
Submitted:
17 October 2025
Posted:
20 October 2025
You are already at the latest version
Abstract
African elephants maintain a delicate balance between high intestinal helminth numbers and symptoms of poor health. However, calves and weaners face survival threats from damage caused by helminth infections and infestations during periods of starvation. Using coprological evaluations, we compared helminth infection patterns and selected risk factors in elephants from Tsavo and Laikipia-Samburu Ecosystem (LSE). Eggs and adult worms were identified and classified into genera using morphological and morphometric characteristics. Overall prevalence rates were high at 95.6% and 98.5% for Tsavo and Laikipia-Samburu respectively. Using the Chi squared test of independence, nematode prevalence rates (97.1%) were significantly different from trematodes (32.6%) ((χ2(1)= 248.84, p< 0.001). Prevalence rates did not differ across age-classes in the two populations. Using the Generalized Linear Model, sex had independent significant effect on the observed mean worm burdens (F(1)= 5.842, p=0.016, CI 95%), but neither age nor location did. However, the interaction had significant effect on mean eggs per gram (EPGs) observed between age and location (F(1)=14.043, p< 0.0005, CI 95%) and between age and sex (F(1)=4.858, p= 0.028, CI 95%). Some of the identified helminth species may potentially lead to life-threatening gut mucosa hemorrhaging and ulcerations, particularly during stressful conditions.
Keywords:
African elephant
; Gastrointestinal parasites
; Helminth prevalence
; Kenya
; Murshidia
; Parasite ecology
; Laikipia
; Tsavo
1. Introduction
Previous studies have evidenced the catastrophic effects that helminths may inflict upon elephant host [1,2,3]. These include pathological lesions, hemorrhages, tissue necrosis and even death [2,3,4]. Helminth infections also suppress the host’s immunity, stagnate growth and decrease reproduction [5].
Due to a well-developed parasite-host equilibrium, the African elephant is usually asymptomatic for gastro-intestinal (GI) parasites [6]. Heavy worm burdens however pose a serious threat to survival. They abrade the epithelial lining of the intestines, causing damage to the rich microvasculature and gaining access to use the host’s ingested nutrients, leading to nutritional deprivation particularly in times of starvation.
Parasite infestations do not always lead to clinical symptoms. In most free-ranging populations, intestinal parasites are neglected with resultant threat to the health and ultimately, survival of especially calves and weaners. Elsheikha and Obanda [7], hypothesized that disease is maintained at subclinical levels as a result of co-evolution between hosts and parasites also cited more recently [6,8]. A parasite-host equilibrium has therefore been established [9]. In particular, helminths produce immune-evasion molecules which help maintain a balance between worm expulsions and virulence. However, clinical signs of disease may begin to show upon destabilizing of the parasite-host equilibrium. This can occur due to factors such as concurrent infection, pregnancy and lactation, or adverse weather changes.
The most common helminths in African elephants are nematodes (Murshidia sp, Khalilia sp, and Quilonia sp) followed by trematodes (Protofasciola sp, and Fasciola sp). Intestinal coccidian infections are common, but have not been associated with any clinical symptoms [1]. In Kenya, the nematode Grammocephalus clathratus, the trematode Protofasciola robusta and a number of unidentified adult worms were recovered from elephant carcasses during the drought of 2009, where 38 young animals aged between 5-8 years died in the Laikipia-Samburu Ecosystem (LSE) [3]. Eleven of the carcasses revealed pathological lesions and hemorrhages that were linked to parasitism. This suggests that helminths could play a potentially important role in regulating wildlife populations.
The aim of this study was to investigate the prevalence of helminth infections and understand species diversity and intensity within two of the largest ecosystems inhabited by elephants in Kenya.
2. Materials and Methods
2.1. Study Area
This study was undertaken in the Tsavo East National Park, the largest in Kenya and considered to be one of the largest in the world with an approximate area of 13,747 Km2, and Laikipia-Samburu ecosystem which covers approximately 25000 Km2 located in the central heartland of the country.
Rivers Galana, Tiva and Voi traverse the park, along which is a narrow riverine forest and thicket dominated by Acacia elatior, Hyphanae compressa and Suaeda monoica. The northern part is more densely vegetated, predominantly Acacia commiphora, while the southern part has been opened up over the years by fire and elephants [10]. Common shrubs here include the genera Premna, Bauhinia and Sericocomopsis, and scattered trees such as Delonix elata, Melia volkensii and baobab (Adansonia digitata). The area experiences two unpredictable rain seasons, between April-May and November-December. The average annual rainfall ranges between 300mm-600mm [11]. Threatened wildlife in this park includes the critically endangered black rhino (Diceros bicornis) and hirola (Damaliscus hunteri), the vulnerable elephants (Loxodonta africana africana), cheetah (Acinonyx jubatus) and leopard (Panthera pardus). Other species in the park include African buffalo (Syncer caffer), lions (Panthera leo), several antelope species including the fringe-eared oryx (Oryx beisa callotis), waterbucks (Kobus ellipsiprymnus) and lesser kudu (Tragelaphus imberbis) and hundreds of bird species, to mention a few [10]. Based on Kenya Wildlife Service [12], this ecosystem is estimated to hold around 7,727 elephants.
The Laikipia-Samburu ecosystem is semi-arid, with a similar bimodal rainfall pattern with an annual average of around 300mm in the north and 700mm in the south. The ecosystem is characterized by hills, plateaus and rough terrain with six major land use types: National reserves, state-protected forest reserves, communal pastoral areas, community conservancies, private ranches and settlements under subsistence production. Wildlife found in this ecosystem include the introduced, near threatened, Northern white rhino (Celatotherium simum cottoni), the black rhino, the endangered reticulated giraffe(Giraffa Camelopardalis reticulata), grevy zebra (Equus grevyi), the vulnerable Somali ostrich (Struthio molybdophanes), elephants, cheetah (Acinonyx jubatus) and leopards (P. pardus); the beisa oryx (Oryx beisa) the gerenuk (Litocranius walleri), several other antelope species, buffaloes (S. caffer), lions (P. leo) and hundreds of bird species. Elephant populations in this area at the time of this study were estimated at 7,166 [13].
2.2. Sample Collection
Freshly voided fecal samples were collected between June and October 2019, following the methods of Baines et al., [1] throughout the week during this whole period. Once spotted, elephant herds or lone males were observed until they had defecated and moved off to a safe distance. We targeted watering holes in the afternoon and opportunistically in the mornings and evenings.
Samples were collected using 50ml and 100ml fecal pots, gloves, wooden splints and 10% formalin as a preservative. Approximately 50g of fecal material was picked from different boluses from the same animal, including some from the top, center and bottom of the bolus. The wooden splints were used to stir the faecal sample with the 10% formalin to ensure homogenization and uniformity in preservation. The fecal pots were then sealed and stored. At the same time demographic variables (age, gender, and family group) were collected and matched with each individual sample. All the samples were collected before the rains in both locations, and were transported to the veterinary laboratory at the KWS headquarters for analysis.
2.3. Sample Processing and Analysis
Parasitological examination and qualitative analysis were done using centrifugal Sheather’s sugar floatation, water sedimentation while quantitative analysis used McMaster egg counting. Fecal material was sorted through to identify presence of adult worms.
2.3.1. Centrifugal Floatation Method
The floatation solution was prepared by mixing 454g of sugar (Kabras Sugar, Kakamega, Kenya) with 355ml of distilled water. The mixture was heated at low temperatures while stirring intermittently until all the sugar dissolved. It was then left to cool to room temperature before use.
Approximately 3g of fecal sample was weighed and placed in a conical flask and 12ml of water added and the mixture strained through a sieve after mixing. The filtrate was transferred to a 15ml centrifuge tube, topped up with water to the 14ml mark, and centrifuged at 1500 rpm for 10 min. The supernatant was poured off and the sediment at the bottom re-suspended with Sheather’s sugar solution; first filled halfway, mixed using a wooden splint and filled to the top until an upper meniscus and a cover slip gently placed on the center. The mixture was then centrifuged at 1500 rpm for 10min after which the cover slip was removed and gently placed on a glass slide for observation under the microscope (X400) (MICROTEC IS300, ISCapture.Ink, Exeter, UK). Micrographs were taken and dimensions of helminth ova measured using the ISCapture micro-imaging software version (MICROTEC IS300, ISCapture.Ink, Exeter, UK)
2.3.2. Sedimentation Method
3g of the faecal sample was weighed and mixed with 45 ml of tap water. The mixture was strained, and the filtrate transferred to a 50ml centrifuge tube. The filtrate was left to sediment for 30 minutes and the supernatant gently decanted out. The sediment was re-suspended with 45ml of tap water and left to stand for at least 10 minutes. Re-suspension and decanting is was repeated until the suspension is clear. 50ul of the sediment is pipetted onto a glass side and covered with a coverslip for observation. Image processing follows the same as that of floatation method.
2.3.3. Mc Master Method
To quantify the burden of helminth infestation, a modified Mc Master method adopted from Foreyt [14] was used, using Sheather’s sugar as the floatation fluid. It follows the same procedure as the sugar floatation, until the point where the sediment is mixed with the Sheather’s sugar solution to form an upper meniscus. No cover slip was placed at this point, instead, the mixture in the 15ml centrifuge tubes were left to stand for about one and a half hours to allow the eggs sufficient time to float to the surface. A pipette was used to transfer suspension from the surface to a McMaster slide, filling both chambers. The slide was placed on a microscope (MICROTEC IS300, ISCapture.Ink, Exeter, UK) and eggs inside the chambers counted under x10 magnification.
For both quantitative and qualitative analysis, 137 faecal samples from TENP and 136 faecal samples from LSE were analysed in each case.
2.3.4. Adult Worm Identification
We carried out manual sorting of faecal samples collected from TENP and LSE to look for adult worms present in the dung. Adult worm processing was conducted following the modified method of McLean et al., [15]. Briefly, worms found were placed in clean sample bottles containing 10% formalin. The worms were later placed on a slide and cleared using glycerol. A coverslip was placed on top of the slide and the sample let to sit for 1 week to achieve clearing before observation [3]. After a week, the slide was placed on the microscope (MICROTEC IS300, ISCapture.Ink, Exeter, UK) for observation under the X100 magnification, and images of anterior and posterior regions were recorded using ISCapture micro-imaging software version (MICROTEC IS300, ISCapture.Ink, Exeter, UK). Whole worms were observed using a dissecting microscope (Leica EZ4D) and images captured using the supporting software LAS EZ (Leica Microsystems, Illinois, USA). To identify the genera present, we focused on studying the morphology of the anterior and posterior ends of identified worms, and compared these with standard published works [16,17,18].
2.4. Data Analysis
Floatation and sedimentation data were used to calculate prevalence of infection for comparison by location, sex and age. Resulting count data on the presence/absence of disease for prevalence was compared based on location, social group and age. Differences were tested for significance using chi square test of independence at 95% CI, using the IBM SPSS version 20 software (IBM, Armonk, USA).
To examine the effect of interaction of location, age, social group and sex on EPG, we utilized the Generalized Linear Model (GLM) with a fitted Poisson distribution function on the IBM SPSS version 20 software.
3. Results
Elephants from TENP and LSE were infected by a wide range of nematodes, mostly strongyles and a few trematodes. Overall, prevalence rates were not significantly different between TENP (95.6%) and LSE (98.5%) (χ2(1)=2.03, p>0.05) (Table 1). Family social group in LSE exhibited prevalence rates that were significantly different from the male social group, (χ2(1) = 7.17, p<0.05) while there were no significant differences observed in the elephant social groups in TENP (χ2(1) = 0.93, p>0.05) (Table 2). Across ages, there were no significant differences observed either; TENP (χ2(2) = 1.54, p>0.05) and LSE (χ2(2) = 1.75, p>0.05) (Table 3). The prevalence of nematode infections was 97.1% while that of trematodes was significantly different at 32.6% (χ2(1) = 248.84, p<0.05), as shown in Table 1. The results indicated that, in general, rates of helminth infection and the egg loads (EPG), were not statistically different between elephants in TENP and in LSE, (F(1)=0.25, p>0.05). Sex had a significant effect on mean worm burden (F(1)=5.84, p<0.05), with females exhibiting higher EPGs than males. However, EPG values within age groups, differed significantly on the basis of sex (F(1)= 4.8, p<0.05) and location (F(1)=14.04, p<0.05, CI 95%) (Figure 1a,b).
Figure 1.
Figure 1. (a) Plot showing the relationship between age and location against worm burden (EPG) in elephant populations from Tsavo East National Park and Laikipia-Samburu ecosystem, Kenya. (b) Plot showing relationship between age and sex against worm burden (EPG) in elephant populations in Tsavo East National Park and Laikipia-Samburu ecosystem, Kenya.
Figure 1.
Figure 1. (a) Plot showing the relationship between age and location against worm burden (EPG) in elephant populations from Tsavo East National Park and Laikipia-Samburu ecosystem, Kenya. (b) Plot showing relationship between age and sex against worm burden (EPG) in elephant populations in Tsavo East National Park and Laikipia-Samburu ecosystem, Kenya.
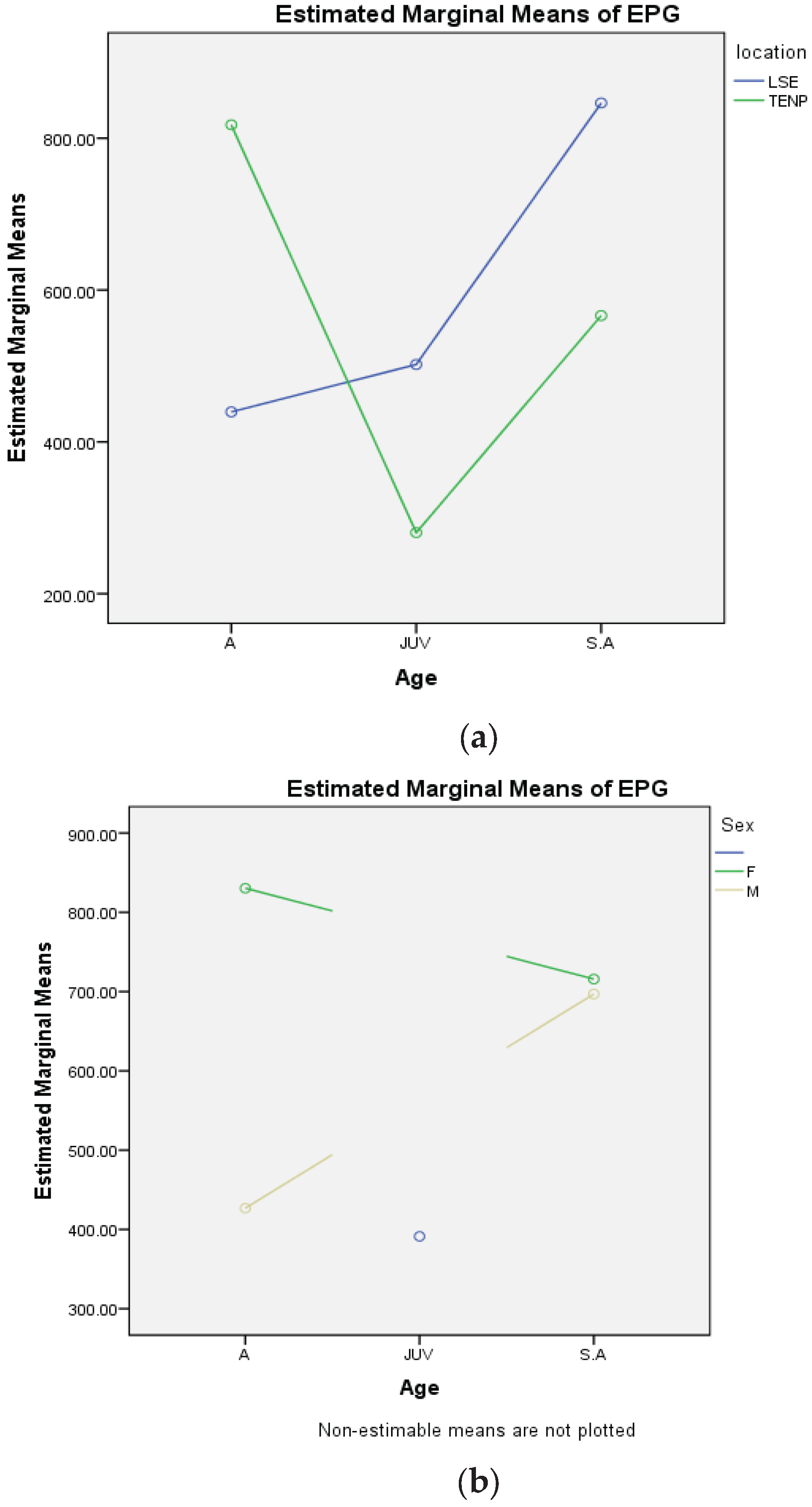

Table 1.
Prevalence of helminth infections in elephants from Tsavo East National Park and Laikipia-Samburu ecosystem, Kenya.
Table 1.
Prevalence of helminth infections in elephants from Tsavo East National Park and Laikipia-Samburu ecosystem, Kenya.
| Category | Variables | TENP | LSE | Total |
|---|---|---|---|---|
| Total Count (N) | 137 | 136 | 273 | |
| Sedimentation | Number infected (n) | 131 | 134 | 265 |
| Prevalence (%) | 95.6 | 98.5 | 97.1 | |
| Trematode infection | Number infected (n) | 54 | 35 | 89 |
| Prevalence (%) | 39.4 | 25.7 | 32.6 |
TENP is Tsavo East National Park and LSE is Laikipia-Samburu ecosystem. All estimation was done based on the sedimentation method.
Table 2.
Prevalence of helminth infections based on elephant social groups in the Tsavo East National Park and Laikipia-Samburu ecosystem, Kenya.
Table 2.
Prevalence of helminth infections based on elephant social groups in the Tsavo East National Park and Laikipia-Samburu ecosystem, Kenya.
| Category | Variables | TENP | LSE | Total |
|---|---|---|---|---|
| Male social group | Count (N) | 65 | 30 | 95 |
| Sedimentation | Number infected (n) | 61 | 28 | 89 |
| Prevalence (%) | 93.9 | 93.3 | 93.7 | |
| Trematode infections | Number infected (n) | 31 | 5 | 36 |
| Prevalence (%) | 47.7 | 16.7 | 37.9 | |
| Family social group | Count (N) | 72 | 106 | 178 |
| Sedimentation | Number infected (n) | 70 | 106 | 176 |
| Prevalence (%) | 97.2 | 100 | 98.9 | |
| Trematode infections | Number infected (n) | 23 | 30 | 53 |
| Prevalence (%) | 31.9 | 28.3 | 29.8 |
TENP is Tsavo East National Park and LSE is Laikipia-Samburu ecosystem. All estimation was done based on the sedimentation method.
Table 3.
Prevalence of helminth infections based on elephant age groups in the Tsavo East National Park and Laikipia-Samburu ecosystem, Kenya.
Table 3.
Prevalence of helminth infections based on elephant age groups in the Tsavo East National Park and Laikipia-Samburu ecosystem, Kenya.
| Category | Variables | TENP | LSE | Total |
|---|---|---|---|---|
| Adult | Count (N) | 87 | 73 | 160 |
| Sedimentation | Number infected (n) | 83 | 71 | 154 |
| Prevalence (%) | 95.4 | 97.3 | 96.3 | |
| Trematode infections | Number infected (n) | 34 | 18 | 52 |
| Prevalence (%) | 39.1 | 24.7 | 32.5 | |
| Sub-adult | Count (N) | 30 | 38 | 68 |
| Sedimentation | Number infected (n) | 28 | 38 | 66 |
| Prevalence (%) | 93.3 | 100 | 97.1 | |
| Trematode infections | Number infected (n) | 10 | 10 | 20 |
| Prevalence (%) | 33.3 | 26.3 | 29.4 | |
| Juvenile | Count (N) | 20 | 25 | 45 |
| Sedimentation | Number infected (n) | 20 | 25 | 45 |
| Prevalence (%) | 100 | 100 | 100 | |
| Trematode infections | Number infected (n) | 10 | 7 | 17 |
| Prevalence (%) | 50 | 28 | 37.8 |
TENP is Tsavo East National Park and LSE is Laikipia-Samburu ecosystem. All estimation was done based on the sedimentation method.
There is no significant difference in overall helminth prevalence rate observed between adults, sub-adults and juveniles in both TENP (χ2(2) = 1.54, p= 0.462) and LSE (χ2(2) = 1.75, p=0.416). Trematode prevalence based on age also shows no significant differences in both TENP (χ2(2) = 1.4q, p= 0.495) and LSE (χ2(2)= 0.12, p= 0.943) (Table 3).
3.1. Intestinal Parasites in Elephants from TENP and LSE Areas
3.1.1. Egg Morphology and Morphometry
A total of 273 elephants were sampled for this analysis, 137 (50.2%) from TENP and 136 from LSE (49.8%). Populations from the sampled areas are infected by nematodes whose eggs were of a typical strongyle-type morphology, as described by [1]. Some nematode genera identified from eggs and their subsequent morphometry are shown in Figure 2a–k. They included those of Quilonia (80-90*40-55μm), Murshidia (70-75*35-50μm), Grammocephalus (65-75*40-50μm), and Khalilia (80-92*44-60μm). Trematode species identified had similar lengths to those of Protofasciola (84-104*56-64μm,) and Fasciola. The eggs of Fasciola hepatica were easily distinguishable from those of the other trematodes due to the absence of an operculum, which is distinct in Protofasciola robusta and Brumptia bicaudata species [9].
3.1.2. Adult Worm Identification
No adult worms were recovered from the TENP samples. However, from the LSE samples, 29 worms were recovered from 8 out of the 136 samples collected. Of the worms recovered, 26 (89.7%) were identified up to genus level. Using keys and descriptions provided by Anderson et al., [16], Monnig, [17] and Van Der Westhuysen, [18], infections from Quilonia sp (Figure 3 a–f) and Murshidia sp (Figure 3 images g-u) were identified. The morphology of the anterior and posterior regions of the worms were found most useful for identification.
3.2. Comparison of Helminth Occurrence in TENP and LSE
Comparison of Prevalence
A total of 273 individual elephants were examined for intestinal helminths. Out of these, 137 (50.2%) individuals were from TENP and 136 (49.8%) individuals from LSE. Based on social grouping, individuals sampled from male social groups were 95 (34.8%) while those samples from family social groups were 178 (65.2%). Adult elephants sampled in this study were 160 (58.6%) of which 87 individuals were from TENP and 73 from LSE. Sub adults were 68 (24.9%), with 30 individuals from TENP and 38 from LSE. Lastly, the total number of juveniles sampled were 45 (16.5%) with 20 individuals from TENP and 25 from LSE. Prevalence rate established from floatation method was 93% while that obtained from sedimentation was 97.1%, this difference proved to be statistically significant (χ2(1)= 4.72, p<0.05) (Table 1).
There was no significant difference in prevalence rates based on location (χ2 (1) = 2.03, p>0.05). The prevalence of trematodes (32.6%) differed from that of nematodes (97.1%) (χ2(1) = 248.84, p<0.001). Elephants from TENP (39.4%) had a trematode prevalence rate that differed from that observed from the population in LSE (25.7%) (χ2(1) =5.81, p<0.05) (Table 1).
There was no significant difference in observed prevalence rates in the male and family social groups of the elephant populations of TENP (χ2 (1)= 0.93, p=0.335). In LSE however the family social group recorded a prevalence rate (100%) that differed from that of the male social group (93.3%) (χ2(1) = 4.715, p= 0.007). There was no significant difference in trematode prevalence based on social groups from both locations: TENP, (χ2(1) = 3.55, p= 0.06) and LSE, (χ2(1) = 1.66, p= 0.198) (Table 2).
3.3. Comparison of Worm Burden
Generalized Linear Modeling at 95% confidence level was used using IBM SPSS Statistics 20, to determine the effect of age sex and location on the worm burden. Based on this model, it was possible to determine the effect of location, age and sex, on the mean worm burden observed. Results obtained from the General Linear Model showed that age alone (F(1)=0.789, p= 0.375 CI 95%) and location alone (F(1)=0.247, p= 0.620, CI 95%) had no significant effect on mean worm burdens observed. Sex had a significant effect on mean worm burden (F(1)= 5.842, p= 0.016, CI 95%), with females exhibiting higher EPGs than males.
Interaction between age and location (F(1)=14.043, p= 0.000219, CI 95%) and age and sex (F(1)=4.858, p= 0.028, CI 95%) did have significant effect on mean EPGs observed as shown in the plots in Figure 1a,b.
Based on location alone, though the difference between them was not statistically significant, adult elephants in TENP exhibited highest EPG means as indicated in Table 4, followed by sub-adults and Juveniles recording the least mean EPG. In LSE, on the other hand, adult elephants recorded the least mean EPG, followed by juveniles and sub-adults recording the highest mean EPG as shown in Table 4. Adults in TENP are more likely to experience higher intensity of worm burdens as compared to adult elephants in LSE. However Juvenile and sub-adult elephants in LSE are more likely to harbor more helminths than those in TENP.
Infection intensity in adult females was higher compared to adult males while sub-adult males and females seem to have similar levels of infection intensity. Mean EPG in adult females is higher than that of sub- adults while adult males have lower egg burdens as compared to sub-adults. It proved difficult to sex elephant calves while collecting samples in the field, hence the missing comparison of sex and age with regard to this group (Figure 1b).
4. Discussion
Our study revealed that African elephants exhibit high prevalence of helminth infections with varied patterns between elephant populations in the TENP and LSE. It should be known that elephants’ habitat loss and fragmentation have led to elephants’ splitting to many sub-populations as a result of movement restriction. King’ori et al [11], have suggested that these elephant sub-populations are likely to suffer different rates of parasite infestation, and in our study, we reported intestinal parasites’ prevalence of 95.6% and 98.5% in elephants from TENP and LSE respectively. Other workers have also reported high prevalence rates in Kenya including 87.5% by Elsheikha et al., [19], and 97.5% by King’ori et al., [11]. These studies established that irrespective of age, social group or location, African elephants are highly susceptible to helminth, especially the nematodes.
In our study, more nematode infection (97.1%) than trematode (32.6%) was observed. Similarly, King’ori et al., [11] in Kenya, and Baines et al., [1] in the Okavango-Delta, have reported more nematodes (97.5% and 73%) than trematodes (39.1% and 26%) respectively. The complex life cycle of trematode may have accounted for the lower prevalence rates, which include the necessity of an intermediate host (aquatic snails), whose presence is largely determined by the presence of a permanent water source. Where these special conditions are not met, trematode infection may be suboptimal compared to nematodes, which undergo a more direct lifecycle [19]. However, during the seasons with abundance of water, elephants do enjoy lengthy periods of time in the water bodies to cool their body temperatures on hot days. Such behavior may serve as risk factor and increase trematode infection.
Apart from the differences in the life cycles of nematodes and trematodes, the abundance and distribution of aquatic snails within the ecosystem serve as risk factors for trematode prevalence. Factors which support aquatic snail distribution include: physico-chemical water quality (water temperature, dissolved oxygen, ions and salts in the water, depth, availability of food), and predation among others, all affect snail species distribution, with resultant effect on trematode prevalence [20]. Although we determined that elephants from TENP recorded higher trematode prevalence than those from LSE, King’ori et al., [11] have observed different result where elephants from LSE recorded higher trematode prevalence than those from TENP. A full scale malacological survey of wildlife habitats in Kenya covering all seasons, would therefore be useful in shedding light on trematode distribution and prevalence in animals in the wild. We hypothesized that location rather than social group influence trematode prevalence.
Elephants’ grazing habits encourage re-infection with nematodes. Grazing areas and the general savannah are characterized by scattered faecal material from elephants and other wild animal species that are found in these areas. In the dry season, most adult elephants preferred to feed on shrubs and trees, due to unavailability of grass. Where grass was found, it was scanty, dry and very close to the ground. Elephants would use their trunk to collect grass, and use their feet to remove dirt and soil from the grass before ingesting. Such feeding habits in contaminated areas will lead to inadvertent ingestion of helminth ova, creating conducive conditions for infection and re-infection, a situation that may account for the high nematode prevalence in elephant populations.
The presence of Protofasciola robusta and Fasciola hepatica can be attributed to the presence of marsh, swamps, streams and even possible watering holes that provide suitable environments for the intermediate snail hosts. Since elephants feed in marshy areas, with the possibilities of ingesting metacercariae, with consequent infection with these parasites. P robusta has been isolated from the duodenum and distal entrance of the bile ducts and small intestines of elephants [2,3], and the parasite has been associated with hemorrhage, intestinal tissue damage and calf fatality [3]. Fasciola hepatica adults have been found occupying the elephant’s bile ducts and can lead to anorexia, constipation, jaundice, anemia and ultimately death. Fowler and Mikota, [9] have explain that chronic infection could lead to obstruction of bile ducts, elevation of intrahepatic blood pressure, hypoproteinemia, hemorrhage and death.
We detected the eggs of elephant hook worm, Grammocephalus clathratus in this study. The adults of this worm are found in the liver and bile ducts of the host and causes hemorrhages and lesions in the liver and bile ducts [3]. Heavy infestation in the bile ducts by these three species can occlude the bile ducts and cause eventual death. Similarly, Murshidia and Quilonia species, whose pathology has not been well defined, were occasionally found in the large intestines and sometimes in the small intestines [2]. While healthy elephants are asymptomatic to helminth infections, starvation, and nutrient deprivation by helminths can cause pathological lesions on the intestinal mucosa [3], and in aggravated helminthosis, elephants’ survival fitness may be greatly compromised.
In this study, we utilized opportunistic non-invasive methods to obtain adult worms that were excreted together with faeces. Because helminth species from same host are subject to extreme variations [18], uncertainties in species classification may exist. We utilized multiple methods to classify the Murshidia and Quilonia spp up to the genus level [16,17,18]. These two species have earlier been found highly concentrated in the caecum and colon of elephant hosts by Condy, [2]. The possible pathological effects of these two species in elephants have been discussed earlier [15,21,22]. Their eggs have also been identified by King’ori et al., [11] from Kenyan elephants across various populations.
We recorded higher mean EPGs in family social groups than in the male social group, an indication that the social structure in elephant populations does have an effect on intensity of helminth infections [11]. Family groups tend to associate in larger herds as compared to bachelor herds and lone bulls. This association in large herds creates an environment of re-infection as they tend to feed for long in the same areas and they defecate in these areas as they feed as explained earlier. In addition, infection intensity in the social groups is a factor of the foraging dynamics found in elephant social groups. Family herds rarely move far from water sources in dry seasons. This is because family herds consist of calves and sub-adults who may not move as fast as adults and are at higher risks of predation and mortality due to exhaustion. Lone bulls and bachelor herds on the other are not held back and can therefore travel further distances, in search of water and even better food in times of drought, with less risk [23]. Thus, in the dry season, when family herds are utilizing dwindled and diminishing resources, the adult bulls are able to acquire better feed and water by traveling further. This therefore means that family herds will undergo nutritive and hydric stress more, leading to increase in intensity of helminth infections due to lowered immunity and thus record higher EPGs as compared to their male counterparts, as observed in this study. Mean EPGs recorded reveal similar patterns of infection intensity, whereby female elephant hosts are more parasitized than male hosts [1,5,11,18]. However, some studies that have looked at the effect of sex on helminth infections in elephants confirmed opposite results [22]. In our case, mean EPGs recorded in female elephants (707) and male elephants (556), was an indication of relatively moderate levels of infection intensity by helminth parasites.
Overall, the patterns observed in the study showed that elephants from TENP have a higher mean EPG (623.06 ± 653.798) compared to elephants from LSE (589.34 ± 589.237) (p> 0.05); and adults in TENP had significantly higher EPGs as compared to adults in LSE. Perhaps, the habitat range and resource distribution in LSE and TENP played a role in this observation. TENP is a gazetted and fenced national park, covering approximately 12,000 Km2. The park has one main source of permanent water, Galana River, with seasonal sources including rivers Tiva and Voi, Aruba dam, scattered ponds, swamps and watering holes [24]. In the dry seasons, elephant home ranges in TENP shrink considerably as water resources become scarce. The elephants retreat to areas along Galana, Voi and Tiva rivers, to increase their chances of survival in the dry season [24]. The reduction in home range therefore increases chances of heavy parasite infestation especially for family social groups, due to foraging in the same grasslands over a prolonged period of time. LSE, on the other hand, covers a much larger area of 33,817Km2 [25]. The ecosystem has a wide range of habitats, associated with climatic gradients within the region: hot and dry lowlands in the north, cool wet highlands to the south, interrupted with rugged mountains and open landscapes. The ecosystem allows for mostly free movement of elephants in between the different land uses due to the wildlife corridors maintained in these areas [25]. Elephant populations in LSE, therefore, have access to a wider range of habitat as compared to those in TENP. During the dry season, elephants in LSE expand their home range in search for water and food, as was observed in Samburu National Reserve during the conducting of this study. Elephant data from Save the Elephants Foundation, included elephant families that were residents of the reserve, migratory herds, and newcomers. This could explain the differences in mean egg burden observed between the two locations. We suggest that elephants in TENP experience more stress in dry periods due to reduction of habitat range and water resources as compared to those in LSE, and therefore experience higher mean egg burden.
Using egg measurements alone for identification of species present presents a few challenges. It has been noted that egg measurements for a single species varies greatly across different elephant populations [11]. To overcome this downfall, it is important, where possible, to study larval stages and adult worm morphology to determine species present. The most assured way however for species determination is through molecular characterization [15].
Whether host species variation exist was not studied in this work, but this knowledge is needed as part of the elephant ecologic system and will be insightful for the parasite-host equilibrium. The patterns observed including sex, age and location may affect mean worm burdens in elephant populations, but more studies are needed to understand the Kenyan ecosystems’ elephants worm burdens and establish ‘high/low worm burden’. This is useful for routine surveillance and health monitoring in wildlife (elephant) health. Ecosystem fragmentation increases pressure on animals (population, movement and resources), which may even be more dire during drought, and these are worsened by high helminth prevalence, especially in calves.
5. Conclusions
Helminth parasite families/species identified in this study are potentially harmful to elephants, especially during stress. The high gastrointestinal parasite burdens should be considered during purposive translocation and during the period of migration, in order to minimize risk of infections and outbreaks. This work should assist wildlife managers to stratify and monitor subpopulations that may be worst afflicted in times of adverse climatic events.
Author Contributions
Conceptualization, S.K.M., E.M.M., and W.W.M.; methodology, S.K.M., E.M.M., W.W.M. and F.O.F.; software, S.K.M., W.W.M. and F.O.F.; validation, S.K.M., E.M.M., W.W.M. and F.O.F.; formal analysis, S.K.M., E.M.M., W.W.M. and F.O.F.; investigation, S.K.M., E.M.M., and W.W.M.; resources, S.K.M., E.M.M., W.W.M. and F.O.F.; data curation, S.K.M., and F.O.F.; writing—original draft preparation, S.K.M., E.M.M., and F.O.F.; writing—review and editing, S.K.M., E.M.M., and W.W.M. and F.O.F.; visualization, S.K.M., E.M.M., W.W.M. and F.O.F.; supervision, E.M.M., and W.W.M.; project administration, S.K.M., E.M.M., and W.W.M.; funding acquisition, S.K.M., and F.O.F. All authors have read and agreed to the published version of the manuscript.
Funding
None.
Institutional Review Board Statement
The ethical review and approval were waived for this study because only the freshly-voided faecal materials were collected and no animal was handled during the study.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data related to the manuscript are available within the manuscript and also on open source on the Repository of the University of Nairobi, Kenya (https://erepository.uonbi.ac.ke/handle/11295/164593).
Acknowledgments
The authors are grateful to the authorities and staff of Kenya Wildlife Services (KWS) and in particular, staff of the Tsavo East National Park and Laikipia-Samburu ecosystem, Kenya.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| CI | Confidence interval |
| EPGs | eggs per gram |
| GI | gastro-intestinal |
| GLM | Generalized Linear Model |
| IBM | International Business Machines Corporation |
| KWS | Kenya Wildlife Service |
| LSE | Laikipia-Samburu ecosystem |
| SPSS | Statistical Package for the Social Sciences |
| TENP | Tsavo East National Park |
References
- Baines, L.; Morgan, E.R.; Ofthile, M.; Evans, K. Occurence and seasonality of internal parasite infectionin elephants, Loxodonta africana, in the Okavango Delta, Botswana. International Journal for Parasitology: Parasites and Wildlife 2015, 4, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Condy, J.B. Observations on internal parasites in Rhodesian elephant, Loxodonta africana Blumenbach 1797. Proceeings and Transactions of the Rhodesia Scientific Association, 1974, 55, 63–99. [Google Scholar]
- Obanda,V. ; Gakuya, F.; Lekolool, I.; Kariuki, J. Composition of intestinal ciliate fauna of free-ranging African elephants in Tsavo West National Park, Kenya. Pachyderm 2007, 42, 90–94. [Google Scholar] [CrossRef]
- King’ori, E.; Obanda, V.; Chiyo, P.I.; Soriguer, R.C.; Morrondo, P.; Angelone, S. Patterns of helminth infection in Kenyan elephant populations. Parasites and vectors 2020, 13. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.M.; Goldenberg, S.Z.; Letitiya, D.; Wittemyer, G. Strongylid infection varies with age, sex, movement and social factors in wild African elephants. Strongylid infection varies with age, sex, movement and social factors in wild African elephants. Parasitology 2020, 147, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Qurratul-Saadah, Z.; Che-Amat, A.; Syed-Hussain, S.S.; Kamaludden, J.; Ariffin, S.M.Z.; Basripuzi, N.H.; Nor-Azlina, A.A. Gastrointestinal Parasites in Asian and African Elephants: A Systematic Review. Tropical Biomedicine 2023, 40, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Elsheikha, H.; Obanda, V. 2010. Parasitic impact on elephant conservation: a Kenyan view. Vet Times, 2010a; 1–6. [Google Scholar]
- Hota, S.R.; Sahoo, S.; Dash, M.; Pahari, A.; Mohanty, B.; Sahoo, N. Molecular detection of Mushidia linstowi in a free-ranging dead elephant calf. Journal of Threatened Taxa 2020, 12, 15359–15363. [Google Scholar] [CrossRef]
- Fowler, M.E.; Mikota, S.K. Biology, Medicine an Surgery of Elephants, 1st ed.; Blackwell Publishing, Iowa, USA, 2006; 592p.
- Kenya Wildlife Service (KWS). Tsavo East National Park. Available at: https://kws.go.ke/tsavo-east-national-park. Accessed 14 October 2025.
- King'ori, E.; Obanda, V.; Chiyo, P.I.; Soriguer, R.C.; Morrondo, P.; Angelone, S. Patterns of helminth infection in Kenyan elephant populations. Parasites and Vectors 2020, 13. [Google Scholar] [CrossRef] [PubMed]
- Kenya Wildlife Service, (2017a). Aerial Total Count of Elephants, Buffaloes and Giraffes in the Tsavo-Mkomazi Ecosystem in February, 2017. Available at: https://www.kws.go.ke/content/aerial-total-count-elephants-buffaloes-and-giraffes-tsav0-mkomazi-ecosystem-february-2017-0. Accessed 20 August 2018.
- Kenya Wildlife Service, (2017b). Results of Censuses of Elephant, Buffalo, Giraffe and Grevy'S Zebra Counted in Five Key Ecosystems Conducted in 2016 and 2017. Available at: https://www.kws.go.ke/content/results-censuses-elephant-buffalo-giraffe-and-grevy's-zebra-counted-five-key-ecosystems. Accessed 20 August 2018.
- Foreyt, W.J. Veterinary Parasitology Reference Manual, 5th ed.; Blackwell Publishing, Iowa, USA, 2001; 235p.
- McLean, E.R.; Kinsella, J.M.; Chiyo, P.; Obanda, V.; Moss, C.; Archie, E.A. Genetic identification of five Strongyle nematode parasites in wild African elephants (Loxodonta africana). Journal of Wildife Diseases 2012, 48, 707–716. [Google Scholar] [CrossRef] [PubMed]
- CIH Keys to the Nematode Parasites of Vertebrates. Farnham Royal, Slough, UK: Commonwealth Agricultural Bureaux, 1974. 1, 17p. Anderson, R.C.; Chabaud, A.G.; Willmott, S.; Bureaux, C.A. (Eds.) 1974. [Google Scholar]
- Monnig, H.O. On some strongylid nematodes of the African elephant. Transactions of the Royal Society of South Africa 1925, 13, 313–321. [Google Scholar] [CrossRef]
- Van Der Westhuysen, O.P. A Monograph of the Helminth Parasites of the Elephant. Onderstepoort Journal of Veterinary Science and Animal Industry 1938, 10, 49–190. [Google Scholar]
- Elsheikha, H.; Patel, J.; Obanda, V. Altered helminth fauna in African elephants in response to resource availability. Vet Times, 2010b; 1–8. [Google Scholar]
- Olkeba, B.K.; Boets, P.; Mereta, S.T.; Yeshigeta, M.; Akessa, G.M.; et al. Environmental and biotic factors affecting freshwater snail intermediate hosts in the Ethiopian Rift Valley region. Parasites and Vectors 2020, 13. [Google Scholar] [CrossRef] [PubMed]
- Thurber, M.I.; O’Connell-Rodwell, C.E.; Turner, W.C.; Nambandi, K.; Kinzley, C.; Rodwell, T.C.; et al. Effects of rainfall, host demography and musth on strongyle fecal egg counts in African elephants (Loxodanta africana) in Namibia. Journal of Wildlife Diseases 2011, 47, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Mbaya, A.W.; Ogwiji, M.; Kumshe, H.A. Effects of Host Demography, Season and Rainfall on the Prevalence and Parasitic Load of Gastrointestinal Parasites of Free-Living Elephants (Loxodonta africana) of the Chad Basin National Park, Nigeria. Pakistan Journal of Biological Sciences 2013, 16, 1152–1158. [Google Scholar] [CrossRef] [PubMed]
- Wato, Y.A.; Prins, H.H.T.; Heitkonig, I.M.A.; Wahungu, G.M.; Ngene, S.M.; Njumbi, S.; Langevelde, F.V. Movement patterns of African elephants (Loxodonta africana) in a semi-arid savanna suggest that they have information on the location of dispersed water sources. Frontiers in Ecology and Evolution 2018, 6, 167. [Google Scholar] [CrossRef]
- Muteti, D.; Ngene, S.M.; Maingi, J. Biophysical and human factors determine the distribution of poached elephants in Tsavo East National Park, Kenya. Pachyderm 2011, 49, 48–60. [Google Scholar] [CrossRef]
- Ihwagi, F.W.; Wang, T.; Wittemyer, G.; Skidmore, A.K.; Toxopeus, A.G.; Ngene, S.; et al. Using Poaching Levels and Elephant Distribution to Assess the Conservation Efficacy of Private, Communal and Government Land in Northern Kenya. PLoS ONE 2015, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
a-k. Photomicrographs of eggs from nematodes and trematodes of 4 genera: a. Fasciola (99.47*52.42 µm), b. and c. are different sizes of Protofasciola: b. (98.39*50.28 µm), c (102.53*53.02µm): d., e. and f. are different sizes of Murshidia d. (71.2*50.46µm) e. (72.27*39.07µm) f. (65.13*37.22µm); g., h. and i. are different sizes of Quilonia; g. (80.02*45.79µm) h. (82.08*43.99µm) i: (76.55*38.63µm); j. Grammocephalus sp (70.36*43.06 µm); k. Khalilia sp (83.52*49.85 µm).
Figure 2.
a-k. Photomicrographs of eggs from nematodes and trematodes of 4 genera: a. Fasciola (99.47*52.42 µm), b. and c. are different sizes of Protofasciola: b. (98.39*50.28 µm), c (102.53*53.02µm): d., e. and f. are different sizes of Murshidia d. (71.2*50.46µm) e. (72.27*39.07µm) f. (65.13*37.22µm); g., h. and i. are different sizes of Quilonia; g. (80.02*45.79µm) h. (82.08*43.99µm) i: (76.55*38.63µm); j. Grammocephalus sp (70.36*43.06 µm); k. Khalilia sp (83.52*49.85 µm).
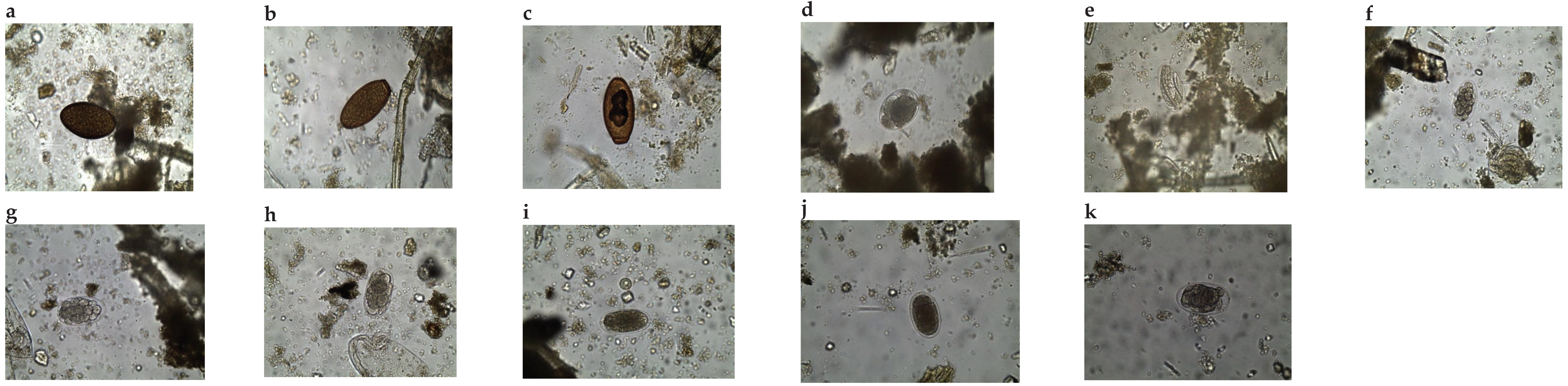
Figure 3.
a-u. Photomicrographs of adult nematode worms isolated from faecal samples. Quilonia sp. male: a. whole worm X8, b. anterior/ head region X100, c. Tail showing copulatory bursa X100, d. Tail region showing spicules X100), Quilonia sp. female: e. anterior region/head X100, f. Tail X100, Murshidia sp. female: whole worm X8: g. anterior, h. posterior, i. anterior region/head X100, j. tail region X100, Murshidia sp. male: k. whole worm X8: l. anterior/ head region X100, m. Tail region with copulatory bursa X100, n. Tail region with arrow showing spiculon X100, Murshidia sp. female: o. worm X8, p. Tail region X100, q. eggs in gravid female X100, r. anterior/ head region X100, Murshidia sp. male: s. whole worm X8: t. anterior/ head region X100, u. tail region showing copulatory bursa and spicules X100.
Figure 3.
a-u. Photomicrographs of adult nematode worms isolated from faecal samples. Quilonia sp. male: a. whole worm X8, b. anterior/ head region X100, c. Tail showing copulatory bursa X100, d. Tail region showing spicules X100), Quilonia sp. female: e. anterior region/head X100, f. Tail X100, Murshidia sp. female: whole worm X8: g. anterior, h. posterior, i. anterior region/head X100, j. tail region X100, Murshidia sp. male: k. whole worm X8: l. anterior/ head region X100, m. Tail region with copulatory bursa X100, n. Tail region with arrow showing spiculon X100, Murshidia sp. female: o. worm X8, p. Tail region X100, q. eggs in gravid female X100, r. anterior/ head region X100, Murshidia sp. male: s. whole worm X8: t. anterior/ head region X100, u. tail region showing copulatory bursa and spicules X100.

Table 4.
Mean helminth burden (epg faeces) for different age group, social group and sex in African elephants from Tsavo East National Park and Laikipia-Samburu ecosystem, Kenya.
Table 4.
Mean helminth burden (epg faeces) for different age group, social group and sex in African elephants from Tsavo East National Park and Laikipia-Samburu ecosystem, Kenya.
| Elephant Population | N | Mean EPG ± SD | Median EPG |
|---|---|---|---|
| TENP | 137 | 623.06 ± 653.798 | 400 |
| Family social group | 72 | 630.68 ± 591.977 | 425 |
| Male social group | 65 | 624.59 ± 741.542 | 400 |
| Adult | 80 | 733.73 ± 737.709 | 500 |
| Sub-adult | 30 | 566.67 ± 545.567 | 375 |
| Juvenile | 20 | 280.45 ± 247.252 | 225 |
| LSE | 136 | 589.34 ± 589.237 | 400 |
| Family social group | 106 | 685.85 ± 623.727 | 525 |
| Male social group | 30 | 248.33 ± 230.996 | 150 |
| Adult | 73 | 469.29 ± 536.717 | 300 |
| Sub-Adult | 38 | 847.56 ± 648.648 | 750 |
| Juvenile | 25 | 502.00 ± 509.591 | 300 |
| Total female EPG | 101 | 706.89 ± 625.963 | 600 |
| Total male EPG | 127 | 555.12 ±650.788 | 350 |
| Total populations’ EPG | 273 | 606.26 ± 621.558 | 400 |
EPG = Eggs per gram; TENP = Tsavo East National Park; LSE = Laikipia-Samburu ecosystem.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



