
Submitted:
29 October 2025
Posted:
29 October 2025
You are already at the latest version
Abstract
Urban stray cats are often associated with ectoparasites and zoonotic pathogens due to their unsanitary living conditions and lack of veterinary care. Fleas, especially Ctenocephalides spp., are competent vectors of Bartonella spp., a genus of emerging bacterial pathogens with both public health and veterinary relevance. This study investigated the presence of Bartonella DNA in ectoparasitic fleas infesting stray cats in various urban habitats within the Klang Valley, Malaysia. A total of 204 fleas were collected from 89 stray cats. Fleas were identified morphologically using established taxonomic keys under a light microscope and further validated through PCR amplification of the mitochondrial cytochrome c oxidase subunit I (cox1) gene. Detection of Bartonella spp. was conducted by targeting the citrate synthase (gltA) gene. All fleas were confirmed as Ctenocephalides felis, with an infestation prevalence of 39.33% among the cats sampled. Of 118 C. felis specimens tested, 86.44% were positive for Bartonella DNA, one of the highest worldwide and significantly surpassing previous Malaysian reports. Sequencing of 12 positive samples showed identities with B. claridgeiae (58.3%), B. henselae (25.0%), an uncultured Bartonella species (8.3%) and a Bartonella isolate from a dog in Chile (8.3%). These results highlight the significant presence of Bartonella, causative agent of cat-scratch disease in stray cats, emphasizing their potential role as urban reservoirs and vectors. The findings underscore the need for ectoparasite surveillance and zoonotic pathogen control as integral components of stray animal management in Malaysia’s urban settings.
Keywords:
Vector
; Zoonosis
; Felis catus
; Infectious disease
; Animal management
1. Introduction
Stray cats are ubiquitous in urban ecosystems, often living in close proximity to humans and other domestic animals. Many of these cats originate from abandoned pets or unplanned litters, forming unregulated populations that thrive in densely populated areas. Due to limited access to veterinary care and poor health management, stray cats are frequently infested with ectoparasites, which play a key role in maintaining and transmitting zoonotic pathogens of public health significance [1]. This human–animal interface, intensified by urbanisation, provides an ideal setting for the emergence and spillover of vector-borne infections under the One Health framework.
Among feline ectoparasites, fleas particularly the cat flea Ctenocephalides felis are the most common and globally widespread [2]. C. felis exhibits remarkable ecological adaptability, infesting a wide range of mammalian hosts across both temperate and tropical regions [3]. Beyond causing dermatological irritation and allergic reactions in animals, C. felis serves as a competent vector for several zoonotic agents, notably Bartonella species. Of these, B. henselae and B. claridgeiae are of major medical relevance, being the primary etiological agents of cat scratch disease (CSD) in humans [4]. Human infection typically results from scratches or bites contaminated with flea faeces [5], and while CSD is usually self-limiting in immunocompetent individuals, severe systemic manifestations such as neuroretinitis, endocarditis, and prolonged fever have been documented, particularly in immunocompromised patients.
Globally, urban stray cats are increasingly recognised as reservoirs for Bartonella spp. and their flea vectors, underscoring the need for integrated surveillance at the animal–vector–human interface. However, in Malaysia, studies on flea infestations and Bartonella infections in stray cats remain limited, often constrained by small sample sizes or narrow geographic coverage. The Klang Valley, encompassing Kuala Lumpur and its surrounding metropolitan districts, is one of the most densely populated and urbanised regions in the country, hosting a large and growing population of free-roaming cats. Despite the frequent human–cat interactions in this region, data on the prevalence of C. felis infestations and their role as vectors of Bartonella spp. are scarce [4,6,7]. This paucity of epidemiological evidence limits public health risk assessment and hinders the development of evidence-based vector control strategies.
The present study aims to address this knowledge gap by investigating the natural infection rates of C. felis and Bartonella spp. in urban stray cats within the Klang Valley, Peninsular Malaysia. By integrating morphological and molecular identification approaches, this research provides updated insights into flea-borne Bartonella circulation in urban environments. The findings are expected to contribute to the growing body of One Health evidence needed to inform zoonotic disease surveillance, risk mitigation, and integrated vector management strategies in tropical urban settings.
2. Materials and Methods
2.1. Study Area and Flea Collection
All procedures involving animal handling and sample collection were conducted in strict accordance with institutional and international ethical standards for the humane treatment of animals. Approval was obtained from the Institutional Animal Care and Use Committee (IACUC), Universiti Malaya (Ethics Reference No: T/22052023/13032023-02/R). Flea sampling was conducted between October and December 2023 across ten urban sites within the Klang Valley, Peninsular Malaysia (Figure 1). This region represents one of the most densely populated and urbanised areas of the country, characterised by a large stray cat population living in close proximity to humans. Each site was visited only once to avoid resampling of individual cats.
Free-roaming cats were approached using food pellets as bait to encourage stationary behaviour. Only cats exhibiting no signs of illness or aggression were sampled. Fleas were collected using a fine-toothed stainless-steel comb, focusing on key anatomical regions such as the head, neck, torso, and tail. Immediately after combing, the flea-laden comb was sealed in a sterile zip-lock bag to prevent escape, and the bag was shaken to dislodge fleas. All animals were released unharmed upon completion of the procedure. Collected fleas were transported to the laboratory and preserved at −80°C until further processing. Morphological identification was performed under a stereomicroscope using standard taxonomic keys [8].
2.2. Study Area and Flea Collection
Genomic DNA was extracted from 118 individual fleas using the DNeasy Blood and Tissue Kit (Qiagen, Germany) following the manufacturer’s protocol, with minor modifications. Each flea was homogenized in 180 µL of ATL buffer using a sterile micropestle, followed by the addition of 20 µL of Proteinase K. The samples were incubated at 56°C with intermittent vortexing until complete tissue digestion was achieved. Subsequently, 200 µL of AL buffer and 200 µL of absolute ethanol were added to each lysate, which was then transferred to a spin column and centrifuged at 6000 × g for 1 minute. The column was washed sequentially with 500 µL of AW1 and AW2 buffers. Finally, DNA was eluted in 50 µL of AE buffer and stored at −20°C until polymerase chain reaction (PCR) analysis.
2.3. Molecular Identification of Fleas
Molecular confirmation of flea species was performed via PCR amplification of the mitochondrial cytochrome c oxidase subunit I (cox1) gene using the primer pair LCO and Cff-R [9] (Table 1), yielding an expected amplicon size of approximately 550 bp. Each 25 µL PCR reaction contained 12.5 µL of DreamTaq Green PCR Master Mix (Thermo Scientific, Lithuania), 1 µL of each primer (10 µM), 2 µL of genomic DNA, and 8.5 µL of nuclease-free water. Amplification was carried out under the following thermocycling conditions: initial denaturation at 95°C for 1 minute, followed by 35 cycles of denaturation at 95°C for 15 seconds, annealing at 55°C for 15 seconds, and extension at 72°C for 10 seconds, with a final extension at 72°C for 5 minutes (Table 2). Positive controls (known C. felis DNA) and negative controls (nuclease-free water) were included in each PCR run. Amplicons were visualized by electrophoresis on 1.0 % agarose gels stained with GelRed and compared against a 100 bp DNA ladder (GeneRuler™, 1st BASE, Malaysia). Positive PCR products were purified and sent for sequencing (Apical Scientific Sdn. Bhd., Malaysia) for species confirmation.
2.4. Detection of Bartonella spp. By PCR
Screening for Bartonella DNA was performed by amplifying a fragment of the citrate synthase (gltA) gene using the primer pair BhCS.781p (5′-GGGGACCAGCTCATGGTGG-3′) and BhCS.1137n (5′-AATGCAAAAAGAACAGTAAACA-3′) as described by Norman et al. [10] (Table 1), generating a 379 bp amplicon. Each PCR reaction (25 µL) contained 12.5 µL of DreamTaq Green PCR Master Mix (Thermo Scientific, Lithuania), 1 µL of each primer (10 µM), 2 µL of flea genomic DNA, and 8.5 µL of nuclease-free water. Thermocycling conditions consisted of an initial denaturation at 94°C for 10 minutes, followed by 35 cycles of denaturation at 94°C for 30 seconds, annealing at 51°C for 45 seconds, and extension at 72°C for 30 seconds, with a final extension at 72°C for 7 minutes (Table 2). Positive and negative controls were included in all PCR assays, using Bartonella henselae DNA as the positive control and sterile nuclease-free water as the negative control. PCR products were visualized via electrophoresis on 1.0 % agarose gels stained with GelRed and compared to a 100 bp molecular size marker. Positive amplicons were purified and submitted for bidirectional sequencing at Apical Scientific Sdn. Bhd. (Selangor, Malaysia).
2.5. Phylogenetic Analysis
Raw chromatograms were examined using SeqScanner v2 (Applied Biosystems) and manually edited for base-calling accuracy in BioEdit v7. Sequences were aligned using MEGA v11 [11] with default parameters, and consensus sequences were compared against the NCBI GenBank database using BLASTn for species identification. Phylogenetic trees were constructed using the Maximum Likelihood (ML) method in MEGA v11. The Tamura 3-parameter (T92) model was applied for cox1 gene sequences of fleas, while the Kimura 2-parameter (K2) model was used for Bartonella gltA sequences. Node reliability was evaluated with 1,000 bootstrap replicates. Pulex irritans [12] and Brucella melitensis [13] were designated as outgroups for flea and Bartonella phylogenies, respectively. Reference sequences retrieved from GenBank were included to confirm genetic clustering and species assignment.
2.6. Statistical Analysis
Flea infestation parameters were quantified following the definitions proposed by [14] to provide a comprehensive assessment of parasite–host interactions. The prevalence of infestation was calculated as the proportion of cats harbouring at least one flea, expressed as a percentage of the total number of cats examined. This parameter reflects the overall likelihood of flea presence within the sampled cat population.
Descriptive statistics were used to summarise flea infestation levels and Bartonella detection rates. Associations between categorical variables and flea infestation status were assessed using Fisher’s Exact Test. A p-value of < 0.05 was considered statistically significant. All statistical analyses were performed using GraphPad Prism v9 (GraphPad Software, USA).
3. Results
3.1. Flea Collection and Prevalence
A total of 204 fleas were recovered from 35 of the 89 stray cats sampled across various Klang Valley localities, yielding an overall infestation prevalence of 39.33 %. Fleas were detected in all surveyed locations except Kampung Baru. The highest flea burden originated from Pantai Dalam (n = 94; 45.19 %), followed by Vista Angkasa Apartment (n = 53; 25.98 %) and Kajang (n = 25; 12.25 %) (Table 3, Table S1). Female cats constituted the majority of the sampled population (n = 69; 77.53 %) compared to males (n = 20; 22.47 %). Of these, 27 females (39 %) and eight males (40 %) were infested (Table 4). Age distribution was similarly consistent, with infestation detected in 39.0 % of adults (33/84) and 40.0 % of juveniles (2/5) (Table 4). Fisher’s Exact Test revealed no significant association between flea infestation and either gender (P = 0.5711) or age group (P = 0.6571). Environmental assessment indicated that infestation prevalence was highest at public transportation areas (100 %), followed by housing (40 %) and eatery zones (34 %) (Table 4). Despite eatery areas contributing the most fleas (n = 136), their prevalence was comparatively low due to a higher number of sampled cats with few infested individuals.
3.2. Flea Identification
All 204 fleas were morphologically identified as Ctenocephalides felis (the cat flea) using established taxonomic keys [8]. Diagnostic characteristics included a long head with an angular frons, genal comb spines 1 and 2 of nearly equal length, and a distinct narrow dorsal incrassation [15]. No other flea species were detected among the examined specimens. For molecular confirmation, DNA from 118 individual fleas was successfully amplified targeting the mitochondrial cox1 gene (~ 550 bp). Ten representative sequences (479 bp) showed 97–100 % similarity with C. felis isolate Malay01 (GenBank accession no.: MT027230.1) in BLAST analyses, confirming morphological identification and demonstrating high sequence conservation among local populations (Table 5).
3.3. Molecular Detection of Bartonella spp.
Of the 118 flea DNA samples, 102 (86.44 %) tested positive for Bartonella spp. via PCR targeting the gltA gene, producing amplicons of the expected 379 bp size (Table 1). Twelve representative positive samples from different locations were sequenced for species identification. BLAST analysis of the 266 bp partial gltA sequences revealed the presence of two recognized species, B. henselae and B. claridgeiae, along with two uncharacterized Bartonella strains (Table 6). Ten sequences exhibited ≥98 % similarity with known type strains, confirming species identity. One sample (F103) displayed 92.51 % similarity to Bartonella sp. isolate Dog_9 (GenBank accession no.: MN233800.1), suggesting a potentially novel or divergent strain, though still above the 95.4 % similarity threshold for genus-level classification [16]. Specifically, three sequences (F01, F11, F33) matched B. henselae isolates from domestic cats in Brazil (MN107415.1), while seven (F23, F38, F44, F46, F83, F91, F109) were closely related to B. claridgeiae isolates PESET LAMADINUFF 23/25 (MH019300.1–MH019301.1). Another sample (F95) showed 98.48 % identity with an uncultured Bartonella sp. clone (MH279890.1) from a crab-eating fox (Cerdocyon thous) in southern Brazil.
3.4. Phylogenetic Analysis
Phylogenetic reconstruction of the cox1 gene sequences confirmed that all flea samples obtained in this study belonged to Ctenocephalides felis (Figure 2). All sequences formed a strongly supported monophyletic cluster (bootstrap = 96 %) together with C. felis isolate Malay01 (GenBank accession no.: MT027230.1), previously reported from cats in Southeast Asia. This cluster was clearly distinct from Ctenocephalides canis and Ctenocephalides orientis, which grouped into a separate, well-supported clade (bootstrap = 94 %). The topology thus corroborates both the morphological and molecular identification of the fleas as C. felis. Phylogenetic analysis of the Bartonella gltA gene sequences revealed the presence of two well-defined clades, corresponding to B. henselae and B. claridgeiae (Figure 3). Sequences F01, F11, and F33 clustered tightly with B. henselae isolates previously reported from domestic cats in Brazil and the United States, supported by a bootstrap value of 99 %. This B. henselae clade formed a sister relationship with B. koehlerae (GenBank accession no.: AF176091.1), indicating close evolutionary relatedness (bootstrap = 81 %). The seven B. claridgeiae sequences (F23, F38, F44, F46, F83, F91, F109) formed a robust monophyletic clade (bootstrap = 98 %) alongside reference strains from Brazil, the USA, and Switzerland. Notably, sequence F103, although related to this cluster, branched independently with relatively lower sequence similarity, suggesting possible intraspecific divergence or geographic variation. Sequence F95 was genetically distinct from both major clades but clustered closely with an uncultured Bartonella sp. clone (GenBank accession no.: MH279890.1) previously detected in a crab-eating fox (Cerdocyon thous) from Brazil (bootstrap = 99 %). This unique positioning may reflect lineage diversification within Bartonella species infecting C. felis in Malaysia.
4. Discussion
This study offers the latest insight into flea infestation patterns among urban stray cats in the Klang Valley, a region marked by close human–animal interactions and limited vector surveillance. The overall flea infestation rate of 39.33 % observed here, although lower than the national average of 71.83 % reported by [17], remains epidemiologically important and aligns with thresholds that raise public health concerns [18]. Similar levels of infestation have been reported in other urbanised tropical region such as Mexico (53.0 %) [19], though they are substantially lower than reports from Thailand (95.8 %) [20], Egypt (85.7 %) [21], and Iran (92.3 %) [22]. Variations in prevalence likely stem from differences in climate, host density, sampling methods, and local veterinary practices.
Urban Malaysia’s tropical humidity and steady temperatures create ideal conditions for continuous flea reproduction [23]. Stray cats frequenting busy areas, housing complexes, eateries, and transport hubs act as sentinels for urban flea persistence. The widespread infestation across these human-related zones highlights the need for integrated ectoparasite surveillance within a One Health approach to reduce zoonotic exposure risks among city populations.
No significant link was found between flea infestation and host sex or age, aligning with previous studies in Asia and the Middle East [20,21,22]. Although juvenile cats showed marginally higher infestation rates, these differences were not statistically significant, supporting findings that grooming behaviour, social structure, and environmental contact have a greater influence than intrinsic host factors [24,25].
The predominance of female cats in the sampling pool aligned with previous studies conducted by [26] and [17]. This situation may be explained behavioural accessibility, as males often avoid human contact due to territorial aggression. Such sex-biased sampling should be considered in future designs to better estimate true infestation demographics.
Morphological and molecular analysis confirmed Ctenocephalides felis as the only flea species parasitising stray cats in the Klang Valley. This finding aligns with global data recognising C. felis as the most widespread flea species associated with both companion and stray animals [27,28,29]. The use of cox1 barcoding improved taxonomic accuracy, confirming species identity and showing close genetic ties to the C. felis isolate Malay01 previously reported from Southeast Asia.
Given its broad host range and vector competence, C. felis acts as a crucial epidemiological link between animals and humans, especially in environments lacking regular veterinary care. The predominance of C. felis supports the hypothesis that this species is the main vector of Bartonella and other flea-borne pathogens in Malaysia’s urban ecosystems [4,8].
The detection of Bartonella DNA in 86.44 % of fleas is remarkably high, one of the highest recorded worldwide, and significantly surpasses previous Malaysian reports (28.0 %) [30] and figures from France (26.2 %) [31], the USA (22.8 %) [32], and Palestine (50.4 %) [33]. Such an elevated prevalence indicates a well-established enzootic cycle of Bartonella within stray cat populations and their flea vectors in the Klang Valley.
Sequencing of the gltA gene identified two pathogenic species, B. henselae and B. claridgeiae, both recognized as causative agents of cat scratch disease (CSD) in humans [5]. B. claridgeiae predominated (58.3 %), mirroring emerging trends in Asia and Latin America where it is increasingly implicated in human bartonellosis [34,35]. The co-circulation of B. henselae and B. claridgeiae reinforces the dual-vector reservoir role of stray cats and their fleas in maintaining zoonotic transmission cycles.
Notably, the detection of two divergent genotypes, Bartonella sp. clone IPVDF_18 and isolate Dog_9, highlights potential novel or regionally adapted Bartonella lineages. The close phylogenetic relationship of these Malaysian sequences to South American isolates [36,37] may suggest global dissemination through companion animal movement or vector migration, an increasingly recognised phenomenon in molecular epidemiology.
Phylogenetic analyses of cox1 and gltA sequences provided molecular confirmation of both flea and bacterial species identity. All C. felis sequences clustered monophyletically with C. felis isolate Malay01, forming a lineage distinct from C. canis and C. orientis, thus validating morphological diagnosis. Similarly, Bartonella sequences formed two well-supported clades representing B. henselae and B. claridgeiae. The branching pattern of sequences F95 and F103, which diverged from these main clades, suggests possible host adaptation or local genetic diversification within Malaysian Bartonella populations. These evolutionary insights strengthen the understanding of regional Bartonella ecology and its zoonotic potential.
The coexistence of high C. felis infestation and elevated Bartonella infection underscores the One Health importance of flea-borne zoonoses in urban Malaysia. Stray cats serve as both reservoirs and amplifiers of vector populations capable of transmitting CSD to humans, particularly in densely populated communities, open eateries, and waste accumulation zones.
Inadequate vector control and the absence of animal health monitoring facilitate ongoing pathogen circulation. Integrating flea surveillance with urban health strategies, particularly in collaboration with municipal authorities and animal welfare organizations, could help mitigate zoonotic risks. This aligns with WHO’s and OIE’s (WOAH) global call to strengthen vector-borne disease surveillance through cross-sectoral approaches [38,39].
While the study provides critical baseline data, limitations include restricted sampling coverage and reliance on a single gene marker (gltA). Expanding future research to include multiple loci (e.g., rpoB, ftsZ, ssrA) and additional pathogens such as Rickettsia felis and Mycoplasma haemofelis would enhance diagnostic resolution [40,41]. Broader spatial sampling across peri-urban and rural gradients, coupled with next-generation sequencing and vector-host interaction modelling, could further elucidate the ecological and evolutionary dynamics of flea-borne Bartonella in Southeast Asia.
Future studies should broaden sampling to include peri-urban and rural areas across Peninsular Malaysia, increase the sample size, and incorporate multi-host analysis involving cats, dogs, and rodents. They should also screen for additional flea-borne pathogens such as Rickettsia, Mycoplasma, and Brucella, and utilise whole-genome sequencing to investigate Bartonella evolution and virulence factors. An integrated surveillance approach that combines molecular diagnostics, vector ecology, and community education is crucial for reducing zoonotic risks. These findings emphasise the urgent need for coordinated One Health interventions that target the urban stray cat population as both a sentinel and reservoir of flea-borne zoonoses in Malaysia.
5. Conclusions
This study presents updated molecular and phylogenetic evidence of Ctenocephalides felis–borne Bartonella species circulating among stray cats in urban areas of the Klang Valley, Malaysia. The high prevalence of C. felis infestations and the notable detection rate of Bartonella DNA highlight a potentially underestimated zoonotic risk within densely populated urban environments. Phylogenetic analyses based on cox1 and gltA genes confirmed that all flea specimens were C. felis and identified B. henselae and B. claridgeiae, with several genetically diverse lineages suggesting possible novel or geographically adapted strains. These findings emphasise the importance of ongoing molecular surveillance of flea-borne pathogens in stray and domestic animal populations, especially in tropical cities where close human–animal contact facilitates transmission. Integrated control strategies combining vector management, stray animal health programmes, and public awareness are crucial to reducing the risk of Bartonella and other flea-borne pathogens spilling over to humans. Future research employing multi-locus or whole-genome approaches will further clarify the evolutionary dynamics and public health implications of these emerging pathogens in Southeast Asia.
Supplementary Materials
Table S1: The number of fleas collected by hosts within Klang Valley area.
Author Contributions
Conceptualization, Z.Y; Methodology, J.T.A.A., Z.Y., M.A.A., and M.R.A.H.; Validation, M.R.A.H., A.A.A.H, M.A.A. and Z.Y.; Formal Analysis, J.T.A.A, M.R.A.H and A.A.A.H; Investigation, J.T.A.A., Z.Y., M.A.A., A.A.A.H. and M.R.A.H.; Resources, Z.Y., M.A.A., M.R.A.H. and S.A.B.; Writing – Original Draft Preparation, J.T.A.A., Z.Y., M.A.A., and A.A.A.H.; Writing – Review and Editing, J.T.A.A., Z.Y. and A.A.A.H.; Visualization, J.T.A.A., Z.Y., and M.A.A.; Supervision, Z.Y., M.A.A., M.R.A.H. and S.A.B.; Project Administration, Z.Y., and M.A.A.; Funding Acquisition, Z.Y., S.A.B. and M.A.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Universiti Malaya Research Grant (RU Grant) (Code: RU006-2025A).
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Acknowledgments
The authors would like to extend their gratitude for the assistance of TIDREC surveillance team during the field sampling. We are grateful for the laboratory accessibility from Department of Microbiology, Universiti Malaya and their staff for the kind assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Centonze, L. A.; Levy, J. K. Characteristics of Free-Roaming Cats and Their Caretakers. J. Am. Vet. Med. Assoc. 2002, 220, 1627–1633. [Google Scholar] [CrossRef]
- Durden, L. A.; Hinkle, N. C. Chapter 10 - Fleas (Siphonaptera). ScienceDirect. https://www.sciencedirect.com/science/article/abs/pii/B9780128140437000108.
- Linardi, P. M.; Santos, J. L. C. Ctenocephalides felis felis vs. Ctenocephalides canis (Siphonaptera: Pulicidae): Some Issues in Correctly Identify These Species. Rev. Bras. Parasitol. Vet. 2012, 21, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Low, V. L.; Prakash, B. K.; Tan, T. K.; Sofian-Azirun, M.; Anwar, F. H. K.; Vinnie-Siow, W. Y.; AbuBakar, S. Pathogens in Ectoparasites from Free-Ranging Animals: Infection with Rickettsia asembonensis in Ticks, and a Potentially New Species of Dipylidium in Fleas and Lice. Vet. Parasitol. 2017, 245, 102–105. [Google Scholar] [CrossRef]
- Goldstein, E. J.; Abrahamian, F. M. Diseases Transmitted by Cats. ASM Press ebook. 2016, 133–150. [Google Scholar] [CrossRef]
- Mokhtar, A. S.; Tay, S. T. Molecular Detection of Rickettsia felis, Bartonella henselae, and B. clarridgeiae in Fleas from Domestic Dogs and Cats in Malaysia. Am. J. Trop. Med. Hyg. 2011, 85(5), 931–933. [Google Scholar] [CrossRef]
- Tay, S. T.; Mokhtar, A. S.; Low, K. C.; Mohd Zain, S. N.; Jeffery, J.; Abdul Aziz, N.; Kho, K. L. Identification of Rickettsiae from Wild Rats and Cat Fleas in Malaysia. Med. Vet. Entomol. 2014, 28, 104–108. [Google Scholar] [CrossRef]
- Che Kamaruddin, N.; Adrus, M.; Wan Ismail, W. N. Prevalence of Ectoparasites on a Stray Cat Population from “Town of Knowledge” Kota Samarahan, Sarawak, Malaysian Borneo. Turk. J. Vet. Anim. Sci. 2020, 44(6), 1212–1221. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M. B.; Hoeh, W. R.; Lutz, R. A.; Vrijenhoek, R. C. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. PubMed 1994, 3, 294–299. [Google Scholar]
- Norman, A.; Regnery, R. L.; Jameson, P. H.; Greene, C. E.; Krause, D. C. Differentiation of Bartonella-like Isolates at the Species Level by PCR-Restriction Fragment Length Polymorphism in the Citrate Synthase Gene. J. Clin. Microbiol. 1995, 33, 1797–1803. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Zhang, Y.; Nie, Y.; Li, L.-Y.; Chen, S.-Y.; Liu, G.-H.; Liu, W. Population Genetics and Genetic Variation of Ctenocephalides felis and Pulex irritans in China by Analysis of Nuclear and Mitochondrial Genes. Parasites Vectors 2022, 15. [Google Scholar] [CrossRef]
- Frank, H. K.; Boyd, S. D.; Hadly, E. A. Global Fingerprint of Humans on the Distribution of Bartonella Bacteria in Mammals. PLoS Negl. Trop. Dis. 2018, 12(11), e0006865–e0006865. [Google Scholar] [CrossRef]
- Krasnov, B.; Khokhlova, I.; Shenbrot, G. The Effect of Host Density on Ectoparasite Distributions: An Example of a Rodent Parasitized by Fleas. Ecol. 2002, 83, 164–175. [Google Scholar] [CrossRef]
- Hii, S. F.; Kopp, S. R.; Abdad, M. Y.; Thompson, M. F.; O’Leary, C. A.; Rees, R. L.; Traub, R. J. Molecular Evidence Supports the Role of Dogs as Potential Reservoirs for Rickettsia felis. Vector-Borne Zoonotic Dis. 2011, 11, 1007–1012. [Google Scholar] [CrossRef]
- Scola, B. L.; Zeaiter, Z.; Khamis, A.; Raoult, D. Gene-Sequence-Based Criteria for Species Definition in Bacteriology: The Bartonella Paradigm. Trends Microbiol. 2003, 11, 318–321. [Google Scholar] [CrossRef] [PubMed]
- Azrizal-Wahid, N.; Sofian-Azirun, M.; Low, V. L. Risk Factors Associated with Flea Infestation on Cats. Trop. Biomed. 2019, 36, 810–821. [Google Scholar] [PubMed]
- Abdullah, S.; Helps, C.; Tasker, S.; Newbury, H.; Wall, R. Pathogens in Fleas Collected from Cats and Dogs: Distribution and Prevalence in the UK. Parasites Vectors 2019, 12. [Google Scholar] [CrossRef]
- Cantó, G. J.; Guerrero, R. I.; Olvera-Ramírez, A. M.; Milián, F.; Mosqueda, J.; Aguilar-Tipacamú, G. Prevalence of Fleas and Gastrointestinal Parasites in Free-Roaming Cats in Central Mexico. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- S. Jittapalapong; A. Sangvaranond; T. Inpankaew; N. Pinyopanuwat; W. Chimnoi; C. Kengradomkij; S. Saengow; S. Wongnakphet. Ectoparasites of Stray Cats in Bangkok Metropolitan Areas, Thailand. Agric. Nat. Res. 2008, 42, 463–470. [Google Scholar]
- ElSeify, M.; Aggour, M.; Sultan, K.; Marey, N. Ectoparasites in Stray Cats in Alexandria Province, Egypt: A Survey Study. Alex. J. Vet. Sci. 2016, 48. [Google Scholar] [CrossRef]
- Nasser Hajipour; Mojtaba Keighobadi; Minas, A. ; Abad, R.; Mostafa Golabi; Badali, A. Prevalence of Flea Infestation in Stray Cats in North West of Iran, Iran. Biol. Forum. 2015, 7, 575–580. [Google Scholar]
- Kreppel, K. S.; Telfer, S.; Rajerison, M.; Morse, A.; Baylis, M. Effect of Temperature and Relative Humidity on the Development Times and Survival of Synopsyllus fonquerniei and Xenopsylla cheopis, the Flea Vectors of Plague in Madagascar. Parasites Vectors 2016, 9. [Google Scholar] [CrossRef]
- Salant, H.; Mumcuoglu, K. Y.; Baneth, G. Ectoparasites in Urban Stray Cats in Jerusalem, Israel: Differences in Infestation Patterns of Fleas, Ticks and Permanent Ectoparasites. Med. Vet. Entomol. 2013, 28, 314–318. [Google Scholar] [CrossRef]
- Farrell, S.; McGarry, J.; Noble, P. M.; Pinchbeck, G. J.; Cantwell, S.; Radford, A. D.; Singleton, D. A. Seasonality and Other Risk Factors for Fleas Infestations in Domestic Dogs and Cats. Med. Vet. Entomol. 2023, 37. [Google Scholar] [CrossRef]
- Mohd Zain, S. N.; Sahimin, N.; Pal, P.; Lewis, J. W. Macroparasite Communities in Stray Cat Populations from Urban Cities in Peninsular Malaysia. Vet. Parasitol. [CrossRef]
- Xhaxhiu, D.; Kusi, I.; Rapti, D.; Visser, M.; Knaus, M.; Lindner, T.; Rehbein, S. Ectoparasites of Dogs and Cats in Albania. Parasitology Res. J. 2009, 105, 1577–1587. [Google Scholar] [CrossRef] [PubMed]
- Zouari, S.; Khrouf, F.; M’ghirbi, Y.; Bouattour, A. First Molecular Detection and Characterization of Zoonotic Bartonella Species in Fleas Infesting Domestic Animals in Tunisia. Parasites Vectors. [CrossRef]
- Jamal; Rabah, S. ; Yacoub, H.; Hajrah, N. H.; Atef, A.; Al-Matary, M.; Edris, S.; Alharbi, M. G.; Magdah Ganash; Jazem Mahyoub; Al-Hindi, R. R.; Al-Ghamdi, K. M.; Hall, N.; Bahieldin, A.; Kamli, M. R.; Rather, I. A. Molecular Evolution of Cytochrome c Oxidase-I Protein of Insects Living in Saudi Arabia. PLOS ONE 2019, 14, e0224336–e0224336. [Google Scholar] [CrossRef]
- Azrizal-Wahid, N.; Sofian-Azirun, M.; Low, V. L. Flea-Borne Pathogens in the Cat Flea Ctenocephalides Felis and Their Association with MtDNA Diversity of the Flea Host. Comp. Immunol. Microbiol. Infect. Dis. 2021, 75, 101621. [Google Scholar] [CrossRef] [PubMed]
- Rolain, J.; Franc, M.; Davoust, B.; Didier Raoult. Molecular Detection of Bartonella quintana, B. koehlerae, B. henselae, B. clarridgeiae, Rickettsia felis, and Wolbachia pipientis in Cat Fleas, France. Emerg. Infect. Dis. 2003, 9, 338–342. [Google Scholar] [CrossRef]
- Lappin, M. R.; Griffin, B.; Brunt, J. D.; Riley, A.; Burney, D. P.; Hawley, J. R.; Brewer, M.; Jensen, W. Prevalence of Bartonella Species, Haemoplasma Species, Ehrlichia Species, Anaplasma phagocytophilum, and Neorickettsia risticii DNA in the Blood of Cats and Their Fleas in the United States. J. Feline. Med. Surg. 2006, 8, 85–90. [Google Scholar] [CrossRef]
- Nasereddin, A.; Risheq, A.; Harrus, S.; Azmi, K.; Ereqat, S.; Baneth, G.; Salant, H.; Mumcuoglu, K. Y.; Abdeen, Z. Bartonella Species in Fleas from Palestinian Territories: Prevalence and Genetic Diversity. J. Vector. Ecol. 2014, 39, 261–270. [Google Scholar] [CrossRef]
- Persichetti, M.-F.; Solano-Gallego, L.; Serrano, L.; Altet, L.; Reale, S.; Masucci, M.; Pennisi, M.-G. Detection of Vector-Borne Pathogens in Cats and Their Ectoparasites in Southern Italy. Parasites Vectors 2016, 9. [Google Scholar] [CrossRef] [PubMed]
- Saengsawang, P.; Kaewmongkol, G.; Phoosangwalthong, P.; Chimnoi, W.; Inpankaew, T. Detection of Zoonotic Bartonella Species in Ticks and Fleas Parasitizing Free-Ranging Cats and Dogs Residing in Temples of Bangkok, Thailand. Vet. Parasitol. Reg. Stud. Reports. 2021, 25, 100612. [Google Scholar] [CrossRef] [PubMed]
- Schott, D.; Souza, U. A.; B. Dall'Agnol; Webster, A.; Doyle, R.; Peters, F.; M. Favarini; F. Mazim; Rosa, A. O.; Jardim, A.; Trigo, T. C.; Reck, J. Detection of Rickettsia spp. And Bartonella spp. In Ctenocephalides felis Fleas from Free-Ranging Crab-Eating Foxes (Cerdocyon thous). Med. Vet. Entomol. 2019, 33, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Weinborn-Astudillo, R. M.; Pau, N.; Tobar, B. Z.; Jaffe, D. A.; Boulouis, H.-J.; Sepúlveda, P.; Müller, A.; Chomel, B. B. Bartonella Infection in Stray Dogs from Central and Southern Chile (Linares and Puerto Montt). Vector-Borne Zoonotic Dis. 2020, 20, 187–192. [Google Scholar] [CrossRef]
- WHO. WHO Results Report 2023 Shows Notable Health Achievements and Calls for Concerted Drive toward Sustainable Development Goals. www.who.int. https://www.who.int/news/item/07-05-2024-who-results-report-2023-shows-notable-health-achievements-and-calls-for-concerted-drive-toward-sustainable-development-goals.
- WOAH. Vector-borne diseases surveillance: a global health imperative, /: World Organisation for Animal Health. https.
- Cruz, T. N. D. A. O.; Gonçalves, L. R.; Eduarda, M.; Marcos Rogério André; Alexandre Diaz Munhoz; Santiago, R. ; Fabiana Lessa Silva. Threat under Cats’ Claws: Molecular Detection and Risk Factors for Zoonotic Bartonella Species in Blood and Claw Samples from Cats in Brazil. Acta. Trop. 2022, 232, 106496–106496. [Google Scholar] [CrossRef]
- Deak, G.; Safarov, A.; Xie, X. C.; Wang, R.; Mihalca, A. D.; Šlapeta, J. Fleas from the Silk Road in Central Asia: Identification of Ctenocephalides canis and Ctenocephalides orientis on Owned Dogs in Uzbekistan Using Molecular Identification and Geometric Morphometrics. Parasites Vectors 2022, 15. [Google Scholar] [CrossRef]
Figure 1.
A map illustrating the distributions of the ten sampling sites for stray cat screening.
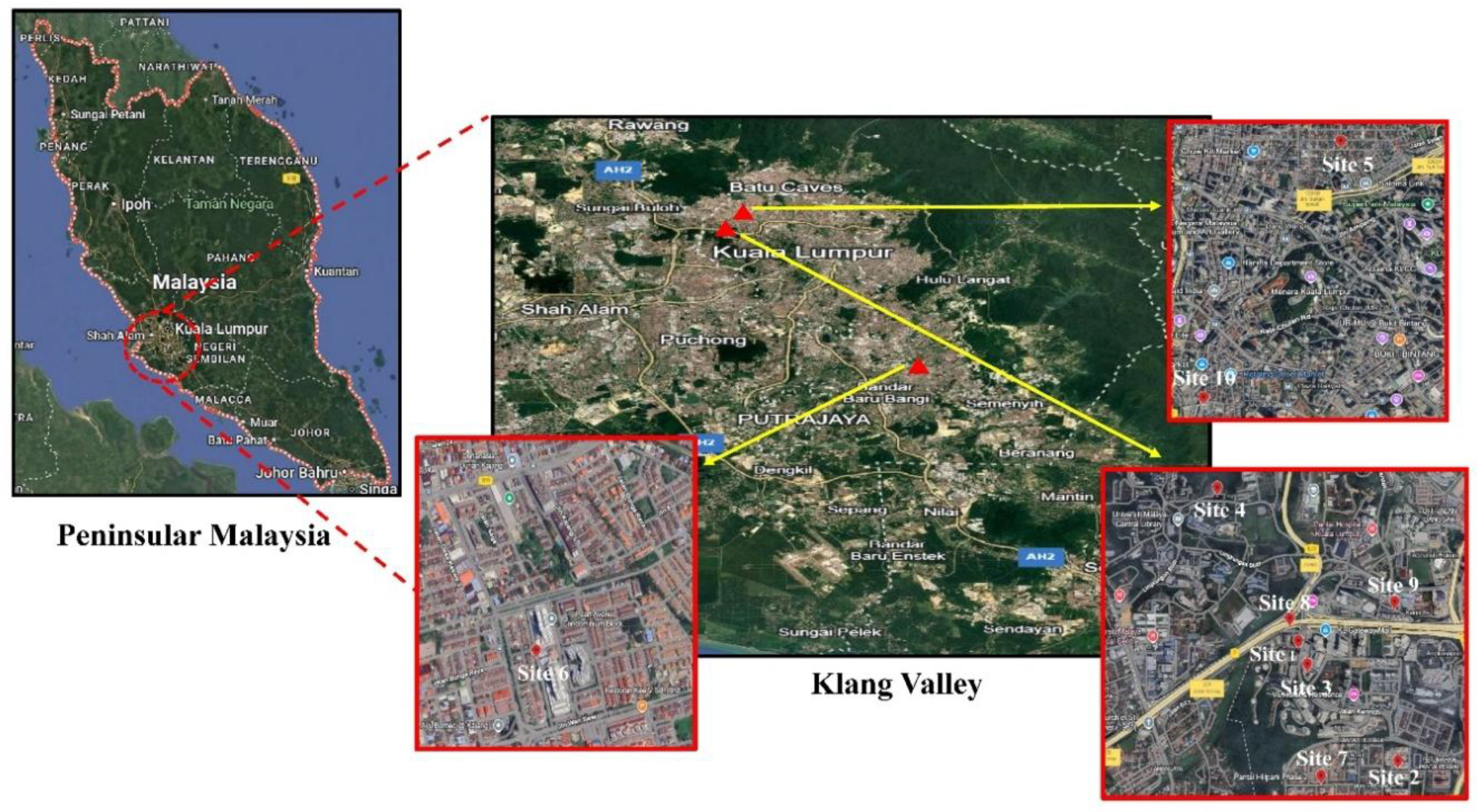
Figure 2.
Phylogenetic tree based on 479 bp of cox1 gene of flea sample from Klang Valley and constructed by using MEGA11 software. The tree was obtained by using the Maximum Likelihood method and Tamura 3-parameter model. Bootstrap support was calculated by using 1000 replicates and >60 % bootstrap values are shown. Pulex irritans isolate CCCF-20 was used as an outgroup for this study. Bold and asterisk (*) indicate the samples in this study.
Figure 2.
Phylogenetic tree based on 479 bp of cox1 gene of flea sample from Klang Valley and constructed by using MEGA11 software. The tree was obtained by using the Maximum Likelihood method and Tamura 3-parameter model. Bootstrap support was calculated by using 1000 replicates and >60 % bootstrap values are shown. Pulex irritans isolate CCCF-20 was used as an outgroup for this study. Bold and asterisk (*) indicate the samples in this study.
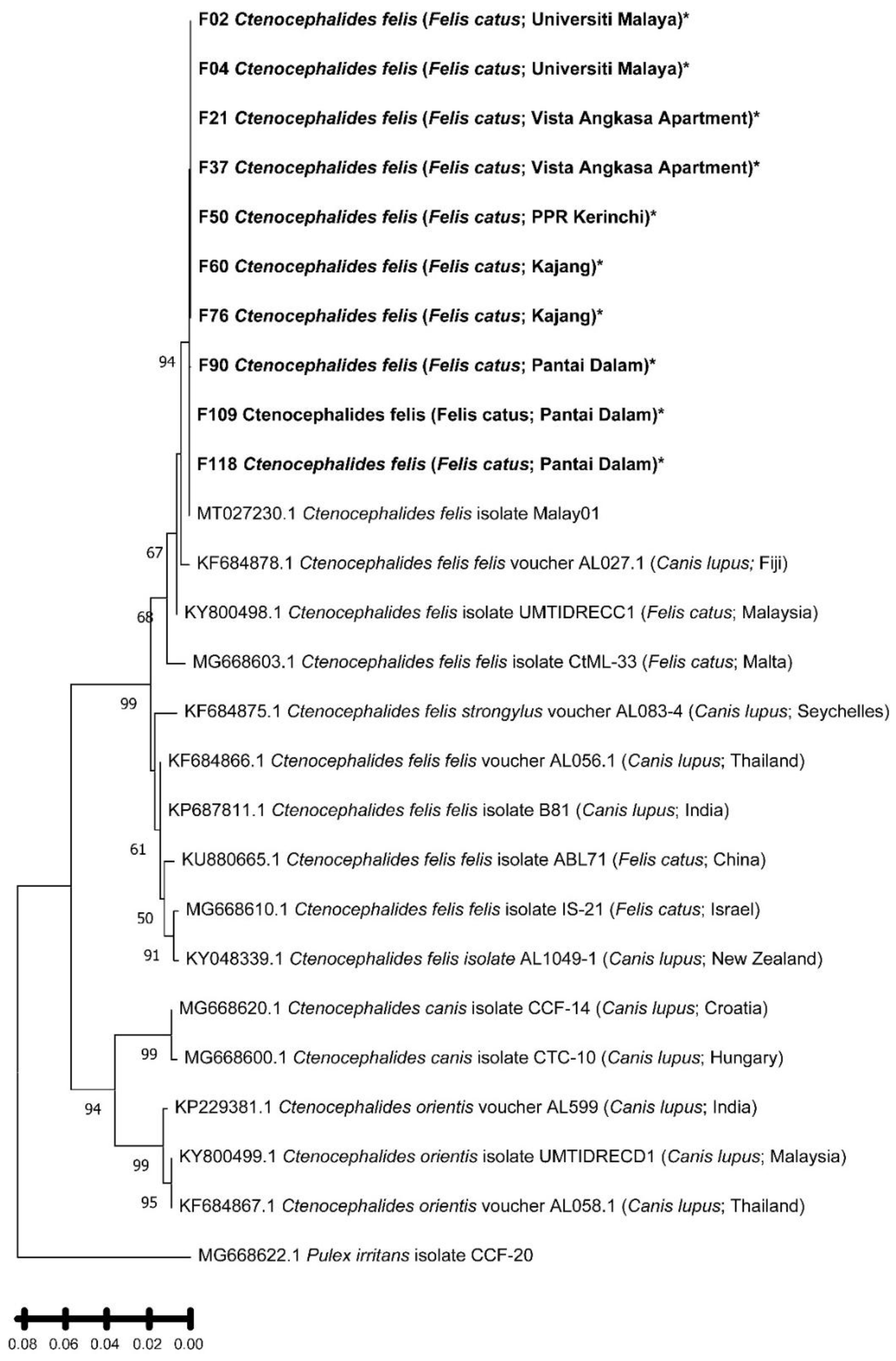
Figure 3.
Phylogenetic tree based on 266 bp citrate synthase (gltA) gene of Bartonella sample from Klang Valley and constructed by using MEGA11 software. The tree was obtained by using the Maximum Likelihood method and Kimura 2-parameter model. Bootstrap support was calculated by using 1000 replicates and >55 % bootstrap values are shown. Brucella melitensis strain 16M was used as an outgroup for this study. Bold and asterisk (*) indicate the samples in this study.
Figure 3.
Phylogenetic tree based on 266 bp citrate synthase (gltA) gene of Bartonella sample from Klang Valley and constructed by using MEGA11 software. The tree was obtained by using the Maximum Likelihood method and Kimura 2-parameter model. Bootstrap support was calculated by using 1000 replicates and >55 % bootstrap values are shown. Brucella melitensis strain 16M was used as an outgroup for this study. Bold and asterisk (*) indicate the samples in this study.
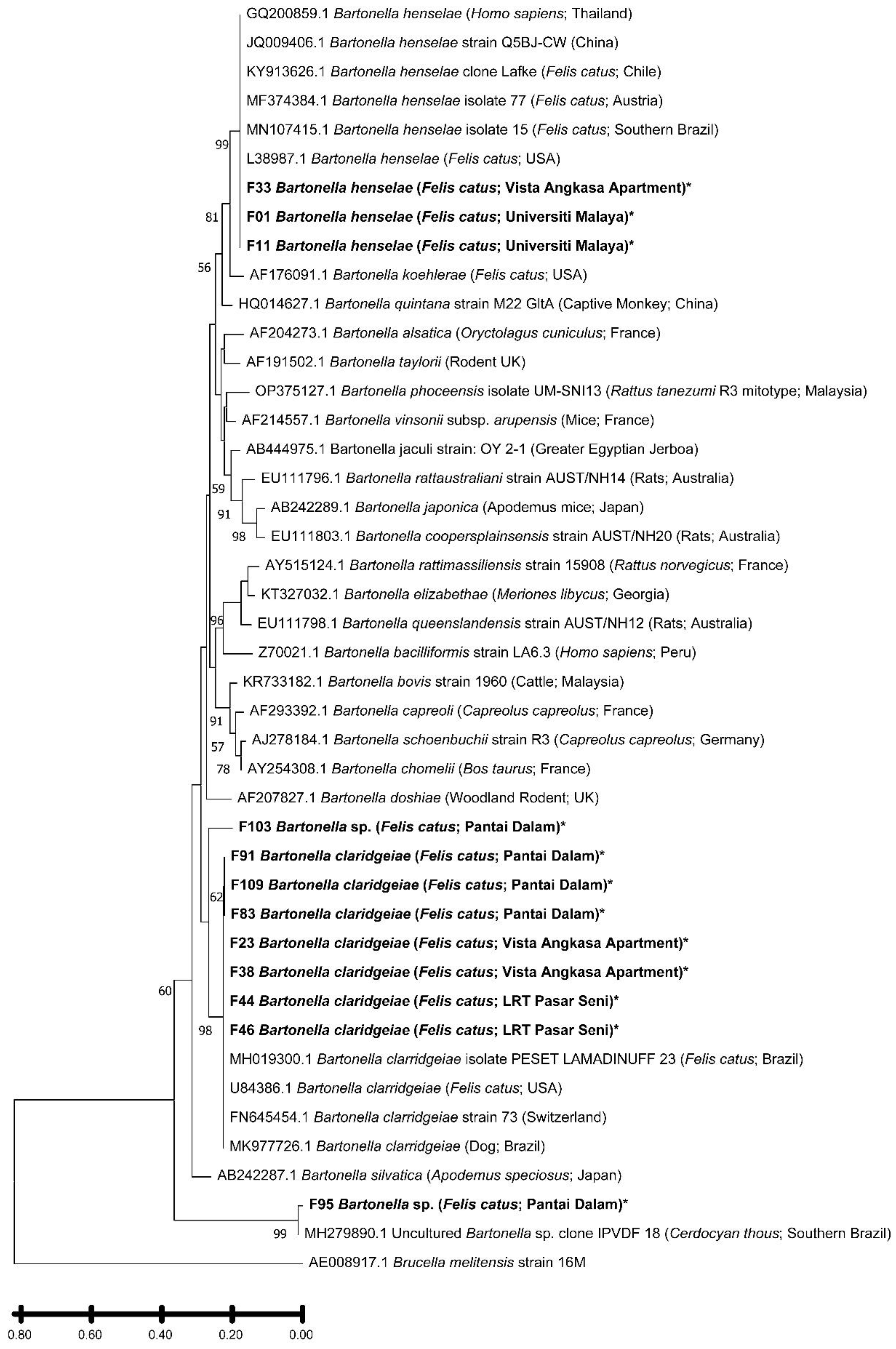

Table 1.
Primers used for PCR assays in this study.
| Gene | Primer Name | Sequence (5′- 3′) | Direction | Detection | Reference |
| cox1 | LCO | GGT CAA CAA ATC ATA AAG ATA TTG G | Forward | Flea species | [9] |
| Cff-R | GAA GGG TCA AAG AAT GAT GT | Reverse | |||
| gltA | BhCS.781p | GGG GAC CAG CTC ATG GTG G | Forward | Bartonella | [10] |
| BhCS.1137n | AAT GCA AAA AGA ACA GTA AAC A | Reverse |
Table 2.
Cycling conditions used for PCR assays in this study.
| Gene | Initial Denaturation | Denaturation | Annealing | Extension | Final Extension | Cycle | Amplicon size (bp) | Reference |
| cox1 | 95 °C 1 min |
95 °C 15 s |
55 °C 15 s |
72 °C 10 s |
72 °C 5 min |
35 | 550 | [9] |
| gltA | 94 °C 10 min |
94 °C 30 s |
51 °C 45 s |
72 °C 30 s |
72 °C 7 min |
35 | 379 | [10] |
Table 3.
Number of fleas collected according to classification of area within Klang Valley.
| Location type | Location Code | Location Name | Number of cats examined | Number of cats infested with fleas (Prevalence) | Total fleas collected |
| Housing Area | Site 1 | Vista Angkasa Apartment | 14 | 7 (50.0) | 53 |
| Site 2 | PPR Kerinchi | 7 | 1 (14.2) | 3 | |
| Site 3 | Pangsapuri 17 Tingkat Kerinchi | 4 | 2 (50.0) | 3 | |
| Eatery Area | Site 4 | Universiti Malaya | 26 | 6 (23.1) | 17 |
| Site 5 | Kampung Baru | 7 | 0 (0.0) | 0 | |
| Site 6 | Kajang | 5 | 3 (60.0) | 25 | |
| Site 7 | Pantai Dalam | 21 | 11 (52.4) | 94 | |
| Public Transportation Area | Site 8 | LRT Universiti Station | 2 | 2 (100.0) | 2 |
| Site 9 | LRT Kerinchi Station | 1 | 1 (100.0) | 3 | |
| Site 10 | LRT Pasar Seni Station | 2 | 2 (100.0) | 4 | |
| Total | 89 | 35 | 204 |
Table 4.
Prevalence of fleas (n = 204) on 89 sampled cats based on identified risk factors in the study areas.
Table 4.
Prevalence of fleas (n = 204) on 89 sampled cats based on identified risk factors in the study areas.
| Risk Factor | Variable | Flea-infested cat | Total fleas | Prevalence of Infestation (%) |
| Area | Housing area (n = 25) | 10 | 59 | 40.0 |
| Eatery area (n = 59) | 20 | 136 | 33.9 | |
| Public transportation area (n = 5) | 5 | 9 | 100.0 | |
| Gender | Male (n = 20) | 8 | 35 | 40.0 |
| Female (n = 69) | 27 | 169 | 39.0 | |
| Age | Adult (n = 84) | 33 | 200 | 39.0 |
| Juvenile (n = 5) | 2 | 4 | 40.0 |
Note: Prevalence of infestation was calculated as the proportion of cats harbouring at least one flea, expressed as a percentage of the total number of cats examined.
Table 5.
BLASTn results for the obtained flea sequences based on the cox1 gene region.
| Flea in this study | Location | Closest GenBank match | Maximum Identity (%) | Query cover (%) | E-value | Origin |
| F02 | University Malaya | C. felis isolate Malay01 (MT027230.1) | 99.83 | 99 | 0.0 | East and Southeast Asia |
| F04 | University Malaya | C. felis isolate Malay01 (MT027230.1) | 97.83 | 98 | 0.0 | East and Southeast Asia |
| F21 | Vista Angkasa Apartment | C. felis isolate Malay01 (MT027230.1) | 100.00 | 98 | 0.0 | East and Southeast Asia |
| F37 | Vista Angkasa Apartment | C. felis isolate Malay01 (MT027230.1) | 100.00 | 98 | 0.0 | East and Southeast Asia |
| F50 | PPR Kerinchi | C. felis isolate Malay01 (MT027230.1) | 100.00 | 100 | 0.0 | East and Southeast Asia |
| F60 | Kajang | C. felis isolate Malay01 (MT027230.1) | 100.00 | 100 | 0.0 | East and Southeast Asia |
| F76 | Kajang | C. felis isolate Malay01 (MT027230.1) | 100.00 | 98 | 0.0 | East and Southeast Asia |
| F90 | Pantai Dalam | C. felis isolate Malay01 (MT027230.1) | 100.00 | 100 | 0.0 | East and Southeast Asia |
| F109 | Pantai Dalam | C. felis isolate Malay01 (MT027230.1) | 100.00 | 100 | 0.0 | East and Southeast Asia |
| F118 | Pantai Dalam | C. felis isolate Malay01 (MT027230.1) | 99.83 | 98 | 0.0 | East and Southeast Asia |
Table 6.
The BLASTn results for the obtained Bartonella sequences based on gltA gene region.
| Sample ID | Location | Closest GenBank match | Maximum Identity (%) | Query cover (%) | E-value | Origin |
| F01 | University Malaya | B. henselae isolate 15 (MN107415.1) | 99.70 | 100 | 5e-172 | Brazil |
| F11 | University Malaya | B. henselae isolate 15 (MN107415.1) | 99.70 | 100 | 5e-172 | Brazil |
| F23 | Vista Angkasa Apartment | B. claridgeiae isolate PESET LAMADINUFF 23 (MH019300.1) | 99.41 | 100 | 2e-170 | Brazil |
| F33 | Vista Angkasa Apartment | B. henselae isolate 15 (MN107415.1) | 98.53 | 100 | 2e-166 | Brazil |
| F38 | Vista Angkasa Apartment | B. claridgeiae isolate PESET LAMADINUFF 23 (MH019300.1) | 99.41 | 100 | 2e-170 | Brazil |
| F44 | LRT Pasar Seni | B. claridgeiae isolate PESET LAMADINUFF 25 (MH019301.1) | 99.69 | 96 | 6e-166 | Brazil |
| F46 | LRT Pasar Seni | B. claridgeiae isolate PESET LAMADINUFF 23 (MH019300.1) | 99.70 | 100 | 2e-171 | Brazil |
| F83 | Pantai Dalam | B. claridgeiae isolate PESET LAMADINUFF 25 (MH019301.1) | 99.11 | 100 | 1e-167 | Brazil |
| F91 | Pantai Dalam | B. claridgeiae isolate PESET LAMADINUFF 25 (MH019301.1) | 98.24 | 100 | 1e-163 | Brazil |
| F95 | Pantai Dalam | Uncultured Bartonella sp. clone IPVDF_18 (MH279890.1) | 98.48 | 98 | 2e-160 | Brazil |
| F103 | Pantai Dalam | Bartonella sp. isolate Dog_9 (MN233800.1) | 92.51 | 99 | 4e-128 | Chile |
| F109 | Pantai Dalam | B. claridgeiae isolate PESET LAMADINUFF 23 (MH019300.1) | 99.41 | 100 | 8e-170 | Brazil |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



