
Submitted:
18 September 2023
Posted:
20 September 2023
You are already at the latest version
Abstract
Nicotinamide riboside (NR) is an NAD+ precursor capable of regulating mammalian cellular metabolism. Phycocyanin oligopeptide (PC), a phytonutrient found in blue-green algae, has antioxidant and anti-inflammatory properties. This study explored the effects of NR, PC, and their combination on telomere length, inflammatory and antioxidant status in rats under chronic stressed conditions (CS). Forty-nine rats were allocated into seven groups: Control, chronic stress (CS), CS with NR (26.44 mg/kg), a low dose of 2.64 mg/kg of PC (PC-LD), or a high dose of 26.44 mg/kg PC (PC-HD), NR+PC-LD and NR+PC-HF. Rats were given daily corticosterone injections (40 mg/kg) to induce stressed conditions, or NR and PC were orally administered for 21 days. NR and PC supplementation, particularly NR plus PC, increased the serum antioxidant enzyme activities, hepatic nicotinamide adenine (NAD+) content, and telomere length (P<0.001 for all) compared with the CS group. The levels of serum malondialdehyde (MDA), liver interleukin-6 (IL-6), tumor necrosis factor α (TNF-α), IL-1β, and IL-8 were reduced under CS condition (P<0.001). In addition, CS decreased the levels of hepatic telomere-related proteins and sirtuin1 (SIRT1-3), while supplementation of NR and PC alone or in combination increased the levels of telomere-related proteins (e.g., POT1b, TRF1, and TRF2), and SIRT3, NAMPT P<0.05. In conclusion, NR and PC, especially their combination, can alleviate metabolic abnormalities by enhancing hepatic cytokines, SIRT3, NAMPT, and NAD+ levels in CS-exposed rats. More research is needed to elucidate further the potential health effects of the combination of NR and PC in humans.
Keywords:
corticosterone
; phycocyanin
; nicotinamide riboside
; NAD
; hepatic metabolism
1. Introduction
Stress is a rapid and coordinated response by an organism to environmental challenges. These responses require the organism to divert energy resources from one tissue to serve the needs of another to counteract the adverse effects of stress. Glucocorticoids (GCs), a class of adrenal steroids released during and after stress, mediate daily physiological changes and coordinate stress. GCs regulate various physiological progressions, mainly by releasing energy stores through gluconeogenesis and suppressing glucose uptake and activity in specific tissues such as adipose and immune tissue [1,2]. Chronic corticosterone (CORT) exposure increases total body fat mass and decreased insulin sensitivity [3]. The formation of reactive oxygen species (ROS), such as hydrogen peroxide (H2O2), hydroxyl radicals (HO), and superoxide anion radicals (O2-), which lead to lipid peroxidation, is a metabolic consequence of the stress-induced increase in energy production [4]. Additionally, the elevated CORT levels induce redox imbalance and ROS generation in several tissues [5]. Oxidative stress mediates antioxidant defense system changes, which is also associated with chronic stress [6]. Therefore, the body primarily uses enzymatic and non-enzymatic antioxidant defense systems to neutralize ROS [7]. Cellular oxidative stress may reduce telomere length by attenuating telomerase activity, which controls the de novo synthesis of telomeres [8]. Various age-related diseases have been shown to cause cellular damage [9], and cellular senescence increases due to ROS-induced oxidative stress [10].
Nicotinamide adenine dinucleotide (NAD+) plays a crucial role in energy metabolism and genome integrity by acting as a substrate for different enzyme families, such as sirtuins (SIRTs) and poly(ADP-ribose) polymerases (PARPs) [11]. NAD+ levels decrease in cellular oxidative reactions [12] that may occur during metabolic and age-related diseases and neurodegeneration [11]. SIRTs regulate DNA damage and improve mitochondrial biogenesis [13]. The increased NAD+ levels may improve SIRT activity [12] and prevent telomere shortening under stress conditions [14]. Natural NAD+ precursors such as nicotinamide riboside (NR) can alleviate telomere damage by inhibiting ROS production and mitochondrial impairment [12]. NR supplementation can prevent NAD+ decline in mitochondria and improves mitochondrial activity and biogenesis in hepatocytes, thus ameliorating oxidative stress-related pathologies and extending the health span [15]. Similarly, phycocyanin (PC), a biliprotein derived from spirulina, has potent antioxidant and anti-inflammatory effects [16,17]. PC could activate nuclear transcription factor erythroid-2-like factor 2 (Nrf2)/heme oxygenase-1 (HO-1) signaling, which attenuates oxidative stress-induced DNA damage in the liver [18]. A recent study by Komorowski et al. [19] indicated that the novel patented PC oligopeptide might be superior to standard PC owing to its high cellular antioxidant capacity. However, the protective effects of PC oligopeptide and its combination with NR are still unclear. Therefore, this study was conducted to test the effects of PC oligopeptide and NR supplementation on the hepatic abnormalities, NAD+ levels, and underlying mechanisms in rats reared under chronic stress conditions.
2. Materials and Methods
2.1. Animals
A total of 49 male Wistar albino rats (8 weeks old, 180±20g) were used, and animals were reared in polypropylene cages conditioned at a temperature of 22±2°C, 55±5% humidity, and a 12/12h light/dark cycle. A regular chow diet and water were provided ad libitum. The experiment was conducted under the protocol approved by the Firat University Animal Experiments Ethics Committee (16/03/2020-384972) according to The European Parliament and the Council directive (2010/63/EU, 2010) and European Economic Community guidelines (Directive 86/609/EEC, 1986). The animal experiments were carried out in the Experimental Research Center of Firat University (FUDAM). All experimental procedures were reported according to the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines.
2.2. Experimental Design
After a one-week adaptation period, 49 rats were randomly separated into seven groups (n=7) as follows: 1) Control; rats were treated with saline containing tween 80 and DMSO in normal conditions; 2) Chronic stress (CS); rats were injected subcutaneously with corticosterone (CORT, 40 mg/kg) daily for 21 consecutive days, 3) NR; rats were injected with CORT and administered NR (26.44 mg/kg), 4) PC-LD; rats were injected with CORT and administered a low dose of 2.64 mg/kg of PC, 5) PC-HD; rats were injected with CORT and administered a high dose of 26.44 mg/kg PC, 6) NR+PC-LD; rats were injected with CORT and administered NR (26.4 mg/kg) and a low dose of 2.64 mg/kg PC, 7) NR+PC-HD; rats were injected with CORT and administered NR (26.44 mg/kg) and a high dose 26.44 mg/kg PC. NR and PC were administered by oral gavage daily for 21 days.
Doses of PC oligopeptide and NB were determined based on the Human Equivalent Dose for Drug Development, according to [20]. Human doses (30 or 300 mg HED) were converted to animal doses based on body surface area. A conversion factor of 6.17 was used to convert human doses to rat doses. It was determined that low-dose PC equals 2.64 mg/kg, high-dose PC, and NR equals 26.44 mg/kg for rats. CORT (Sigma–Aldrich Co., St. Louis, MO, USA) was suspended in saline with 0.1% Tween 80% and 0.2% DMSO.
2.3. Biochemical Analysis
Serum levels of glucose, cholesterol, blood urea nitrogen (BUN), creatinine, and activities of aspartate aminotransferase (AST), and alanine aminotransferase (ALT) were analyzed with a portable automated chemistry analyzer (Samsung LABGEO PT10, Samsung Electronics Co., Suwon, Korea). Total serum corticosterone (LifeSpan Biosciences, Seattle, WA, USA) levels and antioxidant enzyme activities [superoxide dismutase (SOD), glutathione peroxidase (GSH-Px), and catalase (CAT)] were measured using the relevant commercial kits (MyBioSource, Inc. San Diego, CA, USA) by enzyme-linked immunosorbent assay (ELISA, Elx-800, Bio-Tek Instruments Inc, Vermont, USA). Also, liver NAD+, nicotinamide (NAM), nicotinic acid (NA), nicotinamide adenine dinucleotide phosphate (NADPH), and nicotinamide mononucleotide (NMN) GSH (MyBioSource, Inc. San Diego, CA, USA) levels were measured using a commercial kit following the manufacturer’s instructions by ELISA. The serum MDA levels were analyzed by HPLC with an SPD-20A UV detector (Shimadzu). Tissue samples (300 mg) were homogenized and centrifuged, and supernatants were injected (injection volume, 20 µL) into an HPLC system. The mobile phase was 30 mM KH2PO4-methanol (82.5 + 17.5, v/v %, pH 3.6), the flow rate was 1.2 mL/minute, and detection was at 250 nm.
2.4. Relative Telomere Length Measurement
Telomere length in the liver was determined by quantitative real-time PCR (qPCR). Genomic DNA was obtained from the liver tissues of rats using a commercial kit (DNA mini kit, Qiagen, Germany), and the DNA concentration was determined for each sample by a microvolume spectrophotometer (MaestroNano, Maestrogen Inc., USA). For PCR, 20 ng of DNA was diluted in an SYBR Green PCR Kit (QuantiFast, Qiagen, Germany) with AT1 (F: ACGTGTTCTCAGCATCGACCGCTACC, R: AGAATGATAAGGAAAGGGAACAAGAAGCCC) and telomere (tel 1 F: GGTTTTTGAGGGTGAGGGTGAGGGTGAGGGTGAGGG; tel 2 R: TCCCGACTATCCCTA TCCCTATCCCTATCCCTATCCCTA) primers. For telomere primers, PCR was conducted at 95 °C for 3 min, 95 °C for 15 s (40 cycles), and 54 °C for 1 min on a Rotor-Gene Q machine (Qiagen, Germany). Telomeric DNA normalized to AT1 quantity. Relative telomere length expressed as normalized telomere/single gene copy AT 1 receptor. The relative telomere length of the CORT groups was demonstrated as the relative fold change of the control group.
2.5. Western Blotting Analysis
The levels of inflammatory cytokines [interleukin-6 (IL-6), tumor necrosis factor-alpha (TNF-α), IL-1β, and IL-8], telomere shelterin proteins [protection of telomeres protein 1a (POT-1a), Pot-lb, telomeric repeat-binding factor 1 (TRF1), TRF2, and TRF1-interacting nuclear protein 2 (Tin2)], sirtuin-1 (SIRT1), SIRT3, and nicotinamide phosphoribosyltransferase (NAMPT) in the liver were detected by Western blotting. Liver tissues were mechanically homogenized and pooled at 4 °C in tissue lysis buffer and centrifuged at 15.000 × g for 20 min at 4 °C. Protein quantification was performed by a microvolume spectrophotometer (MaestroNano, Maestrogen Inc.). Then, liver homogenates were mixed with 2x Laemmli buffer (1:1 ratio) and boiled for 5 minutes in microtubes. Protein samples were separated using 10% sodium dodecylsulfate–polyacrylamide gel electrophoresis (SDS–PAGE) and transferred onto nitrocellulose (NC) membranes. NC membranes were blocked with 5% bovine serum albumin for 2 hours. The membranes were incubated with rat-specific primary antibodies diluted to 1:1000 [IL-6, TNF-α, IL-1β, IL-8, POT-1a, Pot-lb, TRF2, Tin2, SIRT1, SIRT3, NAMPT (Santa Cruz Biotechnology, Heidelberg, Germany), and TRF1 (Thermo Fisher Scientific, Waltham, MA, USA)] overnight at 4°C. After washing, the membranes were incubated with appropriate secondary antibodies diluted to 1:5000 (Santa Cruz Biotechnology) for 2h. The bands were visualized using a diaminobenzidine substrate solution. Finally, the membranes were scanned, and protein levels were measured densitometrically. Protein loading was checked by β-actin protein (Santa Cruz Biotechnology, Heidelberg, Germany). The blots were repeated at least three times.
2.6. Statistical Analysis
Sample size (N=49) was calculated based on a power of 85%, an effect size of 0.65, and a P value of 0.05 using the G*power program (Version 3.1.9.3) [21]. Given that assumption, a sample size of seven per group was calculated. The data were analyzed using Statistical Analysis Software (IBM SPSS version 22.0). The Shapiro–Wilk and Levene tests were used to determine the normality of the data and homogeneity of variance, respectively. The groups were compared using ANOVA and Tukey posthoc test. P<0.05 was considered to be statistically significant. Data are shown as the mean ± standard deviation.
3. Results
3.1. The Effects of PC and NR on Body Weight and Biochemical Parameters
CORT administration significantly reduced the final body weight compared to the control group (P<0.001, Table 1). Serum glucose levels were not affected by CORT administration in the CS, NR, PC-LD, and NR+PC-HD groups compared to the control group (P>0.05). However, a dose of 26.44 mg/kg NR plus 2.64 mg/kg PC administration decreased the serum glucose level compared to the control (P<0.05), CS (P<0.001), and NR groups (P<0.05, Table 1). The serum creatine, BUN, ALT, and AST levels did not differ among the groups (P>0.05).
Changes in the concentrations of serum corticosterone, MDA, SOD, GSH-Px, and CAT to determine the level of stress and antioxidant capacity were shown in Table 2. As expected, the serum corticosterone level was markedly increased in the CORT-treated groups compared to the control group (P<0.001). Similarly, the serum MDA levels were elevated after CORT administration compared to the control group (P<0.001). The serum MDA levels were reduced by NR and PC supplementation (P<0.001). An apparent decline of the MDA levels was observed in the NR+PC-LD group compared to the NR (P<0.001), PC-LD (P<0.001), and PC-HD (P<0.05). Conversely, serum SOD, GSH-Px, and CAT levels notably decreased after CORT injection (P<0.001, Table 2). NR supplementation increased the GSH-Px (P<0.05) and CAT levels (P<0.01), while it did not change the serum SOD levels (P>0.05) compared to the CS group. The PC- PC-LD and PC-HD groups had higher serum SOD and CAT levels than the NR group (P<0.05). Furthermore, the combination of NR with each dose of PC prominently boosted serum SOD and GSH-Px levels compared to other groups (P<0.05), except the control group (P>0.05).
3.2. The Effects of PC and NR on Liver Nicotinamide Metabolites and GSH
We tested the liver NAD+, NAM, NA, NMN, and NADPH levels to assess hepatic NAD+ metabolism. The level of liver NAD+ and its metabolites decreased in the CS group compared to other groups (P<0.05, Table 3). We found that PC supplementation elevated NAD+ and NAM levels in rat liver compared to NR supplementation (P<0.05). The NR+PC-LD dose group had markedly higher NAD+ and NADPH levels than the NR (P<0.05) and NR+PC-HD groups (P<0.01). Interestingly, liver NAM concentrations were similar between the NR and NR+PC-HD groups (P>0.05), while this metabolite was remarkably elevated in the NR+PC-LD group compared to the NR group (P<0.05). CORT injection effectively diminished liver GSH status compared to the control group (P<0.001). The NR- and PC-supplemented groups significantly reversed the GSH reduction following CORT injection (P<0.001; Table 3). The NR group had a similar GSH activity to the NR+PC-LD and NR+PC-HD groups (P>0.05). The NR+PC-LD group had a higher GSH activity than the NR+PC-HD group (P<0.05).
3.3. The Effects of PC and NR on Telomere Length
As seen in Figure 1, CORT injection noticeably decreased the telomere length, and the NR+PC-LD group had the most protective activity against telomere shortening among the CORT-injected groups (P<0.001). The relative telomere length was similar between the NR+PC-HD and NR groups (P>0.05).
3.4. The Effects of PC and NR on İnflammatory Mediators
As expected, inflammatory cytokine levels were significantly increased in CORT-injected rats compared to other groups (Figure 2A-E; P<0.01). The hepatic IL-6 (Figure 2A) levels were similar between the control, NR, NR+PC-LD, and NR+PC-HD groups (P>0.05). NR plus low-dose PC-supplemented rats had an inhibitory effect on TNF-α (Figure 2B). It was observed that the NR+PC-LD group had the most inhibitory effect on IL-1β activity compared to the other CORT-injected groups (P<0.001, Figure 2C). The NR (P<0.01) and NR+PC-HD (P<0.05) groups had higher IL-8 levels than the control group (Figure 2D), while NR plus low-dose PC supplementation reduced the hepatic IL-8 level compared to the control group level (P>0.05).
3.5. The Effects of PC and NR on Telomere Sheltering, SIRTs, and NAMPT
We next analyzed protein levels of telomere shelterin complexes by western blotting. As shown in Figure 3 and Figure 4. NR or NR+PC supplementation increased the telomere shelterin complex, SIRT1, SIRT3, and NAMPT protein levels, which were reduced after CORT administration (P<0.05). The control and NR+PC-LD groups had similar POT1a protein levels (P>0.05, Figure 3A). Additionally, we detected that the POT1b levels did not differ between the control, NR, and NR+PC-LD groups (P>0.05, Figure 3B). Although the NR and NR+PC-LD groups had significantly higher TRF1 (Figure 3C) and TRF2 (Figure 3D) levels than the other CORT-administered groups (P<0.01 for all), these groups could not reach the level of TRF1 and TRF2 of the control group. Moreover, we found that Tin2 levels in the NR+PC-LD group noticeably increased compared to those in the CS group (P<0.05, Figure 3E). Unfortunately, none of the CORT-injected groups had significantly elevated Tin2, TRF1, or TRF2 levels compared to the control (P<0.05).
In order to explore the effect of NR and PC oligopeptides on telomere lengthening, we measured the levels of SIRT1, SIRT3, and NAMPT, which are NAD+-related enzymes. The NR group had the highest SIRT1 levels among the CORT-injected groups (Figure 4A). Any doses of PC without NR did not alter the SIRT1 level compared to the CS group (P>0.05, Figure 4A). Similarly, the PC-LD and NR+PC-HD did not significantly increase the SIRT3 levels compared to the CS group (P>0.05, Figure 4B). However, the NR and NR+PC-LD groups had the highest SIRT3 levels among the CORT-injected groups (P<0.05, Figure 4B). On the other hand, the NR+PC-LD group remarkably boosted the NAMPT levels compared to the other groups, except the control (P<0.05, Figure 4C). Additionally, the NR+PC-LD group had a higher NAMPT level than the control group, but the difference was not statistically significant (P>0.05, Figure 4C).
4. Discussion
This study showed that chronic CORT exposure increased oxidative stress by increasing MDA content and decreasing the antioxidant activity of SOD, CAT, and GSH-Px. At the same time, it also increased serum corticosterone concentrations and liver anti-inflammatory (TGF-β) and pro-inflammatory (TNF-α, IL-1β, IL-6) cytokines levels. Similar to our results, it has been well documented in previous studies that chronic CORT exposure increases MDA levels [22], stimulates inflammatory cytokines [22,23], suppresses antioxidant enzyme activity [23], and accelerates telomere shortening [23,24]. As in our findings, it has been reported that NR and PC may exert inhibitory effects on oxidative stress-induced tissue damage in animals [16,25] and humans [12,25] by attenuating short telomere-induced DNA damage [12].
CORT-induced oxidative and inflammatory status probably increases PARP1 and cyclic ADP ribose hydrolase (CD38) activation, which are the primary consumers of NAD+, and restricts NAD+ bioavailability to SIRTs [26] that prevent liver damage, fat accumulation, and fibrosis [27]. Therefore, in the present study, the limited NAD+ bioavailability caused by CORT-induced liver inflammation may have reduced SIRT1 and SIRT3 levels in the liver in CORT rats. NR can enter the NAD+ salvage pathway via phosphorylation to NMN by nicotinamide riboside kinases (NRKs), and NMN is converted into NAD+ to feed the NAD+ biosynthesis cycle [28,29]. Oral NR supplementation achieves the replenishment of cellular NAD+ reduced by inflammation and may diminish circulating inflammatory cytokine activities [30]. Thus, NR can attenuate oxidative inflammatory tissue damage by inhibiting mitochondrial ROS production and reducing MDA levels while elevating SOD levels [31,32]. Analogously, PC supplementation boosts antioxidant enzyme levels in the liver and alleviates hepatic oxidative stress owing to its radical scavenging, lipid peroxidation prevention [33], and mitochondrial respiratory complex protective activity [34]. The anti-inflammatory ability of PC may stem from its inhibitory action on activating programmed cell death five protein (PCDP5)/nuclear factor κ-B or tool-like receptor 2/NF-κ-B pathways [35,36].
In the present study, NR plus PC supplementation effectively increased liver NAD+, NAM, and NADPH levels. Similar to our results, It has been shown that PC has inhibitory activity on NADPH oxidase (NOX) [37], producing ROS by utilizing NADPH [26]. Fan et al. [38] showed that NOX inhibition augments NADPH and NAD+ levels. Based on Fan et al. [38]’s report, we assumed that NADPH levels increased following NOX inhibition by PC, and NAD+ and NAM levels were elevated in parallel depending on the reduced NADPH requirement. The elevated NAM level might have stimulated NAMPT upregulation in the NR plus low-dose PC-supplemented group to promote the NAD+ salvage pathway. NAMPT can mediate the activity of NAD+-dependent enzymes and alleviates hepatic steatosis in a SIRT-dependent manner in mice [39]. NAMPT overexpression can independently influence liver regeneration, and this regeneration process may also be restored by NR treatment in NAMPT knockout mice through SIRT1-dependent pathways [40]. In contrast to other spirulina-derived PCs [41], we found that novel PC oligopeptides could not have effectively stimulated SIRT1 while increasing the SIRT3 levels in the liver. Also, NR+PC supplementation was not as effective as NR supplementation in elevating SIRT3 levels. Under inflammatory conditions, increased NAM levels by phytochemical administration may negatively affect SIRT1 activity due to the activation of the PARP1-dependent anti-inflammatory response pathway [42]. Although we could not demonstrate why SIRT1 was not stimulated by PC, our results suggested that the PC oligopeptide exerts SIRT1-independent anti-inflammatory effects in the liver of CORT-treated rats.
We also explored the effects of NR and PC on telomere shelterin complex proteins. Recently, Shen et al. [43] showed that CORT exposure impaired the telomere shelterin protein complex and reduced telomere length in the liver of rats. TRF1 has both lengthening and shortening effects on telomeres to avoid DNA damage and protect telomere stability [44]. Similar to our results, Badmus et al. [45] reported that liver TRF1 levels decreased after four weeks of CORT feeding in broiler chickens. TRF2 positively regulates telomere function and continually interacts with TRF1 and POT1 to control telomere length [46]. An in vitro study showed that oxidative damage could interfere with telomere length by suppressing TRF2 activity [47]. Xie et al. [24] indicated that CORT exposure reduced liver TRF2 activity in rats. In parallel with these results, we found that POT1a and POT1b levels were reduced after CORT injection because POT1 binding to the TRF1 and TRF2 complex is mediated by TIN2-interacting protein (TPP1) and TIN2 [48]. The interaction between POT1, TPP1, and TIN2 orchestrates the functions of telomere shelterin complex proteins and telomere integrity [49]. Shen et al. [43] demonstrated that liver TIN2 and POT1a levels decreased after CORT exposure in rats, and this report supports our findings. Promoting NAD+ metabolism after NR supplementation may prevent ROS-induced DNA damage and mitochondrial impairment in DC fibroblasts [12]. Because NR activates the NAD+/SIRT1 pathway, it increases mitochondrial biogenesis and decreases ROS generation; thus, NR may prevent telomere shortening [12]. On the other hand, PC supplementation presumably regulates telomere shelterin proteins, owing to its potent antioxidant activity [19,50] and protective effect on the mitochondrial respiratory complex [34] rather than the SIRT1-dependent pathway. Although we did not measure the liver mTOR levels, PC might have increased the telomere length by diminishing mTOR complex activation, as reported before by Joly et al. [51]. Furthermore, we elucidate that PC partly stimulated SIRT3 activity, which improved mitochondrial biogenesis and antioxidant defense [52]. Collectively, NR plus PC administration may have concurrently prevented hepatic telomere shortening in CORT rats.
Interestingly, this formulation was less effective than the low-dose PC formulation when combined with the NR high-dose PC oligopeptide. This is because antioxidants exert hormetic effects on the redox state and their mechanisms of action may vary depending on the dose [53]. Thus, the protective effect of NR on hepatic telomere length, inflammation, and NAD+ metabolism may have been attenuated due to the cumulative activity of NR plus high-dose PC supplementation.
5. Conclusions
In conclusion, the present study showed that NR and PC oligopeptide supplementation in CORT-treated rats might be relevant in improving hepatic telomere shortening, energy metabolism by regulating the response of the liver through a metabolic increase in both cellular NAD+ availability and hepatic cytokines, SIRT-3, and NAMPT pathways. However, more preclinical and clinical studies are needed to reveal the molecular mechanisms of action of these combinations.
Author Contributions
Kazim Sahin contributed to study design, analyses, editing and methodology; Cemal Orhan, Emre Sahin, Mehmet Tuzcu, Abdullah Celik and Nurhan Sahin investigation, data curation, and formal analysis; James R. Komorowski, Sara Perez Ojalvo, and Sarah Sylla writing–original draft preparation. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by Nutrition 21 LLC (Harrison, NY, USA) and the Turkish Academy of Sciences (in part, K.S.). The funders were not involved in the study design, collection, analysis, and interpretation of data or the decision to submit it for publication.
Institutional Review Board Statement
The experiment was conducted under the protocol approved by the Firat University Animal Experiments Ethics Committee (16/03/2020-384972).
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets for this study can be found in the article.
Acknowledgments
Authors thank to Nutrition 21 LLC (Harrison, NY, USA) and the Turkish Academy of Sciences.
Conflicts of Interest
James R. Komorowski, Sara Perez Ojalvo, and Sarah Sylla are employee of Nutrition 21 LLC (Harrison, NY, USA). The remaining authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Dhabhar, F.S. Effects of Stress on Immune Function: The Good, the Bad, and the Beautiful. Immunol Res 2014, 58, 193–210. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.G.; Seligsohn, M.; Rubin, T.G.; Griffiths, B.B.; Ozdemir, Y.; Pfaff, D.W.; Datson, N.A.; McEwen, B.S. Stress and Corticosteroids Regulate Rat Hippocampal Mitochondrial DNA Gene Expression via the Glucocorticoid Receptor. Proc. Natl. Acad. Sci. U.S.A. 2016, 113, 9099–9104. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.J.; Batdorf, H.M.; Huang, T.-Y.; Jackson, J.W.; Jones, K.A.; Martin, T.M.; Rohli, K.E.; Karlstad, M.D.; Sparer, T.E.; Burk, D.H.; et al. One Week of Continuous Corticosterone Exposure Impairs Hepatic Metabolic Flexibility, Promotes Islet β-Cell Proliferation, and Reduces Physical Activity in Male C57BL/6 J Mice. The Journal of Steroid Biochemistry and Molecular Biology 2019, 195, 105468. [Google Scholar] [CrossRef] [PubMed]
- Spiers, J.G.; Chen, H.-J.C.; Cuffe, J.S.M.; Sernia, C.; Lavidis, N.A. Acute Restraint Stress Induces Rapid Changes in Central Redox Status and Protective Antioxidant Genes in Rats. Psychoneuroendocrinology 2016, 67, 104–112. [Google Scholar] [CrossRef]
- Daiber, A.; Kröller-Schön, S.; Oelze, M.; Hahad, O.; Li, H.; Schulz, R.; Steven, S.; Münzel, T. Oxidative Stress and Inflammation Contribute to Traffic Noise-Induced Vascular and Cerebral Dysfunction via Uncoupling of Nitric Oxide Synthases. Redox Biology 2020, 34, 101506. [Google Scholar] [CrossRef]
- Jiang, Y.; Botchway, B.O.A.; Hu, Z.; Fang, M. Overexpression of SIRT1 Inhibits Corticosterone-Induced Autophagy. Neuroscience 2019, 411, 11–22. [Google Scholar] [CrossRef]
- Caixeta, D.C.; Teixeira, R.R.; Peixoto, L.G.; Machado, H.L.; Baptista, N.B.; De Souza, A.V.; Vilela, D.D.; Franci, C.R.; Salmen Espindola, F. Adaptogenic Potential of Royal Jelly in Liver of Rats Exposed to Chronic Stress. PLoS ONE 2018, 13, e0191889. [Google Scholar] [CrossRef]
- Mendes-Silva, A.P.; Vieira, E.L.M.; Xavier, G.; Barroso, L.S.S.; Bertola, L.; Martins, E.A.R.; Brietzke, E.M.; Belangero, S.I.N.; Diniz, B.S. Telomere Shortening in Late-life Depression: A Potential Marker of Depression Severity. Brain and Behavior 2021, 11, e2255. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative Stress, Aging, and Diseases. CIA 2018, Volume 13, 757–772. [Google Scholar] [CrossRef]
- Pole, A.; Dimri, M.; P. Dimri, G.; Department of Biochemistry and Molecular Medicine, School of Medicine and Health Sciences, The George Washington University, Washington DC, USA Oxidative Stress, Cellular Senescence and Ageing. AIMS Molecular Science 2016, 3, 300–324. [Google Scholar] [CrossRef]
- Griffiths, H.B.S.; Williams, C.; King, S.J.; Allison, S.J. Nicotinamide Adenine Dinucleotide (NAD+): Essential Redox Metabolite, Co-Substrate and an Anti-Cancer and Anti-Ageing Therapeutic Target. Biochemical Society Transactions 2020, 48, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Wang, K.; Stock, A.J.; Gong, Y.; Demarest, T.G.; Yang, B.; Giri, N.; Harrington, L.; Alter, B.P.; Savage, S.A.; et al. Re-equilibration of Imbalanced NAD Metabolism Ameliorates the Impact of Telomere Dysfunction. The EMBO Journal 2020, 39, e103420. [Google Scholar] [CrossRef] [PubMed]
- Cantó, C.; Menzies, K.J.; Auwerx, J. NAD+ Metabolism and the Control of Energy Homeostasis: A Balancing Act between Mitochondria and the Nucleus. Cell Metabolism 2015, 22, 31–53. [Google Scholar] [CrossRef]
- Lin, J.; Epel, E. Stress and Telomere Shortening: Insights from Cellular Mechanisms. Ageing Research Reviews 2022, 73, 101507. [Google Scholar] [CrossRef] [PubMed]
- Vannini, N.; Campos, V.; Girotra, M.; Trachsel, V.; Rojas-Sutterlin, S.; Tratwal, J.; Ragusa, S.; Stefanidis, E.; Ryu, D.; Rainer, P.Y.; et al. The NAD-Booster Nicotinamide Riboside Potently Stimulates Hematopoiesis through Increased Mitochondrial Clearance. Cell Stem Cell 2019, 24, 405–418. [Google Scholar] [CrossRef]
- Li, Y.-J.; Han, Z.; Ge, L.; Zhou, C.-J.; Zhao, Y.-F.; Wang, D.-H.; Ren, J.; Niu, X.-X.; Liang, C.-G. C-Phycocyanin Protects against Low Fertility by Inhibiting Reactive Oxygen Species in Aging Mice. Oncotarget 2016, 7, 17393–17409. [Google Scholar] [CrossRef]
- Singh, N.K.; Sonani, R.R.; Awasthi, A.; Prasad, B.; Patel, A.R.; Kumar, J.; Madamwar, D. Phycocyanin Moderates Aging and Proteotoxicity in Caenorhabditis Elegans. J Appl Phycol 2016, 28, 2407–2417. [Google Scholar] [CrossRef]
- Liu, Q.; Li, W.; Qin, S. Therapeutic Effect of Phycocyanin on Acute Liver Oxidative Damage Caused by X-Ray. Biomedicine & Pharmacotherapy 2020, 130, 110553. [Google Scholar] [CrossRef]
- Komorowski, J.; Bernsley, D.; Sylla, S.; Ojalvo, S.P. Enhanced Antioxidant Activity of Phycocyanin Oligopeptides. Current Developments in Nutrition 2021, 5, 336. [Google Scholar] [CrossRef]
- 20. Jang-Woo Shin; In-Chan Seol; Chang-Gue Son; Jang-Woo Shin; In-Chan Seol; Chang-Gue Son Interpretation of Animal Dose and Human Equivalent Dose for Drug Development. Journal of Korean Medicine 31.
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A Flexible Statistical Power Analysis Program for the Social, Behavioral, and Biomedical Sciences. Behavior Research Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Song, L.; Wu, X.; Wang, J.; Guan, Y.; Zhang, Y.; Gong, M.; Wang, Y.; Li, B. Antidepressant Effect of Catalpol on Corticosterone-Induced Depressive-like Behavior Involves the Inhibition of HPA Axis Hyperactivity, Central Inflammation and Oxidative Damage Probably via Dual Regulation of NF-κB and Nrf2. Brain Research Bulletin 2021, 177, 81–91. [Google Scholar] [CrossRef]
- Ma, L.; Shen, Q.; Yang, S.; Xie, X.; Xiao, Q.; Yu, C.; Cao, L.; Fu, Z. Effect of Chronic Corticosterone-Induced Depression on Circadian Rhythms and Age-Related Phenotypes in Mice. ABBS 2018, 50, 1236–1246. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Shen, Q.; Ma, L.; Chen, Y.; Zhao, B.; Fu, Z. Chronic Corticosterone-Induced Depression Mediates Premature Aging in Rats. Journal of Affective Disorders 2018, 229, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Chini, C.C.S.; Tarragó, M.G.; Chini, E.N. NAD and the Aging Process: Role in Life, Death and Everything in Between. Molecular and Cellular Endocrinology 2017, 455, 62–74. [Google Scholar] [CrossRef]
- Xie, N.; Zhang, L.; Gao, W.; Huang, C.; Huber, P.E.; Zhou, X.; Li, C.; Shen, G.; Zou, B. NAD+ Metabolism: Pathophysiologic Mechanisms and Therapeutic Potential. Sig Transduct Target Ther 2020, 5, 227. [Google Scholar] [CrossRef]
- Zhao, X.; Xue, X.; Wang, C.; Wang, J.; Peng, C.; Li, Y. Emerging Roles of Sirtuins in Alleviating Alcoholic Liver Disease: A Comprehensive Review. International Immunopharmacology 2022, 108, 108712. [Google Scholar] [CrossRef] [PubMed]
- Garten, A.; Schuster, S.; Penke, M.; Gorski, T.; De Giorgis, T.; Kiess, W. Physiological and Pathophysiological Roles of NAMPT and NAD Metabolism. Nat Rev Endocrinol 2015, 11, 535–546. [Google Scholar] [CrossRef]
- Stock, A.J.; Liu, Y. NAD-Linked Metabolism and Intervention in Short Telomere Syndromes and Murine Models of Telomere Dysfunction. Front. Aging 2021, 2, 785171. [Google Scholar] [CrossRef]
- Elhassan, Y.S.; Kluckova, K.; Fletcher, R.S.; Schmidt, M.S.; Garten, A.; Doig, C.L.; Cartwright, D.M.; Oakey, L.; Burley, C.V.; Jenkinson, N.; et al. Nicotinamide Riboside Augments the Aged Human Skeletal Muscle NAD+ Metabolome and Induces Transcriptomic and Anti-Inflammatory Signatures. Cell Reports 2019, 28, 1717–1728. [Google Scholar] [CrossRef]
- Hong, G.; Zheng, D.; Zhang, L.; Ni, R.; Wang, G.; Fan, G.-C.; Lu, Z.; Peng, T. Administration of Nicotinamide Riboside Prevents Oxidative Stress and Organ Injury in Sepsis. Free Radical Biology and Medicine 2018, 123, 125–137. [Google Scholar] [CrossRef]
- Ma, S.; Feng, J.; Lin, X.; Liu, J.; Tang, Y.; Nie, S.; Gong, J.; Wang, L. Nicotinamide Riboside Alleviates Cardiac Dysfunction and Remodeling in Pressure Overload Cardiac Hypertrophy. Oxidative Medicine and Cellular Longevity 2021, 2021, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.M.A.; Ali, H.A.; Ahmed, M.M. Ameliorative Effects of Phycocyanin against Gibberellic Acid Induced Hepatotoxicity. Pesticide Biochemistry and Physiology 2015, 119, 28–32. [Google Scholar] [CrossRef]
- Fernández-Rojas, B.; Rodríguez-Rangel, D.S.; Granados-Castro, L.F.; Negrette-Guzmán, M.; León-Contreras, J.C.; Hernández-Pando, R.; Molina-Jijón, E.; Reyes, J.L.; Zazueta, C.; Pedraza-Chaverri, J. C-Phycocyanin Prevents Cisplatin-Induced Mitochondrial Dysfunction and Oxidative Stress. Mol Cell Biochem 2015, 406, 183–197. [Google Scholar] [CrossRef]
- Hao, S.; Yan, Y.; Huang, W.; Gai, F.; Wang, J.; Liu, L.; Wang, C. C-Phycocyanin Reduces Inflammation by Inhibiting NF-κB Activity through Downregulating PDCD5 in Lipopolysaccharide-Induced RAW 264.7 Macrophages. Journal of Functional Foods 2018, 42, 21–29. [Google Scholar] [CrossRef]
- Liu, Q.; Li, W.; Lu, L.; Liu, B.; Du, Z.; Qin, S. Phycocyanin Attenuates X-Ray-Induced Pulmonary Inflammation via the TLR2-MyD88-NF-κB Signaling Pathway. J. Ocean. Limnol. 2019, 37, 1678–1685. [Google Scholar] [CrossRef]
- McCarty, M.F.; Barroso-Aranda, J.; Contreras, F. Oral Phycocyanobilin May Diminish the Pathogenicity of Activated Brain Microglia in Neurodegenerative Disorders. Medical Hypotheses 2010, 74, 601–605. [Google Scholar] [CrossRef]
- Fan, C.; Ma, Q.; Xu, M.; Qiao, Y.; Zhang, Y.; Li, P.; Bi, Y.; Tang, M. Ginsenoside Rb1 Attenuates High Glucose-Induced Oxidative Injury via the NAD-PARP-SIRT Axis in Rat Retinal Capillary Endothelial Cells. IJMS 2019, 20, 4936. [Google Scholar] [CrossRef]
- Xiong, X.; Yu, J.; Fan, R.; Zhang, C.; Xu, L.; Sun, X.; Huang, Y.; Wang, Q.; Ruan, H.-B.; Qian, X. NAMPT Overexpression Alleviates Alcohol-Induced Hepatic Steatosis in Mice. PLoS ONE 2019, 14, e0212523. [Google Scholar] [CrossRef]
- Mukherjee, S.; Chellappa, K.; Moffitt, A.; Ndungu, J.; Dellinger, R.W.; Davis, J.G.; Agarwal, B.; Baur, J.A. Nicotinamide Adenine Dinucleotide Biosynthesis Promotes Liver Regeneration. Hepatology 2017, 65, 616–630. [Google Scholar] [CrossRef]
- Ren, Z.; Xie, Z.; Cao, D.; Gong, M.; Yang, L.; Zhou, Z.; Ou, Y. C-Phycocyanin Inhibits Hepatic Gluconeogenesis and Increases Glycogen Synthesis via Activating Akt and AMPK in Insulin Resistance Hepatocytes. Food Funct. 2018, 9, 2829–2839. [Google Scholar] [CrossRef]
- Yanez, M.; Jhanji, M.; Murphy, K.; Gower, R.M.; Sajish, M.; Jabbarzadeh, E. Nicotinamide Augments the Anti-Inflammatory Properties of Resveratrol through PARP1 Activation. Sci Rep 2019, 9, 10219. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Wu, J.; Ni, Y.; Xie, X.; Yu, C.; Xiao, Q.; Zhou, J.; Wang, X.; Fu, Z. Exposure to Jet Lag Aggravates Depression-like Behaviors and Age-Related Phenotypes in Rats Subject to Chronic Corticosterone. ABBS 2019, 51, 834–844. [Google Scholar] [CrossRef] [PubMed]
- McKerlie, M.; Zhu, X.-D. Cyclin B-Dependent Kinase 1 Regulates Human TRF1 to Modulate the Resolution of Sister Telomeres. Nat Commun 2011, 2, 371. [Google Scholar] [CrossRef] [PubMed]
- Badmus, K.A.; Idrus, Z.; Meng, G.Y.; Sazili, A.Q.; Mamat-Hamidi, K. Telomere Length and Regulatory Genes as Novel Stress Biomarkers and Their Diversities in Broiler Chickens (Gallus Gallus Domesticus) Subjected to Corticosterone Feeding. Animals 2021, 11, 2759. [Google Scholar] [CrossRef] [PubMed]
- De Lange, T. Shelterin: The Protein Complex That Shapes and Safeguards Human Telomeres. Genes Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef]
- Coluzzi, E.; Leone, S.; Sgura, A. Oxidative Stress Induces Telomere Dysfunction and Senescence by Replication Fork Arrest. Cells 2019, 8, 19. [Google Scholar] [CrossRef]
- Srinivas, N.; Rachakonda, S.; Kumar, R. Telomeres and Telomere Length: A General Overview. Cancers 2020, 12, 558. [Google Scholar] [CrossRef]
- Pike, A.M.; Strong, M.A.; Ouyang, J.P.T.; Greider, C.W. TIN2 Functions with TPP1/POT1 To Stimulate Telomerase Processivity. Molecular and Cellular Biology 2019, 39, e00593–18. [Google Scholar] [CrossRef]
- Niu, Y.-J.; Zhou, W.; Guo, J.; Nie, Z.-W.; Shin, K.-T.; Kim, N.-H.; Lv, W.-F.; Cui, X.-S. C-Phycocyanin Protects against Mitochondrial Dysfunction and Oxidative Stress in Parthenogenetic Porcine Embryos. Sci Rep 2017, 7, 16992. [Google Scholar] [CrossRef]
- Joly, F.; Branka, J.-E.; Darnis, E.; Lefeuvre, L. Telomere Protective Effects of a Cyanobacteria Phycocyanin against Blue Light and UV Irradiations: A Skin Anti-Aging and Photo-Protective Agent. JCDSA 2019, 09, 336–345. [Google Scholar] [CrossRef]
- Dikalova, A.E.; Pandey, A.; Xiao, L.; Arslanbaeva, L.; Sidorova, T.; Lopez, M.G.; Billings, F.T.; Verdin, E.; Auwerx, J.; Harrison, D.G.; et al. Mitochondrial Deacetylase Sirt3 Reduces Vascular Dysfunction and Hypertension While Sirt3 Depletion in Essential Hypertension Is Linked to Vascular Inflammation and Oxidative Stress. Circ Res 2020, 126, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, V.; Cornelius, C.; Trovato-Salinaro, A.; Cambria, M.; Locascio, M.; Rienzo, L.; Condorelli, D.; Mancuso, C.; De Lorenzo, A.; Calabrese, E. The Hormetic Role of Dietary Antioxidants in Free Radical-Related Diseases. CPD 2010, 16, 877–883. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of NR and PC oligopeptides on liver relative telomere length in rats subject to chronic corticosterone (CORT). Data are expressed as a relative fold change compared to the control. The error bars above the lines indicate the standard deviation of the mean. Different symbols (a-f) indicate significant differences among the groups (ANOVA and Tukey post hoc test; P<0.05). CS: chronic stress; NR: Nicotinamide riboside (26.44 mg/kg); PC-LD: Phycocyanin oligopeptide low dose (2.64 mg/kg); PC-HD: Phycocyanin oligopeptide high dose (26.44 mg/kg). Rats except the control group were given daily corticosterone injections (40 mg/kg) to induce stressed conditions, or NR and PC were orally administered for 21 days.
Figure 1.
Effects of NR and PC oligopeptides on liver relative telomere length in rats subject to chronic corticosterone (CORT). Data are expressed as a relative fold change compared to the control. The error bars above the lines indicate the standard deviation of the mean. Different symbols (a-f) indicate significant differences among the groups (ANOVA and Tukey post hoc test; P<0.05). CS: chronic stress; NR: Nicotinamide riboside (26.44 mg/kg); PC-LD: Phycocyanin oligopeptide low dose (2.64 mg/kg); PC-HD: Phycocyanin oligopeptide high dose (26.44 mg/kg). Rats except the control group were given daily corticosterone injections (40 mg/kg) to induce stressed conditions, or NR and PC were orally administered for 21 days.
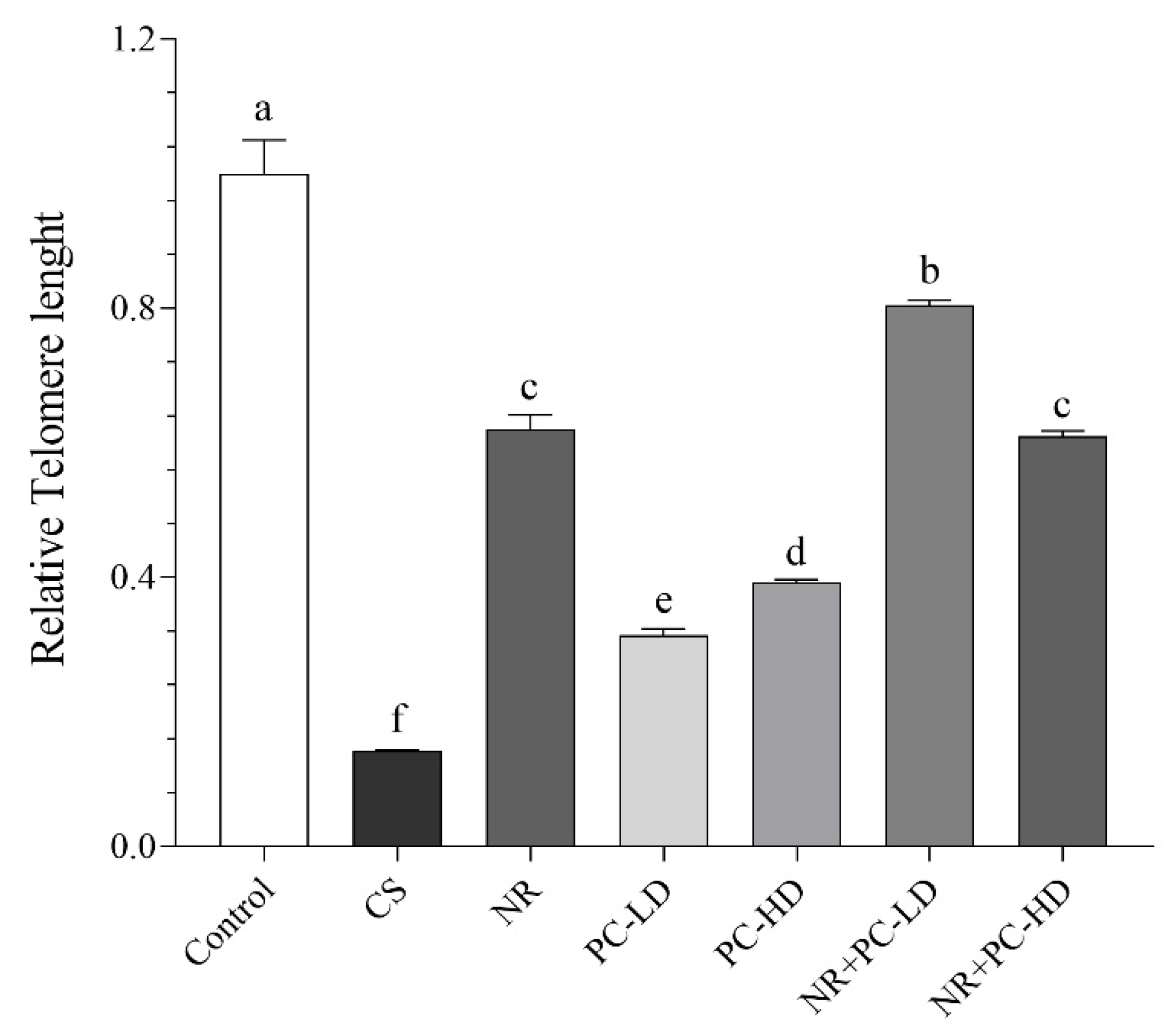
Figure 2.
Effects of NR and PC oligopeptides on liver IL-6 (A), TNF-α (B), IL-1β (C) and IL-8 (D) levels in rats subject to chronic corticosterone (CORT). The densitometric analysis of the relative intensity according to the control group of the western blotting bands was performed with β-actin normalization to ensure equal protein loading (E). Blots were repeated at least three times (n = 3), and a representative blot was shown. Data are expressed as a percent of the control set at 100%. The error bars above the lines indicate the standard deviation of the mean. Different symbols (a-f) indicate significant differences among the groups (ANOVA and Tukey post hoc test; P<0.05). IL-6, interleukin-6; TNF-α, tumor necrosis factor α; IL-1β, interleukin-1β; IL-8, interleukin-8. CS: chronic stress; NR: Nicotinamide riboside (26.44 mg/kg); PC-LD: Phycocyanin oligopeptide low dose (2.64 mg/kg); PC-HD: Phycocyanin oligopeptide high dose (26.44 mg/kg). Rats except the control group were given daily corticosterone injections (40 mg/kg) to induce stressed conditions, or NR and PC were orally administered for 21 days.
Figure 2.
Effects of NR and PC oligopeptides on liver IL-6 (A), TNF-α (B), IL-1β (C) and IL-8 (D) levels in rats subject to chronic corticosterone (CORT). The densitometric analysis of the relative intensity according to the control group of the western blotting bands was performed with β-actin normalization to ensure equal protein loading (E). Blots were repeated at least three times (n = 3), and a representative blot was shown. Data are expressed as a percent of the control set at 100%. The error bars above the lines indicate the standard deviation of the mean. Different symbols (a-f) indicate significant differences among the groups (ANOVA and Tukey post hoc test; P<0.05). IL-6, interleukin-6; TNF-α, tumor necrosis factor α; IL-1β, interleukin-1β; IL-8, interleukin-8. CS: chronic stress; NR: Nicotinamide riboside (26.44 mg/kg); PC-LD: Phycocyanin oligopeptide low dose (2.64 mg/kg); PC-HD: Phycocyanin oligopeptide high dose (26.44 mg/kg). Rats except the control group were given daily corticosterone injections (40 mg/kg) to induce stressed conditions, or NR and PC were orally administered for 21 days.
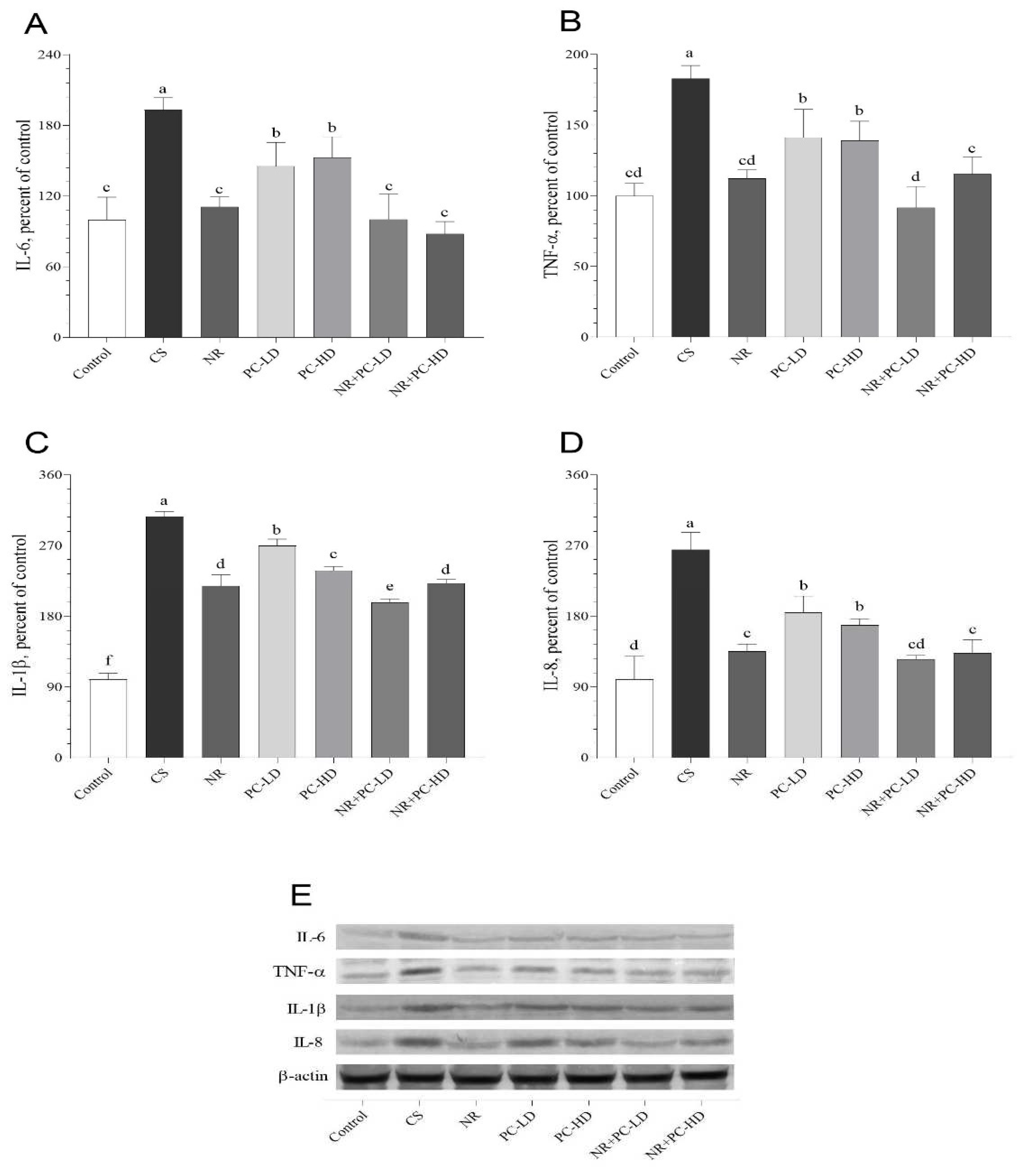
Figure 3.
Effects of NR and PC oligopeptides on liver POT1a (A), POT1b (B), TRF1 (C), TRF2 (D) and Tin2 (E) levels in rats subject to chronic corticosterone (CORT). The densitometric analysis of the relative intensity according to the control group of the western blotting bands was performed with β-actin normalization to ensure equal protein loading (F). Blots were repeated at least three times (n = 3), and a representative blot was shown. Data are expressed as a percent of the control set at 100%. The error bars above the lines indicate the standard deviation of the mean. Different symbols (a-f) indicate significant differences among the groups (ANOVA and Tukey post hoc test; P<0.05). POT1a, protection of telomeres protein 1a; POT1b, protection of telomeres protein 1b; TRF1, telomeric repeat-binding factor 1; TRF2, telomeric repeat-binding factor 1; Tin2, TRF1-interacting protein 2. CS: chronic stress; NR: Nicotinamide riboside (26.44 mg/kg); PC-LD: Phycocyanin oligopeptide low dose (2.64 mg/kg); PC-HD: Phycocyanin oligopeptide high dose (26.44 mg/kg). Rats except the control group were given daily corticosterone injections (40 mg/kg) to induce stressed conditions, or NR and PC were orally administered for 21 days.
Figure 3.
Effects of NR and PC oligopeptides on liver POT1a (A), POT1b (B), TRF1 (C), TRF2 (D) and Tin2 (E) levels in rats subject to chronic corticosterone (CORT). The densitometric analysis of the relative intensity according to the control group of the western blotting bands was performed with β-actin normalization to ensure equal protein loading (F). Blots were repeated at least three times (n = 3), and a representative blot was shown. Data are expressed as a percent of the control set at 100%. The error bars above the lines indicate the standard deviation of the mean. Different symbols (a-f) indicate significant differences among the groups (ANOVA and Tukey post hoc test; P<0.05). POT1a, protection of telomeres protein 1a; POT1b, protection of telomeres protein 1b; TRF1, telomeric repeat-binding factor 1; TRF2, telomeric repeat-binding factor 1; Tin2, TRF1-interacting protein 2. CS: chronic stress; NR: Nicotinamide riboside (26.44 mg/kg); PC-LD: Phycocyanin oligopeptide low dose (2.64 mg/kg); PC-HD: Phycocyanin oligopeptide high dose (26.44 mg/kg). Rats except the control group were given daily corticosterone injections (40 mg/kg) to induce stressed conditions, or NR and PC were orally administered for 21 days.
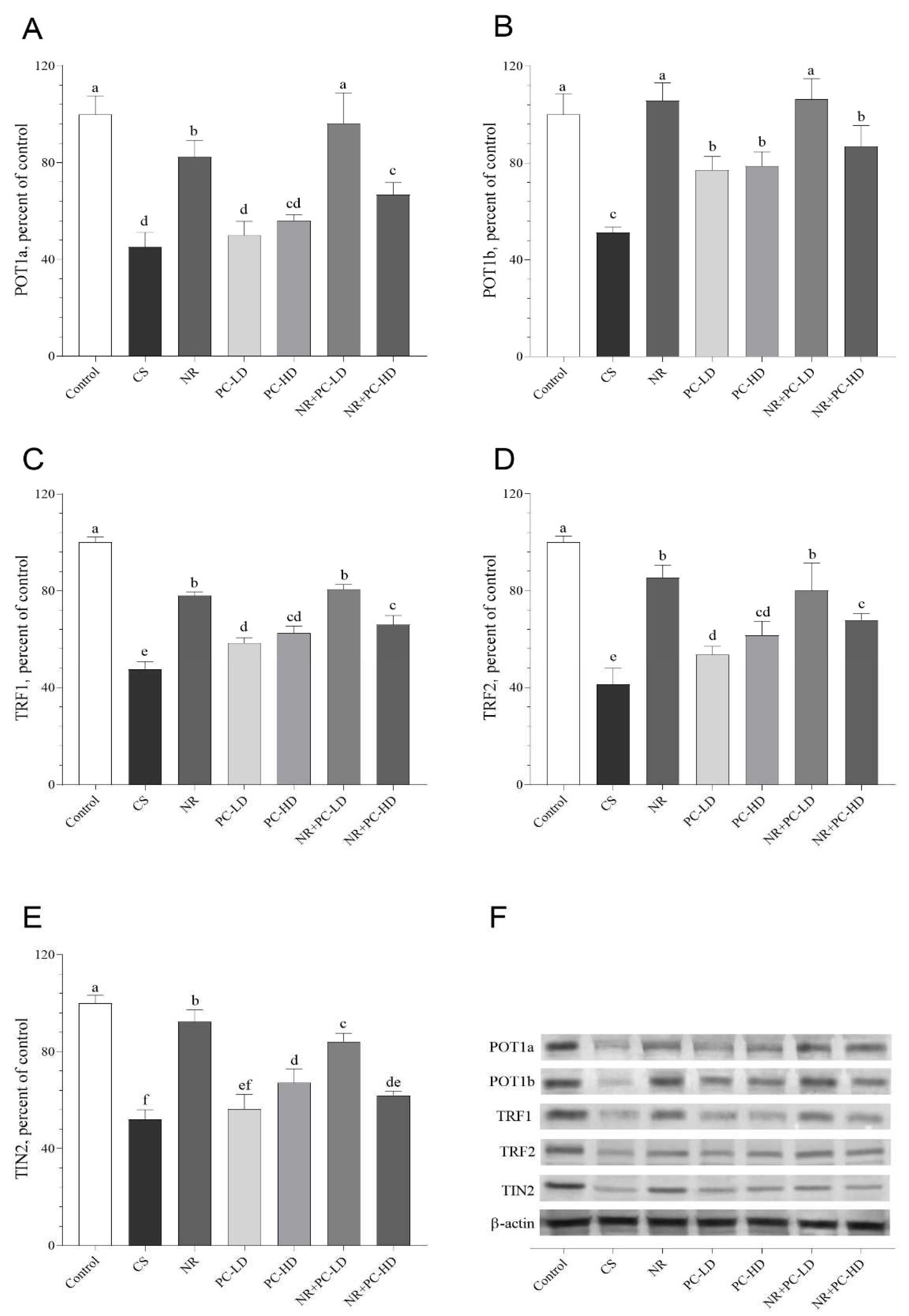
Figure 4.
Effects of NR and PC oligopeptides on liver SIRT1 (A), SIRT3 (B) and NAMPT (C) levels in rats subject to chronic corticosterone (CORT). The densitometric analysis of the relative intensity according to the control group of the western blotting bands was performed with β-actin normalization to ensure equal protein loading (D). Blots were repeated at least three times (n = 3), and a representative blot was shown. Data are expressed as a percent of the control set at 100%. The error bars above the lines indicate the standard deviation of the mean. Different symbols (a-e) indicate significant differences among the groups (ANOVA and Tukey post hoc test; P<0.05). SIRT1, sirtuin 1; SIRT3, sirtuin 3; NAMPT, nicotinamide phosphoribosyltransferase. CS: chronic stress; NR: Nicotinamide riboside (26.44 mg/kg); PC-LD: Phycocyanin oligopeptide low dose (2.64 mg/kg); PC-HD: Phycocyanin oligopeptide high dose (26.44 mg/kg). Rats except the control group were given daily corticosterone injections (40 mg/kg) to induce stressed conditions, or NR and PC were orally administered for 21 days.
Figure 4.
Effects of NR and PC oligopeptides on liver SIRT1 (A), SIRT3 (B) and NAMPT (C) levels in rats subject to chronic corticosterone (CORT). The densitometric analysis of the relative intensity according to the control group of the western blotting bands was performed with β-actin normalization to ensure equal protein loading (D). Blots were repeated at least three times (n = 3), and a representative blot was shown. Data are expressed as a percent of the control set at 100%. The error bars above the lines indicate the standard deviation of the mean. Different symbols (a-e) indicate significant differences among the groups (ANOVA and Tukey post hoc test; P<0.05). SIRT1, sirtuin 1; SIRT3, sirtuin 3; NAMPT, nicotinamide phosphoribosyltransferase. CS: chronic stress; NR: Nicotinamide riboside (26.44 mg/kg); PC-LD: Phycocyanin oligopeptide low dose (2.64 mg/kg); PC-HD: Phycocyanin oligopeptide high dose (26.44 mg/kg). Rats except the control group were given daily corticosterone injections (40 mg/kg) to induce stressed conditions, or NR and PC were orally administered for 21 days.
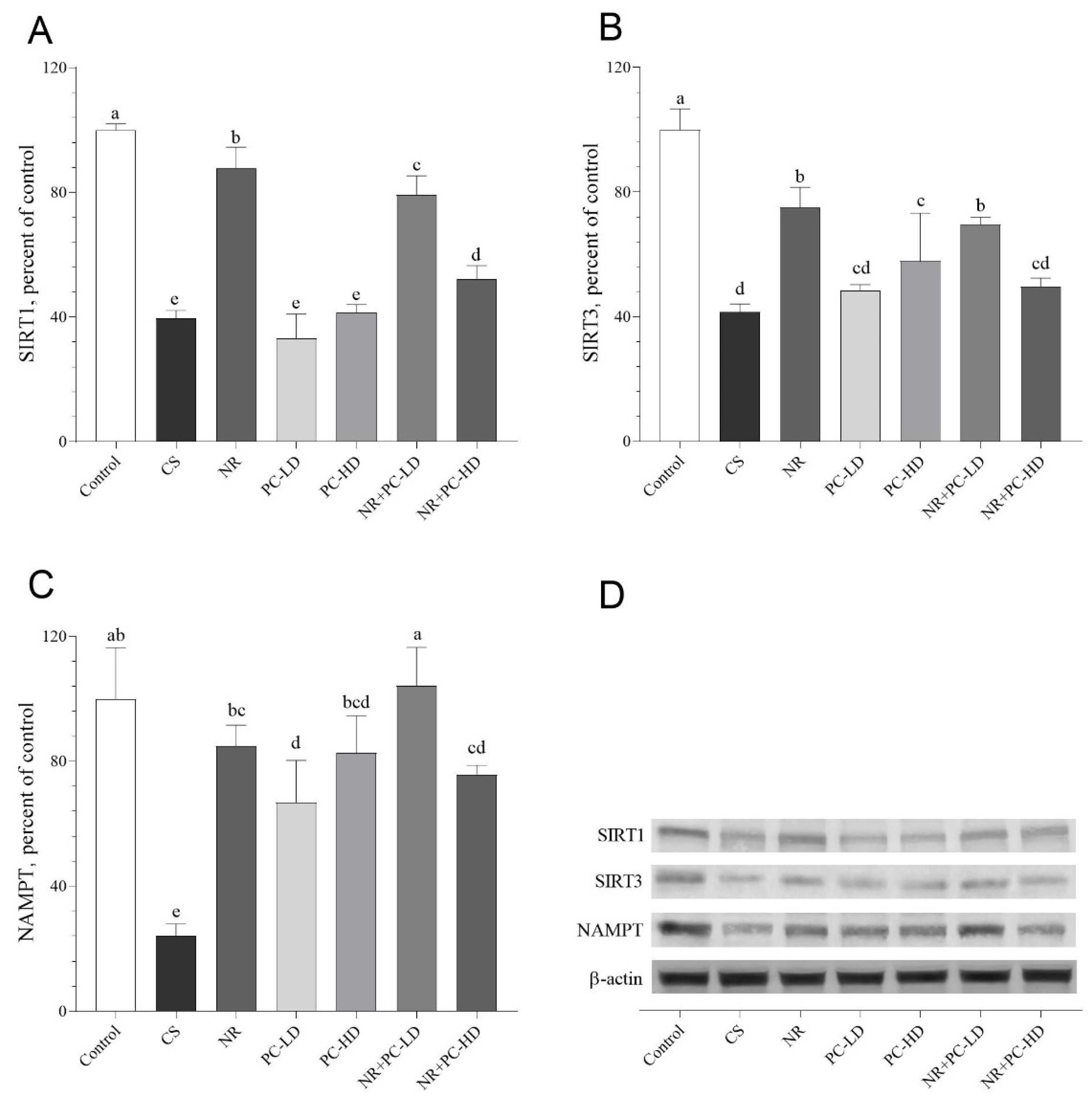

Table 1.
Effects of nicotinamide riboside and phycocyanin oligopeptides on serum biochemical paarmeter in rats subject to chronic corticosterone.
Table 1.
Effects of nicotinamide riboside and phycocyanin oligopeptides on serum biochemical paarmeter in rats subject to chronic corticosterone.
| Groups | |||||||
|---|---|---|---|---|---|---|---|
| Control | CS | NR | PC-LD | PC-HD | NR+PC-LD | NR+PC-HD | |
| Final BW, g | 276.57±9.85a | 210.43±8.28b | 222.86±23.11b | 221.29±11.25b | 221.86±9.92b | 225.14±13.23b | 217.86±19.42b |
| Glucose, mg/dL | 119.14±8.43ab | 127.86±5.05a | 119.86±7.10ab | 118.57±2.64abc | 114.14±5.24bc | 109.29±5.47c | 118.29±4.68bc |
| Creatine, mg/dL | 0.40±0.06 | 0.38±0.07 | 0.37±0.08 | 0.40±0.08 | 0.39±0.06 | 0.36±0.09 | 0.45±0.05 |
| BUN, mg/dL | 21.46±3.57 | 23.01±2.32 | 22.90±1.87 | 22.13±2.52 | 21.59±3.00 | 22.20±4.11 | 23.17±3.65 |
| ALT, U/L | 95.57±8.42 | 98.14±12.81 | 98.00±5.55 | 98.71±6.82 | 98.03±7.36 | 96.10±5.78 | 98.72±4.79 |
| AST, U/L | 110.69±11.79 | 117.46±7.53 | 114.71±10.92 | 112.60±13.13 | 117.43±15.25 | 114.06±12.59 | 116.20±19.06 |
Data are presented as the mean and standard deviation. a-c: Means in the same line without a common superscript differ significantly (P<0.05, ANOVA and Tukey post hoc test). CS: chronic stress induced with corticosterone; NR: Nicotinamide riboside (26.44 mg/kg); PC-LD: Phycocyanin oligopeptide low dose (2.64 mg/kg); PC-HD: Phycocyanin oligopeptide high dose (26.44 mg/kg). Rats except the control group were given daily corticosterone injections (40 mg/kg) to induce stressed conditions, or NR and PC were orally administered for 21 days. BUN: Blood urea nitrogene; ALT: Alanine transaminase; AST: Aspartate aminotransferase.
Table 2.
Effects of nicotinamide riboside and phycocyanin oligopeptides on serum biochemical paarmeter in rats subject to chronic corticosterone.
Table 2.
Effects of nicotinamide riboside and phycocyanin oligopeptides on serum biochemical paarmeter in rats subject to chronic corticosterone.
| Groups | |||||||
|---|---|---|---|---|---|---|---|
| Control | CS | NR | PC-LD | PC-HD | NR+PC-LD | NR+PC-HD | |
| Corticosterone, ng/ml | 46.33±5.82b | 110.22±6.15a | 102.16±8.68a | 104.48±3.67a | 105.91±6.39a | 105.46±8.13a | 104.99±5.34a |
| MDA, nmol/ml | 0.55±0.07f | 1.97±0.13a | 1.74±0.07b | 1.53±0.07c | 1.41±0.07cd | 1.26±0.07e | 1.28±0.09de |
| SOD, U/ml | 128.00±5.48a | 54.54±6.39e | 61.36±4.5e | 72.70±3.05d | 86.41±6.34c | 96.84±6.77b | 98.27±4.65b |
| GSH-Px, U/ml | 64.87±3.08a | 19.62±1.37e | 26.28±1.98d | 31.76±4.96cd | 37.10±2.80c | 44.68±5.58b | 45.98±4.10b |
| CAT, U/ml | 164.98±6.13a | 102.03±4.80e | 114.69±4.82d | 124.55±5.48c | 131.43±3.76bc | 140.55±6.76b | 139.05±7.81b |
Data are presented as the mean and standard deviation. a-f: Means in the same line without a common superscript differ significantly (P<0.05, ANOVA and Tukey post hoc test). CS: chronic stress induced with corticosterone; NR: Nicotinamide riboside (26.44 mg/kg); PC-LD: Phycocyanin oligopeptide low dose (2.64 mg/kg); PC-HD: Phycocyanin oligopeptide high dose (26.44 mg/kg). Rats except the control group were given daily corticosterone injections (40 mg/kg) to induce stressed condition or NR and PC were orally administered for 21 days. MDA: Malondialdehyde; SOD: Superoxide dismutase; GSH-Px: Glutathione peroxidase; CAT: Catalase.
Table 3.
Effects of nicotinamide riboside and phycocyanin oligopeptides on serum biochemical paarmeter in rats subject to chronic corticosterone.
Table 3.
Effects of nicotinamide riboside and phycocyanin oligopeptides on serum biochemical paarmeter in rats subject to chronic corticosterone.
| Groups | |||||||
|---|---|---|---|---|---|---|---|
| Control | CS | NR | PC-LD | PC-HD | NR+PC-LD | NR+PC-HD | |
| NAD+, µmol/g | 0.76±0.05a | 0.33±0.04f | 0.59±0.04c | 0.42±0.05e | 0.49±0.06de | 0.67±0.05b | 0.55±0.06cd |
| NAM, µmol/g | 2.36±0.15a | 1.01±0.08e | 1.60±0.06c | 1.31±0.09d | 1.43±0.13d | 1.78±0.10b | 1.66±0.10cb |
| NA, µmol/g | 1.79±0.09a | 1.22±0.12d | 1.63±0.09abc | 1.46±0.10c | 1.57±0.17bc | 1.71±0.13ab | 1.73±0.12ab |
| NMN, µmol/g | 0.45±0.05a | 0.15±0.02e | 0.30±0.02bc | 0.20±0.02de | 0.24±0.03cd | 0.36±0.05b | 0.32±0.07b |
| NADPH, nmol/g | 62.65±4.13a | 30.84±3.52e | 44.42±2.31c | 39.88±3.04cd | 41.38±3.57cd | 52.52±5.39b | 37.89±2.23d |
Data are presented as the mean and standard deviation. a-f: Means in the same line without a common superscript differ significantly (P<0.05, ANOVA and Tukey post hoc test). CS: chronic stress induced with corticosterone; NR: Nicotinamide riboside (26.44 mg/kg); PC-LD: Phycocyanin oligopeptide low dose (2.64 mg/kg); PC-HD: Phycocyanin oligopeptide high dose (26.44 mg/kg). Rats except the control group were given daily corticosterone injections (40 mg/kg) to induce stressed condition or NR and PC were orally administered for 21 days. Nicotinamide adenine dinucleotide; NAD+: Nicotinamide adenine dinucleotide; NAM: Nicotinamide; NA: Nicotinic acid; NMN:Nicotinamide mononucleotide; NADPH: nicotinamide adenine dinucleotide phosphate; GSH: glutathione.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



