
Submitted:
16 October 2025
Posted:
17 October 2025
You are already at the latest version
Abstract
Haemonchus contortus (H. contortus) is a gastrointestinal parasite that affects small ruminants, causing anemia, edema, and, in severe cases, death, posing a significant threat to livestock production. This study focused on analyzing the parasite’s sexual differentiation to identify potential molecular targets for the development of control strategies. The genes daf-12, fem-1, and sdc-2 were evaluated based on their orthologs in Caenorhabditis elegans. Specific primers were designed, and nucleic acids were extracted from L3 larvae and adult male and female H. contortus. Gene presence and expression were analyzed using PCR and RT-PCR, along with protein structure modeling and phylogenetic analysis. Results showed differential gene expression depending on life stage and sex: daf-12 was highly expressed in L3 larvae, indicating its involvement in early development; fem-1 showed higher expression in males, suggesting a role in male sexual differentiation; while sdc-2 was more expressed in females, implying a function in regulating female characteristics. These findings provide key insights into the molecular mechanisms underlying sexual differentiation in H. contortus, which could contribute to the development of novel control tools and help mitigate the economic losses caused by this parasite in livestock production.
Keywords:
Haemonchus contortus
; sexual differentiation
; gene expression
1. Introduction
Haemonchosis is a parasitic disease caused by the blood-feeding nematode Haemonchus contortus (H. contortus), which primarily infects small ruminants such as sheep and goats. The economic burden of this parasite is considerable, as it reduces meat and milk production and compromises animal health and welfare [1,2].
Sexual reproduction is essential for population growth in most animals and depends on the generation of two distinct sexes. Sex determination usually occurs at early developmental stages and has profound consequences for development and behavior [3]. In insects, sex is determined by different chromosomal systems (e.g., XY/XX in flies and beetles, ZZ/ZW in butterflies and moths, and XO/XX or Z0/ZZ in some groups), while Hymenoptera use a haplodiploid system in which unfertilized eggs develop into haploid males and fertilized eggs into diploid females [4]. In Drosophila melanogaster, sex determination relies on activation of the sex-lethal gene rather than the presence of a Y chromosome. In mammals such as mice (Mus musculus), the testis-determining gene on the Y chromosome triggers testis formation and systemic hormonal regulation of development and behavior [4,5].
In nematodes, Genetic Sex Determination (GSD) occurs, where sex is defined by the presence or absence of sex chromosomes: females are homogametic XX and males heterogametic XO. Nematode species with chromosomal sex determination typically display a 1:1 sex ratio [6,7]. GSD begins with an initial signal based on the ratio of X chromosomes to autosomes (X:A). This signal initiates a cascade of interacting genes that regulate sexual identity and coordinate the development of male or female traits [8,9]. In addition to sex determination, this cascade also regulates X-chromosome dosage compensation [3].
The free-living nematode Caenorhabditis elegans (C. elegans) has been a fundamental model for studying sex determination. Its pathway is divided into two segments: an upstream branch (from X:A signal to sdc-2) that coordinates sex determination and dosage compensation, and a downstream branch (from her-1 to tra-1) that controls sexual fate [10,11]. Key regulators include xol-1, which promotes male development in XO animals, and sdc-2, which directs female/hermaphrodite fate in XX animals. Downstream, the fem genes (fem-1, fem-2, fem-3) promote tra-1 degradation to enable male development, while active tra-1 specifies female/hermaphrodite identity [3,8,12].
In H. contortus, sexual differentiation also follows an XX/XO system, as demonstrated by microsatellite markers. Males are consistently monomorphic, while females are diploid with expected heterozygosity, and polyandry has been reported, increasing offspring genetic variability [13,14]. Comparative analyses suggest that while some regulators are conserved between C. elegans and H. contortus, others differ. For example, fox-1 and sex-1 orthologs are present but autosomal, and regulators such as xol-1 and sdc-1, sdc-2, sdc-3 appear absent. Most downstream components, including orthologs of her-1 and fem-1/fem-2, are present, though tra-2 and fem-3 were not detected [15,16].
These differences suggest that although H. contortus shares the XX/XO system with C. elegans, its regulatory network is only partially conserved. The aim of this study is to identify and characterize orthologs of key sex-differentiation genes in H. contortus, in order to improve understanding of its reproductive biology and to identify potential molecular targets for novel parasite control strategies.
2. Materials and Methods
2.1. Obtaining H. Contortus L3
The L3 were obtained from the feces of Columbia breed sheep previously infected with a pure strain of H. contortus. This strain has been maintained through successive infections in animals free of other nematodes and was donated by the research group of Dr. Fernando Alba Hurtado from the Multidisciplinary Research Unit of FES-Cuautitlán.
2.2. Sucrose Washing and Recovery of H. Contortus L3
A sucrose solution was prepared by dissolving 20 g of sucrose in 200 mL of pre-heated distilled water. Ten milliliters of this solution were dispensed into four 15-mL conical tubes, and 5 mL of a suspension containing H. contortus L3 were added to each tube, reaching a final volume of 15 mL. The sucrose solution created a density gradient, allowing the larvae to remain suspended.
Tubes were centrifuged at 6,000 × g for 5 min, and the supernatant was decanted. One milliliter of the interphase from each tube was transferred to new 15-mL conical tubes, diluted with 14 mL of distilled water, and centrifuged again under the same conditions. This washing step was repeated three times. Larval pellets obtained after centrifugation were resuspended in 5 mL of 1X PBS and transferred to fresh conical tubes.
To prevent bacterial and fungal growth in the suspension containing H. contortus L3, a 20 µL antibiotic–antimycotic treatment was added, consisting of penicillin (10,000 U/mL), streptomycin (10,000 mg/mL), and amphotericin B (25 µg/mL). The treatment was applied for 24 h under refrigeration at 4 °C.
2.3. Exsheathment and Migration of H. Contortus L3
The conical tube containing H. contortus L3 was centrifuged at 6,000 × g for 5 min, and the supernatant was discarded. Ten milliliters of 1X PBS and 500 µL of sodium hypochlorite were added, followed by centrifugation at 6,000 × g for 5 min and decantation. The larvae were washed three times by adding 10 mL of distilled water, centrifuging under the same conditions, and discarding the supernatant.
A 50-mL conical tube was prepared with 45 mL of 1X PBS supplemented with 90 µL of an antibiotic–antimycotic mixture (penicillin, 10,000 U/mL; streptomycin, 10,000 mg/mL; amphotericin B, 25 µg/mL) and 14.4 µL of ceftriaxone. A mini-Baermann apparatus was assembled by cutting the bottom end of a 15-mL conical tube and placing it inside the 50-mL tube. The open end of the 15-mL tube was covered with a lens-cleaning tissue secured with a rubber band, serving as a filter. The 50-mL tube was then filled with 15 mL of 1X PBS containing antibiotics until the liquid reached the lower edge of the 15-mL tube in contact with the tissue [17].
Finally, the suspension of H. contortus L3 was placed inside the 15-mL tube, allowing the larvae to migrate through the tissue by hydrotaxy while shedding their cuticle. The mini-Baermann apparatus was incubated for 24 h at 37 °C.
2.4. Preservation of H. Contortus L3
The conical tube containing the suspension of H. contortus L3 under antibiotic–antimycotic treatment was removed from the incubator, adjusted to 14 mL with 1X PBS, centrifuged at 6,000 × g for 5 min, and the supernatant discarded. This step was repeated twice. Finally, 3 mL of Trizol (Cat. 15596026, Invitrogen) were added to the L3, the solution was homogenized, and aliquoted into three 1.5-mL microtubes (1 mL each), which were stored at –70 °C until further use.
2.5. Collection of H. Contortus Adult Males and Females
Adult male and female H. contortus were kindly provided by the Helminthology Unit, headed by Dr. María Eugenia López Arellano at CENID-SAI. Parasites were collected directly from the abomasum during necropsies of sheep experimentally infected with H. contortus, which had not been exposed to other pathogens or anthelmintic treatments. These animals belonged to the production unit of CENID-SAI. The collected specimens were washed with 1X PBS and placed in Trizol and stored at –70 °C until use.
2.6. Identification of Genes Associated with Sexual Differentiation in C. elegans
An in-silico search was performed across multiple databases (ScienceDirect, Web of Science, PubMed, Scopus, Wiley, and Bidi-UNAM) using keywords such as “C. elegans,” “sex differentiation,” “sex determination,” and “sex-specific gene expression.” Articles from the last 10 years were prioritized, but earlier studies were included if relevant. Selected studies were evaluated for information on gene function, regulation, and expression related to sexual differentiation.
2.7. Identification of Gene Sequences Related to Sexual Differentiation in the H. contortus Genome
A bioinformatic analysis was performed using C. elegans genes as references. Amino acid sequences were aligned via BLASTp in NCBI, Swiss-Prot, and KEGG databases to identify orthologs in H. contortus. Nucleotide sequences were obtained or inferred from amino acid sequences when unavailable. Genes were selected based on sequence availability and functional relevance. The constitutive gene sod was used as a reference to monitor amplification and normalize results.
2.8. Primer Design for Selected Gene Sequences
Oligonucleotides were designed using the DNA sequences obtained from the conversion of amino acid sequences to nucleotide sequences via the Sequence Manipulation Suite. Complementary strands were generated in the 5’–3’ direction. Oligonucleotide length was optimized to 17–20 base pairs (Table 1). The melting temperature (Tm) was calculated to ensure specific annealing to the template DNA, with an ideal range of 55–65 °C. Terminal complementarity was checked to prevent self- or cross-dimer formation. Designed oligonucleotides were synthesized at the Institute of Biotechnology, UNAM.
2.9. Genomic DNA Extraction
Genomic DNA (gDNA) was extracted from a sample of 1000 exsheathed L3 of H. contortus suspended in 500 µL of 1X PBS. The sample was subjected to lysis by thermal shock in a dry bath at 65 °C for 5 min, followed by mechanical maceration and disruption with a sterile pestle, and immersion in liquid nitrogen for 5 min. This cycle was repeated three times. DNA was then purified using the FavorPrep™ Tissue Genomic DNA Extraction kit (Favorgen®).
gDNAquantification was performed using a Nabi-UV/Vis nanophotometer. RNA integrity was assessed by electrophoresis on 2% agarose gels using 6X TriTack DNA Load-ing Dye (Thermo Scientific®). Gels were documented with the EpiChemi3 Darkroom system and ToupView software.
2.10. RNA Extraction
Total RNA was extracted from a sample of 1000 exsheathed L3 and 10 adult males and females of H. contortus suspended in 500 µL of Trizol. Each sample was subjected to lysis by thermal shock in a dry bath at 65 °C for 5 min, followed by mechanical maceration and disruption with a sterile pestle, and immersion in liquid nitrogen for 5 min. This cycle was repeated three times. RNA was then purified using the Direct-zol™ RNA Miniprep Plus kit (Zymo Research®).
RNA quantification was performed using a Nabi-UV/Vis nanophotometer. RNA integrity was assessed by electrophoresis on 2% agarose gels using 6X TriTack DNA Loading Dye (Thermo Scientific®). Gels were documented with the EpiChemi3 Darkroom system and ToupView software.
2.11. PCR
To confirm the presence of genes associated with sexual differentiation in the H. contortus genome, endpoint PCR was performed using the GoTaq® Green Master Mix 2X kit (Promega). The reaction was carried out with previously extracted gDNA, and primers specifically designed for this analysis.
PCR products were resolved on 2% agarose gels stained with ethidium bromide and compared with a 50 bp molecular marker (Jena Bioscience®). Gels were documented with the EpiChemi3 Darkroom system, and amplicon sizes were estimated using a calibration curve based on marker migration distances.
2.12. RT-PCR
Total RNA previously extracted from L3 and adult males and females was used as a template. RT-PCR reactions were carried out with the GoScript™ Reverse Transcription System kit (Promega®), following the manufacturer’s instructions.
RT-PCR products were resolved on 2% agarose gels stained with ethidium bromide and compared with a 50 bp molecular marker (Jena Bioscience®). Band intensities were quantified using ImageJ, averaged from three replicates, and normalized to the constitutive control (100%).
2.13. Statycal Analysis
Statistical analysis was performed using one-way analysis of variance (ANOVA) on optical densitometry values expressed as relative percentages. Analyses were conducted using GraphPad Prism 9 software, and differences were considered statistically significant at p ≤ 0.05.
2.14. Modeling of the daf-12, fem-1, and sdc-2 Gene Structures
3D models of daf-12, fem-1, and sdc-2 were generated with AlphaFold and validated using SAVES. Stereochemical quality was assessed with PROCHECK Ramachandran plots and ERRAT, yielding Overall Quality Factors comparable to experimentally resolved proteins.
2.15. Phylogenetic Analysis
Phylogenetic trees were constructed using amino acid sequences of DAF-12, FEM-1, and SDC-2 obtained from NCBI and Swiss-Prot (Appendix A). Sequences with high identity and conserved domains were aligned with ClustalW (MEGA 12), and a Neighbor-Joining tree was generated with 1500 Bootstrap replicates.
3. Results
3.1. Determination of the Presence of Genes Related to Sexual Differentiation in the H. contortus Genome
3.1.1. daf-12
For daf-12, amplicons of 61 bp were detected, compared with the constitutive sod gene (36 bp). Optical densitometric analysis revealed relative expression levels of 167% in L3, 117% in males, and 101% in females of H. contortus. One-way ANOVA indicated statistically significant differences between L3–males (p ≤ 0.01) and L3–females (p ≤ 0.001) (Figure 2).
3.1.2. fem-1
For fem-1 amplicons of 81 bp were observed in comparison with the constitutive gene sod (36 bp). Optical densitometric analysis revealed relative expression levels of 57% in L3, 140% in males, and 48% in females of H. contortus. Statistical analysis using ANOVA showed significant differences between L3–males (p ≤ 0.001) and males–females (p ≤ 0.0001) (Figure 3).
3.1.3. sdc-2
Amplicons of 74 bp were observed in comparison with the constitutive gene sod (36 bp). Optical densitometric analysis revealed relative expression levels of 72% in L3, 56% in males, and 127% in females of H. contortus. Statistical analysis using ANOVA showed significant differences between L3–females (p ≤ 0.05) and males–females (p ≤ 0.01) (Figure 4).
3.2. Bioinformatic Analysis
3.2.1. 3D Modeling of daf-12
The daf-12 sequence was analyzed in silico and found to encode DAF-12, a protein composed of 686 amino acids (NCBI: QEA03487.1) containing a characteristic nuclear hormone receptor domain. Visual inspection of the 3D model of DAF-12 predicted by AlphaFold revealed a well-defined core composed of α-helices, represented in navy blue, indicating high confidence in the structural prediction for this region (pLDDT > 90). In contrast, orange-colored regions were observed at the termini of the protein, indicating lower confidence (pLDDT < 50) and likely corresponding to flexible or disordered regions (Figure 5a). The 3D model of DAF-12 was further validated using SAVES with the ERRAT and PROCHECK tools. ERRAT generated an overall quality factor of 94.76%, while PROCHECK provided a stereochemical assessment through a Ramachandran plot, which evaluates the orientation of phi (φ) and psi (ψ) angles of amino acids to determine the proper folding and structural stability of the model. The analysis showed that 81.1% of the residues were in the most favored regions of the plot, 9.8% in additionally allowed regions, 3.8% in generously allowed regions, and 5.8% in disallowed regions (Figure 5b).
3.2.2. 3D Modeling of fem-1
The fem-1 sequence was analyzed in silico and found to encode FEM-1, a protein composed of 652 amino acids (NCBI: XGW30565.1) containing an ankyrin repeat domain. Visual inspection of the 3D model of FEM-1 predicted by AlphaFold revealed predominantly α-helices organized in a repetitive and compact pattern, represented in navy blue (pLDDT > 90) and light blue (pLDDT = 90–70), indicating high confidence in the structural prediction for this region. At the protein termini, yellow and orange regions were observed, corresponding to low-confidence predictions (pLDDT < 50) that likely represent flexible segments (Figure 6a). The 3D model of FEM-1 was validated using SAVES with the ERRAT and PROCHECK tools. ERRAT generated an overall quality factor of 76.79%, while PROCHECK provided a stereochemical evaluation through a Ramachandran plot. The analysis showed that 82.4% of residues were in the most favored regions and 17% in additionally allowed regions, with no residues found in generously allowed or disallowed regions (Figure 6b).
3.2.3. 3D Modeling of sdc-2
The sdc-2 sequence was analyzed in silico and found to encode SDC-2, a protein composed of 486 amino acids (NCBI: CDJ84099.1). Visual inspection of the 3D model of SDC-2 predicted by AlphaFold revealed α-helices, some represented in navy blue (pLDDT > 90) and others in light blue (pLDDT = 90–70), indicating high and moderate confidence in the structural prediction. At the protein termini, yellow (pLDDT > 50) and orange (pLDDT < 50) regions were observed, corresponding to low-confidence predictions that are likely to represent flexible regions (Figure 7a). The 3D model of SDC-2 was validated using SAVES with the ERRAT and PROCHECK tools. ERRAT generated an overall quality factor of 86.24%, while PROCHECK provided a stereochemical evaluation through a Ramachandran plot. The analysis showed that 74.8% of residues were located in the most favored regions, 24.5% in additionally allowed regions, 0.5% in generously allowed regions, and 0.2% in disallowed regions (Figure 7b).
3.3. Phylogenetic Analysis
Phylogenetic trees were reconstructed for daf-12, fem-1, and sdc-2 genes using representative protein sequences from invertebrates (insects, arthropods, nematodes, cestodes, trematodes) and vertebrates (reptiles, birds, fish, mammals) available in GenBank and Swiss-Prot. Analyses were performed using the Neighbor-joining method with 1500 bootstrap (BS) replicates.
For daf-12, the tree showed a clear separation among major taxonomic groups, with well-supported clades for insects, arthropods, nematodes, trematodes, cestodes, fish, amphibians, and mammals. H. contortus clustered within the nematode clade together with C. elegans (BS = 100), Oesophagostomum dentatum (BS = 96), and Toxocara canis (BS = 100), indicating conservation of daf-12 among free-living and parasitic nematodes (Figure 8). Lower similarity was observed between nematodes and vertebrates, with BS = 78 for Ovis aries and Capra hircus, the definitive hosts of H. contortus (Figure 8). These results suggest that daf-12 functions in developmental regulation are conserved across taxa.
For fem-1, a similar pattern was observed, with well-defined clades for all major groups. H. contortus grouped with C. elegans (BS = 100), Ancylostoma caninum (BS = 100), and Toxocara canis (BS = 100), confirming conservation of fem-1 in sexual differentiation processes of free-living and parasitic nematodes (Figure 9). In vertebrates, mammalian sequences were strongly supported (BS = 100 for Ovis aries and Capra hircus), suggesting possible roles of fem-1 in the parasite–host interaction (Figure 9).
For sdc-2, the phylogenetic tree also revealed distinct clades for arthropods, nematodes, trematodes, fish, reptiles, birds, and mammals. H. contortus clustered with C. elegans (BS = 92), supporting functional conservation of sdc-2 in developmental regulation, including X-chromosome dosage compensation during sexual differentiation (Figure 10). However, moderate support was obtained for Trichinella spiralis (BS = 56), indicating incomplete conservation of sdc-2 among nematodes. In vertebrates, mammalian sequences showed high support, particularly Ovis aries (BS = 96) (Figure 10).
4. Discussion
Haemonchosis caused by H. contortus is a globally recognized disease due to its pathogenicity in sheep and goats, although it has also been documented in other ruminants such as cattle, deer, antelopes, and camelids. This parasite represents a major challenge to the livestock industry because of the severe economic losses it causes. It is estimated that H. contortus is responsible for global losses exceeding USD 1.3 billion, associated with decreased animal productivity (e.g., weight gain, meat and milk production), management costs, anthelmintic treatments, and mortality [18]. Control currently relies mainly on the use of anthelmintics; however, their intensive and indiscriminate use has resulted in the widespread emergence of resistance among gastrointestinal nematode populations [19]. This situation highlights the urgent need for alternative, sustainable, and specific control strategies that do not rely solely on anthelmintics. A deeper understanding of the biology of H. contortus, particularly essential processes such as development, reproduction, and sexual differentiation, may offer new opportunities for control. Sexual differentiation is of particular interest given the parasite’s life cycle and infective capacity: adult females can produce 1,000–5,000 eggs per day, which are shed into the environment through feces, contaminating pastures and perpetuating infection [20]. Understanding the mechanisms underlying sexual differentiation could therefore provide strategies to interfere with reproduction and persistence of the parasite in animal production systems.
In free-living nematodes such as C. elegans, several genes have been shown to play key roles in sexual differentiation, acting within highly conserved regulatory cascades [21]. However, in H. contortus these genes remain poorly characterized, and their functions in the dynamics of parasitism are largely unknown. Characterizing the presence, expression, and structure of genes associated with sexual differentiation—such as daf-12, fem-1, and sdc-2—provides valuable insights not only into the basic biology of H. contortus, but also into the identification of potential molecular targets for novel antiparasitic therapies or reproductive control strategies.
In C. elegans, daf-12 encodes a nuclear receptor belonging to the steroid hormone receptor family. It plays a central role in the dafachronic acid-mediated signaling pathway, regulating crucial developmental processes such as diapause, reproductive transitions, and longevity [22,23]. Orthologous sequences identified in H. contortus revealed conserved functional domains, particularly the ligand-binding domain (LBD) and the DNA-binding domain (DBD) [24], suggesting functional conservation despite parasitic adaptations. Molecular analyses confirmed the presence and expression of daf-12. Amplicons of different sizes were obtained: 660 bp from genomic DNA and 61 bp from cDNA, differences consistent with intron–exon organization. Relative expression analysis revealed high expression in infective L3 (67%), but markedly lower levels in adult males (17%) and females (1%). This stage-dependent regulation aligns with the infective role of L3 and with observations in C. elegans, where daf-12 controls developmental arrest and latency exit [22,25]. Structural modeling using AlphaFold revealed a compact α-helical core typical of nuclear receptors, validated by SAVES with an Overall Quality Factor of 94.76% and favorable stereochemical parameters. Phylogenetic analysis further confirmed strong clustering of H. contortus with other nematodes, supported by high bootstrap values, while vertebrate orthologs diverged functionally, including homologs such as the vitamin D receptor and bile acid receptor [26,27]. Collectively, these findings highlight daf-12 as a key regulator of development and sexual differentiation in H. contortus and a potential molecular target for novel control strategies.
In C. elegans, fem-1 is essential for male sexual determination. It promotes degradation of the transcription factor TRA-1 via the ubiquitin–proteasome pathway in association with FEM-2 and FEM-3, thereby enabling male development and spermatogenesis [28,29]. The ortholog identified in H. contortus exhibited strong similarity with C. elegans FEM-1, particularly within ankyrin repeat domains, key for protein–protein interactions [30]. PCR confirmed both presence and expression of fem-1, with amplicons of 154 bp from genomic DNA and 81 bp from cDNA, again reflecting intron–exon structure. Expression analysis revealed sex- and stage-dependent patterns: subexpression in L3 (43%), higher expression in adult females (52%), and a relative overexpression in adult males (40%). This expression profile is consistent with a conserved role in male sexual differentiation, as reported in C. elegans. Structural predictions revealed an ankyrin repeat-rich α-helical protein with high-confidence regions, validated by SAVES (Overall Quality Factor 76.79%) and PROCHECK stereochemistry analyses. Phylogenetic analysis placed H. contortus within a well-supported nematode clade alongside C. elegans and other parasites, indicating evolutionary conservation of fem-1 functions. In vertebrates, orthologs were more divergent and associated with broader roles such as apoptosis and neuronal development [31]. Overall, fem-1 appears to play a conserved role in male differentiation and reproductive processes in nematodes, making it a promising target for antiparasitic interventions.
In the case of sdc-2 in C. elegans has dual functions in female sex determination and dosage compensation in XX individuals, initiating assembly of the dosage compensation complex (DCC) and repressing her-1 to promote female development [32,33]. In H. contortus, an ortholog was identified, suggesting partial conservation of dosage compensation and sexual regulation mechanisms. PCR confirmed presence and expression of sdc-2, with amplicons of 469 bp from genomic DNA and 74 bp from cDNA, consistent with intron–exon differences. Expression analysis revealed low levels in L3 (28%) and males (44%), but significant overexpression in females (27%), in agreement with its role in sex determination and X-chromosome regulation. Structural modeling predicted an α-helical protein core with flexible terminal regions, validated by SAVES (Overall Quality Factor 86.24%) and PROCHECK analysis, indicating a reliable structural model. Phylogenetic analysis grouped H. contortus with C. elegans and Trichinella spiralis within a conserved nematode clade, while arthropods, trematodes, and vertebrates formed distinct clades. Notably, vertebrate orthologs showed divergent functions, including roles in transcriptional regulation, angiogenesis, and even tumor invasiveness [34,35]. These results suggest that while sdc-2 retains core regulatory functions in nematodes, its role has diversified in vertebrates.
Together, these findings confirm that daf-12, fem-1, and sdc-2 play crucial and conserved roles in the sexual differentiation and development of H. contortus. Their evolutionary conservation among nematodes, combined with structural and expression analyses, underscores their importance in reproductive regulation. Importantly, these genes represent promising molecular targets for developing novel strategies to disrupt parasite reproduction and control haemonchosis in livestock.
5. Conclusions
This study confirmed the presence and expression of daf-12, fem-1, and sdc-2 in H. contortus, with stage- and sex-specific expression patterns consistent with their described roles in C. elegans. Structural modeling and validation confirmed conserved functional domains, while phylogenetic analyses revealed strong evolutionary conservation among nematodes and divergence from other phyla. These findings underscore the importance of these genes in the development and reproduction of H. contortus and support their potential as molecular targets for novel antiparasitic strategies.
Author Contributions
Conceptualization, V.H.D.R.A. and H.A.D.; methodology, V.H.D.R.A., S.M.O., I.G.D.E., Y.A.C., H.A.D., F.A.H., M.R.C.P. and C.C.V.; validation, V.H.D.R.A., S.M.O., I.G.D.E., Y.A.C., H.A.D., F.A.H., M.R.C.P and C.C.V.; formal analysis, V.H.D.R.A., S.M.O., Y.A.C., H.A.D. and F.A.H.; investigation, V.H.D.R.A., S.M.O., I.G.D.E., Y.A.C., H.A.D., F.A.H., M.R.C.P. and C.C.V.; resources, V.H.D.R.A.; data curation, V.H.D.R.A., Y.A.C. and H.A.D.; writing—original draft preparation, V.H.D.R.A. and S.M.O.; writing—review and editing, V.H.D.R.A., S.M.O., I.G.D.E., Y.A.C., H.A.D., F.A.H., M.R.C.P and C.C.V.; visualization, V.H.D.R.A., and H.A.D.; supervision, V.H.D.R.A., Y.A.C. and H.A.D.; project administration, V.H.D.R.A.; funding acquisition, V.H.D.R.A. and F.A.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Grant IA-207023 from Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica, (PAPIIT), Dirección General de Asuntos del Personal Académico, (DGAPA), Universidad Nacional Autónoma de México, (UNAM) to Víctor Hugo Del Río- Araiza, and by Grant PAPIIT IN-215324 to Fernando Alba-Hurtado.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The datasets generated and analyzed during the current study are included in the present manuscript. Furthermore, they are available from the corresponding author on request.
Acknowledgments
The authors wish to thank Maria Eugenia López Arellano, from CENID-SAI, INIFAP, for providing the adult Haemonchus contortus parasites. We also thank the Secretaría de Ciencia, Humanidades, Tecnología e Innovación (SECIHTI) for supporting this research through graduate scholarships awarded to Selina Montes de Oca-Lagunas and Melodía Rubí Castro-Pérez.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| DNA | Deoxyribonucleic Acid |
| cDNA | Complementary DNA |
| gDNA | Genomic DNA |
| RNA | Ribonucleic Acid |
| mRNA | Messenger RNA |
| BLASTn | Basic Local Alignment Search Tool nucleotide |
| BLASTp | Basic Local Alignment Search Tool protein |
| BS | Bootstrap |
| DCC | Dosage Compensation Complex |
| daf-12 | Dauer formation 12 |
| DBD | DNA-Binding Domain |
| fem-1 | Feminization-1 |
| GSD | Genetic Sex Determination |
| L3 | Larval stage 3 |
| PCR | Polymerase Chain Reaction |
| PBS | Phosphate-Buffered Saline |
| sdc-1 | Sex Determination and Dosage Compensation-1 |
| SOD | Superoxide Dismutase |
| Tm | Melting Temperature |
Appendix A
Appendix A.1. Accession Data of the Sequences Used for the Phylogenetic Analysis of DAF-12
| Accession number | Species | Protein | Data base |
| XP 0539946974.1 | Anastrepha ludens | Nuclear hormone receptor HR96 | NCBI |
| NP 524493.1 | Drosophila melanogaster | Hormone receptor-like in 96 | NCBI |
| XP 037952769.1 | Teleopsis dalmanni | Nuclear hormone receptor HR96-like | NCBI |
| XP 030369834.1 | Scaptodrosophila lebanonensis | Nuclear hormone receptor HR96 isoform X1 | NCBI |
| KAH8407185.1 | Zaprionus bogoriensis | Nuclear hormone receptor HR96 isoform X1 | NCBI |
| XP 013110462.2 | Stomoxys calcitrans | Nuclear hormone receptor HR96 | NCBI |
| AEC03603.1 | Musca domestica | Nuclear receptor HR96 | NCBI |
| XP 065371768.1 | Calliphora vicina | Nuclear hormone receptor HR96 | NCBI |
| KNC27967.1 | Lucilia cuprina | Nuclear hormone receptor HR96 | NCBI |
| XP 001981892.1 | Drosophila erecta | Nuclear hormone receptor HR96 | NCBI |
| XP 016036188.1 | Drosophila simulans | Nuclear hormone receptor HR96 | NCBI |
| XP 033163627.1 | Drosophila mauritiana | Nuclear hormone receptor HR96 | NCBI |
| XP 037570853.1 | Dermacentor silvarum | Vitamin D3 receptor | NCBI |
| XP 037523479.1 | Rhipicephalus sanguineus | Vitamin D3 receptor isoform X1 | NCBI |
| XP 037286542.1 | Rhipicephalus microplus | Vitamin D3 receptor-like | NCBI |
| XP 003748267.1 | Galendromus occidentalis | Vitamin D3 receptor | NCBI |
| KAK5968683.1 | Trichostrongylus colubriformis | Nuclear receptor subfamily 6 group A member 1 | NCBI |
| XSG12237.1 | Toxocara canis | Nuclear hormone receptor DAF-12, partial | NCBI |
| ALF38191.1 | Anisakis simplex | Nuclear hormone receptor, partial | NCBI |
| KHJ95871.1 | Oesophagostomum dentatum | Ligand-binding domain of nuclear hormone receptor, partial | NCBI |
| NP 001041239.1 | Caenorhabditis elegans | Nuclear hormone receptor family member daf-12 | NCBI |
| KJH46651.1 | Dictyocaulus viviparus | Zinc finger, C4 type | NCBI |
| KAK6030423.1 | Ostertagia ostertagi | Zinc finger, C4 type | NCBI |
| QEA03487.1 | Haemonchus contortus | Nuclear hormone receptor DAF-12 | NCBI |
| KAG5443350.1 | Clonorchis sinensis | Nuclear hormone receptor member daf-12 | NCBI |
| XP 035587464.2 | Schistosoma haematobium | Nuclear hormone receptor, partial | NCBI |
| TPP66369.1 | Fasciola gigantica | Nuclear hormone receptor HR96 | NCBI |
| THD27323.1 | Fasciola hepatica | Nuclear hormone receptor HR96 | NCBI |
| KAF8568779.1 | Paragonimus westermani | Nuclear hormone receptor, partial | NCBI |
| KAH9283862.1 | Echinococcus granulosus | Nuclear hormone receptor family member daf-12 | NCBI |
| KAL5964423.1 | Taenia solium | Nuclear hormone receptor family member daf-12 | NCBI |
| CDS25950.1 | Hymenolepis microstoma | Nuclear hormone receptor HR96 | NCBI |
| CDS43059.1 | Echinococcus multilocularis | Nuclear hormone receptor HR96 | NCBI |
| XP 040281812.1 | Bufo bufo | Vitamin D3 receptor isoform X1 | NCBI |
| XP 040196717.1 | Rana temporaria | Vitamin D3 receptor isoform X1 | NCBI |
| XP 002935703.1 | Xenopus tropicalis | Vitamin D3 receptor | NCBI |
| XP 044143545.1 | Bufo gargarizans | Vitamin D3 receptor | NCBI |
| XP 033795349.1 | Geotrypetes seraphini | Vitamin D3 receptor | NCBI |
| XP 069509284.1 | Ambystoma mexicanum | Vitamin D3 receptor isoform X1 | NCBI |
| XP 072262732.1 | Pyxicephalus adspersus | Vitamin D3 receptor | NCBI |
| XP 030054021.1 | Microcaecilia unicolor | Vitamin D3 receptor | NCBI |
| XP 029450950.1 | Rhinatrema bivittatum | Vitamin D3 receptor isoform X1 | NCBI |
| XP 007240874.1 | Astyanax mexicanus | Vitamin D3 receptor A | NCBI |
| XP 026775925.1 | Pangasianodon hypophthalmus | Vitamin D3 receptor A | NCBI |
| NP 570994.1 | Danio rerio | Vitamin D3 receptor A | NCBI |
| XP 028855014.1 | Denticeps clupeoides | Vitamin D3 receptor | NCBI |
| XP 014868079.1 | Poecilia mexicana | Vitamin D3 receptor | NCBI |
| XP 020468294.1 | Monopterus albus | Vitamin D3 receptor | NCBI |
| NP 001121989.1 | Oryzias latipes | Vitamin D3 receptor | NCBI |
| W5PWT0 | Ovis aries | Bile acid receptor | Swiss-Prot |
| Q96RI1 | Homo sapiens | Bile acid receptor | Swiss-Prot |
| Q60641 | Mus musculus | Bile acid receptor | Swiss-Prot |
| Q3SZL0 | Bos taurus | Bile acid receptor | Swiss-Prot |
| A0A8D0VZF6 | Sus scrofa | Nuclear receptor subfamily 1 group H member 4 | Swiss-Prot |
| A0A452FGV9 | Capra hircus | Nuclear receptor subfamily 1 group H member 4 | Swiss-Prot |
| A0A0G2K6D2 | Rattus norvegicus | Nuclear receptor subfamily 1, group H, member 4 | Swiss-Prot |
Appendix A.2. Accession Data of the Sequences Used for the Phylogenetic Analysis of FEM-1
| Accession number | Species | Protein | Data base |
| NP 611508.1 | Drosophila melanogaster | Fem-1, isoform A | NCBI |
| XP 047115103.1 | Schistocerca piceifrons | Protein fem-1 homolog B | NCBI |
| XP 063885865.1 | Scylla paramamosain | Protein fem-1 homolog B-like isoform X1 | NCBI |
| XP 001654238.1 | Aedes aegypti | Protein fem-1 homolog B-like isoform X1 | NCBI |
| XP 023933934.1 | Bicyclus anynana | Protein fem-1 homolog B-like isoform X1 | NCBI |
| XP 011202197.2 | Bactrocera dorsalis | Protein fem-1 homolog B-like isoform X1 | NCBI |
| XP 002138527.2 | Drosophila pseudoobscura | Protein fem-1 homolog B-like isoform X1 | NCBI |
| XP 050712882.1 | Eriocheir sinensis | Protein fem-1 homolog B-like isoform X1 | NCBI |
| XP 036330667.1 | Rhagoletis pomonella | Protein fem-1 homolog B-like isoform X1 | NCBI |
| XP 031341515.1 | Photinus pyralis | Protein fem-1 homolog B | NCBI |
| XP 061507539.1 | Anopheles gambiae | Protein fem-1 homolog B | NCBI |
| XP 022128557.1 | Pieris rapae | Protein fem-1 homolog B-like isoform X1 | NCBI |
| XP 019768800.1 | Dendroctonus ponderosae | Protein fem-1 homolog B-like isoform X1 | NCBI |
| XP 023012557.1 | Leptinotarsa decemlineata | Protein fem-1 homolog B-like isoform X1 | NCBI |
| XP 049701068.1 | Helicoverpa armigera | Protein fem-1 homolog B-like isoform X1 | NCBI |
| XP 068896437.1 | Tenebrio molitor | Protein fem-1 homolog B-like isoform X1 | NCBI |
| XP 050044960.1 | Dermacentor andersoni | Protein fem-1 homolog B | NCBI |
| XP 037563812.1 | Dermacentor silvarum | Protein fem-1 homolog B | NCBI |
| XP 037511638.1 | Rhipicephalus sanguineus | Protein fem-1 homolog B | NCBI |
| XP 037291813.1 | Rhipicephalus microplus | Protein fem-1 homolog B-like | NCBI |
| XP 022670278.1 | Varroa destructor | Protein fem-1 homolog B-like isoform X1 | NCBI |
| XGW30566.1 | Haemonchus contortus | Protein fem-1 homolog B | NCBI |
| KAK5970382.1 | Trichostrongylus colubriformis | Sex-determining protein fem-1 | NCBI |
| RCN43014.1 | Ancylostoma caninum | Ankyrin repeat protein | NCBI |
| KJH51017.1 | Dictyocaulus viviparus | Ankyrin repeat protein | NCBI |
| KAK6025858.1 | Ostertagia ostertagi | Ankyrin repeat protein | NCBI |
| KHN80228.1 | Toxocara canis | Sex-determining protein fem-1 | NCBI |
| PIO65957.1 | Teladorsagia circumcincta | Ankyrin repeat protein | NCBI |
| MCP9265608.1 | Dirofilaria immitis | Protein fem-1 | NCBI |
| NP 500824.1 | Caenorhabditis elegans | Sex-determining protein fem-1 | NCBI |
| THD25860.1 | Fasciola hepatica | Protein fem-1 | NCBI |
| KAA0198803.1 | Fasciolopsis buskii | Fem-1 A | NCBI |
| TPP64208.1 | Fasciola gigantica | Protein fem-1 | NCBI |
| KAG5441328.1 | Clonorchis sinensis | Protein fem-1 C | NCBI |
| TNN08969.1 | Schistosoma japonicum | Protein fem-1 | NCBI |
| XP 035585568.1 | Schistosoma haematobium | Protein fem-1 C | NCBI |
| XP 018647049.1 | Schistosoma mansoni | Sex-determining protein fem-1 | NCBI |
| KAH9279482.1 | Echinococcus granulosus | Protein fem-1 -like protein C | NCBI |
| CDS43292.1 | Echinococcus multilocularis | Sex-determining protein fem-1 | NCBI |
| KAL5961090.1 | Taenia solium | Sex-determining protein fem-1 | NCBI |
| CDS26165.1 | Hymenolepis microstoma | Sex-determining protein fem-1 | NCBI |
| XP 005722081.1 | Pundamilia nyererei | Protein fem-1 homolog C | NCBI |
| XP 003446110.1 | Oreochromis niloticus | Protein fem-1 homolog C | NCBI |
| XP 006789826.1 | Neolamprologus brichardi | Protein fem-1 homolog C | NCBI |
| XP 015828271.1 | Nothobranchius furzeri | Protein fem-1 homolog C | NCBI |
| XP 013870593.1 | Austrofundulus limnaeus | Protein fem-1 homolog C-like | NCBI |
| XP 064901447.1 | Columba livia | Protein fem-1 homolog C isoform X1 | NCBI |
| XP 065717267.1 | Patagioenas fasciata | Protein fem-1 homolog C | NCBI |
| XP 065513711.1 | Caloenas nicobarica | Protein fem-1 homolog C | NCBI |
| NXK13339.1 | Herpetotheres cachinnans | FEM1C protein | Swiss-Prot |
| XP 072716475.1 | Ciconia boyciana | Protein fem-1 homolog C | NCBI |
| NXV77506.1 | Atlantisia rogersi | FEM1C protein, partial | NCBI |
| XP 027827254.2 | Ovis aries | Protein fem-1 homolog C | NCBI |
| XP 017909215.1 | Capra hircus | Protein fem-1 homolog C isoform X1 | NCBI |
| P0C6P7 | Rattus norvegicus | Protein fem-1 homolog B | Swiss-Prot |
| A0A2I3SXM4 | Pan troglodytes | Fem-1 homolog B | Swiss-Prot |
| G7MY25_MACMU | Macaca mulatta | FEM1B | Swiss-Prot |
| I3L6T4 | Sus scrofa | Fem-1 homolog B | Swiss-Prot |
| A0A8C0K439 | Canis lupus dingo | Fem-1 homolog B | NCBI |
| A0A8C9P293_SPEDA | Spermophilus dauricus | Fem-1 homolog B | Swiss-Prot |
| NP 001039691.1 | Bos taurus | Protein fem-1 homolog A | NCBI |
| Q9Z2G0 | Mus musculus | Protein fem-1 homolog B | Swiss-Prot |
| Q9UK73 | Homo sapiens | Protein fem-1 homolog B | Swiss-Prot |
Appendix A.3. Accession Data of the Sequences Used for the Phylogenetic Analysis of SDC-2
| Accession number | Species | Protein | Data base |
| XP 050040600.1 | Dermacentor andersoni | Syndecan 3 | NCBI |
| XP 037507833.1 | Rhipicephalus sanguineus | Syndecan-like | NCBI |
| XP 037507832.1 | Rhipicephalus sanguineus | Syndecan | NCBI |
| XP 037570033.1 | Dermacentor silvarum | Syndecan-like | NCBI |
| XP 065291937.1 | Dermacentor albipictus | Syndecan-like | NCBI |
| CDJ84099.1 | Haemonchus contortus | CBN-SDC-2 protein | NCBI |
| NP 509924.1 | Caenorhabditis elegans | Sex determination and dosage compensation protein sdc-2 | NCBI |
| XP 003379719.1 | Trichinella spiralis | Syndecan-2 | NCBI |
| CAH8611371.1 | Schistosoma bovis | Syndecan | NCBI |
| KAH8873224.1 | Schistosoma japonicum | Syntenin-2 (Syndecan-binding protein 2) | NCBI |
| XP 018653325.1 | Schistosoma mansoni | Putative syntenin-2 (Syndecan-binding protein 2) | NCBI |
| CAH8865890.1 | Trichobilharzia regenti | Syndecan | NCBI |
| XP 062265467.1 | Platichthys flesus | Syndecan-2 | NCBI |
| XP 036793385.1 | Oncorhynchus mykiss | Syndecan-2 isoform X1 | NCBI |
| XP 067257318.1 | Chanodichthys erythropterus | Syndecan-2-A-like isoform X1 | NCBI |
| XP 053302077.1 | Pleuronectes platessa | Syndecan-2 | NCBI |
| XP 060948873.1 | Limanda limanda | Syndecan-2 | NCBI |
| XP 034455044.1 | Hippoglossus hippoglossus | Syndecan-2 | NCBI |
| XP 028255454.1 | Parambassis ranga | Syndecan-2-A-like | NCBI |
| XP 037748165.1 | Chelonia mydas | Syndecan-2-A-like isoform X1 | NCBI |
| XP 038247599.1 | Dermochelys coriacea | Syndecan-2-A-like isoform X2 | NCBI |
| XP 044859986.1 | Mauremys mutica | Syndecan-2 | NCBI |
| XP 053874007.1 | Malaclemys terrapin pileata | Syndecan-2 | NCBI |
| XP 039382872.1 | Mauremys reevessi | Syndecan-2 | NCBI |
| NP 001001462.2 | Gallus gallus | Syndecan-2 precursor | NCBI |
| XP 015711411.1 | Coturnix japonica | Syndecan-2 | NCBI |
| XP 021243681.1 | Numida meleagris | Syndecan-2 | NCBI |
| NP 001290149.1 | Meleagris gallopavo | Syndecan-2 precursor | NCBI |
| XP 005042472.1 | Ficedula albicollis | Syndecan-2 | NCBI |
| XP 042665043.1 | Centrocercus urophasianus | Syndecan-2 | NCBI |
| XP 065609569.1 | Cyrtonyx montezumae | Syndecan-2 | NCBI |
| XP 014953400.1 | Ovis aries | Syndecan-2-A-like isoform X1 | NCBI |
| NP 001029960.2 | Bos taurus | Syndecan-2 precursor | NCBI |
| NP 032330.1 | Mus musculus | Syndecan-2 precursor | NCBI |
| NP 037214.1 | Rattus norvegicus | Syndecan-2 precursor | NCBI |
| NP 001247893.1 | Macaca mulatta | Syndecan-2 precursor | NCBI |
| NP 001302683.1 | Sus scrofa | Syndecan-2 precursor | NCBI |
| NP 002989.2 | Homo sapiens | Syndecan-2 precursor | NCBI |
References
- Arsenopoulos, K. V.; Fthenakis, G.C.; Katsarou, E.I.; Papadopoulos, E. Haemonchosis: A Challenging Parasitic Infection of Sheep and Goats. Anim. 2021, Vol. 11, Page 363 2021, 11, 363. [Google Scholar] [CrossRef]
- Naeem, M.; Iqbal, Z.; Roohi, N. Ovine Haemonchosis: A Review. Trop. Anim. Health Prod. 2020, 53, 1–12. [Google Scholar] [CrossRef]
- Goodwin, E.B.; Ellis, R.E. Turning Clustering Loops: Sex Determination in Caenorhabditis Elegans. Curr. Biol. 2002, 12, R111–R120. [Google Scholar] [CrossRef]
- Kageyama, D. Sex Determination, Insects. Encycl. Reprod. Vol. 1-6, Second Ed. 2018, 6, 198–203. [Google Scholar] [CrossRef]
- Forger, N.G.; de Vries, G.J. Cell Death and Sexual Differentiation of Behavior: Worms, Flies, and Mammals. Curr. Opin. Neurobiol. 2010, 20, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.M.; Grote, A.; Mattick, J.; Tracey, A.; Tsai, Y.C.; Chung, M.; Cotton, J.A.; Clark, T.A.; Geber, A.; Holroyd, N.; et al. Sex Chromosome Evolution in Parasitic Nematodes of Humans. Nat. Commun. 2020 111 2020, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pires-daSilva, A. Evolution of the Control of Sexual Identity in Nematodes. Semin. Cell Dev. Biol. 2007, 18, 362–370. [Google Scholar] [CrossRef]
- Hansen, D.; Pilgrim, D. Sex and the Single Worm: Sex Determination in the Nematode C. Elegans. Mech. Dev. 1999, 83, 3–15. [Google Scholar] [CrossRef]
- Shinya, R.; Sun, S.; Dayi, M.; Tsai, I.J.; Miyama, A.; Chen, A.F.; Hasegawa, K.; Antoshechkin, I.; Kikuchi, T.; Sternberg, P.W. Possible Stochastic Sex Determination in Bursaphelenchus Nematodes. Nat. Commun. 2022, 13. [Google Scholar] [CrossRef]
- Wolff, J.R.; Zarkower, D. Somatic Sexual Differentiation in Caenorhabditis Elegans. Curr. Top. Dev. Biol. 2008, 83, 1–39. [Google Scholar] [CrossRef]
- Meng, K.; Shi, Y.C.; Li, W.X.; Wang, J.; Cheng, B.J.; Li, T.L.; Li, H.; Jiang, N.; Liu, R. Testosterone Mediates Reproductive Toxicity in Caenorhabditis Elegans by Affecting Sex Determination in Germ Cells through Nhr-69/ Mpk-1/ Fog-1/ 3. Toxics 2024, 12. [Google Scholar] [CrossRef]
- Ellis, R.E. Sex Determination in Nematode Germ Cells. Sex Dev. 2022, 16, 305–322. [Google Scholar] [CrossRef] [PubMed]
- Redman, E.; Grillo, V.; Saunders, G.; Packard, E.; Jackson, F.; Berriman, M.; Gilleard, J.S. Genetics of Mating and Sex Determination in the Parasitic Nematode Haemonchus Contortus. Genetics 2008, 180, 1877–1887. [Google Scholar] [CrossRef] [PubMed]
- Sallé, G.; Doyle, S.R.; Cortet, J.; Cabaret, J.; Berriman, M.; Holroyd, N.; Cotton, J.A. The Global Diversity of Haemonchus Contortus Is Shaped by Human Intervention and Climate. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Doyle, S.R.; Tracey, A.; Laing, R.; Holroyd, N.; Bartley, D.; Bazant, W.; Beasley, H.; Beech, R.; Britton, C.; Brooks, K.; et al. Genomic and Transcriptomic Variation Defines the Chromosome-Scale Assembly of Haemonchus Contortus, a Model Gastrointestinal Worm. Commun. Biol. 2020, 3, 656. [Google Scholar] [CrossRef]
- Zheng, Y.; Young, N.D.; Wang, T.; Chang, B.C.H.; Song, J.; Gasser, R.B. Systems Biology of Haemonchus Contortus – Advancing Biotechnology for Parasitic Nematode Control. Biotechnol. Adv. 2025, 81. [Google Scholar] [CrossRef]
- Huynh, T.; McKean, E.L.; Hawdon, J.M. Mini-Baermann Funnel, a Simple Device for Cleaning Nematode Infective Larvae. J. Parasitol. 2022, 108, 403–407. [Google Scholar] [CrossRef]
- Brinzer, R.A.; McIntyre, J.R.; Britton, C.; Laing, R. The Parasitic Nematode Haemonchus Contortus Lacks Molybdenum Cofactor Synthesis, Leading to Sulphite Sensitivity and Lethality in Vitro. Int. J. Parasitol. 2025, 55, 117–128. [Google Scholar] [CrossRef]
- Tuerhong, R.; Tuersong, W.; Maimaiti, A.; Maimaitiyiming, H.; Zhang, Y.; Xuekelaiti, D.; Tuoheti, A.; Xin, L.; Abula, S. Genetic Diversity Analysis of Benzimidazole Resistance-Associated Genes in Haemonchus Contortus from Four Regions in Southern Xinjiang. Vet. Parasitol. 2025, 336, 110426. [Google Scholar] [CrossRef]
- Okino, C.H.; Bello, H.J.S.; Niciura, S.C.M.; Melito, G.R.; Cunha, A.F. da; Costa, E.C. da; de Campos, E.M.; Kapritchkoff, R.T.I.; Minho, A.P.; Esteves, S.N.; et al. Haemonchus Contortus Parasitic Stages Development and Host Immune Responses in Lambs of Different Sheep Breeds. Vet. Immunol. Immunopathol. 2025, 284, 110936. [Google Scholar] [CrossRef]
- Zarkower, D. Somatic Sex Determination. WormBook 2006, 1–12. [Google Scholar] [CrossRef]
- Antebi, A.; Yeh, W.H.; Tait, D.; Hedgecock, E.M.; Riddle, D.L. Daf-12 Encodes a Nuclear Receptor That Regulates the Dauer Diapause and Developmental Age in C. Elegans. Genes Dev. 2000, 14, 1512. [Google Scholar] [CrossRef]
- Antebi, A. Nuclear Receptor Signal Transduction in C. Elegans. Wormbook 2015, 1. [Google Scholar] [CrossRef] [PubMed]
- Blum, M.; Chang, H.Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro Protein Families and Domains Database: 20 Years On. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Wang, T.; Korhonen, P.K.; Young, N.D.; Nie, S.; Ang, C.S.; Williamson, N.A.; Reid, G.E.; Gasser, R.B. Dafachronic Acid Promotes Larval Development in Haemonchus Contortus by Modulating Dauer Signalling and Lipid Metabolism. PLoS Pathog. 2019, 15. [Google Scholar] [CrossRef] [PubMed]
- Mcdonnell, D.P.; Mangelsdorf, D.J.; Pike, J.W.; Haussler, M.R.; O’Malley, B.W. Molecular Cloning of Complementary DNA Encoding the Avian Receptor for Vitamin D. Science (80-. ). 1987, 235, 1214–1217. [Google Scholar] [CrossRef]
- Makishima, M.; Okamoto, A.Y.; Repa, J.J.; Tu, H.; Learned, R.M.; Luk, A.; Hull, M. V.; Lustig, K.D.; Mangelsdorf, D.J.; Shan, B. Identification of a Nuclear Receptor for Bile Acids. Science (80-. ). 1999, 284, 1362–1365. [Google Scholar] [CrossRef]
- Doniach, T.; Hodgkin, J. A Sex-Determining Gene, Fem-1, Required for Both Male and Hermaphrodite Development in Caenorhabditis Elegans. Dev. Biol. 1984, 106, 223–235. [Google Scholar] [CrossRef]
- Spence, A.M.; Coulson, A.; Hodgkin, J. The Product of Fem-1, a Nematode Sex-Determining Gene, Contains a Motif Found in Cell Cycle Control Proteins and Receptors for Cell-Cell Interactions. Cell 1990, 60, 981–990. [Google Scholar] [CrossRef]
- Mehra, A.; Gaudet, J.; Heck, L.; Kuwabara, P.E.; Spence, A.M. Negative Regulation of Male Development in Caenorhabditis Elegans by a Protein-Protein Interaction between TRA-2A and FEM-3. Genes Dev. 1999, 13, 1453–1463. [Google Scholar] [CrossRef]
- Ventura-Holman, T.; Lu, D.; Si, X.; Izevbigie, E.B.; Maher, J.F. The Fem1c Genes: Conserved Members of the Fem1 Gene Family in Vertebrates. Gene 2003, 314, 133–139. [Google Scholar] [CrossRef]
- Dawes, H.E.; Berlin, D.S.; Lapidus, D.M.; Nusbaum, C.; Davis, T.L.; Meyer, B.J. Dosage Compensation Proteins Targeted to X Chromosomes by a Determinant of Hermaphrodite Fate. Science (80-. ). 1999, 284, 1800–1804. [Google Scholar] [CrossRef]
- Meyer, B.J. Targeting X Chromosomes for Repression. Curr. Opin. Genet. Dev. 2010, 20, 179–189. [Google Scholar] [CrossRef]
- Chen, E.; Hermanson, S.; Ekker, S.C. Syndecan-2 Is Essential for Angiogenic Sprouting during Zebrafish Development. Blood 2004, 103, 1710–1719. [Google Scholar] [CrossRef]
- Tsoyi, K.; Osorio, J.C.; Chu, S.G.; Fernandez, I.E.; De Frias, S.P.; Sholl, L.; Cui, Y.; Tellez, C.S.; Siegfried, J.M.; Belinsky, S.A.; et al. Lung Adenocarcinoma Syndecan-2 Potentiates Cell Invasiveness. Am. J. Respir. Cell Mol. Biol. 2019, 60, 659–666. [Google Scholar] [CrossRef]
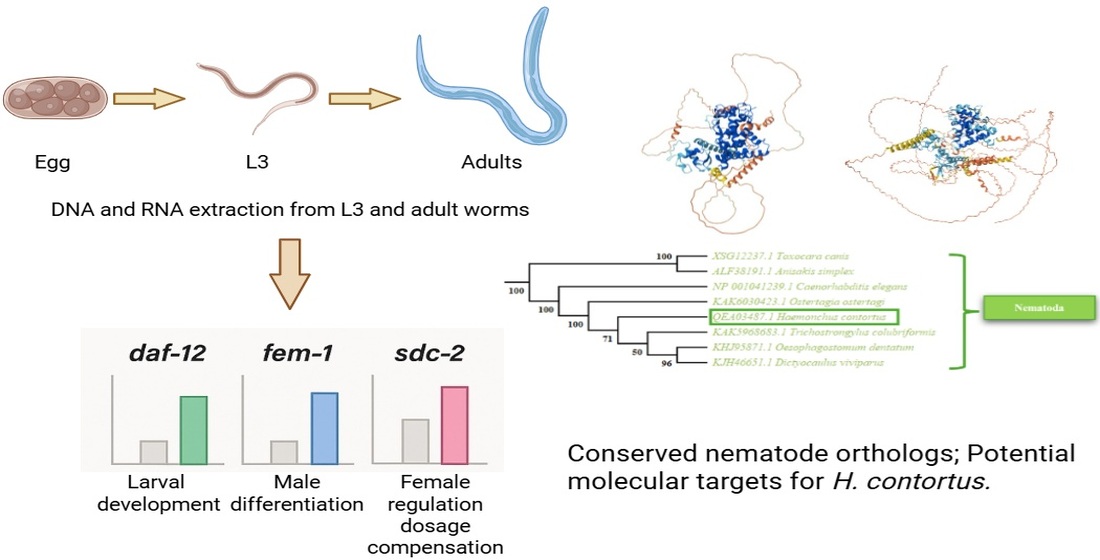
Figure 1.
Endpoint PCR detection of daf-12 (660 bp), sdc-2 (469 bp), and fem-1 (154 bp) in the H. contortus genome. C1: sdc-2; C2: daf-12; C 3: fem-1.
Figure 1.
Endpoint PCR detection of daf-12 (660 bp), sdc-2 (469 bp), and fem-1 (154 bp) in the H. contortus genome. C1: sdc-2; C2: daf-12; C 3: fem-1.
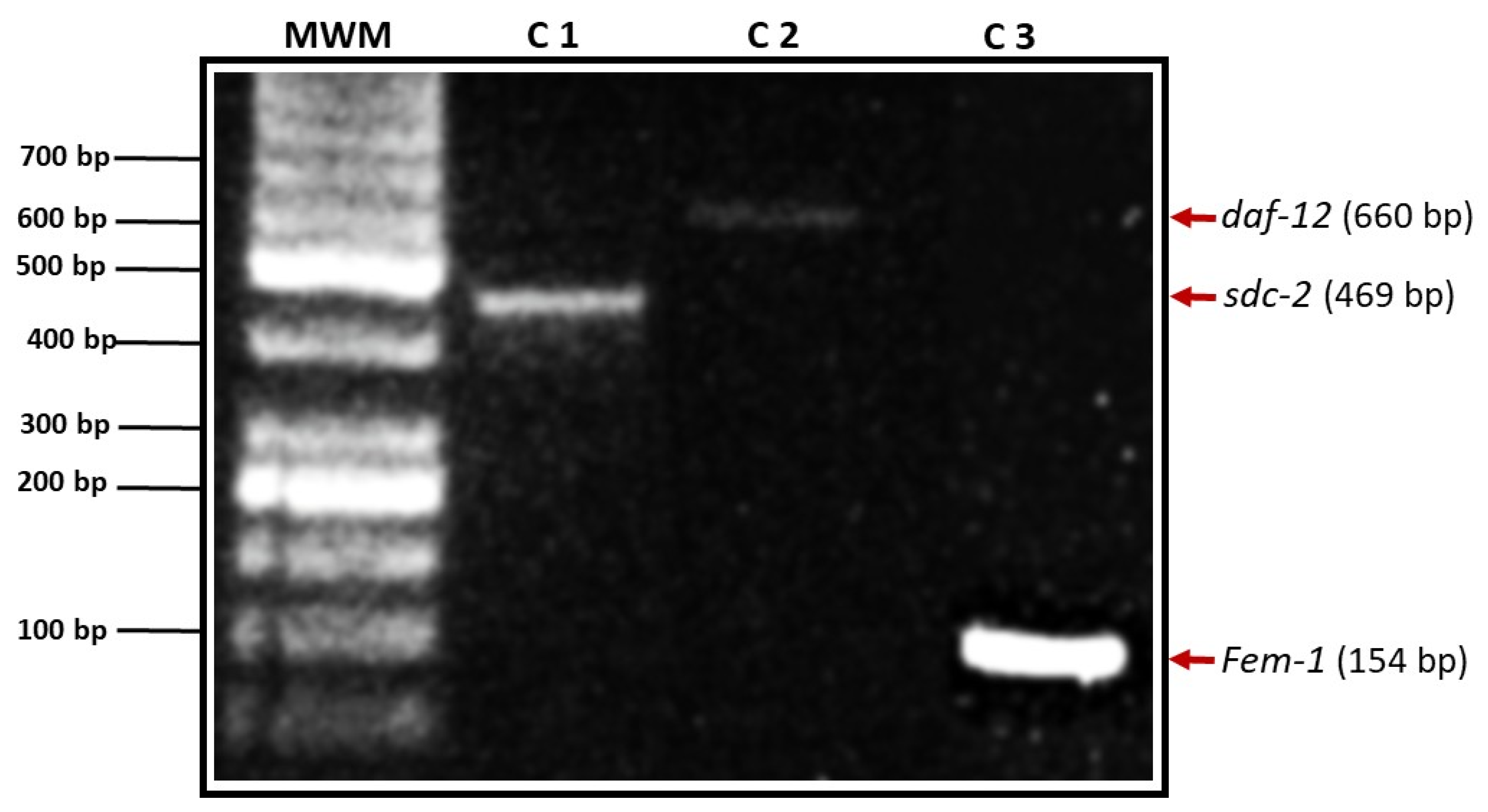
Figure 2.
RT-PCR for daf-12. (a) Relative expression of the daf-12 gene using the constitutive gene sod as control. The figure shows agarose gel electrophoresis with a 50 bp molecular weight marker (MPM). Samples correspond to third-stage larvae (L3), adult males, and adult females of H.contortus. (b) Optical densitometric analysis of daf-12. Statistical analysis using ANOVA revealed significant differences, indicated with **(p ≤ 0.01) and ***(p ≤ 0.001).
Figure 2.
RT-PCR for daf-12. (a) Relative expression of the daf-12 gene using the constitutive gene sod as control. The figure shows agarose gel electrophoresis with a 50 bp molecular weight marker (MPM). Samples correspond to third-stage larvae (L3), adult males, and adult females of H.contortus. (b) Optical densitometric analysis of daf-12. Statistical analysis using ANOVA revealed significant differences, indicated with **(p ≤ 0.01) and ***(p ≤ 0.001).
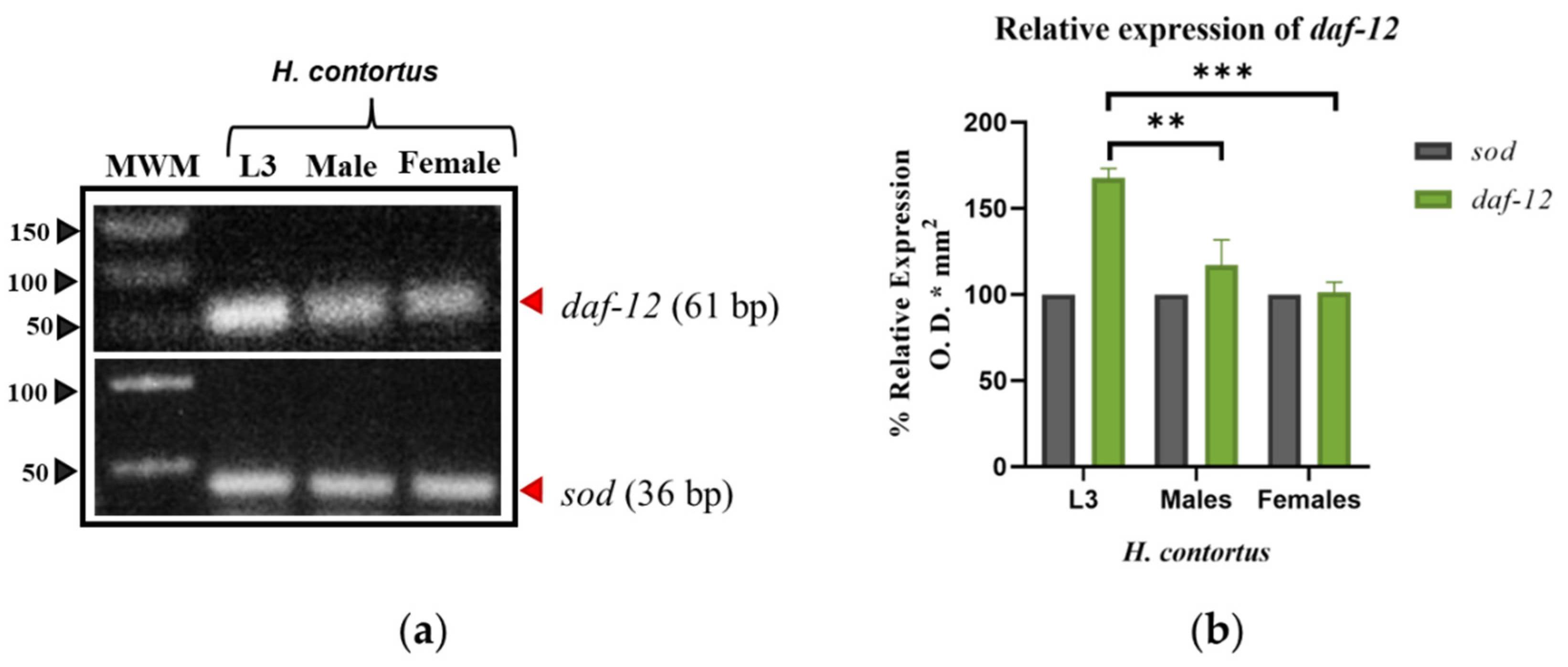
Figure 3.
RT-PCR for fem-1. (a) Relative expression of the fem-1 gene using the constitutive gene sod as control. The figure shows agarose gel electrophoresis with a 50 bp molecular weight marker (MPM). Samples correspond to third-stage larvae (L3), adult males, and adult females of H.contortus. (b) Optical densitometric analysis of fem-1. Statistical analysis using ANOVA revealed significant differences, indicated with ***(p ≤ 0.001) and ****(p ≤ 0.0001).
Figure 3.
RT-PCR for fem-1. (a) Relative expression of the fem-1 gene using the constitutive gene sod as control. The figure shows agarose gel electrophoresis with a 50 bp molecular weight marker (MPM). Samples correspond to third-stage larvae (L3), adult males, and adult females of H.contortus. (b) Optical densitometric analysis of fem-1. Statistical analysis using ANOVA revealed significant differences, indicated with ***(p ≤ 0.001) and ****(p ≤ 0.0001).
Figure 4.
RT-PCR for sdc-2. (a) Relative expression of the sdc-2 gene using the constitutive gene sod as control. The figure shows agarose gel electrophoresis with a 50 bp molecular weight marker (MPM). Samples correspond to third-stage larvae (L3), adult males, and adult females of H. contortus. (b) Optical densitometric analysis of sdc-2. Statistical analysis using ANOVA revealed significant differences, indicated with *(p ≤ 0.05) and **(p ≤ 0.01).
Figure 4.
RT-PCR for sdc-2. (a) Relative expression of the sdc-2 gene using the constitutive gene sod as control. The figure shows agarose gel electrophoresis with a 50 bp molecular weight marker (MPM). Samples correspond to third-stage larvae (L3), adult males, and adult females of H. contortus. (b) Optical densitometric analysis of sdc-2. Statistical analysis using ANOVA revealed significant differences, indicated with *(p ≤ 0.05) and **(p ≤ 0.01).
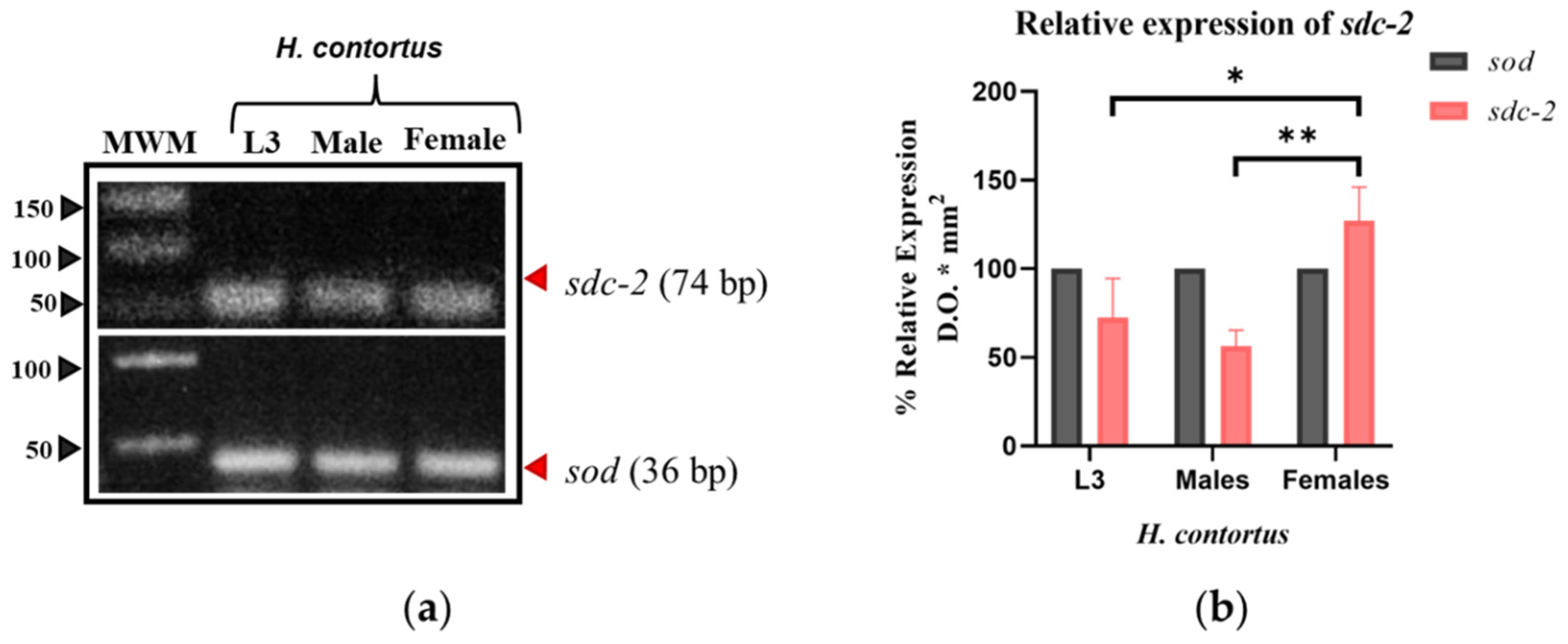
Figure 5.
In silico 3D model of DAF-12 predicted by AlphaFold and its structural validation. (a) The model displays a well-defined α-helical core (navy blue) with high confidence in the structural prediction (pLDDT > 90), while orange-colored termini indicate lower confidence regions (pLDDT < 50), likely corresponding to flexible or disordered segments. (b) ERRAT yielded an overall quality factor of 94.76%, and PROCHECK Ramachandran analysis confirmed that most residues were in favored regions.
Figure 5.
In silico 3D model of DAF-12 predicted by AlphaFold and its structural validation. (a) The model displays a well-defined α-helical core (navy blue) with high confidence in the structural prediction (pLDDT > 90), while orange-colored termini indicate lower confidence regions (pLDDT < 50), likely corresponding to flexible or disordered segments. (b) ERRAT yielded an overall quality factor of 94.76%, and PROCHECK Ramachandran analysis confirmed that most residues were in favored regions.
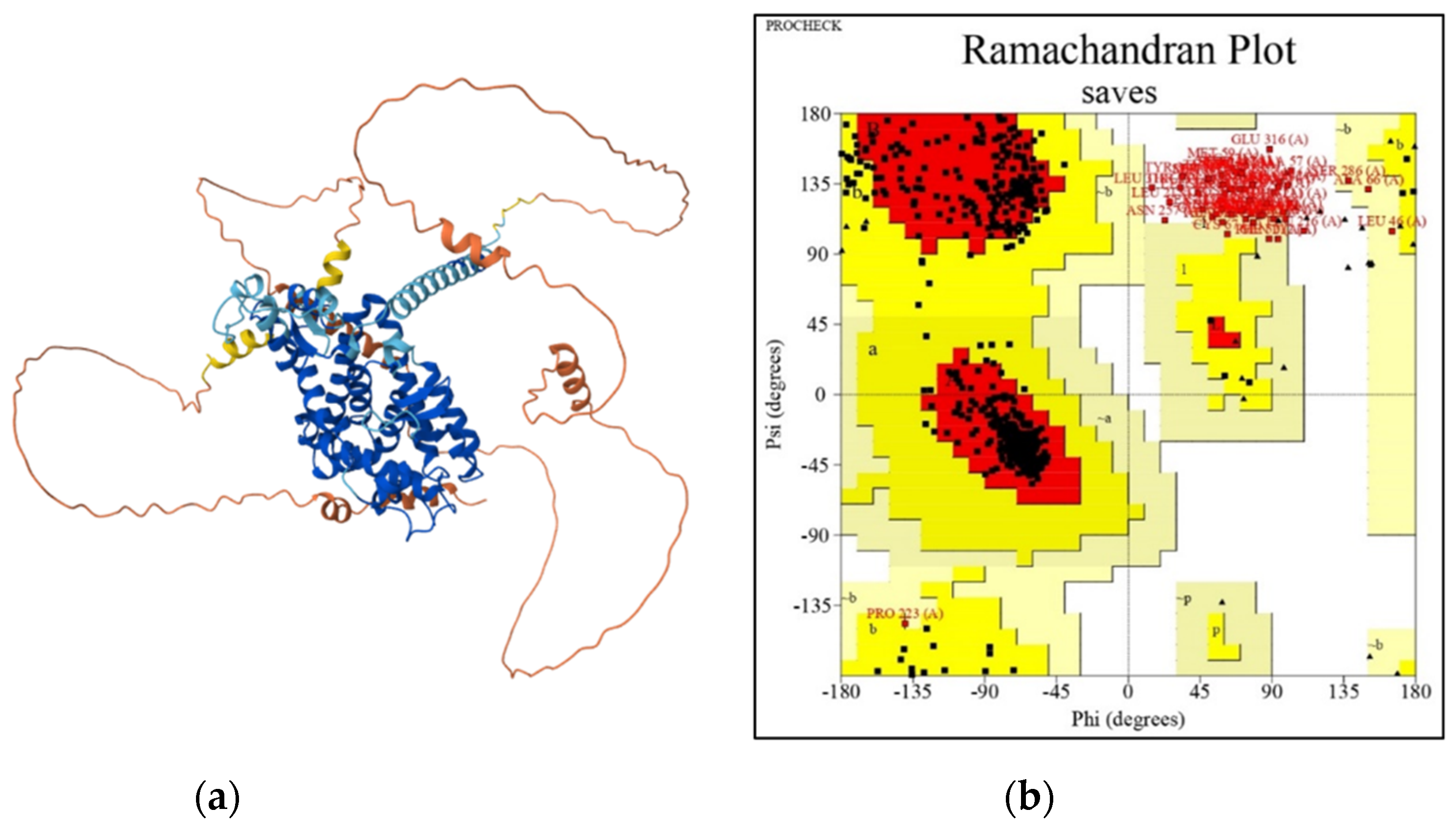
Figure 6.
In silico 3D model of FEM-1 predicted by AlphaFold and its structural validation. (a) The model revealed a repetitive and compact arrangement of α-helices with high confidence (pLDDT > 70), while terminal regions showed low-confidence segments (pLDDT < 50). (b) Validation with SAVES tools yielded an overall quality factor of 76.79% in ERRAT, and PROCHECK Ramachandran analysis showed 82.4% of residues in the most favored regions and 17% in additionally allowed regions.
Figure 6.
In silico 3D model of FEM-1 predicted by AlphaFold and its structural validation. (a) The model revealed a repetitive and compact arrangement of α-helices with high confidence (pLDDT > 70), while terminal regions showed low-confidence segments (pLDDT < 50). (b) Validation with SAVES tools yielded an overall quality factor of 76.79% in ERRAT, and PROCHECK Ramachandran analysis showed 82.4% of residues in the most favored regions and 17% in additionally allowed regions.
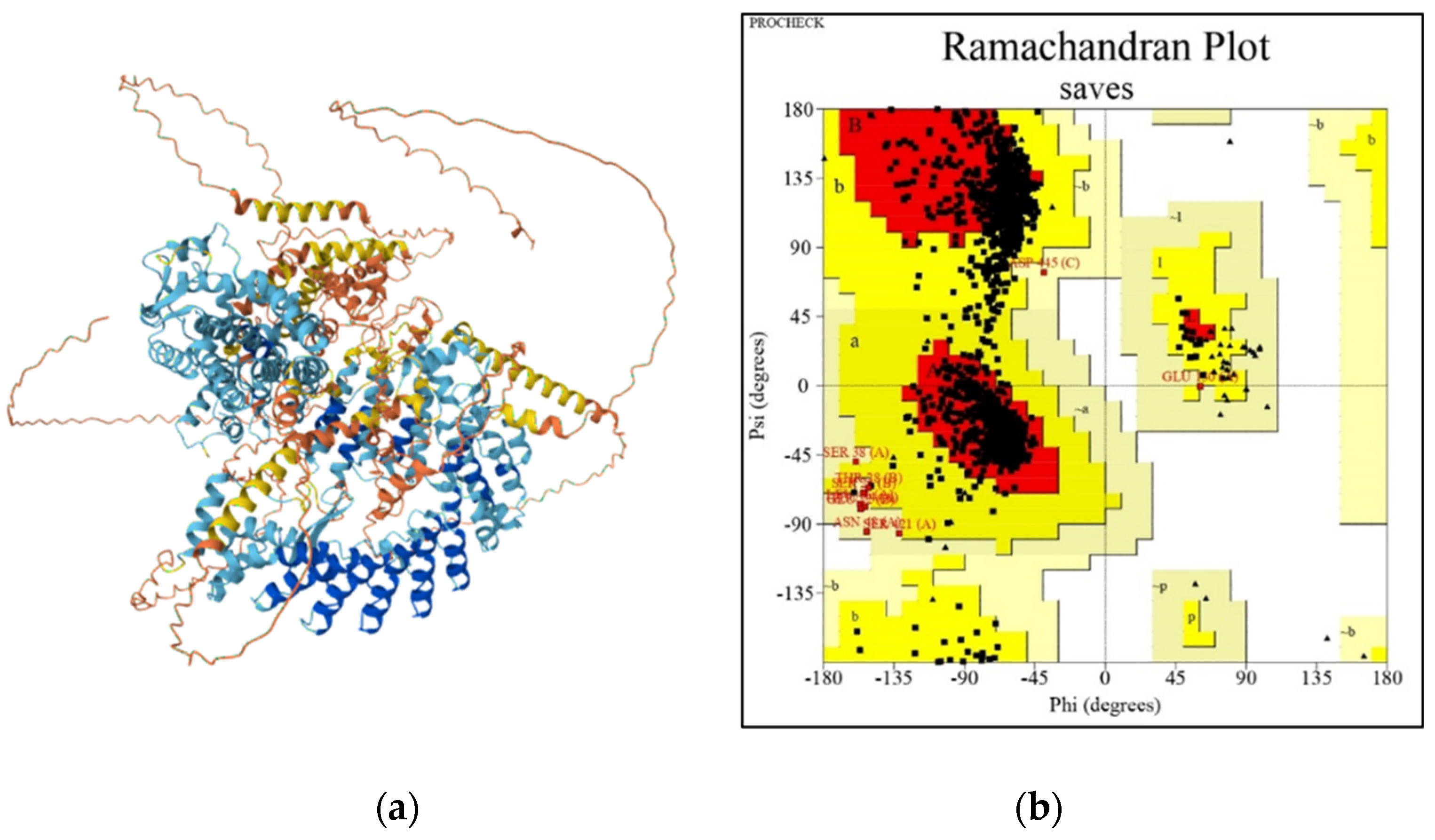
Figure 7.
In silico 3D model of SDC-2 predicted by AlphaFold and its structural validation. (a) The model revealed a repetitive and compact arrangement of α-helices with high confidence (pLDDT > 90), while terminal regions showed low-confidence segments (pLDDT < 90-70). (b) Validation with SAVES tools yielded an overall quality factor of 86.24% in ERRAT, and PROCHECK Ramachandran analysis showed 74.8% of residues in the most favored regions and 24.5% in additionally allowed regions.
Figure 7.
In silico 3D model of SDC-2 predicted by AlphaFold and its structural validation. (a) The model revealed a repetitive and compact arrangement of α-helices with high confidence (pLDDT > 90), while terminal regions showed low-confidence segments (pLDDT < 90-70). (b) Validation with SAVES tools yielded an overall quality factor of 86.24% in ERRAT, and PROCHECK Ramachandran analysis showed 74.8% of residues in the most favored regions and 24.5% in additionally allowed regions.
Figure 8.
Phylogenetic tree of DAF-12. Constructed by Neighbor-Joining with 1500 bootstrap replicates using MEGA 12; bootstrap support values are shown at the nodes.
Figure 8.
Phylogenetic tree of DAF-12. Constructed by Neighbor-Joining with 1500 bootstrap replicates using MEGA 12; bootstrap support values are shown at the nodes.
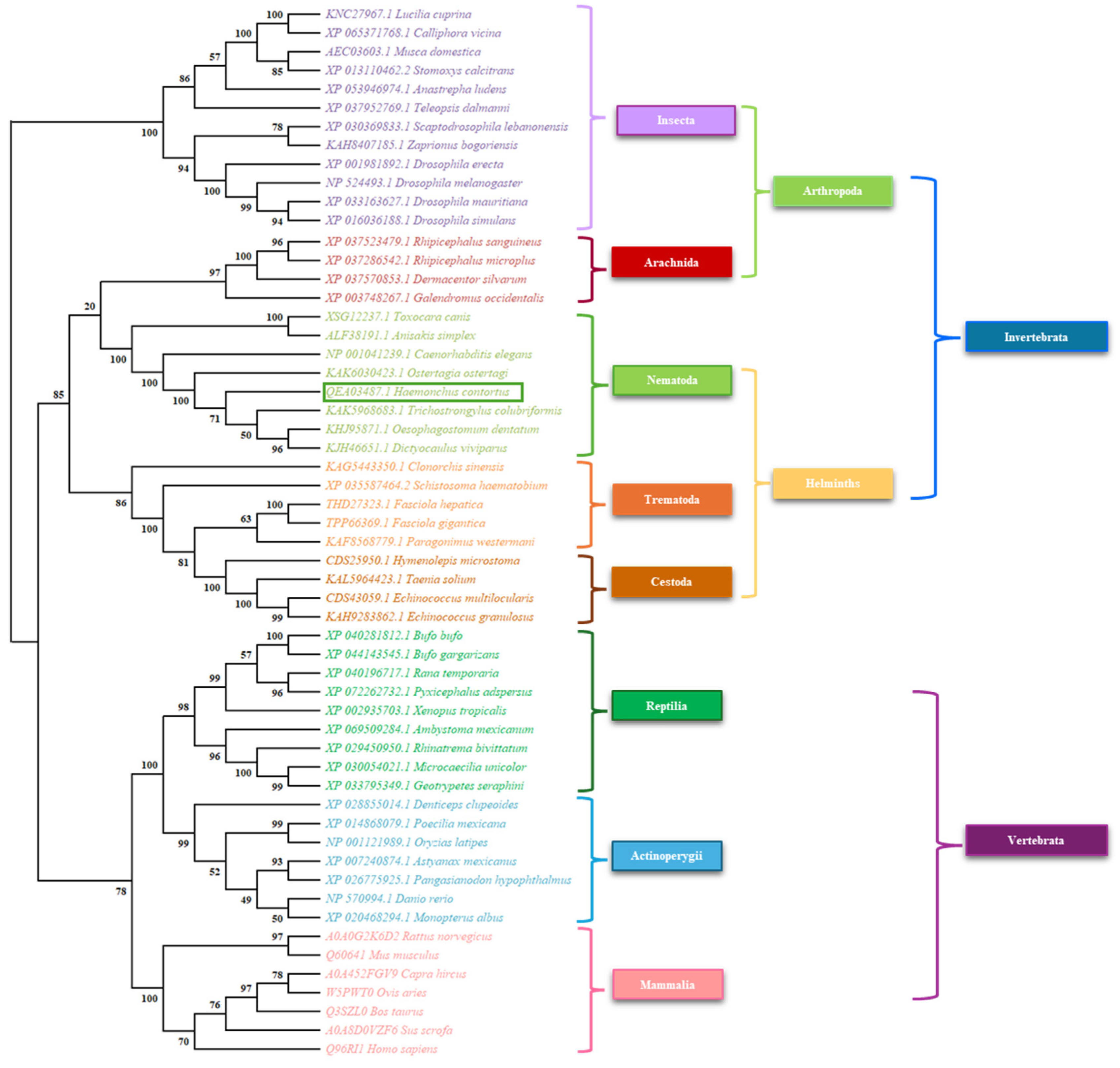
Figure 9.
Phylogenetic tree of FEM-1. Constructed by Neighbor-Joining with 1500 bootstrap replicates using MEGA 12; bootstrap support values are shown at the nodes.
Figure 9.
Phylogenetic tree of FEM-1. Constructed by Neighbor-Joining with 1500 bootstrap replicates using MEGA 12; bootstrap support values are shown at the nodes.
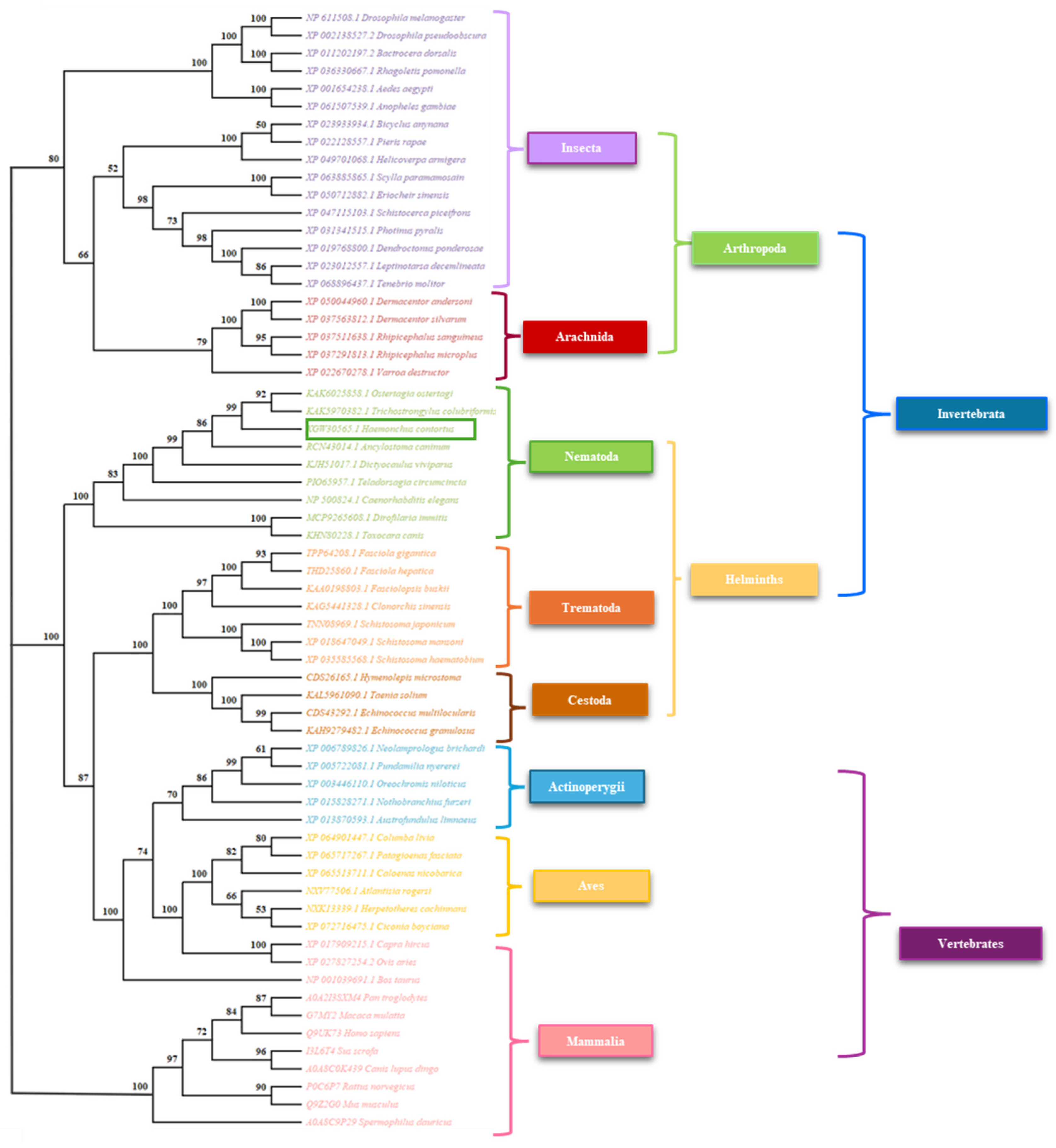
Figure 10.
Phylogenetic tree of SDC-2. Constructed by Neighbor-Joining with 1500 bootstrap replicates using MEGA 12; bootstrap support values are shown at the nodes.
Figure 10.
Phylogenetic tree of SDC-2. Constructed by Neighbor-Joining with 1500 bootstrap replicates using MEGA 12; bootstrap support values are shown at the nodes.

Table 1.
Oligonucleotides for the amplification of selected sexual differentiation genes of H. contortus.
Table 1.
Oligonucleotides for the amplification of selected sexual differentiation genes of H. contortus.
| Genes | Sequence Forward | Sequence reverse | Tm (°C) |
|---|---|---|---|
| sod | GCTGGCACTGATGATTTGGG | CGCCAGCATTTCCTGTCTTC | 62 |
| daf-12 | ATGCAAGGCGTTTTTCCGTC | CGACCATCGGCGAAATTGAC | 60 |
| fem-1 | GATGCGTTGAAGCTGTTGG | CACGAAGGTTACGGTTTGC | 60 |
| sdc-2 | CGAGTCCAGCGATTCATCCA | AGCTGTCGTTTGCGTCACTA | 60 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



