
Submitted:
28 October 2025
Posted:
30 October 2025
You are already at the latest version
Abstract
Powdery mildew poses a persistent threat to global vegetable and fruit production, particularly affecting leafy crops such as lettuce, spinach, and cucurbits. Conventional control strategies including chemical fungicides, biological agents, and resistant cultivars face limitations due to resistance development, environmental toxicity, and inconsistent field efficacy. This review explores the emerging role of nanotechnology, specifically nanoparticles and nanosuspensions, in managing powdery mildew. Metallic nanoparticles and non-metallic variants demonstrate potent antifungal activity through mechanisms such as membrane disruption, reactive oxygen species (ROS) generation, and gene regulation. Encapsulated nano-fungicides and sprayable essential oils represent potential application methods that could enhance delivery precision and activate plant defense mechanisms against powdery mildew. The integration of smart delivery systems and digital agriculture platforms offers promising avenues for precision disease management. Integrating the application of nanoparticles (NPs) and nanosuspensions (NSs) with smart and digital delivery systems could be a promising strategy for managing powdery mildew infestation in fruits and vegetables. Despite their potential, challenges including ecotoxicity, formulation stability, scalability, and regulatory gaps must be addressed. This review under-scores the need for interdisciplinary research to advance safe, effective, and sustainable nano-enabled solutions for powdery mildew control.
Keywords:
fruits
; fungicides
; nanoparticles
; nanosuspension
; powdery mildew
; vegetables
1. Introduction
Vegetables including lettuce (Lactuca sativa), spinach (Spinacia oleracea), kale (Brassica oleracea), and arugula are pivotal to global nutrition, supplying essential nutrients and fiber to diverse populations [1]. These crops, however, face recurring threats from powdery mildew, a fungal disease that impairs photosynthesis and reduces marketable yield by 30–60% under severe outbreaks [1]. Powdery mildew significantly impacts leafy produce in the U.S., particularly affecting crops like lettuce and cucurbits. Furthermore, powdery mildew poses a persistent and significant threat to fruit production on a global scale in general, and US in particular [2]. The effects manifest as decreased fruit size and number, premature leaf senescence, and lower market quality due to sunburned or poorly ripened fruit [3,4]. Moreover, the presence of powdery mildew on harvested leaves reduces their visual appeal and shelf life, making them unsuitable for fresh markets.
The disease powdery mildew thrives in warm, dry climates with high humidity and is especially problematic in greenhouse and field-grown leafy greens. Key pathogens include Erysiphe cruciferarum in crucifers, Golovinomyces cichoracearum and Podosphaera xanthii in cucurbits and Podosphaera fusca in lettuce [5]. In plant species, these pathogens cause considerable damage to leave, stems, and fruits during development, leading to substantial economic losses [6,7]. Additionally, in cucumbers, powdery mildew causes malformed fruit and lowers sugar content; in tomatoes and peppers, it results in defoliation and sunscald [8]. In cucurbits, the disease shortens harvest periods and lowers fruit quality, affecting flavor and storage [3,4]. Infected plants are also more vulnerable to secondary diseases [4]. Powdery mildew impairs photosynthesis by disrupting chlorophyll fluorescence, electron transport, and lipid metabolism in chloroplasts which diminishes plant productivity and vigor [9,10,11]. The severity of impact depends on infection level and crop species. A variety of strategies such as chemical fungicides, integrated pest management, biological controls, resistant cultivars, and nanotechnology have been implemented to combat powdery mildew infestations.
Chemical fungicides have been the cornerstone of powdery mildew management for decades. Commonly used classes include demethylation inhibitors (DMIs), quinone outside inhibitors (QoIs), and succinate dehydrogenase inhibitors (SDHIs) [12]. These fungicides target specific biochemical pathways in the fungal cells, disrupting their growth and reproduction. The application of fungicides on vegetables showed significantly lower powdery mildew infection rates, with some treatments reducing incidence to 0.7% compared to 55.3% in untreated fields [5]. Traditional interventions, such as sulfur or copper fungicides and integrated pest management (IPM), confront obstacles like development of fungal resistance, biochemical residues, and environmental and health risks and limited efficacy under certain environmental conditions [8]. Despite the initial effectiveness of such fungicides, several challenges have emerged, including the rapid development of resistance in powdery mildew pathogens [12]. Due to their high reproductive rates and genetic variability, powdery mildew fungi can quickly adapt to selective pressures imposed by fungicides. For instance, resistance to QoIs has been widely reported in Blumeria graminis and Podosphaera xanthii, rendering these fungicides ineffective in many regions [12]. Most fungicides have a single-site mode of action, which increases the risk of resistance development. Once a mutation occurs in the target site, the entire class of fungicides may become ineffective. This is particularly problematic in obligate biotrophs like powdery mildew fungi, which cannot be cultured easily for resistance screening, making early detection difficult [12]. Besides, one of the primary issues is poor water solubility, which reduces the bioavailability of many active ingredients, reducing their effectiveness and requiring higher, more frequent applications. Moreover, conventional fungicides often degrade quickly and harm non-target organisms, prompting the need for safer, more effective alternatives.
To address these persistent challenges, the deployment of biological control agents, notably Ampelomyces quisqualis, a mycoparasite of powdery mildew, as well as microbial formulations comprising Trichoderma spp. and Bacillus subtilis, has garnered considerable attention. These biocontrol strategies suppress pathogenic fungi via mechanisms such as resource competition, antibiosis, and direct parasitism, though field efficacy remains variable [13]. In addition, breeding powdery mildew-resistant cultivars is a sustainable alternative to chemical control. Resistance genes (R genes) such as Pm3, Pm21, and Pm55 have been introduced into various crops, offering race-specific or broad-spectrum resistance [14]. However, this strategy is not without limitations including pathogens that can overcome host resistance through mutation or recombination, especially when a single R gene is deployed over large areas. This phenomenon, known as the “boom and bust” cycle, has been observed in cucumber, where initially resistant cultivars became susceptible within a few growing seasons [14]. Resistance genes may be linked to undesirable agronomic traits, such as reduced yield or delayed maturity. Moreover, pyramiding multiple R genes to enhance durability can be technically challenging and time-consuming. In many crops, the molecular basis of powdery mildew resistance remains poorly understood. For example, in cucumber, proteomic studies have revealed complex responses involving redox homeostasis and photosynthesis, but the specific pathways and genes involved are still being elucidated [15,16]. This knowledge gap hampers the development of durable resistance strategies.
Integrated Pest Management (IPM) combines chemical, biological, and cultural practices to control powdery mildew, though its effectiveness depends on local conditions, grower expertise, and available infrastructure [17,18]. Cultural methods like crop rotation and pruning can reduce disease incidence but are often labor-intensive and only partially effective [19]. IPM's success is further challenged by evolving fungal resistance, environmental hazards, and stricter residue regulations, while breeding resistant cultivars is complicated by pathogen variability. Overuse of fungicides increase resistance risk, and chemical controls pose environmental and health concerns, leading to tighter regulations and fewer available options [12, 20, 21).
Given the limitations of existing solutions, nanotechnology has emerged as a promising frontier in plant disease management. Nano-fungicides (nanosuspension or nanoparticles formulated), which incorporate nanoparticles with antifungal properties or serve as carriers for conventional fungicides, offer several advantages compared to the traditional fungicides [22]. Nanotechnology reduces pesticide usage, addressing the 90% loss of applied pesticides and promoting eco-friendly practices [23]. Nanosensors facilitate early detection of pathogens, enabling timely interventions [24]. Emerging nanotechnology presents a novel plant protection paradigm. By leveraging nanoparticles materials ≤100 nm and nanosuspensions, which are colloidal dispersions of these particles, researchers are developing tools that enhance delivery efficiency and achieve targeted antifungal actions [25,26]. This review assesses biology, mechanisms, field applications, and limitations of nanoparticle-based approaches for controlling powdery mildew in fruits and leafy vegetables.
2. Powdery Mildew: Biology and Impact
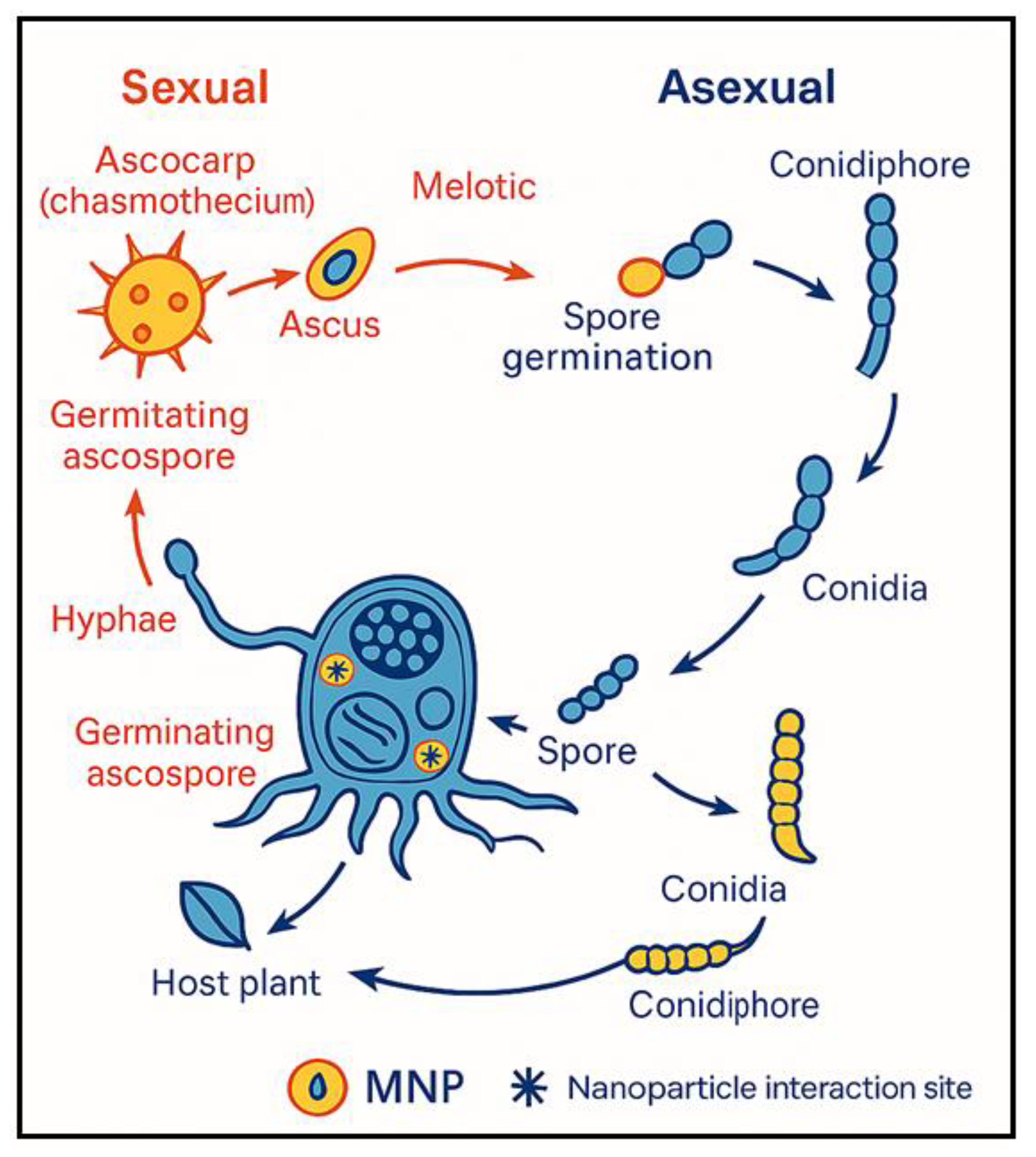
Powdery mildew, caused by obligate biotrophic fungi such as Erysiphe, Golovinomyces, and Podosphaera, poses a persistent threat to leafy crops including lettuce, spinach, arugula, and melon. Powdery mildew fungi have both asexual and sexual stages (Figure 1). In the asexual phase, a conidium lands on a susceptible host, germinates, and develops an appressorium that penetrates the plant’s cuticle and cell wall but leaves the plasma membrane intact [27]. The appressorium’s turgor pressure and lytic enzymes allow a hyphal peg to enter the epidermal cell and form a primary haustorium to extract nutrients [28]. Successful infections lead to branching hyphae and the emergence of conidiophores, which generate conidia in numbers that vary by genus [29,30]. After several days, these fungal colonies appear as white spots on plant surfaces, indicating infection. During sexual reproduction, two compatible hyphae join to produce a chasmothecium, a fruiting structure that holds one or more asci containing sexual spores (ascospores), depending on the genus [29]. Chasmothecia play a significant epidemiological role in certain species like grape powdery mildew (Erysiphe necator) [31], but are rarely or never seen in others, such as cucurbit powdery mildew (Podosphaera xanthii), leaving the epidemiological impact of their sexual cycle uncertain [32]. The disease is characterized by white, powdery growth on foliage, leading to chlorosis, premature senescence, and substantial yield and quality losses [1]. Pathogen development begins with wind-dispersed conidia that germinate under moderate temperatures and high humidity, forming appressoria to penetrate the plant epidermis. Haustoria then extract nutrients, enabling extensive mycelial growth and repeated cycles of conidia production, which perpetuate infection throughout the season [33].
Powdery mildew fungi can reproduce rapidly, especially in monoculture systems with limited genetic diversity. Notably, the pathogen does not require free water for germination, thriving in dry climates with high relative humidity, and often evading conventional systemic fungicides due to its superficial colonization [21]. For effective control, interventions must target early infection stages, particularly spore germination and appressorium formation [14,21]. Epidemics are driven by environmental factors like moderate temperatures (15–27°C), elevated humidity (above 60%), and poor ventilation that favor rapid disease spread. In leafy vegetables, where foliage is the main commodity, the resulting symptoms translate directly to economic losses [34]. A comprehensive understanding of the biology and epidemiology of powdery mildew is essential for developing advanced and sustainable control strategies.
3. Nanoparticles and Nanosuspensions Applications on Powdery Mildew
Products of nanotechnology including nanoparticles and nanosuspensions have emerged as innovative and promising approaches for controlling powdery mildew in agricultural crops [35,36,37]. Unlike traditional pesticides, the minuscule size of nanoparticles typically ranging from 1 to 100 nm grants them increased surface reactivity. This unique property allows for more efficient delivery and targeted action against pathogens, which in turn means lower doses of active agents are needed. By incorporating nanoparticles or dispersing them as nanosuspensions, researchers have been able to enhance the solubility and stability of fungicides, further improving their efficacy and minimizing their environmental footprint [38]. Moreover, nanoparticles can be engineered to carry active ingredients, such as fungicides or plant defense inducers, and release them in a controlled manner. This targeted delivery reduces the required dosage and minimizes off-target effects [39]. Nanosuspensions improve the solubility, stability, and targeted delivery of fungicides, enabling better adhesion to plant surfaces, penetration into fungal structures, and controlled release for sustained action and fewer applications [39]. Application methods include foliar sprays, soil drenching, and encapsulated slow-release formulations. Studies show sulfur nanosuspensions provide enhanced retention and antifungal efficacy, while copper-based nanosuspensions in grapevines improve uptake and disease control. Nano-formulations also increase the bioavailability of active ingredients and display intrinsic antifungal activities, such as ROS generation, membrane disruption, and gene expression modulation [39]. These advanced formulations can target critical points in the pathogen’s lifecycle, work in synergy with existing control strategies, and potentially reduce the environmental impact associated with conventional fungicides [37]. Extensive research demonstrates the applications of different metallic, non-metallic and polymeric nanoparticles and/or nano-formulations against powdery mildew and enhancing crop quality [37,40,41].
3.1. Metallic Nanoparticles (MNPs) Effect on Powdery Mildew
Metal-based nanoparticles (MNPs) such as silver (AgNPs), copper (CuNPs), and zinc oxide (ZnONPs) have emerged as potent antifungal agents due to their unique physicochemical properties (Table 1). These nanoparticles exhibit high surface area-to-volume ratios, enabling enhanced interaction with fungal cells. Numerous field and greenhouse studies have validated the effectiveness of nano-based strategies against powdery mildew. These case studies highlight the practical benefits and real-world applicability of the innovations discussed. These findings demonstrate that nano-based treatments are not only effective but also safe and environmentally compatible. They offer a viable alternative to traditional fungicides, especially in high-value crops where disease control and residue management are critical.
Recent investigations have highlighted the efficacy of AgNPs as foliar sprays in the management of powdery mildew; for instance, application to cucumber plants infected with Podosphaera xanthii has led to marked reductions in disease severity, enhanced foliar health, and increased yields, with microscopy confirming membrane disruption and ROS-mediated cellular damage in the pathogen [42]. In controlled greenhouse trials, neem-extract-synthesized AgNPs applied to zucchini plants infected by Podosphaera xanthii reduced disease severity by over 70% and promoted plant vigor without observable phytotoxic effects [35]. Furthermore, foliar application of AgNPs at concentrations of 100 ppm has been shown to significantly inhibit powdery mildew in cucurbits (cucumber and pumpkin), impeding both hyphal development and conidial germination [35]. Studies also have further corroborated the potential of AgNPs as highly effective agents in the management of powdery mildew. For example, Vizitiu et al. [40] showed that AgNPs synthesized from Dryopteris filix-mas extract, as well as the extract itself, serve as promising alternatives for controlling Uncinula necator in vineyards. The antifungal efficacy of AgNPs is attributed to their ability to penetrate fungal cell walls and membranes, resulting in structural damage and the leakage of cellular contents, while also catalyzing the generation of ROS that induce oxidative stress and programmed cell death in fungal cells [41]. Empirical studies have reported that foliar application of AgNPs can reduce the severity of Podosphaera xanthii infections in cucurbits by up to 90%, with minimal observed phytotoxicity [8,41]. Collectively, these findings underscore the value of AgNPs as safe and potent tools for the sustainable management of powdery mildew pathogens.
Copper-based nanoparticles, such as copper nanoparticles (CuNPs) and nanosuspensions, have revealed potent antifungal activity against powdery mildew pathogens by interacting with fungal enzymes and disrupting essential metabolic pathways. Their application in grapevines led to improved control of Erysiphe necator, with studies reporting superior adhesion to leaf surfaces, enhanced uptake, and prolonged protection compared to conventional copper formulations [39]. In this case, notably, nano-copper treatments sustained disease suppression for up to 21 days, exceeding the efficacy period of traditional copper sprays, and contributed to higher chlorophyll content and improved fruit quality. Besides, copper-based fungicides, including those containing nanoparticles, remain among the most effective biorational options for managing powdery mildew in crops like winter squash, outperforming many biological and botanical alternatives [43].
ZnO nanoparticles (ZnONPs) have demonstrated robust antifungal activity, notably inhibiting spore germination and hyphal development through mechanisms involving ROS generation and membrane disruption. Their deployment in tomato and pepper systems has resulted in marked declines in powdery mildew prevalence, while concomitantly enhancing chlorophyll concentration and overall plant vigor, thus highlighting their dual functionality as anti-fungal agents and growth promoters [35,37,44]. Investigations into ZnONPs applied to tomato plants revealed induction of pathogenesis-related proteins and antioxidant synthesis, evidencing strengthened plant defense responses. Comparative analyses of green-synthesized and chemically synthesized ZnO NPs in pepper have reported antifungal efficacy ranging from 78.74% to 82.45% at 300 mg/L, alongside augmented pigment levels [37,44]. Further, greenhouse trials incorporating ZnO and MgO nanoparticles at 200 mg/L in pepper plants achieved significant reductions in disease severity, which correlated with elevated chlorophyll and polyphenol oxidase activity; cytotoxicity assessments indicated no substantial alterations in mitotic index, supporting the safety of these formulations as alternatives to standard fungicides such as penconazole [45].

Table 1.
Metallic nanoparticles size, concentration and effectiveness on powdery mildew treatment Where GS: green synthesis, CS=chemical synthesis.
Table 1.
Metallic nanoparticles size, concentration and effectiveness on powdery mildew treatment Where GS: green synthesis, CS=chemical synthesis.
| Nanoparticle Type |
Crop | Size (nm) | Concentration | Synthesis Method | Application Method | Effectiveness | Additional Benefits | Reference |
|---|---|---|---|---|---|---|---|---|
| Ag | Eggplant | 7–25 | 10–100 ppm | CS | Foliar | Effective in reducing powdery mildew | SEM: Mycelial and spore deformation | [35] |
| Ag | Beans | ~25 | 10–100 ppm | CS and GS | Foliar | Effective against powdery mildew and Botrytis cinerea | ↓ Disease incidence, ↑ yield potential | [35] |
| Ag | Melons | 7–25 | 10–100 ppm | CS | Foliar | 100 ppm: 20% disease incidence | SEM: Spore deformation, ↑ efficacy pre-infection | [35] |
| Ag | Radish | 7–25 | 10–100 ppm | CS | Foliar | Effective in reducing powdery mildew (extrapolated) | SEM: Spore deformation, safe for leafy crops | [35] |
| Ag | cucumber & pumpkin | ~10–50 | 100 ppm | GS | Foliar | Highest inhibition rate | Damages fungal hyphae and conidia; SEM confirmed effects | [35] |
| Ag | Grapevine | ~20–23 | ND | GS | Foliar | Improved control of E. necator | Superior leaf adhesion, enhanced uptake, prolonged protection vs. copper formulations | [39] |
| Ag | Grapevine | ~17 | crude | GS | Foliar | Protective effect against Uncinula necator | enhanced sugar, starch, water content; increased shoot length and grape yield | [40] |
| Cu | Squash | ~40–60 | ~300 mg·L⁻¹ | CS | Foliar | Highest among tested | Outperformed biological and botanical alternatives; consistent disease suppression | [43] |
| ZnO | Tomato, Pepper | ~40 | 50–250 mg·L⁻¹ | CS | Foliar & Soil | Significant reduction in powdery mildew | Enhanced chlorophyll, lycopene, β-carotene, sugar content; reduced oxidative stress (MDA) | [44] |
| ZnO | Pepper | 79.5 | 100, 150, 200 mg·L⁻¹ | CS | Foliar | Significant reduction in disease severity | Increased chlorophyll; no substantial cytotoxicity (mitotic index unaffected); alternative to penconazole | [45] |
| MgO | Pepper | 53 | 100, 150, 200 mg·L⁻¹ |
CS |
Foliar | Significant reduction in disease severity | Increased chlorophyll; no substantial cytotoxicity (mitotic index unaffected); alternative to penconazole | [45] |
| Fe₃O₄ | Lettuce | ~20–50 | ~200 mg·L⁻¹ | CS | Foliar | Significant reduction in disease severity | Increased chlorophyll, carotene, phenolics, protein; elevated CAT and PPO enzyme activity | [46] |
| Se | Melons | ~50–100 | 25–75 mg·L⁻¹ | CS | Foliar | ~21–45% reduction | Enhances antioxidant enzymes; alters polyamine, phenylpropanoid, and hormone pathways | [47] |
| Se | Various crops | ~50–100 | 25–100 mg·L⁻¹ | GS | Foliar | High antifungal activity; effective against resistant strains | Antioxidant, biocompatible, low toxicity; safe fungicide alternative | [48] |
| Ag, ZnO, TiO₂ | Tomato | 10–100 | Varies | GS | Foliar | Effective against fungal & insect pests | ↑ Photosynthesis | [49] |
| CuO and ZnO | Mustard | ~50–80 | 100–300 mg·L⁻¹ | GS | Foliar | Promising antifungal activity | Eco-friendly alternative to fungicides | [50] |
| TiO₂ | Spinach | ~20 | 50–100 mg·L⁻¹ | Sol-gel/GS | Foliar | ↑ Photosynthesis, ↓ fungal stress | ↑ Biomass, ↑ Chlorophyll content | [50] |
| CuO | Lettuce | 230–400 | 100 mg·L⁻¹ | GS | Foliar | ↓ Fungal colonization | ↑ Leaf health, ↓ oil evaporation | [50] |
| Ag and ZnO | Grapes | 10–50 | 50–200 ppm | GS | Foliar | Effective against Erysiphe necator | ↑ Fruit quality, ↓ chemical residues | [50] |
| ZnO and CuO | Oranges | 20–80 | 100–300 ppm | GS | Foliar | Antifungal & antibacterial | ↑ Shelf life, ↑ Disease resistance | [50] |
| ZnO | Beetroot (Sugar beet) | — | 10, 50, 100 ppm | Engineered | Foliar | ↓ Disease severity, ↑ chlorophyll, ↑ PPO & POD | Induced resistance via ROS and phenolics | [51] |
| Ag | Cucurbits | ~10–50 | 25–100 mg·L⁻¹ | CS/GS | Foliar | Up to 90% reduction | Minimal phytotoxicity; eco-friendly; strong antifungal activity | [52] |
In a recent comparative study, Masoud et al. [46] assessed the efficacy of Fe₃O₄ nanoparticles, yeast, and Bio-Arc alongside conventional fungicides such as azoxystrobin and penconazole for the management of powdery mildew in lettuce. While azoxystrobin exhibited the highest level of disease suppression, applications of Fe₃O₄ nanoparticles and yeast also resulted in significant reductions in disease severity and notable improvements in key growth parameters. Furthermore, treatments were associated with enhanced chlorophyll, protein, and phenolic contents, as well as increased activities of catalase and polyphenol oxidase, collectively supporting the potential of biocontrol agents and nanoparticle applications as safe and effective alternatives for powdery mildew management in lettuce production systems [46].
Selenium nanoparticles (SeNPs) have emerged as potent antifungal agents, with applications in melon demonstrating significant decreases in powdery mildew incidence and notable enhancements in both fresh and dry biomass [47]. In this study, SeNPs treatments at 5 mg/L yielded disease index reductions between 21–45%, attributed to simultaneous activation of host defense pathways and improved plant growth parameters. These authors also presented mechanistic analogies and cross-crop efficacy observed in melon and cucurbits suggest promising prospects for future deployment of SeNPs. Biologically synthesized SeNPs particularly via Trichoderma-mediated processes exhibit broad-spectrum antifungal activity, including potent suppression of powdery mildew [12]. Critically, the antifungal effectiveness of SeNPs is strongly influenced by particle size, with smaller nanoparticles demonstrating enhanced inhibition of fungal proliferation, sporulation, and zoospore viability. This inverse relationship underscores the value of SeNPs as advanced alternatives or adjuncts to conventional fungicide regimes, especially in scenarios characterized by fungicide resistance among powdery mildew pathogens [48].
3.2. Non-Metallic Nanoparticles (NMNPs) Effect on Powdery Mildew
Farhat et al. [53] highlighted the substantial efficacy of silicon nanoparticles (SiNPs) biosynthesized by Pseudomonas putida and Trichoderma harzianum for powdery mildew suppression with greenhouse trials reporting up to 93.5% disease reduction. Similarly, Elsharkawy et al. 54] observed that nanosilica at 500 mg/L matched the effectiveness of penconazole in mitigating rose powdery mildew, while also enhancing vegetative growth, floral attributes, and vase longevity. Microscopy revealed marked ultrastructural hyphal damage, accompanied by upregulation of the defense-associated gene PAL. In parallel, Rashad et al. [55] reported that foliar application of SiNPs at 150 ppm curtailed mildew in grapevines by up to 81.5% and improved both yield and berry quality. This intervention was associated with increased expression of defense-related genes, including β-1,3-glucanase and peroxidase, and improved physiological indices such as chlorophyll content and antioxidant enzyme activity. Notably, higher nanoparticle concentrations elicited cytotoxic and genotoxic responses, underscoring the necessity for comprehensive safety evaluation.
Foliar application of sulfur nanoparticles (SNPs) at 100 ppm to mango cv. Keitt resulted in a 14.6% reduction in powdery mildew incidence and a remarkable 342% increase in productivity, alongside improved fruit quality and upregulated antioxidant enzyme activity [56]. However, raising the concentration to 500 ppm induced phytotoxic symptoms and diminished yield, underscoring the necessity for precise dosage optimization. Efficacy trials by Gogoi et al. [56] with SNPs against Erysiphe cichoracearum in okra revealed that the nano-formulation outperformed commercial sulfur products, substantially inhibiting conidial germination and cleistothecial integrity while exhibiting minimal phytotoxicity being fourfold more effective than conventional alternatives. Abdel-Rahman and Alkolaly [57] compared SNPs (12.2–23.5 nm), bulk sulfur, and systemic fungicides (azoxystrobin, diniconazole) in cucumber powdery mildew management, finding that SNPs delivered superior disease suppression relative to bulk sulfur and matched the efficacy of systemic fungicides, while achieving the greatest fruit yield and quality. Residue analyses in this study also confirmed SNPs treated produce was safe for harvest within two days post-application, supporting its suitability as an eco-friendly intervention. Collectively, these findings establish SNPs formulations as potent, safe, and sustainable alternatives for powdery mildew control [56]. Overall, these studies highlight the promise of biosynthesized nanomaterials as sustainable and environmentally benign alternatives to conventional treatment of powdery mildew.
3.3. Nano-Encapsulated Fungicides and Essential Oils
Recent advancements in polymeric and lipid-based nanocarriers have revolutionized fungicide delivery systems, enabling precise, environmentally responsive release and superior bioavailability [58]. Encapsulation within nanoscale matrices offers a dynamic approach for enhancing efficacy and reducing ecological risk, particularly in crop protection [59]. For example, Ruano-Rosa et al. [60] demonstrated the utility of chitosan oligomer-based bioformulations, incorporating Streptomyces spp. metabolites and hydrolyzed gluten, in mitigating powdery mildew under both field and laboratory conditions. Soleimani et al. [16] further expanded this paradigm by encapsulating celery seed essential oil (CSEO) in chitosan nanoparticles, achieving significant reductions in cucumber powdery mildew, improved chlorophyll and phenolic content, and pronounced activation of defense-related genes. The release profile adhered to Fickian diffusion, ensuring sustained antifungal activity. Likewise, chitosan-encapsulated spinach seed essential oil nanoparticles (SSEO-LCNPs) exhibited high encapsulation efficiency and spherical structure; at 400 µg/mL, these nanoparticles markedly curtailed disease severity while stimulating systemic resistance pathways, evidenced by elevated phenolic and flavonoid levels and enhanced antioxidant enzyme activity [61]. These findings affirm the efficacy of biopesticide-loaded nanocarriers as sustainable alternatives to conventional synthetic fungicides.
By integrating essential oils with synthetic fungicides within nanoscale carriers, researchers have also achieved synergistic antifungal effects that disrupt multiple fungal metabolic pathways. Commercial innovations such as Nutragreen® have enabled substantial reductions in pesticide use, and across numerous studies, nano-formulated agents have consistently delivered 70–90% decreases in powdery mildew severity [36]. Moreover, nano-formulated thyme oil has exhibited robust suppression of Erysiphe cichoracearum in lettuce without adversely impacting beneficial microbial communities [62].
Leveraging nanomaterials for encapsulating volatile plant-derived compounds represents a promising trajectory in crop protection, effectively mitigating traditional limitations of instability and rapid dissipation [5,63,64]. Nevertheless, the search for sustainable, biodegradable carrier materials remains a critical consideration. Chitosan, noted for its antioxidant properties, biocompatibility, and encapsulation capacity, has garnered extensive interest in bioactive delivery in multiple nanoscale formulations [63,65]. Chitosan-coated AgNPs demonstrate enhanced foliar retention and sustained Ag⁺ release, conferring extended antifungal efficacy at reduced dosages compared to uncoated counterparts [66]. Also, empirical evidence substantiates the efficacy of essential oil encapsulation in chitosan nanoparticles for antifungal applications [64,67,68,69,70]. Soleimani et al. [16] evaluated the performance of CSEO-loaded chitosan nanoparticles (approx. 113 nm diameter) against Podosphaera fusca in cucumber, observing significant suppression of disease severity and robust induction of host defense mechanisms, as indicated by increased phenolic and flavonoid synthesis and activation of defense-related enzymes. Gene expression analysis further revealed upregulation of resistance pathways, highlighting the potential of CSEO-LCNPs as eco-friendly alternatives in powdery mildew management.
Table 2.
Nonmetallic Nanoparticles for Powdery Mildew Control in Fruits and Vegetables. NS = not specified; CS = Chemical synthesis.
Table 2.
Nonmetallic Nanoparticles for Powdery Mildew Control in Fruits and Vegetables. NS = not specified; CS = Chemical synthesis.
| Nanoparticle | Crop Name | Size (nm) | Concentration / Dose | Synthesis Method | Application Method | Effectiveness | Additional Benefits | References |
|---|---|---|---|---|---|---|---|---|
| Chitosan + CSEO | Cucumber | 300 | 1–3 mg·mL⁻¹ | Ionic gelation + encapsulation | Foliar | Significant reduction in powdery mildew severity | Increased chlorophyll, phenolics, flavonoids, defense enzyme activity, and gene expression | [16] |
| Nutragreen® nanoscale carrier | Hop | NS | 30% v/v | CS | Foliar | ~70–90% reduction in powdery mildew severity | Reduced pesticide use; improved cone yield and α-acid content; enhanced leaf and cone protection | [36] |
| Sulfur | Okra | 50–90 | 1000 ppm | Liquid synthesis | Foliar | 100% inhibition of conidial germination | Disrupted cleistothecia, reduced phytotoxicity | [56] |
| Sulfur | Apple | 50–90 | 1000 ppm | Liquid synthesis | Foliar | Effective at lower doses, avoids phytotoxicity | Safer than conventional sulfur for fruit crops | [56] |
| Sulfur | Mango | 85 | 100ppm | CS | Foliar | 14.6% reduction in powdery mildew | 342% increase in productivity; improved fruit quality (TSS, vitamin C); enhanced POD & PPO enzyme activity | [56] |
| Sulfur | Cucumber | 12.2–23.5 | 500 mg·L⁻¹ | CS | Foliar | 60.9% reduction in powdery mildew | Matched azoxystrobin (74%) and diniconazole (68.8%) in efficacy; highest fruit yield and quality | [57] |
| Chitosan oligomers + Streptomyces metabolites / hydrolyzed gluten | Grapevine | <2 kDa | ~40 mL | Enzymatic hydrolysis / fermentation | Foliar & root | Comparable to conventional fungicides | Effective against Erysiphe necator; biostimulant effects; reduced overwintering inoculum | [60] |
| Chitosan NPs + SSEO | Cucumber | ~116.2 | 400 µg·mL⁻¹ | Ionic gelation + encapsulation | Foliar | Significant reduction in powdery mildew severity | High encapsulation efficiency; spherical morphology; elevated phenolics, flavonoids, POD, PPO, PAL activity | [61] |
| Thyme oil nanoemulsion | Lettuce | ~83 | 10% (v/v) | Ultrasonic emulsification | Foliar | Maintains beneficial microbes; stable for >3 months; effective even when diluted | [62] | |
| Chitosan | Cucumber | 150–250 | 0.1–0.2% (w/v) | Ionic gelation | Foliar | ~70% disease reduction | Induces SAR; enhances chlorophyll and defense enzymes | [74] |
| Chitosan | Tomato | 20-100 | 0.1–1%(w/v) | Ionic gelation | Foliar | Effective against powdery mildew at early stage | Induces systemic resistance, enhances growth | [75] |
| Chitosan | Cucumber | 20-100 | 0.1–1%(w/v) | Emulsion cross-linking | Foliar | Effective against fungal pathogens including powdery mildew | Improved resistance, growth promotion | [75] |
| SiO₂- | Grape | 50–80 | 50–100 mg L⁻¹ | Sol–gel | Foliar | 85–90% reduction | Strengthens epidermis; Si-mediated resistance | [76] |
| SiO₂ | Cucumber | 40–60 | 50 mg L⁻¹ | Sol–gel | Foliar | 80–90% mildew reduction | Reinforces cuticle; nontoxic | [76] |
| SiO₂ | Watermelon | Mesoporous SNPs | NS | Modified Stöber method | Root dip | 40% reduction in disease severity | Downregulation of stress genes (CSD1, PAO, PPO, RAN1) | [77] |
| SiO₂ | Cucumber | 10-100 | NS | Sol–gel, GS | Foliar | High efficacy | Improved photosynthesis, enzyme activity, stomatal conductance | [77] |
| SiO₂ | Cucumber | 10-100 | 1.7–56 mM | Sol–gel | Foliar | Up to 87% reduction in powdery mildew | Improved resistance, structural strength | [78] |
| Silica–alginate nanocomposite | Pumpkin | 70–150 | 25–75 mg L⁻¹ | Sol–gel + biopolymer | Foliar | 80% mildew control | Reinforces epidermis; water balance | [79] |
| Silica–chitosan | Spinach | 80–150 | 0.1% (w/v) | Sol–gel + ionic gelation | Foliar | 65% infection reduction | Improves leaf turgidity; safe | [79] |
| Silica–pectin | Apple | 40–90 | 50 mg L⁻¹ | Sol–gel hybridization | Foliar | 75% reduction | Biodegradable; strengthens cuticle | [80] |
| Silica–pectin | Peach | 40–100 | 50 mg L⁻¹ | Sol–gel | Foliar | 78% mildew suppression | Strengthens fruit epidermis | [80] |
| Carbon nanotubes | Tomato | 10–40 | 10–25 mg L⁻¹ | CS | Foliar | ~55% mildew reduction | Boosts antioxidants | [81] |
| Graphene oxide nanosheets (GO) | Cucumber | 30–200 | 25–50 mg L⁻¹ | Modified Hummers | Foliar | ~60% reduction | Activates enzymes; nutrient uptake | [82] |
| Nano-encapsulated lemongrass EO (Alginate) | Strawberry | 150–300 | 2–4 mg mL⁻¹ | Emulsion crosslinking | Foliar | ~85% infection reduction | Antioxidant; flavor-safe | [83] |
| Nanobubble water | Papaya | 70–130 | 5×10⁸ to 5×10⁹ bubbles/mL | Nanobubble generator | Foliar | Effective against powdery mildew (patented method) | Non-toxic, enhances root zone health | [84] |
Overall, comparing encapsulated and non-encapsulated nanoformulations for powdery mildew prevention reveal distinct advantages and limitations for each approach, depending on environmental conditions and application goals. Encapsulated nanoformulations are widely recognized for their ability to provide controlled release, targeted delivery, and enhanced stability of active ingredients [71,72]. In contrast, non-encapsulated nanoformulations, such as AgNPs, offer rapid antifungal effects but are more susceptible to environmental degradation [35]. Additionally, Adnan and Waleed [73] found that biologically synthesized copper oxide nanoparticles achieved 81.71% effectiveness against powdery mildew in roses under greenhouse conditions, suggesting that non-encapsulated forms may be suitable for controlled environments [73].
4. Mechanisms of Action Against Powdery Mildew
4.1. Direct Antifungal Effects
Metallic nanoparticles (MNPs), notably, have emerged as potent antifungal agents against powdery mildew, acting through coordinated mechanisms that include cell membrane disruption, cell wall perturbation, and the induction of oxidative stress via reactive oxygen species (ROS) [8,22,85]. These nanoparticles possess high surface reactivity and positive charge, facilitating electrostatic interactions with fungal membrane phospholipids and proteins. Such interactions compromise membrane architecture, promoting increased permeability, leakage of intracellular solutes, and ultimately, cell lysis. Transmission and scanning electron microscopy (SEM) studies revealed rapid spore collapse, hyphal distortion, membrane thinning, and pore formation, underscoring their acute impact on fungal morphology [35,42] and rendering them highly vulnerable to osmotic stress and host defenses. Complementing these effects, nanoparticles interfere with fungal cell wall integrity by binding to chitin, β-glucans, and glycoproteins, as well as inhibiting enzymes critical for wall maintenance, such as chitin synthase and glucanase [85].
The generation of ROS such as superoxide anions, hydroxyl radicals, and hydrogen peroxide by metal and metal oxide nanoparticles further exacerbates cellular damage by attacking membrane lipids, cytoplasmic proteins, and nucleic acids. This multifaceted oxidative assault leads to denaturation of enzymes, DNA fragmentation, and overall impairment of vital metabolic processes. Notably, ROS production is dose-dependent and can be amplified by environmental factors such as light and pH, increasing antifungal potency [22,85].
Spore germination and hyphal elongation are pivotal events in the powdery mildew fungal lifecycle, underpinning host colonization and resource acquisition. Nanoparticle interventions disrupt these processes through several sophisticated mechanisms. Primarily, nanoparticles can establish a physical barrier on spore surfaces, impeding essential gas and nutrient exchanges necessary for germination, a phenomenon documented with silica and titanium dioxide nanomaterials. Furthermore, nanoparticle-induced ROS perturb critical intracellular signaling, notably calcium flux and ATP production, thereby impairing fungal development [53]. Experimental data indicate that ZnO and CuO nanoparticles markedly reduce spore germination rates in pathogens such as Erysiphe necator and Podosphaera xanthii; treated spores display aberrant morphology, delayed germ tube emergence, and diminished viability [22]. These inhibitory effects manifest as reduced infection success and disease severity in plants. Importantly, by targeting the incipient stages of pathogen establishment, nanoparticle-based strategies provide a robust preventive modality, complementing curative interventions and reinforcing integrated disease management paradigms.
4.2. Indirect Effects: Induced Resistance and Gene Regulation
Nanoparticles exert multifaceted effects against powdery mildew, extending beyond physical damage to include transcriptional and transcriptomic modulation. Exposure to metal-based nanoparticles such as AgNPs and CuO NPs triggers pronounced alterations in fungal gene expression, notably upregulating stress response genes (e.g., catalase and superoxide dismutase) while concurrently repressing those implicated in cell wall biosynthesis, ergosterol production, hyphal elongation, and efflux pump activity [85]. Such transcriptomic interference destabilizes core processes essential for fungal growth, reproduction, and virulence, as demonstrated by the suppression of chitin synthase and β-glucan synthase in Blumeria graminis upon CuO NPs treatment. This gene-level disruption not only impairs pathogen development but also diminishes the likelihood of resistance emerging by targeting regulatory networks rather than single metabolic pathways. Additionally, nanoparticle-based modalities, exemplified by SeNPs, can potentiate host plant defenses through coordinated activation of antioxidant enzymes, enhancement of polyamine levels (putrescine, spermine), and upregulation of genes within phenylpropanoid and hormone signaling cascades [47].
4.3. Synergy with Conventional Fungicides
The integration of nanoparticles with conventional fungicides presents a promising strategy for managing powdery mildew, a widespread and economically damaging fungal disease. When combined with traditional fungicides, these nanoparticles enhance the efficacy of disease control by improving solubility, bioavailability, and targeted delivery, while reducing the required chemical dosage and environmental toxicity [86]. For instance, AgNPs amplify membrane-targeted fungicide activity, whereas SeNPs augments systemic plant defenses; together, these modalities enable robust disease suppression while diminishing overall chemical inputs [66]. The integration of nanoparticles in agriculture is particularly compelling due to their capacity to enhance the performance of traditional fungicides via complementary mechanisms of action. Such synergy addresses challenges like limited solubility, resistance emergence, and environmental toxicity. Empirical research confirms that copper-based nanoparticles, when combined with azole fungicides, significantly lower the minimum inhibitory concentration necessary for effective targeting of Erysiphe necator. Similarly, the co-application of AgNPs with sulfur-based fungicides yields superior suppression of powdery mildew in cucurbits [22]. Combining nanoparticles with natural extracts or essential oils (e.g., CSEO-LCNPs) enhances disease control and stimulates plant defense mechanisms [16]. Their integration with nanoparticles or oils could further improve efficacy [87]. These synergistic approaches not only improve therapeutic efficacy but also align with integrated pest management (IPM) principles by reducing reliance on chemical inputs and promoting environmental sustainability. Nano-based interventions can be seamlessly incorporated into IPM frameworks by serving as preventive agents, complementing resistant cultivars and cultural controls, minimizing fungicide application frequency through synergy, and ensuring ecological safety through continuous assessment of phytotoxicity and environmental persistence.
5. Challenges and Limitations
Nanoparticles, particularly those composed of metals such as AgNPs, CuNPs, and ZnONPs, present notable ecotoxicological risks for non-target organisms [88]. Their ability to generate ROS may compromise both plant tissues and beneficial soil microorganisms, thereby perturbing ecosystem equilibrium. Environmental persistence further exacerbates concerns regarding bioaccumulation and the progressive degradation of soil health [88]. The pronounced toxicity associated with nanoparticle exposure is largely attributable to their diminutive dimensions and heightened surface reactivity, which facilitate complex, and often unintended, biological interactions [89]. A major physicochemical challenge is the propensity of nanoparticles to aggregate, driven by elevated surface energy, resulting in diminished effective surface area and altered functionalities. Such aggregation undermines antifungal performance and complicates formulation and storage protocols. To counteract this, stabilizers and surfactants are routinely employed, yet these additives may introduce supplementary toxicity or adversely affect delivery mechanisms [90]. Additionally, biological instability manifested through protein adsorption and enzymatic degradation can hasten clearance and curtail therapeutic efficacy [91]. Although recommended application rates typically avoid acute toxicity, the cumulative impacts of chronic exposure and high-dosage scenarios demand comprehensive investigation [21,37,92]. To mitigate these risks, research into biodegradable, plant-derived carriers including chitosan and lignin-based systems is advancing. Controlled-release formulations, such as hydrogels or polymeric matrices, not only enhance persistence and prophylactic efficacy but also minimize off-target dispersion [35].
While laboratory synthesis of nanosuspensions is well established, scaling these protocols for agricultural deployment demands intricate, resource-intensive processes, including high-pressure homogenization and wet milling [88]. The necessity for precise particle size control, combined with the substantial cost of excipients and solvents, can render such applications prohibitive to smaller agricultural operations [90]. Optimizing nanosuspension formulations necessitates stringent management of particle size distribution, zeta potential, and crystallinity as these parameters susceptible to fluctuation during scale-up. Advanced preservation techniques, notably lyophilization and spray drying, are frequently required to maintain particle stability, increasing both operational complexity and cost [90]. The efficient removal of residual solvents and surfactants remains an unresolved technical bottleneck.
Regulatory oversight of nanopesticides remains incipient, with limited harmonization in protocols evaluating safety, efficacy, and environmental ramifications. This regulatory ambiguity impedes timely product approval and market integration, stalling widescale adoption [88]. Moreover, public apprehension rooted in perceived health risks and opaque technological processes compounds regulatory inertia [25].
Attaining high-fidelity targeting powdery mildew pathogens, while preserving host plant integrity and safeguarding beneficial microbiota, poses a significant technical challenge. Controlled release modalities remain suboptimal, often yielding inconsistent dosing and potentiating pathogen resistance [25]. Although surface engineering strategies, such as PEGylation and ligand conjugation, show promise for enhanced specificity, their translation to agronomic settings is still under development [93].
Nanoparticles interact with plant metabolic networks in multifaceted and sometimes unpredictable manners. While certain formulations can induce systemic resistance, others may disrupt essential physiological processes including nutrient assimilation, photosynthesis, and hormonal signaling. The chronic effects of repeated nanoparticle exposure are insufficiently elucidated and necessitate comprehensive field validation and toxicological assessment [25]. Such interactions are highly contingent upon the physicochemical attributes of the nanoparticles, necessitating tailored, crop-specific investigations.
6. Future Perspectives and Directions
The intersection of nanotechnology and precision agriculture has ushered in sophisticated modalities for controlling powdery mildew across diverse cropping systems. Smart nanoparticles, which respond to environmental stimuli such as pH, temperature, and pathogen presence, are redefining targeted fungicide delivery, enabling precise temporal and spatial disease intervention. This technological leap is complemented by biodegradable carriers derived from natural polymers like chitosan, cellulose, and starch, which not only shield plants against powdery mildew but also contribute to improved nutrition and vigor.
Recent advancements in digital agriculture encompassing drone-based multispectral sensing and AI-powered diagnostics are streamlining the deployment and monitoring of nano-formulations [94,95,96]. Such integration facilitates site-specific treatment, optimizes use of antifungal agents, and enhances both efficacy and resource efficiency. Real-time analytics further enable adaptive management, aligning disease control with sustainability imperatives.
Translating laboratory breakthroughs to field environments necessitates robust interdisciplinary collaboration, encompassing plant pathology, chemistry, toxicology, and regulatory science. Rigorous field trials under varied agroecological contexts, combined with harmonized regulatory frameworks, remain pivotal for validating safety, efficacy, and environmental impact [85]. The development of biodegradable stabilizers, optimization of delivery platforms, and comprehensive risk assessments underpin the pathway to widespread adoption.
Emerging nano-enabled strategies offer multifaceted solutions to powdery mildew, a persistent threat exacerbated by climate variability and resistance evolution. Next-generation interventions such as rationally designed multi-site fungicides and genomic-assisted breeding are enhancing host resilience. Innovative tools, including RNA interference and CRISPR-mediated gene editing, promise precise manipulation of host–pathogen interactions, while real-time diagnostics inform strategic fungicide deployment and resistance stewardship.
Smart nano-delivery systems mark a substantial advancement in powdery mildew suppression. These platforms, engineered to release antifungal payloads in response to specific environmental or pathogen-derived signals (e.g., shifts in pH, temperature, or early infection markers), achieve unparalleled precision. pH-responsive nanoparticles target acidic microenvironments at infection loci; temperature-sensitive carriers activate antifungal release under conditions favoring disease proliferation; and photo-triggered systems enable on-demand payload dispersion via controlled light exposure [71].
Additionally, functionalized nanoparticles equipped with ligands for selective binding to pathogen molecules or plant receptors increase delivery specificity, minimizing collateral impacts. For example, chitosan-based nanoparticles conjugated with lectins have demonstrated superior affinity for Blumeria graminis spores, yielding enhanced control over conventional formulations [71].
The convergence of smart nano-formulations with digital agriculture platforms, including drone-assisted application and automated sprayers, is catalyzing a paradigm shift toward precision, sustainability, and regulatory compliance in powdery mildew management. Continued multidisciplinary research, comprehensive field validation, and internationally harmonized guidelines are essential for mainstreaming these innovations and securing future food systems. In sum, the integration of advanced nanotechnological approaches with precision agricultural practices holds significant promise for transformative powdery mildew management, balancing effectiveness with environmental stewardship and regulatory rigor.
7. Conclusions
In conclusion, this review highlights the promising role of nanotechnology in managing powdery mildew across vegetable and fruit crops. Metallic and non-metallic nanoparticles including AgNPs, CuNPs, ZnONPs, SiNPs, and SNPs demonstrate potent antifungal activity through mechanisms such as membrane disruption, ROS generation, and gene regulation, while nano-encapsulated fungicides and essential oils enhance targeted delivery and plant defense activation. These nano-enabled strategies offer synergistic benefits when combined with conventional fungicides, reducing chemical inputs and environmental risks. However, challenges remain, including ecotoxicity, formulation stability, scalability, and regulatory gaps. Future directions emphasize smart delivery systems, biodegradable carriers, and integration with digital agriculture platforms to achieve precision, sustainability, and regulatory compliance. Interdisciplinary research and harmonized safety frameworks are essential to realize the full potential of these innovations in crop protection.
Author Contributions
Conceptualization, A.G, L.C. and A.S.; validation, A.G, L.C. and A.S.; resources, A.G. and L.C..; writing—original draft preparation, A.G.; writing—review and editing, A.G, L.C. and A.S..; visualization, A.G..; supervision. L.C.; project administration, L.C. funding acquisition, A.G. and L.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by United States Department of Agriculture National Institute of Food and Agriculture (USDA-NIFA) Evans-Allen Grant 180835-82601.
Institutional Review Board Statement
Not applicable for this study.
Informed Consent Statement
Not applicable for this study
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
We are grateful to the CAFNR Research in College of Agriculture, Food and Natural resources (CAFNR) at the Cooperative Agriculture Research Center at Prairie View A&M University.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ghalehgolabbehbahani, A.; Zinati, G.; Hamido, S.; Bhusal, N.; Dhakal, M. , Afshar, R. K.; Contina, J. B.; Caetani, R.; Smith, A.; Panday, D. Integrated control of powdery mildew using UV light exposure and OMRI-certified fungicide for greenhouse organic lettuce production. Horticulturae, 2025; 11, 246. [Google Scholar] [CrossRef]
- Rehman, A. U.; Davik, J.; Karisto, P.; Astakhova, T.; Sønsteby, A. A major QTL region associated with powdery mildew resistance in leaves and fruits of the reconstructed garden strawberry. Theoretical and Applied Genetics. 2025; 138, 93. [Google Scholar] [CrossRef]
- Amsalem, L.; Freeman, S.; Rav-David, D.; Nitzani, Y.; Sztejnberg, A.; Pertot, I.; Elad, Y. Effect of climatic factors on powdery mildew caused by Sphaerotheca macularis f. sp. fragariae on strawberry. European Journal of Plant Pathology, 2006, 114, 283–292. [Google Scholar] [CrossRef]
- McGrath, M.T.; Staniszewska, H.; Shishko, N. Fungicide Sensitivity of Sphaerotheca fuliginea populations in the United States. Plant Dis, 1996; 80, 697–703. [Google Scholar]
- Saharan, G. S.; Mehta, N.; Saharan, M. S. (2019). Powdery Mildew Disease of Crucifers: Biology, Ecology and Disease Management (Chapter 3: The Pathogen). Springer Nature Singapore.
- Nie, J.; Yuan, Q.; Zhang, W.; Pan, J. Genetics, resistance mechanism, and breeding of powdery mildew resistance in cucumbers (Cucumis sativus L.). Horticultural Plant Journal, 2023; 9, 603–615. [Google Scholar] [CrossRef]
- Qi, Q.; Fan, C.; Wu, H.; Sun, L.; Cao, C. Preparation of Trichoderma asperellum microcapsules and biocontrol of cucumber powdery mildew. Microbiology Spectrum, 2023; 11, 9e05084-22. [Google Scholar] [CrossRef]
- Elagamey, E.; Abdellatef, M.A.; Haridy, M.S.; Abd El-aziz, E.S.A. Evaluation of natural products and chemical compounds to improve the control strategy against cucumber powdery mildew, Eur. J. Plant Pathol. 2023, 165, 385–400. [Google Scholar] [CrossRef]
- Magyarosy, A. C.; Malkin, R. Photosynthetic impairment in plants infected with powdery mildew. Physiological Plant Pathology, 1978; 13, 183–188. [Google Scholar]
- Jaenisch, J.; Xue, H.; Schläpfer, J.; McGarrigle, E.R.; Louie, K.; Northen, T.R.; Wildermuth, M.C. Powdery mildew infection induces a non-canonical route to storage lipid formation at the expense of host thylakoid lipids to fuel its spore production. 2023. [Google Scholar] [CrossRef]
- Percival, G.C.; Fraser, G.A. The influence of powdery mildew infection on photosynthesis, chlorophyll fluorescence, leaf chlorophyll and carotenoid content of three woody species. Arboric. J. 2002, 26(4):333–347.
- Vielba-Fernández, A.; Polonio, Á.; Ruiz-Jiménez, L.; de Vicente, A.; Pérez-García, A.; Fernández-Ortuño, D. Fungicide resistance in powdery mildew fungi. Microorganisms, 2020; 8, 1431. [Google Scholar] [CrossRef]
- Lin, Z.; Pu, X.; Du, W. The effect of powdery mildew on the cell structure and cell wall composition of red clover. European Journal of Plant Pathology, 2024, 171, 603–614. [Google Scholar] [CrossRef]
- Lu, C.; Du, J.; Chen, H.; Gong, S.; Jin, Y.; Meng, X.; Zhang, T.; Fu, B.; Molnár, I.; Holušová, K.; Said, M.; Xing, L.; Kong, L.; Doležel, J.; Li, G.; Wu, J.; Chen, P.; Cao, A.; Zhang, R. Wheat Pm55 alleles exhibit distinct interactions with an inhibitor to cause different powdery mildew resistance. Nature Communications, 2024; 15. [Google Scholar] [CrossRef]
- Xu, X.; Liu, X.; Yan, Y.; Wang, W.; Gebretsadik, K.; Qi, X.; Xu, Q.; Chen, X. Comparative proteomic analysis of cucumber powdery mildew resistance between a single-segment substitution line and its recurrent parent. Hortic Res 2019, 6:115.
- Soleimani, H.; Reza Mostowfizadeh-Ghalamfarsa, R.; Zarei, A. Chitosan nanoparticle encapsulation of celery seed essential oil: Antifungal activity and defense mechanisms against cucumber powdery mildew. Carbohydrate Polymer Technologies and Applications, 2024, 8, 100531. [Google Scholar] [CrossRef]
- Németh, M. Z.; Kiss, L.; Pintye, A. Environmental factors influencing the efficacy of Ampelomyces quisqualis as a biocontrol agent against powdery mildew. Frontiers in Plant Science, 2023, 14, 1176542. [Google Scholar]
- Gilardi, G.; Gullino, M. L.; Garibaldi, A. Efficacy of biological and chemical products against powdery mildew on zucchini. Journal of Plant Diseases and Protection, 2008; 115, 241–246. [Google Scholar]
- MSU Extension. (2023). Integrated pest management (IPM) for powdery mildew in horticultural crops. Michigan State University (MSU) Extension. https://www.canr.msu.edu/news/integrated-pest-management-ipm-for-powdery-mildew.
- Lebeda, A.; Mieslerová, B. Taxonomy, distribution and biology of lettuce powdery mildew (Golovinomyces cichoracearum sensu stricto). Plant Pathol, 2011; 60, 400-15. [Google Scholar]
- Aldeeb, M.M.E.; Wilar, G.; Suhandi, C.; Elamin, K.M.; Wathoni, N. Nanosuspension-Based Drug Delivery Systems for Topical Applications, International Journal of Nanomedicine 2024, 19, 825-844. [CrossRef]
- Parada, J.; Tortella, G.; Seabra, A. B.; Fincheira, P.; Rubilar, O. Potential antifungal effect of copper oxide nanoparticles combined with fungicides against Botrytis cinerea and Fusarium oxysporum. Antibiotics, 2024,13(3), 215.
- Sangeeta, M. K.; Gunagambhire, V. M.T.; Bhat, M. P.; Nagaraja, S. K.; Gunagambhire, P. V.; Kumar, R. S.; Mahalingam, S. M. In-vitro evaluation of Talaromyces islandicus mediated zinc oxide nanoparticles for antibacterial, anti-inflammatory, bio-pesticidal and seed growth promoting activities. Waste and Biomass Valorization, 2024, 15, 1901–1915. [Google Scholar] [CrossRef]
- Bruno, A.; Tripodi, F.; Armanni, A.; Barbieri, L.; Colombo, A.; Fumagalli, S.; Moukham, H.; Tomaino, G.; Kukushkina, E.; Lorenzi, R.; Marchesi, L.; Monguzzi, A.; Paleari, A.; Ronchi, A.; Secchi, V.; Sironi, L.; Colombo, M. (2024). Advancements in nanosensors for detecting pathogens in healthcare environments. Environmental Science: Nano, 2024, 11(12), 4449–4474. [CrossRef]
- Ly, P-D.; Ly, K-N.; Phan, H-L.; Nguyen, HHT.; Duong, V-A.; Nguyen, H.V. Recent advances in surface decoration of nanoparticles in drug delivery. Front. Nanotechnol. 2024, 6:1456939.
- Xu, Y.; Du, S.; Li, M.; Tan, H.; Sohail, X.; Liu, X.; Qi, X.; Yang, X.; Chen, A. Genome- wide association study reveals molecular mechanism underlying powdery mildew resistance in cucumber, Genome Biol. 2024, 25:252. [CrossRef]
- Eichmann, R.; Hückelhoven, R. Accommodation of powdery mildew fungi in intact plant cells. J. Plant Physiol. 2008, 165, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Tucker, S.L.; Talbot, N.J. Surface attachment and pre-penetration stage development by plant pathogenic fungi. Annu. Rev. Phytopathol. 2002, 39, 385–417. [Google Scholar] [CrossRef]
- Heffer, V.; Johnson, K. B.; Powelson, M. L.; Shishkoff, N. Identification of powdery mildew fungi. The Plant Health Instructor, 2006, 6. [CrossRef]
- Gadoury, D.M.; Cadle-Davidson, L.; Wilcox, W.F.; Dry, I.B.; Seem, R.C.; Milgroom, M.G. Grapevine powdery mildew (Erysiphe necator): A fascinating system for the study of the biology, ecology and epidemiology of an obligate biotroph. Mol. Plant Pathol. 2012, 13, 1–16. [Google Scholar] [CrossRef]
- Pearson, R.C.; Gadoury, D.M. Cleistothecia, the source of primary inoculum for grape powdery mildew in New York. Phytopathology 1987, 77, 1509–1514. [Google Scholar] [CrossRef]
- Pirondi, A.; Vela-Corcía, D.; Dondini, L.; Brunelli, A.; Pérez-García, A.; Collina, M. Genetic diversity analysis of the cucurbit powdery mildew fungus Podosphaera xanthii suggests a clonal population structure. Fungal Biol. 2015, 119, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Polonio, Á; Pérez-García, A.; Martínez-Cruz, J.; Fernández-Ortuño, D.; de Vicente, A. (2020). The haustorium of phytopathogenic fungi: A short overview of a specialized cell of obligate biotrophic plant parasites. In Progress in Botany (Vol. 82, pp. 337–355). Springer. [CrossRef]
- Ma, X.; Wang, Z.; Liu, R.; Jiang, Y. Effect of powdery mildew on interleaf microbial communities and leaf antioxidant enzyme systems. For. Res. 2023, 34:1535–1547. [CrossRef]
- Lamsal, K.; Kim, S. ; Jung, Kim, Y.S.; Kim, K.S.; Lee, Y.S. Inhibition Effects of Silver Nanoparticles against Powdery Mildews on Cucumber and Pumpkin. Mycobiology,2011, 39(1): 26-32. [CrossRef]
- Porteous-Álvarez, A.J.; Maldonado-González, M.M.; Mayo-Prieto, S.; Lorenzana, A.; Paniagua-García, A.I.; Casquero, P.A. Green Strategies of Powdery Mildew Control in Hop: From Organic Products to Nanoscale Carriers. J. Fungi 2021, 7, 490. [Google Scholar] [CrossRef]
- Almusallam, A.H.; Ismail, A.M.; Elshewy, E.S.; El-Gawad, M.E.A. Green synthesized zinc oxide nanoparticles: Antifungal activity against powdery mildew disease of pepper (Capsicum annuum L.) and genotoxicity potentials. Biopestic. Int.; 2024, 20, 193–206. [Google Scholar] [CrossRef]
- Zhu, Z.; Shao, C.; Guo, Y.; Feng, J.; Chen, C.; Yang, H. Facile pathway to generate agrochemical nanosuspensions integrating super-high load, eco-friendly excipients, intensified preparation process, and enhanced potency. Nano Res. 2021, 14, 2179–2187. [Google Scholar] [CrossRef]
- Rossier, B.; Jordan, O.; Allémann, E.; Roderguez-Nogales, C. Nanocrystals and nanosuspensions: an exploration from classic formulations to advanced drug delivery systems. Drug Deliv. and Transl. Res. 2024, 14, 3438–3451. [Google Scholar] [CrossRef]
- Vizitiu, D.E.; Sardarescu, D.I.; Fierascu, I.; Fierascu, R.C.; Soare, L.C.; . Ungureanu, C.; Buciumeanu, E.C.; Guta, I.C.; Pandelea, L.M. Grapevine plants management using natural extracts and phytosynthesized silver nanoparticles. Materials, 2022, 15, 8188. [Google Scholar] [CrossRef]
- Shandila, P.; Mahatmanto, T.; Hsu, J.-L. Metal-Based nanoparticles as nanopesticides: Opportunities and challenges for sustainable Crop Protection. Processes 2025, 13, 1278. [Google Scholar] [CrossRef]
- Ruiz-Jiménez, L.; Polonio, Á.; Vielba-Fernández, A.; Pérez-García, A.; Fernández-Ortuño, D. Gene mining for conserved, non-annotated proteins of Podosphaera xanthii identifies novel target candidates for controlling powdery mildews by spray-induced gene silencing. Journal of Fungi, 2021, 7(9), 735. [CrossRef]
- Indermaur, E.J.; Day, C.T.C.; Dunn-Silver, A.R.; Smar, C.D. Biorational Fungicides to manage cucurbit powdery mildew on winter squash in New York. 2024, 25. [CrossRef]
- Włodarczyk, K.; Smoli´nska, B.; Majak, I. How Nano-ZnO Affect Tomato Fruits (Solanum lycopersicum L.)? Analysis of Selected Fruit Parameters. Int. J. Mol. Sci. 2024, 25, 8522. [Google Scholar] [CrossRef]
- Ismail, A.M.; Abd El-Gawed, M.E. Antifungal activity of MgO and ZnO nanoparticles against powdery mildew of pepper under greenhouse condition. Egypt. J. Agric. Res.; 2021, 4: 421–434.
- Masoud, S.A.; Morsy, S.M.; El-Dein, M.M.Z. Controlling powdery mildew using Fe3O4 NPs, yeast, and Bio-Arc and their effects on the performance of lettuce Lactuca sativa L. Egypt. J. Chem. 2023, 66 (11), 393- 401. [CrossRef]
- Kang, L.; Wu, Y.; Jia, Y.; Chen, Z.; Kang, D.; Zhang, L.; Pan, C. Nano-selenium enhances melon resistance to Podosphaera xanthii by enhancing the antioxidant capacity and promoting alterations in the polyamine, phenylpropanoid and hormone signaling pathways. J Nanobiotechnol. 2023, 16;21(1):377. [CrossRef]
- Nandini, B.; Hariprasad, P.; Prakash, H. S.; Shetty, H. S.; Geetha, N. Trichogenic-selenium nanoparticles enhance disease suppressive ability of Trichoderma against downy mildew disease caused by Sclerospora graminicola in pearl millet. Scientific Reports, 2017, 7, 2737. [Google Scholar] [CrossRef]
- Haq, I. U.; Cai, X.; Ali, H.; Akhtar, M. R.; Ghafar, M. A.; Hyder, M.; Hou, Y. Interactions between nanoparticles and tomato plants: Influencing host physiology and the tomato leaf miner’s molecular response. Nanomaterials, 2024, 14(22), 1788. [CrossRef]
- Al-Goul, S. T. The impact of eco-friendly nanoparticles on the management of phytopathogenic fungi: A comprehensive review. Journal of Plant Pathology, 2024, 107, 265–290. [Google Scholar] [CrossRef]
- Amin, H. H.; Elsayed, A. B.; Maswada, H. F.; Elsheery, N. I. Enhancing sugar beet plant health with zinc nanoparticles: A sustainable solution for disease management. Journal of Soil, Plant and Environment, 2023, 2(1), 1–20. [CrossRef]
- Elagamey, E.; Abdellatef, M.A.; Haridy, M.S.; Abd El-aziz, E.S.A. Evaluation of natural products and chemical compounds to improve the control strategy against cucumber powdery mildew, Eur. J. Plant Pathol. 2023, 165, 385–400. [Google Scholar] [CrossRef]
- Farhat, M.; Thabet, M.; Haggag, W.; Mosa, A. Efficacy of silicon and titanium nanoparticles biosynthesis by some antagonistic fungi and bacteria for controlling powdery mildew disease of wheat plants. Int. J. Agric. Technol. 2018, 14, 661–674. [Google Scholar]
- Elsharkawy, M. M.; El-Kot, G. A. N.; Hegazi, M. Management of rose powdery mildew by nanosilica, diatomite and bentocide. Egyptian Journal of Biological Pest Control, 25(3), 545–553. Proceedings of the 4th International Conference, ESPCP2015, Cairo, Egypt, 19–22 October 2015.
- Rashad, Y. M, El-Sharkawy, H.H.A.; Belal, B.E.A., Abdel Razik, E.S., Eds.; Galilah, D.A. Silica nanoparticles as a probable anti-oomycete compound against downy mildew, and yield and quality enhancer in grapevines: Field Evaluation, Molecular, Physiological, Ultrastructural, and Toxicity Investigations. Front. Plant Sci. 2021, 12:763365. [Google Scholar] [CrossRef]
- R. ; Singh, P.K.; Kumar, R.; Nair, K.K.; Alam, I.; Sirvasava, C.; Yadav, S.; Gopal, M.; Choudhury, S.R.; Goswaml, A. Suitability of nano-sulphur for biorational Management of Powdery mildew of Okra (Abelmoschus esculentus Moench) caused by Erysiphe cichoracearum. J. Plant Pathol. Microbiol. 2013, 4, 1–4. [Google Scholar] [CrossRef]
- Abdel-Rahman, H. R.; Alkolaly, A. M. A. Comparative studies on nanosulfur and certain fungicides to control cucumber powdery mildew disease and their residues in treated fruits. Science Exchange Journal, 2020, 41(1), 54–70. [CrossRef]
- Chen, H.; Yada, R. Y. Nanotechnologies in agriculture: New tools for sustainable development. Trends in Food Science & Technology, 2011, 22(11), 585–594. [CrossRef]
- Rathore, A.; Hasan, W.; Ramesha N, M.; Pujar, K.; Singh, R.; Panotra, N.; Satapathy, S.N. Nanotech for Crop Protection: Utilizing Nanoparticles for Targeted Pesticide Delivery Uttar Pradesh J. Zool.; 2024, 45 (6), 46-71, 2024. [CrossRef]
- Ruano-Rosa, D.; Sánchez-Hernández, E.; Baquero-Foz, R.; Martín-Ramos, P.; Martín-Gil, J.; Torres-Sánchez, S.; Casanova-Gascón, J. Chitosan-based bioactive formulations for the control of powdery mildew in viticulture. Agronomy, 2022, 12, 495. [Google Scholar] [CrossRef]
- Soleimani, H.; Mostowfizadeh-Ghalamfarsa, R.; Ghanadian, M. (2025). Characterization, biochemical defense mechanisms, and antifungal activities of chitosan nanoparticle-encapsulated spinach seed essential oil. J. Agr. Food Research, 2025, 22: 102016. [CrossRef]
- Patel, A.; Ghosh, V. Thyme (Thymus vulgaris) Essential Oil–Based Antimicrobial Nanoemulsion Formulation for Fruit Juice Preservation. In: Sadhukhan, P.; Premi, S. (eds) Biotechnological Applications in Human Health. Springer, Singapore, 2020. [CrossRef]
- Detsi, A.; Kavetsou, E.; Kostopoulou, I.; Pitterou, I.; Pontillo, A. R. N.; Tzani, A. ; Christodoulou; P. Siliachli, A.; Zoumpoulakis, P. Nanosystems for the encapsulation of natural products: The case of chitosan biopolymer as a matrix. Pharmaceutics, 2020; 12, 669. [Google Scholar] [CrossRef]
- Upadhyay, N.; Singh, V. K.; Dwivedy, A. K.; Chaudhari, A. K.; Dubey, N. K. Assessment of nanoencapsulated Cananga odorata essential oil in chitosan nanopolymer as a green approach to boost the antifungal, antioxidant and in situ efficacy. Int. J. Biol. Macromol., 2021, 171, 480–490. [Google Scholar] [CrossRef]
- Sreelatha, S.; Kumar, N.; Yin, T. S.; Rajani, S. Evaluating the antibacterial activity and mode of action of thymol-loaded chitosan nanoparticles against plant bacterial pathogen Xanthomonas campestris pv. campestris. Frontiers in Microbiology, 2022; 12. [Google Scholar] [CrossRef]
- Desouky, M.M.; Abou-Saleh, R.H.; Moussa, TA.A.; Fahmy; H. M. Nano-chitosan-coated, green-synthesized selenium nanoparticles as a novel antifungal agent against Sclerotinia sclerotiorum: in vitro study. Sci Rep 2025, 15, 1004. [Google Scholar] [CrossRef]
- Keawchaoon, L.; Yoksan, R. (2011). Preparation, characterization and in vitro release study of carvacrol-loaded chitosan nanoparticles. Colloids and Surfaces B: Biointerfaces, 2011, 84, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Feyzioglu, G. C.; Tornuk, F. Development of chitosan nanoparticles loaded with summer savory (Satureja hortensis L.) essential oil for antimicrobial and antioxidant delivery applications. LWT, 2016; 70, 107–110. [Google Scholar] [CrossRef]
- Granata, G.; Stracquadanio, S.; Leonardi, M.; Napoli, E.; Malandrino, G.; Cafiso, V.; Stefani, S.; Geraci, C. Oregano and thyme essential oils encapsulated in chitosan nanoparticles as effective antimicrobial agents against foodborne pathogens. Molecules, 2021. 26, 4055. [CrossRef]
- Gasti, T.; Dixit, S.; Hiremani, V. D. Chougale, R. B.; Masti, S. P.; Vootla, S. K.; Mudigoudra, BS. Chitosan/pullulan-based films incorporated with clove essential oil loaded chitosan-ZnO hybrid nanoparticles for active food packaging. Carbohydrate Polymers, 2022, 277,118866. [Google Scholar] [CrossRef]
- Zhang, H.; Yuan. ; Gao, Y.; Su, P.; Jia, H.; Tang, C.; Meng, H.; Wu, L. Nano protective membrane coated wheat to resist powdery mildew. Frontiers in Plant Science, 2024, 15, 1369330. [Google Scholar] [CrossRef]
- Chowdhury, S.; Kar, K.; Mazumder, R. Exploration of different strategies of nanoencapsulation of bioactive compounds and their ensuing approaches. Future Journal of Pharmaceutical Sciences, 2024, 10, 644. [Google Scholar] [CrossRef]
- Adnan, I.; Waleed, M. Evaluation of the effectiveness of biologically synthesized copper and zinc nanoparticles in controlling powdery mildew on Rose hybrida. Natural and Engineering Sciences, 2025; 10, 552–562. [Google Scholar] [CrossRef]
- Jiménez-Pérez, O.; GallegosMorales, G.; Espinoza-Ahumada, C.A.; Delgado-Luna, C.; Rangel, P.; Espinosa-Palomeque, B. Potential of chitosan for the control of powdery mildew (Leveillula taurica (Lév.) Arnaud) in a Jalapeño Pepper (Capsicum annuum L.) cultivar. Plants, 2024, 13, 915. [Google Scholar] [CrossRef]
- Nandhini, R.; Rajeswari, E.; Harish, S.; Sivakumar, V.; Selvi, R.G.; Sundrasharmila, D. J. Role of chitosan nanoparticles in sustainable plant disease management. Journal of Nanoparticle Research, 2025; 27. [Google Scholar] [CrossRef]
- Ahmed, F.; Adnan, F.; Bazai, M. J.; Fareed, S. R.; Tareen, J. K.; Uddin, M.; Kakar, H. Powdery mildew: A disease of grapes and the fungicides’ mode of action—A review. Bioscience Research, 2022; 3, 78. [Google Scholar] [CrossRef]
- Naaz, H.; Rawat, K.; Saffeullah, P.; Umar, S. Silica nanoparticles synthesis and applications in agriculture for plant fertilization and protection: A review. International Journal of Molecular Sciences, 2023; 23, 1947. [Google Scholar] [CrossRef]
- Laane, H.-M. The effects of foliar sprays with different silicon compounds. Plants, 2018; 7, 45. [Google Scholar] [CrossRef]
- Krishnani, K. K.; Boddu, V. M.; Chadha, N. K.; Saffeullah, P.; Umar, S. Metallic and non-metallic nanoparticles from plant, animal, and fisheries wastes: Potential and valorization for application in agriculture. Environmental Science and Pollution Research, 2022; 29, 81130–81165. [Google Scholar] [CrossRef]
- Li, H.; Chen, X.; Shen, D.; Wu, F.; Pleixats, R.; Pan, J. Functionalized silica nanoparticles: Classification, synthetic approaches and recent advances in adsorption applications. Nanoscale, 2021; 13, 16200–16230. [Google Scholar] [CrossRef]
- González-García, Y.; Cadenas-Pliego, G.; Alpuche-Solís, Á.G.; Cabrera, R.I.; Juárez-Maldonado, A. Carbon Nanotubes Decrease the Negative Impact of Alternaria solani in Tomato Crop. Nanomaterials 2021, 11, 1080. [Google Scholar] [CrossRef]
- Zhang, X.; Cao, H.; Wang, J.; Li, F.; Zhao, J. Graphene Oxide Exhibits Antifungal Activity against Bipolaris sorokiniana In Vitro and In Vivo. Microorganisms 2022, 10, 1994. [Google Scholar] [CrossRef]
- Nair, A.; Mallya, R.; Suvarna, V.; Khan, T.A.; Momin, M.; Omri, A. Nanoparticles—Attractive Carriers of Antimicrobial Essential Oils. Antibiotics 2022, 11, 108. [Google Scholar] [CrossRef]
- Tanaka, H. (2019). Method of controlling powdery mildew using nanobubble water (WO2019230789A1). AQUASOLUTION Corp.
- Slavin, Y.N.; Bach, H. Mechanisms of antifungal properties of metal nanoparticles. Nanomaterials 2022, 12, 4470. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, E.; Krishna, S.B.N.; Golla, N.; Patil, S.J. Nanofungicides: A Promising Solution for Climate-Resilient Plant Disease Management. In: Abd-Elsalam, K.A.; Abdel-Momen, S.M. (eds) Plant Quarantine Challenges under Climate Change Anxiety. Springer, Cham, 2024, pp 513-532. [CrossRef]
- Mohammadi, A.; Hashemi, M.; Hosseini, S. M. Nanoencapsulation of Zataria multiflora essential oil preparation and characterization with enhanced antifungal activity for controlling Botrytis cinerea, the causal agent of gray mould disease. Innovative Food Science and Emerging Technologies 2015, 28, 73–80. [Google Scholar] [CrossRef]
- Hanwate, R.M.; Babu, R.H.; Wadhave, A.A.; Mishra, V. Advancements in Nanosuspension Technology for Drug Delivery. Biomedical Materials & Devices. [CrossRef]
- Kyriakides, T.R.; Raj, A.; Tseng, T.H.; Xiao, H.; Ngu, R.R. ; MoF.S.; F.S.; Halder, S.; Xu, M.; Wu, M.J.; Bao, S.; Sheu, W.C. Biocompatibility of nanomaterials and their immunological properties. Biomed Mater. 2021, 16(4):10. [CrossRef]
- Yin, J.; Su, X.; Yan, S.; Shen, J. Multifunctional nanoparticles and nanopesticides in agricultural application. Nanomaterials, 2023, 13(7), 1255. [CrossRef]
- Guerrini, L.; Alvarez-Puebla, R. A.; Pazos-Perez, N. Surface modifications of nanoparticles for stability in biological environments. Chemical Society Reviews, 2018, 47(10): 3862–3871. [CrossRef]
- Wohlmuth, J.; Tekielska, D.; Cˇechová, J.; Baránek, M. Interaction of the Nanoparticles and Plants in selective growth stages—usual effects and resulting impact on usage perspectives. Plants 2022, 11, 2405. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Lu, H. Protein PEPylation: A New Paradigm of Protein–Polymer Conjugation Bioconjugate Chemistry, 2019, 30 (6), 1604-1616. [CrossRef]
- Lanke, N. P.; Chandak, M. B. Recent trends in deep learning and hyperspectral imaging for fruit quality analysis: An overview. J. Opt. 2025. [Google Scholar] [CrossRef]
- Singh, J.; Kashyap, R.; Bansal, K.; Das, R.; Sandhu, K.; Singh, G.; Saini, D. K. Transition from conventional to AI-based methods for detection of foliar disease symptoms in vegetable crops: A comprehensive review. Journal of Plant Pathology, 2025. [Google Scholar] [CrossRef]
- Vu Thanh, C.; Gooding, J. J.; Kah, M. Learning lessons from nanomedicine to improve the design and performances of nano-agrochemicals. Nature Communications. 2025, 16, 2306. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Sexual and asexual phases off powdery mildew life cycle and the interaction site of the nanoparticles within the stage of its life cycle. The figure is generated using Copilot.
Figure 1.
Sexual and asexual phases off powdery mildew life cycle and the interaction site of the nanoparticles within the stage of its life cycle. The figure is generated using Copilot.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



