
Submitted:
14 October 2025
Posted:
15 October 2025
You are already at the latest version
Abstract
The serpin (serine protease inhibitor) superfamily is the largest class of protease inhibitors, involved in proteolytic cascades and mostly serving as serine/cysteine proteinase inhibitors. Serpins are involved in various biological functions: coagulation, fibrinolysis, angiogenesis, and have crucial roles in various diseases, including cardiac fibrosis, Alzheimer’s, emphysema, obesity, and diabetes. Based on the subcellular distribution profile, vertebrate serpins are classified as intracellular and extracellular serpins. Clade B serpins are considered ancestral serpins, mainly found inside the cells. Clade C serpins were the first to appear extracellularly and most likely bridge intracellular and extracellular serpins. Surprisingly, few reports indicated that secretory serpins such as Plasminogen activator inhibitor-1 (PAI-1) are localized in various subcellular compartments. The intracellular localization of such serpins prompted us to investigate whether the other members of the human serpin superfamily were also inside the cells. Here, we showed for the first time that various secretory serpins from clades A, D, E, H, and I are not obligatory extracellular and were also found in different subcellular compartments, such as the nucleus and centrosome. Surprisingly, secretory serpins tend to shuttle between the nucleus and the cytoplasm as they possess either a nuclear localization signal, a nuclear export signal, or both. Intriguingly, the intracellular localization of secretory serpins is not in line with the evolution-based origin of secretory serpins, suggesting that the secretory serpins acquired additional extracellular functions during evolution, in addition to the intracellular functions. These findings will help decipher the novel intracellular functions of secretory serpins.
Keywords:
Serpin
; Protease
; PAI-1
; nuclear localization signal
; nuclear export signal
; centrosome
Introduction
The serpin (serine protease inhibitor) superfamily is the largest class of protease inhibitors (PIs), comprised of ~1500 members, and is found in all living organisms, i.e., from viruses and prokaryotes (bacteria and archaea) to humans [1,2]. Most of the serpins are involved in proteolytic cascades and serve as serine/cysteine proteinase inhibitors, but a few have novel functions too, such as hormone transport and blood pressure regulation [2]. Serpins are involved in various biological functions: coagulation, fibrinolysis, angiogenesis, and have crucial roles in various diseases, including cardiac fibrosis, Alzheimer’s, emphysema, obesity, and diabetes [3]. They have a conserved secondary structure consisting of three β sheets and nine α-helices and utilize an irreversible suicide-substrate mechanism of inhibition. Serpins contain an exposed reactive center loop (RCL) that imitates the protease substrate [4]. It interacts with target proteases, and the serpin and the protease are irreversibly inactivated [4]. Serpins are classified into 17 clades, of which 9 belong to vertebrate serpins (clades A-I) [5]. In another type of classification considering various biological criteria, i.e., genomic organization, amino acid sites, and indels, vertebrate serpins are further classified into six groups (V1-V6) [6]. Based on the subcellular distribution profile, vertebrate serpins are classified as intracellular and extracellular serpins. Clade B serpins are considered ancestral serpins, mainly found inside the cells [7]. The remaining clades (clade A, clade C-I) of vertebrate serpins are extracellular serpins, of which the largest clade is clade A [8]. Clade C serpins topologically belong to a monophyletic clade that separates the last common ancestors of clade B and other clades (A, D-I). Clade C serpins were the first to appear extracellularly and most likely bridge intracellular and extracellular serpins.
Interestingly, some reports indicated that secretory serpins such as PAI-1 [9], SERPINA2 [10], and SERPINH1 [11] are localized in various subcellular compartments. Recently, a study showed that secretory PAI-1 has been found in the Promyelocytic Leukemia Nuclear Bodies (PML bodies) of endothelial cells and might regulate the endothelial cell growth [9]. The unconventional localization of such serpins tantalized us to explore whether the other members of the human serpin superfamily were also inside the cells. If yes, are they localized in the nucleus or other cell organelles?
To reside in the nucleus, a protein requires a nuclear localization signal (NLS), a basic residue-rich motif responsible for the nuclear import of protein [12]. NLSs are basic amino acid-rich (R and K) short peptides that mediate the nuclear import of proteins by binding to their receptors, known as importins (karyopherins). The cargo protein forms a complex with importin α, importin β1 [13,14]. Importin α serves as an adaptor that links cargos and importin β1 and recognizes NLSs within the cargos [13,14]. The classical best-characterized NLSs (cNLS) are either monopartite, a single cluster of basic residues, or bipartite, two clusters separated by a stretch of 4-10 residues. Additionally, nonclassical NLSs were also reported earlier [15]. One of the subclasses of nonclassical NLSs is comprised of proline-tyrosine-rich NLS (PY-NLS), which follows the consensus R/K/H(X)2-5PY, and they bind with transportin-1 and 2 (importin Kapβ members), mediating the import. These motifs are present in a disordered region of protein possessing an N-terminal stretch of either hydrophobic or basic amino acids and a typical C-terminal R/K/H(X)2-5PY consensus pattern [15].
The nuclear export of proteins is mainly governed by the classical Chromosome region maintenance 1-dependent (CRM1)-dependent nuclear export signals (NES) motifs [16]. Classical CRM1-dependent NES motifs are 10–15 hydrophobic amino acid-long residues (mainly leucine-rich), followed by a specific consensus pattern (Φ0xxΦ1x(2,3)Φ2 x(2,3)Φ3xΦ4, where x is any amino acid, and Φ are hydrophobic amino acids), and are categorized into various classes [17,18]. Various studies showed that in addition to the consensus patterns, the NES motifs should be located in the disordered region of the protein for better accessibility to bind with CRM-1, and possess a helical or helix-to-extended confirmation [17].
In this study, we showed for the first time that various secretory serpins from clades A, D, E, H, and I are not obligatory extracellular and were also found in different subcellular compartments, such as the nucleus and centrosome. Notably, most of the members of secretory serpins possess either NLS or NES or both, suggesting that they might have crucial nuclear functions. These findings will help in identifying the functions of uncharacterized serpins and might open avenues for future functional studies of secretory serpins.
Materials and Methods
Collection of Primary Sequences and 3D Structures of Human Serpin Superfamily Members
Primary reference sequences of human serpins belonging to different clades (Serpin clade A-I) were retrieved from Gene hub, NCBI (https://www.ncbi.nlm.nih.gov/), and the Ensemble database (https://www.ensembl.org/index.html) (Table S1). Protein structures were obtained from PDB (https://www.rcsb.org/) and the UniProt databases (https://www.uniprot.org/)[19] (Table S1). The crystal structures of a few serpins were not available. In such cases, the structures were predicted from AlphaFold and used in the study.
Multiple Sequence Alignment
Subcellular Distribution Data Retrieval of Human Serpins
Two major subcellular resource databases that are based on high-throughput studies of global subcellular distribution profiles of proteins, i.e., the Human Protein Atlas database (HPA) (https://www.proteinatlas.org/) (Table S3) [23] and Subcell Barcode (www.subcellbarcode.org)[24], were used to analyze the intracellular localization of human serpins.
NLS and NES Analysis
cNLS mapper, ELM database (http://elm.eu.org/), and manual curation methods were used to identify NLSs in human serpins. In the cNLS mapper (https://nls-mapper.iab.keio.ac.jp/cgi-bin/), a cut-off score of 0.6 was used to identify NLSs [13]. We manually searched for basic amino acids (R and K) clusters to identify classical NLS and non-classical NLS (PY-NLS). NESs were analyzed using the ELM server (http://elm.eu.org/) and the LocNES tool (http://prodata.swmed.edu/LocNES). NES having a cut-off score > 0.1 were selected from the LocNES predicted motifs for further analysis. The serpin sequences were also manually analyzed by searching the consensus patterns and structural features of different NES classes described in Method S1-2 [25].
Assessment of the Solvent Accessibility of the NES Motif
The solvent-accessible area of identified NESs was assessed using BIOVIA Discovery Studio 2024 Client Visualizer (https://www.3ds.com/products/biovia/discovery-studio) [26]. Serpins were analyzed using the software to calculate the residue solvent accessibility (RSA), sidechain solvent accessibility (SSA), i.e., restricted to side chains, and percent solvent accessibility (PSA). PSA was calculated with a probe radius of 1.4 Å and 240 grid points. The identified motifs showing PSA >25 % were considered solvent-accessible and surface-exposed, while those with PSA <10% were considered buried.
Cell Culture and Immunofluorescence Staining
The human umbilical vein endothelial cells (HUVECs) were isolated from the umbilical cords as described previously [7] as per the guidelines and approval of the Institutional Ethical Committee at the J.L.N. Medical College, Ajmer, India, and the University of Greifswald, Germany. HeLa cells were cultured in DMEM media supplemented with 10% fetal bovine serum and cultured at 5% CO2 and 37°C. Immunofluorescence studies were done as described earlier [9]. Briefly, cells were fixed for 10 min with 3.7% PFA, permeabilized for 20 min using 0.2% Triton-X100 in PBS/T, and stained with Hoechst 33258 at a 10 µg/ml concentration. Non-specific binding was blocked with 2% fatty acid-free BSA in PBS/T for 40 min, followed by an incubation of the primary antibodies in 2% fatty acid-free BSA in PBS/T for 60 min. Secondary antibody incubation was performed using 2% fatty acid-free BSA in PBS/T for 45 min.
The primary antibodies were used at the dilutions: PAI-1 (C-09) antibody (1:75), PML antibody (1:150). The secondary antibodies were used at a dilution of 1:200. Images were captured using a Zeiss fluorescence microscope with Zeiss Axio Vision 4.8 software for data analysis.
Mitochondrial Isolation and Western Blotting
Hela and HUVEC cells’ mitochondria were isolated using the Qproteome Mitochondrial Isolation Kit from Qiagen as per the manufacturer’s instructions. In brief, cell pellet of 1 × 107 cells were resuspended in ice-cold lysis buffer and incubated for 10 min at 4 °C. The lysate was centrifuged, and then the cell pellet was resuspended in 1.5ml ice-cold disruption buffer. Cell disruption was completed by passing the cell pellet through a blunt-ended needle several times. The supernatant was collected after centrifugation of the lysate at 1000xg for 10 min at 4 °C. and then centrifuged again at 6000xg for 10 min at 4 °C The mitochondrial-containing pellet was collected, washed, resuspended, and stored in mitochondrial storage buffer. The mitochondrial samples were prepared using 2X Laemmli buffer, and Western blotting was performed with PAI-1 mouse (C-09) antibodies as described before [7].
Results
Various Secretory Serpins from Different Clades are Not Obligatory Extracellular and Are Found to Be Localized Inside the Cells
Out of 36 human serpins (Table S1), immunofluorescence data of 16 serpins in various cell lines were found in HPA and retrieved (Table 1, Table S3). Interestingly, the secretory serpins from different clades (A1, A10, H1, I1, D1, and E2) were intracellularly localized (Figure 1A). In agreement with the previous reports, the Clade B serpins (B1, B3-B10, and B13) were also found to be intracellular (Figure 2). For further investigation, the subcellular localization information of 20 serpins was retrieved from SubCell BarCode, in which clade A secretory serpins (A4, A6, A7, and A10) and the serpins from other clades (F1 and G1) were found in the cytosol (Table 1). Consistent with HPA data, serpins D1, E2, H1, and I1 were also found intracellularly. Additionally, classical intracellular serpins of Clade B (serpin B1-4, B8, B9, and B11-B13) were localized inside the cells (Table 1). A total of 11 serpins (A2, A3, A5, A8, A9, A11, A12, C1, E3 F2 and I2) were not found in either database. These data showed for the first time that various secretory serpins from different clades are not obligatory extracellular and might have novel intracellular functions.
Secretory PAI-1 was previously reported to be present in the nucleus of endothelial cells [9]. Here, to explore the subcellular distribution of PAI-1 in HeLa cells. Indeed, we identified that PAI-1 is present in the nucleus, especially in the PML bodies (Figure 1B). Interestingly, in the SubCell Barcode database, the PAI-1 protein was also identified in the mitochondrial fraction in the U251 cell line. To verify the localization of PAI-1, we isolated the mitochondria from HUVECs and HeLa cell lines. Interestingly, PAI-1 was observed in the mitochondria of HUVECs but not in HeLa cells. It suggests that mitochondrial localization of PAI-1 is highly regulated and dependent on cell type and specific environment.
Various Serpins are Localized in the Nucleus and Have Potential Nuclear Localization Signals
This study found that serpins A10, B2, B8-B11, B13, and H1 are mainly localized in the nucleus, at least as shown by one subcellular resource (Table 1), suggesting that these serpins might have a nuclear localization signal (NLS). We used the ELM database and the cNLS mapper tool to identify NLS in human serpins. Also, we manually screened for basic amino acids (R and K) clusters in the human serpins’ primary sequences to identify classical NLSs. We identified a single classical NLS motif in serpin B2. Two motifs were identified in A10 and B5 (Table 2). Using the ELM server, we couldn’t identify any NLS in these identified nuclear serpins (Table 2). These data showed that the identified nuclear serpins A10, B2, and B5 had potential classical NLS motifs. Interestingly, these tools couldn’t identify NLS in serpins B8-B11, B13, and H1. To explore this further, we manually analyzed the serpin sequences. We found one basic amino acid-rich NLS motif in serpin B8 and B13 and two NLS motifs in B10 (Table 2), suggesting that these serpins have potential classical NLS.
We also found a specific nonclassical NLS (PY-NLS) in 8 serpins (A2, A3, A5, A10, A12, B1, B3, and E1). Previously, we reported that serpin E1 (PAI-1) didn’t have any classical NLS [9]. Notably, in this study, we identified a nonclassical PY-NLS motif in serpins A3 and E1. None of the used methods was able to identify a potential NLS in sepin B9, B11, and H1, suggesting that these serpins may be transported to the nucleus via an indirect mechanism, such as by interacting with NLS-containing proteins. Interestingly, we found that 9 serpins (A2, A3, A5, B1, B3, B4, D1, E1, and I1) had potential NLSs. But, these serpins were not shown in the nucleus by any subcellular databases (Table 2). Notably, the serpins B1, B3, B4, D1, and I1 were localized in the cytosol (Table 1). Additionally, we found that serpin A9 and A12 have potential NLS; however, we couldn’t find the subcellular data from the databases. Serpins A5 contains 2 classical NLS motifs, and serpins B1 and I1 have a single classical NLS motif identified using the ELM server. Serpins B1 had 4 NLS (3 classical NLS and 1 nonclassical NLS). Serpin A3 and E1 had one nonclassical NLS motif. Serpin A9 (2 motifs), B4 (1 motif), and D1 (1 motif) had classical NLS. Serpin I1 had one NLS predicted by the ELM database only. Serpin A2 and A5 had 2 classical NLSs and 1 nonclassical NLS. A12 has 1 classical NLS and 2 nonclassical NLSs. B3 has 1 classical NLS and 1 nonclassical NLS. These data suggest that the localization of these serpins is highly regulated and may require specific signaling to import them into the nucleus using identified potential NLSs.
Identification of Potential Nuclear Export Signals in the Serpin Superfamily
Here, we used the ELM database and the LocNES tool to identify NESs in human serpins, followed by manual curation of the predicted NES motifs. The LocNES tool predicted multiple putative NES motifs in 24 human serpins. Notably, the ELM database confirmed the presence of NESs in 18 of these serpins, with the exception of six (B1, E3, I1, I2, A10, and A11), for which no NES motifs were identified. This substantial overlap highlights the robustness of the predictions while also underscoring tool-specific differences. This divergence may be attributed to differences in the underlying prediction algorithms and sensitivity thresholds of each tool. (Table S2).
All identified motifs were subsequently subjected to manual curation, and those that failed to meet the established criteria for known NES classes were excluded from the final analysis. It has been depicted that the NES should follow consensus patterns, possess solvent-accessibility, and have a helical/coiled secondary structure [17]. The secondary structure and surface accessibility of the predicted NES motifs were analyzed within the structural models of human serpins. NES motifs identified in four serpins (B3, B9, B11, and I2) exhibited a solvent-accessible surface area (PSA) of less than 25%, suggesting they are likely buried and not accessible on the protein surface. Additionally, these motifs adopted β-sheet-like conformations, which deviate from the typical helical/coil structure commonly associated with functional NESs. Hence, these motifs were rejected. In the remaining 20 serpins from/assigned to different clades, a total of 27 NES motifs were retained, as they met all established criteria for potential NES (Table 3). The surface accessibility scores and the secondary structure conformations of these selected NESs are shown in Figure S1. No NES motifs were identified in serpins belonging to clades D, F, and G. Interestingly, we found that 10 serpins (A4, A6, A11, B6, B7, B12, C1, E2, E3, and H1) contained NES motifs but lacked identifiable NLS motifs. Another group of 10 serpins (A10, B1, B2, B4, B5, B8, B10, B13, E1, and I1) harbored both NES and NLS motifs, whereas seven other serpins (A2, A3, A5, A9, A12, B3, and D1) possessed only NLS motifs (Table 4). These findings suggest that this subset of serpins may actively shuttle between the nucleus and cytoplasm, potentially engaging in specific functions in the nucleus.
Conservation of Identified NES Motifs in Human SERPIN Superfamily
We investigated whether the identified NES motifs are conserved across members of the human serpin superfamily. Interestingly, multiple sequence alignments (MSA) of human serpin sequences revealed that most NES consensus motifs were conserved and localized within the same regions across the majority of serpins. Notably, each conserved region contained only one type of NES consensus class. Based on this observation, we classified these regions into six distinct groups (Group A to Group F) (Figure 3). In addition, several NES motifs were found in serpins that did not align with any of the defined groups. These were categorized separately as unique groups, referred to as Group U. Among all groups, Group F emerged as the most prominent and conserved region, encompassing 11 of the 27 predicted NES motifs. In comparison, Group U contained six NES motifs (Table 3). Group F is characterized by a typical class 1a NES consensus pattern (Φ0xxΦ1xxxΦ2xxΦ3xΦ4) and is located in the region of 270-370 amino acids.
Discussion
It is believed that vertebrate serpins, except Clade B, function exclusively in the extracellular environment [2,27,28]. The bioinformatic analysis of human serpin superfamily members revealed that Clade B members may shuttle between the nucleus and cytoplasm. This study identifies the unique behavior of many extracellular serpins to be present inside the cell, e.g., the mitochondria, nucleus, and possesses either NLS or NES or both. The subcellular localization of secretory serpins in the cells gives a strong hint about their intracellular functions and alternative protein trafficking, which might be secretory signal-independent and regulated by novel regulatory pathways.
We explored two major subcellular resource databases based on high-throughput studies of global subcellular distribution profiles of proteins, i.e., the HPA and Subcell Barcode [23,24]. In the HPA database, we could retrieve the immunofluorescence data of 16 serpins. Surprisingly, we found that various secretory serpins from clades A, D, E, H, and I are not obligatory extracellular and are present inside the various subcellular compartments of the cells. Notably, the identified secretory serpins were not only found in the cytoplasm and nucleus but also in other subcompartments such as mitochondria, cytoskeleton, and ER. Interestingly, PAI-1 was present in the mitochondria of endothelial cells but absent in HeLa cells, suggesting its cell-specific intracellular localization of PAI-1 that should be explored in future.
Moreover, the subcellular localization data of Serpin A1, A2, and I1 were unavailable in either subcellular resource. Some previous studies showed that they might be present in the ER [10,29], but we couldn’t detect the classical ER motifs in these serpins. Serpin A2 was co-localized with CANX and PDIA3, two chaperones involved in ER quality control processing [30,31]. Interestingly, in COS cells, serpin I1 (neuroserpin) was reported in the ER or Golgi bodies [10]. These data suggest that these serpins might have functions in the ER and the Golgi.
Together, these data suggest the functions of these serpins inside the cell, and they might regulate cellular functions such as cell adhesion [32], apoptosis [33], and cell growth [9]. It was intriguing that the intracellular localization of secretory serpins is not in line with the evolution-based origin of secretory serpins, suggesting that the secretory serpins acquired additional extracellular functions during evolution, in addition to the intracellular functions, due to an increase in body complexity and biological functions. The novel regulatory pathway may have evolved to coordinate its sub-cellular distribution in accordance with specific cell types and a cell state-dependent manner.
In this study, we observed that most of the members of clade B and a few secretory serpins are mainly localized in the nucleus, at least as shown by one subcellular database. Most nuclear serpins contain basic residues (R, K) rich motifs with specific conserved patterns of NLS motifs. NLS analysis revealed that, except for serpin H1, we could identify NLS in all the identified nuclear serpins (Table 2), which indicates that they have an inherent property to localize in the nucleus. Consistent with previous studies, our findings also show that most intracellular serpins, particularly those from clade B, contain identifiable NLS motifs and are predominantly localized in the nucleus [34,35,36]. Surprisingly, we could not identify any NLS in serpin B9 (PI-9). Still, it is actively imported into the nucleus (Table 1) [37], suggesting that PI-9 may interact with an NLS-containing protein to enter the nucleus. It is unlikely that PI-9 passively diffuses in the nucleus due to its molecular weight (~40 kDa) [14,37,38].
Notably, some of the members of secretory serpins contain the potential NLS sequences. It is surprising that these secretory serpins were not only found inside the cells but also tend to shuttle between the nucleus and the cytoplasm, suggesting their functions in the nucleus that are yet to be explored. Previous reports showed that the nuclear localization of serpin A3 depended on the N-glycosylation pattern [39,40]. In this study, we found a potential PY-NLS motif in serpin A3, suggesting that N-glycosylation might help in translocating it into the nucleus.
We identified 27 NES motifs in 20 human serpins. Remarkably, 10 serpins possess only NES motifs, and the other 10 serpins contain both NLS and NES motifs. Serpin A4 and A6 have only NES and are found in the cytosol and the ER, respectively (Table 1). Also, it is interesting to note that serpins B6, B7, and B12 are found in the centrosome, ER, and mitochondria, respectively, and contain only NESs. Other secretory serpins, serpin A1, I1, and E1, have an N-terminal signal peptide, and we found that they contain both NLS and NES motifs, indicating that they might have functions in the nucleus. Furthermore, we could identify 7 serpins that possess only NLS and were not found in the nucleus. In subcellular resources, few serpins were cytosolic (Table 1) but had at least one potential NLS (Table 2). It is also observed in other proteins, such as LIMK1 and LIMK2, which possess functional NLS but are mainly localized in the cytoplasm [41]. The possible explanation is that these proteins might shuttle between the nucleus and cytoplasm, and active NES motifs are responsible for their cytoplasmic presence. Also, NLS might be exposed in specific conditions, and the protein may be localized in the nucleus.
Notably, we identified 6 regions (Group A to F) in serpins sequences where most identified NESs were present and possess a region-specific consensus class of NES. It was very interesting to observe that amongst these identified groups, group F contains the highest number of NES motifs from different serpins. Therefore, Group F is the largest group found in the region of 270-370 amino acids and follows the “1a” consensus pattern (Table 3), suggesting that this conservation might have an evolutionary significance.
In conclusion, this study might open avenues for future functional studies of secretory serpins in various physiological conditions and diseases. Some serpins (Table 5) have functions that do not require protease inhibition and have paradoxical physiological and pathophysiological functions. These secretory serpins identified inside the cells may be used as a reference in identifying the intracellular functions to decipher the precise role in various human diseases.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Acknowledgments
The work was supported by the grants from the DBT, Govt. of India (DBT Builder Project (BT/INF/22/SP44383/2021), DBT-PG (BT/HRD/01/59/2020), ICMR, Govt. of India (6/9-7(234)2020/ECD-II)), and DST-SERB, Govt. of India (DST-SERB CRG/2022/007356). PR. G. and A.Y. are the recipients of fellowships from CSIR, Govt. of India (CSIR-NET SRF; 09/1131(0039)/2019-EMR-I) and DBT, Govt. of India (DBT/2020/CUR/1470), respectively.
Conflict of interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of the data; in the writing of the manuscript; or in the decision to publish the results.
References
- Law, R.H.P.; Zhang, Q.; McGowan, S.; Buckle, A.M.; Silverman, G.A.; Wong, W.; Rosado, C.J.; Langendorf, C.G.; Pike, R.N.; Bird, P.I.; et al. An overview of the serpin superfamily. Genome Biol. 2006, 7, 216. [Google Scholar] [CrossRef]
- Silverman, G.A.; Whisstock, J.; Askew, D.J.; Pak, S.C.; Luke, C.J.; Cataltepe, S.; Irving, J.A.; Bird, P.I. Human clade B serpins (ov-serpins) belong to a cohort of evolutionarily dispersed intracellular proteinase inhibitor clades that protect cells from promiscuous proteolysis. Cell. Mol. Life Sci. 2004, 61, 301–325. [Google Scholar] [CrossRef]
- van Gent, D.; Sharp, P.; Morgan, K.; Kalsheker, N. Serpins: structure, function and molecular evolution. Int. J. Biochem. Cell Biol. 2003, 35, 1536–1547. [Google Scholar] [CrossRef]
- Gettins, P.G.W. Serpin Structure, Mechanism, and Function. Chem. Rev. 2002, 102, 4751–4804. [Google Scholar] [CrossRef]
- Irving, J.A.; Pike, R.N.; Lesk, A.M.; Whisstock, J.C. Phylogeny of the Serpin Superfamily: Implications of Patterns of Amino Acid Conservation for Structure and Function. Genome Res. 2000, 10, 1845–1864. [Google Scholar] [CrossRef]
- Kumar, A. Bayesian phylogeny analysis of vertebrate serpins illustrates evolutionary conservation of the intron and indels based six groups classification system from lampreys for ∼500 MY. PeerJ 2015, 3, e1026. [Google Scholar] [CrossRef]
- Kubala, M.H.; DeClerck, Y.A. The plasminogen activator inhibitor-1 paradox in cancer: a mechanistic understanding. Cancer Metastasis Rev. 2019, 38, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Spence, M.A.; Mortimer, M.D.; Buckle, A.M.; Minh, B.Q.; Jackson, C.J. A Comprehensive Phylogenetic Analysis of the Serpin Superfamily. Mol. Biol. Evol. 2021, 38, 2915–2929. [Google Scholar] [CrossRef]
- Gehlot, P.; Brünnert, D.; Kaushik, V.; Yadav, A.; Bage, S.; Gaur, K.; Saini, M.; Ehrhardt, J.; Manjunath, G.K.; Kumar, A.; et al. Unconventional localization of PAI-1 in PML bodies: A possible link with cellular growth of endothelial cells. Biochem. Biophys. Rep. 2024, 39, 101793. [Google Scholar] [CrossRef] [PubMed]
- Marques, P.I.; Ferreira, Z.; Martins, M.; Figueiredo, J.; Silva, D.I.; Castro, P.; Morales-Hojas, R.; Simões-Correia, J.; Seixas, S. SERPINA2 Is a Novel Gene with a Divergent Function from SERPINA1. PLOS ONE 2013, 8, e66889. [Google Scholar] [CrossRef]
- Ito, S.; Nagata, K. Biology of Hsp47 (Serpin H1), a collagen-specific molecular chaperone. Semin. Cell Dev. Biol. 2017, 62, 142–151. [Google Scholar] [CrossRef]
- Lu, J.; Wu, T.; Zhang, B.; Liu, S.; Song, W.; Qiao, J.; Ruan, H. Types of nuclear localization signals and mechanisms of protein import into the nucleus. Cell Commun. Signal. 2021, 19, 60. [Google Scholar] [CrossRef]
- Kosugi, S.; Hasebe, M.; Matsumura, N.; Takashima, H.; Miyamoto-Sato, E.; Tomita, M.; Yanagawa, H. Six Classes of Nuclear Localization Signals Specific to Different Binding Grooves of Importin α. J. Biol. Chem. 2009, 284, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Görlich, D.; Kutay, U. Transport Between the Cell Nucleus and the Cytoplasm. Annu. Rev. Cell Dev. Biol. 1999, 15, 607–660. [Google Scholar] [CrossRef]
- Tessier, T.M.; MacNeil, K.M.; Mymryk, J.S. Piggybacking on Classical Import and Other Non-Classical Mechanisms of Nuclear Import Appear Highly Prevalent within the Human Proteome. Biology 2020, 9, 188. [Google Scholar] [CrossRef] [PubMed]
- Fung, H.Y.J.; Fu, S.-C.; Chook, Y.M. Nuclear export receptor CRM1 recognizes diverse conformations in nuclear export signals. eLife 2017, 6, RP89040. [Google Scholar] [CrossRef]
- Lee, Y.; Pei, J.; Baumhardt, J.M.; Chook, Y.M.; Grishin, N.V. Structural prerequisites for CRM1-dependent nuclear export signaling peptides: accessibility, adapting conformation, and the stability at the binding site. Sci. Rep. 2019, 9, 6627. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, S.; Hasebe, M.; Tomita, M.; Yanagawa, H. Nuclear Export Signal Consensus Sequences Defined Using a Localization-Based Yeast Selection System. Traffic 2008, 9, 2053–2062. [Google Scholar] [CrossRef]
- UniProt Consortium UniProt: the Universal Protein Knowledgebase in 2025. Nucleic Acids Res. 2025, 53, D609–D617. [CrossRef]
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. 2017, 27, 135–145. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; The UGENE Team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef]
- Thul, P.J.; Åkesson, L.; Wiking, M.; Mahdessian, D.; Geladaki, A.; Ait Blal, H.; Alm, T.; Asplund, A.; Björk, L.; Breckels, L.M.; et al. A subcellular map of the human proteome. Science 2017, 356, eaal3321. [Google Scholar] [CrossRef]
- Orre, L.M.; Vesterlund, M.; Pan, Y.; Arslan, T.; Zhu, Y.; Woodbridge, A.F.; Frings, O.; Fredlund, E.; Lehtiö, J. SubCellBarCode: Proteome-wide Mapping of Protein Localization and Relocalization. Mol. Cell 2019, 73, 166–182.e7. [Google Scholar] [CrossRef]
- Otlewski, J.; Jelen, F.; Zakrzewska, M.; Oleksy, A. The many faces of protease–protein inhibitor interaction. EMBO J. 2005, 24, 1303–1310. [Google Scholar] [CrossRef]
- Kaushik, V.; Brünnert, D.; Hanschmann, E.-M.; Sharma, P.K.; Anand, B.G.; Kar, K.; Kateriya, S.; Goyal, P. The intrinsic amyloidogenic propensity of cofilin-1 is aggravated by Cys-80 oxidation: A possible link with neurodegenerative diseases. Biochem. Biophys. Res. Commun. 2021, 569, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Remold-O'DOnnell, E. The ovalbumin family of serpin proteins. FEBS Lett. 1993, 315, 105–108. [Google Scholar] [CrossRef]
- Silverman, G.A.; Bird, P.I.; Carrell, R.W.; Church, F.C.; Coughlin, P.B.; Gettins, P.G.; Irving, J.A.; Lomas, D.A.; Luke, C.J.; Moyer, R.W.; et al. The serpins are an expanding superfamily of structurally similar but functionally diverse proteins. Evolution, mechanism of inhibition, novel functions, and a revised nomenclature. J. Biol. Chem. 2001, 276, 33293–33296. [Google Scholar] [CrossRef]
- Ishigami, S.; Sandkvist, M.; Tsui, F.; Moore, E.; Coleman, T.A.; Lawrence, D.A. Identification of a novel targeting sequence for regulated secretion in the serine protease inhibitor neuroserpin. Biochem. J. 2007, 402, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Raykhel, I.; Alanen, H.; Salo, K.; Jurvansuu, J.; Nguyen, V.D.; Latva-Ranta, M.; Ruddock, L. A molecular specificity code for the three mammalian KDEL receptors. J. Cell Biol. 2007, 179, 1193–1204. [Google Scholar] [CrossRef] [PubMed]
- Alanen, H.I.; Williamson, R.A.; Howard, M.J.; Hatahet, F.S.; Salo, K.E.H.; Kauppila, A.; Kellokumpu, S.; Ruddock, L.W. ERp27, a New Non-catalytic Endoplasmic Reticulum-located Human Protein Disulfide Isomerase Family Member, Interacts with ERp57. J. Biol. Chem. 2006, 281, 33727–33738. [Google Scholar] [CrossRef]
- Wu, S.; Yang, Y.; Zhang, M.; Khan, A.U.; Dai, J.; Ouyang, J. Serpin peptidase inhibitor, clade E, member 2 in physiology and pathology: recent advancements. Front. Mol. Biosci. 2024, 11, 1334931. [Google Scholar] [CrossRef]
- Balsara, R.D.; Ploplis, V.A. Plasminogen activator inhibitor-1: The double-edged sword in apoptosis. Thromb. Haemost. 2008, 100, 1029–1036. [Google Scholar] [CrossRef]
- Medcalf, R.L.; Stasinopoulos, S.J. The undecided serpin. The ins and outs of plasminogen activator inhibitor type 2. FEBS J. 2005, 272, 4858–4867. [Google Scholar] [CrossRef]
- Goulet, B.; Kennette, W.; Ablack, A.; O Postenka, C.; Hague, M.N.; Mymryk, J.S.; Tuck, A.B.; Giguère, V.; Chambers, A.F.; Lewis, J.D. Nuclear localization of maspin is essential for its inhibition of tumor growth and metastasis. Lab Invest 2011, 91, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Longhi, M.T.; Silva, L.E.; Pereira, M.; Magalhães, M.; Reina, J.; Vitorino, F.N.L.; Gumbiner, B.M.; da Cunha, J.P.C.; Cella, N. PI3K-AKT, JAK2-STAT3 pathways and cell–cell contact regulate maspin subcellular localization. Cell Commun. Signal. 2021, 19, 86. [Google Scholar] [CrossRef] [PubMed]
- Bird, C.H.; Blink, E.J.; Hirst, C.E.; Buzza, M.S.; Steele, P.M.; Sun, J.; Jans, D.A.; Bird, P.I. Nucleocytoplasmic Distribution of the Ovalbumin Serpin PI-9 Requires a Nonconventional Nuclear Import Pathway and the Export Factor Crm1. Mol. Cell. Biol. 2001, 21, 5396–5407. [Google Scholar] [CrossRef]
- Andreu, I.; Granero-Moya, I.; Chahare, N.R.; Clein, K.; Molina-Jordán, M.; Beedle, A.E.M.; Elosegui-Artola, A.; Abenza, J.F.; Rossetti, L.; Trepat, X.; et al. Mechanical force application to the nucleus regulates nucleocytoplasmic transport. Nat. Cell Biol. 2022, 24, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Soman, A.; Nair, S.A. Unfolding the cascade of SERPINA3: Inflammation to cancer. Biochim Biophys Acta Rev Cancer 2022, 1877, 188760. [Google Scholar] [CrossRef]
- Santamaria, M.; Pardo–Saganta, A.; Alvarez–Asiain, L.; Di Scala, M.; Qian, C.; Prieto, J.; Avila, M.A. Nuclear α1-Antichymotrypsin Promotes Chromatin Condensation and Inhibits Proliferation of Human Hepatocellular Carcinoma Cells. Gastroenterology 2013, 144, 818–828.e814. [Google Scholar] [CrossRef]
- Goyal, P.; Pandey, D.; Behring, A.; Siess, W. Inhibition of Nuclear Import of LIMK2 in Endothelial Cells by Protein Kinase C-dependent Phosphorylation at Ser-283. J. Biol. Chem. 2005, 280, 27569–27577. [Google Scholar] [CrossRef]
- Lonardo, F.; Li, X.; Siddiq, F.; Singh, R.; Al-Abbadi, M.; Pass, H.I.; Sheng, S. Maspin nuclear localization is linked to favorable morphological features in pulmonary adenocarcinoma. Lung Cancer 2006, 51, 31–39. [Google Scholar] [CrossRef]
- Jakobsen, L.; Vanselow, K.; Skogs, M.; Toyoda, Y.; Lundberg, E.; Poser, I.; Falkenby, L.G.; Bennetzen, M.; Westendorf, J.; A Nigg, E.; et al. Novel asymmetrically localizing components of human centrosomes identified by complementary proteomics methods. EMBO J. 2011, 30, 1520–1535. [Google Scholar] [CrossRef] [PubMed]
- Sauk JJ, Nikitakis N, and Siavash H. Hsp47 a novel collagen binding serpin chaperone, autoantigen and therapeutic target. Front. Biosci. 2005, 10, 107–18. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.-M.; Mi, Y.-S.; Yu, F.-D.; Han, Y.; Liu, X.-S.; Lu, S.; Zhang, Y.; Zhao, S.-L.; Ye, L.; Liu, T.-T.; et al. SERPINA4 is a novel independent prognostic indicator and a potential therapeutic target for colorectal cancer. Am J Cancer Res 2016, 6, 1636–1649. [Google Scholar] [PubMed]
- Sanchez-Navarro, A.; González-Soria, I.; Caldiño-Bohn, R.; Bobadilla, N.A. Integrative view of serpins in health and disease: the contribution of SerpinA3. Am. J. Physiol. Physiol. 2021, 320, C106–C118. [Google Scholar] [CrossRef]
- Janciauskiene, S.; Lechowicz, U.; Pelc, M.; Olejnicka, B.; Chorostowska-Wynimko, J. Diagnostic and therapeutic value of human serpin family proteins. Biomed. Pharmacother. 2024, 175, 116618. [Google Scholar] [CrossRef]
- Van de Water, N.; Tan, T.; Ashton, F.; O'Grady, A.; Day, T.; Browett, P.; Ockelford, P.; Harper, P. Mutations within the protein Z-dependent protease inhibitor gene are associated with venous thromboembolic disease: a new form of thrombophilia. Br. J. Haematol. 2004, 127, 190–194. [Google Scholar] [CrossRef]
- Song, Y.; Li, Z.; Li, L.; Zhou, H.; Zeng, T.-T.; Jin, C.; Lin, J.-R.; Gao, S.; Li, Y.; Guan, X.-Y.; et al. SERPINA11 Inhibits Metastasis in Hepatocellular Carcinoma by Suppressing MEK/ERK Signaling Pathway. J. Hepatocell. Carcinoma 2021, 8, 759–771. [Google Scholar] [CrossRef]
- Sillen, M.; Declerck, P.J. A Narrative Review on Plasminogen Activator Inhibitor-1 and Its (Patho)Physiological Role: To Target or Not to Target? Int. J. Mol. Sci. 2021, 22, 2721. [Google Scholar] [CrossRef]
- Madjene, C.; Boutigny, A.; Bouton, M.-C.; Arocas, V.; Richard, B. Protease Nexin-1 in the Cardiovascular System: Wherefore Art Thou? Front. Cardiovasc. Med. 2021, 8, 652852. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Subcellular localization of various extracellular serpins. A) The immunofluorescence confocal images were retrieved from the HPA subcellular resource. The upper panel (green) shows the staining of the respective serpins. Middle panel: the cell nuclei were stained with DAPI (blue), and the ER was stained with calreticulin antibody, which is shown in yellow. The overlay is shown in the lower panel. B) HeLa cells were stained with specific PAI-1 antibody (left panel, green), nuclei with Hoechst 33258 (middle panel, blue), PML bodies with specific PML antibody, and overlay picture (right panel). PAI-1 was found in the nucleus. C) The mitochondria of HUVECs and HeLa cells were isolated, and then a Western blot was performed using the PAI-1 antibody. A band of PAI-1 is observed in the mitochondria of HUVECs but not in HeLa cells.
Figure 1.
Subcellular localization of various extracellular serpins. A) The immunofluorescence confocal images were retrieved from the HPA subcellular resource. The upper panel (green) shows the staining of the respective serpins. Middle panel: the cell nuclei were stained with DAPI (blue), and the ER was stained with calreticulin antibody, which is shown in yellow. The overlay is shown in the lower panel. B) HeLa cells were stained with specific PAI-1 antibody (left panel, green), nuclei with Hoechst 33258 (middle panel, blue), PML bodies with specific PML antibody, and overlay picture (right panel). PAI-1 was found in the nucleus. C) The mitochondria of HUVECs and HeLa cells were isolated, and then a Western blot was performed using the PAI-1 antibody. A band of PAI-1 is observed in the mitochondria of HUVECs but not in HeLa cells.
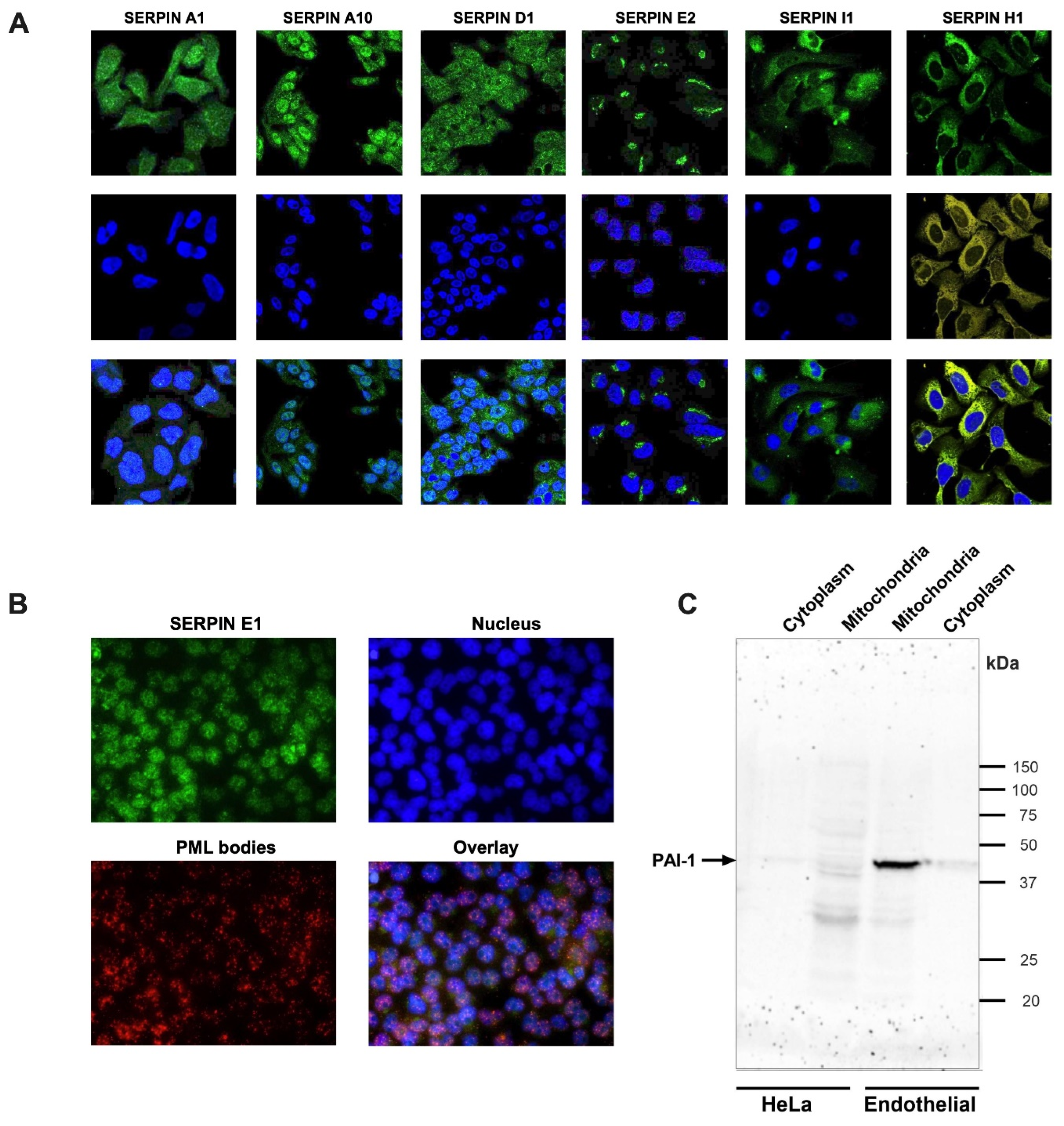
Figure 2.
Subcellular localization of Clade B serpins. The immunofluorescence confocal images were retrieved from the HPA subcellular resource. The upper panel (green) shows the staining of the respective serpins. Middle panel: the cell nuclei were stained with DAPI (blue). The overlay is shown on the lower panel.
Figure 2.
Subcellular localization of Clade B serpins. The immunofluorescence confocal images were retrieved from the HPA subcellular resource. The upper panel (green) shows the staining of the respective serpins. Middle panel: the cell nuclei were stained with DAPI (blue). The overlay is shown on the lower panel.
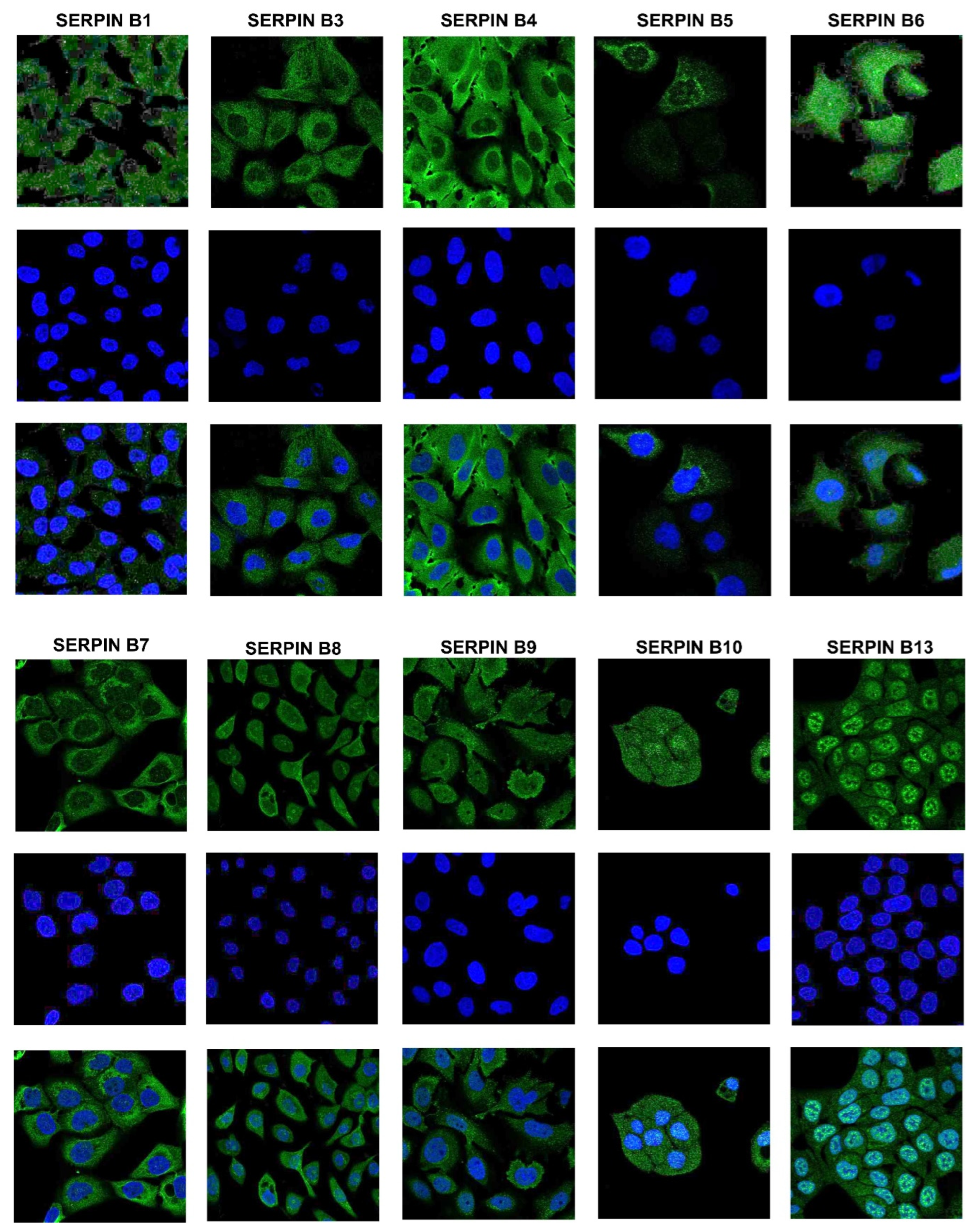
Figure 3.
The multiple sequence alignment of NES motifs from clades A-I of human serpins. The multiple sequence alignment of identified NES motifs in members of the serpin superfamily. The sequences of the identified NES motifs in human serpins from clades A-I of vertebrates were aligned. The specific class of consensus pattern in each NES motif is shown here. The conserved hydrophobic amino acids are shown in boxes. Notably, most of the NES consensus sequences were conserved, and only one type of NES consensus class was present in one region. These regions are divided into 6 groups (group A to group F).
Figure 3.
The multiple sequence alignment of NES motifs from clades A-I of human serpins. The multiple sequence alignment of identified NES motifs in members of the serpin superfamily. The sequences of the identified NES motifs in human serpins from clades A-I of vertebrates were aligned. The specific class of consensus pattern in each NES motif is shown here. The conserved hydrophobic amino acids are shown in boxes. Notably, most of the NES consensus sequences were conserved, and only one type of NES consensus class was present in one region. These regions are divided into 6 groups (group A to group F).
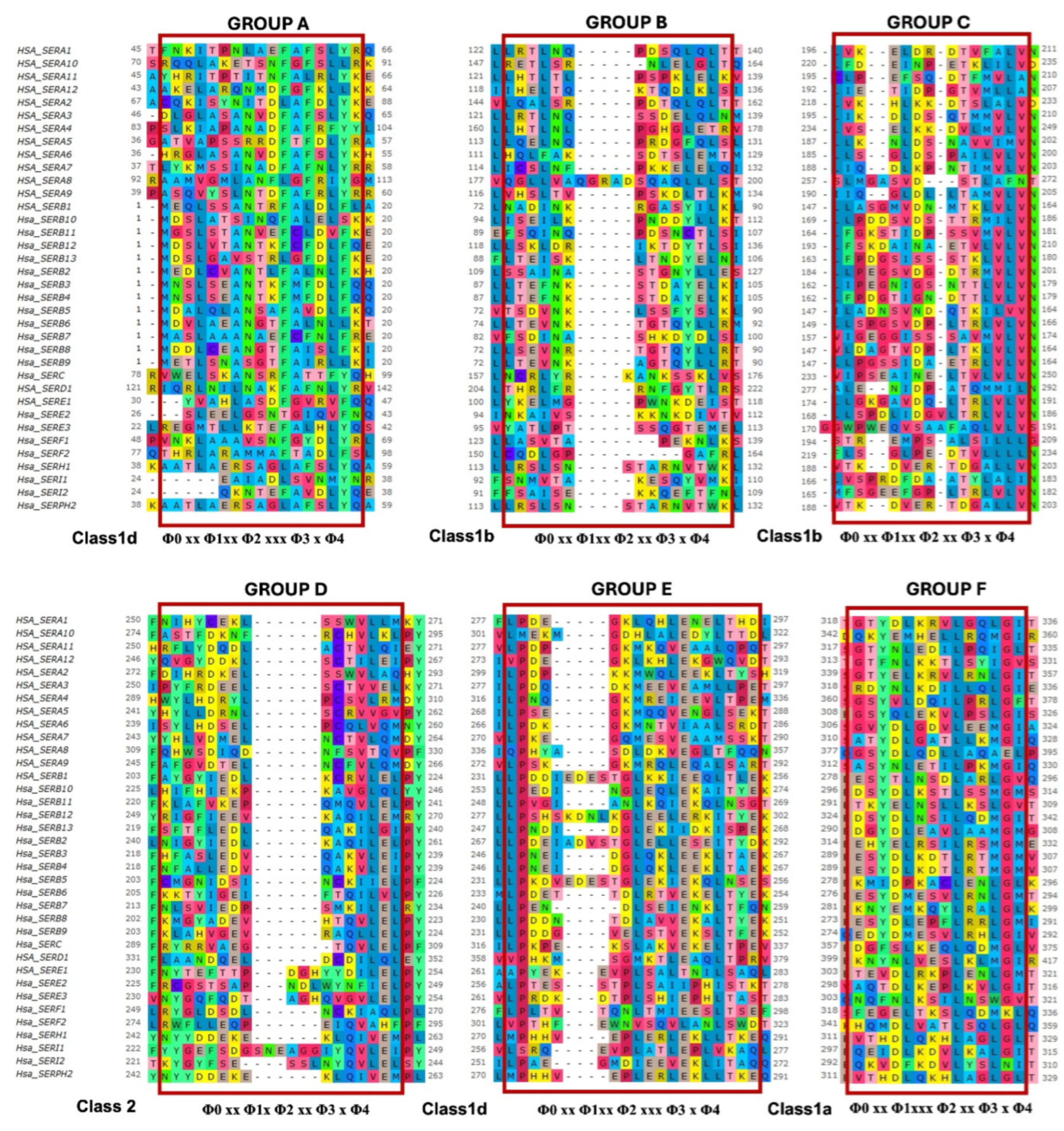

Table 1.
Subcellular localization of human serpins. The subcellular distributions of human serpins were analyzed using the subcellular resource of the Human Protein Atlas (HPA) and Subcell Barcode resources. PM: plasma membrane; ER: endoplasmic reticulum.
Table 1.
Subcellular localization of human serpins. The subcellular distributions of human serpins were analyzed using the subcellular resource of the Human Protein Atlas (HPA) and Subcell Barcode resources. PM: plasma membrane; ER: endoplasmic reticulum.
| S.No. | Clade | Protein code | HPA | Cell line | Subcell Barcode | Cell line | Literature |
| 1. | A | SERPINA1 | Vesicles | HepG2 | N/A | N/A | ER [10] |
| 2. | SERPINA2 | N/A | N/A | N/A | N/A | ER [10] | |
| 3. | SERPINA3 | N/A | N/A | N/A | N/A | Nucleus [40], [39] |
|
| 4. | SERPINA4 | N/A | N/A | Cytoplasm | MCF7 | N/A | |
| 5. | SERPINA6 | N/A | N/A | Cytoplasm | MCF7 | N/A | |
| 6. | SERPINA7 | N/A | N/A | Cytoplasm | MCF7 | N/A | |
| 7. | SERPINA10 | Cytosol, Nucleus | HepG2 | Cytoplasm | H322 | N/A | |
| 8. | B | SERPINB1 | Cytoplasm | SK-MEL-30 | Cytoplasm, cytoskeleton | H322 | N/A |
| 9. | SERPINB2 | N/A | N/A | Cytoplasm | A431, H322 | Nucleus [34] | |
| 10. | SERPINB3 | Cytoplasm, PM | A431 | Cytoplasm | H322 | N/A | |
| 11. | SERPINB4 | Cytoplasm, PM | U2OS | Cytoplasm | H322, HCC827 | N/A | |
| 12. | SERPINB5 | Vesicles | A431 | Cytoplasm | A431, H322, MCF7, HCC827 | Nucleus [35,36,42] | |
| 13. | SERPINB6 | Centrosome | U251-MG | NA | A431 | Centrosome [43] | |
| 14. | SERPINB7 | Mitochondria, ER | HaCaT | NA | H322 | N/A | |
| 15. | SERPINB8 | Golgi bodies, Nucleus | HBEC3KT | Cytoplasm, cytoskeleton | A431, HCC827 | N/A | |
| 16. | SERPINB9 | Cytoplasm, Nucleus | SiHa | Cytoplasm, cytoskeleton | HCC827 | Nucleus, cytoplasm [37] | |
| 17. | SERPINB10 | Cytoplasm, Nucleus | HaCaT | N/A | N/A | N/A | |
| 18. | SERPINB11 | N/A | N/A | Nuclear | HCC827 | N/A | |
| 19. | SERPINB12 | N/A | N/A | Mitochondria | HCC827 | N/A | |
| 20. | SERPINB13 | Cytoplasm, Nuclear speckles | RT-4 | Cytoplasm | H322 | N/A | |
| 21. | D | SERPIND1 | Vesicles | HepG2 | Cytoplasm | U251 | N/A |
| 22. | E | SERPINE1 | N/A | N/A | Mitochondria, ER | U251 | Nucleus [9] |
| 23. | SERPINE2 | Golgi bodies | SK-MEL-30 | PM | U251 | N/A | |
| 24. | F | SERPINF1 | N/A | N/A | Cytoplasm cytoskeleton | H322 | N/A |
| 25. | G | SERPING1 | N/A | N/A | Cytoplasm | H322, MCF7 | N/A |
| 26. | H | SERPINH1 | ER | U2OS | Nucleosol, ER | U251, A431 | ER [11,44] |
| 27 | I | SERPINI1 | Cytoplasm, vesicles | U251-MG | N/A | N/A | ER, Golgi bodies, vesicles [29] |
Table 2.
Prediction of NLS motifs in human serpins based on primary sequence analysis. The cNLS Mapper tool, ELM database, and manual curation method were used to identify putative NLS motifs in all human serpins.
Table 2.
Prediction of NLS motifs in human serpins based on primary sequence analysis. The cNLS Mapper tool, ELM database, and manual curation method were used to identify putative NLS motifs in all human serpins.
| S. No. | Name | Position | cNLS Mapper | ELM Database | Manual curation | ||
| Sequence | Score | Classical NLS | Non classical NLS (PY-NLS) | ||||
| 1 | SERPINA2 | 11-16 350-378 203-220 |
RNLGITKIFSNEADLSGVSQEAPLKLSKA |
5.3 | N/A |
EKRTGRKVVDLVKHLKKD |
HRLGPY |
| 2 | SERPINA3 | 247-253 | N/A | N/A | HHLTIPY | ||
| 3 | SERPINA5 | 254-260 294-300 292-299 |
N/A |
FKKRQLE KMFKKRQL |
RVVGVPY | ||
| 4 | SERPINA9 | 171-189 280-301 |
N/A | N/A | AQARINSHVKKKTQGKVV RQLEQALSARTLRKWSHSLQKR |
||
| 5 | SERPINA10 | 179-206 238-267 286-292 |
FNLSKRYFDTECVPMNFRNASQAKRLMN FKGKWLTPFDPVFTEVDTFHLDKYKTIKVP |
5.4 6.4 |
N/A |
HVLKLPY |
|
| 6 | SERPINA12 | 39-66 339-345 385-390 |
WKQRMAAKELARQNMDFGFKLLKKLAFN | 5.35 | N/A | N/A |
LTKIAPY KIDKPY |
| 7 | SERPINB1 | 168-197 189-205 185-200 215-221 |
KGNWKDKFMKEATTNAPFRLNKKDRKTVKM | 5.1 |
KKDRKTVKMMYQKKKFA |
FRLNKKDRKTVKMMYQ |
RVLELPY |
| 8 | SERPINB2 | 143-172 165-181 |
RLCQKYYSSEPQAVDFLECAEEARKKINSW | 6.3 | N/A |
ARKKINSWVKTQTKGKI |
|
| 9 | SERPINB3 | 143-160 229-236 |
N/A | N/A | SRKKINSWVESQTNEKIK |
AKVLEIPY |
|
| 10 | SERPINB4 | 142-150 | N/A | N/A | ESRKKINSW | ||
| 11 | SERPINB5 | 83-114 87-114 |
FYSLKLIKRLYVDKSLNLSTEFISSTKRPYAK KLIKRLYVDKSLNLSTEFISSTKRPYAK |
5.1 6.1 |
N/A | N/A | |
| 12 | SERPINB8 | 176-191 | N/A | N/A | N/A | RKYTRGMLFKTNEEKK | |
| 13 | SERPINB10 | 73-83 222-233 |
N/A | N/A | N/A | EKKRKMEFNLS MKKKLHIFHIEK |
|
| 14 | SERPINB13 | 143-155 | N/A | N/A | ESRKKINSWVESK | ||
| 15 | SERPIND1 | 203-219 | N/A | N/A | RKLTHRLFRRNFGYTLR | ||
| 16 | SERPINE1 | 54-61 | N/A | N/A | N/A | RNVVFSPY | |
| 17 | SERPINI1 | 283-289 | N/A | VKKQKVE | N/A | ||
Table 3.
Summary of the potential NESs identified in the various human serpins.
| Clade | Position | Sequence | Serpin | Group | Class | Consensus class |
| A | 116-130 | DVHRGFQHLLHTLNL | A4 | U | 1a | Φ0 xx Φ1xxx Φ2 xx Φ3 x Φ4 |
| 118-133 | SDTSLEMTMGNALFL | A6 | U | 1c | Φ0 xx Φ1xxx Φ2 xxx Φ3 x Φ4 | |
| 307-321 | SGVYDLGDVLEEMGI | A6 | F | 1a | Φ0 xx Φ1xxx Φ2 xx Φ3 x Φ4 | |
| 300-312 | VVLMEKMGDHLAL | A10 | U | 1d | Φ0 xx Φ1xx Φ2 xxx Φ3 x Φ4 | |
| 92-106 | QANTSALILEGLGFN | A11 | D | 2 | Φ0 xx Φ1x Φ2 xx Φ3 x Φ4 | |
| B | 238-252 | DESTGLKKIEEQLTL | B1 | E | 1d | Φ0 xx Φ1xx Φ2 xxx Φ3 x Φ4 |
| 279-293 | EESYTLNSDLARLGV | B1 | ||||
| 259-273 | TKPENLDFIEVNVSL | B1 | F | 1a | Φ0 xx Φ1xxx Φ2 xx Φ3 x Φ4 | |
| 268-282 | LPDEIADVSTGLELL | B2 | E | 1d | Φ0 xx Φ1xx Φ2 xxx Φ3 x Φ4 | |
| 292-306 | ESYDLKDTLRTMGMV | B4 | F | 1a | Φ0 xx Φ1xxx Φ2 xx Φ3 x Φ4 | |
| 74-88 | TSDVNKLSSFYSLKL | B5 | B | 1b | Φ0 xx Φ1xx Φ2 xx Φ3 x Φ4 | |
| 149-163 | LLSPGSVDPLTRLVL | B6 | C | 1b | Φ0 xx Φ1xx Φ2 xx Φ3 x Φ4 | |
| 241-255 | LPENDLSEIENKLTF | B7 | E | 1d | Φ0 xx Φ1xx Φ2 xxx Φ3 x Φ4 | |
| 283-297 | KNYEMKQYLRALGLK | B7 | F | 1a | Φ0 xx Φ1xxx Φ2 xx Φ3 x Φ4 | |
| 274-288 | EESYDLEPFLRRLGM | B8 | ||||
| 297-311 | ENSYDLKSTLSSMGM | B10 | ||||
| 305-319 | EDSYDLNSILQDMGI | B12 | ||||
| 218-232 | QMMFMKKKLHIFHIE | B10 | U | 1a | Φ0 xx Φ1xxx Φ2 xx Φ3 x Φ4 | |
| 221-235 | SFTFLEDLQAKILGI | B13 | D | 2 | Φ0 xx Φ1x Φ2 xx Φ3 x Φ4 | |
| 250-262 | NDIDGLEKIIDKI | B13 | U | 1aR | Φ0 x Φ1xx Φ2 xxx Φ3 xx Φ4 | |
| C | 358-372 | EDGFSLKEQLQDMGL | C1 | F | 1a | Φ0 xx Φ1xxx Φ2 xx Φ3 x Φ4 |
| E | 306-318 | ETEVDLRKPLENLGM | E1 | |||
| 150-164 | LSPDLIDGVLTRLVL | E2 | U | 1a | Φ0 xx Φ1xxx Φ2 xx Φ3 x Φ4 | |
| 175-189 | DGVLTRLVLVNAVYF | E2 | C | 1b | Φ0 xx Φ1xx Φ2 xx Φ3 x Φ4 | |
| 318-332 | VTDLFDPLKANLKGI | E3 | U | 1aR | Φ0 x Φ1xx Φ2 xxx Φ3 xx Φ4 | |
| H | 312-326 | EVTHDLQKHLAGLGL | H1 | F | 1a | Φ0 xx Φ1xxx Φ2 xx Φ3 x Φ4 |
| I | 298-312 | EQEIDLKDVLKALGI | I1 | F | 1a | Φ0 xx Φ1xxx Φ2 xx Φ3 x Φ4 |
Table 4.
Summary of identified NLS and NES in human serpins.
| S.no. | Clade | Protein code | NLS | NES | S.no. | Clade | Protein code | NLS | NES |
| 1. | A | SERPINA2 | NLS | N/A | 15. | B. | SERPINB6 | N/A | NES |
| 2. | SERPINA3 | NLS | N/A | 16. | SERPINB7 | N/A | NES | ||
| 3. | SERPINA4 | N/A | NES | 17. | SERPINB8 | NLS | NES | ||
| 4. | SERPINA5 | NLS | N/A | 18. | SERPINB10 | NLS | NES | ||
| 5. | SERPINA6 | N/A | NES | 19. | SERPINB12 | N/A | NES | ||
| 6. | SERPINA9 | NLS | N/A | 20. | SERPINB13 | NLS | NES | ||
| 7. | SERPINA10 | NLS | NES | 21. | C. | SERPINC1 | N/A | NES | |
| 8. | SERPINA11 | N/A | NES | 22. | D. | SERPIND1 | NLS | N/A | |
| 9. | SERPINA12 | NLS | N/A | 23. | E. | SERPINE1 | NLS | NES | |
| 10. | B | SERPINB1 | NLS | NES | 24. | SERPINE2 | N/A | NES | |
| 11. | SERPINB2 | NLS | NES | 25. | SERPINE3 | N/A | NES | ||
| 12. | SERPINB3 | NLS | N/A | 26. | H. | SERPINH1 | N/A | NES | |
| 13. | SERPINB4 | NLS | NES | 27. | I. | SERPINI1 | NLS | NES | |
| 14. | SERPINB5 | NLS | NES | ||||||
Table 5.
List of the secretory serpins that are of interest and recommended for future studies.
| S.No. | Clade | Protein code | Localization signal | Functions (protease inhibition-independent) and associated diseases | References | |
| NLS | NES | |||||
| 1. | A | SERPINA3 | NLS | N/A | Inflammation, apoptosis, cancer diagnosis, Alzheimer’s and emphysema | [39,40] |
| 2. | SERPINA4 | N/A | NES | Inflammation, Diabetic retinopathy, novel prognostic indicator and therapeutic target for Colorectal cancer. | [45,46] | |
| 3. | SERPINA5 | NLS | N/A | Coagulation, sperm development, tumor cell invasion and metastasis | [28] | |
| 4. | SERPINA6 | N/A | NES | Hormone transport | [28] | |
| 5. | SERPINA9 | NLS | N/A | B cell development | [28] | |
| 6. | SERPINA10 | NLS | NES | Venous thromboembolic disease | [47,48] | |
| 7. | SERPINA11 | N/A | NES | Hepatocellular carcinoma | [47,49] | |
| 8. | SERPINA12 | NLS | N/A | Anti-insulin resistance and Obesity | [28] | |
| 9. | E | SERPINE1 | NLS | NES | Cell Growth , aging, cancer prognosis | [7,9,50] |
| 10. | SERPINE2 | N/A | NES | Neurotrophic factors, emphysema, thoracic aortic aneurysms and atherosclerosis. | [28,46,47,51] | |
| 11. | H | SERPINH1 | N/A | NES | Rheumatoid arthritis, cancer and aortic stenosis | [11,44] |
| 12. | I | SERPINI1 | NLS | NES | Neurotrophic factors | [28] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



