Submitted:
08 November 2025
Posted:
10 November 2025
You are already at the latest version
Abstract
How does the complex, adaptive, and autonomous organization of life emerge from the laws of physics and information? This review argues that the answer lies in a convergent set of universal organizational principles that constitute a physical and informational grammar of the living. Living sys-tems are dissipative structures that achieve organizational clo-sure—materially and energetically open, yet causally closed—thereby at-taining genuine autonomy and agency. Their architecture exhibits fractal and modular scaling laws that maximize energy flow, robustness, and evolv- ability under universal physical constraints. Critically, organisms operate at critical transitions—zones of controlled instability where fluctuations amplify information processing, transforming noise into adaptive signal. This self-organized criticality enables functional degeneracy, relational redun-dancy, and evolutionary antifragility. Cognition emerges as distributed ac-tive inference—a predictive-corrective cycle that integrates perception, ac-tion, and learning in accordance with the Free Energy Prin- ciple. From molecular networks to ecosystems, the same physico-informational gram- mars unfold recursively, revealing a deep organizational holography: the principles of organization are replicated across scales. Evolution under the Law of Increasing Functional Information is not random drift, but a direc-tional expansion of functional complexity—a thermodynamic gradient to-wards greater agency. This synthesis chal- lenges biological exceptionalism: the trajectory from thermodynamics to cognition is continuous, physically constrained, and potentially inevitable. Life does not violate physical laws—it fulfills them in regimes of high informational complexity, instan- tiating fundamental principles in self-organized architectures capable of prediction, memory, and purpose.
Keywords:
physics of life
; complex systems
; self-organization
; emergence
; information theory
; biological organization
; criticality
Introduction
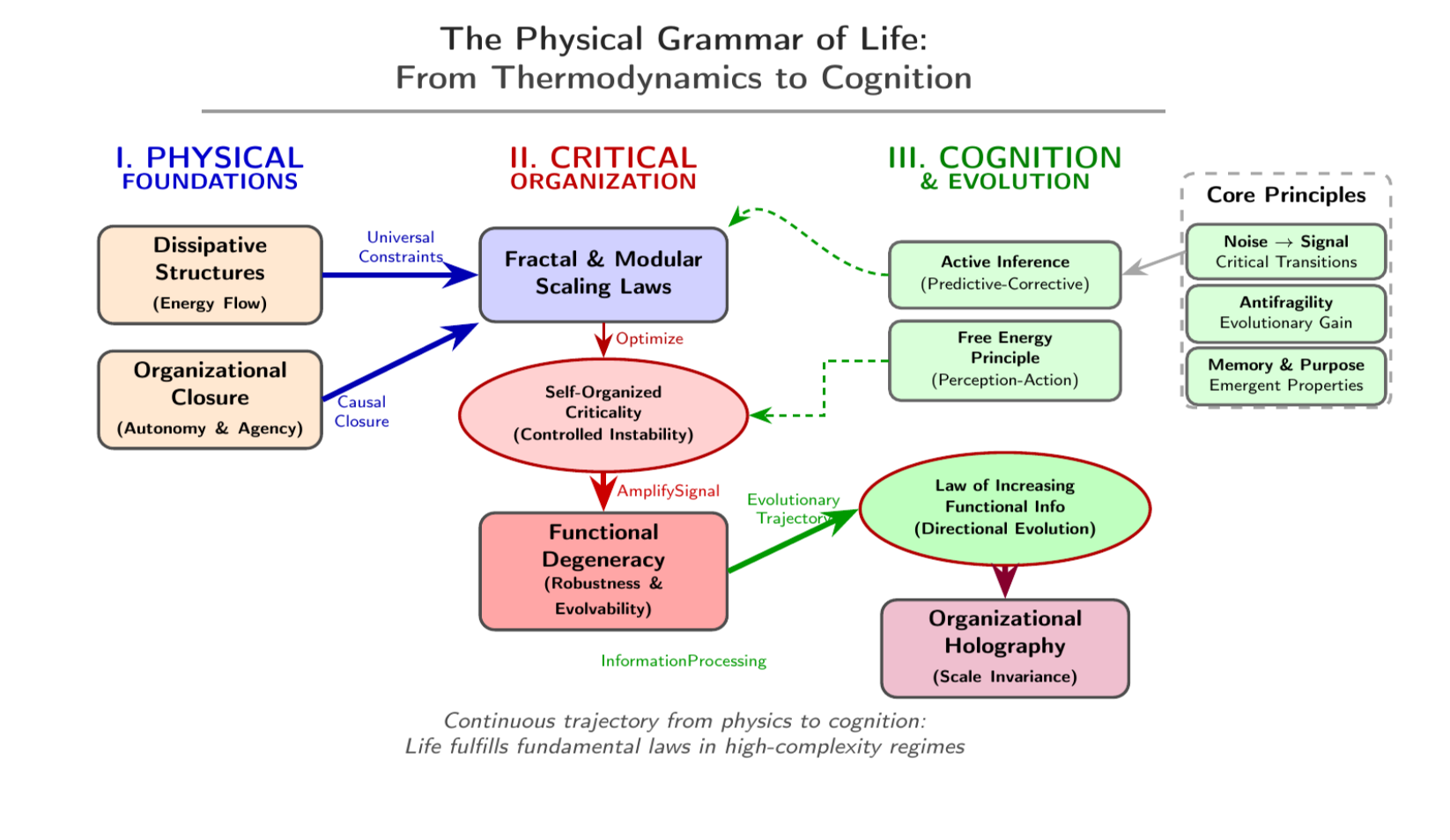
The Essence of Universal Principles of Biological Organization Life is distin- guished from inert matter by its capacity to maintain organization, adaptability, and autonomy in an environment subject to fluctuations and increasing entropy. This organization is not arbitrary; it responds to universal principles that govern the emergence, stability, and evolution of complex systems far from equilibrium. Under- standing these principles is the central goal of a physics of life, in which biology is interpreted as a particular manifestation of the laws governing the self-organization of matter, energy, and information [1,2]. Unlike closed systems that tend toward disorder (increasing entropy), organisms are open entities that constantly exchange energy and matter with their surroundings. This exchange allows them to export entropy and maintain an internal state of high organization and low entropy, an in- dispensable condition for life (dissipative structures) [3,4,5]. The continuous flow of energy not only sustains this order but also drives the system far from thermody- namic equilibrium, where non-linear dynamics arise. Under these critical conditions, small internal fluctuations can be amplified, generating instabilities that culminate in the spontaneous breaking of symmetries. This process is the engine that orig- inates complex patterns and new levels of spatiotemporal organization, a funda- mental mechanism explained by the theories of Prigogine and Turing [6]. Consequently, biological stability should not be understood as a static homeostasis (maintenance of constancy), but as a homeodynamics: a dynamic equilibrium that allows the persistence of organization through constant change. Life, therefore, is not
maintained despite disequilibrium, but because of it [4]. However, the key distin- guishing feature of the living does not reside solely in its dissipative structure, but in its organizational closure: the system’s capacity to actively produce and maintain its own components [6,7,8]. This principle, formalized by Maturana and Varela’s autopoiesis, Rosen’s (M,R) systems, and Kauffman’s autocatalytic sets, defines bi- ological autonomy. A system that is causally closed but materially open is capable of sustaining its identity and purpose, generating an internal normativity: action oriented toward its own viability [8]. Biological organization is further expressed in fractal and modular architectures, where self-similarity and hierarchy optimize energy and information exchanges[9]. Models such as that of West, Brown, and Enquist (WBE) demonstrate that fractality constitutes an optimal physical solution for transport ef- ficiency and functional robustness [10,11]. Modularity, in turn, favors evolvability and the capacity for structural adaptation to perturbations [10]. At a dynamic level, living systems operate near self-organized criticality—the “edge of chaos”—a regime that maximizes information processing and flexible response capacity in changing environments [8,9,10,11,12,13,14]. This balance between order and fluctuation is a universal property of biological complex systems: from neural networks to ecosystems, criticality confers adaptability, memory, and functional creativity [15,16,17,18]. Adaptive behavior is sustained by functional degeneracy—the existence of multiple structural configurations capable of performing the same function—and by antifragility, the ability to improve in the face of perturbations [18,19] . These properties make life a system that learns from disorder and transforms variability into a source of innovation[20]. Evolution can be understood as a process directed by physical-informational constraints. The Law of Increasing Functional Information (LIFC) posits that evolutionary systems tend toward an increase in functional information—the capacity to sustain and execute complex functions—which introduces a natural directionality into biological evolution [20]. This review addresses a fundamental question: how does the complex, adaptive, and autonomous organization of life emerge from the laws of physics and information? We argue that the answer lies not in the molecular components, but in a convergent set of organizational principles. These principles—dissipation, organizational closure, fractality, modularity, criticality, degeneracy, antifragility, and increasing functional information—constitute a universal grammar of life. Through them, living matter is revealed as a physical-informational system capable of generating order, meaning, and cognition from the continuous flow of energy and data. The objective of this work is, therefore, to articulate how the synthesis of these principles not only unifies physics and biology but also illuminates the profound continuity between thermodynamics, organization, and mind.
Results
State of the Art and Main Formal Models
Understanding the universal organizational principles that sustain life requires integrating three conceptual domains: non-equilibrium thermodynamics, the theory of self-organization, and the logic of organizational closure. Together, these approaches explain how living systems emerge, maintain themselves, and evolve as coherent structures far from equilibrium, transforming energy and information to sustain their functional identity [21]. Specifically, the thermodynamic domain by stating that the second law of thermodynamics, acting as a fundamental constraint, drives the emergence of structure in dissipative systems. This driving force is mani- fested that processes emerge to reduce the applied gradient, a phenomenon which, if dynamic conditions permit, is recognized as self-organization. Consequently, the expected outcomes of this thermodynamic mandate: as biosystems grow and develop, they must increase their total dissipation, leading to greater complexity, energy flow, diversity, and hierarchical levels[21]. Furthermore, this thermodynamic imperative provides the critical lens for interpreting biological success and failure, which di- rectly supports the conceptual domain of organizational closure mentioned in the paragraph. The summary’s corollary—that organisms which do not increase total dissipation are essentially organisms “dedicated to death” (like during aging)—and its observation that surviving species are those that funnel energy into reproduction and contribute to autocatalytic processes that increase the ecosystem’s total dissipation, provides the practical, observable evidence for a system maintaining its “functional identity” far from equilibrium. In essence, the opening paragraph lays out the required intellectual toolkit, while the summary translates that toolkit into a set of universal organizational principles that govern the growth, survival, aging, and complexity of all biosystems[21].
Thermodynamic Foundation: Dissipation, Instability, and Organization
Living organisms are thermodynamically open systems that maintain their state of low entropy (high order) thanks to a continuous flow of energy and matter, ex- porting disorder to the environment. This principle, already stated by Schrödinger , constitutes the physical basis of life as a process of maintained negentropy [22].
The theory of dissipative structures developed by Ilya Prigogine and collaborators demonstrated that, far from equilibrium, non-linearity in system dynamics allows for the amplification of small fluctuations under conditions of energetic forcing, leading to the emergence of new spatiotemporal organizational states. This process can be represented as a fundamental causal sequence [23]:
energy gradient → dissipation → instability → symmetry breaking → emergent order
This mechanism not only explains self-organization in physical systems but also biological morphogenesis, where Turing patterns emerge as a consequence of diffusive-reactional instabilities that break spatial symmetries and give rise to ordered structures.
From this perspective, biological stability does not correspond to a static homeostasis, but to a homeodynamics, in which equilibrium is redefined as a continuous oscillation around dynamic attractors. Thus, life is interpreted as a form of organized matter that is maintained “on the edge of instability,” using energy dissipation not as a cost, but as the engine of its own organization [24].
Non-equilibrium thermodynamics therefore provides the physical framework that explains how living systems can maintain order through the flow of entropy. Self- organization emerges as a natural consequence of the critical conditions imposed by energy gradients, establishing the material bases for the emergence of functional hierarchies, metabolic networks, and biological rhythms (Figure 1).
Logic of Self-Fabrication: Organizational Closure and Autonomy
What distinguishes living systems from other dissipative structures is not just their ability to sustain order, but their organizational closure : the property of actively producing, repairing, and maintaining the components that constitute them. This principle defines biological autonomy, understood as a system’s ability to regenerate the processes that make its own existence possible[25].
Three convergent formalisms have modeled this property with remarkable theoretical consistency:
(a) Autopoiesis (Maturana and Varela): Living beings are networks of production processes that continuously regenerate the components that constitute the network itself and its boundary, thereby defining an autonomous operational identity. This approach emphasizes that organization, not material composition, is what distinguishes the living[12,26].
(b) (M,R) Systems (Rosen): Life is characterized as a system “closed to efficient causality,” where each functional component is, in turn, a product of another component of the system. This causal closure implies an essential noncomputability, which differentiates organisms from machines, as an organism cannot be fully described or predicted from an external algorithmic description.
(c) Autocatalytic Sets (Kauffman): Beyond a critical threshold of complexity, networks of chemical reactions can become catalytically closed and self- sustaining. This model offers a plausible formalization for the emergence of coherent metabolism and the initial molecular self-organization of life[27].
In all these frameworks, biological agency the ability to act with purpose emerges as a thermodynamic consequence of organizational closure. A system that must maintain itself under conditions of viability necessarily acts to preserve its internal structure, generating behaviors oriented towards its own persistence[28].
Organizational closure, therefore, explains not only autonomy but also the teleonomy inherent in living systems: their tendency to preserve identity through processes of self-fabrication and self-regulation. From a physical-informational viewpoint, these systems can be conceived as inferential entities that process information to maintain their internal coherence against external entropy, which anticipates the later articulation with the principles of active inference and free energy (Figure 2).
Universal Organizational Principles
Life can be understood as a particular expression of organizational principles that govern matter far from equilibrium. These principles autopoiesis, fractality, modularity, criticality, distributed cognition, antifragility, and the increase of functional information act as organizational invariants that cut across biological scales, from molecular networks to cognitive systems and ecosystems[29]. Together, they constitute a physical and informational grammar of the living, where structure and dynamics are inseparable
Autopoiesis, (M,R) Systems, and Autocatalytic Sets
system actively maintains its identity through the production and repair of its own components. In the autopoiesis of Maturana and Varela, living beings are conceived as networks of production processes that regenerate the network itself and its boundary, establishing an internal operational identity [7]. Rosen formalized this idea in (M,R) systems, showing that organisms are “closed to efficient causality,” meaning that each function within the system is a product of another internal function, generating a causal circularity not reducible to external mechanisms [8]. Kauffman, for his part, demonstrated that when chemical reaction networks reach a critical density, closed and self-sustaining autocatalytic sets emerge, capable of maintaining a coherent metabolism [6]. These models converge on a fundamental thesis: life is self-referential, a system that produces the conditions of its own persistence. Causal closure implies internal normativity: the system acts to keep itself within a domain of viable states, giving rise to an immanent form of biological purpose.
Fractality and Modularity
Biological organization is not random: it follows hierarchical, modular, and fractal architectures that reflect universal principles of optimization under physical constraints. Modularity implies that systems are composed of subsystems that are densely interconnected internally but weakly coupled to each other. This pattern confers local robustness against perturbations and global evolvability, by allowing the functional recombination of “building blocks” [9]. Fractality, on the other hand, reflects structural self-similarity across scales. It constitutes an optimal solution for maximizing exchange surface area within a limited volume, as occurs in lungs, vascular systems, or neural networks [13]. The West, Brown, and Enquist (WBE) model formalized this principle, showing that allometric laws—such as the metabolic rate M*)—emerge from the fractal optimization of energy flow [10]. Modularity and fractality, together, define the universal architecture of living networks: hierarchical structures that balance efficiency, resilience, and the capacity for innovation.
Criticality
Numerous biological systems—from gene regulation to cortical activity—operate near a critical point, at the “edge of chaos.” Self-organized criticality (SOC) posits that complex systems can spontaneously evolve toward this intermediate state, where stability and variability coexist[10,11]. In this regime, life exploits the statistical properties of phase transitions to optimize its performance:
▪ It maximizes the dynamic range and sensitivity to stimuli.
▪ It optimizes information processing and transmission.
▪ It maintains a balance between robustness and adaptive flexibility.
Evidence of critical dynamics has been observed in neural avalanches that follow power laws, in self-regulated gene networks, and in ecosystems showing scaling of population fluctuations [14,30]. Criticality, in this context, is not an evolutionary accident but a boundary condition where information and energy are optimally coupled, allowing the system to respond creatively to perturbations without losing coherence.
Distributed Cognition
The functional coherence of living systems emerges from distributed self-organization rather than centralized control. This principle is manifest from insect colonies and immune systems to the brain. Models like Kuramoto’s describe the spontaneous synchronization of coupled oscillators, a universal mechanism by which systems achieve temporal coherence from local interactions [15]. On a more abstract level, Friston’s Free Energy Principle (FEP) formalizes cognition as active inference: living systems maintain an internal model of the world and act to minimize “surprise” or prediction error, unifying perception, action, and learning under a Bayesian principle of self-evidence [16]. Thus, cognition ceases to be an exclusive attribute of the brain and is redefined as an extended and multi-scalar property: the continuous process by which life models, anticipates, and regulates its states in an uncertain environment.
Antifragility and Degeneracy
Living systems do not just resist disorder: they benefit from it. Antifragility, as stated by Taleb [17,18], describes systems whose structure improves in the face of perturbations, volatility, or stress. Its biological foundation lies in functional degeneracy—the existence of multiple structurally different components capable of fulfilling the same function—[18]. This diverse redundancy allows for the absorption of variations without loss of performance and enables evolutionary innovation: new functions can emerge without sacrificing existing ones. Degeneracy, therefore, constitutes the structural substrate of antifragility, while criticality acts as its dynamic condition. Together, these properties make life a robust, evolutionary, and creative system that transforms uncertainty into a source of adaptation.
Law of Increasing Functional Information (LIFC)
Biological evolution, far from being a random drift, shows directional trends in the accumulation of organized complexity. The Law of Increasing Functional Information (LIFC) proposes that functional information—understood as a system’s ability to maintain its integrity and execute functions—tends to increase in open systems subjected to selection for function [31]. This law is analogous to the Second Law of Thermodynamics but oriented toward information: while entropy measures energy disorder, functional information measures operational order [29,32]. Throughout evolution, systems tend to explore configurations that increase their ability to process energy and information more efficiently. The LIFC offers a quantitative formulation of evolutionary directionality: life progresses toward states of greater integration, coordination, and informational processing, which can be observed in the expansion of metabolic networks, cellular specialization, and the emergence of cognition[19].
Conceptual Synthesis and Structural Analogies: The Holographic View
The principles described—dissipation, organizational closure, fractality, modularity, criticality, degeneracy, antifragility, and increasing functional information—do not act in isolation. They manifest in a self-similar and recursive manner at all levels of biological organization, from macromolecules to the biosphere. This holographic convergence suggests that each level of organization reflects the logic of the others: the same patterns of self-organization, distributed control, and energy optimization are repeated, adjusting to the specific constraints of each scale.
The Immune System: Autonomy, Fractality, and Distributed Cognition
The immune system is a paradigmatic expression of this holographic view. It is, first and foremost, a dissipative structure that maintains the organizational closure of the “self” through the continuous production of cells, receptors, and signals that sustain immunological identity[33]. Its fractal and modular architecture—from lymph nodes to antigenic recognition microdomains—optimizes the transport of information and cells throughout the body, maximizing efficiency under energy constraints. The immune system operates near a dynamic criticality, where small stimuli can be amplified into coordinated responses without loss of stability, analogous to the “neural avalanches” observed in the brain [34]. This condition allows it to respond to an unpredictable universe of pathogens without collapsing into systemic inflammation. Its antifragility is manifested in immunological memory: exposure to perturbations (pathogens or antigens) strengthens the network, expanding its functional repertoire. Immunological degeneracy—multiple clones, pathways, and molecular redundancies that converge on equivalent functions—confers resilience against mutations or antigenic evasion [35]. Finally, the immune system can be understood as a distributed cognitive system: it learns, remembers, and makes decisions without a control center, operating under a principle of active inference that minimizes surprise in the face of the pathogenic environment, as formalized by the Free Energy Principle [16]. Its dynamics thus conform to the same principles that govern the brain, but in a different topology: that of the extended body.
The Brain: Criticality, Modularity, and Informational Efficiency
The brain is another instance where holographic self-organization becomes evident. Neural networks exhibit hierarchical modularity and fractal geometry, optimizing signal transmission under metabolic constraints, analogously to vascular or lymphatic networks [36]. Experimental evidence indicates that cortical activity operates near a critical point, where neural connectivity oscillates between order and chaos, maximizing computational capacity and adaptive plasticity [37]. The brain maintains functional closure in its dynamics of synaptic self-fabrication, generating and reinforcing connections that preserve the internal coherence of the system, a neurobiological form of the principle of autopoiesis. In terms of the Free Energy Principle [16], the brain acts as an active inference system that minimizes the discrepancy between its internal predictions and sensory reality, i.e., an information processor that maintains its viability by reducing uncertainty.
The Microbiome: Critical Ecological Network and Co-autonomy
The microbiome represents an intermediate scale between the cellular and the ecological, where the principles of modularity, degeneracy, and antifragility are clearlyexpressed. The microbial community constitutes a distributed and redundant metabolic network: different taxa can perform similar functions (functional degeneracy), ensuring stability in the face of environmental changes[38]. These networks exhibit critical properties, adapting rapidly to perturbations through phase transitions that reorganize the relative abundance of species without collapsing the ecosystem’s functionality. Their ecological fractality is reflected in the self-similarity of connectivity and co-occurrence patterns at different scales—from intestinal microhabitats to global communities—a trait shared with intracellular metabolic networks[39]. The microbiome, moreover, participates in the extended cognition of the organism: it modulates behavior, the immune system, and neuroendocrine homeodynamics. In this sense, the biological individual cannot be separated from its symbiotic ecosystem: autonomy is always co-autonomy[40].
Tissues: Self-organization and Morphodynamic Patterns
In multicellular tissues, the principles of self-organization are expressed in morphogenesis and tissue homeodynamics. Turing models [6] and Prigogine’s dissipative structures [41] explain how chemical gradients and local fluctuations are amplified to generate self-organized spatial patterns (e.g., skin pigmentation, glandular architecture). Cellular interactions exhibit critical feedbacks that maintain the balance between proliferation and differentiation, and the fractal geometry of the vascular system optimizes nutrient diffusion. As a whole, tissues operate as multi-scalar systems, where cellular organization reflects—in a reduced version—the dynamics of the entire organism[42].
Ecosystems and Biosphere: Critical Flow Networks and Resilience
At the macroscopic level, ecosystems also show the signature of universal organizational principles. Trophic networks and energy transport networks are modular and fractal, which allows for local stability and global flexibility in the face of perturbations. Population fluctuations follow power laws, indicative of self-organized critical dynamics, where ecosystems are maintained in dynamic equilibrium at the edge of instability [43]. Ecological antifragility is observed in adaptive succession: environmental perturbations not only select but also induce functional reorganization, increasing the informational complexity of the system. Thus, the entire biosphere can be understood as an autopoietic meta-system that maintains its organization through dissipative cycles, energy transfers, and informational couplings at multiple scales[44].
Organizational Holography: A Universal Grammar of Life At All Scales, Life Obeys a Universal Organizational Grammar
▪ Dissipation maintains the distance from equilibrium.
▪ Organizational closure ensures autonomy.
▪ Fractality and modularity optimize structural efficiency.
▪ Criticality maximizes adaptability and information processing.
▪ Degeneracy and antifragility confer evolutionary robustness.
▪ Increasing functional information orients evolution toward greater degrees of integration.
This self-similar repetition of principles generates a holographic view of life, in which each level—molecular, cellular, organic, cognitive, and ecological—reflects the totality of the system. Biological organization is, thus, a fractal manifestation of the laws of informationally active matter: a continuity between thermodynamics, information, and cognition.
Falsifiable Predictions, Controversies, Limits, and Experimental Projections
Although the framework of a physics of life offers a unifying theoretical synthesis, its validity depends on its ability to generate testable predictions and to confront its own conceptual limits. The strength of the approach lies in its falsifiability: the possibility of subjecting its universal organizational principles—criticality, organizational closure, and the Law of Increasing Functional Information (LIFC)—to rigorous empirical tests that can distinguish between real universality and superficial analogy[45].
Controversies and Limits
Despite its integrative power, this framework faces three main objections:
(a) Context Dependency and Historical Contingency. The “small world” approach and the search for universal laws have been criticized for ignoring the irreducible specificity of biological contexts. Evolution does not always converge to optimal structures; it is shaped by historical contingencies, ecological constraints, and stochastic drift. Thus, the universality of evolutionary criticality or the directionality of the LIFC may be modulated by historical factors that break the idealization of global invariance [46].
(b) Incompleteness and Partial Unobservability. Concepts such as “elastic states” and “irreparable incompleteness” [20] suggest that it is not possible to derive universal axioms without incorporating the perspective of the system itself that enunciates them. Living systems possess degrees of self-reference—for example, cognition that models its own model—that hinder complete formalization. This imposes an intrinsic epistemological limit: the observer cannot be completely outside the system being studied.
(c) Ambiguity in the Definitions of Function and Information. Although the LIFC offers an attractive conceptual directionality, the objective measurement of functional information remains an open challenge. There are multiple metrics (effective information, Kolmogorov complexity, predictive capacity, structural redundancy), none of which fully captures the relationship between information and biological function. This methodological indefiniteness currently limits the predictive capacity of the law[47].
Falsifiable Predictions
(1) Fractal Scaling (WBE model). Metabolic rates and network geometries in extremophilic organisms, or in microbial ecosystems under high pressure, should conform to the scaling exponents predicted by the WBE model (body mass3/4). Systematic deviations under conditions of energy boundaries—e.g., organisms living in extremely low energy conditions or with non-branching cellular architectures— would refute the universality of the model [13].
(2) Neuronal and Genetic Criticality. Cultured neural networks (brain organoids) or in vitro gene regulation systems should exhibit distributions of “avalanches” or expression fluctuations that follow power laws—a signature of self-organized criticality [10,14]. Genetic manipulations that alter connectivity (e.g., overexpression of synaptic channels or suppression of coupling proteins) should shift the system to subcritical or supercritical regimes, reducing information processing capacity and functional plasticity. This would offer a direct falsifiable test of the principle of functional criticality[48].
(3) Law of Increasing Functional Information (LIFC). Comparative paleogenomics allows testing the LIFC hypothesis. If biological evolution follows a trend toward greater functional information, ancient genomes should show a systematic increase in effective information density, measurable by Kolmogorov complexity or integrated information in conserved genes [19]. A lack of net increase, or the presence of long-term reversible oscillations, would refute the directionality of the LIFC as a universal principle.
(4) Organizational Closure and Self-Fabrication. Synthetic biology offers an experimental avenue to evaluate organizational closure[49]. Protocellular systems designed with partially autocatalytic metabolic networks should show abrupt transitions to self-sustainment once a connectivity threshold is reached. If no such transition is observed—or if organizational closure requires external control—the universal applicability of the principle of autopoiesis and (M,R) systems would be called into question[50].
Conceptual Integration and Interdisciplinary Applications
The integrative framework of the universal organizational principles of biological systems transcends theoretical explanation to offer an operational language that connects physics, biology, and engineering. These principles—dissipation, organizational closure, fractality, modularity, criticality, degeneracy, antifragility, and increasing functional information—not only describe life[51]: they provide criteria for diagnosing, modeling, and designing living and artificial systems. Their value lies in the ability to translate theory into prediction and prediction into intervention.
Precision Medicine and Dynamic Pathophysiology
From this perspective, disease can be defined as a pathological phase transition: a deviation from the critical dynamic regime that characterizes healthy homeodynamics. Normal biological systems operate near the “edge of chaos,” where information processing is maximal and variability is functional; when the system shifts toward rigidity or disorder, pathological states emerge. Cancer can be reinterpreted as a breach of organizational closure: cells lose integration with the organism’s normative domain and acquire metabolic autonomy, breaking the system’s coherence [8]. Epilepsy represents a case of subcritical hypersynchronization: a loss of neural criticality that suppresses the dynamic diversity necessary for cognitive function [30]. Autoimmune diseases arise from a topological rupture of the self’s boundary, where the immune system confuses its own internal space with the environment. From this framework, precision medicine is redefined as a physics of living network control: therapies do not seek merely to inhibit molecules, but to restore critical dynamics and organizational closure. Therapeutic intervention becomes an engineering of functional phase, aimed at redirecting the system’s dynamics toward viable attractors, assessable through metrics of entropy, criticality, and effective information[52].
Bioengineering, Synthetic Biology, and the Design of Life
Contemporary bioengineering faces the challenge of designing systems that not only function, but self-organize. The universal principles of biological organization provide a framework for this: Hierarchical modularity and functional degeneracy offer design criteria for building robust and adaptive biological circuits, capable of maintaining functionality in the face of perturbations [9]. Fractal architecture inspires the design of tissues and microstructures that optimize transport, diffusion, and signaling, mimicking the geometries of vascular, neural, or lymphatic networks [10]. Synthetic autocatalytic and autopoietic systems offer a platform to explore the transition between complex chemistry and autonomous metabolism, experimentally testing the limits of organizational closure [12]. The synthetic biology of the future will be predictive if it adopts a thermodynamic and informational paradigm, designing organisms or ecosystems not by trial and error, but by following principles of dynamic stability, energetic coupling, and maximization of functional information.
Artificial Intelligence, Cognition, and Complex Systems
In the domain of cognitive sciences and artificial intelligence, the organizational principles of life offer a radically different model from the classic computational approach. Instead of networks trained on external data, life operates as a self-generating system of information, which actively learns by maintaining its internal coherence. The Free Energy Principle (FEP) [16] provides a unifying framework between biology and cognition: neurons, cells, and organisms all minimize surprise (prediction error) through active inference. From this perspective: Biological cognition is redefined as a physical process of informational self-organization; Biologically inspired artificial intelligence must be antifragile, that is, improve with perturbation and learn not only from data, but from errors and fluctuations; Distributed AI systems—based on decentralized control, adaptive synchronization, and structural plasticity—represent the practical convergence between autopoiesis, criticality, and distributed cognition. This framework suggests a transition from “artificial intelligence” to organizational intelligence, where algorithms do not replicate the human mind, but the universal principles of life’s adaptation[53].
Towards a Technobiological Synthesis
The confluence of these fields points toward a new discipline: a predictive technobiology, where the physical principles of living organization guide both the understanding of biology and the creation of artificial systems that share its essential properties: autonomy, plasticity, degeneracy, and antifragility. In this context, the Physics of Life is not a return to mechanism, but the expansion of physics into domains where information acquires causal power. Its interdisciplinary application does not seek to control life, but to learn from its organizational grammar to design systems—medical, technological, or ecological—that share its capacity to persist, learn, and evolve[54].
Discussion
Critical Conclusions and Prospects for a Physics of Life
This review proposes that life is not an exception to physical laws, but a manifestation of them under conditions of flow, instability, and organizational closure. In this sense, the Physics of Life represents the most coherent contemporary attempt to formulate a unified framework that explains the emergence, persistence, and evolution of biological organization through general principles. From this effort, three fundamental theses emerge: Life as a Dissipative Structure with Organizational Closure. Organisms are materially open systems that maintain their identity through closed causal loops of production and regulation. Their teleonomy—the apparent purpose of living—is not a metaphysical attribute, but a physical consequence of the causal closure between metabolism, information, and boundary. Adaptation as a Critical and Antifragile Homeodynamics. Living systems self-regulate in a dynamic regime close to criticality, where stability and plasticity coexist. Functional degeneracy—multiple pathways for the same function—allows them to absorb perturbations and improve under stress, making antifragility a constitutive property of living. Cognition as Distributed Active Inference. Cognition emerges as a physical process of uncertainty minimization, extending from molecules to societies. It does not require a central controller: coherence arises from the synchronization and dynamic coupling between agents, formalized by the Free Energy Principle[55].
Epistemological Implications
The program of a “Physics of Life” requires abandoning the dichotomy between reductionism and holism. The challenge is not to derive biology from classical physics, but to expand physics to include systems where information and organization have causal power. This epistemological shift transforms the object of study: from passive matter to organizing matter. Rather than seeking closed universal laws, the physics of life pursues principles of compatibility between scales, which explain how biological systems preserve their identity in fluctuating environments. This introduces a methodological change: laws are replaced by regimes of organization, defined by interaction topologies, information flows, and critical transitions.
Future Scientific Agenda
The development of this discipline will require coordinated efforts on three fronts: Multi-scale Formal Models. It is a priority to advance the unification of nonequilibrium thermodynamics, information theory, and network dynamics. Models that integrate organizational closure, energy flow, and active inference will allow the formulation of equations of state for living systems. Experimental Validation and Falsification. The experimental agenda must be directed at testing the falsifiable predictions derived from the theoretical framework: the universality of criticality in organoids, the increase of functional information in evolutionary lineages, and the emergence of autocatalytic closure in synthetic systems. Technobiological Applications. The future of bioengineering and artificial intelligence will depend on incorporating these principles into the design of systems that learn, adapt, and evolve on their own. The engineering of the 21st century will not only build machines that work, but systems that live.
Towards a Physics of Creative Order
The Physics of Life is emerging as the science of creative order: the study of how matter, by dissipating energy, generates organization, meaning, and operational purpose. Its consolidation will mark a new phase in the history of knowledge: a truly integrative science where life, mind, and evolution are understood as expressions of the same organizational grammar. The challenge is not minor: to understand how nature turns disorder into meaning. But in that quest lies the promise of a theory capable of reconciling physics with biology—and, ultimately, with the very experience of being alive. The future lies not only in finding more universal laws, but in understanding the principles by which life navigates uncertainty and creates its own order.
Limitations of the Study
While this theoretical framework offers a unifying perspective on biological organization, several limitations should be acknowledged:
Theoretical and Conceptual Limitations: The proposed universal principles, while compelling, remain largely theoretical and require more extensive empirical validation across diverse biological systems. The framework’s broad scope necessarily sacrifices some biological specificity, potentially oversimplifying the intricate details of particular organisms or evolutionary contexts.
Measurement Challenges: Quantitative assessment of key concepts such as “functional information,” “organizational closure,” and “criticality” in real biological systems presents significant methodological challenges. Current metrics may not fully capture the multidimensional nature of these properties across different scales of biological organization.
Historical Contingency vs. Universal Principles: The tension between universal physical principles and historical evolutionary contingency remains unresolved. While the framework emphasizes universal organizational patterns, it may underweight the role of stochastic events, path dependence, and unique historical circumstances in shaping biological systems.
Scale Integration Challenges: Although the holographic principle suggests self-similar organization across scales, the specific mechanisms translating organizational principles from molecular to ecosystem levels require more detailed elaboration. The framework currently lacks comprehensive mathematical formalisms bridging these scale transitions.
Experimental Validation Gaps: Many predictions derived from this framework, particularly those concerning evolutionary directionality and critical transitions, require long-term experimental studies that are challenging to implement. The falsifiability of some principles remains limited by current technological capabilities.
Computational and Modeling Limitations: The complex, non-linear dynamics described in this framework pose significant challenges for computational modeling and simulation. Current modeling approaches may be insufficient to capture the full richness of multi-scale biological organization described by these principles.
These limitations highlight the need for continued interdisciplinary research to refine, test, and potentially revise the proposed framework through integration of theoretical insights with empirical data from diverse biological systems..
Resource Availability
Lead contact: Further information and requests for resources should be directed to and will be fulfilled by the lead contact, Elkin Navarro Quiroz (elkin.navarro@unisimon.edu.co).
Materials Availability
- This study did not generate new unique reagents.
Data and Code Availability
-
This paper analyzes existing, publicly available data. All data and theoretical models discussed are available from the cited references.
- This paper does not report original code.
- Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.
Author Contributions
Conceptualization, E.N.Q. and R.N.Q.; Methodology, E.N.Q., A.G., and C.F.P.; Investigation, E.N.Q., Y.B.L., E.Z.P., L.P.L., L.P.L., M.R.C., and D.P.G.; Writing – Original Draft, E.N.Q., E.Z.P., R.F.C., and I.S.G.; Writing – Review & Editing, R.N.Q., K.N.Q., A.G., C.F.P., and N.G.D.; Visualization, E.N.Q. and Y.B.L.; Supervision, E.N.Q.; Funding Acquisition, E.N.Q.; Resources, A.A.H., Y.B.L., L.P.L., and V.N.Q.
Acknowledgments
The authors acknowledge the collaborative support of the Center for Research in Critical Dynamics (Colombia), the INiBICA Institute (Spain), and the Center for Life Sciences Research (CICV, Universidad Simón Bolívar). Their shared commitment to transdisciplinary integration between physics and biology has made this conceptual synthesis possible. We also thank the Instituto Nacional de Salud de Colombia and the Colombian Association of Immunology (ACOI) for fostering a systemic and integrative vision of immunology that inspired and strengthened this work. This work was supported by the General System of Royalties of Colombia (Sistema General de Regalías, SGR) under the research Project Code BPIN: 2025000100150. This study was financed in part by the São Paulo Research Foundation (FAPESP), Brazil, Process Number 2025/14843-7.
Declaration Of Interests
The authors declare no competing interests.
Declaration Of Generative Ai And Ai-Assisted Technologies
During the preparation of this work, the authors used GPT-4 and Gemini Pro to assist in literature search, conceptual synthesis, structuring, and drafting of the manuscript. After using these tools, the authors reviewed, critically evaluated, and edited the content as needed and take full responsibility for the content of the publication.
References
- Woese, C.R. (2004). A New Biology for a New Century. Microbiol Mol Biol Rev 68, 173–186. [CrossRef]
- Goldenfeld, N., and Woese, C. (2011). Life is Physics: Evolution as a Collective Phenomenon Far From Equilibrium. Annu. Rev. Condens. Matter Phys. 2, 375–399. [CrossRef]
- Prigogine, I., and Nicolis, G. (1985). Self-Organisation in Nonequilibrium Systems: Towards A Dynamics of Complexity. In Bifurcation Analysis, M. Hazewinkel, R. Jurkovich, and J. H. P. Paelinck, eds. (Springer Netherlands), pp. 3–12. [CrossRef]
- Turing, A.M. (1952). The chemical basis of morphogenesis. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 237, 37–72. [CrossRef]
- Lloyd, D., Aon, M.A., and Cortassa, S. (2001). Why Homeodynamics, Not Homeostasis? The Scientific World JOURNAL 1, 133–145. [CrossRef]
- Maturana, H.R., and Varela, F.J. (1980). Autopoiesis and cognition: the realization of the living (D. Reidel Publishing).
- Rosen, R. (1991). Life itself: a comprehensive inquiry into the nature, origin, and fabrication of life (Columbia University Press).
- Varela, F.J. (1981). Describing the logic of the living: The adequacy and limitations of the idea of autopoiesis. In Autopoiesis: A theory of living organization, pp. 36–48.
- Kauffman, S.A. (1993). The origins of order: self-organization and selection in evolution (Oxford University Press).
- West, G.B., Brown, J.H., and Enquist, B.J. (2007). A general model for the origin of allometric scaling laws in biology. Science 276, 122–126. [CrossRef]
- West, G.B., and Brown, J.H. (2000). The origin of allometric scaling laws in biology. In Scaling in biology (Oxford University Press), pp. 87–112.
- Fleischaker, G.R. (1988). Autopoiesis: the status of its system logic. Biosystems 22, 37–49. [CrossRef]
- Ravasz, E., Somera, A.L., Mongru, D.A., Oltvai, Z.N., and Barabási, A.-L. (2002). Hierarchical organization of modularity in metabolic networks. Science 297, 1551–1555. [CrossRef]
- Bak, P., Tang, C., and Wiesenfeld, K. (1987). Self-organized criticality: an explanation of 1/f noise. Physical Review Letters 59, 381–384. [CrossRef]
- Muñoz, M.A. (2018). Colloquium: Criticality and dynamical scaling in living systems. Reviews of Modern Physics 90, 031001. [CrossRef]
- Kuramoto, Y. (1984). Chemical oscillations, waves, and turbulence (Springer-Verlag).
- Friston, K. (2010). The free-energy principle: a unified brain theory? Nature Reviews Neuroscience 11, 127–138. [CrossRef]
- Taleb, N.N. (2012). Antifragile: things that gain from disorder (Random House).
- Edelman, G.M., and Gally, J.A. (2001). Degeneracy and complexity in biological systems. Proceedings of the National Academy of Sciences 98, 13763–13768. [CrossRef]
- Hazen, R.M., Grew, E.S., Downs, R.T., Golden, J., and Hystad, G. (2023). On the roles of function and selection in evolving systems. Proceedings of the National Academy of Sciences 120, e2310223120. [CrossRef]
- Toussaint, O., and Schneider, E.D. (1998). The thermodynamics and evolution of complexity in biological systems. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 120, 3–9. [CrossRef]
- Kauffman, S. (2020). Answering Schrödinger’s “What Is Life?” Entropy 22, 815. [CrossRef]
- Kondepudi, D.K., De Bari, B., and Dixon, J.A. (2020). Dissipative structures, organisms and evolution. Entropy 22, 1305. [CrossRef]
- Painter, K.J., Hunt, G.S., Wells, K.L., Johansson, J.A., and Headon, D.J. (2012). Towards an integrated experimental-theoretical approach for assessing the mechanistic basis of hair and feather morphogenesis. Interface Focus 2, 433–450. [CrossRef]
- Mossio, M. (2024). Introduction: Organization as a Scientific Blind Spot. In Organization in Biology History, Philosophy and Theory of the Life Sciences., M. Mossio, ed. (Springer International Publishing), pp. 1–22. [CrossRef]
- Jureček, M., and Švorcová, J. (2025). Flowing boundaries in autopoietic systems and microniche construction. BioSystems 254, 105477. [CrossRef]
- Hordijk, W., Kauffman, S.A., and Steel, M. (2011). Required Levels of Catalysis for Emergence of Autocatalytic Sets in Models of Chemical Reaction Systems. IJMS 12, 3085–3101. [CrossRef]
- Yolles, M., and Frieden, B.R. (2021). Autopoiesis and Its Efficacy—A Metacybernetic View. Systems 9, 75. [CrossRef]
- Huitzil, S., and Huepe, C. (2024). Life’s building blocks: the modular path to multiscale complexity. Front Syst Biol 4, 1417800. [CrossRef]
- Hoffmann, H., and Payton, D.W. (2018). Optimization by Self-Organized Criticality. Sci Rep 8, 2358. [CrossRef]
- Shojaie, A., Jauhiainen, A., Kallitsis, M., and Michailidis, G. (2014). Inferring Regulatory Networks by Combining Perturbation Screens and Steady State Gene Expression Profiles. PLOS ONE 9, e82393. [CrossRef]
- Nourisa, J. and others (2025). geneRNIB: a living benchmark for gene regulatory network inference. bioRxiv. [CrossRef]
- Miller, W.B., Torday, J.S., and Baluška, F. (2020). The N-space Episenome unifies cellular information space-time within cognition-based evolution. Progress in Biophysics and Molecular Biology 150, 112–139. [CrossRef]
- Chang, J.E., and Turley, S.J. (2015). Stromal infrastructure of the lymph node and coordination of immunity. Trends in Immunology 36, 30–39. [CrossRef]
- Bahl, A., Pandey, S., Rakshit, R., Kant, S., and Tripathi, D. (2025). Infection-induced trained immunity: a twist in paradigm of innate host defense and generation of immunological memory. Infect Immun 93, e00472-24. [CrossRef]
- Singh, S.S., Haobijam, D., Malik, Md.Z., Ishrat, R., and Singh, R.K.B. (2018). Fractal rules in brain networks: Signatures of self-organization. Journal of Theoretical Biology 437, 58–66. [CrossRef]
- Heiney, K., Huse Ramstad, O., Fiskum, V., Christiansen, N., Sandvig, A., Nichele, S., and Sandvig, I. (2021). Criticality, Connectivity, and Neural Disorder: A Multifaceted Approach to Neural Computation. Front Comput Neurosci 15, 611183. [CrossRef]
- Berg, G., Rybakova, D., Fischer, D., Cernava, T., Vergès, M.-C.C., Charles, T., Chen, X., Cocolin, L., Eversole, K., Corral, G.H., et al. (2020). Microbiome definition re-visited: old concepts and new challenges. Microbiome 8, 103. [CrossRef]
- Baruah, G., and Lakämper, T. (2024). Stability, resilience and eco-evolutionary feedbacks of mutualistic networks to rising temperature. Journal of Animal Ecology 93, 989–1002. [CrossRef]
- Fung, T.C., Olson, C.A., and Hsiao, E.Y. (2017). Interactions between the microbiota, immune and nervous systems in health and disease. Nat Neurosci 20, 145–155. [CrossRef]
- Schrödinger, E. (1944). What is life? The physical aspect of the living cell (Cambridge University Press).
- Nguyen, L.L., and D’Amore, P.A. (2001). Cellular interactions in vascular growth and differentiation. In International Review of Cytology (Elsevier), pp. 1–48. [CrossRef]
- Lane, P.A., and Paulin, N. (2025). The Reality of Constraint and the Illusion of Control in Ecological Networks. Preprint at Computer Science and Mathematics. [CrossRef]
- Equihua, M., Espinosa Aldama, M., Gershenson, C., López-Corona, O., Munguía, M., Pérez-Maqueo, O., and Ramírez-Carrillo, E. (2020). Ecosystem antifragility: beyond integrity and resilience. PeerJ 8, e8533. [CrossRef]
- Goenner, H.F.M. (2014). On the History of Unified Field Theories. Part II. (ca. 1930–ca. 1965). Living Rev. Relativ. 17, 5. [CrossRef]
- Gintis, H. (2007). A framework for the unification of the behavioral sciences. Behav Brain Sci 30, 1–16. [CrossRef]
- Parker, M.C., Jeynes, C., and Walker, S.D. (2025). A Metric for the Entropic Purpose of a System. Entropy 27, 131. [CrossRef]
- Tavoni, G., Ferrari, U., Battaglia, F.P., Cocco, S., and Monasson, R. (2017). Functional coupling networks inferred from prefrontal cortex activity show experience-related effective plasticity. Network Neuroscience 1, 275–301. [CrossRef]
- Rubin, S. (2023). Cartography of the multiple formal systems of molecular autopoiesis: from the biology of cognition and enaction to anticipation and active inference. Biosystems 230, 104955. [CrossRef]
- Solé, R., Kempes, C., and Stepney, S. (2025). Origins of life: the possible and the actual. Phil. Trans. R. Soc. B 380, 20240281. [CrossRef]
- Wolf, Y.I., Katsnelson, M.I., and Koonin, E.V. (2018). Physical foundations of biological complexity. Proc. Natl. Acad. Sci. U.S.A. 115. [CrossRef]
- Toro-Domínguez, D., and Alarcón-Riquelme, M.E. (2021). Precision medicine in autoimmune diseases: fact or fiction. Rheumatology 60, 3977–3985. [CrossRef]
- Dehghani, N., and Levin, M. (2024). Bio-inspired AI: Integrating Biological Complexity into Artificial Intelligence. Preprint at arXiv. [CrossRef]
- Krauss, A. (2024). Science of science: A multidisciplinary field studying science. Heliyon 10, e36066. [CrossRef]
- Boisvert, R.F. (2016). Applied and Computational Mathematics Division: Summary of Activities for Fiscal Year 2015 (National Institute of Standards and Technology). [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



