Submitted:
15 October 2025
Posted:
16 October 2025
You are already at the latest version
Abstract
The immune system plays a critical role in the development and progression of tumors. In recent years, despite the remarkable clinical successes achieved with immunotherapies such as CAR-T cell therapy and checkpoint inhibitors, treatment resistance continues to be a pervasive and critical barrier for a substantial portion of patients. The gut microbiota has been established as a critical determinant of responses to immunotherapy. Enriched with bioactive components such as polysaccharides, polyphenols, and flavonoids, edible and medicinal plants (EMPs) exhibit significant potential to enhance host immunity by reshaping the gut microbiota, increasing the production of microbiota-derived metabolites (e.g., short-chain fatty acids), strengthening the intestinal barrier, and reducing intestinal inflammation. The bioactive components derived from EMPs not only demonstrate substantial pharmacological activities but also serve dual roles: functioning either as inherent drug carriers or as effective modifiers for existing carrier systems, which facilitates targeted drug delivery to specific sites such as the liver and intestinal, enhancing therapeutic efficacy. In summary, this review highlights that bioactive components from EMPs hold significant promise for enhancing cancer immunotherapy by modulating complex interactions with the gut microbiota.
Keywords:
cancer immunotherapy
; gut microbiota
; bioactive components
; edible and medicinal plants
; drug carriers
1. Introduction
According to the GLOBOCAN 2022 data, approximately 20 million new cancer cases and 9.7 million cancer deaths were reported in world, which show that cancer seriously threaten human health [1]. The conventional treatment for cancer includes surgery, radiotherapy, chemotherapy and targeted therapy. However, the conventional treatment increases tumor recurrence and poor prognosis. Immunotherapy targets immune cells and their interactions with tumor cells [2] and can treat metastatic malignant tumors and prolong the survival time of patients with good prognosis. However, due to the immunosuppressive tumor microenvironment (TME) and the immune escape mechanism of tumor cells, immunotherapy has limited efficacy in tumors [3]. At the same time, in the process of cancer immunotherapy, the emergence of drug resistance, immune-related adverse reactions and heterogeneity make immunotherapy face great clinical challenges.
The gastrointestinal tract contains 60-80% of the body’s immune cells and plays a crucial role in maintaining immune homeostasis [4]. Research demonstrates that gut microbiota and metabolites not only modulate intestinal immunity but also play a role in systemic cancer immunomodulation [4,5]. Recent studies show that cancer immunotherapy, including immune checkpoint blockade (ICB), CAR-T and cancer vaccines, seems to have different therapeutic effects in different individuals, which is related to the gut microbiome [6,7,8,9,10]. Therefore, gut microbiota, as the “second genome” of the human body, represents a highly promising target for precision cancer immunotherapy.
Edible and medicinal plants (EMPs), as an indispensable and important part of traditional Chinese medicine, have functions of both nutritional and pharmacological. Due to high safety, minimal side effect, abundant functional components including polysaccharides, flavonoids, and saponins and multi-pathway pharmacological, EMPs display unique advantages in the field of cancer treatment, improving quality of life for patients [11]. Emerging evidence demonstrates that EMPs can modulate antitumor immune responses through gut microbiota regulation [12]. In this review, we emphasize the crosstalk among the active compounds from EMPs, gut microbiome, cancer immune response, and cancer immunotherapy, providing insights for the development of novel immunotherapeutic strategies. Furthermore, the article examines current research advancements in utilizing active compounds from EMPs to engineer drug delivery systems for tumor therapy, offering translational insights for clinical applications.
2. Bioactive Compounds from EMPs and Regulation of Gut Microbiome
EMPs have been widely utilized in China, Korea, Japan, and other countries for treating various diseases, including cancer [13], inflammatory disorders [14], and neurodegenerative disease [15,16]. According to chemical structures, bioactive compounds from EMPs are categorized as polysaccharides, flavonoids, saponins, and polyphenols, alkaloids, terpenoid, phenolic compounds, coumarin, essential oil and volatile oil. Extensive research has shown that polysaccharides, saponins, and polyphenols found in MFH plants enhance anti-tumor immunity by directly or indirectly modulating immune cell activity and the gut microbiota [12,17,18,19] (Figure 1).
2.1. Polysaccharides
Polysaccharides are a class of natural bioactive compounds abundantly present in EMPs, including Astragalus membranaceus [20], Panax ginseng C. A. Meyer [21,22], and Lycium chinense Miller [23,24], demonstrating unique anti-tumor activities. Increasing evidence demonstrates that polysaccharides can effectively regulates both the structural composition and metabolic activities of gut microbiota, maintaining microbial homeostasis, preserving intestinal barrier integrity, and ultimately enhancing gut immunity [25,26]. Ginseng polysaccharides (GPs), bioactive components of Panax ginseng, enhanced PD-1 blockade therapy by increasing the microbial metabolites valeric acid and decreasing the ratio of Kyn/Trp, which collectively increased CD8+ T cell recruitment while inhibiting Treg-mediated immunosuppression. Furthermore, GPs modulated gut microbial metabolism and composition, particularly by promoting the abundance of Parabacteroides distasonis and Bacteroides vulgatus, key bacterial genus involved in ginsenoside biotransformation, which was significantly associated with therapeutic response and prognosis. Clinical investigations have demonstrated that GPs improves therapeutic response to PD-1 checkpoint blockade, potentially overcoming immunotherapy resistance [27]. The water-soluble polysaccharides extracted from the spore bodies of Ganoderma lucidum polysaccharides (GLP) inhibit the TLR4/MyD88/NF-κB signaling pathway, improving the microbial flora imbalance induced by AOM/DSS, increasing the production of short-chain fatty acids, and alleviating endotoxemia. In addition, GLP can significantly improve the intestinal barrier function, inhibit the infiltration of macrophages, and suppress colonic inflammation and tumor occurrence [22]. In addition, emerging research indicates that polysaccharides such as Lycium barbarum polysaccharides[28], Lentinan edodes polysaccharides [29] and Astragalus polysaccharides (APS) [30] modulate gut microbiota and their derived metabolites, promoting the restoration of immune function. These findings demonstrate that polysaccharides can serve as adjuvants to improve intestinal and immunosuppression.
2.2. Polyphenols
Polyphenols, known for their potent antioxidant and anti-inflammatory properties, are one of the most important and widely occurring bioactive components in EMPs. Flavonoids, phenolic acids, lignans, and stilbenes are the four major classes of polyphenols [31]. Among these, flavonoids are the most abundant, followed by phenolic acids.
Flavonoids exhibit diverse biological activities and pharmacological effects, including potent antioxidant, anti-inflammatory, and anti-tumor effects. Based on their core structures and substituents, flavonoids can be classified into isoflavones, neoflavonoids, flavones, flavonols, flavanones, flavanonols, flavanols, chalcones [32]. Accumulating research suggests that flavonoids extracted from EMPs, such as kaempferol (a flavonol) and calycosin (an isoflavone from Astragalus membranaceus), can enhance cancer immunotherapy by activating antitumor immune responses through remodeling the gut microbial diversity and modulating microbial metabolites [33,34,35]. Inflammatory bowel disease (IBD) is a chronic intestinal inflammatory disorder with the potential to progress to colorectal cancer (CRC) [36]. Luteolin, a bioactive flavonoid derived from EMPs such as honeysuckle (Lonicera japonica), astragalus (Astragalus membranaceus), and angelica (Angelica sinensis), demonstrates significant therapeutic effects against IBD [37,38]. Its mechanisms of action include: (1) modulating T-cell subset differentiation [39], (2) downregulating pro-inflammatory mediators, (3) activating antioxidant pathways [40,41], (4) enhancing gut microbiota composition [42,43], and (5) restoring intestinal mucosal barrier function through specific molecular signaling pathways [44]. These multifaceted actions collectively ameliorate IBD symptoms and consequently suppress CRC development. However, the majority of polyphenols that escape absorption in the upper gastrointestinal tract travel directly to the colon, where they are metabolized by gut microbiota into a spectrum of smaller phenolic acids. These metabolites exhibit greater biological activity and higher absorption efficiency. For example, ellagic acid is converted into urolithins, which significantly enhance gut barrier function, reduce inflammation, and create a systemically favorable environment for antitumor immunity [45]. Furthermore, resveratrol, a natural stilbenes compound, enhance antitumor immune responses by modulating the abundance and diversity of gut microbiota, which contributes to improve the efficacy of tumor immunotherapy [46].
Additionally, other polyphenolic compounds such as curcumin can remodel the gut microbiota composition by enriching beneficial bacteria, which are closely associated with the maintenance of intestinal barrier integrity, anti-inflammatory effects, the production of beneficial metabolites and tumor microenvironment remodeling [47,48,49].
2.3. Saponins
Saponins are a class of bioactive compounds ubiquitously present in EMPs. These structurally complex phytochemicals exhibit diverse biological activities attributable to their intricate chemical architectures, including immunomodulation [50,51], cardiovascular protection [52,53], antioxidant effects [54,55], and antitumor properties[56,57,58]. Based on their aglycone structures, saponins are primarily classified into two major categories: triterpenoid saponins (such as ginseng, licorice, astragalus, and platycodon) and steroidal saponins (like ophiopogon japonicus and Chinese yam). Among these, triterpenoid saponins are particularly notable for their immunomodulatory and antitumor activities, such as ginsenoside and astragaloside. Emerging evidence suggests that multiple ginsenosides (such as Rh2 [59], Rg1 [60] and Rk3 [61]) regulate gut microbial diversity and promote anti-tumor immune responses, inhibiting tumor growth. For example, Rk3 ameliorates gut microbiota dysbiosis by enriching beneficial bacteria such as Akkermansia muciniphila and Barnesiella intestinihominis while eliminating pathogenic Desulfovibrio species, restoring intestinal barrier integrity. Mechanistically, Rk3 modulates chemokine and inflammatory cytokine production through regulation of innate lymphoid cells and T helper 17 (Th17) cell signaling pathways, significantly suppressing colonic tumorigenesis [61]. Moreover, recent studies revealed that Astragaloside IV(AS-IV) exhibit potent immunomodulatory and gastrointestinal protective effects by alleviating mucosal inflammation and restoring intestinal barrier integrity during different stages of inflammatory bowel disease (IBD) progression to colorectal cancer. AS-IV exerts their therapeutic effects through multiple mechanisms, including targeting key immune signaling pathways (e.g., NF-κB and PPAR) and promoting M1 macrophage polarization, suppressing tumor growth and metastasis via multi-targeted actions, which offers novel strategies for future drug development [62].
3. Synergistic Effects of the Bioactive Compounds from EMPs on Immunotherapy via Gut Microbiota Modulation
To counteract tumor immune evasion during cancer progression, various immunotherapies have been developed [2]. However, tumor immunotherapy faces substantial limitations and challenges. Immune checkpoint blockade therapies, particularly PD-1/PD-L1 inhibitors, demonstrate clinical efficacy in only approximately <30% of patients, with a subset of responders eventually experiencing disease recurrence [63]. Meanwhile, chimeric antigen receptor (CAR) T-cell therapy exhibits limited therapeutic potential against solid tumors [64]. Furthermore, immunotherapeutic approaches are frequently associated with severe immune-related adverse events and the development of treatment resistance. In recent years, accumulating evidence from both preclinical and clinical research highlights the crucial role of the gut microbiota in modulating patient response to immunotherapy [8,65,66]. Research indicates that the gut microbiome-derived metabolites and secreted molecules enhance immunotherapy efficacy [67,68]. With advantages including natural derivation, multi-target modulation, and low toxicity, bioactive compounds from EMPs can synergize with immunotherapy by modulating gut microbiota, which reduces treatment toxicity while improving antitumor efficacy, leading to better patient outcomes. Emerging research reveals that active ingredients from medicinal foods potentiate cancer immunotherapy via modulating gut microbiota composition, promoting the production of beneficial metabolites such as short-chain fatty acids (SCFAs) (Figure 2), and improving intestinal barrier function to reduce inflammation (Figure 3).
3.1. Gut Microbiota Composition Remodeling
Emerging evidence suggests that specific gut bacteria, notably Bacteroides fragilis, Bacteroides thetaiotaomicron, Bifidobacterium species, Akkermansia muciniphila, and Faecalibacterium spp., may enhance the efficacy of cancer immunotherapy, as observed in preclinical studies and patient cohorts [8,9,69,70]. Consequently, therapeutic strategies targeting gut microbiota modulation have been proposed to enhance treatment efficacy in oncology. Immune checkpoint inhibitors (ICIs) represent the most extensively studied form of immunotherapy [71,72,73]. However, their low response rates in cancer patients remain a major therapeutic challenge.
As the dominant effector population in anti-tumor immunity, CD8+ T cells have shown remarkable therapeutic efficacy in various immunotherapeutic strategies. Anti-PD-1 antibodies blockade activated CD8+ T cells to secrete perforin and granzyme B, mediating tumor cell death [74,75]. Bioactive compounds from EMPs have been shown to modulate the microbiota-immune axis, ultimately enhancing the efficacy of ICIs. For instance, GPs can enhance immune surveillance by increasing the abundance of Lachnospiraceae bacteria, which in turn suppresses myeloid-derived suppressor cells (MDSCs) and promotes the infiltration and activation of CD8⁺ T cells [76,77]. Furthermore, when combined with αPD-1 monoclonal antibody (mAb) therapy, GPs reshape the gut microbiota and improve the sensitivity of non-small cell lung cancer (NSCLC) patients to anti-PD-1 immunotherapy [27]. In addition, recent research showed that Chinese yam polysaccharide (CYPs), derived from medicinal food plants, exhibit immunomodulatory and antitumor properties. 16S rRNA sequencing revealed that combined treatment with CYPs and αPD-1 monoclonal antibody (mAb) enriched beneficial bacteria (e.g., *Clostridium_UCG-014* and Actinobacteria) while reducing pathogenic taxa (including Enterobacteriaceae and Desulfovibrionaceae). Furthermore, this combination therapy remodeled the tumor microenvironment (TME) by suppressing immunosuppressive M2 macrophages (CD206+ subset) and enhancing cytotoxic CD8+ T-cell infiltration, improving the response to anti-PD-1 immunotherapy in CRC patients [78]. Bilberry anthocyanin extracts significantly increased the relative abundance of Clostridia and Lactobacillus johnsonii while enhancing effective microbial community diversity, consequently improving the efficacy of ICIs [79].
Based on the above research, it is suggested that bioactive compounds from EMPs can enhance the efficacy of tumor immunotherapy and inhibit tumor progression by modulating the composition of the gut microbiota (promoting the abundance of beneficial bacteria while suppressing harmful bacteria) and regulating the tumor immunosuppressive microenvironment. This highlights the potential of bioactive compounds from EMPs in immune modulation for microbiota-targeted cancer therapies.
3.2. Microbial Metabolites
Gut microbiota-derived metabolites modulate the development and function of various immune cells while reshaping the tumor microenvironment [80,81,82]. Immunotherapy combined with microbial metabolites can effectively activate the immune system, eliminate tumor cells and overcome drug resistance, mitigating severe treatment-related side effects [83]. Therefore, targeting gut microbiota-derived metabolites represents a promising therapeutic strategy to enhance tumor immunotherapy response rates in patients.
Research indicates that non-digestible bioactive compounds from EMPs, such as polysaccharides, are fermented by specific gut microbiota (e.g., Bifidobacterium and Lactobacillus), leading to the production of short-chain fatty acids (SCFAs) like propionate and butyrate [84,85,86]. SCFAs regulate immune cells function to augment antitumor immune responses and maintain intestinal homeostasis through mechanisms involving the activation of G-protein-coupled receptors (GPRs) and inhibition of histone deacetylases (HDAC) [87,88,89,90]. In addition, polyphenols-EMPs components, such as pomegranate ellagitanninsare, are metabolized by gut microbiota into ellagic acid and further converted into urolithins that can reshape the gut microbiota and modulate the tumor microenvironment. Moreover, when combined with anti-PD-1 antibody therapy, they have been shown to inhibit the growth of colon cancer, thereby supporting more effective anticancer treatment [91,92]. Together, these findings reveal how interactions between gut microbiota and EMPs may offer novel strategies for developing natural adjuvants to enhance cancer immunotherapy.
3.3. Intestinal Barrier Dysfunction and Intestinal Inflammation
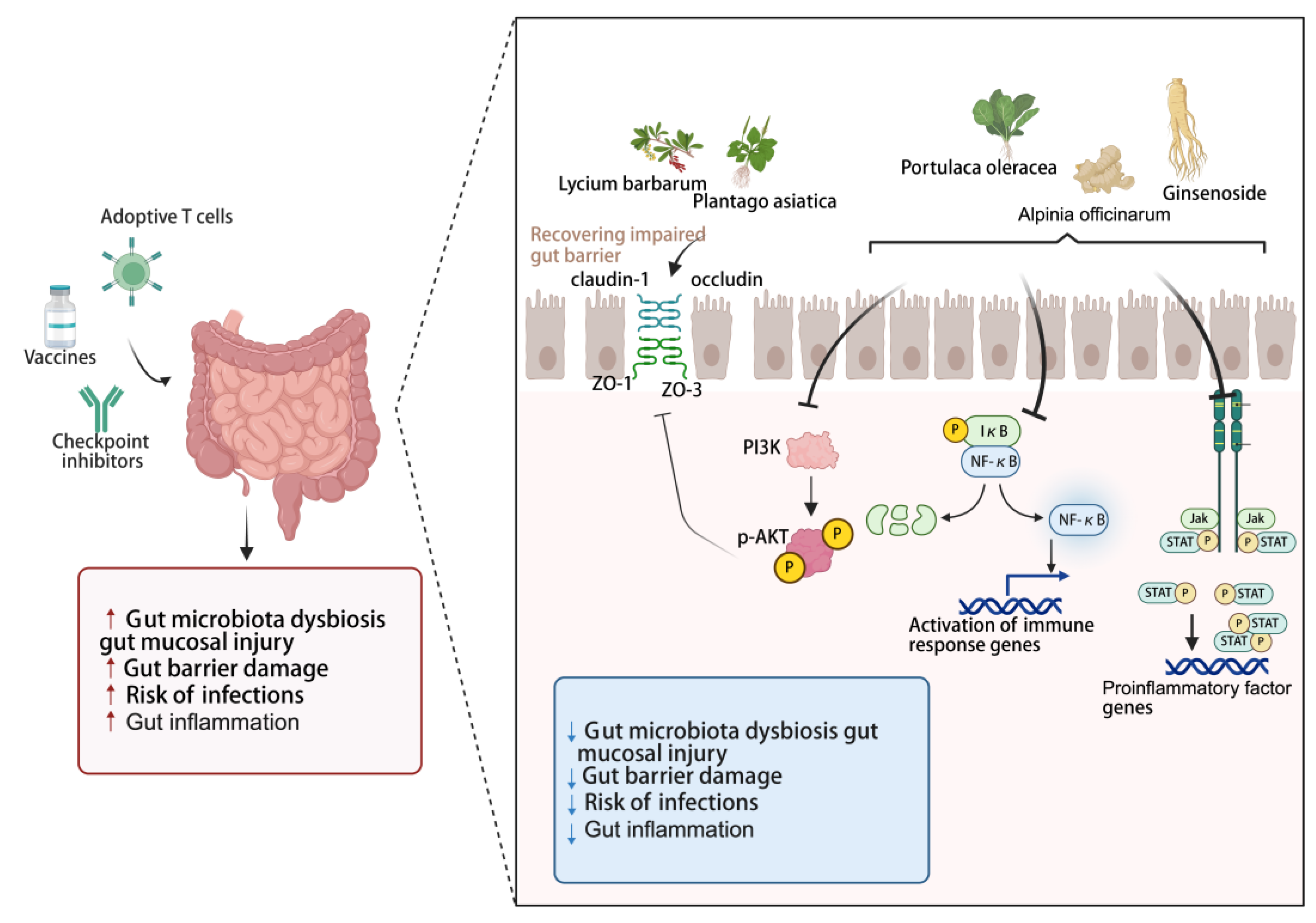
Cancer immunotherapy, particularly ICIs, frequently induces immune-related adverse events (irAEs), many of which are strongly linked to both gut microbiota dysbiosis gut mucosal injury, intestinal barrier damage, bacterial translocation resulting from increased gut permeability and systemic inflammation [93,94,95,96]. A healthy gut microbiome and an intact intestinal barrier are essential for maintaining immune homeostasis [97,98]. Once the barrier is compromised, systemic chronic inflammation is triggered, which promotes the expansion of immunosuppressive cells—such as myeloid-derived suppressor cells (MDSCs) and regulatory T cells (Tregs). This process fosters an immunosuppressive TME, ultimately leading to either primary or adaptive resistance to ICIs [99,100,101]. Therefore, targeting the intestinal barrier and inflammation represents a viable strategy for sensitizing tumors to immunotherapy.
The intestinal barrier acts as a crucial frontline defense against external pathogens and toxins [102]. It is well established that numerous components from EMPs directly or indirectly contribute to the intestinal barrier integrity [103,104,105]. Polysaccharides, known for their favorable safety profile, are effective in reducing inflammation and suppressing the growth of malignant tumors, which represent a promising adjuvant therapy for a variety of disorders associated with intestinal barrier injury [104]. Numerous bioactive polysaccharides, including those derived from Plantago asiatica [106], Lycium barbarum [107] and Spirulina [108] have been shown to modulate the expression of tight junction proteins (including occludin, claudin-1, ZO-1, and ZO-3), which collectively contribute to the amelioration of epithelial barrier dysfunction.
Additionally, Studies indicate that approximately 60% of cancer patients treated with ICIs develop severe treatment-limiting toxicities [109,110], which are closely associated with inflammation-related adverse events induced during the therapy [111,112,113]. Colitis is a common and severe adverse effect of ICIs therapy. Therefore, developing targeted therapeutic strategies for intestinal inflammation—aimed at effectively managing toxicity while preserving or even enhancing the antitumor efficacy of ICIs—represents a promising approach. As potent natural anti-inflammatory agents, various bioactive compounds from EMPs serve such as astragalosides [114], curcumin [115,116], ginsenoside Rh2 [117], Alpinia officinarum Hance polysaccharides [118], and Portulaca oleracea L. polyphenols [119], work synergistically through multiple targets and pathways, precisely modulating key signaling pathways associated with intestinal inflammation, including PI3K/AKT, NF-κB, MAPK, and JAK/STAT—thereby effectively alleviating gut barrier disruption and reducing intestinal inflammation. Clinical studies have also shown that certain medicinal food homologous ingredients, such as curcumin, can serve as an adjunctive therapy to significantly improve the clinical remission rate and endoscopic improvement scores in patients with mild to moderate ulcerative colitis [120].
Based on these findings, it has been elucidated that natural bioactive compounds from EMPs can enhance tumor immunotherapy by modulating the gut microbiota composition, regulating microbial metabolites, improving intestinal barrier dysfunction, and alleviating intestinal inflammation, which work synergistically to mitigate adverse effects induced by immunotherapy, increase patient response rates, and improve overall treatment efficacy.
4. Construct DDS Based on Components from EMPs
EMPs compounds including polysaccharides, polyphenols, flavonoids and saponins exhibit considerable potential in enhancing tumor immunotherapy by modulating the gut microbiota–immune axis. However, their widespread clinical translation and application are substantially limited by inherently low bioavailability, which results from poor water solubility, chemical instability, and inefficient intestinal absorption. Recent studies have confirmed that advanced drug delivery systems (DDS) strategies, such as nanotechnology, targeted delivery systems, and biotransformation, can significantly enhance the anti-tumor efficacy of EMPs bioactive compounds [121,122,123]. These approaches not only reduce side effects and improve patient compliance but also hold great promise for developing these compounds into effective adjuvant interventions for tumor immunotherapy. However, DDS still face numerous challenges and problems. EMPs components are now being extensively explored as “smart materials” for constructing or functionalizing drug delivery systems (DDS), rather than merely serving as the encapsulated “cargo,” owing to their inherent bioactivity, excellent biocompatibility, and unique physicochemical properties [124,125]. Additionally, their ability to modulate immune cell function enables an integrated strategy of “delivery and therapy in one.
4.1. Advantages of Bioactive Compounds from EMPs for DDS Carriers
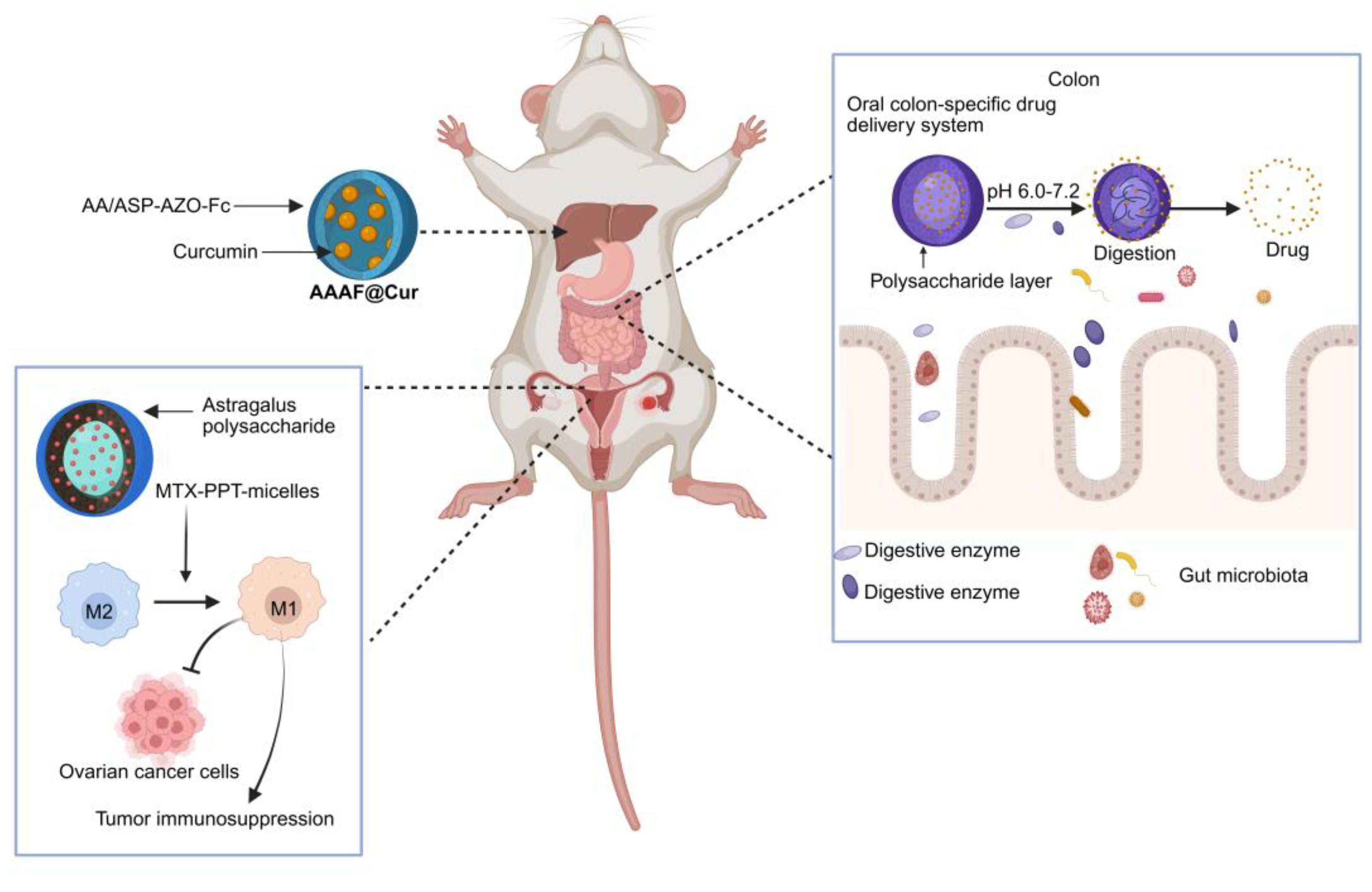
An ideal DDS should be capable of precisely delivering drugs to the diseased site. Nevertheless, Dai et al. revealed that due to target heterogeneity, the delivery efficiency of nanoparticles to cancer cells is abysmally low, with only approximately 0.0014% of the ligand-modified nanoparticles successfully binding to them [126]. Utilizing the inherent targeting capabilities of natural plant active ingredients such as polysaccharides, or further functionalizing them, constructed novel DDS, which significantly enhance drug targeting precision and increase drug accumulation at the site of disease. For example, Liu et al. developed a novel nanocarrier material, AA/ASP-AZO-Fc, by leveraging the intrinsic liver-targeting property of Angelica sinensis polysaccharide (ASP). The resulting polymeric micelle system effectively delivered curcumin to hepatic tissues, demonstrating a targeted therapeutic strategy for hepatocellular carcinoma [127]. Furthermore, nanotechnology can be integrated with immunotherapy to enhance tumor-targeted treatment [128]. Liu et al. designed novel methotrexate (MTX)-modified PPT nanoparticles loaded with Astragalus polysaccharide micelles. This system can safely and effectively target ovarian cancer cells and achieve precise drug release. Additionally, it exerts immunomodulatory effects on tumor-associated macrophages to combat tumor cells, thereby enhancing the efficacy of immunotherapy for ovarian cancer [129].
Indeed, the long-term biosafety of many nanocarrier materials remains to be fully elucidated. Nel et al. highlighted that the intrinsic properties of nanomaterials could lead to adverse consequences, primarily through harmful interactions with biological systems and the environment, potentially resulting in toxicity [130]. Among these, liposomes have been successfully utilized as nanocarriers in drug delivery systems. With various formulations available, they have entered clinical applications for targeted cancer therapy [131,132]. However, clinical studies have revealed that the application of liposomes often results in low targeting efficiency and elevated systemic toxicity, resulting in adverse side effects [133,134,135,136]. Natural biopolymers are recognized for their established safety profile, excellent biodegradability, non-toxic degradation products and favorable tolerability, make them promising candidates for replacing synthetic materials in DDS [125,137]. The enhancement of the physicochemical properties of DDS by bioactive compounds such as ginsenosides can increase the bioavailability of active pharmaceutical ingredients, potentiating their antitumor therapeutic effects [125,138]. For instance, ginsenoside Rh2-liposomes can replace cholesterol to enhance stability and reduce potential side effects, while significantly prolonging their circulation time in vivo. Their key advantages include active targeting of tumor cells, promoted drug accumulation at tumor sites, and effective remodeling of the TME to reverse immunosuppression, potently inhibiting tumor growth [139]. Bioactive food-derived components serving as DDS carriers offer a multifunctional and innovative platform for anticancer drug delivery.
4.2. Intestinal-Targeting Release of the Medication
Oral administration is the most preferred route of drug delivery due to its convenience and high patient compliance [140]. However, the highly acidic environment of gastric acid can lead to its inactivation, degradation by gastrointestinal enzymes may reduce its bioavailability, and its premature release in non-target areas could potentially cause systemic side effects [141]. In order to achieve efficient and precise delivery of these active ingredients to specific intestinal sites and control their release behavior, colon targeted drug delivery have emerged and become a research hotspot in the field of pharmaceutics. It is crucial for treating inflammatory bowel disease (IBD) and colon cancer, as well as for enhancing the bioavailability of easily degradable drugs for systemic absorption, making it a vital approach [142,143]. Delivery systems utilizing edible bioactive compounds which offer excellent biocompatibility, biodegradability, low toxicity, and ease of functional modification have emerged as ideal carriers for colon-targeted drug delivery [143]. Edible bioactive ingredients exhibit diverse responsive properties, allowing them to be naturally engineered to respond to specific intestinal conditions such as pH and microbial enzymes, enabling targeted drug release at disease sites. Due to their high safety profile, structural versatility, and the fact that they can only be degraded by specific enzymes produced by colonic microbiota after reaching the intestine, natural polysaccharides are widely used in the construction of oral colon-specific drug delivery systems for targeted colonic release of therapeutics [144]. In addition, leveraging the pH sensitivity of polysaccharides, nano-sized polysaccharide-based drug carriers can be utilized to construct pH-responsive delivery systems, enabling targeted intestinal drug release and controlled therapeutic delivery [145]. Moreover, orally administered nano-drugs, constructed from bioactive components of EMPs, enter the systemic circulation via the colon and exert antitumor effects by directly killing tumor cells and modulating the gut microbiota [146]. Based on EMPs compounds, natural, safe, and robust delivery systems are expected to lead the field of oral drug delivery into a new era characterized by sustainability, precision, and personalization, thereby offering innovative solutions for nutritional intervention and disease treatment.
Figure 4.
Drug delivery systems constructed from bioactive components of medicinal-edible plants enable precise drug release at specific sites and modulate immune responses against cancer. Created with biorender.com.
Figure 4.
Drug delivery systems constructed from bioactive components of medicinal-edible plants enable precise drug release at specific sites and modulate immune responses against cancer. Created with biorender.com.
5. Conclusions
In recent years, cancer immunotherapy, particularly ICBs, has become a hotspot in cancer treatment. However, during immunotherapy, due to tumor heterogeneity and inter-patient variability, irAEs might arise, including issues such as drug resistance and poor prognosis. Gut microbiota and their metabolites can modulate tumor immune responses. In turn, cancer can reshape the gut microbial composition, leading to the modulation of TME and the promotion of immune suppression [147]. This review has detailed that EMPs, which are abundant in polysaccharides, polyphenols, and saponins (exemplified by traditional sources like ginseng, astragalus, angelica, and turmeric), exhibit favorable characteristics: high safety, stability, low toxicity, and multi-targeting capability. Notably, EMPs can enhance anti-tumor immune responses and synergize with cancer immunotherapy via bidirectional interactions with the gut microbiota, constituting a highly promising interdisciplinary research field. Beyond their direct use as anti-cancer agents, the bioactive compounds of EMPs can also participate in the assembly of drug delivery systems, which enables targeted transport of therapeutic agents to specific sites, facilitating precise cancer treatment. Additionally, DDS assembled from EMPs compounds offer a promising alternative to synthetic materials, mitigating the long-term toxicity concerns associated with their use, while also boosting the bioavailability and anti-tumor potency of the therapeutic agents. Nevertheless, substantial challenges persist in effectively translating these findings into clinical practice. The inherent variability in the origin, processing, and extraction of EMPs leads to significant inconsistencies in the composition and concentration of their bioactive compounds, resulting in poor reproducibility and unstable research outcomes. This complexity is compounded by the fact that, unlike single-compound drugs, their extracts are multi-component mixtures, making it difficult to determine whether their overall interaction with cancer immunotherapy is synergistic or antagonistic. Taken together, these advancements underscore the substantial potential of leveraging the interplay between bioactive components from EMPs and the gut microbiota to enhance the efficacy of cancer immunotherapy.
Author Contributions
Conceptualization: L.P., and H.Y.; literature search: L.P.; writing—original draft preparation: L.P.; writing—review and editing: H.Y., L.W. and Y.Z.; supervision: H.Y. and P.Y. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
During the preparation of this review, the authors used https://www.biorender.com for the purposes of creating figures. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.S.; Mellman, I. Oncology meets immunology: the cancer-immunity cycle. Immunity. 2013, 39, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: the next generation. Cell. 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- McDermott, A.J.; Huffnagle, G.B. The microbiome and regulation of mucosal immunity. Immunology. 2014, 142, 24–31. [Google Scholar] [CrossRef]
- Wong, S.H.; Yu, J. Gut microbiota in colorectal cancer: mechanisms of action and clinical applications. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 690–704. [Google Scholar] [CrossRef]
- Asokan, S.; Cullin, N.; Stein-Thoeringer, C.K.; Elinav, E. CAR-T Cell Therapy and the Gut Microbiota. Cancers. 2023, 15. [Google Scholar] [CrossRef]
- Frankel, A.E.; Coughlin, L.A.; Kim, J.; Froehlich, T.W.; Xie, Y.; Frenkel, E.P.; Koh, A.Y. Metagenomic Shotgun Sequencing and Unbiased Metabolomic Profiling Identify Specific Human Gut Microbiota and Metabolites Associated with Immune Checkpoint Therapy Efficacy in Melanoma Patients. Neoplasia. 2017, 19, 848–855. [Google Scholar] [CrossRef]
- Vétizou, M.; Pitt, J.M.; Daillère, R.; Lepage, P.; Waldschmitt, N.; Flament, C.; Rusakiewicz, S.; Routy, B.; Roberti, M.P.; Duong, C.P.; et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science. 2015, 350, 1079–1084. [Google Scholar] [CrossRef]
- Routy, B.; Gopalakrishnan, V.; Daillère, R.; Zitvogel, L.; Wargo, J.A.; Kroemer, G. The gut microbiota influences anticancer immunosurveillance and general health. Nat. Rev. Clin. Oncol. 2018, 15, 382–396. [Google Scholar] [CrossRef]
- Jing, N.; Wang, L.; Zhuang, H.; Jiang, G.; Liu, Z. Enhancing therapeutic effects of murine cancer vaccine by reshaping gut microbiota with Lactobacillus rhamnosus GG and jujube powder. Front. Immunol. 2023, 14, 1195075. [Google Scholar] [CrossRef]
- Tan, W.; Pan, T.; Wang, S.; Li, P.; Men, Y.; Tan, R.; Zhong, Z.; Wang, Y. Immunometabolism modulation, a new trick of edible and medicinal plants in cancer treatment. Food Chem. 2022, 376, 131860. [Google Scholar] [CrossRef]
- Xia, L.; Li, C.; Zhao, J.; Sun, Q.; Mao, X. Rebalancing immune homeostasis in combating disease: The impact of medicine food homology plants and gut microbiome. Phytomedicine. 2025, 136, 156150. [Google Scholar] [CrossRef]
- Zhou, Z.; Nan, Y.; Li, X.; Ma, P.; Du, Y.; Chen, G.; Ning, N.; Huang, S.; Gu, Q.; Li, W.; et al. Hawthorn with “homology of medicine and food”: a review of anticancer effects and mechanisms. Front. Pharmacol. 2024, 15, 1384189. [Google Scholar] [CrossRef]
- Lu, Q.; Li, R.; Yang, Y.; Zhang, Y.; Zhao, Q.; Li, J. Ingredients with anti-inflammatory effect from medicine food homology plants. Food Chem. 2022, 368, 130610. [Google Scholar] [CrossRef]
- Wen, S.; Han, Y.; Li, Y.; Zhan, D. Therapeutic Mechanisms of Medicine Food Homology Plants in Alzheimer’s Disease: Insights from Network Pharmacology, Machine Learning, and Molecular Docking. Int. J. Mol. Sci. 2025, 26. [Google Scholar] [CrossRef] [PubMed]
- Li, M.J.; Xu, J.Y.; Zhang, H.Y.; Guo, M.; Lan, M.N.; Kong, J.; Liu, S.W.; Zheng, H.J. A medicine and food homology formula prevents cognitive deficits by inhibiting neuroinflammation and oxidative stress via activating AEA-Trpv1-Nrf2 pathway. Inflammopharmacology. 2024, 32, 3745–3759. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Xu, C.; Zhou, W.; Li, Y.; Xie, Y.; Hu, H.; Wang, Z. Polygonati Rhizoma with the homology of medicine and food: A review of ethnopharmacology, botany, phytochemistry, pharmacology and applications. J. Ethnopharmacol. 2023, 309, 116296. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Wang, H.; Chen, Y. Prunella vulgaris L. - A Review of its Ethnopharmacology, Phytochemistry, Quality Control and Pharmacological Effects. Front. Pharmacol. 2022, 13, 903171. [Google Scholar] [CrossRef]
- Liu, C.; He, P.; Qiao, R.; Yang, X.; Ding, C.; He, F. Mechanistic study of Lonicerae Japonicae Flos (Caprifoliaceae) in non-small cell lung cancer prevention and treatment through integrative pharmacology, multi-machine learning, artificial intelligence, and in vitro experiments. J. Ethnopharmacol. 2025, 348, 119832. [Google Scholar] [CrossRef]
- Xu, Q.; Cheng, W.; Wei, J.; Ou, Y.; Xiao, X.; Jia, Y. Synergist for antitumor therapy: Astragalus polysaccharides acting on immune microenvironment. Discover oncology. 2023, 14, 179. [Google Scholar] [CrossRef]
- Lee, J.S.; Lee, H.Y. Ginseng-derived compounds as potential anticancer agents targeting cancer stem cells. J. Ginseng Res. 2024, 48, 266–275. [Google Scholar] [CrossRef]
- Guo, C.; Guo, D.; Fang, L.; Sang, T.; Wu, J.; Guo, C.; Wang, Y.; Wang, Y.; Chen, C.; Chen, J.; et al. Ganoderma lucidum polysaccharide modulates gut microbiota and immune cell function to inhibit inflammation and tumorigenesis in colon. Carbohydr. Polym. 2021, 27, 118231. [Google Scholar] [CrossRef]
- Cheng, J.; Zhou, Z.W.; Sheng, H.P.; He, L.J.; Fan, X.W.; He, Z.X.; Sun, T.; Zhang, X.; Zhao, R.J.; Gu, L.; et al. An evidence-based update on the pharmacological activities and possible molecular targets of Lycium barbarum polysaccharides. Drug Des. Devel. Ther. 2015, 9, 33–78. [Google Scholar] [CrossRef]
- Tian, X.; Liang, T.; Liu, Y.; Ding, G.; Zhang, F.; Ma, Z. Extraction, Structural Characterization, and Biological Functions of Lycium Barbarum Polysaccharides: A Review. Biomolecules. 2019, 9. [Google Scholar] [CrossRef]
- Guo, H.; Gibson, S.A.; Ting, J.P.Y. Gut microbiota, NLR proteins, and intestinal homeostasis. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef]
- Ahmadi, S.; Mainali, R.; Nagpal, R.; Sheikh-Zeinoddin, M.; Soleimanian-Zad, S.; Wang, S.; Deep, G.; Kumar Mishra, S.; Yadav, H. Dietary Polysaccharides in the Amelioration of Gut Microbiome Dysbiosis and Metabolic Diseases. Obes Control Ther. Open Access. 2017, 4. [Google Scholar] [CrossRef]
- Huang, J.; Liu, D.; Wang, Y.; Liu, L.; Li, J.; Yuan, J.; Jiang, Z.; Jiang, Z.; Hsiao, W.W.; Liu, H.; et al. Ginseng polysaccharides alter the gut microbiota and kynurenine/tryptophan ratio, potentiating the antitumour effect of antiprogrammed cell death 1/programmed cell death ligand 1 (anti-PD-1/PD-L1) immunotherapy. Gut. 2022, 71, 734–745. [Google Scholar] [CrossRef]
- Cao, C.; Wang, Z.; Gong, G.; Huang, W.; Huang, L.; Song, S.; Zhu, B. Effects of Lycium barbarum Polysaccharides on Immunity and Metabolic Syndrome Associated with the Modulation of Gut Microbiota: A Review. Foods. 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Wu, Z.; Chen, H.; Liu, W.; Gu, F.; Li, J. Extraction and Identification of Polysaccharide from Lentinus edodes and Its Effect on Immunosuppression and Intestinal Barrier Injury Induced by Cyclophosphamide. Int. J. Mol. Sci. 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhu, W.; Hong, Y.; Wei, W.; Zheng, N.; He, X.; Bao, Y.; Gao, X.; Huang, W.; Sheng, L.; et al. Astragalus polysaccharides attenuate chemotherapy-induced immune injury by modulating gut microbiota and polyunsaturated fatty acid metabolism. Phytomedicine. 2024, 128, 155492. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.P.; Li, S.; Chen, Y.M.; Li, H.B. Natural Polyphenols for Prevention and Treatment of Cancer. Nutrients. 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Khan, A.; Ahmad, I.; Alghamdi, S.; Rajab, B.S.; Babalghith, A.O.; Alshahrani, M.Y.; Islam, S.; Islam, M.R. Flavonoids a Bioactive Compound from Medicinal Plants and Its Therapeutic Applications. Biomed Res. Int. 2022, 2022, 5445291. [Google Scholar] [CrossRef] [PubMed]
- Guan, M.; Xu, W.; Bai, H.; Geng, Z.; Yu, Z.; Li, H.; Liu, T. Potential mechanisms underlying inhibition of xenograft lung cancer models by kaempferol: modulation of gut microbiota in activating immune cell function. J. Cancer. 2024, 15, 1314–1327. [Google Scholar] [CrossRef] [PubMed]
- Bratkov, V.M.; Shkondrov, A.M.; Zdraveva, P.K.; Krasteva, I.N. Flavonoids from the Genus Astragalus: Phytochemistry and Biological Activity. Pharmacogn. Rev. 2016, 10, 11–32. [Google Scholar] [CrossRef]
- Che, Y.; Li, L.; Kong, M.; Geng, Y.; Wang, D.; Li, B.; Deng, L.; Chen, G.; Wang, J. Dietary supplementation of Astragalus flavonoids regulates intestinal immunology and the gut microbiota to improve growth performance and intestinal health in weaned piglets. Front. Immunol. 2024, 15, 1459342. [Google Scholar] [CrossRef]
- Hirsch, D.; Hardt, J.; Sauer, C.; Heselmeyer-Hadded, K.; Witt, S.H.; Kienle, P.; Ried, T.; Gaiser, T. Molecular characterization of ulcerative colitis-associated colorectal carcinomas. Mod. Pathol. 2021, 34, 1153–1166. [Google Scholar] [CrossRef]
- Imran, M.; Rauf, A.; Abu-Izneid, T.; Nadeem, M.; Shariati, M.A.; Khan, I.A.; Imran, A.; Erdogan Orhan, I.; Rizwan, M.; Atif, M.; et al. Corrigendum to “Luteolin, a flavonoid, as an anticancer agent: A review” [Biomed. Pharmacother. 112 (2019) 108612]. Biomed. Pharmacother. 2019, 116, 109084. [Google Scholar] [CrossRef]
- Liu, F.; Guo, C.; Liu, X.; Gu, Z.; Zou, W.; Tang, X.; Tang, J. Luteolin in Inflammatory Bowel Disease and Colorectal Cancer: A Disease Continuum Perspective. Curr. Issues Mol. Biol. 2025, 47. [Google Scholar] [CrossRef]
- Tang, L.; Gao, J.; Li, X.; Cao, X.; Zhou, B. Molecular Mechanisms of Luteolin Against Atopic Dermatitis Based on Network Pharmacology and in vivo Experimental Validation. Drug Des. Devel. Ther. 2022, 16, 4205–4221. [Google Scholar] [CrossRef]
- Li, Y.; Shen, L.; Luo, H. Luteolin ameliorates dextran sulfate sodium-induced colitis in mice possibly through activation of the Nrf2 signaling pathway. Int. Immunopharmacol. 2016, 40, 24–31. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Li, B.; Du, P.; Du, Y.; Zhao, D.; Cai, Y.; Yang, Q.; Guo, Z. Luteolin alleviates inflammation and modulates gut microbiota in ulcerative colitis rats. Life Sci. 2021, 269, 119008. [Google Scholar] [CrossRef] [PubMed]
- Neurath, M.F.; Vieth, M. Different levels of healing in inflammatory bowel diseases: mucosal, histological, transmural, barrier and complete healing. Gut. 2023, 72, 2164–2183. [Google Scholar] [CrossRef] [PubMed]
- Li, B.L.; Zhao, D.Y.; Du, P.L.; Wang, X.T.; Yang, Q.; Cai, Y.R. Luteolin alleviates ulcerative colitis through SHP-1/STAT3 pathway. Inflamm. Res. 2021, 70, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Chandrashekharappa, S.; Bodduluri, S.R.; Baby, B.V.; Hegde, B.; Kotla, N.G.; Hiwale, A.A.; Saiyed, T.; Patel, P.; Vijay-Kumar, M.; et al. Enhancement of the gut barrier integrity by a microbial metabolite through the Nrf2 pathway. Nat. Commun. 2019, 10, 89. [Google Scholar] [CrossRef]
- Luo, B.; An, Q.; Lei, J.; Tan, D.; Liu, X.; Li, H.; Zhao, Y.; Qin, J.; Zhang, C.; Zhang, Y.; et al. Resveratrol amplifies the anti-tumor effect of α-PD-1 by altering the intestinal microbiome and PGD2 content. Gut Microbes. 2025, 17, 2447821. [Google Scholar] [CrossRef]
- Zhou, H.; Zhuang, Y.; Liang, Y.; Chen, H.; Qiu, W.; Xu, H.; Zhou, H. Curcumin exerts anti-tumor activity in colorectal cancer via gut microbiota-mediated CD8(+) T Cell tumor infiltration and ferroptosis. Food Funct. 2025, 16, 3671–3693. [Google Scholar] [CrossRef]
- Shabbir, U.; Rubab, M.; Daliri, E.B.; Chelliah, R.; Javed, A.; Oh, D.H. Curcumin, Quercetin, Catechins and Metabolic Diseases: The Role of Gut Microbiota. Nutrients. 2021, 13. [Google Scholar] [CrossRef]
- McFadden, R.M.; Larmonier, C.B.; Shehab, K.W.; Midura-Kiela, M.; Ramalingam, R.; Harrison, C.A.; Besselsen, D.G.; Chase, J.H.; Caporaso, J.G.; Jobin, C.; et al. The Role of Curcumin in Modulating Colonic Microbiota During Colitis and Colon Cancer Prevention. Inflamm. Bowel Dis. 2015, 21, 2483–2494. [Google Scholar] [CrossRef]
- Qi, Y.; Gao, F.; Hou, L.; Wan, C. Anti-Inflammatory and Immunostimulatory Activities of Astragalosides. Am. J. Chin. Med. 2017, 45, 1157–1167. [Google Scholar] [CrossRef]
- Luo, H.; Vong, C.T.; Chen, H.; Gao, Y.; Lyu, P.; Qiu, L.; Zhao, M.; Liu, Q.; Cheng, Z.; Zou, J.; et al. Naturally occurring anti-cancer compounds: shining from Chinese herbal medicine. Chin. Med. 2019, 14, 48. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Huang, Y.; Zheng, H.; Li, S.; Li, Z.; Yuan, L.; Cheng, X.; He, C.; Sun, J. Ginsenosides for the treatment of metabolic syndrome and cardiovascular diseases: Pharmacology and mechanisms. Biomed. Pharmacother. 2020, 132, 110915. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Li, M.; Zhang, Z.; Song, Z.; Xu, J.; Zhang, M.; Gong, M. Overview of Panax ginseng and its active ingredients protective mechanism on cardiovascular diseases. J. Ethnopharmacol. 2024, 334, 118506. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Tang, Y.; Keep, R.F.; Ma, X.; Xiang, J. Antioxidative effects of Panax notoginseng saponins in brain cells. Phytomedicine. 2014, 21, 1189–1195. [Google Scholar] [CrossRef]
- Baik, I.H.; Kim, K.H.; Lee, K.A. Antioxidant, Anti-Inflammatory and Antithrombotic Effects of Ginsenoside Compound K Enriched Extract Derived from Ginseng Sprouts. Molecules. 2021, 26. [Google Scholar] [CrossRef]
- Khan, M.I.; Karima, G.; Khan, M.Z.; Shin, J.H.; Kim, J.D. Therapeutic Effects of Saponins for the Prevention and Treatment of Cancer by Ameliorating Inflammation and Angiogenesis and Inducing Antioxidant and Apoptotic Effects in Human Cells. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- Zhou, R.; Guo, T.; Li, J. Research progress on the antitumor effects of astragaloside IV. Eur. J. Pharmacol. 2023, 938, 175449. [Google Scholar] [CrossRef]
- Choi, Y.H.; Yoo, D.S.; Cha, M.R.; Choi, C.W.; Kim, Y.S.; Choi, S.U.; Lee, K.R.; Ryu, S.Y. Antiproliferative effects of saponins from the roots of Platycodon grandiflorum on cultured human tumor cells. J. Nat. Prod. 2010, 73, 1863–1867. [Google Scholar] [CrossRef]
- Xia, T.; Zhang, B.; Li, Y.; Fang, B.; Zhu, X.; Xu, B.; Zhang, J.; Wang, M.; Fang, J. New insight into 20(S)-ginsenoside Rh2 against T-cell acute lymphoblastic leukemia associated with the gut microbiota and the immune system. Eur. J. Med. Chem. 2020, 203, 112582. [Google Scholar] [CrossRef]
- Yousuf, S.; Liu, H.; Yingshu, Z.; Zahid, D.; Ghayas, H.; Li, M.; Ding, Y.; Li, W. Ginsenoside Rg1 modulates intestinal microbiota and supports re-generation of immune cells in dexamethasone-treated mice. Acta Microbiol. Immunol. Hung. 2022, 69, 259–269. [Google Scholar] [CrossRef]
- Bai, X.; Fu, R.; Liu, Y.; Deng, J.; Fei, Q.; Duan, Z.; Zhu, C.; Fan, D. Ginsenoside Rk3 modulates gut microbiota and regulates immune response of group 3 innate lymphoid cells to against colorectal tumorigenesis. J. Pharm. Anal. 2024, 14, 259–275. [Google Scholar] [CrossRef] [PubMed]
- Ran, J.; Ai, Y.; Ni, J.; Zhang, Y.; Chen, J.; Wang, T.; Ma, J.; Zheng, J.; Li, R.; Ma, X.; et al. Astragaloside IV: A Promising Drug to Prevent the Transition from Colitis to Colorectal Cancer. Am. J. Chin. Med. 2025, 53, 1065–1091. [Google Scholar] [CrossRef] [PubMed]
- Hamid, O.; Robert, C.; Daud, A.; Hodi, F.S.; Hwu, W.J.; Kefford, R.; Wolchok, J.D.; Hersey, P.; Joseph, R.W.; Weber, J.S.; et al. Safety and tumor responses with lambrolizumab (anti-PD-1) in melanoma. N. Engl. J. Med. 2013, 369, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Li, X.; Wang, X.; Cheng, L.; Li, Z.; Zhang, C.; Ye, Z.; Qian, Q. Current Progress in CAR-T Cell Therapy for Solid Tumors. Int. J. Biol. Sci. 2019, 15, 2548–2560. [Google Scholar] [CrossRef]
- Chrysostomou, D.; Roberts, L.A.; Marchesi, J.R.; Kinross, J.M. Gut Microbiota Modulation of Efficacy and Toxicity of Cancer Chemotherapy and Immunotherapy. Gastroenterology. 2023, 164, 198–213. [Google Scholar] [CrossRef]
- Li, Z.; Xiong, W.; Liang, Z.; Wang, J.; Zeng, Z.; Kołat, D.; Li, X.; Zhou, D.; Xu, X.; Zhao, L. Critical role of the gut microbiota in immune responses and cancer immunotherapy. J. Hematol. Oncol. 2024, 17, 33. [Google Scholar] [CrossRef]
- Fernandes, M.R.; Aggarwal, P.; Costa, R.G.F.; Cole, A.M.; Trinchieri, G. Targeting the gut microbiota for cancer therapy. Nat. Rev. Cancer. 2022, 22, 703–722. [Google Scholar] [CrossRef]
- Park, E.M.; Chelvanambi, M.; Bhutiani, N.; Kroemer, G.; Zitvogel, L.; Wargo, J.A. Targeting the gut and tumor microbiota in cancer. Nat. Med. 2022, 28, 690–703. [Google Scholar] [CrossRef]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.L.; et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science. 2015, 350, 1084–1089. [Google Scholar] [CrossRef]
- Marcos-Kovandzic, L.; Avagliano, M.; Ben Khelil, M.; Srikanthan, J.; Abdallah, R.; Petrocelli, V.; Rengassamy, J.; Alfaro, A.; Bied, M.; Fidelle, M.; et al. Gut microbiota modulation through Akkermansia spp. supplementation increases CAR-T cell potency. 2025. [Google Scholar] [CrossRef]
- Pardoll, D.M. The blockade of immune checkpoints in cancer immunotherapy. Nature reviews. Cancer. 2012, 12, 252–264. [Google Scholar] [CrossRef]
- Webb, E.S.; Liu, P.; Baleeiro, R.; Lemoine, N.R.; Yuan, M.; Wang, Y.H. Immune checkpoint inhibitors in cancer therapy. J. Biomed. Res. 2018, 32, 317–326. [Google Scholar] [CrossRef]
- Granier, C.; De Guillebon, E.; Blanc, C.; Roussel, H.; Badoual, C.; Colin, E.; Saldmann, A.; Gey, A.; Oudard, S.; Tartour, E. Mechanisms of action and rationale for the use of checkpoint inhibitors in cancer. ESMO open. 2017, 2, e000213. [Google Scholar] [CrossRef] [PubMed]
- Wolchok, J.D.; Chiarion-Sileni, V.; Gonzalez, R.; Rutkowski, P.; Grob, J.J.; Cowey, C.L.; Lao, C.D.; Wagstaff, J.; Schadendorf, D.; Ferrucci, P.F.; et al. Overall Survival with Combined Nivolumab and Ipilimumab in Advanced Melanoma. N. Engl. J. Med. 2017, 377, 1345–1356. [Google Scholar] [CrossRef] [PubMed]
- Ahmadzadeh, M.; Johnson, L.A.; Heemskerk, B.; Wunderlich, J.R.; Dudley, M.E.; White, D.E.; Rosenberg, S.A. Tumor antigen-specific CD8 T cells infiltrating the tumor express high levels of PD-1 and are functionally impaired. Blood. 2009, 114, 1537–1544. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yu, D.; Wu, D.; Gao, X.; Shao, F.; Zhao, M.; Wang, J.; Ma, J.; Wang, W.; Qin, X.; et al. Tissue-resident Lachnospiraceae family bacteria protect against colorectal carcinogenesis by promoting tumor immune surveillance. Cell Host Microbe. 2023, 31, 418–432.e418. [Google Scholar] [CrossRef]
- Zong, G.; Deng, R.; Pan, Y.; Liu, M.; Zhu, H.; Tao, R.; Shan, Y.; Wei, Z.; Lu, Y. Ginseng polysaccharides ameliorate colorectal tumorigenesis through Lachnospiraceae-mediated immune modulation. Int. J. Biol. Macromol. 2025, 307, 142015. [Google Scholar] [CrossRef]
- Zhang, G.; Pan, J.; Xu, X.; Nie, S.; Lu, L.; Jing, Y.; Yang, F.; Ji, G.; Xu, H. Chinese yam polysaccharide enhances anti-PD-1 immunotherapy in colorectal cancer through alterations in the gut microbiota and metabolites. Int. J. Biol. Macromol. 2025, 310, 143323. [Google Scholar] [CrossRef]
- Wang, L.; Jiang, G.; Jing, N.; Liu, X.; Li, Q.; Liang, W.; Liu, Z. Bilberry anthocyanin extracts enhance anti-PD-L1 efficiency by modulating gut microbiota. Food Funct. 2020, 11, 3180–3190. [Google Scholar] [CrossRef]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nature reviews. Immunology. 2016, 16, 341–352. [Google Scholar] [CrossRef]
- Krautkramer, K.A.; Fan, J.; Bäckhed, F. Gut microbial metabolites as multi-kingdom intermediates. Nat. Rev. Microbiol. 2021, 19, 77–94. [Google Scholar] [CrossRef]
- Davar, D.; Dzutsev, A.K.; McCulloch, J.A.; Rodrigues, R.R.; Chauvin, J.M.; Morrison, R.M.; Deblasio, R.N.; Menna, C.; Ding, Q.; Pagliano, O.; et al. Fecal microbiota transplant overcomes resistance to anti-PD-1 therapy in melanoma patients. Science. 2021, 371, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wang, B.; Zheng, Q.; Li, H.; Meng, X.; Zhou, F.; Zhang, L. A Review of Gut Microbiota-Derived Metabolites in Tumor Progression and Cancer Therapy. Adv. Sci. 2023, 10, e2207366. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jia, L.; Ma, Q.; Zhang, X.; Chen, M.; Liu, F.; Zhang, T.; Jia, W.; Zhu, L.; Qi, W.; et al. Astragalus Polysaccharide Modulates the Gut Microbiota and Metabolites of Patients with Type 2 Diabetes in an In Vitro Fermentation Model. Nutrients. 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Wu, J.; Zheng, Y.; Li, M.; Xu, X.; Chen, H.; Rui, W. Structural changes of polysaccharides from Astragulus after honey processing and their bioactivities on human gut microbiota. J. Sci. Food Agric. 2023, 103, 7241–7250. [Google Scholar] [CrossRef]
- Yue, B.; Zong, G.; Tao, R.; Wei, Z.; Lu, Y. Crosstalk between traditional Chinese medicine-derived polysaccharides and the gut microbiota: A new perspective to understand traditional Chinese medicine. Phytother. Res. 2022, 36, 4125–4138. [Google Scholar] [CrossRef]
- Chang, P.V.; Hao, L.; Offermanns, S.; Medzhitov, R. The microbial metabolite butyrate regulates intestinal macrophage function via histone deacetylase inhibition. Proceedings of the National Academy of Sciences of the United U.S.A. 2014, 111, 2247–2252. [Google Scholar] [CrossRef]
- Thangaraju, M.; Cresci, G.A.; Liu, K.; Ananth, S.; Gnanaprakasam, J.P.; Browning, D.D.; Mellinger, J.D.; Smith, S.B.; Digby, G.J.; Lambert, N.A.; et al. GPR109A is a G-protein-coupled receptor for the bacterial fermentation product butyrate and functions as a tumor suppressor in colon. Cancer Res. 2009, 69, 2826–2832. [Google Scholar] [CrossRef]
- Singh, N.; Thangaraju, M.; Prasad, P.D.; Martin, P.M.; Lambert, N.A.; Boettger, T.; Offermanns, S.; Ganapathy, V. Blockade of dendritic cell development by bacterial fermentation products butyrate and propionate through a transporter (Slc5a8)-dependent inhibition of histone deacetylases. J. Biol. Chem. 2010, 285, 27601–27608. [Google Scholar] [CrossRef]
- Sun, M.; Wu, W.; Chen, L.; Yang, W.; Huang, X.; Ma, C.; Chen, F.; Xiao, Y.; Zhao, Y.; Ma, C.; et al. Microbiota-derived short-chain fatty acids promote Th1 cell IL-10 production to maintain intestinal homeostasis. Nat. Commun. 2018, 9, 3555. [Google Scholar] [CrossRef]
- Wang, L.; Chen, J.; Chen, Q.; Song, H.; Wang, Z.; Xing, W.; Jin, S.; Song, X.; Yang, H.; Zhao, W. The Gut Microbiota Metabolite Urolithin B Prevents Colorectal Carcinogenesis by Remodeling Microbiota and PD-L1/HLA-B. Oxid. Med. Cell. Longev. 2023, 2023, 6480848. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; González-Sarrías, A.; García-Villalba, R.; Núñez-Sánchez, M.A.; Selma, M.V.; García-Conesa, M.T.; Espín, J.C. Urolithins, the rescue of “old” metabolites to understand a “new” concept: Metabotypes as a nexus among phenolic metabolism, microbiota dysbiosis, and host health status. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Lo, B.C.; Kryczek, I.; Yu, J.; Vatan, L.; Caruso, R.; Matsumoto, M.; Sato, Y.; Shaw, M.H.; Inohara, N.; Xie, Y.; et al. Microbiota-dependent activation of CD4(+) T cells induces CTLA-4 blockade-associated colitis via Fcγ receptors. Science. 2024, 383, 62–70. [Google Scholar] [CrossRef]
- Zhou, Y.; Medik, Y.B.; Patel, B.; Zamler, D.B.; Chen, S.; Chapman, T.; Schneider, S.; Park, E.M.; Babcock, R.L.; Chrisikos, T.T.; et al. Intestinal toxicity to CTLA-4 blockade driven by IL-6 and myeloid infiltration. J. Exp. Med. 2023, 220. [Google Scholar] [CrossRef]
- Simpson, R.C.; Shanahan, E.R.; Batten, M.; Reijers, I.L.M.; Read, M.; Silva, I.P.; Versluis, J.M.; Ribeiro, R.; Angelatos, A.S.; Tan, J.; et al. Diet-driven microbial ecology underpins associations between cancer immunotherapy outcomes and the gut microbiome. Nat. Med. 2022, 28, 2344–2352. [Google Scholar] [CrossRef] [PubMed]
- Michot, J.M.; Bigenwald, C.; Champiat, S.; Collins, M.; Carbonnel, F.; Postel-Vinay, S.; Berdelou, A.; Varga, A.; Bahleda, R.; Hollebecque, A.; et al. Immune-related adverse events with immune checkpoint blockade: a comprehensive review. Eur. J. Cance. 2016, 54, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Takiishi, T.; Fenero, C.I.M.; Câmara, N.O.S. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue barriers. 2017, 5, e1373208. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Littman, D.R. The microbiota in adaptive immune homeostasis and disease. Nature. 2016, 535, 75–84. [Google Scholar] [CrossRef]
- Qi, P.; Yin, L.; Wei, R.; Yang, S.; Liu, Z.; Huang, P.; Yu, Q.; Xiong, S.; Wang, M.; Deng, Y.; et al. Chronic stress synergizes with Listeria monocytogenes to promote intestinal adenomagenesis via myeloid-derived suppressor cells. Front. Immunol. 2025, 16. [Google Scholar] [CrossRef]
- Jin-quan, Z.; Jian-song, W.; Hui, Z. Immunosuppression Mechanism of MDSCs and Regulatory T Cells in Inflammation and Tumor Microenvironment. J. Int. Oncol. 2013. [Google Scholar]
- Gill, G.S.; Kharb, S.; Goyal, G.; Das, P.; Kurdia, K.C.; Dhar, R.; Karmakar, S. Immune checkpoint inhibitors and immunosuppressive tumor microenvironment: current challenges and strategies to overcome resistance. Immunopharmacol. Immunotoxicol. 2025, 47, 485–507. [Google Scholar] [CrossRef]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Huang, J.; Sun, M.; Jiang, Y.; Wang, S.; Wang, L.; Yu, N.; Peng, D.; Wang, Y.; Chen, W.; et al. Poria cocos polysaccharide improves intestinal barrier function and maintains intestinal homeostasis in mice. Int. J. Biol. Macromol. 2023, 249, 125953. [Google Scholar] [CrossRef] [PubMed]
- Huo, J.; Wu, Z.; Sun, W.; Wang, Z.; Wu, J.; Huang, M.; Wang, B.; Sun, B. Protective Effects of Natural Polysaccharides on Intestinal Barrier Injury: A Review. J. Agric. Food Chem. 2022, 70, 711–735. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.S.; He, H.; Wang, J.; Gehr, T.W.; Ghosh, S. Curcumin-mediated regulation of intestinal barrier function: The mechanism underlying its beneficial effects. Tissue Barriers. 2018, 6, e1425085. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Du, P.; Yang, W.; Huang, D.; Nie, S.; Xie, M. Polysaccharide from the seeds of Plantago asiatica L. alleviates nonylphenol induced intestinal barrier injury by regulating tight junctions in human Caco-2 cell line. Int. J. Biol. Macromol. 2020, 164, 2134–2140. [Google Scholar] [CrossRef]
- Li, W.; Gao, M.; Han, T. Lycium barbarum polysaccharides ameliorate intestinal barrier dysfunction and inflammation through the MLCK-MLC signaling pathway in Caco-2 cells. Food Funct. 2020, 11, 3741–3748. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, F.; Chen, X.; Yang, Z.; Cao, Y. Effects of the polysaccharide SPS-3-1 purified from Spirulina on barrier integrity and proliferation of Caco-2 cells. Int. J. Biol. Macromol. 2020, 163, 279–287. [Google Scholar] [CrossRef]
- Dougan, M.; Pietropaolo, M. Time to dissect the autoimmune etiology of cancer antibody immunotherapy. J. Clin. Invest. 2020, 130, 51–61. [Google Scholar] [CrossRef]
- Dougan, M. Checkpoint Blockade Toxicity and Immune Homeostasis in the Gastrointestinal Tract. Front. Immunol. 2017, 8, 1547. [Google Scholar] [CrossRef]
- Larkin, J.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.J.; Cowey, C.L.; Lao, C.D.; Schadendorf, D.; Dummer, R.; Smylie, M.; Rutkowski, P.; et al. Combined Nivolumab and Ipilimumab or Monotherapy in Untreated Melanoma. N. Engl. J. Med. 2015, 373, 23–34. [Google Scholar] [CrossRef]
- Postow, M.A.; Chesney, J.; Pavlick, A.C.; Robert, C.; Grossmann, K.; McDermott, D.; Linette, G.P.; Meyer, N.; Giguere, J.K.; Agarwala, S.S.; et al. Nivolumab and ipilimumab versus ipilimumab in untreated melanoma. N. Engl. J. Med. 2015, 372, 2006–2017. [Google Scholar] [CrossRef] [PubMed]
- Postow, M.A.; Sidlow, R.; Hellmann, M.D. Immune-Related Adverse Events Associated with Immune Checkpoint Blockade. N. Engl. J. Med. 2018, 378, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, F.; Li, Y.; Fan, N.; Zhao, K.; Zhang, A.; Kang, J.; Lin, Y.; Xue, X.; Jiang, X. Blockade of PI3K/AKT signaling pathway by Astragaloside IV attenuates ulcerative colitis via improving the intestinal epithelial barrier. J. Transl. Med. 2024, 22, 406. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Ao, M.; Dong, B.; Jiang, Y.; Yu, L.; Chen, Z.; Hu, C.; Xu, R. Anti-Inflammatory Effects of Curcumin in the Inflammatory Diseases: Status, Limitations and Countermeasures. Drug Des. Devel. Ther. 2021, 15, 4503–4525. [Google Scholar] [CrossRef]
- Zhao, J.; Jia, W.; Zhang, R.; Wang, X.; Zhang, L. Improving curcumin bioavailability: Targeted delivery of curcumin and loading systems in intestinal inflammation. Food Res. Int. 2024, 196, 115079. [Google Scholar] [CrossRef]
- Long, D.; Mao, C.; Zhang, W.; Zhu, Y.; Xu, Y. Natural products for the treatment of ulcerative colitis: focus on the JAK/STAT pathway. Front. Immunol. 2025, 16, 1538302. [Google Scholar] [CrossRef]
- Jia, X.; Huang, Y.; Liu, G.; Li, Z.; Tan, Q.; Zhong, S. The Use of Polysaccharide AOP30 from the Rhizome of Alpinia officinarum Hance to Alleviate Lipopolysaccharide-Induced Intestinal Epithelial Barrier Dysfunction and Inflammation via the TLR4/NfκB Signaling Pathway in Caco-2 Cell Monolayers. Nutrients. 2024, 16. [Google Scholar] [CrossRef]
- Li, Z.; Chu, T.; Sun, X.; Zhuang, S.; Hou, D.; Zhang, Z.; Sun, J.; Liu, Y.; Li, J.; Bian, Y. Polyphenols-rich Portulaca oleracea L. (purslane) alleviates ulcerative colitis through restiring the intestinal barrier, gut microbiota and metabolites. Food Chem. 2025, 468, 142391. [Google Scholar] [CrossRef]
- Hanai, H.; Iida, T.; Takeuchi, K.; Watanabe, F.; Maruyama, Y.; Andoh, A.; Tsujikawa, T.; Fujiyama, Y.; Mitsuyama, K.; Sata, M.; et al. Curcumin maintenance therapy for ulcerative colitis: randomized, multicenter, double-blind, placebo-controlled trial. Clin. Gastroenterol. Hepatol. 2006, 4, 1502–1506. [Google Scholar] [CrossRef]
- Shang, Q.; Liu, W.; Leslie, F.; Yang, J.; Guo, M.; Sun, M.; Zhang, G.; Zhang, Q.; Wang, F. Nano-formulated delivery of active ingredients from traditional Chinese herbal medicines for cancer immunotherapy. Acta Pharm. Sin. B. 2024, 14, 1525–1541. [Google Scholar] [CrossRef]
- Sun, D.; Zou, Y.; Song, L.; Han, S.; Yang, H.; Chu, D.; Dai, Y.; Ma, J.; O’Driscoll, C.M.; Yu, Z.; et al. A cyclodextrin-based nanoformulation achieves co-delivery of ginsenoside Rg3 and quercetin for chemo-immunotherapy in colorectal cancer. Acta Pharm. Sin. B. 2022, 12, 378–393. [Google Scholar] [CrossRef]
- Man, S.; Liu, W.; Bi, J.; Bai, J.; Wu, Q.; Hu, B.; Hu, J.; Ma, L. Smart Mesoporous Silica Nanoparticles Loading Curcumin Inhibit Liver Cancer. J. Agric. Food Chem. 2024, 72, 25743–25754. [Google Scholar] [CrossRef] [PubMed]
- Peng, P.; Chen, Z.; Wang, M.; Wen, B.; Deng, X. Polysaccharide-modified liposomes and their application in cancer research. Chem. Biol. Drug Des. 2023, 101, 998–1011. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zheng, Y.; Sun, Q.; Zhang, Z.; Zhao, M.; Peng, C.; Shi, S. Ginsenosides emerging as both bifunctional drugs and nanocarriers for enhanced antitumor therapies. J. Nanobiotechnology. 2021, 19, 322. [Google Scholar] [CrossRef] [PubMed]
- Dai, Q.; Wilhelm, S.; Ding, D.; Syed, A.M.; Sindhwani, S.; Zhang, Y.; Chen, Y.Y.; MacMillan, P.; Chan, W.C.W. Quantifying the Ligand-Coated Nanoparticle Delivery to Cancer Cells in Solid Tumors. ACS Nano. 2018, 12, 8423–8435. [Google Scholar] [CrossRef]
- Liu, X.; Wu, Z.; Guo, C.; Guo, H.; Su, Y.; Chen, Q.; Sun, C.; Liu, Q.; Chen, D.; Mu, H. Hypoxia responsive nano-drug delivery system based on angelica polysaccharide for liver cancer therapy. Drug Deliv. 2022, 29, 138–148. [Google Scholar] [CrossRef]
- Lynam, S.; Lugade, A.A.; Odunsi, K. Immunotherapy for Gynecologic Cancer: Current Applications and Future Directions. Clin. Obstet. Gynecol. 2020, 63, 48–63. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, Z.X.; Wang, J.H.; Guo, R.B.; Zhang, L.; Kong, L.; Yu, Y.; Zang, J.; Liu, Y.; Li, X.T. Immunomodulatory and anti-ovarian cancer effects of novel astragalus polysaccharide micelles loaded with podophyllotoxin. Int. J. Biol. Macromol. 2025, 290, 138960. [Google Scholar] [CrossRef]
- Nel, A.; Xia, T.; Mädler, L.; Li, N. Toxic potential of materials at the nanolevel. Science. 2006, 311, 622–627. [Google Scholar] [CrossRef]
- Sercombe, L.; Veerati, T.; Moheimani, F.; Wu, S.Y.; Sood, A.K.; Hua, S. Advances and Challenges of Liposome Assisted Drug Delivery. Front. Pharmacol. 2015, 6, 286. [Google Scholar] [CrossRef]
- Oude Blenke, E.; Mastrobattista, E.; Schiffelers, R.M. Strategies for triggered drug release from tumor targeted liposomes. Expert Opin. Drug Deliv. 2013, 10, 1399–1410. [Google Scholar] [CrossRef]
- Moein Moghimi, S.; Hamad, I.; Bünger, R.; Andresen, T.L.; Jørgensen, K.; Hunter, A.C.; Baranji, L.; Rosivall, L.; Szebeni, J. Activation of the human complement system by cholesterol-rich and PEGylated liposomes-modulation of cholesterol-rich liposome-mediated complement activation by elevated serum LDL and HDL levels. J. Liposome Res. 2006, 16, 167–174. [Google Scholar] [CrossRef]
- Szebeni, J.; Baranyi, L.; Savay, S.; Bodo, M.; Morse, D.S.; Basta, M.; Stahl, G.L.; Bünger, R.; Alving, C.R. Liposome-induced pulmonary hypertension: properties and mechanism of a complement-mediated pseudoallergic reaction. Am. J. Physiol. Heart Circ. Physiol. 2000, 279, H1319–1328. [Google Scholar] [CrossRef]
- Su, Y.; Wang, L.; Liang, K.; Liu, M.; Liu, X.; Song, Y.; Deng, Y. The accelerated blood clearance phenomenon of PEGylated nanoemulsion upon cross administration with nanoemulsions modified with polyglycerin. Asian J. Pharm. Sci. 2018, 13, 44–53. [Google Scholar] [CrossRef]
- Verhoef, J.J.; Anchordoquy, T.J. Questioning the Use of PEGylation for Drug Delivery. Drug Deliv. Transl. Res. 2013, 3, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Atanase, L.I. Micellar Drug Delivery Systems Based on Natural Biopolymers. Polymers. 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, A.; Baskaran, R.; Maeng, H.J.; Yoo, B.K. Ginsenoside improves physicochemical properties and bioavailability of curcumin-loaded nanostructured lipid carrier. Arch. Pharm. Res. 2017, 40, 864–874. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.; Liang, J.; Xia, J.; Zhu, Y.; Guo, Y.; Wang, A.; Lu, C.; Ren, H.; Chen, C.; Li, S.; et al. One Stone Four Birds: A Novel Liposomal Delivery System Multi-functionalized with Ginsenoside Rh2 for Tumor Targeting Therapy. Nano-Micro Lett. 2020, 12, 129. [Google Scholar] [CrossRef]
- Fox, C.B.; Kim, J.; Le, L.V.; Nemeth, C.L.; Chirra, H.D.; Desai, T.A. Micro/nanofabricated platforms for oral drug delivery. J. Control. Release. 2015, 219, 431–444. [Google Scholar] [CrossRef]
- Laffleur, F.; Mayer, A.H. Oral nanoparticulate drug delivery systems for the treatment of intestinal bowel disease and colorectal cancer. Expert Opin. Drug Deliv. 2023, 20, 1595–1607. [Google Scholar] [CrossRef]
- Krishnaiah, Y.S.; Khan, M.A. Strategies of targeting oral drug delivery systems to the colon and their potential use for the treatment of colorectal cancer. Pharm. Dev. Technol. 2012, 17, 521–540. [Google Scholar] [CrossRef] [PubMed]
- Chourasia, M.K.; Jain, S.K. Polysaccharides for colon targeted drug delivery. Drug Deliv. 2004, 11, 129–148. [Google Scholar] [CrossRef]
- Gong, T.; Liu, X.; Wang, X.; Lu, Y.; Wang, X. Applications of polysaccharides in enzyme-triggered oral colon-specific drug delivery systems: A review. Int. J. Biol. Macromol. 2024, 275, 133623. [Google Scholar] [CrossRef]
- Meng, Y.; Qiu, C.; Li, X.; McClements, D.J.; Sang, S.; Jiao, A.; Jin, Z. Polysaccharide-based nano-delivery systems for encapsulation, delivery, and pH-responsive release of bioactive ingredients. Crit. Rev. Food Sci. Nutr. 2024, 64, 187–201. [Google Scholar] [CrossRef]
- Gao, Q.; Chen, N.; Li, B.; Zu, M.; Ma, Y.; Xu, H.; Zhu, Z.; Reis, R.L.; Kundu, S.C.; Xiao, B. Natural lipid nanoparticles extracted from Morus nigra L. leaves for targeted treatment of hepatocellular carcinoma via the oral route. J. Nanobiotechnology. 2024, 22, 4. [Google Scholar] [CrossRef]
- Jin, C.; Lagoudas, G.K.; Zhao, C.; Bullman, S.; Bhutkar, A.; Hu, B.; Ameh, S.; Sandel, D.; Liang, X.S.; Mazzilli, S.; et al. Commensal Microbiota Promote Lung Cancer Development via γδ T Cells. Cell. 2019, 176, 998–1013.e1016. [Google Scholar] [CrossRef]
Figure 1.
The Interaction between bioactive compounds from edible and medicinal plants and gut microbiota in cancer immunomodulation. Created with biorender.com.
Figure 1.
The Interaction between bioactive compounds from edible and medicinal plants and gut microbiota in cancer immunomodulation. Created with biorender.com.
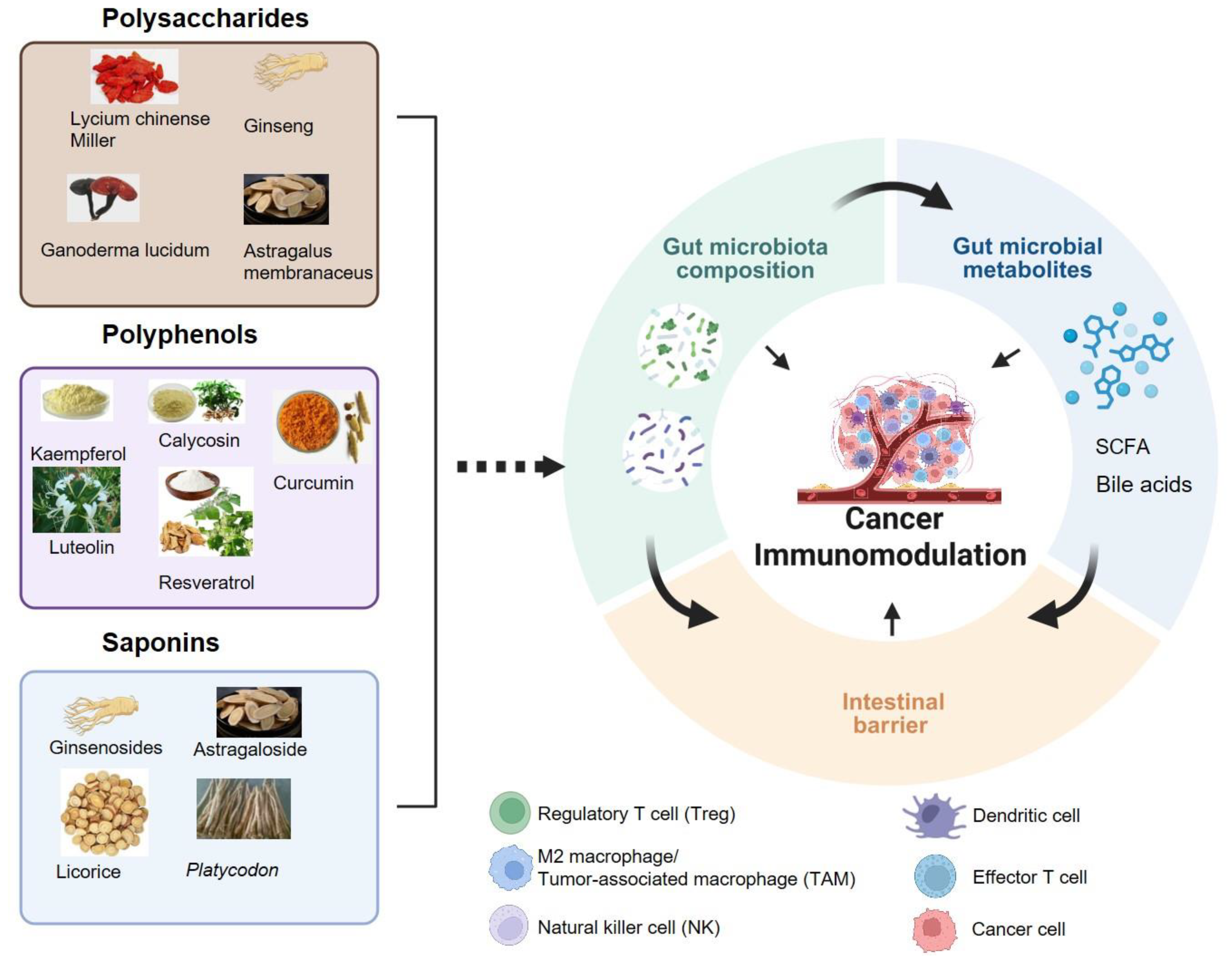
Figure 2.
The interaction bioactive compounds from edible and medicinal plants and gut microbiota in cancer immune response. Created with biorender.com.
Figure 2.
The interaction bioactive compounds from edible and medicinal plants and gut microbiota in cancer immune response. Created with biorender.com.
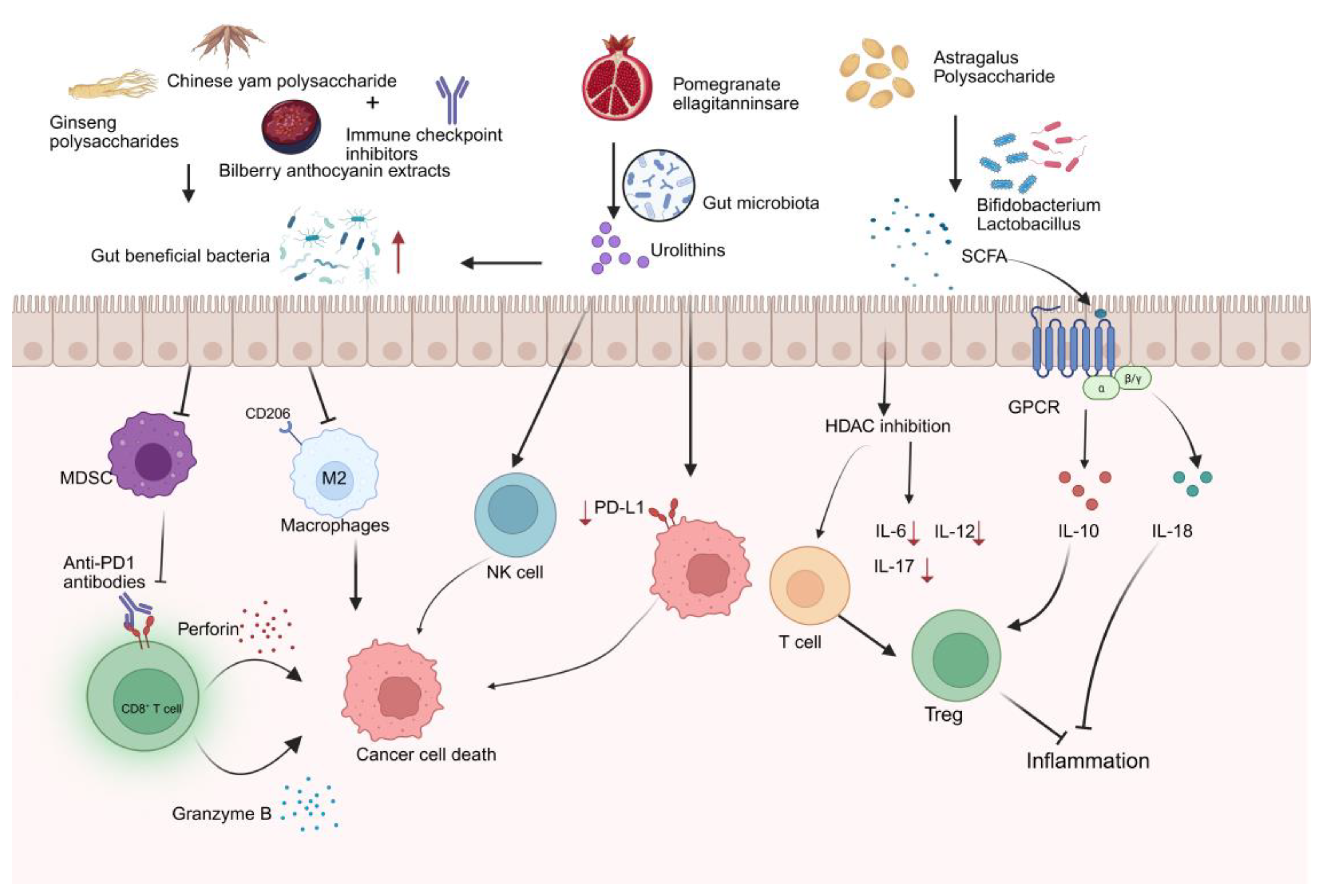
Figure 3.
Natural bioactive compounds from edible and medicinal plants improve intestinal barrier dysfunction and alleviate intestinal inflammation in cancer immunotherapy. Created with biorender.com.
Figure 3.
Natural bioactive compounds from edible and medicinal plants improve intestinal barrier dysfunction and alleviate intestinal inflammation in cancer immunotherapy. Created with biorender.com.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



