order where applicable and transitioning from molecular mechanisms to ecological and industrial applications.
4.1. Core NAD⁺ Metabolism in Fish Nutrition
As previously discussed, NAD+ is crucial not only as a redox cofactor in metabolic processes but also as a substrate in stress-responsive signaling pathways. Under stressful conditions, NAD+ synthesis diminishes as NAD+ consumption increases, leading to a reduction in the cellular levels. The administration of dietary precursors to enhance biosynthesis has been proposed as a potential therapeutic strategy to counteract physiological decline and prevent the onset of pathologies. Consequently, numerous fish studies have focused on these subjects. For example, the enzymatic activities of hepatic NAD- and NADP-isocitrate dehydrogenase (NADP-ICD) in carp (Cyprinus carpio) and tilapia (Oreochromis niloticus) have been investigated under various dietary conditions. Fish that consumed carbohydrate-rich diets and exhibited increased feeding rates demonstrated elevated levels of soluble NADP-ICD, presumably due to enhanced lipogenesis. In contrast, enzymatic activity was diminished in fish subjected to starvation or a lipid-rich diet. Notably, mitochondrial NADP-ICD activity remains constant irrespective of the dietary regimen [56].
Numerous studies have focused on NAD+ metabolism from a nutritional perspective, specifically by examining SIRTs and NAMPT. SIRTs are NAD+-dependent histone deacetylases that regulate important metabolic pathways in prokaryotes and eukaryotes and are involved in many biological processes, such as cell survival, senescence, proliferation, apoptosis, DNA repair, cell metabolism, and caloric restriction. The mammalian SIRT family of evolutionarily conserved proteins belonging to class III histone deacetylases (HDACs) comprises seven members [57]. These enzymes have been studied in several fish species. For example, a thorough spatiotemporal examination of SIRT expression patterns throughout the aging process in short-lived annual turquoise killifish (Nothobranchius furzeri), a significant model for aging studies, has been conducted. These results indicate distinct tissue-specific and age-related regulation of sirtuin isoforms (SIRT1-7), with significant reduction in NAD+-dependent sirtuins (SIRT1, SIRT3, and SIRT6) in metabolically active tissues as aging progresses. The consistent link between SIRT downregulation and aging suggests an evolutionarily conserved function of the NAD+-sirtuin axis in vertebrate aging. These findings suggest that N. furzeri is an effective model for investigating the temporal dynamics of SIRT expression and its functional roles in aging while emphasizing the potential significance of sustaining NAD+ bioavailability for healthy aging [58]. A reliable quantitative PCR (qPCR) reference gene framework has been established for microRNA (miRNA) analysis in Wucham bream (Megalobrama amblycephala), enabling the exploration of the miR-34a/Sirt1 regulatory axis in energy metabolism. This study showed that miR-34a targets Sirt1 mRNA, inhibits the NAD+-dependent Sirt1 pathway, and disrupts lipid and glucose homeostasis in hepatic tissues. The use of stable reference genes, such as 5S rRNA, enabled the precise quantification of miR-34a upregulation during metabolic stress, showing an inverse correlation with Sirt1 expression. These findings reveal that miR-34a-mediated inhibition of Sirt1 compromises mitochondrial function and promotes metabolic dysfunction, highlighting the role of the NAD+/Sirt1 axis in energy regulation. This study advances both miRNA methodologies in aquaculture species and the understanding of SIRT pathway regulation [59].
Epigenetic regulation of muscle SIRT1 expression was explored in gilthead seabream (Sparus aurata), revealing how local DNA methylation dynamically influences SIRT1 transcription in response to seasonal changes and aging. These findings show that decreased promoter methylation is associated with increased SIRT1 expression during high metabolic demand, whereas age-related hypermethylation leads to transcriptional decline, linking epigenetics to NAD+-dependent metabolic adaptation. These results reveal how environmental cues epigenetically regulate SIRT1 expression for muscle homeostasis, demonstrating a conserved mechanism that integrates epigenetic and SIRT-mediated metabolic control in vertebrates [60]. In the same fish species, an expanded repertoire of SIRT genes includes three Sirt3 copies (sirt3.1a, sirt3.1b, sirt3.2) and two Sirt5 copies (sirt5a, sirt5b), in addition to seven canonical paralogs (SIRT1-7), shaped by vertebrate 2R and teleost-specific 3R duplications. While sirt3.1 and sirt5a are mainly expressed in skeletal muscle, sirt3.2 and sirt5b show higher expression in immune tissues and gills, suggesting tissue-specific adaptation [61]. Similarly, the first comprehensive genome-wide analysis of the SIRT gene family in Nile tilapia revealed the conserved and teleost-specific features of these NAD+-dependent proteins. Seven SIRT orthologs (SIRT1-7) were identified, showing distinct tissue expression patterns were identified. Sirt1 and sirt3 are highly expressed in metabolically active tissues such as the liver and muscle. Fasting increased sirt1 and sirt5 expression in the liver, indicating their role in NAD+-mediated nutrient stress adaptation, whereas sirt2 responded to oxidative stress in brain tissue. This study also identified teleost-specific gene duplications (sirt3 and sirt5), suggesting adaptation to aquatic environments. These findings help to understand how NAD+-sirtuin signaling networks regulate metabolism and stress responses in this important aquaculture species [62].
Several studies have focused on NAMPT (also known as pre-B-cell colony-enhancing factor or visfatin), a rate-limiting enzyme in the NAD⁺ salvage pathway. NAMPT plays a vital role in maintaining intestinal integrity and immune competence in hybrid crucian carp [originated from White crucian carp (Carassius cuvieri, WCC, female) × Red crucian carp (C. auratus red var., RCC, male), [63]. NAMPT sustains intracellular NAD+ levels by transforming nicotinamide into NMN, supporting energy metabolism, redox balance, and NAD⁺-dependent enzymes such as SIRTs and PARPs. Research has shown that increased Nampt expression improves barrier function and bacterial resistance, as observed in hybrid crucian carp specimens injected with Aeromonas hydrophila. These findings underscore the pivotal role of NAD⁺ metabolism in fish health and immune defense. Higher Nampt levels lead to a marked increase in the number of goblet cells in the distal intestine. Additionally, Nampt significantly increased the expression of antimicrobial molecules, such as interleukin 22 (IL-22), hepcidin-1 (an antimicrobial peptide), liver-expressed antimicrobial peptide 2 (LEAP-2), and mucin 2 (MUC2), as well as tight junction proteins, such as zonule 1 and occludin. In line with these observations, fish treated with NAMPT showed a significant decrease in intestinal permeability and apoptosis, thereby strengthening host defense against bacterial infections [64]. In addition, we investigated the role of NAD+ in feeding regulation in goldfish (C. auratus). The results demonstrated that NAMPT influenced appetite-regulating neuropeptides, suggesting that NAD⁺ metabolism is not only central to energy production and redox control but is also tightly integrated with neuroendocrine circuits controlling feeding behavior in fish [65]. Some studies have focused on appetite and its possible regulation, leading to a better understanding of feeding behavior, which is of great interest in aquaculture. Unfortunately, very little attention has been paid to studies investigating their effects on NAD+ metabolism. For example, ceramides have been shown to suppress food intake in the rainbow trout (Oncorhynchus mykiss) by modulating appetite-regulating neuropeptides. Although NAD+ has not been directly studied, emerging evidence suggests mechanistic links: ceramides impair mitochondrial function and increase oxidative stress, depleting NAD+ levels, which are cofactors of SIRT1, and regulating metabolism and feeding. NAD+ decline reduces SIRT1 activity, amplifying ceramide-induced anorexia by disrupting neuropeptide balance. Ceramide-driven inflammation activates NAD+-consuming enzymes, worsening NAD+ depletion and energy deficits [66]. Although unexplored, ceramide-NAD+ interactions may drive metabolic dysregulation in fish, similar to that observed in mammals. Future studies should test whether NAD+ restoration reduces the anorexigenic effects of ceramide. Another study on transcription factors, such as SIRT1, in goldfish suggested a link between NAD+-dependent metabolic sensing and feeding behavior. Findings show that fasting and re-feeding significantly affect hypothalamic expression, with SIRT1 being strongly associated with orexigenic neuropeptides (e.g., neuropeptide Y). Although NAD+ levels were not measured, SIRT1 activation during fasting indicated its role as a nutrient sensor connecting energy status and appetite regulation, aligning with its NAD+-dependent function in other vertebrates [67]. These findings provide evolutionary insights into conserved feeding regulation mechanisms, suggesting that SIRT-mediated metabolic adaptation may connect the peripheral energy status with central appetite pathways in fish.
4.2. NAD+ and SIRTs as Molecular Hubs for Environmental Stress Adaptation in Fish
NAD+ is integral to the oxidative stress response and pathways related to environmental adaptation, which are crucial for fish to cope with various environmental stressors including pollutants, pesticides, and nanoparticles. These pathways are essential for maintaining the redox balance, energy metabolism, and cellular homeostasis under adverse conditions.
4.2.1. NAD+ Role in Stress Responses
There is evidence linking thermal physiology and cellular stress responses, highlighting the significance of NAD+/SIRT pathways in environmental adaptation. For example, seasonal acclimatization induces muscle plasticity in carp through epigenetic regulation of ribosomal RNA (rRNA) synthesis, involving NAD+-dependent sirtuin pathways as environmental sensors. Cold acclimation upregulated ribosomal biogenesis genes while altering DNA methylation at the rDNA loci, suggesting temperature-sensitive epigenetic reprogramming. The observed metabolic remodeling aligns with SIRT1's role in linking environmental cues to ribosomal synthesis. These findings demonstrate how fish optimize muscle function across seasons through epigenetic mechanisms, where sirtuin-mediated sensing may translate thermal signals into adaptive expressions [68]. Cold acclimation in sticklebacks (Gasterosteus aculeatus) triggers a stress response through the upregulation of heat shock proteins (HSP70 and HSP90) and NAD+-dependent SIRTs (SIRT1 and SIRT3), showing a conserved mechanism linking proteostasis and metabolic adaptation. These findings reveal tissue-specific responses and suggest that SIRT-mediated mitochondrial changes and HSP-facilitated protein stability enhance cold tolerance, providing insights into environmental adaptation through NAD+-sensitive pathways [69].
Another study examined the function of SIRT2 in the regulation of adipocyte maturation during hypoxic adaptation in fish, emphasizing its role as a NAD+-dependent regulatory mechanism. These findings indicate that hypoxia induces the differential expression of sirt2, which constrains adipocyte maturation by modulating lipid metabolism and cellular differentiation pathways. This adaptive response suggests that SIRT2 plays a pivotal role in metabolic reprogramming under low oxygen conditions, potentially preserving energy homeostasis. This study offers novel insights into how hypoxia influences lipid storage and metabolic flexibility in aquatic species through NAD+-dependent sirtuin signaling, providing broader implications for understanding vertebrate stress adaptation mechanisms [70].
It was also demonstrated that stress disrupts the hypothalamic circadian system and appetite regulation in rainbow trout through the coordinated actions of cortisol and NAD+-dependent SIRT1, thereby linking endocrine stress responses with metabolic regulation. Stress induced elevation of cortisol suppresses sirt1 expression in the hypothalamus, correlating with disruptions in circadian clock genes and altered orexigenic/anorexigenic peptide expression. These findings revealed a mechanism by which stress-mediated NAD+/SIRT1 suppression impairs circadian timing and energy homeostasis, suggesting that SIRT1 functions as a metabolic integrator of stress responses. The results showed that environmental challenges may dysregulate feeding behavior in fish through SIRT-mediated pathways, with implications for aquaculture welfare [71].
The physiological and molecular responses of roughskin sculpin (Trachidermus fasciatus) to osmotic stress were analyzed, revealing significant alterations in Na<sup>+</sup>/K<sup>+</sup>-ATPase activity, caspase 3/7 activity, and expression of stress-related genes (e.g., sirt1 and hsp70). These findings indicate that osmotic stress induces cellular stress and apoptosis while modulating SIRT1 expression, suggesting a role for NAD+-dependent SIRT signaling in adaptation. The upregulation of sirt1 and hsp70 implies activation of stress response pathways involving SIRT1-mediated regulation. These results provide insights into osmoregulatory mechanisms in euryhaline fish, highlighting the connections between ion transport, cellular stress, and NAD+-sensitive responses [72]. The molecular characterization and stress-responsive expression of sirt2, sirt3, and sirt5 in Wuchang bream were analyzed under temperature and ammonia nitrogen stress. These findings reveal that SIRTs play crucial roles in metabolic regulation and stress adaptation. Under thermal and ammonia-induced stress, SIRT2, SIRT3, and SIRT5 showed tissue-specific expression patterns, suggesting their roles in cellular homeostasis and mitochondrial function. This study highlights NAD+-dependent sirtuin activity in stress responses, providing insights into stress tolerance mechanisms in fish [73].
A novel dual regulatory mechanism has been identified in grass carp (Ctenopharyngodon idella), wherein SIRT1 modulates p53-mediated apoptosis through both KAT8-dependent and-independent pathways, underscoring the functional complexity of NAD+-dependent deacetylation in fish. SIRT1 suppresses apoptosis by deacetylating p53 at K382 (KAT8-dependent) and inhibiting p53 transcriptional activity through KAT8-independent interactions, revealing an evolutionary innovation in teleost stress response. These NAD+-sensitive mechanisms enhance cell survival under oxidative stress, providing the first evidence that fish sirtuins regulate apoptotic thresholds via multiple p53-targeting strategies. The results showed that SIRT1 metabolic sensing via NAD+ integrates conserved and lineage-specific anti-apoptotic mechanisms [74].
In summary, these studies underscore the role of SIRTs as pivotal molecular hubs in environmental adaptation, effectively linking metabolic and stress response networks in fishes. SIRTs function as metabolic stress sensors, connecting cellular energy status to adaptive responses, including mitochondrial modifications, HSP induction, and metabolic reprogramming (Table 1). Although certain pathways are evolutionarily conserved, teleosts demonstrate unique adaptations. The integration of SIRTs with other stress pathways is evident, as evidenced by their interactions with cortisol, circadian clock, apoptosis regulators, and metabolic enzymes to maintain homeostasis under stress. A comprehensive understanding of these mechanisms could enhance fish welfare and facilitate the prediction of adaptive capacities.

Table 1.
Role of NAD+/SIRT pathways in fish stress and environmental adaptation.
| Species / Model | Stressor / Condition | Sirtuins (SIRT) involved | Main findings | References |
|---|---|---|---|---|
| Carp (Cyprinus carpio) |
Seasonal acclimatization (thermal cycles) | SIRT1 | Cold acclimation induced upregulation of rRNA biogenesis genes and DNA methylation changes at rDNA loci; SIRT1 linked environmental sensing to ribosomal synthesis | [68] |
| Stickleback (Gasterosteus aculeatus) | Cold acclimation | SIRT1, SIRT3 | Upregulation of heat shock proteins (HSP) HSP70/HSP90 and NAD+-dependent SIRTs; mitochondrial adaptation and proteostasis maintenance | [69] |
| Fish adipocytes (in vitro model) |
Hypoxia | SIRT2 | Hypoxia induced differential sirt2 expression; constrained adipocyte maturation and lipid metabolism | [70] |
| Rainbow trout (Oncorhynchus mykiss) | Stress (cortisol elevation) |
SIRT1 | Stress-induced cortisol suppressed sirt1 expression in hypothalamus; disrupted circadian clock and appetite-regulating peptides | [71] |
| Roughskin sculpin (Trachidermus fasciatus) | Osmotic stress | SIRT1 | Altered Na⁺/K⁺-ATPase, caspase 3/7, and stress-related genes (sirt1, hsp70), apoptosis and cellular stress regulation | [72] |
| Wuchang bream (Megalobrama amblycephala) | Temperature and ammonia stress | SIRT2, SIRT3, SIRT5 | Tissue-specific expression changes under stress, regulating mitochondrial function and metabolic homeostasis | [73] |
| Grass carp (Ctenopharyngodon idella) | Oxidative stress | SIRT1 | Dual regulation of p53 apoptosis: KAT8-dependent deacetylation (p53 K382) and KAT8-independent suppression of p53 transcription | [74] |
4.2.2. Disruption of NAD+-Dependent Pathways in Aquatic Organisms Exposed to Environmental Contaminants
Numerous studies have focused on the role and modulation of metabolic pathways involving NAD+ in animals exposed to various contaminants ( Table 2). A previous study investigated the biochemical responses of the wild chub (Leuciscus cephalus) to environmental pollutants, showing oxidative stress and metabolic enzyme changes. Alterations in the biochemical markers of aquatic pollution effects in chub liver tissues from various river sites were characterized by different pollution types and levels. These findings were compared to the concentrations of organochlorine compounds, polycyclic aromatic hydrocarbons, and heavy metals. Enzymes associated with NAD⁺ metabolism and redox balance are affected, suggesting that NAD⁺-dependent pathways are critical for the response of fish to pollutant stress. The disruption of NAD⁺ metabolism highlights the importance of maintaining the cellular energy balance when exposed to environmental toxicants [75].
Polychlorinated biphenyls (PCB) are a family of synthetic compounds that are widely used in various industrial applications (such as refrigerants and lubricants in electrical equipment) but are now banned because of their persistence in the environment and toxicity. The effects of PCB 153 exposure on the brain proteome of Atlantic cod (Gadus morhua) revealed significant alterations in proteins involved in energy metabolism, oxidative stress response, and disruptions in NAD⁺-dependent pathways. These findings suggest that PCB 153 exposure impairs NAD⁺-linked metabolic functions in the brain, which could contribute to neurotoxicity and compromise neuronal health in fish [76]. Bifenthrin is a pyrethroid insecticide that is widely used to treat ant infestations. Bifenthrin toxicity and endocrine disruption have been studied in the rainbow trout. Bifenthrin exposure also induces oxidative stress, disrupting the NAD+/NADH balance and impacting energy production and redox homeostasis. NAD-dependent enzymes involved in DNA repair and hormone biosynthesis may be affected by pesticide-induced endocrine disruptions. In addition, salinity acclimation influences metabolic demands and stress responses, potentially altering NAD metabolism and fish sensitivity to bifenthrin [77].
Table 2.
Disruption of NAD⁺-dependent pathways in fish exposed to contaminants.
| Fish species / Model | Contaminant / Stressor | Main affected pathway | Main findings | References |
|---|---|---|---|---|
| Chub (Leuciscus cephalus) |
Environmental pollutants (organochlorines, PAHs, heavy metals) | NAD⁺ metabolism / redox enzymes | Oxidative stress, metabolic enzyme disruption, pollutant-type dependent responses | [75] |
| Atlantic cod (Gadus morhua) |
PCB 153 (Polychlorinated biphenyl 153) |
NAD⁺-linked energy metabolism | Brain proteome alterations, neurotoxicity risk | [76] |
| Rainbow trout (Oncorhynchus mykiss) | Bifenthrin (pesticide) | NAD⁺/NADH balance | Oxidative stress, endocrine disruption, salinity interaction | [77] |
| Rainbow trout (O. mykiss) |
Molybdo-flavoenzymes (AOX, XOR) | NAD⁺-dependent oxidoreduction | XOR exclusively NAD⁺-dependent, detoxification role | [79] |
| Crucian carp (Carassius carassius) | 17α-ethinylestradiol (EE2) | NAD⁺ metabolism | Disrupted energy/redox homeostasis, endocrine disruption | [81] |
| Common carp (Cyprinus carpio) | Lufenuron & Flonicamide (pesticides) | NAD⁺-linked antioxidant/immune pathways | Altered antioxidant gene expression and immune response | [82] |
| Mosquitofish (Gambusia affinis) | Triclosan | NAD⁺/SIRT/Nrf2 signaling | Downregulated SIRT, impaired antioxidant defenses | [84] |
| Yellowstripe goby (Mugilogobius chulae) | Paracetamol | NAD⁺/SIRT/PXR pathway | SIRT1/3 activation, oxidative stress mitigation, xenobiotic defense | [86] |
| Yellowstripe goby | Atorvastatin | NAD⁺/SIRT/PXR pathway | Altered sirt1/3 expression, antioxidant/inflammatory regulation | [88] |
| Common carp | Triclocarban (TCC) | NAD⁺/SIRT3 / redox balance | Neutrophil extracellular traps formation via sirt3 inhibition, ROS accumulation | [90] |
| Atlantic cod | Wastewater treatment plant effluents | NAD⁺-SIRT / neuronal related genes | Transcriptomic disruption, impaired mitochondrial defense | [91] |
| EPC fish cells | Fluorene-9-bisphenol (Bisphenol A substitute) | NAD⁺/SIRT3 / mitophagy | Quercetin protection, restored mitochondrial homeostasis | [92] |
| Delta smelt (Hypomesus transpacificus) | Ammonia | NAD⁺-redox pathways | Oxidative stress, metabolic resilience disruption | [93] |
| Common carp | Hydrogen peroxide | NAD⁺ redox/DNA repair | Neuronal oxidative damage, impaired NAD⁺ regeneration | [94] |
| Atlantic cod | Methylmercury | NAD⁺-linked mitochondrial pathways | Brain proteome disruption, neurotoxicity | [95] |
| Common carp | Cadmium | miR-217 / NAD⁺-SIRT1 axis | Immune dysregulation, NF-κB hyperacetylation | [96] |
| Fish (various, incl. Tachysurus sinensis) | Zinc (Zn) & Copper (Cu) | NAD⁺-SIRT1/3-autophagy | Zn activates lipophagy, Cu disrupts it, co-deficiency worsens steatosis | [97,98,99,100,101] |
| Crucian carp | ZnO nanoparticles | NAD⁺ redox / immune NETs | Oxidative stress, immune toxicity, NAD⁺ disruption | [98] |
| Tilapia (Oreochromis mossambicus) and Gibel carp (Carassius gibelio) |
Resveratrol (polyphenol) |
NAD⁺-SIRT1 / stress response | Enhanced antioxidant capacity, cold/ammonia stress protection | [102,103] |
| Grass carp (Ctenopharyngodon idella) | Astilbin (flavonoid) | NAD⁺-SIRT1/Nrf2 | Protection against PCB126-induced apoptosis | [104] |
Molybdo-flavoenzymes (MFEs) [such as aldehyde oxidase (AOX) and xanthine oxidoreductase (XOR)] are involved in the oxidation of N-heterocyclic compounds and aldehydes, including environmental pollutants, drugs, and vitamins. This biotransformation generates more polar compounds for easier excretion; thus, MFEs are classified as detoxification enzymes [78]. XOR activity has been demonstrated in fish liver species using xanthine as a substrate and O2 and NAD+ as electron acceptors, indicating both oxidase and dehydrogenase activities. Studies on the properties and specificities of MFE in non-mammalian vertebrates are limited. MFEs were examined in rainbow trout, which have a single AOX (AOXβ) and XOR. Unlike mammalian XOR, which uses both NAD+ and O2 as electron acceptors, trout XOR is exclusively NAD+-dependent with no O2 activity [79].
17α-ethinylestradiol is a synthetic estrogen primarily utilized in oral contraceptives, although it is also present in other contraceptive forms such as patches. It is recognized for its estrogenic effects and occurrence in the environment, particularly in treated wastewater [80]. A metabolomic study showed that exposure to environmentally relevant levels of 17α-ethinylestradiol causes metabolic disruptions in crucian carp, including alterations in NAD⁺ metabolic pathways. The results showed changes in energy metabolism and redox homeostasis, indicating that NAD⁺-dependent processes were sensitive to endocrine disruptors. Such disturbances could impair the cellular energy balance and contribute to toxic effects on fish physiology [81]. Another study examined how lufenuron and flonicamide affect the immune system and antioxidant gene expression in common carp gills. Since antioxidant defense and immune function are linked to cellular redox balance, NAD⁺ acts as a coenzyme in redox reactions and as a substrate for PARPs and SIRTs involved in oxidative stress responses. Altered NAD⁺ metabolism could affect antioxidant genes and immune responses under pesticide stress in this fish species [82]. Another study investigated triclosan (TCS), a chemical compound with antibacterial and antifungal properties, which is used in various consumer products such as soaps, toothpastes, and cosmetics [83]. TCS affects redox-sensitive microRNAs (RedoximiRs) / SIRT / Nrf2 / ARE signaling pathway in mosquitofish (Gambusia affinis), revealing disruption of NAD+-dependent antioxidant defenses. TCS exposure alters redox imiRs, downregulates sirtuin activity, and impairs Nrf2/ARE-mediated antioxidant responses leading to oxidative stress. TCS compromised the NAD+/SIRT axis, which is crucial for maintaining redox homeostasis through Nrf2 regulation. The results showed TCS's ecotoxicological risks of TCS by disrupting cellular defense mechanisms in aquatic organisms while highlighting the protective role of NAD+-dependent SIRT signaling. This study revealed the interference of contaminants with antioxidant pathways in wildlife [84]. Paracetamol is a medication widely used as an analgesic and antipyretic and it is present in the environment [85]. A study investigated the SIRT/PXR signaling pathway response in yellowstripe goby (Mugilogobius chulae) exposed to paracetamol, highlighting NAD+-dependent mechanisms in detoxification and adaptation. These findings show that paracetamol exposure modulated SIRTs, particularly SIRT1 and SIRT3, which are involved in oxidative stress mitigation through NAD+-mediated pathways. Activation of the pregnane X receptor (PXR) suggests a defense mechanism for xenobiotic metabolism. This study demonstrated the role of NAD+-linked SIRT activity in cellular resilience to pharmaceutical pollutants, providing insights into the molecular adaptations of aquatic organisms to environmental contaminants [86]. Atorvastatin is a type of medication used to lower cholesterol in humans [87]. The effects of atorvastatin on the SIRT/PXR signaling pathway were also studied in the same fish species (yellowstripe goby), revealing its role in NAD+-dependent cellular defence. Atorvastatin exposure altered sirt1 and sirt3 expression and activates PXR, suggesting an adaptive response to xenobiotic stress. These findings demonstrate that atorvastatin enhances antioxidant capacity through NAD+-mediated SIRT activity, thereby regulating oxidative stress and inflammatory pathways. These results highlight the role of the SIRT/PXR axis in mediating pharmaceutical stress responses in aquatic organisms, providing insights into NAD+ metabolism and xenobiotic detoxification while showing the protective role of NAD+-linked pathways [88]. Triclocarban (TCC), a polychlorinated antimicrobial agent, has been used in toys, clothing, packaging, medical supplies, and personal care products, such as soaps and toothpaste. Although used for more than 50 years, concerns regarding its endocrine disruptive properties have recently emerged [89]. TCC triggers neutrophil extracellular trap (NET) formation in common carp by disrupting SIRT3-mediated redox homeostasis, demonstrating a novel immunotoxic mechanism. TCC exposure suppressed NAD+-dependent SIRT3 activity, leading to mitochondrial ROS accumulation and ERK1/2/p38 MAPK signaling pathway activation, driving NETosis. These effects were mitigated by SIRT3 overexpression or antioxidant treatment, confirming the SIRT3-ROS crosstalk in TCC-induced NET formation. These findings provide the first evidence that environmental pollutants can hijack NAD+-SIRT networks to dysregulate innate immunity in fish, thereby linking metabolic stress to excessive inflammatory response. This work highlights ecological risks, while elucidating a conserved mechanism by which NAD+ depletion predisposes to immunopathology [90]. Another study investigated the neurotoxic effects of wastewater treatment plant (WWTP) effluents on juvenile Atlantic cod, showing transcriptomic disruptions in the brain tissue related to oxidative stress, neurotransmission, and NAD+-dependent pathways. The downregulation of genes involved in mitochondrial function and antioxidant defense suggests impaired NAD+-SIRT signaling, which is a key regulator of neuronal homeostasis. Pharmaceuticals and personal care products alter stress-responsive genes and neurotransmitter receptors with effects persisting after depuration. These findings show marine fish vulnerability to mixtures of these compounds, suggesting the disruption of NAD + metabolism as a neurotoxic mechanism. This study showed the sublethal effects of pollutant mixtures on commercial species [91]. Fluorene-9-bisphenol (BHPF) is a substitute or alternative for bisphenol A (BPA), which is used in the manufacture of plastics, including materials that are in contact with food. A study has elucidated the protective mechanism of quercetin against BHPF-induced apoptosis in epithelioma papulosum cyprini (EPC) cells through SIRT3-mediated mitophagy. BHPF is increasingly being used in plastic products. Quercetin upregulates NAD+-dependent SIRT3 expression, activates mitophagy to remove damaged mitochondria, and reduces oxidative stress-induced apoptosis. Quercetin counteracts BHPF cytotoxicity by restoring mitochondrial homeostasis via NAD+/SIRT3/mitophagy. These results show the potential of flavonoids to reduce toxicant damage in aquatic organisms while demonstrating the role of sirtuins in stress responses. This study revealed the mechanisms linking phytochemical protection and xenobiotic toxicity in fish cells [92].
Several studies have examined the adverse effects of ammonia and hydrogen peroxide. These basic chemical compounds are composed of small, lightweight elements that are commonly used as cleaning and disinfecting agents. Both substances function as oxidizing or reactive agents, making them effective in eliminating bacteria and stains. Because of these characteristics, ammonia and hydrogen peroxide are frequently used in industrial cleaning products. Sublethal ammonia exposure in delta smelt (Hypomesus transpacificus) has revealed physiological disruptions that affect its survival. The observed oxidative stress aligns with ammonia-induced disruption of NAD+-dependent pathways. These results indicated that ammonia triggers cellular stress through a redox imbalance associated with NAD+ homeostasis, suggesting population threats by impairing metabolic resilience linked to NAD+-sirtuin networks [93]. A transcriptomic study showed that hydrogen peroxide exposure disrupts brain function in common carp by inducing oxidative stress and altering neurotransmission pathways. Given that NAD⁺ is a central cofactor in redox reactions and DNA repair, oxidative damage increases NAD⁺ consumption, while impairing regeneration. These findings suggest that maintenance of NAD⁺ homeostasis is critical for neuronal energy metabolism under oxidative stress in aquatic vertebrates [94].
Other studies have focused on the negative impacts of heavy metals on fish environments. One study examined how methylmercury, a neurotoxic pollutant, affects the brains of Atlantic cod. Focusing on proteomic changes, the affected proteins were found to be related to NAD-dependent pathways, including mitochondrial function and energy metabolism. Brain proteomic alterations in methylmercury-exposed cod likely disrupt NAD-associated processes, contributing to neurotoxicity [95]. Another study revealed that cadmium (Cd) exposure induces immunotoxicity in common carp by dysregulating the miR-217/SIRT1/NF-κB axis through NAD-dependent SIRT1 suppression. Cd upregulates miR-217, which inhibits sirt1 expression, leading to NF-κB hyperacetylation and overproduction of pro-inflammatory cytokines. SIRT1 reactivation or miR-217 inhibition restored immune homeostasis, highlighting the therapeutic potential of targeting the miR-217/SIRT1 axis to mitigate Cd-induced inflammation in aquatic organisms [96]. Environmentally relevant concentrations of zinc (Zn) induce hepatic lipophagy in fish through SIRT1-mediated deacetylation of Beclin1 while alleviating copper (Cu)-induced lipotoxicity. Zn activates NAD+-dependent SIRT1, which deacetylates Beclin1 to promote autophagic flux and lipid clearance, whereas Cu disrupts this mechanism. Zn supplementation restored SIRT1/Beclin1 signaling and mitigated Cu-induced lipid accumulation, highlighting the role of metal homeostasis in regulating NAD+-sirtuin-autophagy pathways. These results provide the first evidence that trace metals modulate lipid metabolism through post-translational regulation of autophagy in aquatic species [97]. ZnO nanoparticles induce oxidative stress and immune toxicity in crucian carp, triggering NETs. Oxidative stress affects NAD⁺/NADH balance, impacting NAD⁺-dependent metabolic pathways for redox homeostasis and immune function. These disturbances in NAD⁺ metabolism contribute to immune dysregulation in fish exposed to pollutants [98]. In contrast, another study demonstrated that Cu and Zn deficiencies induced hepatic lipotoxicity in fish through mitochondrial oxidative stress-mediated inhibition of the Sirt3/Foxo3/PPARα pathway, revealing NAD+ homeostasis dependence. Cu and Zn are essential trace elements for terrestrial organisms. Studies have suggested that a single dietary Cu or Zn deficiency leads to liver lipid deposition and causes metabolic dysfunction-associated steatotic liver disease (MASLD). Fish and mammals share similarities in uptake, storage, utilization, excretion, and interaction of metal ions [99]. However, the metal-detoxifying protein metallothionein is expressed at lower levels in fish than in mammals [100]. Trace element co-deficiency suppresses NAD+-dependent Sirt3 activity, leading to Foxo3 hyperacetylation, impaired PPARα signaling, disruption of lipid oxidation, and exacerbation of hepatic steatosis in yellow catfish (Tachysurus sinensis). Mitochondrial ROS accumulation has been identified as both a cause and consequence of NAD+/Sirt3 axis dysfunction, leading to metabolic dysregulation. These results establish the role of Cu/Zn in maintaining the mitochondrial redox balance and sirtuin function, providing insights into how micronutrient deficiencies affect NAD+-sensitive pathways in vertebrates. This study suggests Sirt3 activation as an intervention for trace element-related hepatotoxicity [101].
To address and mitigate the adverse effects experienced by fish due to the presence of these environmental contaminants, various compounds have been evaluated. Resveratrol, a polyphenol that acts as a powerful antioxidant, is found in the skins of grapes, blueberries, raspberries, blackberries, and peanuts. Resveratrol enhances cold stress tolerance in tilapia (Oreochromis mossambicus) by activating NAD+- dependent SIRT1 signaling, which upregulates metabolic and antioxidant machinery for cellular homeostasis. Resveratrol increases mitochondrial biogenesis, antioxidant enzyme activity, and HSP70 expression while reducing oxidative damage under cold stress. These effects occur via SIRT1 activation, which modulates the energy metabolism. These results demonstrate the potential of resveratrol to improve fish resilience through NAD+-SIRT pathways, offering insights into stress adaptation in ectotherms [102]. Resveratrol mitigates ammonia-induced oxidative stress, inflammation, and mitochondrial dysfunction in Gibel carp (Carassius gibelio) by activating NAD+-dependent SIRT1/PGC-1α signaling. The results showed that resveratrol enhanced antioxidant capacity, reduced pro-inflammatory cytokines, and improved mitochondrial bioenergetics by upregulating SIRT1-mediated deacetylation of PGC-1α and restoring redox balance. These findings highlight the protective role of the NAD+/SIRT1 axis against ammonia toxicity, and suggest that resveratrol acts as a potential therapeutic agent via sirtuin-mediated pathways. This study revealed how polyphenols counteract xenobiotic-induced damage by modulating NAD+-sensitive stress response networks in fish [103]. Another study demonstrated that astilbin (a flavonoid) alleviates PCB126-induced hepatocyte apoptosis in grass carp by targeting the SIRT1/Nrf2 acetylation axis to restore mitochondrial function. Astilbin activates NAD+-dependent SIRT1, which deacetylates Nrf2 to enhance its antioxidant activity, thereby counteracting PCB126-induced oxidative stress and mitochondrial disruption. By modulating SIRT1/Nrf2 signaling, astilbin reduced cytochrome c release and caspase-3 activation, thereby mitigating the apoptotic cascades. These results highlight the therapeutic potential of flavonoid compounds in aquaculture by targeting NAD+-sirtuin pathways to protect against xenobiotic-induced hepatotoxicity while showing a link between mitochondrial metabolism and cell survival in fish [104]. Although these results are encouraging, further research is essential to achieve results that can be broadly applied to various types of pollutants and fish species.
4.2.3. NAD+-sensitive mechanisms in fish metabolic adaptations: Insights for managing aquaculture-associated disorders
The involvement of NAD+ in metabolic homeostasis and its pathological disruption is a subject of extensive research in humans. However, its specific functions in fish metabolic adaptation remain unexplored. Nevertheless, the available results elucidate conserved yet distinct NAD+/Sirt1-mediated mechanisms that underpin dietary adaptability in teleosts, revealing a compelling therapeutic avenue for the management of metabolic syndromes in commercially relevant fish species (Table 3).
Table 3.
Role of NAD+ in fish metabolic regulation and disruption.
| Fish species | Main findings related to NAD⁺/SIRTUINS (SIRTs) metabolism | References |
|---|---|---|
| Rainbow trout (Oncorhynchus mykiss) |
Glucokinase-independent glucose sensing in liver and Brockmann bodies; metabolic regulation linked to NAD⁺-dependent SIRTs; suggests alternative nutrient-sensing pathways | [105] |
| Several fish species | Link between adipose triglyceride lipase, lipid metabolism, inflammation and NAD⁺ depletion; low NAD⁺ impairs SIRT1 activity affecting lipid metabolism and inflammation | [106] |
| Wuchang bream (Megalobrama amblycephala) | Feeding restriction activates NAD⁺-dependent AMPK-SIRT1 pathway, suppressing nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB)-mediated inflammation and oxidative stress, improving glucose metabolism and mitochondrial function | [107] |
| Large yellow croaker (Larimichthys crocea) | n-3 PUFAs activate NAD⁺-SIRT1 pathway, reducing NF-κB-mediated inflammation, improving lipid metabolism and redox balance | [108] |
| Black seabream (Acanthopagrus schlegelii), juvenile | Fenofibrate activates peroxisome proliferator-activated receptor α (PPARα) /SIRT1 axis, enhancing fatty acid oxidation and reducing lipogenesis and inflammation, alleviating high-fat diet-induced hepatic dysfunction | [109] |
| Largemouth bass (Micropterus salmoides) | SIRT1 regulates lipid catabolism, inhibits lipogenesis, and enhances antioxidant defenses via NAD⁺/SIRT1/FOXO1 (Forkhead Box O3a) signaling; upregulated under nutrient deprivation. | [111] |
| Black seabream | Betaine supplementation restores NAD⁺, activates SIRT1/ Sterol Regulatory Element-Binding Protein 1 (SREBP-1)/PPARα pathway, reduces lipogenesis, enhances fatty acid oxidation, lowers inflammation, improves mitochondrial function. | [113] |
| Black seabream | SIRT1 protects against hepatic lipotoxicity through NAD⁺-dependent deacetylation of Ire1α, alleviating endoplasmic reticulum (ER) stress and lipid accumulation. | [114], [115] |
| Black seabream | Fucoidan activates SIRT1, modulating PERK-eIF2α-ATF4 axis, reducing ER stress, enhancing fatty acid oxidation, and improving redox homeostasis. | [117] |
Some studies have focused on the role of NAD⁺ as a regulator of carbohydrate and lipid metabolism as well as its involvement in endocrine disruption. A study revealed that the rainbow trout liver and Brockmann bodies employ glucokinase-independent mechanisms for glucose sensing, implicating alternative nutrient-sensing pathways, such as NAD+-dependent SIRTs, in metabolic regulation. Although not directly measuring SIRT activity, metabolic responses to glucose fluctuations, including changes in glycolytic enzymes and insulin/glucagon secretion, align with the NAD+-sensitive regulatory networks in mammals. These findings challenge glucokinase as the primary glucose sensor in fish, and suggest that SIRT-mediated metabolic adaptation plays a prominent role in teleost energy homeostasis. These results provide a foundation for investigating NAD+-linked nutrient sensing in aquatic species, with implications for vertebrate metabolic flexibility [105].
NAD⁺ has not been directly investigated in many metabolic studies in fish, but these findings suggest that NAD⁺ is involved in many processes. Connections between adipose triglyceride lipase (ATGL), lipid metabolism, inflammation (TNF-α), and NAD+ biology can be established. ATGL is the rate-limiting enzyme in triglyceride breakdown and releases fatty acids (FFAs) and glycerol. FFAs oxidize mitochondria, generate NADH, and affect the NAD+/NADH ratios. Impaired lipolysis disrupts mitochondrial function and NAD+ homeostasis. LPS induces TNF-α expression associated with metabolic dysfunction. Chronic inflammation activates PARP-1 and CD38, thereby depleting NAD+. Low NAD+ levels impair SIRT1 activity, which regulates lipid metabolism. Metabolic stress can alter NAD+ levels in aquaculture. Boosting NAD+ improves metabolic resilience under stress [106]. Future research should explore whether LPS-induced TNF-α affects NAD+ levels and whether NAD+ supplementation modulates ATGL activity or inflammatory responses in fish.
Another study demonstrated that controlled feeding restriction mitigated high-carbohydrate diet-induced oxidative stress and inflammation in Wuchang bream by activating the NAD+-dependent AMPK-SIRT1 pathway. The findings revealed that feeding restriction enhances SIRT1 activity, which suppresses NF-κB-mediated inflammation and upregulates antioxidant defenses (SOD and catalase), while concurrently improving glucose metabolism and mitochondrial function. These effects are mediated by AMPK-dependent NAD+ elevation, confirming the central role of the AMPK-SIRT1 axis in metabolic adaptation to dietary stress. These results highlight feeding restriction as a practical aquaculture strategy to counteract diet-induced metabolic disorders by activating conserved NAD+-sensitive pathways that optimize energy homeostasis and inflammatory responses in fish [107].
Dietary omega-3 polyunsaturated fatty acids (n-3 PUFAs) mitigate hepatic steatosis-induced inflammation in large yellow croaker (Larimichthys crocea) by activating the NAD+-dependent SIRT1 pathway, which also suppresses NF-κB-mediated inflammatory responses. The results showed that n-3 PUFAs upregulated SIRT1 expression and activity, leading to deacetylation and cytoplasmic retention of the NF-κB p65 subunit, thereby reducing pro-inflammatory cytokine production (e.g., interleukin 1β). These anti-inflammatory effects are associated with improved lipid metabolism and redox homeostasis, highlighting the dual role of n-3 PUFAs in alleviating both metabolic dysfunction and inflammation through the SIRT1-mediated NAD+-sensitive pathways. These findings provide mechanistic insights into how dietary lipids modulate hepatocyte inflammation in marine fish, supporting the use of n-3 PUFAs as functional nutrients to enhance metabolic health in aquaculture [108]. Another study demonstrated that dietary fenofibrate alleviates high-fat diet-induced hepatic lipid accumulation and inflammation in juvenile black seabream by activating the PPARα/SIRT1 axis, revealing a conserved NAD+-dependent metabolic regulatory mechanism in fish. Fenofibrate is a lipid-lowering pharmaceutical compound. In an ecotoxicological context, dietary fenofibrate refers to the exposure of fish to fenofibrate through feed supplementation, which simulates the dietary uptake of fenofibrate from contaminated environments. Fenofibrate is widely prescribed in human medicine to regulate lipid metabolism [109]. However, when introduced into aquatic systems via wastewater effluents, it may affect non-target organisms such as fish. Fenofibrate treatment significantly upregulated PPARα and SIRT1 expression, enhanced fatty acid oxidation, and suppressed lipogenic gene and pro-inflammatory cytokine expression. Coordinated activation of these pathways reduces ER stress and improves mitochondrial function, highlighting the dual role of fenofibrate in modulating lipid metabolism and immune responses through NAD+-sensitive SIRT signaling. These findings provide the first evidence that pharmacological PPARα activation can rescue diet-induced metabolic dysfunction in marine fish by synergizing with SIRT1, thereby offering potential therapeutic strategies for aquaculture-associated metabolic disorders [110].
The role of SIRT1 has been studied in the largemouth bass (Micropterus salmoides), emphasizing its critical NAD+-dependent functions in regulating lipid metabolism and antioxidant responses. The activation of SIRT1 promotes lipid catabolism while inhibiting lipogenesis, and it significantly enhances resistance to oxidative stress through the NAD+/SIRT1/FOXO1 signaling pathway. Notably, nutrient deprivation resulted in increased hepatic SIRT1 expression, suggesting its evolutionarily conserved role as a metabolic sensor in fishes. This study provides the first comprehensive evidence that SIRT1 serves as a key regulator of energy balance and redox stability in this economically important aquaculture species, highlighting the potential of targeting the NAD+/SIRT1 pathway to improve stress resistance and metabolic health in farmed fish [111]. These findings advance our understanding of the conserved functions of SIRT in vertebrates.
Betaine, also known as trimethylglycine (TMG), is a naturally occurring quaternary ammonium compound derived from glycine. It functions as an important osmolyte, contributing to the cellular osmotic balance and protecting cells against environmental stress. Additionally, betaine serves as a methyl group donor in one-carbon metabolism, and plays a critical role in the remethylation of homocysteine to methionine. Owing to these properties, it has been widely studied for its implications in liver function, cardiovascular health, and metabolic regulation [112]. A previous study demonstrated that dietary betaine supplementation alleviated high-fat diet-induced hepatic steatosis and inflammation in juvenile black seabream by activating the NAD+-dependent Sirt1/SREBP-1/PPARα pathway. Betaine restored hepatic NAD+ levels by enhancing Sirt1-mediated deacetylation of SREBP-1 (Sterol regulatory element-binding protein 1), which suppresses lipogenesis while upregulating PPARα-driven fatty acid oxidation. These effects were accompanied by reduced inflammatory markers and improved mitochondrial function, highlighting the role of betaine as a methyl donor that concurrently modulates NAD+/Sirt1 signaling to rebalance lipid metabolism and immune responses. These findings reveal a conserved mechanism in which betaine integrates one-carbon metabolism with sirtuin-mediated metabolic regulation, offering a nutritional strategy to combat diet-induced metabolic disorders in aquaculture [113]. Sirt1 also plays a crucial protective role against hepatic lipotoxicity in fish fed a high-fat diet through NAD+-dependent deacetylation of inositol-requiring enzyme 1 alpha (Ire1α). Ire1α is a transmembrane sensor protein located in the endoplasmic reticulum that plays a central role in the unfolded protein response, helping cells adapt to ER stress [114]. These findings demonstrated that Sirt1 activation alleviates high-fat diet-induced ER stress and lipid accumulation by deacetylating Ire1α, thereby restoring hepatic metabolic homeostasis in black sea bream. Notably, this study identified a conserved NAD+/Sirt1/Ire1α axis in fish that modulated lipid metabolism under nutritional stress, mirroring mammalian pathways. This study established an evolutionary link between SIRT-mediated metabolic regulation and nutritional stress responses in vertebrates [115]. Fucoidan is a polysaccharide mainly consisting of l-fucose and sulfate groups. It is highly valued worldwide, especially in the food and pharmaceutical sectors, owing to its potential therapeutic benefits. The remarkable biological activity of fucoidan is attributed to its unique molecular structure. Fucoidan is known for its antioxidant, antitumor, anticoagulant, antithrombotic, immunoregulatory, antiviral, and anti-inflammatory properties [116]. Fucoidan alleviates hepatic lipid deposition in black seabream by activating SIRT1-mediated modulation of the PERK-eIF2α-ATF4 axis, revealing a novel NAD+-dependent mechanism linking ER stress to lipid metabolism. Fucoidan enhanced SIRT1 activity, attenuating ER stress by suppressing the PERK pathway, reducing lipogenic gene expression, and promoting fatty acid oxidation. These effects improved mitochondrial function and redox homeostasis, highlighting the role of fucoidan in alleviating ER stress and lipid accumulation through NAD+-sensitive SIRT1 signaling. This is the first evidence that marine polysaccharides regulate hepatic lipid metabolism in fish by targeting the SIRT1/PERK axis, thereby offering potential dietary strategies to combat metabolic disorders in aquaculture [117].
5. NAD⁺ Metabolism and Neuromodulation in Fish: From Muscle Innervation to Cognitive Function
The main results of this study are summarized in Table 4. The presence of different putative neuromodulators in the nerves innervating the skeletal muscles of teleosts has been previously investigated. Morphological investigation involved histochemical staining of cryostat sections from the epaxial, hypaxial, and adductor mandibulae muscles of the gilthead seabream and eel (Anguilla anguilla) to reduce NADPH-diaphorase activity. While focusing on neuromodulators, this study relates to NAD metabolism, as neuromodulatory enzymes such as NADPH-diaphorase produce nitric oxide (NO), a key signaling molecule. NADPH acts as a cofactor in these reactions, linking NAD metabolism to neuromodulation of fish muscle function [118].
Table 4.
NAD metabolism and neuromodulation in fish.
| Species / Model | Focus / Pathway | NAD⁺/NADPH Role | Main Findings | References |
|---|---|---|---|---|
| Gilthead seabream (Sparus aurata), and eel (Anguilla anguilla) |
Neuromodulators & NADPH-diaphorase | NADPH as cofactor for nitric oxide (NO) production | Histochemical staining revealed NADPH-diaphorase activity in skeletal muscle nerves, linking NAD metabolism to NO-mediated neuromodulation of muscle function. | [118] |
| General vertebrate model | cAMP Response Element-Binding protein (CREB) transcription factor | Indirect NAD⁺/SIRT1 (sirtuin 1) regulation | CREB integrates extracellular signals into gene expression changes, regulating survival, metabolism, and circadian rhythms. | [119] |
| Goldfish (Carassius auratus) |
CREB in learning & memory | NAD⁺-SIRT1 regulation of CREB | Cognitive activity triggers CREB phosphorylation in memory-related brain areas; NAD⁺-SIRT1 likely modulates CREB-dependent plasticity. | [120] |
| Goldfish | miRNA-132/212 & fear memory | NAD⁺ in neuroplasticity & epigenetics | miRNAs regulate neuronal plasticity; altered NAD⁺ metabolism may affect memory formation and synaptic function. | [121] |
| Mediterranean farmed fish | Somatotropic axis & growth regulation | NAD⁺/SIRT1 metabolic regulation | Nutrition and environment modulate hepatic sirtuin activity; diet enhances NAD⁺-SIRT1 signaling, stress impairs growth via metabolic disruption. | [122] |
| Swordtail fish (Xiphophorus helleri) | NADPH-diaphorase atlas & escape reflex | NADPH as NOS cofactor | Mapped NADPH-d in Mauthner cells; linked NADPH-dependent NO signaling to escape reflex pathways. | [123] |
| Dogfish (Triakis scyllia) |
Vagal afferent NADPH-d activity | NADPH in sensory NO signaling | NADPH-d in vagal afferents suggests NADPH-dependent NO production in sensory/autonomic pathways. | [124] |
| Cichlid (Tilapia mariae) |
NADPH-d in central nervous system | NADPH in neural development | Histochemistry showed NADPH-d activity essential for NO-mediated maturation of neuronal pathways. | [125] |
| African cichlid (Haplochromis burtoni) | Brain regional NADPH-d mapping | NADPH turnover from NAD⁺ | Enrichment in entopeduncular nucleus suggests localized NAD⁺/NADP⁺ demand for NO signaling. | [126] |
| Goldfish (Carassius auratus) |
Nitric oxide synthase (NOS) and NADPH-d distribution | NADPH as cofactor for NO | Broad distribution in brain regions for sensory, motor, and neuroendocrine regulation. | [127] |
| Grass puffer (Takifugu niphobles) | NOS in branchial innervation | NADPH-dependent (NOS) activity | NOS activity in glossopharyngeal/vagal afferents links NAD⁺ metabolism to vascular regulation in gills. | [128] |
| Atlantic salmon (Salmo salar) | NAD⁺ in acoustic stress response | NAD⁺/NADH redox in auditory stress | Genes linked to NAD⁺ metabolism and oxidative stress protect auditory tissues from loud sound damage. | [129] |
The cAMP Response Element-Binding protein (CREB) functions as a transcription factor. Its main function is to convert signals from outside the cell into lasting alterations in the gene expression within the cell nucleus. Acting as a pivotal switch, it activates genes essential for processes such as learning, memory, cell survival, metabolism, and circadian rhythms when triggered by various signals [119]. One study explored CREB signaling in spatial learning and memory in goldfish, showing that cognitive activities trigger CREB phosphorylation in brain areas linked to learning. Although it does not directly address NAD+ or SIRTs, CREB activation is associated with NAD+-dependent regulation by SIRT1 in vertebrates. These results suggested that goldfish use conserved mechanisms of spatial cognition, including sirtuin-mediated regulation. These findings provide insights into fish neurobiology and metabolic effects on cognition across species [120]. In goldfish, spatial learning and stimulus responses have been studied in relation to cAMP response element-binding protein (CREB) signaling activation. CREB is crucial for memory formation and synaptic plasticity. NAD acts as a cofactor for enzymes, such as sirtuins (SIRT1), which modulate CREB activity and gene expression in learning and memory. Thus, NAD-dependent pathways may influence CREB activation, linking metabolism and neuronal plasticity during spatial learning [120]. In this fish species, changes in microRNA-132/212 expression affect fear memory. Because miRNAs regulate key pathways involved in neuronal plasticity, NAD⁺ metabolism, which supports neuronal energy homeostasis and sirtuin-mediated epigenetic regulation, could intersect with miRNA processes that influence memory. Altered NAD⁺ levels may contribute to cognitive deficits by affecting synaptic function and neuroprotective mechanisms [121].
Another study investigated the regulatory mechanisms of the somatotropic axis in Mediterranean marine farmed fish, and demonstrated how nutritional and environmental factors modulate growth through NAD+-sensitive metabolic pathways. These findings revealed that diet and environmental stressors alter hepatic sirtuin activity and NAD+ bioavailability, which influences growth hormone signaling and energy allocation. Optimal nutrition enhances the NAD+/SIRT1 axis and improves metabolic efficiency. However, environmental challenges disrupt this pathway, leading to growth retardation. These results highlight the interface between nutrient sensing and endocrine growth regulation in fish, providing a framework for optimizing aquaculture practices by targeting metabolic sensors that coordinate growth and environmental adaptation [122].
NADPH-diaphorase (NADPH-d) activity, which is linked to nitric oxide (NO) signaling, has been mapped in fish species, highlighting the importance of NAD-related cofactors in neural function, particularly in sensory processing and motor control. A detailed atlas mapping NADPH-d activity in the brains of the swordtail fish (Xiphophorus helleri) was developed. This enzyme has been studied in Mauthner cells, which are neurons involved in the fish escape reflexes. NADPH-d is a histochemical marker of nitric oxide synthase (NOS), which uses NADPH to produce NO from L-arginine. This finding links the NADPH-dependent activity to brain scape responses, indicating NADPH's importance of NADPH in NO-mediated neurotransmission. This study demonstrated the role of NADPH-dependent NO signaling in fish brains [123]. NADPH-d activity has also been studied in the vagal afferent pathway of dogfish (Triakis scyllia). The presence of NADPH-d activity in the vagal afferent pathway suggests that NADPH-dependent NO production plays a significant role in neural signaling within this sensory pathway in dogfish. This highlights the importance of NADPH in modulating neural functions related to autonomic control via NO synthesis [124]. NADPH-d activity was investigated in the central nervous system of cichlid fish (Tilapia mariae). These results highlight the importance of NADPH in the maturation and functioning of neuronal pathways in fish, emphasizing its critical role in NO-mediated neurodevelopment and neural communication [125]. Another histochemical study mapped NADPH-d activity in the brains of African cichlid fish (Haplochromis burtoni), with notable enrichment in the entopeduncular nucleus. NADPH is generated from NADP⁺, which is ultimately derived from NAD⁺ via phosphorylation, indicating that these patterns reflect the localized metabolic demand for NAD⁺-derived cofactors. These results suggest that specific brain regions have elevated NAD⁺/NADP⁺ turnover to sustain NO signaling, highlighting the integration of redox cofactor metabolism with neuromodulatory functions in teleosts [126]. The distribution of NADPH-d activity and NOS reactivity in the central nervous system of goldfish were analyzed. This study showed that NADPH-diaphorase, which reflects NOS activity, is widely distributed in multiple brain regions involved in sensory processing, motor control, and neuroendocrine regulation. This distribution highlights the role of NADPH as a critical cofactor in the enzymatic production of NO, a key neuromodulator in teleost fish brain function that influences neural signaling pathways and physiological processes [127]. On the other hand, NOS was studied in the glossopharyngeal and vagal afferent pathways of the grass puffer (Takifugu niphobles), focusing on branchial vascular innervation. This study links NAD metabolism directly to NO signaling pathways in fish, highlighting the role of NADPH-dependent NOS activity in regulating vascular function and neural signaling in teleost gills [128]. Another study underscored NAD's critical role in cellular resilience and neuroprotection of fish exposed to acoustic stress. Specifically, a specific gene set was identified in the ears of fish, using Atlantic salmon (Salmo salar) as a model to assess the potential impact of loud sounds, such as those from seismic surveys. Research has focused on genes involved in auditory function, stress responses, and cellular repair mechanisms. Among these, genes related to NAD⁺/NADH-dependent processes and oxidative stress pathways have been highlighted, reflecting the importance of redox balance and energy metabolism in protecting auditory tissues from sound-induced damage [129]. Given the unique sensory characteristics of marine fish and the vast diversity of species, further research on the effects of NAD+ metabolism on these sensory systems and their relationship with the central nervous system is recommended.
6. Dietary Interventions and NAD+ Homeostasis: Implications for Fish Health and Product Quality
Numerous in vivo assays have been developed for fish, in which the diet is enriched with a variety of compounds. In this review, we focus on studies utilizing biologically active compounds that are essential for metabolic processes. These compounds are involved in biochemical pathways including energy production, tissue repair, and cellular regulation, highlighting their importance in nutrition and physiological functions. Interestingly, there is a scarcity of studies on this subject that have examined the role of NAD+ and its associated metabolites (Table 5).
Table 5.
Modulation of NAD⁺ pathways in fish through the use of dietary supplements to improve growth, immunity, and product quality.
Table 5.
Modulation of NAD⁺ pathways in fish through the use of dietary supplements to improve growth, immunity, and product quality.
| Fish species | Supplements/Context | Key Findings | References |
|---|---|---|---|
|
Gilthead seabream (Sparus aurata) |
Sirtuins (SIRTs), genes & fasting | Fasting upregulated hepatic sirt1/sirt3, showing NAD+-dependent roles in nutrient deprivation. SIRT functions tissue-specific, with gene duplications suggesting subfunctionalization. | [130] |
| Wuchang bream (Megalobrama amblycephala) | Mulberry leaf meal | Dietary supplementation (6–9%) enhanced growth, feed efficiency, antioxidant capacity, and immune genes. Likely influences NAD+-dependent SIRT-mediated regulation. | [131] |
|
Gilthead seabream |
Chitosan-tripolyphosphate-DNA nanoparticles | Gene delivery enhanced carbohydrate-to-lipid conversion; NAD+/NADH and NADPH involved in lipogenesis. Suggests central role of NADPH-dependent pathways in lipid biosynthesis | [132] |
|
Tilapia GIFT (Oreochromis niloticus) |
Branched-chain amino acids (BCAA) supplementation | Leucine/valine enhanced growth, glycolipid metabolism, immune function via NAD+-SIRT1/AMPK pathways. Improved insulin sensitivity, antioxidant capacity, and disease resistance | [133] |
|
Grass carp (Ctenopharyngodon idella) |
Niacin deficiency | Deficiency caused poor flesh quality, glycolysis increase, mitochondrial dysfunction. Niacin is a precursor for NAD+/NADP+, essential for energy metabolism | [134] |
| Meagre (Argyrosomus regius) and gilthead seabream |
Fish by-products | By-products rich in niacin, tryptophan, proteins – contribute to NAD+ biosynthesis via de novo/salvage pathways. Implications for aquafeeds and functional foods | [135] |
|
Wuchang bream |
NAD⁺ precursors (hyperglycemia) | NA, NAM, NR, NMN tested against high-glucose damage. NR most effective: restored NAD+, activated SIRT1/SIRT3, reduced oxidative stress/inflammation, improved glucose metabolism | [138] |
|
Nile tilapia (Oreochromis niloticus) |
Zophobas atratus larval meal | Replacing soybean meal improved flavor quality and energy metabolism. Enhanced NADH, acetyl-CoA, ATP, fatty acid accumulation; increased umami compounds, reduced off-flavors | [143] |
|
Zig-zag eel (Mastacembelus armatus) |
Spirulina supplementation and Aeromonas hydrophila infection | Improved liver immune/metabolic responses under infection. Likely acts through NAD-dependent enzymes (SIRTs, PARPs) regulating oxidative stress and inflammation | [144] |
|
Killifish (Nothobranchius guentheri) |
Resveratrol in reproductive aging |
Activated NAD+/SIRT1 axis, reduced inflammation, improved lipid metabolism, delayed ovarian aging. Highlighted role of SIRT1 in gut senescence, hepatic steatosis, and reproduction | [145] |
|
Nile tilapia |
Resveratrol |
Improved hepatic lipid metabolism in red tilapia by activating NAD+/SIRT1/AMPK signaling, enhancing lipolysis, and suppressing lipogenesis | [146] |
| Killifish |
Resveratrol in short-lived fish | Delayed ovarian aging | [147] |
|
Black seabream (Acanthopagrus schlegelii) |
Arachidonic acid |
Optimal 0.76% diet improved growth, lipid metabolism via SIRT1 activation. Promoted FA oxidation, reduced lipogenesis/oxidative stress. Linked NAD+ pathways with eicosanoid signaling | [148] |
|
Coho salmon (Oncorhynchus kisutch) |
Vitamin K3 + nicotinamide |
VK3 + nicotinamide improved growth, antioxidant capacity, tissue composition. Nicotinamide component supports NAD+ salvage pathway and redox balance | [149] |
The tissue-specific expression and nutritional regulation of SIRT genes (SIRT1-7) were elucidated in gilthead seabream, revealing that fasting significantly upregulates hepatic sirt1 and sirt3 expression, highlighting their NAD+-dependent roles in metabolic adaptation to nutrient deprivation. These findings demonstrate conserved yet specialized SIRT functions across tissues, with SIRT1/SIRT3 dominating in metabolically active organs (liver and muscle) and SIRT2 dominating in the brain, while identifying teleost-specific gene duplications (SIRT3 and SIRT5), suggesting potential subfunctionalization. These results provide critical insights into how sirtuins orchestrate energy homeostasis in marine fish through NAD+-sensitive pathways, thereby establishing an evolutionary framework for understanding metabolic stress responses in aquatic vertebrates [130].
Another study demonstrated that dietary supplementation with mulberry leaf meal enhanced the growth performance and immune function of juvenile Wuchang bream by modulating key metabolic and immune pathways, including the potential activation of NAD+-dependent processes. The findings revealed that mulberry leaf inclusion (optimal at 6-9% of diet) significantly improved growth rate, feed utilization, and antioxidant capacity, while upregulating immune-related genes (e.g., TNF-α and IL-1β) and intestinal barrier function, suggesting its role as a functional feed additive that may influence SIRT-mediated metabolic regulation. These results highlight the potential of phytogenic feed supplements to enhance aquaculture productivity through multi-targeted effects on fish physiology, possibly involving NAD+-sensitive pathways that link nutrition with immune-metabolic homeostasis [131].
Dietary modulation of NAD+ precursors and related metabolic pathways influence fish growth, product quality, and disease resistance. For example, the delivery of chitosan-tripolyphosphate-DNA nanoparticles expressing the transcription factor SREBP1a enhanced the conversion of dietary carbohydrates into lipids in the liver of gilthead seabream. Although NAD was not explicitly mentioned in this study, it is likely to be involved in this metabolic shift, because NAD+/NADH is a crucial cofactor in carbohydrate metabolism and lipid biosynthesis. Increased lipogenesis driven by SREBP1a expression would requires NADPH as the key reducing agent for fatty acid synthesis. Therefore, modulation of NADPH-dependent pathways is central to the enhanced lipid production observed following nanoparticle-mediated gene expression in fish livers [132]. Dietary supplementation with branched-chain amino acids (BCAAs) leucine and valine enhances growth performance, glycolipid metabolism, and immune function in genetically improved farmed tilapia (GIFT, O. niloticus), potentially via the NAD+-sirtuin pathway. These findings show that BCAAs improve hepatic insulin sensitivity and antioxidant capacity while reducing inflammation, which is associated with SIRT1/AMPK activation. Optimal BCAA ratios enhance nutrient utilization and disease resistance, suggesting their role in metabolic-immune homeostasis via NAD+-dependent regulatory networks. These results provide dietary guidelines for tilapia aquaculture and suggest associations between amino acid metabolism and SIRT-mediated regulation in teleosts [133]. Niacin (vitamin B3) deficiency in grass carp results in poor flesh quality and is associated with metabolic disruptions. Niacin is a precursor of NAD+ and NADP+, which are essential cofactors in cellular metabolism. Deficiency causes increased glycolysis and mitochondrial dysfunction, reflecting an imbalance between NAD+/NADH ratio and energy homeostasis. This highlights the crucial role of NAD biosynthesis in fish health, growth, and product quality [134]. In a study that primarily analyzed the nutritional composition of fish by-products [head, gills, intestines, trimmings, bones, and skin from meagre (Argyrosomus regius) and gilthead seabream], the findings have indirect implications for NAD⁺ metabolism. The by-products were rich in niacin (vitamin B₃) precursors, amino acids such as tryptophan, and high-quality proteins, all of which contribute to NAD⁺ biosynthesis through the de novo and salvage pathways. The utilization of such byproducts in aquafeeds or functional foods could help sustain NAD⁺ levels in fish and humans, supporting metabolic health, oxidative stress resistance, and cellular energy production [135].
Wuchang bream is a cyprinid fish with a significant commercial value [35]. Owing to its herbivorous diet, this species is susceptible to hepatocyte injury induced by high glucose levels, which is a consequence of consuming high-carbohydrate diets [136]. Notably, NAD+ depletion has been observed in individuals of this species who experience hyperglycemia [137]. Consequently, M. amblycephala serves as an appropriate model for studying liver damage induced by high glucose levels. In light of these considerations, one study investigated the protective effects of four NAD⁺ precursors (NA, NAM, NR, and NMN) against high-glucose-induced hepatocyte damage in this fish species. NR was the most effective in restoring NAD⁺ homeostasis, activating Sirt1/Sirt3, reducing oxidative stress and inflammation, and improving glucose metabolism both in vitro and in vivo. NR and NMN were particularly effective in improving glucose metabolism, suggesting their therapeutic potential for mitigating hyperglycemia-related liver damage. These findings highlight NAD+ supplementation as a promising strategy for addressing diabetes-related hepatic dysfunction [138]. However, the efficacy of NAD+ precursors for the treatment of metabolic disorders remains unclear. A comparison of NAD+ precursors focused on their effectiveness in reducing high glucose-induced hepatocyte damage. Each precursor has distinct properties that affect the NAD+ levels and pathway activation. Identifying the most effective precursor (NR) can help develop strategies to combat metabolic dysfunctions. This study identified NR as the most potent precursor for protecting hepatocytes and improving metabolic health.
Another popular topic in aquaculture is the use of alternative sources of protein in aquatic feed derived from agricultural waste, which can reduce the feed production costs in developing areas. Agricultural waste is used to raise insects for dry meal production in animal feed and human consumption [139]. Zophobas atratus Fab. (superworms) from the order Coleoptera and family Pyrethidae serve as food and feed sources [140]. Z. atratus grows faster and adapts better than Tenebrio molitor L. [141]. Z. atratus larval meal (ZLM) can improve fish growth and meat quality, though its metabolic effects are understudied. Although tilapia production has increased through factory farming, consumer focus has shifted from quantity to quality, emphasizing fish meat flavors [142]. One study examined how ZLM improved the flavor of tilapia meat. Soybean meal in the basal diet was replaced with 15, 30, or 60% ZLM. After 30 days, the dorsal muscles of tilapia underwent sensory evaluation, whereas the liver samples were subjected to metabolomic analysis. ZLM enhanced liver energy metabolism enzymes, including NADP-malate dehydrogenase (NAD-MDH), increasing NADH, acetyl-CoA, and ATP levels, leading to fatty acid accumulation. Flavor nucleotides and umami metabolites increased, whereas off-flavor metabolites decreased. This study showed that ZLM diets enhanced tilapia muscle flavor by reducing earthy taste and increasing flavor compounds, thereby improving meat quality [143].
In a recent study, replacing fishmeal with Spirulina platensis affected the liver of a zig-zag eel (Mastacembelus armatus) infected with A. hydrophila. NAD is likely involved here because liver metabolic and immune responses depend heavily on the redox balance and energy metabolism, both of which are regulated by NAD/NADH and NADP/NADPH pools. Spirulina is known for its antioxidant and immunomodulatory properties, which can influence NAD-dependent enzymes (e.g., SIRTs and PARPs) involved in the oxidative stress response and inflammation. Thus, dietary Spirulina may modulate NAD-related pathways in the eel liver, enhancing resistance to bacterial infection and supporting liver function during immune challenges [144].
As mentioned in this review, several studies have reported the use of resveratrol as a dietary supplement in fish. Resveratrol attenuates the senescence-associated secretory phenotype (SASP) in the gut of Günther's killifish (Nothobranchius guentheri) through activation of the NAD+-dependent SIRT1/NF-κB pathway. Resveratrol treatment reduced age-related inflammation by enhancing SIRT1-mediated deacetylation, suppressing NF-κB signaling, and decreasing pro-inflammatory cytokine production. These results highlight the role of the NAD+/SIRT1 axis in modulating gut senescence, and suggest that SIRT1 activation may counteract age-related gut dysfunction. This study provides insights into the mechanisms linking SIRT activity, inflammation, and aging in vertebrates, supporting the potential of NAD+-boosting compounds in age-related diseases [145]. Resveratrol supplementation improved hepatic lipid metabolism in red tilapia by activating NAD+/SIRT1/AMPK signaling, enhancing lipolysis, and suppressing lipogenesis. The results showed that resveratrol reduced hepatic lipid accumulation by upregulating fatty acid oxidation genes (e.g., PPARα) and downregulating adipogenic markers through SIRT1-dependent deacetylation. These findings highlight the potential of resveratrol in managing hepatic steatosis in aquaculture species by modulating NAD+-sensing pathways that regulate energy homeostasis, thereby providing insights into polyphenol-mediated metabolic reprogramming in teleost fish [146]. In addition, resveratrol significantly delayed ovarian aging in short-lived Günther's killifish by alleviating inflammation and ER stress through the SIRT1/NRF2 signaling pathway. These findings reveal that the anti-aging effects of resveratrol are mediated by SIRT1, an NAD+-dependent deacetylase that enhances antioxidant defenses. Resveratrol mitigates age-related ovarian dysfunction and preserves follicular integrity by boosting NAD+ levels and SIRT1 activity These results highlight the role of NAD+/SIRT1 signaling in combating ovarian aging and suggest resveratrol as a promising therapeutic agent for age-related reproductive decline. This study provides insights into the mechanisms linking NAD+ metabolism and ovarian senescence [147].
The optimal dietary level of ARA (an omega-6 fatty acid involved in inflammation) in black seabream was found to be 0.76% of dry weight, underscoring its role in regulating growth and lipid metabolism through the NAD+-sirtuin pathways. ARA supplementation enhances growth and hepatic lipid homeostasis by promoting fatty acid oxidation (PPARα) and inhibiting lipogenesis, aligning with SIRT1 activation. The anti-steatotic effects of ARA were linked to improved mitochondrial function and reduced oxidative stress, suggesting an interaction between NAD+-dependent regulation and eicosanoid signaling. These findings provide dietary guidelines and indicate that ARA may optimize NAD+/SIRT1 signaling for metabolic flexibility in marine fish [148]. A recent study investigated the effects of dietary supplementation of menadione nicotinamide bisulfite (VK3) on Coho salmon alevins. VK3 is a synthetic form of vitamin K3 that integrates vitamin K₃ with nicotinamide. The nicotinamide component supports the redox balance and energy metabolism through the NAD+ salvage pathway. The findings showed that Optimal VK3 levels improved growth performance, antioxidant capacity, and tissue composition, indicating that dietary nicotinamide enhances NAD⁺-dependent processes essential for early fish development [149].
7. NAD⁺ Related Metabolites and Their Implications in Fish Skin
Despite the importance of the skin as a primary barrier against external pathogen entry in fish, and as a vital organ for defense, few studies have been conducted on its relationship with NAD+ metabolism (Table 6).
Knowledge of the differences in innate immunity between the wild-type (WR_S) and yellow mutant rainbow trout (YR_S) (O. mykiss) remains limited. Transcriptome analysis of the skin was performed. Pathway enrichment analyses revealed that differentially expressed mRNAs were enriched in innate immune-related GO terms and pathways, including NAD+ADP-ribosyltransferase activity. GO functional enrichment analysis comprised biological processes, cellular components, and molecular functions. Five molecular function sub-categories were identified: 'NAD+ADP-ribosyltransferase activity' (GO:0003950), 'complement binding' (GO:0001848), 'nicotinate-nucleotide diphosphorylase activity' (GO:0004514), 'lipid transporter activity' (GO:0005319) and 'endopeptidase inhibitor activity' (GO:0004866). Nampt, nicotinate phosphoribosyltransferase (naprt), and cyclic ADP-ribose hydrolase 1 (cd38) were upregulated in WR_S, functioning in nicotinate and nicotinamide metabolism [150]. Two GO terms were assigned to NAD+ADP-ribosyltransferase activity (molecular function) and NAD biosynthetic processes (biological processes), with the former being the most significantly enriched. NAMPT and NAPRT are rate-limiting enzymes catalyzing NAD synthesis from NA and NAM precursors, and their increased activity accelerates NAD synthesis [151,152].
Table 6.
NAD⁺ related metabolites and their implications in fish skin.
| Fish species | Context | Key Findings | References |
|---|---|---|---|
|
Rainbow trout (Oncorhynchus mykiss) |
Wild-type vs yellow mutant |
Transcriptome analysis revealed NAD+ADP-ribosyltransferase activity and NAD biosynthetic processes enriched |
[150] |
|
Blass bloched rockfish (Sebastes pachycephalus) |
Skin pigmentation |
Nicotinamide Riboside Kinase 2 (NMRK2) differentially expressed across skin color types; involved in NAD biosynthesis. Suggests NAD pathways affect pigmentation via cellular energy metabolism | [153] |
|
Cichlids |
Aquaculture implications of pigmentation | Skin coloration linked to marketability, health indicators, selective breeding, mate selection, and survival. NAD+-related genes (e.g., NMRK2) influence pigmentation processes | [155] |
Another study identified differentially expressed genes associated with skin color patterns in blass-bloched rockfish (Sebastes pachycephalus), focusing on NAD-related pigmentation pathways. Nicotinamide Riboside Kinase 2 (NMRK2) was differentially expressed among skin color types and plays a role in NAD biosynthesis, suggesting that NAD metabolic pathways affect pigmentation through cellular energy metabolism in pigmented cells. These findings indicate a connection between nicotinamide metabolism and the development of color pattern in fish. Because NMRK2 is involved in NAD+ biosynthesis, which is a crucial cofactor for cellular metabolism, its varied expression suggests a metabolic influence on skin coloration. This advances our understanding of the molecular mechanisms underlying body color patterns in this fish species, supporting future genomic research and applications in aquaculture breeding, while offering insights into pigmentation processes across vertebrates [153]. In aquaculture, fish skin coloration affects marketability, consumer preferences, and economic value. Bright skin colors indicate health and quality, attract buyers, and boosting sales [154]. Specific skin colors are essential for selective breeding programs to enhance aesthetic appeal, camouflage, and species identification. Skin coloration influences social interactions, mate selection, and survival and affects fish behavior and reproductive success [155]. Understanding skin pigmentation is crucial to improve farmed fish production and commercial success. Research indicates that metabolic genes such as NMRK2 affect pigmentation processes in fish. Studies have shown that genes involved in NAD+ biosynthesis and energy metabolism are linked to pigmentation and pigmented cell functions. In zebrafish, metabolic pathway mutations can alter melanophore development and melanosome function [156]. Research on cichlids suggests that the metabolic state influences pigment cell behavior [155]. These findings suggest that NMRK2 may regulate pigment cell activity and color patterns in fish.
8. NAD⁺ Metabolism in Fish: Implications For Immune Defense and Cellular Homeostasis
NAD+-related metabolic pathways were found to be significantly altered during bacterial and parasitic infections, underscoring their roles in orchestrating immune defenses and maintaining cellular homeostasis (Table 7). Similar results have been reported in studies on bacterial infections. For example, another study used metabolomic profiling to explore the inflammatory and oxidative stress responses in common coral trout (Plectropomus leopardus) infected with Vibrio sp. Key findings indicated significant alterations in NAD⁺-dependent metabolic pathways, including shifts in the redox balance and energy metabolism. The modulation of NAD⁺-linked enzymes and cofactors under infection stress suggests that NAD⁺ metabolism plays a central role in orchestrating immune defense and maintaining cellular homeostasis during bacterial challenge [157].
Aptamers are artificial single-stranded oligonucleotide molecules that bind to specific target molecules with a high affinity and specificity [158]. Aptamers are widely utilized in various fields, such as disease diagnosis and treatment, analysis and detection, and targeted therapy [159]. Transcriptomic approximations were used to determine how aptamer B4 inhibited Pseudomonas plecoglossicida (a fish pathogen). While this work mainly focuses on antibacterial mechanisms, transcriptomic shifts likely involve metabolic pathways, including NAD+/NADH redox reactions, which are key to bacterial energy metabolism and stress responses. Understanding these pathways could reveal targets in NAD-dependent processes for therapeutic intervention [160].
Table 7.
NAD⁺ related metabolic pathways in fish infection, immunity, and therapy.
| Fish species | Context | Key Findings | Reference |
|---|---|---|---|
| Coral trout (Plectropomus leopardus) |
Bacterial infection (Vibrio sp.) |
Metabolomic profiling revealed alterations in NAD+-dependent pathways, affecting redox balance and energy metabolism | [157] |
| Large yellow croakers (Pseudosciaena crocea) | Aptamers and bacterial infection (Pseudomonas plecoglossicida) | Aptamer B4 inhibits pathogen; transcriptomic shifts involve NAD+/NADH redox reactions; potential therapeutic targets in NAD-dependent processes | [160] |
| Grass carp (Ctenopharyngodon idella) | Viral infection (IRF9, interferon regulator factor 9) | IRF9 inhibits SIRT1, enhances p53 acetylation & apoptosis; demonstrates trade-off between metabolic regulation and immune defense | [161] |
| Killifish (Nothobranchius guentheri) |
Metformin and Poly I:C | Metformin attenuates gut aging via NAD+-dependent AMP-activated protein kinase activation; reduces inflammation, oxidative stress, enhances mitochondrial function | [163] |
| Chinese perch (Siniperca chuatsi) |
Sirtuin 6 (SIRT6) in antiviral defense | SIRT6 enhances interferon-stimulated genes; viral infections increase NAD+; highlights SIRT6 role in NAD+-dependent antiviral defense | [164] |
| Grouper hybrid (Epinephelus fuscogutatus × Epinephelus lanceolatus) | Parasite resistance | Transcriptomic analysis revealed NAD+-dependent enzymes involved in immune signaling and redox balance, contributing to parasite resistance | [165] |
Another study revealed a mechanism by which interferon regulatory factor 9 (IRF9) promotes apoptosis and enhances innate immunity in grass carp by suppressing the SIRT1-p53 axis. IRF9 directly inhibited SIRT1 activity, leading to increased p53 acetylation and apoptosis during viral infection, thereby strengthening the antiviral response. This study showed an evolutionarily conserved trade-off between metabolic regulation and immune defense, providing evidence that IRF9 prioritizes immune activation over cellular survival. These results advance our understanding of immunity and NAD+-mediated metabolic regulation in vertebrates, with implications for immunostimulatory strategies in aquaculture [161].
Metformin is a first-line therapy for the treatment of type 2 diabetes, due to its robust glucose-lowering effects, well-established safety profile, and relatively low cost [162]. One study explored the protective effects of metformin against inflammation and oxidative stress in the Günther's killifish gut following polyinosinic:polycytidylic acid (poly I:C)-induced aging-like phenomena. These findings indicate that metformin treatment attenuates gut aging by reducing inflammation and oxidative stress and enhancing mitochondrial function. Metformin activates AMPK signaling, contributing to anti-aging effects through NAD+-dependent pathways. These results suggest that metformin can counteract age-related gut dysfunction by mitigating inflammation and oxidative damage [163]. Another study revealed a novel immunoregulatory role of SIRT6 in antiviral defense mechanisms in Chinese perch (Siniperca chuatsi), demonstrating NAD+-dependent regulation of the host antiviral response. SIRT6 enhances interferon-stimulated gene expression and promotes antiviral immunity through RIG-I–like receptor signaling. Viral infections increase cellular NAD+ levels, suggesting a link between NAD+ metabolism and innate immunity in vertebrates. These results showed that SIRT6, an NAD+-sensing protein, plays a crucial role in antiviral defense in fish, highlighting its therapeutic applications in aquaculture. This study advances our understanding of the SIRT-mediated immunity in vertebrates [164].
A transcriptomic analysis of a parasite-resistant grouper hybrid (Epinephelus fuscogutatus × E. lanceolatus) was performed to elucidate the innate immune mechanisms underlying resistance. This study revealed the differential expression of genes involved in immune signaling, inflammation, and oxidative stress responses. Genes associated with NAD⁺-dependent enzymes and redox balance were implicated, showing NAD's role in modulating metabolism and immune function during pathogen defense in fish. This indicates that NAD-related pathways contribute to the immune resilience against parasites [165].
These findings collectively underscore the fundamental significance of NAD+ metabolism in fish biology, encompassing cellular energy production and complex physiological processes such as immune function and environmental adaptation. This study highlights the potential of targeting NAD+-related pathways in aquaculture and fish health management strategies.
9. NAD⁺ Influence in Fish Eggs and Declining in Muscle Post-Mortem
Different enzymes related to NAD+ metabolism have been studied in fish eggs and postmortem muscle. In both cases, the same idea was used, to determine the fundamental enzymatic principles that could be investigated in both systems. Both are highly useful, albeit very different, model systems for understanding enzyme kinetics, regulation, and functions under specific conditions.
Enzymatic activities within eggs have been proposed as indicators of quality. The measurement of biochemical components, such as the NADH/NAD ratio and egg respiration rate, provided reliable insights into the viability of lake trout (Salmo trutta lacustris) eggs [166]. The potential application of these parameters for predicting egg quality during short-term (4 hours) storage was examined. The studied species included common carp, silver carp (Hypophthalmichthys molitrix), grass carp, and bleak (Chalcalburnus chalcoides). Many changes were observed in the eggs, although their biochemical composition and enzymatic activities remained unchanged. The parameters examined for their correlation with fertilization rate included egg enzyme activities crucial for energy metabolism (such as NAD-dependent malate dehydrogenase) and biosynthetic processes NADP-dependent isocitrate dehydrogenase) [167].
An investigation examined the influence of factors such as nucleoside triphosphates, inorganic salts, NADH, catecholamines, and oxygen saturation on nitrite-induced oxidation of rainbow trout hemoglobin (Hb). NADH acts as a reducing agent affecting hemoglobin's oxidative state. This study shows NADH's importance in modulating hemoglobin's vulnerability to nitrite-induced oxidation, which is essential for oxygen transport and preventing methemoglobin formation in fish [168]. Physiological studies have demonstrated that hypoxia-tolerant species, such as carp, employ NADH-dependent mechanisms to preserve Hb functionality under oxygen-limited conditions [169], whereas Antarctic fish have evolved enhanced Hb redox stability as an adaptation to extreme environments [170]. The redox behavior of fish hemoproteins, such as myoglobin (Mb) and Hb, is crucial for both physiological adaptations and seafood quality after death. This review provides significant insights from both foundational and recent research to clarify the distinct redox characteristics of fish hemoproteins and their practical applications. Expanding on the foundational research by Brown and Snyder regarding NADH/flavin-driven redox processes in mammalian hemoproteins [171], studies have demonstrated that fish Mb undergoes unique oxidation processes that occur two to three times faster than those in mammals. This significantly affects the color stability in species such as sardine (Sardinella gibbosa) and mackerel (Rastrelliger kanagurta) [172]. The NADH-cytochrome b₅ reductase system has been identified as crucial for maintaining the reduced state of Mb in tuna muscle, directly impacting shelf life and visual quality [173]. These fundamental insights have driven innovations in seafood preservation, including the development of CO-treated tuna products that reduce metmyoglobin formation by 60% [174], and optimized freezing protocols to minimize oxidation in mackerel, highlighting the need for rapid freezing and antioxidant treatments to preserve fish quality [175].
Reduction in NAD+ and NADH levels following death has been associated with muscle breakdown and a decline in the quality of stored fish, highlighting the crucial importance of NAD+ in sustaining cellular energy and redox equilibrium. This investigation employed NMR spectroscopy to monitor biochemical alterations in the muscle of Atlantic salmon post-mortem at various storage temperatures. After death, a decrease in NAD⁺ and its reduced counterpart, NADH, disrupts cellular energy metabolism, resulting in changes in glycolysis and oxidative processes. The study underscores how shifts in NAD⁺ metabolism contribute to muscle deterioration and quality degradation in stored fish, emphasizing the vital role of NAD⁺ in preserving cellular energy and redox balance, even after death [176]. Collectively, these studies bridge molecular redox chemistry with ecological adaptation and food technology, offering a deeper understanding of fish physiology and actionable strategies in the seafood industry. The integration of these findings presents new opportunities for optimizing aquaculture practices, improving seafood preservation methods, and understanding evolutionary adaptations in aquatic vertebrates.
10. Concluding Remarks and Future Research
Several key conclusions can be drawn based on this comprehensive review. NAD+ homeostasis is integral to fish physiology, influencing processes such as energy metabolism, oxidative stress responses, and cellular signaling. Besides this, NAD+ depletion has been linked to various pathological conditions in fish, paralleling observations in mammals, which suggests conserved mechanisms across vertebrates. Zebrafish models have been pivotal in investigating NAD+-related pathways and their implications in disease processes and potential therapeutic interventions. NAD+ metabolism is intricately connected to fish responses to environmental stressors including pollutants and temperature fluctuations, underscoring its significance in ecological adaptation. The dietary modulation of NAD+ precursors and related pathways can affect fish growth, immunity, and product quality, thereby offering potential applications in aquaculture. On the other hand, NAD+-dependent signaling, particularly through SIRTs, plays a significant role in fish neurobiology and sensory processing. NAD+ metabolism is also involved in fish skin health and pigmentation, which has implications for both ecological and commercial aspects of fish biology. The role of NAD+ in immune regulation and host-pathogen interactions in fish is emerging as a critical area of study, with potential applications in disease management in aquaculture. Furthermore, NAD+ levels and related enzymatic activities in fish eggs can serve as indicators of egg quality, which is crucial for aquaculture and conservation. Finally, postmortem changes in NAD+ levels affect the muscle quality in fish, which has significant implications for the seafood industry. Research on NAD+ in fish biology spans from molecular mechanisms to ecological adaptations and commercial applications, thereby demonstrating the importance of this molecule in aquatic vertebrates. These conclusions underscore the fundamental significance of NAD+ metabolism in fish biology and highlight potential areas for future research and application in various fields related to aquatic sciences and industries.
Further research is necessary to fully understand species-specific variations in NAD+ metabolism and to translate these findings into practical applications in aquaculture, conservation, and seafood technology. Extant research on NAD+-related metabolites in fish underscores their significance in various physiological processes. NAD can be synthesized via both de novo and salvage pathways, both of which have potential benefits for aquaculture through dietary supplementation aimed at enhancing NAD synthesis, although such studies are still in the nascent stages. Strategies to augment NAD+ synthesis include supplementation with NAD+ precursors (e.g., NAM, NA, NR, and NMN), stimulation of NAD+ synthesis enzymes (such as NAMPT), and activation of NQO1. To avert NAD+ depletion, inhibiting NAD+-consuming enzymes (e.g., SIRTs, PARP-1, and CD38) is also feasible. These strategies warrant further exploration in fish to ascertain their efficacy under various conditions. The optimal dosage for each species, timing of administration of possible supplements, long-term effects, and many other factors remain undetermined.
The restoration of NAD+ levels, which has been demonstrated to enhance mitochondrial function and confer protection in animal models, presents a promising therapeutic strategy for mitigating adverse conditions affecting farmed fish production, such as stress, unfavorable environmental conditions, and periods of increased energy expenditure. Additionally, strategies to boost NAD+ concentrations, which are often consistently diminished in pathological conditions leading to ongoing economic challenges in this domain, are of interest.
Several studies have focused on appetite and its possible regulation, thus enhancing our understanding of feeding behavior, which is a crucial subject in aquaculture. These findings offer evolutionary insights into the conserved mechanisms of feeding regulation, suggesting that SIRT-mediated metabolic adaptation may link the peripheral energy status with central appetite pathways in fish. Unfortunately, little attention has been devoted to investigating the effects of these studies on NAD+ metabolism. Similarly, understanding skin pigmentation is vital for enhancing farmed fish production and commercial success, with NAD+ and SIRTs implicated in skin pigmentation. Unfortunately, research focusing on this topic remains scarce.
Research findings suggest that SIRTs function as metabolic coordinators of stress responses. These studies underscore the importance of SIRTs as pivotal molecular hubs in environmental adaptation, effectively linking metabolic and stress response systems in fish. Augmenting NAD+ levels enhances metabolic resilience under stressful conditions. The activation of PPARα can mitigate diet-induced metabolic issues in marine fish by synergizing with SIRT1, thereby offering potential therapeutic strategies for metabolic disorders associated with aquaculture. Specifically, SIRT1 is crucial for regulating the energy balance and maintaining redox stability in economically significant fish species. Given the unique sensory characteristics of marine fish and their extensive species diversity, further investigation of how NAD+ metabolism influences these sensory systems and their interactions with the central nervous system is recommended.
Additionally, the fish by-products analyzed were always rich in niacin (vitamin B₃) precursors, amino acids such as Try, and high-quality proteins, all of which contribute to NAD⁺ biosynthesis through both de novo and salvage pathways. Utilizing these by-products in aquafeeds or functional foods could help sustain NAD⁺ levels in both fish and humans, supporting metabolic health, resistance to oxidative stress, and cellular energy production.
There is still research to be done on the involvement of NAD+ metabolism in various fields of interest, such as reproduction, growth, and metamorphosis in fish. There are multiple reasons to study all these molecules related to NAD metabolism in fish, in order to improve our understanding of the physiological responses of these animals and to develop new strategies to optimize their health and production.
Author Contributions
Conceptualization, M.A.E. and A.S.F.; formal analysis, M.A.E. and A.S.F.; writing—original draft preparation, M.A.E.; writing—review and editing, M.A.E. and A.S.F. Both authors read and agreed to the published version of the manuscript.
Funding
This work was financed by the Spanish MCIN/AEI (grant number PID2024-156529NB-I00).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
No new data were created.
Acknowledgments
This work is part of the ThinkInAzul program supported by MCIN with funding from European Union Next Generation EU (PRTR-C17.I1) and by the Comunidad Autónoma de la Región de Murcia-Fundación Séneca (Spain).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Katsyuba, E.; Romani, M.; Hofer, D.; Auwerx, J. NAD+ homeostasis in health and disease. Nat Metab. 2020, 2, 9–31. [Google Scholar] [CrossRef] [PubMed]
- Houtkooper, R. H.; Cantó, C.; Wanders, R. J.; Auwerx, J. The secret life of NAD+ is an old metabolite that controls new metabolic signaling pathways. Endocr. Rev. 2010, 31, 194–223. [Google Scholar] [CrossRef]
- Davila, A.; Liu, L.; Chellappa, K.; Redpath, P.; Nakamaru-Ogiso, E.; Paolella, L. M.; Zhang, Z.; Migaud, M. E.; Rabinowitz, J. D.; Baur, J. A. Nicotinamide adenine dinucleotide is transported into the mammalian mitochondria. eLife 2018, 7, e33246. [Google Scholar] [CrossRef]
- Cantarow, W.; Stollar, B.D. Nicotinamide mononucleotide adenylyltransferase, a non-histone chromatin protein. Arch. Biochem. Biophys. 1977, 180, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.; Niere, M.; Ziegler, M. The NMN/NaMN adenylyltransferase (NMNAT) protein family. Front. Biosci. (Landmark Ed). 2009, 14, 410–431. [Google Scholar] [CrossRef]
- Su, M.; Qiu, F.; Li, Y.; Che, T.; Li, N.; Zhang, S. Mechanisms of the NAD+ salvage pathway in enhancing skeletal muscle function. Front. Cell Dev. Biol. 2024, 12, 1464815. [Google Scholar] [CrossRef]
- Shahgaldi, S.; Kahmini, F.R. A comprehensive review of Sirtuins: With a major focus on redox homeostasis and metabolism. Life Sci. 2021, 282, 119803. [Google Scholar] [CrossRef]
- Houtkooper, R.H.; Pirinen, E.; Auwerx, J. Sirtuins regulate metabolism and health. Nat. Rev. Mol. Cell Biol. 2012, 13, 225–238. [Google Scholar] [CrossRef]
- Tannous, C.; Booz, G.W.; Altara, R.; Muhieddine, D.H.; Mericskay, M.; Refaat, M.M.; Zouein, F.A. Nicotinamide adenine dinucleotide: biosynthesis, consumption, and therapeutic roles in cardiac diseases. Acta Physiol. 2021, 231, e13551. [Google Scholar] [CrossRef]
- Gariani, K.; Menzies, K.J.; Ryu, D.; Wegner, C.J.; Wang, X.; Ropelle, E.R.; Moullan, N.; Zhang, H.; Perino, A.; Lemos, V.; Kim, B.; Park, Y.K.; Piersigilli, A.; Pham, T.X.; Yang, Y.; Ku, C.S.; Koo, S.I.; Fomitchova, A.; Cantó, C.; Schoonjans, K.; Sauve, A.A.; Lee, J.Y.; Auwerx, J. Eliciting the mitochondrial unfolded protein response by nicotinamide adenine dinucleotide repletion reverses fatty liver disease in mice. Hepatology 2016, 63, 1190–1204. [Google Scholar] [CrossRef] [PubMed]
- Asher, G.; Reinke, H.; Altmeyer, M.; Gutierrez-Arcelus, M.; Hottiger, M. O.; Schibler, U. Poly (ADP-ribose) polymerase 1 participates in phase entrainment of circadian clocks during feeding. Cell 2010, 142, 943–953. [Google Scholar] [CrossRef]
- Abbas, K.; Sharf, R.; Alam, M.; Sharf, Y.; Usmani, N. Chronotherapeutic and epigenetic regulation of circadian rhythms: Nicotinamide adenine dinucleotide-sirtuin axis. J. Sleep Med. 2024, 21, 127–133. [Google Scholar] [CrossRef]
- Araki, T.; Sasaki, Y.; Milbrandt, J. Increased nuclear NAD biosynthesis and SIRT1 activation prevented axonal degeneration. Science 2004, 305, 1010–1013. [Google Scholar] [CrossRef]
- Yang, Y.; Mohammed, F. S.; Zhang, N.; Sauve, A. A. Dihydronicotinamide riboside is a potent NAD+ concentration enhancer both in vitro and in vivo. J. Biol. Chem. 2019, 294, 9295–9307. [Google Scholar] [CrossRef]
- Bian, C.; Zhang, C.; Luo, T.; Vyas, A.; Chen, S. H.; Liu, C.; Kassab, M. A.; Yang, Y.; Kong, M.; Yu, X. NADP+ is an endogenous PARP inhibitor involved in DNA damage response and tumor suppression. Nat. Commun. 2019, 10, 693. [Google Scholar] [CrossRef]
- Katsyuba, E.; Mottis, A.; Zietak, M.; De Franco, F.; van der Velpen, V.; Gariani, K.; Ryu, D.; Cialabrini, L.; Matilainen, O.; Liscio, P.; Giacchè, N.; Stokar-Regenscheit, N.; Legouis, D.; de Seigneux, S.; Ivanisevic, J.; Raffaelli, N.; Schoonjans, K.; Pellicciari, R.; Auwerx, J. De novo NAD+ synthesis enhances mitochondrial function and improves health. Nature 2018, 563, 354–359. [Google Scholar] [CrossRef]
- Canto, C.; Houtkooper, R.H.; Pirinen, E.; Youn, D.Y.; Oosterveer, M.H.; Cen, Y.; Fernandez-Marcos, P.J.; Yamamoto, H.; Andreux, P.A.; Cettour-Rose, P.; et al. The NAD+ precursor nicotinamide riboside enhances oxidative metabolism and protects against high-fat diet-induced obesity. Cell Metab. 2012, 15, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Pitta, M.; Jiang, H.; Lee, J.H.; Zhang, G.; Chen, X.; Kawamoto, E.M.; Mattson, M.P. Nicotinamide forestalls pathology and cognitive decline in Alzheimer mice: Evidence for improved neuronal bioenergetics and autophagy progression. Neurobiol. Aging 2013, 34, 1564–1580. [Google Scholar] [CrossRef]
- Yamamoto, T.; Byun, J.; Zhai, P.; Ikeda, Y.; Oka, S.; Sadoshima, J. Nicotinamide mononucleotide, an intermediate of NAD+ synthesis, protects the heart from ischemia and reperfusion. PLoS One 2014, 9, e98972. [Google Scholar] [CrossRef]
- Khan, N.A.; Auranen, M.; Paetau, I.; Pirinen, E.; Euro, L.; Forsström, S.; Pasila, L.; Velagapudi, V.; Carroll, C.J.; Auwerx, J.; Suomalainen, A. Effective treatment of mitochondrial myopathy by nicotinamide riboside and vitamin B3. EMBO Mol. Med. 2014, 6, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Morigi, M.; Perico, L.; Rota, C.; Longaretti, L.; Conti, S.; Rottoli, D.; Novelli, R.; Remuzzi, G.; Benigni, A. Sirtuin 3-dependent mitochondrial dynamic improvements protect against acute kidney injury. J. Clin. Invest. 2015, 125, 715–726. [Google Scholar] [CrossRef]
- Gariani, K.; Menzies, K.J.; Ryu, D.; Wegner, C.J.; Wang, X.; Ropelle, E.R.; Moullan, N.; Zhang, H.; Perino, A.; Lemos, V.; et al. Eliciting the mitochondrial unfolded protein response by nicotinamide adenine dinucleotide repletion reverses fatty liver disease in mice. Hepatology 2016, 63, 1190–1204. [Google Scholar] [CrossRef]
- Verdin, E. NAD+ in aging, metabolism, and neurodegeneration. Science 2015, 350, 1208–1213. [Google Scholar] [CrossRef]
- Amjad, S.; Nisar, S.; Bhat, A.A.; Shah, A.R.; Frenneaux, M.P.; Fakhro, K.; Haris, M.; Reddy, R.; Patay, Z.; Baur, J.; et al. Role of NAD+ in regulating cellular and metabolic signaling pathways. Mol. Metab. 2021, 49, 101195. [Google Scholar] [CrossRef]
- Yang, S.J.; Choi, J.M.; Kim, L.; Park, S.E.; Rhee, E.J.; Lee, W.Y.; Oh, K.W.; Park, S.W.; Park, C.Y. Nicotinamide improves glucose metabolism and affects the hepatic NAD-sirtuin pathway in a rodent model of obesity and type 2 diabetes. J. Nutr. Biochem. 2014, 25, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Canto, C. NAD+ Precursors: A questionable redundancy. Metabolites 2022, 12, 630. [Google Scholar] [CrossRef]
- Trammell, S.A.J.; Schmidt, M.S.; Weidemann, B.J.; Redpath, P.; Jaksch, F.; Dellinger, R.W.; Li, Z.; Abel, E.D.; Migaud, M.E.; Brenner, C. Nicotinamide riboside is uniquely and orally bioavailable in mice and humans. Nat. Commun. 2016, 7, 12948. [Google Scholar] [CrossRef] [PubMed]
- Alegre, G.F.S.; Pastore, G.M. NAD+ precursors nicotinamide mononucleotide (NMN) and nicotinamide riboside (NR), are potential dietary contributors to health. Curr Nutr Rep. 2023, 12, 445–464. [Google Scholar] [CrossRef]
- Elhassan, Y.S.; Kluckova, K.; Fletcher, R.S.; Schmidt, M.S.; Garten, A.; Doig, C.L.; Cartwright, D.M.; Oakey, L.; Burley, C.V.; Jenkinson, N.; Wilson, M.; Lucas, S.J.E.; Akerman, I.; Seabright, A.; Lai, Y.C.; Tennant, D.A.; Nightingale, P.; Wallis, G.A.; Manolopoulos, K.N.; Brenner, C.; Lavery, G.G. Nicotinamide riboside augments the human skeletal muscle NAD+ metabolome and induces transcriptomic and anti-inflammatory signatures in aged subjects: a placebo-controlled, randomized trial. Cell Rep. 2019, 28, 1717–1728. [Google Scholar] [CrossRef]
- Dollerup, O.L.; Christensen, B.; Svart, M.; Schmidt, M.S.; Sulek, K.; Ringgaard, S.; Stødkilde-Jørgensen, H.; Møller, N.; Brenner, C.; Treebak, J.T.; Jessen, N. A randomized placebo-controlled clinical trial of nicotinamide riboside in obese men: safety, insulin sensitivity, and lipid-mobilizing effects. Am. J. Clin. Nutr. 2018, 108, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Obrador, E.; Salvador-Palmer, R.; Pellicer, B.; López-Blanch, R.; Sirerol, J.A.; Villaescusa, J.I.; Montoro, A.; Dellinger, R.W.; Estrela, J.M. The combination of natural polyphenols with a precursor of NAD+ and a TLR2/6 ligand lipopeptide protects mice against lethal γ-radiation. J. Adv. Res. 2023, 45, 73–86. [Google Scholar]
- Espinoza, S.E.; Khosla, S.; Baur, J.A.; de Cabo, R.; Musi, N. Drugs targeting mechanisms of aging to delay age-related disease and promote health span: Proceedings of a National Institute on Aging Workshop. J. Gerontol. A Biol. Sci. Med. Sci. 2023, 78, 53–60. [Google Scholar] [CrossRef]
- Yang, X.; Lu, A.; Guan, X.; Ying, T.; Pan, J.; Tan, M.; Lu, J. An updated review on the mechanisms, pre-clinical and clinical comparisons of Nicotinamide Mononucleotide (NMN) and Nicotinamide Riboside (NR). Food Front. 2024, 6, 630–643. [Google Scholar]
- Schlegel, A.; Stainier, D.Y.R. Lessons from "lower" organisms: What worms, flies, and zebrafish can teach us about human energy metabolism. PLoS Genet. 2007, 3, 2037–2048. [Google Scholar]
- Prisingkorn, W.; Prathomya, P.; Jakovlic, I.; Liu, H.; Zhao, Y.H.; Wang, W.M. Transcriptomics, metabolomics and histology indicate that high-carbohydrate diet negatively affects the liver health of blunt snout bream (Megalobrama amblycephala). BMC Genom. 2017, 18, 856. [Google Scholar] [CrossRef]
- Kamel, M.; Ninov, N. Catching new targets in metabolic disease with a zebrafish. Curr. Opin. Pharm. 2017, 37, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Zang, L.; Maddison, L.A.; Chen, W. Zebrafish as a model for obesity and diabetes. Front. Cell Dev. Biol. 2018, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- Asaoka, Y.; Terai, S.; Sakaida, I.; Nishina, H. The expanding role of fish models in understanding non-alcoholic fatty liver disease. Dis. Model. Mech. 2013, 6, 905–914. [Google Scholar] [CrossRef]
- Oka, T.; Nishimura, Y.; Zang, L.; Hirano, M.; Shimada, Y.; Wang, Z.; Umemoto, N.; Kuroyanagi, J.; Nishimura, N.; Tanaka, T. Diet-induced obesity in zebrafish shares common pathophysiological pathways with mammalian obesity. BMC Physiol. 2010, 10, 21. [Google Scholar] [CrossRef]
- Goody, M.F.; Kelly, M.W.; Lessard, K.N.; Khalil, A.; Henry, C.A. Nrk2b-mediated NAD+ production regulates cell adhesion and is required for muscle morphogenesis in vivo: Nrk2b and NAD+ in muscle morphogenesis. Dev. Biol. 2010, 344, 809–826. [Google Scholar] [CrossRef] [PubMed]
- Goody, M.F.; Henry, C.A. A need for NAD+ in muscle development, homeostasis, and aging. Skeletal Muscle 2018, 8, 9. [Google Scholar] [CrossRef]
- Goody, M.F.; Kelly, M.W.; Reynolds, C.J.; Khalil, A.; Crawford, B.D.; Henry, C.A. NAD+ biosynthesis ameliorates a zebrafish model of muscular dystrophy. PLoS Biol. 2012, 10, e1001409. [Google Scholar] [CrossRef]
- Ran, G.; Ying, L.; Li, L.; Yan, Q.; Yi, W.; Ying, C.; Wu, H.; Ye, X. Resveratrol ameliorates diet-induced dysregulation of lipid metabolism in zebrafish (Danio rerio). PLoS One 2017, 12, e0180865. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.C.; Gregório, C.; Uribe-Cruz, C.; Guizzo, R.; Malysz, T.; Faccioni-Heuser, M.C.; Longo, L.; da Silveira, T.R. Chronic exposure to ethanol causes steatosis and inflammation in zebrafish liver. World J. Hepatol. 2017, 9, 418–426. [Google Scholar] [CrossRef]
- Park, K.H.; Kim, S.H. Adult zebrafish as an in vivo drug-testing model for ethanol-induced acute hepatic injury. Biomed. Pharmacother. 2020, 132, 110836. [Google Scholar] [CrossRef]
- Martínez-Morcillo, F.J.; Cantón-Sandoval, J.; Martínez-Navarro, F.J.; Cabas, I.; Martínez-Vicente, I.; Armistead, J.; Hatzold, J.; López-Muñoz, A.; Martínez-Menchón, T.; Corbalán-Vélez, R.; Lacal, J.; Hammerschmidt, M.; García-Borrón, J.C.; García-Ayala, A.; Cayuela, M.L.; Pérez-Oliva, A.B.; García-Moreno, D.; Mulero, V. NAMPT-derived NAD+ fuels PARP1 to promote skin inflammation via parthanatos. PLoS Biol. 2021, 19, e3001455. [Google Scholar] [CrossRef]
- Ding, Q.; Zhang, Z.; Li, Y.; Liu, H.; Hao, Q.; Yang, Y.; Ringø, E.; Olsen, R.E.; Clarke, J.L.; Ran, C.; Zhou, Z. Propionate induces intestinal oxidative stress via Sod2 propionylation in zebrafish. iScience. 2021, 24, 102515. [Google Scholar] [CrossRef]
- Zhang, D.; Zhao, L.; Wong, L. β-Nicotinamide mononucleotide supplement with astaxanthin and blood orange enhanced NAD+ bioavailability and mitigated age-associated physiological decline in zebrafish. Curr Dev Nutr. 2022, 6, 54. [Google Scholar] [CrossRef]
- Yang, L.; Zhu, B.; Zhou, S.; Zhao, M.; Li, R.; Zhou, Y.; Shi, X.; Han, J.; Zhang, W.; Zhou, B. Mitochondrial dysfunction is involved in decabromodiphenyl ethane-induced lipid metabolism disorders and neurotoxicity in zebrafish larvae. Environ. Sci. Technol. 2023, 57, 11043–11055. [Google Scholar] [CrossRef] [PubMed]
- Pandaram, A.; Paul, J.; Wankhar, W.; Thakur, A.; Verma, S.; Vasudevan, K.; Wankhar, D.; Kammala, A.K.; Sharma, P.; Jaganathan, R.; Iyaswamy, A.; Rajan, R. Aspartame causes developmental defects and teratogenicity in zebrafish embryo: Role of impaired SIRT1/FOXO3a axis in neuron cells. Biomedicines 2024, 12, 855. [Google Scholar] [CrossRef]
- Tsurho, V.; Gilliland, C.; Ensing, J.; Vansickle, E.; Lanning, N.J.; Mark, P.R.; Grainger, S. A zebrafish model of nicotinamide adenine dinucleotide (NAD+) deficiency-derived congenital disorders. Preprint https://www.researchgate.net/publication/387907417 [accessed Aug 06 2025]. 2025.
- Xu, H.; Mao, X.; Zhang, S.; Ren, J.; Jiang, S.; Cai, L.; Miao, X.; Tao, Y.; Peng, C.; Lv, M.; Li, Y. Perfluorooctanoic acid triggers premature ovarian insufficiency by impairing NAD+ synthesis and mitochondrial function in adult zebrafish. Toxicol. Sci. 2024, 201, 118–128. [Google Scholar] [CrossRef]
- Wan, Y.; Li, J.; Guo, Y.; Guo, F.; Zhao, Y.; Li, Y.; Yang, X.; Chen, H.; Xie, S.; Wang, M.; Guan, G.; Zhu, Y.; Li, X. Identification and functional characterization of a novel PRPS1 variant in X-linked nonsyndromic hearing loss: Insights from zebrafish and cellular models. Hum. Mutat. 2025, 2025, 6690588. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Yang, Y.; Xian, C.; Zhou, P.; Zhang, H.; Lv, Z.; Liu, J. Nicotinamide riboside ameliorates survival time and motor dysfunction in an MPTP-Induced Parkinson's disease zebrafish model through effects on glucose metabolism and endoplasmic reticulum stress. Chem. Biol. Interact. 2024, 399, 111118. [Google Scholar] [CrossRef]
- Raas, Q.; Haouy, G.; de Calbiac, H.; Pasho, E.; Marian, A.; Guerrera, I.C.; Rosello, M.; Oeckl, P.; Del Bene, F.; Catanese, A.; Ciura, S.; Kabashi, E. TBK1 is involved in programmed cell death and ALS-related pathways in novel zebrafish models. Cell Death Discovery 2025, 11, 98. [Google Scholar] [CrossRef]
- Shimeno, S.; Saida, Y.; Tabata, T. Response of hepatic NAD- and NADP-isocitrate dehydrogenase activities to several dietary conditions in fishes. Nippon Suisan Gakk. 1996, 62, 642–648. [Google Scholar] [CrossRef]
- Carafa, V.; Rotili, D.; Forgione, M.; Cuomo, F.; Serretiello, E.; Hailu, G.S.; Jarho, E.; Lahtela-Kakkonen, M.; Mai, A.; Altucci, L. Sirtuin functions and modulation: from chemistry to the clinic. Clin Epigenetics 2016, 8, 61. [Google Scholar] [CrossRef] [PubMed]
- Kabiljo, J.; Murko, C.; Pusch, O.; Zupkovitz, G. Spatio-temporal expression profile of sirtuins during aging of the annual fish Nothobranchius furzeri. Gene Expr. Patterns. 2019, 33, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jia, E.; Shi, H.; Li, X.; Jiang, G.; Chi, C.; Liu, W.; Zhang, D. Selection of reference genes for miRNA quantitative PCR and its application in miR-34a/Sirtuin-1 mediated energy metabolism in Megalobrama amblycephala. Fish Physiol. Biochem. 2019, 45, 1663–1681. [Google Scholar] [CrossRef]
- Simó-Mirabet, P.; Perera, E.; Calduch-Giner, J.A.; Pérez-Sánchez, J. Local DNA methylation helps to regulate muscle sirtuin 1 gene expression across seasons and advancing age in gilthead sea bream (Sparus aurata). Front. Zool. 2020, 17, 15. [Google Scholar] [CrossRef]
- Simó-Mirabet, P.; Naya-Català, F.; Calduch-Giner, J.A.; Pérez-Sánchez, J. The expansion of sirtuin gene family in gilthead sea bream (Sparus aurata). Phylogenetic, syntenic, and functional insights across the vertebrate/fish lineage. Int. J. Mol. Sci. 2024, 25, 6273. [Google Scholar] [CrossRef]
- Peng, J.; Zou, J.; Zhao, J.; Chen, A. Genome-wide identification and expression analysis of the SIRT gene family in Nile tilapia (Oreochromis niloticus). Comp Biochem Physiol Part D Genomics Proteomics. 2025, 54, 101425. [Google Scholar] [CrossRef]
- Liu, Q.; Qi, Y.; Liang, Q.; et al. The chimeric genes in the hybrid lineage of Carassius auratus cuvieri (♀) × Carassius auratus red var. (♂). Sci. China Life Sci. 2018, 6, 1079–1089. [Google Scholar] [CrossRef]
- Tang, Y.; Liu, X.; Feng, C.; Zhou, Z.; Liu, S. Nicotinamide phosphoribosyl transferase (Nampt) of hybrid crucian carp protects intestinal barrier and enhances host immune defense against bacterial infection. Dev. Comp. Immunol. 2022, 128, 104314. [Google Scholar] [CrossRef]
- Butler, M.J.; Volkoff, H. The role of visfatin / NAMPT in the regulation of feeding in goldfish (Carassius auratus). Peptides 2023, 160, 170919. [Google Scholar] [CrossRef]
- Velasco, C.; Librán-Pérez, M.; Otero-Rodiño, C.; López-Patiño, M.A.; Míguez, J.M.; Soengas, J.L. Ceramides are involved in the regulation of food intake in rainbow trout (Oncorhynchus mykiss). Am. J. Physiol. Regul. Integr. Comp Physiol. 2016, 311, R658–R668. [Google Scholar] [CrossRef]
- Vinnicombe, K.R.T.; Volkoff, H. Possible role of transcription factors (BSX, NKX2.1, IRX3 and SIRT1) in the regulation of appetite in goldfish (Carassius auratus). Comp Biochem Physiol A Mol Integr Physiol. 2022, 268, 111189. [Google Scholar] [CrossRef]
- Fuentes, E.N.; Zuloaga, R.; Nardocci, G.; Fernandez de la Reguera, C.; Simonet, N.; Fumeron, R.; Valdes, J.A.; Molina, A.; Alvarez, M. Skeletal muscle plasticity induced by seasonal acclimatization in carp involves differential expression of rRNA and molecules that epigenetically regulate its synthesis. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2014, 172-173, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Teigen, L.E.; Orczewska, J.I.; McLaughlin, J.; O'Brien, K.M. Cold acclimation increases levels of some heat shock protein and sirtuin isoforms in threespine stickleback. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2015, 188, 139–147. [Google Scholar] [CrossRef]
- Ekambaram, P.; Parasuraman, P. Differential expression of sirtuin 2 and adipocyte maturation restriction: an adaptation process during hypoxia in fish. Biol Open. 2017, 6, 1375–1382. [Google Scholar] [CrossRef]
- Naderi, F.; Hernández-Pérez, J.; Chivite, M.; Soengas, J.L.; Míguez, J.M.; López-Patiño, M.A. Involvement of cortisol and sirtuin1 during the response to stress of hypothalamic circadian system and food intake-related peptides in rainbow trout, Oncorhynchus mykiss. Chronobiol. Int. 2018, 35, 1122–1141. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Kuang, J.; Liu, X.; Li, A.; Feng, W.; Zhuang, Z. Effects of osmotic stress on Na<sup>+</sup>/K<sup>+</sup>-ATPase, caspase 3/7 activity, and the expression profiling of sirt1, hsf1, and hsp70 in the roughskin sculpin (Trachidermus fasciatus). Fish Physiol. Biochem. 2020, 46, 135–144. [Google Scholar] [CrossRef]
- Qian, L.; Miao, L.; Abba, B.S.A.; Lin, Y.; Jiang, W.; Chen, S.; Luo, C.; Liu, B.; Ge, X. Molecular characterization and expression of sirtuin 2, sirtuin 3, and sirtuin 5 in the Wuchang bream (Megalobrama amblycephala) in response to acute temperature and ammonia nitrogen stress. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2021, 252, 110520. [Google Scholar] [CrossRef]
- Li, M.; Hu, J.; Zhou, J.; Wu, C.; Li, D.; Mao, H.; Kong, L.; Hu, C.; Xu, X. Grass carp (Ctenopharyngodon idella) deacetylase SIRT1 targets p53 to suppress apoptosis in a KAT8 dependent or independent manner. Fish Shellfish Immunol. 2024, 144, 109264. [Google Scholar] [CrossRef]
- Machala, M.; Dusek, L.; Hilscherová, K.; Kubínová, R.; Jurajda, P.; Neca, J.; Ulrich, R.; Gelnar, M.; Studnicková, Z.; Holoubek, I. Determination and multivariate statistical analysis of biochemical responses to environmental contaminants in feral freshwater fish Leuciscus cephalus, L. Environ. Toxicol. Chem. 2001, 20, 1141–1148. [Google Scholar]
- Berg, K.; Puntervoll, P.; Klungsøyr, J.; Goksøyr, A. Brain proteome alterations of Atlantic cod (Gadus morhua) exposed to PCB 153. Aquat. Toxicol. 2011, 105, 206–217. [Google Scholar] [CrossRef]
- Riar, N.; Crago, J.; Jiang, W.; Maryoung, L.A.; Gan, J.; Schlenk, D. Effects of salinity acclimation on the endocrine disruption and acute toxicity of bifenthrin in freshwater and euryhaline strains of Oncorhynchus mykiss. Environ. Toxicol. Chem. 2013, 32, 2779–2785. [Google Scholar] [CrossRef]
- Garattini, E.; Mendel, R.; Romão, M.J.; Wright, R.; Terao, M. Mammalian molybdo-flavoenzymes, an expanding family of proteins: structure, genetics, regulation, function and pathophysiology. Biochem. J. 2003, 372, 15–32. [Google Scholar] [CrossRef]
- Aburas, O.A. Investigation of aldehyde oxidase and xanthine oxidoreductase in rainbow trout (Oncorhynchus mykiss). Doctoral Thesis, University of Huddersfield. 2014. Available at http://eprints.hud.ac.uk/id/eprint/23543/.
- Almeida, Â.; Silva, M.G.; Soares, A.M.V.M.; Freitas, R. Concentrations levels and effects of 17alpha-Ethinylestradiol in freshwater and marine waters and bivalves: A review. Environ. Res. 2020, 185, 109316. [Google Scholar] [CrossRef]
- Zhou, X.; Li, Y.; Li, H.; Yang, Z.; Zuo, C. Responses in the crucian carp (Carassius auratus) exposed to environmentally relevant concentration of 17α-Ethinylestradiol based on metabolomics. Ecotoxicol. Environ. Saf. 2019, 183, 109501. [Google Scholar] [CrossRef]
- Taheri Mirghaed, A.; Baes, M.; Hoseini, S.M. Humoral immune responses and gill antioxidant-related gene expression of common carp (Cyprinus carpio) exposed to lufenuron and flonicamide. Fish Physiol. Biochem. 2020, 46, 739–746. [Google Scholar] [CrossRef]
- Jabłońska-Trypuć, A. A review on triclosan in wastewater: Mechanism of action, resistance phenomenon, environmental risks, and sustainable removal techniques. Water Environ Res. 2023, 95, e10920. [Google Scholar] [CrossRef]
- Bao, S.; He, C.; Ku, P.; Xie, M.; Lin, J.; Lu, S.; Nie, X. Effects of triclosan on the RedoximiRs/Sirtuin/Nrf2/ARE signaling pathway in mosquitofish (Gambusia affinis). Aquat. Toxicol. 2021, 230, 105679. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, L.; Chen, J. Paracetamol in the environment and its degradation by microorganisms. Appl. Microbiol. Biotechnol. 2012, 96, 875–884. [Google Scholar] [CrossRef]
- Meinan, X.; Yimeng, W.; Chao, W.; Tianli, T.; Li, J.; Peng, Y.; Xiangping, N. Response of the Sirtuin/PXR signaling pathway in Mugilogobius chulae exposed to environmentally relevant concentration Paracetamol. Aquat. Toxicol. 2022, 249, 106222. [Google Scholar] [CrossRef]
- Vargas, R.; Ponce-Canchihuamán, J. Emerging various environmental threats to brain and overview of surveillance system with zebrafish model. Toxicol. Rep. 2017, 4, 467–473. [Google Scholar] [CrossRef]
- Zhao, Y.; Xie, M.; Wang, C.; Wang, Y.; Peng, Y.; Nie, X. Effects of atorvastatin on the Sirtuin/PXR signaling pathway in Mugilogobius chulae. Environ. Sci. Pollut. Res. Int. 2023, 30, 60009–60022. [Google Scholar] [CrossRef]
- Iacopetta, D.; Catalano, A.; Ceramella, J.; Saturnino, C.; Salvagno, L.; Ielo, I.; Drommi, D.; Scali, E.; Plutino, M.R.; Rosace, G.; Sinicropi, M.S. The Different facets of triclocarban: A review. Molecules. 2021, 26, 2811. [Google Scholar] [CrossRef]
- Li, S.; Wang, Y.; Yu, D.; Zhang, Y.; Wang, X.; Shi, M.; Xiao, Y.; Li, X.; Xiao, H.; Chen, L.; Xiong, X. Triclocarban evoked neutrophil extracellular trap formation in common carp (Cyprinus carpio L.) by modulating SIRT3-mediated ROS crosstalk with ERK1/2/p38 signaling. Fish Shellfish Immunol. 2022, 129, 85–95. [Google Scholar] [CrossRef]
- Magnuson, J.T.; Sydnes, M.O.; Ræder, E.M.; Schlenk, D.; Pampanin, D.M. Transcriptomic profiles of brains in juvenile Atlantic cod (Gadus morhua) exposed to pharmaceuticals and personal care products from a wastewater treatment plant discharge. Sci. Total Environ. 2024, 912, 169110. [Google Scholar] [CrossRef]
- Chen, S.; Wang, Y.; Chen, K.; Xing, X.; Jiang, Q.; Xu, T. Unraveling the mechanism of quercetin alleviating BHPF-induced apoptosis in epithelioma papulosum cyprini cells: SIRT3-mediated mitophagy. Fish Shellfish Immunol. 2024, 154, 109907. [Google Scholar] [CrossRef]
- Connon, R.E.; Deanovic, L.A.; Fritsch, E.B.; D'Abronzo, L.S.; Werner, I. Sublethal responses to ammonia exposure in the endangered delta smelt; Hypomesus transpacificus (Fam. Osmeridae). Aquat Toxicol. 2011, 105, 369–377. [Google Scholar] [CrossRef]
- Jia, R.; Du, J.; Cao, L.; Feng, W.; He, Q.; Xu, P.; Yin, G. Application of transcriptome analysis to understand the adverse effects of hydrogen peroxide exposure on brain function in common carp (Cyprinus carpio). Environ. Pollut. 2021, 286, 117240. [Google Scholar] [CrossRef]
- Berg, K.; Puntervoll, P.; Valdersnes, S.; Goksøyr, A. Responses in the brain proteome of Atlantic cod (Gadus morhua) exposed to methylmercury. Aquat Toxicol. 2010, 100, 51–65. [Google Scholar] [CrossRef]
- Li, H.; Di, G.; Zhang, Y.; Liang, J.; Wang, X.; Xu, Z.; Kong, X. miR-217 through SIRT1 regulates the immunotoxicity of cadmium in Cyprinus carpio. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 248, 109086. [Google Scholar] [CrossRef]
- Wei, X.; Hogstrand, C.; Chen, G.; Lv, W.; Song, Y.; Xu, Y.; Luo, Z. Zn Induces lipophagy via the deacetylation of beclin1 and alleviates Cu-induced lipotoxicity at their environmentally relevant concentrations. Environ. Sci. Technol. 2021, 55, 4943–4953. [Google Scholar] [CrossRef]
- Hong, H.; Liu, Z.; Li, S.; Wu, D.; Jiang, L.; Li, P.; Wu, Z.; Xu, J.; Jiang, A.; Zhang, Y.; Wei, Z.; Yang, Z. Zinc oxide nanoparticles (ZnO-NPs) exhibit immune toxicity to crucian carp (Cyprinus carpio). by neutrophil extracellular traps (NETs) release and oxidative stress. Fish Shellfish Immunol. 2022, 129, 22–29. [Google Scholar] [CrossRef]
- Lall, S.P.; Kaushik, S. Nutrition and metabolism of minerals in fish. Animal (Basel) 2021, 11, 2711. [Google Scholar] [CrossRef]
- Wagemann, R.; Dick, J.G.; Klaverkamp, J.F. Metallothionein estimates in marine mammal and fish tissues by three methods: 203Hg displacement, polarography and metal-Summation. Int. J. Environ. Anal. Chem. 1994, 54, 147–160. [Google Scholar] [CrossRef]
- Zhong, C.C.; Zhang, X.; Pantopoulos, K.; Song, C.C.; Yang, H.; Wei, X.L.; Luo, Z. Mitochondrial oxidative stress inhibited Sirt3/Foxo3/PPARα pathway and aggravated copper and zinc co-deficiency-induced hepatic lipotoxicity in a fish model. Cell Mol. Life Sci. 2025, 82, 226. [Google Scholar] [CrossRef]
- Wang, M.C.; Wang, Y.C.; Peng, H.W.; Hseu, J.R.; Wu, G.C.; Chang, C.F.; Tseng, Y.C. Resveratrol induces expression of metabolic and antioxidant machinery and protects tilapia under cold stress. Int. J. Mol. Sci. 2020, 21, 3338. [Google Scholar] [CrossRef]
- Wu, L.; Chen, Q.; Dong, B.; Han, D.; Zhu, X.; Liu, H.; Yang, Y.; Xie, S.; Jin, J. Resveratrol attenuated oxidative stress and inflammatory and mitochondrial dysfunction induced by acute ammonia exposure in gibel carp (Carassius gibelio). Ecotoxicol. Environ. Saf. 2023, 251, 114544. [Google Scholar] [CrossRef]
- Xia, Y.; Li, S.; Wang, X.; Zhao, B.; Chen, S.; Jiang, Q.; Xu, S.; Li, S. Astilbin targeted Sirt1 to inhibit acetylation of Nrf2 to alleviate grass carp hepatocyte apoptosis caused by PCB126-induced mitochondrial kinetic and metabolism dysfunctions. Fish Shellfish Immunol. 2023, 141, 109000. [Google Scholar] [CrossRef]
- Otero-Rodiño, C.; Librán-Pérez, M.; Velasco, C.; Álvarez-Otero, R.; López-Patiño, M.A.; Míguez, J.M.; Soengas, J.L. Glucosensing in liver and Brockmann bodies of rainbow trout through glucokinase-independent mechanisms. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2016, 199, 29–42. [Google Scholar] [CrossRef]
- Dai, Y.J.; Liu, W.B.; Li, X.F.; Zhou, M.; Xu, C.; Qian, Y.; Jiang, G.Z. Molecular cloning of adipose triglyceride lipase (ATGL) gene from blunt snout bream and its expression after LPS-induced TNF-α factor. Fish Physiol. Biochem. 2018, 44, 1143–1157. [Google Scholar] [CrossRef]
- Xu, C.; Liu, W.B.; Remø, S.C.; Wang, B.K.; Shi, H.J.; Zhang, L.; Liu, J.D.; Li, X.F. Feeding restriction alleviates high carbohydrate diet-induced oxidative stress and inflammation of Megalobrama amblycephala by activating the AMPK-SIRT1 pathway. Fish Shellfish Immunol. 2019, 92, 637–648. [Google Scholar] [CrossRef]
- Wang, T.; Yang, B.; Ji, R.; Xu, W.; Mai, K.; Ai, Q. Omega-3 polyunsaturated fatty acids alleviate hepatic steatosis-induced inflammation through Sirt1-mediated nuclear translocation of NF-κB p65 subunit in hepatocytes of large yellow croaker (Larmichthys crocea). Fish Shellfish Immunol. 2017, 71, 76–82. [Google Scholar] [CrossRef]
- Saha, M.; Dhiman, S.; Gupta, G.D.; Asati, V. An investigative review for pharmaceutical analysis of fenofibrate. J. Chromatogr. Sci. 2023, 61, 494–504. [Google Scholar] [CrossRef]
- Jin, M.; Zhu, T.; Tocher, D.R.; Luo, J.; Shen, Y.; Li, X.; Pan, T.; Yuan, Y.; Betancor, M.B.; Jiao, L.; Sun, P.; Zhou, Q. Dietary fenofibrate attenuated high-fat-diet-induced lipid accumulation and inflammation response partly through regulation of pparα and sirt1 in juvenile black seabream (Acanthopagrus schlegelii). Dev. Comp. Immunol. 2020, 109, 103691. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, S.; Meng, X.; Chen, N.; Li, S. Molecular cloning and characterization of sirtuin 1 and its potential regulation of lipid metabolism and antioxidant response in largemouth bass (Micropterus salmoides). Front Physiol. 2021, 12, 726877. [Google Scholar] [CrossRef]
- Arumugam, M.K.; Paal, M.C.; Donohue, T.M. Jr.; Ganesan, M.; Osna, N.A.; Kharbanda, K.K. Beneficial effects of betaine: A comprehensive review. Biology (Basel) 2021, 10, 456. [Google Scholar] [CrossRef]
- Jin, M.; Shen, Y.; Pan, T.; Zhu, T.; Li, X.; Xu, F.; Betancor, M.B.; Jiao, L.; Tocher, D.R.; Zhou, Q. Dietary betaine mitigates hepatic steatosis and inflammation induced by a high-fat-diet by modulating the Sirt1/Srebp-1/Pparɑ pathway in juvenile black seabream (Acanthopagrus schlegeli). Front. Immunol. 2021, 12, 694720. [Google Scholar] [CrossRef] [PubMed]
- Raymundo, D.P.; Doultsinos, D.; Guillory, X.; Carlesso, A.; Eriksson, L.A.; Chevet, E. Pharmacological targeting of IRE1 in cancer. Trends Cancer 2020, 6, 1018–1030. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Shen, Y.; Monroig, Ó.; Zhao, W.; Bao, Y.; Zhu, T.; Tocher, D.R.; Zhou, Q. Sirt1 mitigates hepatic lipotoxic injury induced by high-fat-diet in fish through Ire1α deacetylation. J. Nutr. 2024, 154, 3210–3224. [Google Scholar] [CrossRef]
- Luthuli, S.; Wu, S.; Cheng, Y.; Zheng, X.; Wu, M.; Tong, H. Therapeutic effects of fucoidan: A review on recent studies. Mar. Drugs. 2019, 17, 487. [Google Scholar] [CrossRef]
- Zhao, W.; Shen, Y.; Bao, Y.; Monroig, Ó.; Zhu, T.; Sun, P.; Tocher, D.R.; Zhou, Q.; Jin, M. Fucoidan alleviates hepatic lipid deposition by modulating the Perk-Eif2α-Atf4 axis via Sirt1 activation in Acanthopagrus schlegelii. Int. J. Biol. Macromol. 2024, 282, 137266. [Google Scholar] [CrossRef]
- Radaelli, G.; Domeneghini, C.; Arrighi, S.; Mascarello, F.; Veggetti, A. Different putative neuromodulators are present in the nerves which distribute to the teleost skeletal muscle. Histol. Histopathol. 1998, 13, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xu, J.; Lazarovici, P.; Quirion, R.; Zheng, W. cAMP response element-binding protein (CREB): A possible signaling molecule link in the pathophysiology of schizophrenia. Front. Mol. Neurosci. 2018, 11, 255. [Google Scholar] [CrossRef]
- Rajan, K.E.; Thangaleela, S.; Balasundaram, C. Spatial learning associated with stimulus response in goldfish Carassius auratus: relationship to activation of CREB signalling. Fish Physiol. Biochem. 2015, 41, 685–694. [Google Scholar] [CrossRef]
- Thangaleela, S.; Shanmugapriya, V.; Mukilan, M.; Radhakrishnan, K.; Rajan, K.E. Alterations in MicroRNA-132/212 expression impairs fear memory in goldfish Carassius auratus. Ann. Neurosci. 2018, 25, 90–97. [Google Scholar] [CrossRef]
- Pérez-Sánchez, J.; Simó-Mirabet, P.; Naya-Català, F.; Martos-Sitcha, J.A.; Perera, E.; Bermejo-Nogales, A.; Benedito-Palos, L.; Calduch-Giner, J.A. Somatotropic axis regulation unravels the differential effects of nutritional and environmental factors in growth performance of marine farmed fishes. Front. Endocrinol. 2018, 9, 687. [Google Scholar] [CrossRef]
- Anken, R.H.; Rahmann, H. An atlas of the distribution of NADPH-diaphorase in the brain of the highly derived swordtail fish Xiphophorus helleri (Atherinoformes: Teleostei). J. Hirnforsch. 1996, 37, 421–449. [Google Scholar] [PubMed]
- Funakoshi, K.; Kadota, T.; Atobe, Y.; Goris, R.C.; Kishida, R. NADPH-diaphorase activity in the vagal afferent pathway of the dogfish, Triakis scyllia. Neurosci. Lett. 1997, 237, 129–132. [Google Scholar] [CrossRef]
- Villani, L. Development of NADPH-diaphorase activity in the central nervous system of the cichlid fish, Tilapia mariae. Brain Behav. Evol. 1999, 54, 147–158. [Google Scholar] [CrossRef]
- Jadhao, A.G.; Malz, C.R. Nicotinamide adenine dinucleotide phosphate (NADPH)-diaphorase activity in the brain of a cichlid fish, with remarkable findings in the entopeduncular nucleus: a histochemical study. J. Chem. Neuroanat. 2004, 27, 75–86. [Google Scholar] [CrossRef]
- Giraldez-Perez, R.M.; Gaytan, S.P.; Ruano, D.; Torres, B.; Pasaro, R. Distribution of NADPH-diaphorase and nitric oxide synthase reactivity in the central nervous system of the goldfish (Carassius auratus). J. Chem. Neuroanat. 2008, 35, 12–32. [Google Scholar] [CrossRef]
- Funakoshi, K.; Kadota, T.; Atobe, Y.; Nakano, M.; Goris, R.C.; Kishida, R. Nitric oxide synthase in the glossopharyngeal and vagal afferent pathway of a teleost, Takifugu niphobles. The branchial vascular innervation. Cell Tissue Res. 1999, 298, 45–54. [Google Scholar] [CrossRef]
- Andrews, C.D.; Payne, J.F.; Rise, M.L. Identification of a gene set to evaluate the potential effects of loud sounds from seismic surveys on the ears of fishes: a study with Salmo salar. J. Fish Biol. 2014, 84, 1793–1819. [Google Scholar] [CrossRef]
- Simó-Mirabet, P.; Bermejo-Nogales, A.; Calduch-Giner, J.A.; Pérez-Sánchez, J. Tissue-specific gene expression and fasting regulation of sirtuin family in gilthead sea bream (Sparus aurata). J. Comp. Physiol. B 2017, 187, 153–163. [Google Scholar] [CrossRef]
- Jiang, W.; Lin, Y.; Qian, L.; Miao, L.; Liu, B.; Ge, X.; Shen, H. Mulberry leaf meal: A potential feed supplement for juvenile Megalobrama amblycephala "Huahai No. 1". Fish Shellfish Immunol. 2022, 128, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Silva-Marrero, J.I.; Villasante, J.; Rashidpour, A.; Palma, M.; Fàbregas, A.; Almajano, M.P.; Viegas, I.; Jones, J.G.; Miñarro, M.; Ticó, J.R.; Baanante, I.V.; Metón, I. The administration of chitosan-tripolyphosphate-DNA nanoparticles to express exogenous SREBP1a enhances conversion of dietary carbohydrates into lipids in the liver of Sparus aurata. Biomolecules 2019, 9, 297. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Huang, X.P.; Guan, J.F.; Chen, Z.M.; Ma, Y.C.; Xie, D.Z.; Ning, L.J.; Li, Y.Y. Effects of dietary leucine and valine levels on growth performance, glycolipid metabolism and immune response in Tilapia GIFT Oreochromis niloticus. Fish Shellfish Immunol. 2022, 121, 395–403. [Google Scholar] [CrossRef]
- Sun, S.S.; Feng, L.; Jiang, W.D.; Liu, Y.; Ren, H.M.; Jin, X.W.; Zhou, X.Q.; Wu, P. Declined flesh quality resulting from niacin deficiency is associated with elevated glycolysis and impaired mitochondrial homeostasis in grass carp (Ctenopharyngodon idella). Food Chem. 2024, 451, 139426. [Google Scholar] [CrossRef] [PubMed]
- Kandyliari, A.; Mallouchos, A.; Papandroulakis, N.; Golla, J.P.; Lam, T.T.; Sakellari, A.; Karavoltsos, S.; Vasiliou, V.; Kapsokefalou, M. Nutrient composition and fatty acid and protein profiles of selected fish by-products. Foods 2020, 9, 190. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Liu, W.B.; Shi, H.J.; Mi, H.F.; Li, X.F. Benfotiamine ameliorates high-carbohydrate diet-induced hepatic oxidative stress, inflammation and apoptosis in Megalobrama amblycephala. Aquacult. Res. 2021, 52, 3174–3185. [Google Scholar] [CrossRef]
- Shi, H.J.; Xu, C.; Liu, M.Y.; Wang, B.K.; Liu, W.B.; Chen, D.H.; Zhang, L.; Xu, C.Y.; Li, X.F. Resveratrol improves the energy sensing and glycolipid metabolism of blunt snout bream Megalobrama amblycephala fed high-carbohydrate diets by activating the AMPK-SIRT1-PGC-1 network. Front. Physiol. 2018, 9, 1258. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, X.; Wei, L.; Liu, Z.; Chu, X.; Xiong, W.; Liu, W.; Li, X. The effectiveness of four Nicotinamide Adenine Dinucleotide (NAD+) precursors in alleviating the high-glucose-induced damage to hepatocytes in Megalobrama amblycephala: Evidence in NAD+ homeostasis, Sirt1/3 activation, redox defense, inflammatory response, apoptosis, and glucose metabolism. Antioxidants 2024, 13, 385. [Google Scholar]
- Wachira, M.N.; Osuga, I.M.; Munguti, J.M.; Ambula, M.K.; Subramanian, S.; Tanga, C.M. Efficiency and improved profitability of insect-based aquafeeds for farming Nile tilapia fish (Oreochromis niloticus L.). Animals 2021, 11, 2599. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kwak, K.W.; Park, E.S.; Yoon, H.J.; Kim, Y.S.; Park, K.; et al. Evaluation of subchronic oral dose toxicity of freeze-dried skimmed powder of Zophobas atratus larvae (frpfdZAL) in rats. Foods (Basel, Switzerland). 2020, 9, 995. [Google Scholar] [CrossRef]
- van Broekhoven, S.; Oonincx, D.G.A.B.; van Huis, A.; van Loon, J.J.A. Growth performance and feed conversion efficiency of three edible mealworm species (Coleoptera: Tenebrionidae) on diets composed of organic by-products. J. Insect Physiol. 2015, 73, 1–10. [Google Scholar] [CrossRef]
- Hong, H.; Regenstein, J.M.; Luo, Y. The importance of ATP-related compounds for the freshness and flavor of post-mortem fish and shellfish muscle: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 1787–1798. [Google Scholar]
- Li, Y.; Li, H.; Zhang, G.; Liu, J.; Drolma, D.; Ye, B.; Yang, M. Boosted meat flavor by the metabolomic effects of Nile tilapia dietary inclusion of Zophobas atratus larval meal. Front. Biosci. (Landmark Ed) 2024, 29, 382. [Google Scholar] [CrossRef]
- Wang, C.; Sun, D.; Hu, X.; Chen, W.; Zou, C.; Zou, J. Effects of Spirulina platensis as a substitute for fishmeal on the liver of zig-zag eel (Mastacembelus armatus) infected with Aeromonas hydrophila. Comp Biochem Physiol Part D Genomics Proteomics. 2025, 56, 101563. [Google Scholar] [CrossRef]
- Liu, S.; Zheng, Z.; Ji, S.; Liu, T.; Hou, Y.; Li, S.; Li, G. Resveratrol reduces senescence-associated secretory phenotype by SIRT1/NF-κB pathway in gut of the annual fish Nothobranchius guentheri. Fish Shellfish Immunol. 2018, 80, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Shi, Y.; Yang, X.; Gao, J.; Nie, Z.; Xu, G. Effects of resveratrol on lipid metabolism in liver of red tilapia Oreochromis niloticus. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2022, 261, 109408. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Li, X.; Qiao, M.; Sun, X.; Li, G. Resveratrol alleviates inflammation and ER stress through SIRT1/NRF2 to delay ovarian aging in a short-lived fish. J. Gerontol. A Biol. Sci. Med. Sci. 2023, 78, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Shen, Y.; Zhao, W.; Yang, B.; Zhao, X.; Tao, S.; Sun, P.; Monroig, Ó.; Zhou, Q.; Jin, M. Evaluation of the optimum dietary arachidonic acid level and its essentiality for black seabream (Acanthopagrus schlegelii): Based on growth and lipid metabolism. Aquac. Nutr. 2024, 2024, 5589032. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, L.; Rahman, A.; Govindharajan, S.; Li, L.; Yu, H.; Waqas, M. Effects of varying dietary concentrations of menadione nicotinamide bisulphite (VK3) on growth performance, muscle composition, liver and muscle menaquinone-4 concentration, and antioxidant capacities of coho salmon (Oncorhynchus kisutch) alevins. Biology (Basel) 2025, 14, 447. [Google Scholar] [CrossRef]
- Wu, S.; Huang, J.; Li, Y.; Liu, Z.; Zhao, L. Integrated analysis of lncRNA and circRNA mediated ceRNA regulatory networks in skin reveals innate immunity differences between wild-type and yellow mutant rainbow trout (Oncorhynchus mykiss). Front. Immunol. 2022, 13, 802731. [Google Scholar] [CrossRef]
- Li, X.Q.; Lei, J.; Mao, L.H.; Wang, Q.L.; Xu, F.; Ran, T.; et al. NAMPT and NAPRT, key enzymes in NAD salvage synthesis pathway, are of negative prognostic value in colorectal cancer. Front. Oncol. 2019, 9, 736. [Google Scholar] [CrossRef]
- Rajman, L.; Chwalek, K.; Sinclair, D.A. Therapeutic potential of NAD-boosting molecules: The in vivo evidence. Cell. Metab. 2018, 27, 529–547. [Google Scholar] [CrossRef]
- Jang, Y.S. Skin transcriptome profiling of the blass bloched rockfish (Sebastes pachycephalus) with different body color patterns. Korean J. Ichthyol. 2020, 32, 117–129. [Google Scholar] [CrossRef]
- Protas, M.E. and N.H. Patel. 2008. Evolution of coloration patterns. Annu. Rev. Cell Dev. Biol. 2008, 24, 425–446. [Google Scholar] [CrossRef]
- Hubbard, J.K.; Uy, J.A.; Hauber, M.E.; Hoekstra, H.E.; Safran, R.J. Vertebrate pigmentation: from underlying genes to adaptive function. Trends Genet. 2010, 26, 231–239. [Google Scholar] [CrossRef]
- Lamason, R.L.; Mohideen, M.A.P.; Mest, J.R.; Wong, A.C.; Norton, H.L.; Aros, M.C.; Jurynec, M.J.; Mao, X.; Humphreville, V.R.; Humbert, J.E.; et al. SLC24A5, a putative cation exchanger, affects pigmentation in zebrafish and humans. Science, 2005, 310, 1782–1786. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, J.; Shi, M.; Ma, X.; Chen, S.; Zhou, Q.; Zhu, C. Metabolomic analysis revealed the inflammatory and oxidative stress regulation in response to Vibrio infection in Plectropomus leopardus. J. Fish Biol. 2024, 105, 1694–1702. [Google Scholar] [CrossRef] [PubMed]
- Jeddi, I.; Saiz, L. Computational design of single-stranded DNA hairpin aptamers immobilized on a biosensor substrate. Sci. Rep. 2021, 11, 10984. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Liu, M.; Wei, S.; Qin, X.; Qin, Q.; Li, P. Research progress and prospects for the use of aptamers in aquaculture biosecurity. Aquaculture 2021, 534, 736257. [Google Scholar] [CrossRef]
- Tan, Y.; Lin, X.; Huang, L.; Yan, Q.; Wang, J.; Weng, Q.; Zhengzhang, Y.; Chen, Y.; Ma, Y.; Zheng, J. Transcriptomic analysis of the inhibition mechanisms against Pseudomonas plecoglossicida by antibacterial aptamer B4. Front Vet Sci. 2024, 11, 1511234. [Google Scholar] [CrossRef]
- Jiang, Z.; Weng, P.; Xu, X.; Li, M.; Li, Y.; Lv, Y.; Chang, K.; Wang, S.; Lin, G.; Hu, C. IRF9 promotes apoptosis and innate immunity by inhibiting SIRT1-p53 axis in fish. Fish Shellfish Immunol. 2020, 103, 220–228. [Google Scholar] [CrossRef]
- LaMoia, T.E.; Shulman, G.I. Cellular and molecular mechanisms of metformin action. Endocr. Rev. 2021, 42, 77–96. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Hou, Y.; Liu, K.; Zhu, H.; Qiao, M.; Sun, X.; Li, G. Metformin protects against inflammation, oxidative stress to delay Poly I:C-Induced aging-like phenomena in the gut of an annual fish. J Gerontol A. Biol Sci Med Sci. 2022, 77, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.Y.; Zhang, Z.W.; Chen, S.N.; Pang, A.N.; Peng, X.Y.; Li, N.; Liu, L.H.; Nie, P. SIRT6 positively regulates antiviral response in a bony fish, the Chinese perch Siniperca chuatsi. Fish Shellfish Immunol. 2024, 150, 109662. [Google Scholar] [CrossRef] [PubMed]
- Mo, Z.Q.; Wu, H.C.; Hu, Y.T.; Lu, Z.J.; Lai, X.L.; Chen, H.P.; He, Z.C.; Luo, X.C.; Li, Y.W.; Dan, X.M. Transcriptomic analysis reveals innate immune mechanisms of an underlying parasite-resistant grouper hybrid (Epinephelus fuscogutatus × Epinephelus lanceolatus). Fish Shellfish Immunol 2021, 119, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Lahnsteiner, F.; Weismann, T.; Patzner, R.A. Physiological and biochemical parameters for egg quality determination in lake trout, Salmo trutta lacustris. Fish Physiol. Biochem. 1999, 20, 375–388. [Google Scholar] [CrossRef]
- Lahnsteiner, F.; Urbanyi, B.; Horvath, A.; Weismann, T. Bio-markers for egg quality determination in cyprinid fish. Aquaculture 2001, 195, 331–352. [Google Scholar] [CrossRef]
- Jensen, F.B. Influence of nucleoside triphosphates, inorganic salts, NADH, catecholamines, and oxygen saturation on nitrite-induced oxidation of rainbow trout haemoglobin. Fish Physiol. Biochem. 1993, 12, 111–117. [Google Scholar] [CrossRef]
- Jensen, F.B.; Nikinmaa, M.; Weber, R.E. Environmental perturbations of oxygen transport in teleost fishes: Causes, consequences, and compensations. J. Exp. Biol. 1993, 179, 153–166. [Google Scholar]
- Cashon, R.E.; Vayda, M.E.; Sidell, B.D. Kinetic characterization of myoglobins from vertebrates with vastly different body temperatures. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol. 1997, 118, 613–619. [Google Scholar] [CrossRef]
- Brown, W.D.; Snyder, H.E. Nonenzymatic reduction and oxidation of myoglobin and hemoglobin by nicotinamide adenine dinucleotides and flavins. J. Biol. Chem. 1969, 244, 6702–6706. [Google Scholar] [CrossRef]
- Chaijan, M.; Benjakul, S.; Visessanguan, W.; Faustman, C. Characteristics and gel properties of muscles from sardine (Sardinella gibbosa) and mackerel (Rastrelliger kanagurta) caught in Thailand. Food Chem. 2006, 97, 231–239. [Google Scholar] [CrossRef]
- Richards, M.P.; Hultin, H.O. Contributions of blood and blood components to lipid oxidation in fish muscle. J. Agric. Food Chem. 2002, 50, 7413–7419. [Google Scholar] [CrossRef]
- Baron, C.P.; Kjærsgård, I.V.H.; Jessen, F.; Jacobsen, C. Protein and lipid oxidation during frozen storage of rainbow trout (Oncorhynchus mykiss). J. Agric. Food Chem. 2007, 55, 8118–8125. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, M. J.; Børresen, T. Effect of freezing on biochemical properties of Atlantic mackerel (Scomber scombrus). J Aquat Food Prod Technol 1995, 4, 5–24. [Google Scholar]
- Shumilina, E.; Ciampa, A.; Capozzi, F.; Rustad, T.; Dikiy, A. NMR approach for monitoring post-mortem changes in Atlantic salmon fillets stored at 0 and 4°C. Food Chemistry, 2015, 184, 12–22. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation illustrating the central role of NAD⁺ metabolism as a key component of a network that links metabolism, neuronal function, and environmental responsiveness in teleosts. NAD⁺ acts as a critical hub connecting 1. Dietary precursor availability. 2. Regulation of synaptic plasticity and memory through activation of sirtuins (SIRT1) and transcription factors such as CREB. 3. Nitric oxide (NO) synthesis, a key signaling molecule, via the NADPH-diaphorase (NOS) enzyme, relies on NAD⁺-derived cofactor (NADPH). NO mediates processes such as neurotransmission and vasoregulation. 4. Protective mechanisms against external stressors such as acoustic stress, maintain redox homeostasis, and promote cellular repair. 5. Integration of nutritional and environmental signals to regulate metabolic efficiency and organismal growth.
Figure 1.
Schematic representation illustrating the central role of NAD⁺ metabolism as a key component of a network that links metabolism, neuronal function, and environmental responsiveness in teleosts. NAD⁺ acts as a critical hub connecting 1. Dietary precursor availability. 2. Regulation of synaptic plasticity and memory through activation of sirtuins (SIRT1) and transcription factors such as CREB. 3. Nitric oxide (NO) synthesis, a key signaling molecule, via the NADPH-diaphorase (NOS) enzyme, relies on NAD⁺-derived cofactor (NADPH). NO mediates processes such as neurotransmission and vasoregulation. 4. Protective mechanisms against external stressors such as acoustic stress, maintain redox homeostasis, and promote cellular repair. 5. Integration of nutritional and environmental signals to regulate metabolic efficiency and organismal growth.
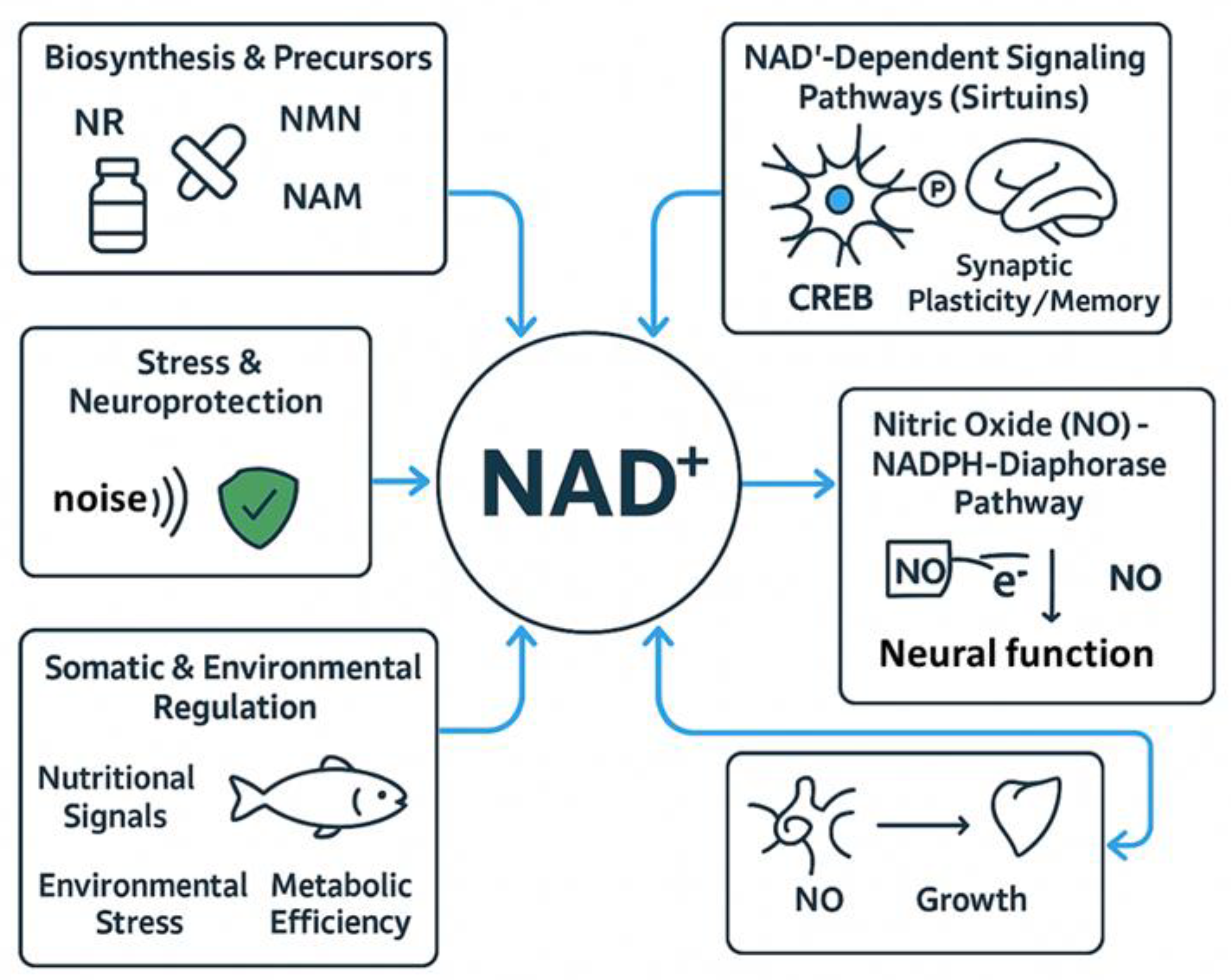
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



