Submitted:
18 December 2024
Posted:
19 December 2024
You are already at the latest version
Abstract
The longevity gene-associated protein sirtuin, which has NAD-dependent deacetylation enzyme activity, plays an important role in genomic stability, antioxidant potential, DNA repair, mitochondrial and energy metabolism all of which are associated with a healthy state. NAD+ and sirtuin levels can be increased by supplementing with NAD precursors including nicotinamide mononucleotide (NMN) and some polyphenols. If meals contain NAD precursors and NAD synthesis activators, a daily diet, as well as NMN supplements, may support NAD+ augmentation and the resultant sirtuin activation. This can then lead to a resulting improvement in mitochondrial metabolism and health. Because tuna meat contains NAD precursors, we postulate that a daily diet of fish has the potential to increase the levels of sirtuins which are NAD-dependent regulators of mitochondria in human cells.
In the present study, we investigated whether tuna meat regulates mitochondrial function and energy metabolism in human peripheral blood mononuclear cells (PBMCs). A digestive enzyme tuna meat hydrolysate was prepared from tuna dark red meat. Mitochondrial activity (NAD+/NADH level), sirtuin expression, energy production (ATP level) and antioxidant activity in PBMCs were analyzed after the treatment of cells with the digestive enzyme tuna hydrolysate or controls in vitro. Our results demonstrate that the digestive enzyme tuna hydrolysate has the potential to increase the expression of longevity gene-related proteins sirtuin 1 and 2, NAD+/NADH levels, and ATP production by mitochondria in PBMCs. Furthermore, tuna meat showed high antioxidant capacity in vitro. These findings indicate the favorable effect of tuna meat on mitochondrial function and homeostasis. Our study underscores the benefits of tuna meat as a dietary choice and provides scientific support for promotion of healthy aging.
Keywords:
tuna meat
; digestive enzyme hydrolysates
; longevity gene-related protein
; sirtuin
; mitochondria
; antioxidant
; nicotinamide adenine dinucleotide (NAD)
1. Introduction
Population aging is now a global phenomenon especially in developing countries. An increase in life expectancy and a decline in fertility rates have resulted in accelerated aging of the population in many countries [1,2]. In particular, in Asian countries, a quarter of the population is predicted to be ≥60 years old by 2050, which will inevitably lead to an increased number of older adults with chronic diseases and disability, and have profound consequences for health, health systems, the workforce, and budgeting for many countries [1,2,3].
The determinants of longevity are not fully understood. It is commonly believed that lifestyle factors, such as physical activity, smoking, and food habits, play an important role in human health and longevity [4]. Of these, nutrition and diet have been established as being among the most important influences [2,5,6,7]. It is now thought that altering the levels, type, and timing of food consumption is the most potent, feasible and safest intervention for improvement of health and extension of longevity and the time in which health and functional capacity are maintained (i.e., health span) [2,8,9,10,11]. However, despite extensive research, the type, quantity, and combination of nutrients that optimize healthy longevity remains highly controversial. Understanding mechanisms of dietary interventions will be key for optimizing human health span and longevity.
Diet is one of the major determinates of health [2,12]. The beneficial effects of fruit and vegetable intake are well known including a reduction in the risk of mortality [13,14,15]. In addition, fish meats are commonly recommended for health in the general population [16,17]. Numerous studies have already demonstrated that omega-3 fats, docosahexaenoic acid (DHA) and eicosatetraenoic acid (EPA), in fish meat can help reduce the risk of chronic diseases, such as heart and inflammatory disease, and help maintain brain function and eye health [17]. Like the omega-3 fats, EPA and DHA are plentiful in oily fish, such as salmon and anchovies [18]. Since our body can only make a small amount of omega-3 fat DHA from other fatty acids, we need to replenish it directly from food or supplements. Although fish meats are recommended for health and longevity in the general population, the association between fish meat and health outcomes still remains unclear.
Recently, attention has been attracted by the selenium-containing compound, selenoneine, found in tuna meat which may have the potential to improve the health condition and extend longevity in the general population [19]. Yamashita Y et. al. identified a novel selenium-containing imidazole compound, selenoneine, in tuna meats [19,20]. They clarified that selenoneine is an ergothioneine analog with greater antioxidant activity and is the major form of organic selenium in the blood, muscles, and other tissues of tuna [21]. Also, recent studies demonstrated that the selenoneine-containing tuna meats, especially dark muscle meat, may prevent carcinogenesis and liver disorder [21,22]. Since selenoneine shows a stronger antioxidative activity than other antioxidants such as vitamin E (500-fold) and ergothioneine (1000-fold), an intake of tuna meats could improve the cellular and organism response to oxidative stress in the body [21].
Our recent research has focused on mitochondrial metabolism and its key regulator sirtuin, a longevity gene-related deacetylase, that is closely involved in mitochondrial function, cellular energy metabolism, health condition and longevity. The sirtuin protein, which has nicotinamide adenine dinucleotide (NAD)-dependent deacetylation enzyme activity, plays an important role in a broad range of mammalian processes such as genomic stability, cellular antioxidant potential, DNA repair and cellular energy metabolism [23,24]. Sirtuins perform deacetylation on target proteins and genes and play a central role in various cellular responses to extrinsic stresses [25]. In addition, sirtuins have important roles in the control of mitochondrial function and energy metabolism as well as cellular stress tolerance. It is now thought that aging symptoms of various tissues and organs may be suppressed by sirtuin activation [26,27].
Numerous studies have demonstrated that boosting NAD+ levels increase sirtuin activity and the resultant mitophagy (reverse mitochondrial dysfunction), by which damaged mitochondria are targeted and degraded, suggesting that activation of NAD+, as well as sirtuin activation, is essential for mitochondrial balance and healthy aging [27,28,29]. NAD+ and sirtuin levels can be increased by supplementing with NAD precursors including nicotinamide mononucleotide (NMN) and some polyphenols [30,31,32]. Since downregulation of mitophagy has also been suggested to be implicated in the impediment of health status and healthy aging, enhancing mitophagy with NAD precursors, such as NMN, could be an effective strategy to boost health and longevity [27,29]. To date, many studies have evaluated how NAD+ precursors support healthy aging in people [31,32,33,34]. If the daily diet contains NAD precursors and NAD synthesis activators, meals as well as NMN supplements may support NAD+ augmentation and the resultant sirtuin activation, consequently resulting in improvement in mitochondrial metabolism and health condition.
We aim to explore the connections between dietary choice and its impact on longevity-related cellular mechanisms. Currently, our attention is directed towards exploring potential effects of tuna meat, which is rich in nutrients. In the present study, to verify how tuna meat consumption impacts longevity-related cellular mechanisms, we studied the effects of digestive enzyme tuna hydrolysates on the level of longevity gene-related sirtuin proteins, mitochondrial activity and antioxidant capacity in peripheral blood mononuclear cells (PBMCs) in vitro. The results of this study could provide important evidence to help better dietary choice aimed at promoting healthy ageing.
2. Materials and Methods
2.1. Fish Hydrolysate Preparation
Dark red meat from yellow fin tuna (Misaki Megumi Suisan Co., Ltd., Kanagawa, Japan) was used for hydrolysate preparation. The raw material was minced, vacuum packaged, and stored at − 20 °C until use. Digestive enzyme hydrolysates were prepared following the cited methods with slight modifications [35,36,37].
Briefly, minced tuna meat was mixed with a 2-fold volume of diluted water and homogenized in a blender. Homogenates were then incubated at 70 °C for 30 minutes. The homogenized tuna meat was adjusted to pH 1.8 using HCl. The digestive enzyme pepsin (Fujifilm Wako Pure Chemical Ltd., Osaka, Japan) was added at 1 g/kg (1/1000 w/w) of the total volume of the raw material. All reactions were performed in a shaking incubator at 37 °C for 2 hours. After adjusting to pH 6.8 using NaOH, the reactions were terminated by heating the solution to 90–100 °C for 10 minutes to deactivate enzyme. The tuna hydrolysates were treated with the digestive enzymes trypsin and pancreatin (Fujifilm Wako Pure Chemical Ltd., Osaka, Japan) for 2 hours using the same method as described above. After terminating by heating the solution to 90–100 °C for 10 minutes to deactivate enzyme, the samples were cooled on ice to room temperature, and then filtered through a 0.45 μm membrane filter. They were used as tuna meat digestive enzyme hydrolysate in the following experiment.
2.2. Treatment of PBMCs with the Digestive Enzyme Hydrolysate of Tuna Meat
Human peripheral blood mononuclear cells (PBMCs; Fujifilm Wako Pure Chemical Ltd., Osaka, Japan) were incubated in the presence or absence of digestive enzyme tuna hydrolysate for 24 hours at 37 °C in a humidified atmosphere of 95% air and 5% CO2.
As a positive control group, PBMCs were also incubated with 10 μM NMN, an activator of sirtuins and mitochondria [31,32]. As another positive control group, a hot water extract of tuna meat was prepared by the method described previously [36,37]. PBMCs were treated with the hot water tuna extract for 24 hours at 37 °C in a humidified atmosphere of 95% air and 5% CO2.
2.3. Viability of PBMCs
Effects of digestive enzyme tuna hydrolysate on cell viability, proliferation and cytotoxicity in PBMCs were analyzed using a Cell Counting Kit (CCK)-8 assay kit (Dojindo Molecular Technologies Inc., Kumamoto, Japan), after treatment of PBMCs with tuna hydrolysates or controls.
2.4. Effect of Digestive Enzyme Tuna Hydrolysate on the Level of Nicotinamide Adenine Dinucleotide (NAD+) in PBMCs
To verify whether digestive enzyme tuna hydrolysate influences the level of NAD+ in PBMCs, concentrations of NAD/NADH in the PBMCs were analyzed by a colorimetric assay (NAD/NADH colorimetric assay kit, Dojindo Molecular Technologies Inc., Kumamoto, Japan), after treatment of cells with tuna hydrolysates or controls.
2.5. Western Blot Assay for Sirtuin 1 and Sirtuin 2 in PBMCs
To study the effect of digestive enzyme tuna hydrolysate on the expression of sirtuin 1 and sirtuin 2 in PBMCs, the levels of these two proteins in cells were analyzed by western blotting as previously described [38,39], after treatment of cells with tuna hydrolysates or controls. For western blotting, the antibodies used were a polyclonal antibody against human sirtuin 1 (1:5,000, dilution; Abcam Inc., Cambridge, UK), sirtuin 2 (1:2,000, dilution; Abcam Inc.), beta-actin (1:10,000 dilution; Proteintech Group Inc., IL) and the corresponding secondary antibody conjugated with horseradish peroxidase [Dako, rabbit IgG P0448 for both primary antibodies (1:5,000 dilution). The antibody-bound protein bands were visualized, and densitometry of the signal bands was analyzed using an extended cavity laser system (GE Healthcare Bio-sciences KK, Tokyo, Japan).
2.6. ATP Production by PBMCs
To study the effect of digestive enzyme tuna hydrolysate on the production of ATP by PBMCs, the level of ATP production of cells was analyzed using an ATP assay for blood (Toyo be-net Co., Ltd., Tokyo, Japan), after treatment of cells with tuna hydrolysates or controls.
2.7. Antioxidant Activity of Digestive Enzyme Tuna Hydrolysate
The antioxidative activity of digestive enzyme tuna hydrolysate was analyzed using an antioxidant assay kit, [2,2-Diphenyl-1-picrylhydrazyl (DPPH) Antioxidant Assay Kit, Dojindo Molecular Technologies].
2.8. Concentrations of NAD+ Precursors, Nicotinamide Mononucleotide (NMN) and Nicotineamide (NAM), in Tuna Red and Dark Meats
To study whether tuna meat contains NAD+ precursors, NMN and NAM, concentrations of NMN and NAM in tuna meat was analyzed by liquid chromatography-mass spectrometry. Analysis was performed by the Japan Foods Inspection Corporation, Tokyo, Japan.
2.9. Statistical Analysis
The results are presented as the mean ± standard deviations. All data are representative of at least three individual experiments and analyzed with the Mann–Whitney U-test. Differences were considered significant at p < 0.05.
3. Results
3.1. Cell Viability of PBMCs
The effect of digestive enzyme tuna hydrolysate on cell viability in PBMCs was analyzed by CCK8 assay. Results demonstrated that the treatment of PBMCs with tuna hydrolysate resulted in the downregulation of cell viability at protein concentrations greater than 9.4 μg/mL (Figure 1). Subsequently, in the present study, PBMCs were treated with digestive enzyme tuna hydrolysate at a concentration of 4.0 μg/mL in all experiments.
3.2. Effect of Digestive Enzyme Tuna Hydrolysate on the Level of Nicotinamide Adenine Dinucleotide (NAD+) in PBMCs
As shown in Figure 2, both digestive enzyme hydrolysate and the hot water extract (4 μM) and NMN (10 μM) increased the levels of NAD+/NADH activity in PBMCs (% ratio versus negative control, digestive enzyme hydrolysate: 108%, hot water extract: 112%, NMN: 111%) (Figure 2A). Both tuna extracts and NMN also induced the oxidized form of NAD (Figure 2B).
3.3. Effects of Digestive Enzyme Tuna Hydrolysate on the Expression of Sirtuin 1 and Sirtuin 2 in PBMCs
As shown in Figure 3A, B, treatment with digestive enzyme tuna hydrolysate (4.0 μg/mL) upregulated the expression of sirtuin 1 in PBMCs, as well as in two different positive controls, namely NMN (10 μM) treatment and addition of a hot water extract of tuna meat (4.0 μg/mL) (ratio versus negative control, digestive enzymatic hydrolysate: 110 %, hot water extract: 120%, NMN: 116 %).
Both tuna extracts (4.0 μg/mL), the digestive enzyme hydrolysate and the hot water extract, induced the expression of sirtuin 2 in PBMCs, and NMN (10 μM) also increased the expression of sirtuin 2 in PBMCs (ratio versus negative control, digestive enzymatic hydrolysate: 114 %, hot water extract: 129%, NMN: 111 %) ((Figure 3C).
3.4. Effect of Digestive Enzyme Tuna Hydrolysate on ATP Production by PBMCs
Both tuna extracts (4μM), the digestive enzyme hydrolysate and the hot water extract, and NMN (10 μM) upregulated the production of ATP by PBMCs (% ratio versus negative control, digestive enzyme hydrolysate: 108%, hot water extract: 110%, NMN: 115%), although no significant differences were observed between them (Figure 4).
3.5. Antioxidative Activity of Digestive Enzyme Tuna Hydrolysate
The antioxidant activity of the sample was evaluated as Torolox equivalent activity (TEAC) which was based on antioxidant standard material Trolox. The TEAC value of digestive enzyme hydrolysate and the hot water extract were 0.313 and 0.297 TE μg/mg, respectively.
3.6. Concentrations of NAD+ Precursors, NMN and NAM, in Tuna Red And Dark Meats
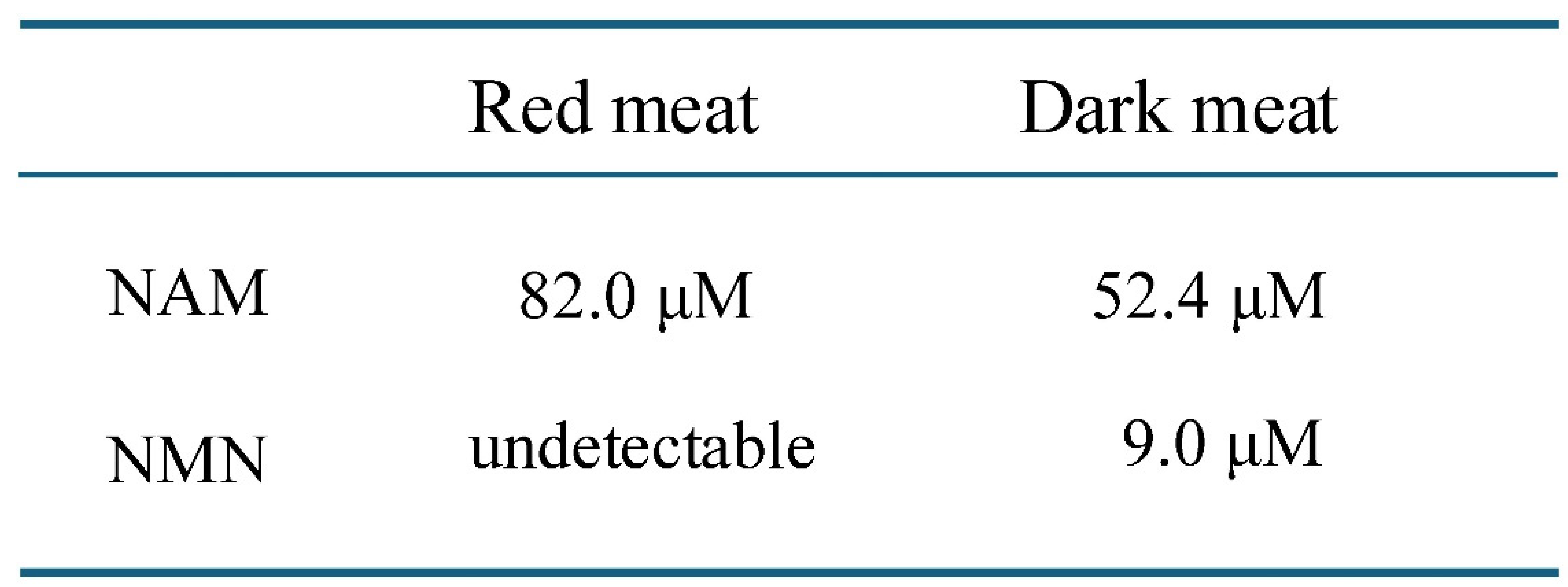
Both red and dark tuna meat contain 5 ~8 μM of NAM (red meat: 82.0 μM, dark meat: 52.4 μM), although no detectable levels of NMN were observed in both tuna meats (Table 1).
4. Discussion
Our study elucidates the effects of digestive enzyme tuna hydrolysate on the expression of the longevity gene-related sirtuin protein, on mitochondrial activity, and on the antioxidant capacity of PBMCs. The results reveal for the first time important evidence to clarify the connections between a specific dietary choice, namely “tuna meat”, and its impact on longevity-related cellular mechanisms and oxidative stress tolerance. We demonstrate in vitro that digestive enzyme tuna hydrolysate has the potential to increase the expression of longevity gene-related sirtuin 1and 2, the NAD+/NADH level and ATP production by mitochondria in PBMCs. These findings suggest that tuna meat is a healthy dietary choice which may function in health status and longevity via the upregulation of mitochondrial activity, since both activation of sirtuin and NAD+ are essential for mitochondrial balance and function [26,27,28,29].
Downregulation of mitophagy (reverse mitochondrial dysfunction) has been suggested to be implicated in health status and healthy aging [40,41]. Numerous studies have demonstrated that boosting NAD+ levels can increase sirtuin activity and the resultant mitophagy (by which damaged mitochondria are targeted and degraded)., This indicates that activation of sirtuin, as well as NAD+ activation, is essential for mitochondrial balance and healthy aging [40,41,42,43,44]. It has also been demonstrated that NAD+ and sirtuin levels can be increased by supplementing with NAD precursors including NMN and some polyphenols such as resveratrol [30,31,32]. Enhancing mitophagy with NAD precursors may be an effective strategy to boost health and longevity. To date, many studies have evaluated how NAD+ precursors support healthy aging in people [31]. If tuna meats contain NAD precursors and NAD synthesis activators, tuna as part of a daily diet, as well as NMN supplements, may support NAD+ augmentation and the resulting sirtuin activation. Consequently, this could result in improvement of mitochondrial metabolism and increased health status. Indeed, our present study indicates that both red and dark tuna meats have NAM levels of 50 ~80 μM (red meat: 82 μM, dark meat: 52 μM) which as a NAD+ precursor suggests that NAM in tuna meat has the potential to upregulate the level of NAD+ and induce the activation of sirtuins in mitochondria.
In mammals, it is well known that the NAD precursor, NMN, is synthesized in vivo by NAM phosphoribosyl transferase (NAMPT), using NAM as the main reaction substrate in the salvage NAD+ biosynthesis pathway [45,46]. As shown in Figure 5A, NAM, an intermediate metabolite of NAD+, is thought to be metabolized to NMN, which enters the cell, is converted to NAD+, and activates sirtuins [47,48]. NAD+ is recycled from both NAM and NMN in the salvage pathway to maintain the cellular NAD+ levels. Our findings of micromolar levels of NAM in the digestive enzyme tuna hydrolysate provide evidence to support the efficacy of tuna meats in cellular energy production via mitochondria activation (Figure 5B).
Our recent research has focused on mitochondrial metabolism and its key regulator sirtuin, a longevity gene-related deacetylase, that is closely involved in the mitochondrial function, cellular energy metabolism, health condition and longevity. Sirtuin, which has NAD-dependent deacetylation enzyme activity, plays an important role in a broad range of mammalian processes including genomic stability, cellular antioxidant potential, DNA repair and cellular energy metabolism [23,24,25]. In addition, sirtuin protein plays an important part in the control of mitochondrial function and energy metabolism through the NAD-dependent mechanism of its activation [27]. Its activation depends on NAD metabolism in mitochondria. Once activated, sirtuin performs deacetylation of target proteins and genes, and plays a central role in various cellular processes. In mammalian cells, the sirtuin family consists of seven members (sirtuin 1 to 7) as shown in Table 1. Members of the sirtuin family have different target proteins in different cellular compartments [24]. The sirtuin family, proteins related to the longevity gene Sir2, activate or suppress approximately 100 aging-related factors and genes [23,24]. It is now thought that the aging symptoms of various tissues and organs may be suppressed by sirtuin activation [23]. Consequently, sirtuin levels are considered to affect the health condition and life expectancy. Our data demonstrate that the digestive enzyme tuna hydrolysate has the potential to increase the expression of sirtuin 1 and sirtuin 2 in PBMCs. As mentioned above, we found that tuna meats contain the NAS+ precursor NAM which is the main reaction substrate in the NAD+ biosynthesis salvage pathway. The NAM in tuna meats may be converted to NAD+ and activate sirtuin in PBMCs. Our findings highlight important evidence to help better dietary choices, such as tuna meat, aimed at promoting healthy ageing.
Selenoneine is a selenium compound that was initially identified in the blood of bluefin tuna and has subsequently been shown to be present in various animals of marine origin, including multiple species of fish, dolphins, sea turtles, and seabirds [19,20,21,49]. Selenoneine has also been identified in red blood cells of human populations whose diet comprises large amounts of marine foods, including in Japan [50]. Selenoneine has strong antioxidant activity and may play a role in the protection and function of organs/tissues [20,51,52]. We have postulated that tuna meats may show beneficial effects on mitochondrial function and cellular energy metabolism (ATP production). It is well known that mitochondria are the location of the anabolic pathway which produces cellular energy [53]. Also, as a location of catabolic pathways, the mitochondria produces reactive oxygen species (ROS) [54,55]. By intake of tuna meats, selenoneine, as a free radical scavenger, may influence mitochondrial functions in terms of both anabolic ATP production and catabolic ROS production in the human body. Indeed, our data indicate that digestive enzyme tuna hydrolysate showed high levels of antioxidative potential. The digestive enzyme tuna hydrolysate as an antioxidant may therefore have the potential to protect PBMC against oxidative stress and to maintain mitochondria function. These findings suggest beneficial effects of tuna meat on cellular molecules involved in energy metabolism and oxidative stress tolerance.
5. Conclusions
In conclusion, our findings reveal that digestive enzyme tuna hydrolysate has the potential to accelerate the expression of longevity gene-related sirtuin 1 and 2, the NAD+/NADH level and ATP production by mitochondria in PBMCs. Also, tuna meat shows high antioxidant capacity in vitro, indicating a possible favorable effect of tuna meat on mitochondrial function and homeostasis. These observations underscore the usefulness of tuna meat as a dietary choice and provide scientific support for the promotion of healthy ageing.
Author Contributions
Formal analysis, visualization, writing—original draft preparation, writing—review and editing, Y.S. and K.Y.; methodology, Y.S-T.; data curation, K.Y.; investigation, Y.S., Y.S-T., K.Y.; resources, Y.S.; validation, Y.S.; conceptualization, K.Y.; funding acquisition and supervision, K.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by JSPS KAKENHI grant number 17K11035 from the Japan Society for the Promotion of Science (JSPS).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding author.
Acknowledgments
We would like to thank Junko Tamate, Tomoko Sato, Megumi Tanaka of the Institute of Medical Science, St. Marianna University School of Medicine, for excellent technical assistance.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- ESCAP. Ageing in Asia and the Pacific: Overview; ESCAP: Bangkok, Thailand, 2017.
- Yan-Feng Zhou, Xing-Yue Song, An Pan, Woon-Puay Koh. Nutrition and Healthy Ageing in Asia: A Systematic Review. Nutrients, 15, 3153. 2023. [CrossRef]
- Bautmans I, Knoop V, Amuthavalli Thiyagarajan J, Maier AB, Beard JR, Freiberger E, Belsky D, Aubertin-Leheudre M, Mikton C, Cesari M, Sumi Y, Diaz T, Banerjee A; WHO Working Group on Vitality Capacity. WHO working definition of vitality capacity for healthy longevity monitoring. Lancet Healthy Longev. 3(11):e789-e796, 2022. [CrossRef]
- Wang J, Chen C, Zhou J, Ye L, Li Y, Xu L, Xu Z, Li X, Wei Y, Liu J, Lv Y, Shi X. Healthy lifestyle in late-life, longevity genes, and life expectancy among older adults: a 20-year, population-based, prospective cohort study. Lancet Healthy Longev. 4(10):e535-e543, 2023. [CrossRef]
- Molendijk M., Molero P., Ortuno Sanchez-Pedreno, F., Van der Does, W.Angel Martinez-Gonzalez M. Diet quality and depression risk: A systematic review and dose-response meta-analysis of prospective studies. J. Affect. Disord. 226, 346–354, 2018.
- Chen L.K., Arai H., Assantachai P., Akishita M., Chew S.T.H., Dumlao L.C., Duque G., Woo J. Roles of nutrition in muscle health of community-dwelling older adults: Evidence-based expert consensus from Asian Working Group for Sarcopenia. J. Cachexia Sarcopenia Muscle 13, 1653–1672, 2022.
- Zhang J., Zhao A. Dietary Diversity and Healthy Aging: A Prospective Study. Nutrients, 13, 1787, 2021. [CrossRef]
- Zhou, Y.F.; Song, X.Y.; Wu, J.; Chen, G.C.; Neelakantan, N.; van Dam, R.M.; Feng, L.; Yuan, J.M.; Pan, A.; Koh, W.P. Association Between Dietary Patterns in Midlife and Healthy Ageing in Chinese Adults: The Singapore Chinese Health Study. J. Am. Med. Dir. Assoc., 22, 1279–1286, 2021.
- Zhou, Y.F.; Lai, J.S.; Chong, M.F.; Tong, E.H.; Neelakantan, N.; Pan, A.; Koh, W.P. Association between changes in diet quality from mid-life to late-life and healthy ageing: The Singapore Chinese Health Study. Age Ageing, 51, afac232, 2021. [CrossRef]
- Aihemaitijiang, S.; Zhang, L.; Ye, C.; Halimulati, M.; Huang, X.; Wang, R.; Zhang, Z. Long-Term High Dietary Diversity Maintains Good Physical Function in Chinese Elderly: A Cohort Study Based on CLHLS from 2011 to 2018. Nutrients, 14, 1730, 2022. [CrossRef]
- Hata, T.; Seino, S.; Yokoyama, Y.; Narita, M.; Nishi, M.; Hida, A.; Shinkai, S.; Kitamura, A.; Fujiwara, Y. Interaction of Eating Status and Dietary Variety on Incident Functional Disability among Older Japanese Adults. J. Nutr. Health Aging, 26, 698–705, 2022.
- Zhang, J.; Zhao, A.; Wu, W.; Ren, Z.X.; Yang, C.L.; Wang, P.Y.; Zhang, Y.M. Beneficial Effect of Dietary Diversity on the Risk of Disability in Activities of Daily Living in Adults: A Prospective Cohort Study. Nutrients, 12, 3263, 2022. [CrossRef]
- Lee, A.T.C.; Richards, M.; Chan, W.C.; Chiu, H.F.K.; Lee, R.S.Y.; Lam, L.C.W. Lower risk of incident dementia among Chinese older adults having three servings of vegetables and two servings of fruits a day. Age Ageing, 46, 773–779, 2017.
- Yeung, S.S.Y.; Kwok, T.; Woo, J. Higher fruit and vegetable variety associated with lower risk of cognitive impairment in Chinese community-dwelling older men: A 4-year cohort study. Eur. J. Nutr., 61, 1791–1799, 2022.
- Wang, R.S.; Wang, B.L.; Huang, Y.N.; Wan, T.T.H. The combined effect of physical activity and fruit and vegetable intake on decreasing cognitive decline in older Taiwanese adults. Sci. Rep., 12, 9825, 2022. [CrossRef]
- M. Fotuhi, P. Mohassel, K. Yaffe. Fish consumption, long-chain omega-3 fatty acids and risk of cognitive decline or Alzheimer disease: a complex association. Nat Clin Pract Neurol, 5 (3), 140-152, 2009. [CrossRef]
- Gao, Q.; Niti, M.; Feng, L.; Yap, K.B.; Ng, T.P. Omega-3 Polyunsaturated Fatty Acid Supplements and Cognitive Decline: Singapore Longitudinal Aging Studies. J. Nutr. Health Aging, 15, 32–35, 2011.
- Matsuoka, Y.J.; Sawada, N.; Mimura, M.; Shikimoto, R.; Nozaki, S.; Hamazaki, K.; Uchitomi, Y.; Tsugane, S. Dietary fish, n-3 polyunsaturated fatty acid consumption, and depression risk in Japan: A population-based prospective cohort study. Transl. Psychiatry, 7, e1242, 2017. [CrossRef]
- Yumiko Yamashita, Michiaki Yamashita. Identification of a novel selenium-containing compound, selenoneine, as the predominant chemical form of organic selenium in the blood of bluefin tuna. J Biol Chem, 285(24):18134-8, 2010. [CrossRef]
- Yumiko Yamashita, Takeshi Yabu, Michiaki Yamashita. Discovery of the strong antioxidant selenoneine in tuna and selenium redox metabolism. World J Biol Chem., 1(5):144-50, 2010. [CrossRef]
- Junko Masuda, Chiho Umemura, Miki Yokozawa, Ken Yamauchi, Takuya Seko, Michiaki Yamashita, Yumiko Yamashita. Dietary Supplementation of Selenoneine-Containing Tuna Dark Muscle Extract Effectively Reduces Pathology of Experimental Colorectal Cancers in Mice. Nutrients, 10(10):1380, 2018. [CrossRef]
- Masaaki Miyata, Koki Matsushita, Ryunosuke Shindo, Yutaro Shimokawa, Yoshimasa Sugiura, Michiaki Yamashita. Selenoneine Ameliorates Hepatocellular Injury and Hepatic Steatosis in a Mouse Model of NAFLD. Nutrients, 12(6):1898, 2020. [CrossRef]
- Wan X, Garg NJ. Sirtuin Control of Mitochondrial Dysfunction, Oxidative Stress, and Inflammation in Chagas Disease Models. Front Cell Infect Microbiol., 11:693051. eCollection 2021. [CrossRef]
- Wan W, Hua F, Fang P, Li C, Deng F, Chen S, Ying J, Wang X. Regulation of Mitophagy by Sirtuin Family Proteins: A Vital Role in Aging and Age-Related Diseases. Front Aging Neurosci. 9;14:845330. 845330. eCollection 2022. [CrossRef]
- Wu QJ, Zhang TN, Chen HH, Yu XF, Lv JL, Liu YY, Liu YS, Zheng G, Zhao JQ, Wei YF, Guo JY, Liu FH, Chang Q, Zhang YX, Liu CG, Zhao YH. The sirtuin family in health and disease. Signal Transduct Target Ther. 7(1):402, 2022. [CrossRef]
- Imai S, Guarente L. NAD+ and sirtuins in aging and disease. Trends Cell Biol. 24(8):464-71. Epub, 2014. [CrossRef]
- Ji Z, Liu GH, Qu J. Mitochondrial sirtuins, metabolism, and aging. J Genet Genomics. 49(4):287-298. Epub, 2022. [CrossRef]
- Zapata-Pérez R, Wanders RJA, van Karnebeek CDM, Houtkooper RH. NAD(+) homeostasis in human health and disease. EMBO Mol Med. 13(7):e13943. Epub, 2021. [CrossRef]
- Chini CCS, Cordeiro HS, Tran NLK, Chini EN. NAD metabolism: Role in senescence regulation and aging. Aging Cell. 23(1):e13920. Epub, 2023. [CrossRef]
- Yaku K, Nakagawa T. NAD(+) Precursors in Human Health and Disease: Current Status and Future Prospects. Antioxid Redox Signal. 39(16-18):1133-1149. Epub 2023. [CrossRef]
- Alegre GFS, Pastore GM. NAD+ Precursors Nicotinamide Mononucleotide (NMN) and Nicotinamide Riboside (NR): Potential Dietary Contribution to Health. Curr Nutr Rep. 12(3):445-464. Epub 2023. [CrossRef]
- Loreto A, Antoniou C, Merlini E, Gilley J, Coleman MP. NMN: The NAD precursor at the intersection between axon degeneration and anti-ageing therapies. Neurosci Res. 197:18-24. Epub, 2023. [CrossRef]
- Fang EF, Kassahun H, Croteau DL, Scheibye-Knudsen M, Marosi K, Lu H, Shamanna RA, Kalyanasundaram S, Bollineni RC, Wilson MA, Iser WB, Wollman BN, Morevati M, Li J, Kerr JS, Lu Q, Waltz TB, Tian J, Sinclair DA, Mattson MP, Nilsen H, Bohr VA. NAD(+) Replenishment Improves Lifespan and Healthspan in Ataxia Telangiectasia Models via Mitophagy and DNA Repair. Cell Metab. 24(4):566-581, 2016. [CrossRef]
- Wilson N, Kataura T, Korsgen ME, Sun C, Sarkar S, Korolchuk VI. The autophagy-NAD axis in longevity and disease. Trends Cell Biol. 33(9):788-802. Epub, 2023. [CrossRef]
- Abdulatef Mrghni Ahhmed, Michio Muguruma.A review of meat protein hydrolysates and hypertension. Meat Sci., Sep;86(1):110-118.Epub 2010. [CrossRef]
- Zhou J, Yang M, Han J, Lu C, Li Y, Su X. Effects of dietary tuna dark muscle enzymatic hydrolysis and cooking drip supplementations on growth performance, antioxidant activity and gut microbiota modulation of Bama mini-piglets. RSC Adv. 9(43):25084-25093. eCollection, 2019. [CrossRef]
- M. Ovissipour, A. Abedian Kenari, A. Motamedzadegan, R.M. Nazari. Optimization of enzymatic hydrolysis of visceral waste proteins of Yellowfin Tuna (Thunnus albacares), Food Bioprocess Technol., 5: 696-705, 2021.
- Terauchi K, Kobayashi H, Yatabe K, Yui N, Fujiya H, Niki H, Musha H, Yudoh K. The NAD-Dependent deacetylase sirtuin-1 regulates the expression of osteogenic transcriptional activator runt-related transcription factor 2 (Runx2) and production of matrix metalloproteinase (MMP)-13 in chondrocytes in osteoarthritis. Int. J Molecular Science, 17(7), 2016. [CrossRef]
- Shu Somemura, Takanori Kuma Kanaka Yatabe, Chizuko Sasaki, Hiroto Fujiya 1, Hisateru Niki, Kazuo Yudoh. Physiologic Mechanical Stress Directly Induces Bone Formation by Activating Glucose Transporter 1 (Glut 1) in Osteoblasts, Inducing Signaling via NAD+-Dependent Deacetylase (Sirtuin 1) and Runt-Related Transcription Factor 2 (Runx2). International J. Molecular Science, 22(16), 2021. [CrossRef]
- Guo J, Chiang WC. Mitophagy in aging and longevity. IUBMB Life. 74(4):296-316. Epub, 2021. [CrossRef]
- Onishi M, Yamano K, Sato M, Matsuda N, Okamoto K. Molecular mechanisms and physiological functions of mitophagy. EMBO J. 40(3):e104705. Epub, 2021. [CrossRef]
- Leduc-Gaudet JP, Hussain SNA, Barreiro E, Gouspillou G. Mitochondrial Dynamics and Mitophagy in Skeletal Muscle Health and Aging. Int J Mol Sci. 22(15):8179, 2021. [CrossRef]
- Zhang Y, Bai J, Cui Z, Li Y, Gao Q, Miao Y, Xiong B. Polyamine metabolite spermidine rejuvenates oocyte quality by enhancing mitophagy during female reproductive aging. Nat Aging. 3(11):1372-1386. Epub, 2023. [CrossRef]
- Jiménez-Loygorri JI, Villarejo-Zori B, Viedma-Poyatos Á, Zapata-Muñoz J, Benítez-Fernández R, Frutos-Lisón MD, Tomás-Barberán FA, Espín JC, Area-Gómez E, Gomez-Duran A, Boya P. Mitophagy curtails cytosolic mtDNA-dependent activation of cGAS/STING inflammation during aging. Nat Commun. 15(1):830, 2024. [CrossRef]
- Mercurio L, Morelli M, Scarponi C, Scaglione GL, Pallotta S, Avitabile D, Albanesi C, Madonna S. Enhanced NAMPT-Mediated NAD Salvage Pathway Contributes to Psoriasis Pathogenesis by Amplifying Epithelial Auto-Inflammatory Circuits. Int J Mol Sci. 22(13):6860, 2021. [CrossRef]
- Sharma P, Xu J, Williams K, Easley M, Elder JB, Lonser R, Lang FF, Lapalombella R, Sampath D, Puduvalli VK. Inhibition of nicotinamide phosphoribosyltransferase (NAMPT), the rate-limiting enzyme of the nicotinamide adenine dinucleotide (NAD) salvage pathway, to target glioma heterogeneity through mitochondrial oxidative stress. Neuro Oncol. 24(2):229-244, 2022. [CrossRef]
- Guo X, Tan S, Wang T, Sun R, Li S, Tian P, Li M, Wang Y, Zhang Y, Yan Y, Dong Z, Yan L, Yue X, Wu Z, Li C, Yamagata K, Gao L, Ma C, Li T, Liang X. NAD + salvage governs mitochondrial metabolism, invigorating natural killer cell antitumor immunity. Hepatology. 78(2):468-485. Epub, 2023. [CrossRef]
- Su M, Qiu F, Li Y, Che T, Li N, Zhang S. Mechanisms of the NAD(+) salvage pathway in enhancing skeletal muscle function. Front Cell Dev Biol. 12:14648155. eCollection 2024. [CrossRef]
- Seko T, Uchida H, Sato Y, Imamura S, Ishihara K, Yamashita Y, Yamashita M. Selenoneine Is Methylated in the Bodies of Mice and then Excreted in Urine as Se-Methylselenoneine. Biol Trace Elem Res. 202(8):3672-3685. Epub 2023. [CrossRef]
- Yamashita M, Yamashita Y, Ando T, Wakamiya J, Akiba S. Identification and determination of selenoneine, 2-selenyl-Nα, Nα, Nα-trimethyl-L-histidine, as the major organic selenium in blood cells in a fish-eating population on remote Japanese islands. Biol Trace Elem Res 156:36–44, 2013. [CrossRef]
- Rohn I, Kroepfl N, Aschner M, Bornhorst J, Kuehnelt D, Schwerdtle T. Selenoneine ameliorates peroxide-induced oxidative stress in C. elegans. J Trace Elem Med Biol. 55:78-81. Epub, 2019. [CrossRef]
- Tohfuku T, Ando H, Morishita N, Yamashita M, Kondo M. Dietary Intake of Selenoneine Enhances Antioxidant Activity in the Muscles of the Amberjack Seriola dumerili Grown in Aquaculture. Mar Biotechnol (NY). 23(6):847-853. Epub 2021. [CrossRef]
- Yang Y, Karakhanova S, Hartwig W, D’Haese JG, Philippov PP, Werner J, Bazhin AV. Mitochondria and Mitochondrial ROS in Cancer: Novel Targets for Anticancer Therapy. J Cell Physiol. 231(12):2570-81. Epub, 2016. [CrossRef]
- Angelova PR, Abramov AY. Role of mitochondrial ROS in the brain: from physiology to neurodegeneration. FEBS Lett. 592(5):692-702. Epub, 2018. [CrossRef]
- Annesley SJ, Fisher PR. Mitochondria in Health and Disease. Cells. 5;8(7):680, 2019. [CrossRef]
Figure 1.
The treatment of PBMCs with tuna hydrolysate showed no significant change of the cell viability at protein concentrations less than 9.4 μg/mL. In the present study, PBMCs were treated with digestive enzymatic tuna hydrolysate at the concentration of 4.0 μg/mL in each experiment.
Figure 1.
The treatment of PBMCs with tuna hydrolysate showed no significant change of the cell viability at protein concentrations less than 9.4 μg/mL. In the present study, PBMCs were treated with digestive enzymatic tuna hydrolysate at the concentration of 4.0 μg/mL in each experiment.
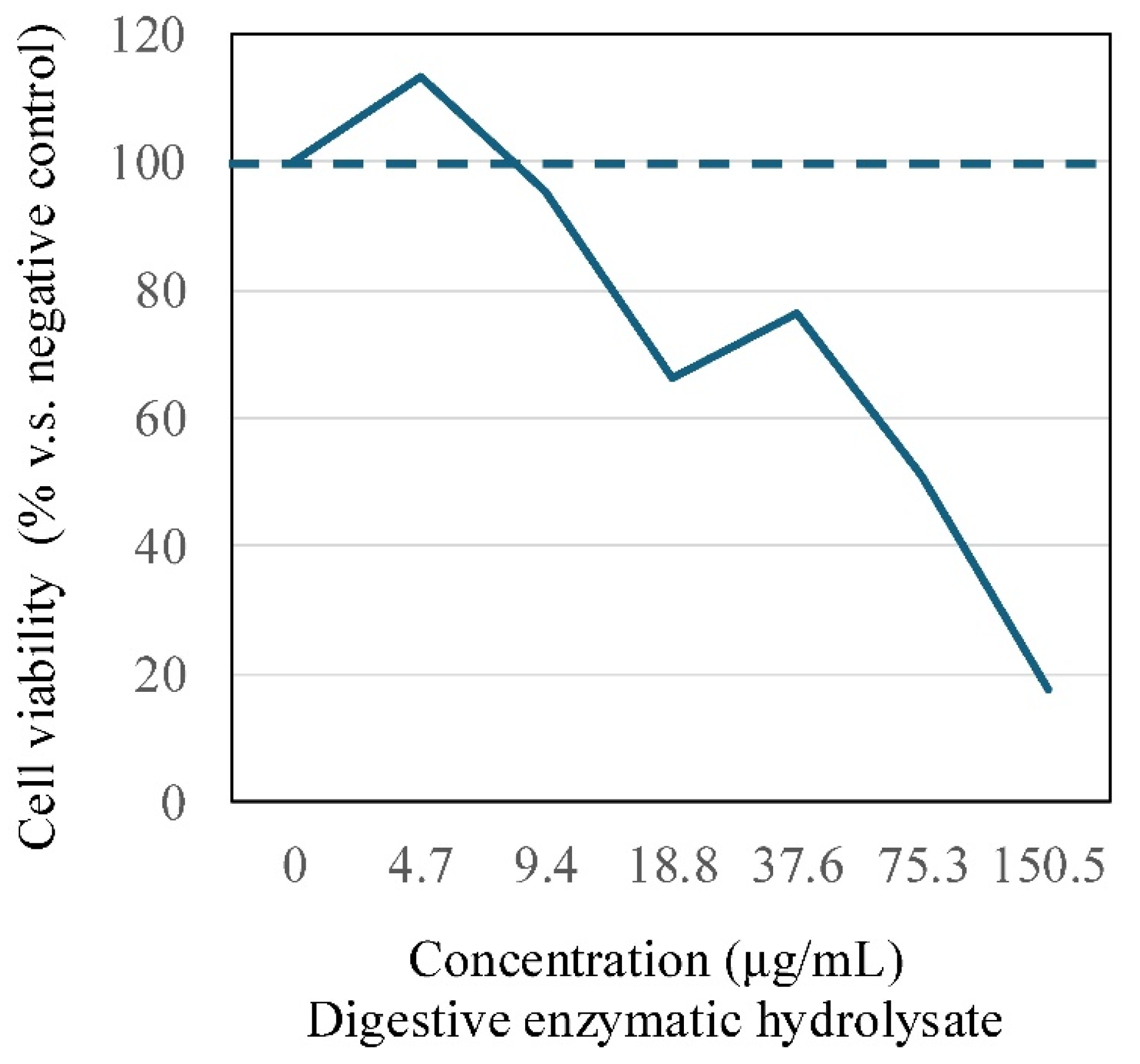
Figure 2.
Effect of digestive enzymatic tuna hydrolysate on the level of nicotinamide adenine dinucleotide (NAD+)/NADH in PBMCs. Both digestive enzymatic hydrolysate and the hot water extract (4 μM), and positive control NMN (10 μM) increased the levels of NAD+/NADH activity in PBMCs [A: oxidized NAD + reduced NAD (NAHD), B: oxidized NAD] (ratio versus negative control, digestive enzymatic hydrolysate: 108 %, hot water extract: 112 %, NMN: 111 %). These findings indicated that tuna meats could activate mitochondria function and NAD-dependent sirtuin proteins in PBMCs.
Figure 2.
Effect of digestive enzymatic tuna hydrolysate on the level of nicotinamide adenine dinucleotide (NAD+)/NADH in PBMCs. Both digestive enzymatic hydrolysate and the hot water extract (4 μM), and positive control NMN (10 μM) increased the levels of NAD+/NADH activity in PBMCs [A: oxidized NAD + reduced NAD (NAHD), B: oxidized NAD] (ratio versus negative control, digestive enzymatic hydrolysate: 108 %, hot water extract: 112 %, NMN: 111 %). These findings indicated that tuna meats could activate mitochondria function and NAD-dependent sirtuin proteins in PBMCs.
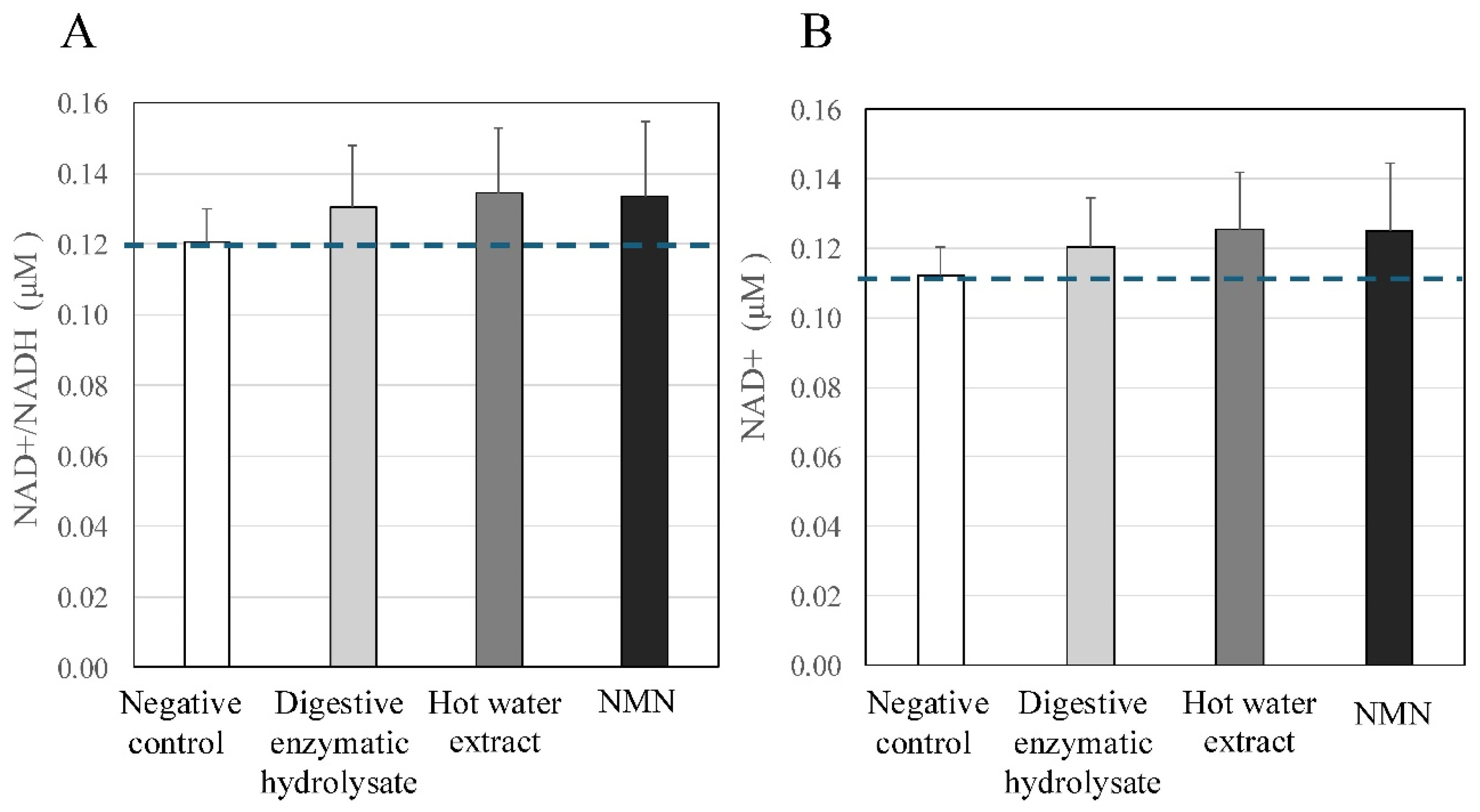
Figure 3.
Effects of mechanical loading on expression of Glut1, SIRT1 and Runx2 in osteoblasts. A: Representative images of western blot analyses for expressions of sirtuin 1 and sirtuin 2 in PBMCs. B: Expression relative to b-actin. Sirtuin 1 protein showed an increasing trend in PBMCs, after treatments with both digestive enzymatic hydrolysate and the hot water extract (4 μM), and positive control NMN (10 μM) (ratio versus negative control, digestive enzymatic hydrolysate: 110 %, hot water extract: 120%, NMN: 116 %). C: Expression relative to b-actin. Sirtuin 2 protein showed an increasing trend in PBMCs, after treatments with both digestive enzymatic hydrolysate and the hot water extract (4 μM), and positive control NMN (10 μM) (ratio versus negative control, digestive enzymatic hydrolysate: 114 %, hot water extract: 129%, NMN: 111 %).
Figure 3.
Effects of mechanical loading on expression of Glut1, SIRT1 and Runx2 in osteoblasts. A: Representative images of western blot analyses for expressions of sirtuin 1 and sirtuin 2 in PBMCs. B: Expression relative to b-actin. Sirtuin 1 protein showed an increasing trend in PBMCs, after treatments with both digestive enzymatic hydrolysate and the hot water extract (4 μM), and positive control NMN (10 μM) (ratio versus negative control, digestive enzymatic hydrolysate: 110 %, hot water extract: 120%, NMN: 116 %). C: Expression relative to b-actin. Sirtuin 2 protein showed an increasing trend in PBMCs, after treatments with both digestive enzymatic hydrolysate and the hot water extract (4 μM), and positive control NMN (10 μM) (ratio versus negative control, digestive enzymatic hydrolysate: 114 %, hot water extract: 129%, NMN: 111 %).
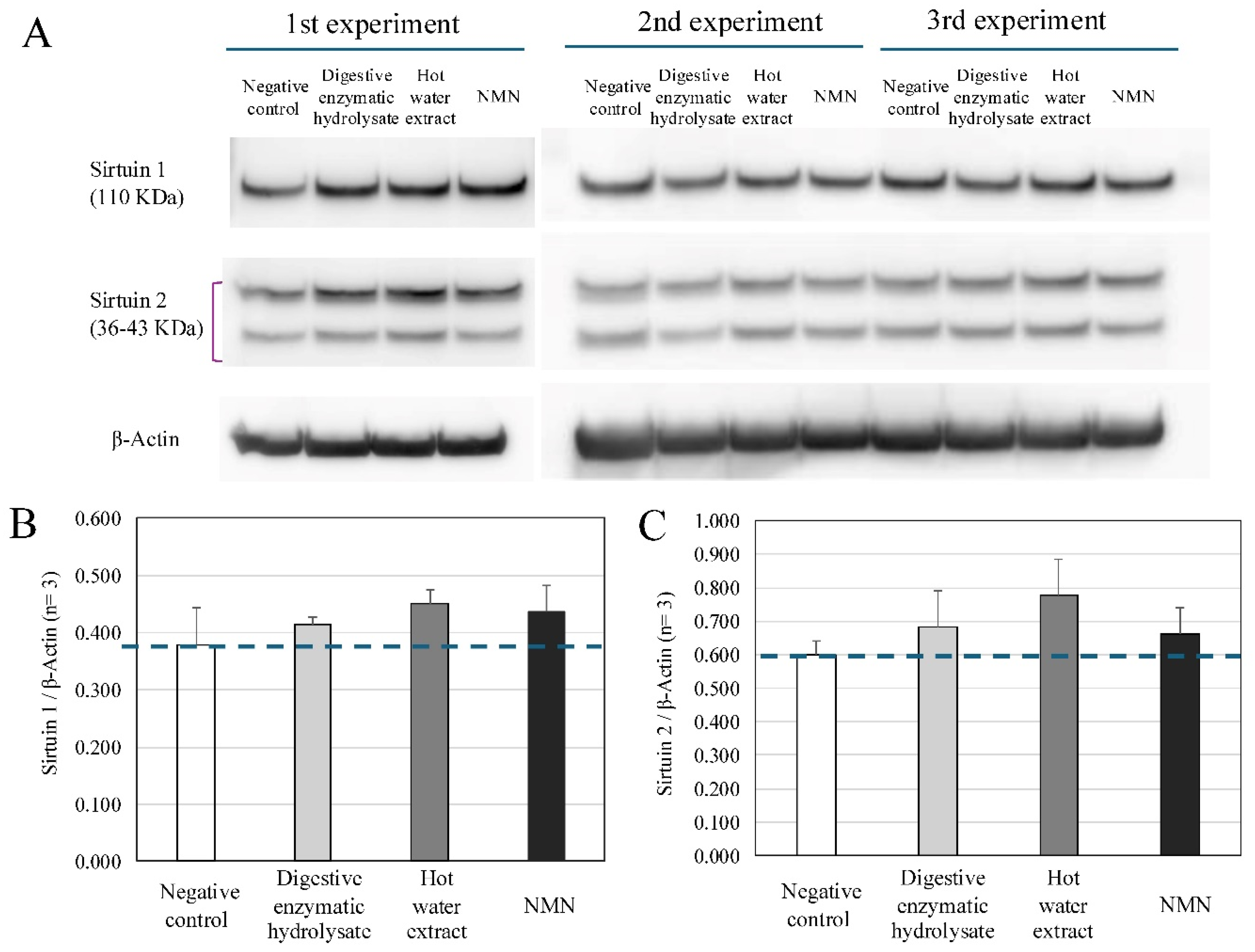
Figure 4.
Effect of digestive enzymatic tuna hydrolysate on the ATP production by PBMCs. Treatments with both tuna extracts (4 μM), the digestive enzymatic hydrolysate and the hot water extract showed a tendency to increase the production of ATP in PBMCs, and treatment of positive control, NMN (10 μM), also accelerated the production of ATP by PBMCs [% ratio versus negative control, digestive enzymatic hydrolysate: 109 %, hot water extract: 111 %, NMN: 115 %]. These findings indicated that tuna meats have a potential to upregulate the energy metabolism of PBMCs.
Figure 4.
Effect of digestive enzymatic tuna hydrolysate on the ATP production by PBMCs. Treatments with both tuna extracts (4 μM), the digestive enzymatic hydrolysate and the hot water extract showed a tendency to increase the production of ATP in PBMCs, and treatment of positive control, NMN (10 μM), also accelerated the production of ATP by PBMCs [% ratio versus negative control, digestive enzymatic hydrolysate: 109 %, hot water extract: 111 %, NMN: 115 %]. These findings indicated that tuna meats have a potential to upregulate the energy metabolism of PBMCs.
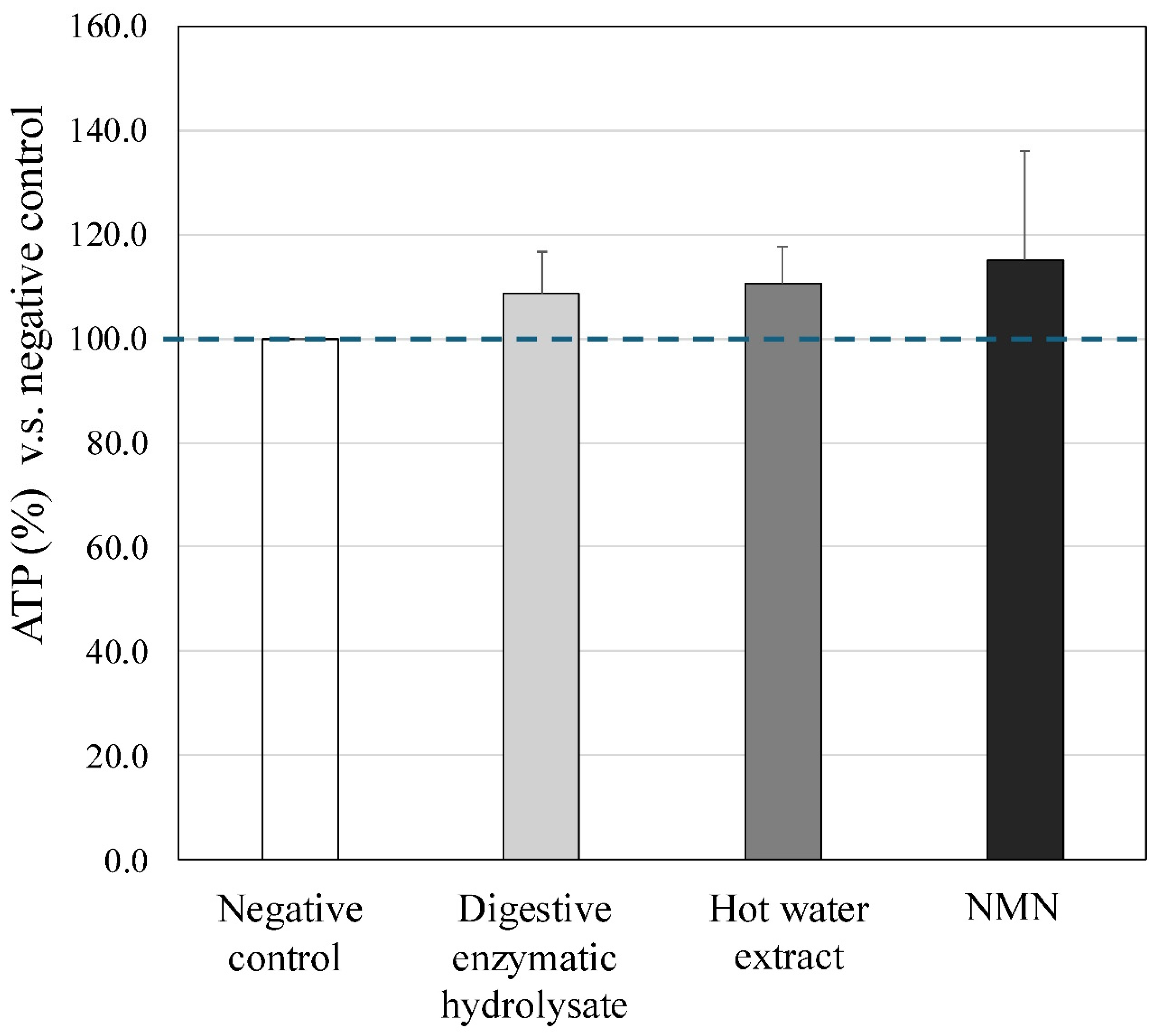
Figure 5.
NAD precursors in tuna meat induces the NAD+ level, NAD-dependent sirtuin activation and energy metabolism. A: NAM phosphoribosyl transferase (NAMPT) -Mediated NAD Salvage Pathway. The NAD precursor NAM, an intermediate metabolite of NAD+, is metabolized to NMN, which enters the cell, is converted to NAD+, and activates sirtuins. The NMN, is synthesized in vivo by NAM phosphoribosyl transferase (NAMPT), using NAM as the main reaction substrate in the salvage NAD+ biosynthesis pathway. B. Effects of tuna meat on expression of the NAD+ level, sirtuin activity, mitochondrial activity, in peripheral blood mononuclear cells. NAD precursors, NAM, in tuna meat has a potential to induce the NAD+ level, the NAD-dependent sirtuin activation, and energy metabolism. NAM: nicotineamide, NMN: nicotinamide mononucleotide, NAD: nicotinamide adenine dinucleotide,.
Figure 5.
NAD precursors in tuna meat induces the NAD+ level, NAD-dependent sirtuin activation and energy metabolism. A: NAM phosphoribosyl transferase (NAMPT) -Mediated NAD Salvage Pathway. The NAD precursor NAM, an intermediate metabolite of NAD+, is metabolized to NMN, which enters the cell, is converted to NAD+, and activates sirtuins. The NMN, is synthesized in vivo by NAM phosphoribosyl transferase (NAMPT), using NAM as the main reaction substrate in the salvage NAD+ biosynthesis pathway. B. Effects of tuna meat on expression of the NAD+ level, sirtuin activity, mitochondrial activity, in peripheral blood mononuclear cells. NAD precursors, NAM, in tuna meat has a potential to induce the NAD+ level, the NAD-dependent sirtuin activation, and energy metabolism. NAM: nicotineamide, NMN: nicotinamide mononucleotide, NAD: nicotinamide adenine dinucleotide,.
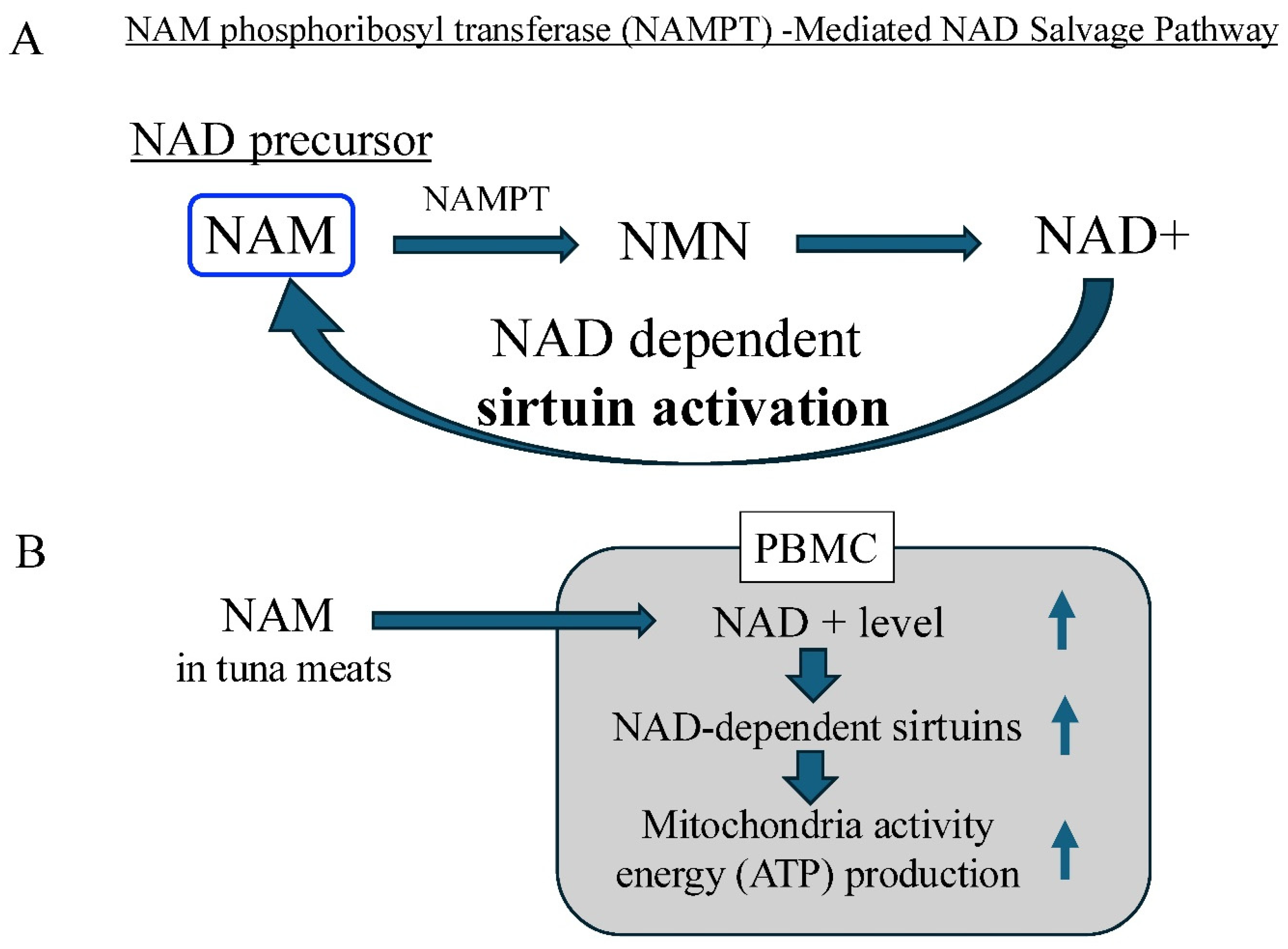

Table 1.
Concentrations of NAD+ precursors, NMN andNAM, in tuna red and dark meats.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



