
Submitted:
14 October 2025
Posted:
15 October 2025
You are already at the latest version
Abstract
Hepatocellular carcinoma (HCC) is a leading cause of cancer mortality worldwide, with chronic hepatitis B virus (HBV) infection a major etiological factor. While microRNAs have been extensively studied in HCC, the role of PIWI-interacting RNAs (piRNAs) is at the beginning. Here, a comprehensive analysis was applied on small RNA sequencing data to compare piRNA expression in HBV-related HCC tumors, HBV-infected non-tumor liver, and normal liver tissue. RNA reads were aligned with Rsubread and quantified piRNAs using featureCounts, identifying differentially expressed piRNAs with DESeq2. In HBV-HCC versus normal liver 11 piRNAs found to be significantly dysregulated (5 up, 6 down), including a 9.9-fold upregulation of hsa-piR-32885 and 5.3-fold downregulation of hsa-piR-28734. Comparing HBV-HCC to HBV-infected liver revealed 17 differentially expressed piRNAs, with hsa-piR-432 most strongly upregulated. HBV-infected versus normal tissue showed 22 dysregulated piRNAs (21 down, 1 up), especially hsa-piR-432 was 21-fold down. These stage-specific changes suggest dynamic piRNA regulation during HBV-driven hepatocarcinogenesis. Many of the identified piRNAs are novel in the HCC context. Accordingly, hsa-piR-32885 has not been previously linked to liver cancer, though piR-Hep1 was found upregulated in ~47% of HCCs. The switch in hsa-piR-432 (down in HBV infection, up in HCC) highlights candidates for biomarkers or functional regulators. This study concludes that piRNA profiling in HBV-HCC uncovers novel markers and pathways. Future work will validate these piRNAs in larger cohorts and explore their regulatory targets. These findings support piRNAs as emerging players and biomarkers in HBV-associated HCC.
Keywords:
piRNA
; Hepatitis B virus
; Hepatocellular carcinoma
; biomarkers
; RNA sequencing
1. Introduction
Hepatocellular carcinoma (HCC) is among the most common and lethal cancers worldwide [1]. Chronic hepatitis B virus (HBV) infection is a predominant risk factor for HCC [2,3,4,5], yet early diagnosis remains challenging. Conventional biomarkers like alpha-fetoprotein lack sensitivity and specificity, underscoring the need for novel molecular markers [6]. In recent years, noncoding RNAs (ncRNAs) such as microRNAs and long noncoding RNAs have been implicated in HCC development [7,8,9]. Among small ncRNAs, PIWI-interacting RNAs (piRNAs) are emerging as important regulators of gene expression [10,11]. Originally characterized in germline cells for transposon silencing, piRNAs bind PIWI proteins and have been found in somatic tissues, where dysregulation is increasingly linked to cancer [10,12]. Indeed, a growing body of evidence indicates that piRNAs and PIWI proteins are abnormally expressed across diverse tumor types, acting in oncogenic or tumor suppressive pathways [13,14,15].
In HCC specifically, early studies have identified aberrant piRNA expression. Law et al. (2013) performed deep sequencing of small RNAs in HCC tissues and discovered a novel piRNA, dubbed piR-Hep1, which was upregulated in 46.6% of tumors [16]. Functional assays showed that silencing piR-Hep1 reduced hepatoma cell proliferation, motility and invasion, coincident with downregulation of phosphorylated AKT in the PI3K/AKT pathway [16]. Similarly, Liu et al. (2019) reviewed piRNAs in cancer and noted that the PIWI–piRNA complex can modulate signaling like PI3K/AKT in tumors [15]. Recently, comprehensive profiling has revealed many piRNAs in liver disease. Accordingly, Koduru et al. (2018) sequenced small RNAs in cirrhosis and HCC and reported 128 differentially expressed piRNAs in HCC (83 up, 45 down) versus normal liver [17]. Rizzo et al. (2016) performed piRNA profiling of dysplastic liver nodules and HCC, identifying 24 piRNAs with stage-specific expression patterns and suggesting their utility as biomarkers for distinguishing premalignant lesions [18]. Despite these advances, the roles of specific piRNAs in HBV-driven HCC remain largely unexplored.
In parallel, there is growing interest in piRNAs as minimally invasive biomarkers. Circulating and exosomal piRNAs have been detected in cancer patients. In this regard, Rui et al. (2023) identified 253 differentially expressed piRNAs in serum exosomes of HCC patients versus controls, with the top candidates robustly distinguishing tumors even at early stages [19]. These findings highlight the potential of piRNA signatures in blood for HCC diagnosis. In the context of viral infection, recent work indicates that piRNA/PIWI complexes can be dysregulated during viral hepatitis. Li et al. (2024) reviewed piRNA involvement in viral infections and noted that virus-induced changes in host piRNAs can influence disease progression and viral replication [20]. Such interactions suggest that HBV infection might imprint characteristic piRNA alterations in the liver, some of which could promote tumorigenesis.
Despite these indications, systematic profiling of piRNAs in HBV-associated HCC is lacking. Here, an RNA-seq-based analysis of piRNA expression was performed across normal liver, HBV-infected non-tumor liver, and HBV-positive HCC tissues. Using bioinformatic pipelines, including alignment and counting with Rsubread/featureCounts [21] and differential analysis with DESeq2 [22], it was aimed to identify piRNAs that are specifically dysregulated during HBV-driven hepatocarcinogenesis. Such piRNAs may clarify molecular mechanisms and serve as novel biomarkers. This study thus places HBV-HCC piRNA profiling within the current knowledge of ncRNAs in liver cancer, underscoring its novelty and clinical relevance. By integrating new sequencing data with existing literature, one of the first comprehensive piRNA landscape of HBV-related HCC was conducted and discuss its implications for biomarker discovery and cancer biology.
2. Materials and Methods
2.1. Sample and Data Acquisition
The dataset for this study was obtained from the Gene Expression Omnibus (GEO) repository under accession number GSE21279 [23]. This dataset comprises small RNA sequencing data from a cohort of patients with different liver conditions, including Normal Liver, HBV-infected Liver, and HBV-related HCC. The raw sequencing data, associated with Sequence Read Archive (SRA) project SRP002272, consisted of single-end sequencing runs. The metadata, including clinical characteristics and sample conditions, was retrieved from the GEO database.
2.2. piRNA Annotation and Reference Preparation
A human-specific piRNA database [24], piRNAdb.hsa.v1_7_6, was downloaded from the piRNA database (https://www.pirnadb.org/). To enable read counting with Rsubread, a custom Gene Transfer Format (GTF)-like file in Simplified Annotation Format (SAF) was created. This file contained GeneID, Chr, Start, End, and Strand. Each piRNA sequence from the FASTA file was treated as a single genomic feature, with its GeneID and Chr set to its unique identifier, and the Start and End positions defined by its sequence length. This SAF file served as the primary annotation for subsequent read quantification.
2.3. Sequence Alignment and Read Counting
The raw sequencing data in SRA format was first converted to FASTQ format using the fasterq-dump tool from the SRA Toolkit v3.2.1. To accurately quantify piRNA expression, a dedicated reference genome index was constructed. The piRNAdb.hsa.v1_7_6 was used as the reference to build a custom index using the Rsubread package's buildindex function. The processed FASTQ files were then aligned to this custom piRNA index using the align function from Rsubread with default parameters for RNA-seq data. This approach ensures that only reads mapping to known piRNA sequences are considered. Following alignment, the number of reads mapping to each piRNA was quantified using the featureCounts function with the previously generated SAF annotation file. The resulting output was a raw count matrix where rows represent piRNAs and columns represent samples.
2.4. Differential Expression Analysis
The raw count matrix and the sanitized metadata were used as input for differential expression analysis (DEA) using the DESeq2 package v1.38.3. To ensure robust statistical analysis, samples corresponding to patient groups with a sample size of less than two (HCV_HCC and Non_Viral_HCC) were excluded from the analysis. The raw counts were rounded to integers, and a DESeqDataSet object was constructed with the design formula.
The standard DESeq2 workflow was then applied, including, size factor estimation to normalize for sequencing depth, dispersion estimation to model biological and technical variability, and generalized linear model (GLM) fitting and Wald's statistical test to identify differentially expressed piRNAs. Differential expression was determined by comparing the following groups: HBV-related HCC versus Normal, HBV-related HCC versus HBV-infected, and HBV-infected versus Normal. piRNAs with an absolute log2-fold change greater than 1 and an adjusted p-value (padj) less than 0.05 were considered statistically significant.
2.5. Data Visualization
To visualize the findings, high-quality plots were generated using the ggplot2 and pheatmap packages. Volcano plots were created for each comparison to simultaneously display the log2-fold change and the adjusted p-value. The plots were color-coded to highlight up-regulated and down-regulated piRNAs, with labels for the top 10 most significant ones. For these plots, the normalized counts were used, and samples were manually ordered by their clinical condition to improve interpretability. The heatmap were scaled by row and included hierarchical clustering on piRNAs to group those with similar expression patterns.
3. Results
3.1. Overview of Differentially Expressed piRNAs
Differential expression analysis identified a total of 11, 17, and 22 statistically significant piRNAs in the HBV-related HCC vs. Normal, HBV-related HCC vs. HBV-infected, and HBV-infected vs. Normal comparisons, respectively. 31 significantly down- or up-regulated piRNAs were highlighted (Figure 1). A comprehensive summary of the findings, including log2-fold change and adjusted p-values, is detailed in the sections below. All raw data regarding to the count of piRNAs mapped to the respective SRAs is provided in Additional file 1.
3.2. HBV-Related HCC vs. Normal Liver Comparison
In the comparison of HBV-related HCC and Normal liver samples, 11 differentially expressed piRNAs identified (Figure 2). Among these, 5 were significantly up-regulated and 6 were significantly down-regulated. The most up-regulated piRNA was hsa-piR-32885, with a log2-fold change of 9.91, while the most down-regulated was hsa-piR-28734, with a log2-fold change of -5.32. A summary of the top differentially expressed piRNAs for this comparison is provided in Table 1.
3.3. HBV-Related HCC vs. HBV-Infected Liver Comparison
A total of 17 piRNAs were found to be significantly differentially expressed when comparing HBV-related HCC and HBV-infected samples (Figure 3). The analysis revealed 7 up-regulated and 10 down-regulated piRNAs. hsa-piR-432 was the most up-regulated piRNA, with an exceptionally high log2-fold change of 20.14. Conversely, the most down-regulated piRNA was hsa-piR-33001, exhibiting a log2-fold change of -12.75. Especially, hsa-piR-5969, hsa-piR-32993, and hsa-piR-28734 were identified as differentially expressed in both the HCC vs. Normal and HCC vs. HBV-infected comparisons, suggesting their potential role in the progression of liver disease. Table 2 provides a detailed list of these findings.
3.4. HBV-Infected vs. Normal Liver Comparison
This comparison yielded 22 differentially expressed piRNAs, with 19 being down-regulated and 3 being up-regulated. The most significantly down-regulated piRNA was hsa-piR-432, with a log2-fold change of -21.28 (Figure 4). This is a particularly important finding, as hsa-piR-432 was also identified as the most up-regulated piRNA in the HCC vs. HBV-infected comparison. The most up-regulated piRNA in this group was hsa-piR-1900, with a log2-fold change of 3.13. A detailed summary of these findings is presented in Table 3.
4. Discussion
This study aimed to profile the most recent annotated human piRNAs in instances of HBV-related HCC for the first time, to the author's knowledge. The RNA-seq profiling revealed a distinct piRNA landscape in HBV-related HCC. In HCC versus normal liver, 11 significantly dysregulated piRNAs were identified (5 upregulated, 6 downregulated). This is fewer than the 128 dysregulated piRNAs reported by Koduru et al. [17], likely reflecting differences in sample size, filtering stringency, and data origin. Nonetheless, the results of the present study highlight key candidates. The most upregulated in HBV-HCC was hsa-piR-32885 with 9.9-fold increase, and the most downregulated was hsa-piR-28734 with 5.3-fold decrease relative to normal. Particularly, hsa-piR-32885 has not been previously implicated in liver cancer; piRNAdb lists it from lung tissue with targets including MIR182 and several zinc-finger genes. By contrast, the well-studied piR-Hep1 was not detected in the dataset, perhaps due to differences in patient cohorts or sequencing depth. However, the pattern of findings, with specific piRNAs elevated in tumors, is consistent with Law et al. showing piR-Hep1 upregulation in HCC [16]. Functional parallels may exist. for instance, piR-Hep1 promotes HCC cell growth via AKT phosphorylation. If hsa-piR-32885 similarly modulates oncogenic pathways by repressing tumor-suppressive miRNAs like MIR182, it could represent a novel driver of hepatocarcinogenesis.
Conversely, downregulated piRNAs in HCC may normally act as tumor suppressors. hsa-piR-28734, strongly depleted in HCC, could be of this type. Its predicted length (~31 nt) and sequence are unique, and to the author’s knowledge it has not been studied in cancer. The loss of such piRNAs might release repression of oncogenes or impair genomic stability. General loss of piRNA silencing may activate transposons or dysregulate chromatin, as piRNAs are known to protect genome integrity [10,14]. The dysregulation of particular piRNAs in HBV-HCC thus suggests new biological roles. In this regard, hsa-piR-32885 may promote oncogenic signaling, similar to piR-Hep1 [15], while hsa-piR-28734 and others may normally suppress malignancy and are suppressed themselves during tumorigenesis.
Comparing HBV-HCC to HBV-infected non-tumor liver, 17 dysregulated piRNAs were found (9 up in HCC, 8 down). Importantly, hsa-piR-32885 and hsa-piR-28734 again showed similar changes, reinforcing their HCC association. Critically, hsa-piR-432 emerged as the most upregulated (20-fold) in HCC versus infected liver. Conversely, in HBV-infected versus normal tissue, hsa-piR-432 was the most downregulated (~21-fold). This bidirectional change suggests that hsa-piR-432 may repressed by HBV infection but then dramatically induced during malignant transformation. The nature of this switch is intriguing. It may reflect virus-host interactions. Recent studies report that viral infections alter piRNA expression [20], and HBV integration or HBx protein could suppress certain piRNAs in chronic infection. Upon oncogenic progression, tumor cells might re-express hsa-piR-432, perhaps to cope with HBV-induced genomic stress or to activate proliferation pathways. The dramatic fold-changes also make hsa-piR-432 an attractive biomarker candidate. Accordingly, its absence in infected liver but abundance in HBV-HCC could distinguish malignant from pre-malignant disease. To the author’s knowledge, hsa-piR-432 has not been reported in cancer before; its sequence, ACAGTAGCATTGGTGGTTCAGTGGTA, is correspond to piR-hsa-624 in piRBase, but functional data is lacking. Follow-up functional studies will be essential to understand how hsa-piR-432 might influence hepatocyte biology or viral interactions.
For the HBV-infected versus normal comparison, 22 differentially expressed piRNAs were observed, mostly downregulated. This indicates that HBV infection alone perturbs the hepatic piRNome. Such broad downregulation could mirror the virus’s impact on host epigenetic regulators. It was noted that viral infection can disrupt host piRNA/PIWI expression [20]. Chronic HBV may suppress piRNAs that normally maintain genome stability, potentially facilitating integration or oncogenic mutations. In this regard, hsa-piR-1900 was modestly upregulated in infected liver versus normal, while many others were lost. The loss of piRNAs during infection might prime the liver for malignant transformation, consistent with HBV’s carcinogenic role. These changes complement reports in other systems. For instance, influenza and SARS-CoV-2 viruses have been shown to interact with host piRNAs [25,26,27,28]. The data of the present study suggest a similar phenomenon in HBV-HCC, although the exact mechanisms remain to be elucidated.
In contextualizing the findings, it is noted that piRNA profiling in HCC is an emerging field. Previous reports have similarly highlighted stage-specific piRNA expression. It was found that piRNAs were specifically expressed in dysplastic versus neoplastic liver lesions and proposed these as diagnostic markers [18]. Here, it was also observed that stage-specific piRNAs like hsa-piR-432 that may serve analogous roles. Moreover, it was demonstrated that serum exosomal piRNAs strongly discriminate HCC from non-tumor, implying clinical utility [19]. It will be important to test whether any of the identified piRNAs such as hsa-piR-432, hsa-piR-32885, or hsa-piR-28734 are detectable in blood and predictive of HBV-HCC. The overlap between tissue and circulating piRNAs remains largely unexplored.
Technically, the presented analysis employed current best practices for small RNA-seq. Accordingly, reads were aligned to a custom piRNA reference and counted with featureCounts, followed by DESeq2 normalization and testing. Specialized pipelines like piPipes have been developed for piRNA analysis [29], though the presented approach was effective in quantifying known piRNAs. The use of the piRNAdb catalog ensured capture of both annotated and novel piRNAs. Here, it was also validated that sample clustering separated tumor, infected, and normal groups based on piRNA expression, supporting the biological relevance of the identified differences.
Biologically, the key question is how these piRNAs influence hepatocarcinogenesis. Emerging models suggest piRNAs can regulate gene networks via mRNA degradation or epigenetic modification [10]. In this regard, piRNAs in germline silence transposons, but in somatic cells they may target oncogenic or tumor-suppressor transcripts. In HCC, upregulated piRNAs like piR-Hep1 appear to activate growth pathways. It is plausible that hsa-piR-32885 or hsa-piR-432 target genes involved in proliferation or apoptosis; this could be tested by target prediction and reporter assays. The downregulated piRNAs like hsa-piR-28734 might normally repress pro-metastatic genes. Indeed, one piRNA, piR1/97, was previously identified that promoted migration of HCC cell lines [30], showing that individual piRNAs can drive malignancy. The differentially expressed piRNAs, reported here, warrant similar functional studies. Additionally, PIWI proteins themselves (PIWIL1-4) have been noted as prognostic markers in HCC; for instance, nuclear PIWIL2/4 co-expression portends poor outcome [31]. The relationship between PIWI protein levels and the presented piRNA signatures will be of interest.
In summary, the findings extend the current understanding of the piRNA/PIWI axis in liver cancer. They corroborate the concept that distinct piRNA signatures mark different disease stages and may regulate HCC progression. Importantly, the identified piRNAs, highlighting hsa-piR-32885, hsa-piR-432, and hsa-piR-28734 are largely novel in the context of liver disease and represent testable biomarkers or therapeutic targets. The results suggest a model in which HBV infection initially alters host piRNAs, and subsequent malignant transformation leads to additional changes that promote tumorigenesis. Verification of these piRNAs in independent HBV-HCC cohorts and in circulation is a key next step.
5. Conclusions
In this study, one of the first comparative profiling of piRNAs in human liver across normal, HBV-infected non-tumor, and HBV-associated HCC tissues was performed. The analyses uncovered piRNAs whose expression differs with disease stage, revealing a dynamic piRNA landscape in HBV-driven hepatocarcinogenesis. The findings include the strong upregulation of hsa-piR-32885 in HCC and downregulation of hsa-piR-28734, neither previously reported in HCC, as well as the switch of hsa-piR-432, down in infected liver and up in HCC. These stage-specific piRNAs point to potentially distinct roles. hsa-piR-32885 may act as an oncogenic piRNA analogous to piR-Hep1, while loss of hsa-piR-28734 suggests a tumor-suppressive function. The fold-changes of hsa-piR-432 imply its involvement in the transition from chronic HBV infection to malignancy. Overall, The results indicate that chronic HBV infection broadly suppresses many piRNAs, reflected by the 22 piRNAs downregulated in infected versus normal liver, and that malignant transformation induces a distinct set of changes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Name: Additional file 1. Description: Raw piRNA counts mapped to the SRA data. File: .csv.
Author Contributions
Conceptualization: A.M.; Data curation: A.M.; Formal analysis: A.M.; Funding acquisition: A.M.; Investigation: A.M.; Methodology: A.M.; Project administration: A.M.; Resources: A.M.; Software: A.M.; Supervision: A.M.; Validation: A.M.; Visualization: A.M.; Writing – original draft: A.M.; Writing – review and editing: A.M.
Funding
Not applicable.
Declaration of Generative AI Use
During the preparation of this work the author used GPT 5 in order to improve the manuscript’s sentence wording and English grammar. After using this tool/service, the author reviewed and edited the content as needed and takes full responsibility for the content of the published article.
Informed Consent and Patient Details
Not applicable.
Clinical Trials
Not applicable.
Data Statement
Data supporting this study are included within the article and supporting material.
Declaration of Competing Interests
I have nothing to declare.
References
- Li, Q.; Ding, C.; Cao, M.; Yang, F.; Yan, X.; He, S.; Cao, M.; Zhang, S.; Teng, Y.; Tan, N.; et al. Global epidemiology of liver cancer 2022: An emphasis on geographic disparities. Chin. Med J. 2024, 137, 2334–2342. [Google Scholar] [CrossRef]
- Mohebbi, A.; Lorestani, N.; Tahamtan, A.; Kargar, N.L.; Tabarraei, A. An Overview of Hepatitis B Virus Surface Antigen Secretion Inhibitors. Front. Microbiol. 2018, 9, 662. [Google Scholar] [CrossRef]
- Naderi, M.; Salavatiha, Z.; Gogoi, U.; Mohebbi, A. An overview of anti-Hepatitis B virus flavonoids and their mechanisms of action. Front. Cell. Infect. Microbiol. 2024, 14, 1356003. [Google Scholar] [CrossRef]
- Gonzalez, S. ; A, S. Novel biomarkers for hepatocellular carcinoma surveillance: has the future arrived? Hepatobiliary Surg Nutr 2014, 3, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Xiang, X.; Feng, B.; Zhou, H.; Wang, T.; Chu, X.; Wang, R. Targeting Long Non-Coding RNAs in Hepatocellular Carcinoma: Progress and Prospects. Front. Oncol. 2021, 11. [Google Scholar] [CrossRef]
- Tian, Y.; Zhang, M.; Liu, L.-X.; Wang, Z.-C.; Liu, B.; Huang, Y.; Wang, X.; Ling, Y.-Z.; Wang, F.; Feng, X.; et al. Exploring non-coding RNA mechanisms in hepatocellular carcinoma: implications for therapy and prognosis. Front. Immunol. 2024, 15, 1400744. [Google Scholar] [CrossRef]
- Zhou, H.; Xu, Q.; Ni, C.; Ye, S.; Xu, X.; Hu, X.; Jiang, J.; Hong, Y.; Huang, D.; Yang, L. Prospects of Noncoding RNAs in Hepatocellular Carcinoma. BioMed Res. Int. 2018, 2018, 1–9. [Google Scholar] [CrossRef]
- Mei, Y.; Clark, D.; Mao, L. Novel dimensions of piRNAs in cancer. Cancer Lett. 2013, 336, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yang, X.; Zhou, Q.; Zhuang, J.; Han, S. Biological significance of piRNA in liver cancer: a review. Biomarkers 2020, 25, 436–440. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Wang, Q.; Jiang, W.; Bian, Y.; Zhou, Y.; Gou, A.; Zhang, W.; Fu, K.; Shi, W. Emerging roles of piRNAs in cancer: challenges and prospects. Aging 2019, 11, 9932–9946. [Google Scholar] [CrossRef] [PubMed]
- Cai, A.; Hu, Y.; Zhou, Z.; Qi, Q.; Wu, Y.; Dong, P.; Chen, L.; Wang, F. PIWI-Interacting RNAs (piRNAs): Promising Applications as Emerging Biomarkers for Digestive System Cancer. Front. Mol. Biosci. 2022, 9, 848105. [Google Scholar] [CrossRef]
- Garcia-Borja, E.; Siegl, F.; Mateu, R.; Slaby, O.; Sedo, A.; Busek, P.; Sana, J. Critical appraisal of the piRNA-PIWI axis in cancer and cancer stem cells. Biomark. Res. 2024, 12, 1–24. [Google Scholar] [CrossRef]
- Liu, Y.; Dou, M.; Song, X.; Dong, Y.; Liu, S.; Liu, H.; Tao, J.; Li, W.; Yin, X.; Xu, W. The emerging role of the piRNA/piwi complex in cancer. Mol. Cancer 2019, 18, 1–15. [Google Scholar] [CrossRef]
- Law, P.T.-Y.; Qin, H.; Ching, A.K.-K.; Lai, K.P.; Na Co, N.; He, M.; Lung, R.W.-M.; Chan, A.W.-H.; Chan, T.-F.; Wong, N. Deep sequencing of small RNA transcriptome reveals novel non-coding RNAs in hepatocellular carcinoma. J. Hepatol. 2013, 58, 1165–1173. [Google Scholar] [CrossRef]
- Koduru, S.V.; Leberfinger, A.N.; Kawasawa, Y.I.; Mahajan, M.; Gusani, N.J.; Sanyal, A.J.; Ravnic, D.J. Non-coding RNAs in Various Stages of Liver Disease Leading to Hepatocellular Carcinoma: Differential Expression of miRNAs, piRNAs, lncRNAs, circRNAs, and sno/mt-RNAs. Sci. Rep. 2018, 8, 1–48. [Google Scholar] [CrossRef]
- Rizzo, F.; Rinaldi, A.; Marchese, G.; Coviello, E.; Sellitto, A.; Cordella, A.; Giurato, G.; Nassa, G.; Ravo, M.; Tarallo, R.; et al. Specific patterns of PIWI-interacting small noncoding RNA expression in dysplastic liver nodules and hepatocellular carcinoma. Oncotarget 2016, 7, 54650–54661. [Google Scholar] [CrossRef] [PubMed]
- Rui, T.; Wang, K.; Xiang, A.; Guo, J.; Tang, N.; Jin, X.; Lin, Y.; Liu, J.; Zhang, X. Serum Exosome-Derived piRNAs Could Be Promising Biomarkers for HCC Diagnosis. Int. J. Nanomed. 2023, ume 18, 1989–2001. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, K.; Liu, W.; Zhang, Y. The potential emerging role of piRNA/PIWI complex in virus infection. Virus Genes 2024, 60, 333–346. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. feature Counts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- M. I. Love, W. Huber, S. Anders, Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol 2014, 15, 1–21. [Google Scholar] [CrossRef]
- Hou, J.; Lin, L.; Zhou, W.; Wang, Z.; Ding, G.; Dong, Q.; Qin, L.; Wu, X.; Zheng, Y.; Yang, Y.; et al. Identification of miRNomes in Human Liver and Hepatocellular Carcinoma Reveals miR-199a/b-3p as Therapeutic Target for Hepatocellular Carcinoma. Cancer Cell 2011, 19, 232–243. [Google Scholar] [CrossRef]
- R. Piuco, P.A.F. R. Piuco, P.A.F. Galante, piRNAdb: A piwi-interacting RNA database, (n.d.). [CrossRef]
- Kondratov, K.A.; Artamonov, A.A.; Nikitin, Y.V.; Velmiskina, A.A.; Mikhailovskii, V.Y.; Mosenko, S.V.; Polkovnikova, I.A.; Asinovskaya, A.Y.; Apalko, S.V.; Sushentseva, N.N.; et al. Revealing differential expression patterns of piRNA in FACS blood cells of SARS-CoV−2 infected patients. BMC Med Genom. 2024, 17, 1–11. [Google Scholar] [CrossRef]
- Akimniyazova, A.; Yurikova, O.; Pyrkova, A.; Rakhmetullina, A.; Niyazova, T.; Ryskulova, A.-G.; Ivashchenko, A. In Silico Study of piRNA Interactions with the SARS-CoV-2 Genome. Int. J. Mol. Sci. 2022, 23, 9919. [Google Scholar] [CrossRef]
- Rakhmetullina, A.; Akimniyazova, A.; Niyazova, T.; Pyrkova, A.; Kamenova, S.; Kondybayeva, A.; Ryskulova, A.-G.; Ivashchenko, A.; Zielenkiewicz, P. Endogenous piRNAs Can Interact with the Omicron Variant of the SARS-CoV-2 Genome. Curr. Issues Mol. Biol. 2023, 45, 2950–2964. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Gu, W. Small RNA Plays Important Roles in Virus–Host Interactions. Viruses 2020, 12, 1271. [Google Scholar] [CrossRef] [PubMed]
- Han, B.W.; Wang, W.; Zamore, P.D.; Weng, Z. piPipes: a set of pipelines for piRNA and transposon analysis via small RNA-seq, RNA-seq, degradome- and CAGE-seq, ChIP-seq and genomic DNA sequencing. Bioinformatics 2014, 31, 593–595. [Google Scholar] [CrossRef]
- Miao, P.Z.; Yang, Y.; Chen, E.B.; Zhu, G.Q.; Wang, B.; Dai, Z. [Differential expressions analysis of piwi-interacting RNAs in hepatocellular carcinoma]. . 2018, 26, 842–846. [Google Scholar] [PubMed]
- Zeng, G.; Zhang, D.; Liu, X.; Kang, Q.; Fu, Y.; Tang, B.; Guo, W.; Zhang, Y.; Wei, G.; He, D. Co-expression of Piwil2/Piwil4 in nucleus indicates poor prognosis of hepatocellular carcinoma. Oncotarget 2016, 8, 4607–4617. [Google Scholar] [CrossRef]
Figure 1.
Comprehensive Heatmap of All Significantly Expressed piRNAs. Heatmap showing the row-scaled normalized expression of all unique, significant piRNAs across all samples. The samples are manually ordered and grouped by their condition.
Figure 1.
Comprehensive Heatmap of All Significantly Expressed piRNAs. Heatmap showing the row-scaled normalized expression of all unique, significant piRNAs across all samples. The samples are manually ordered and grouped by their condition.
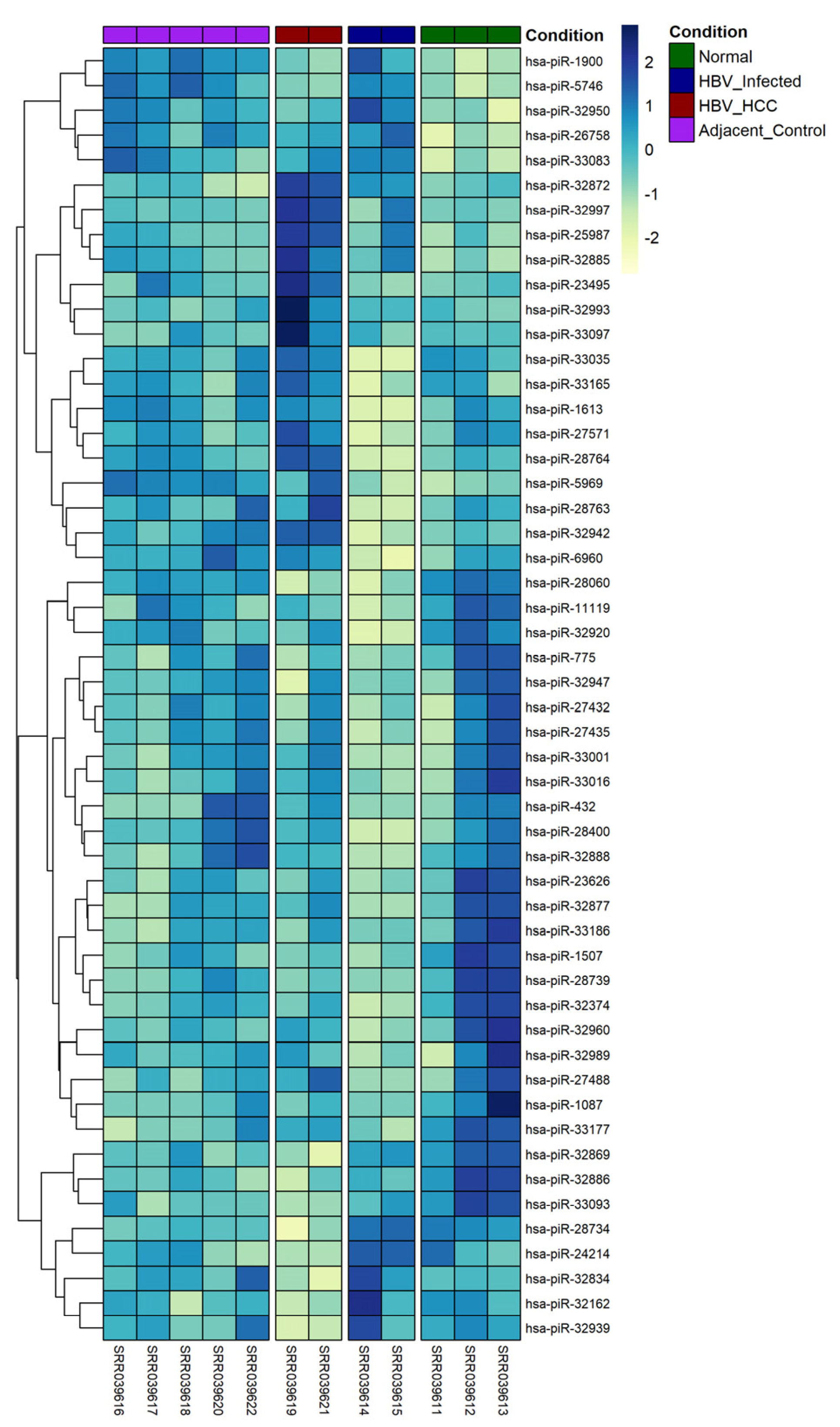
Figure 2.
Volcano Plot for HBV-related HCC vs. Normal Liver. A scatter plot visualizing differential piRNA expression between HBV-related HCC and Normal liver samples. Red dots indicate significantly up-regulated piRNAs, blue dots indicate significantly down-regulated piRNAs, and grey dots represent piRNAs that are not significant. The horizontal dashed line marks the significance threshold of padj = 0.05, and the vertical dashed lines indicate a log2-fold change of ±1. The top 10 most significant piRNAs are labeled.
Figure 2.
Volcano Plot for HBV-related HCC vs. Normal Liver. A scatter plot visualizing differential piRNA expression between HBV-related HCC and Normal liver samples. Red dots indicate significantly up-regulated piRNAs, blue dots indicate significantly down-regulated piRNAs, and grey dots represent piRNAs that are not significant. The horizontal dashed line marks the significance threshold of padj = 0.05, and the vertical dashed lines indicate a log2-fold change of ±1. The top 10 most significant piRNAs are labeled.
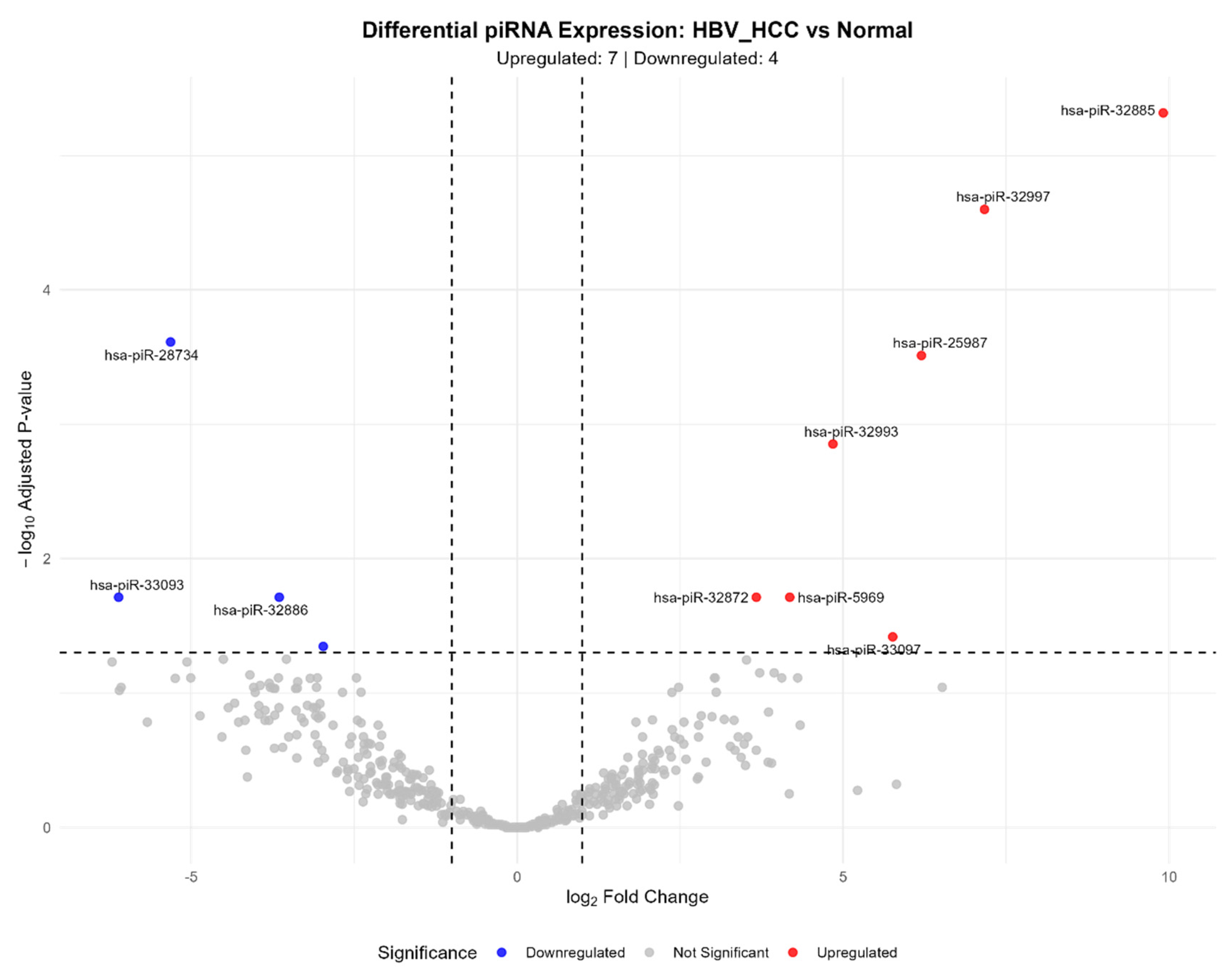
Figure 3.
Volcano Plot for HBV-related HCC vs. HBV-infected Liver. A scatter plot visualizing differential piRNA expression between HBV-related HCC and HBV-infected liver samples. Each dot corresponds to a piRNA. Red dots indicate significantly up-regulated piRNAs, blue dots indicate significantly down-regulated piRNAs, and grey dots represent piRNAs that are not significant. The horizontal dashed line marks the significance threshold of padj = 0.05, and the vertical dashed lines indicate a log2-fold change of ±1. The top 10 most significant piRNAs are labeled.
Figure 3.
Volcano Plot for HBV-related HCC vs. HBV-infected Liver. A scatter plot visualizing differential piRNA expression between HBV-related HCC and HBV-infected liver samples. Each dot corresponds to a piRNA. Red dots indicate significantly up-regulated piRNAs, blue dots indicate significantly down-regulated piRNAs, and grey dots represent piRNAs that are not significant. The horizontal dashed line marks the significance threshold of padj = 0.05, and the vertical dashed lines indicate a log2-fold change of ±1. The top 10 most significant piRNAs are labeled.
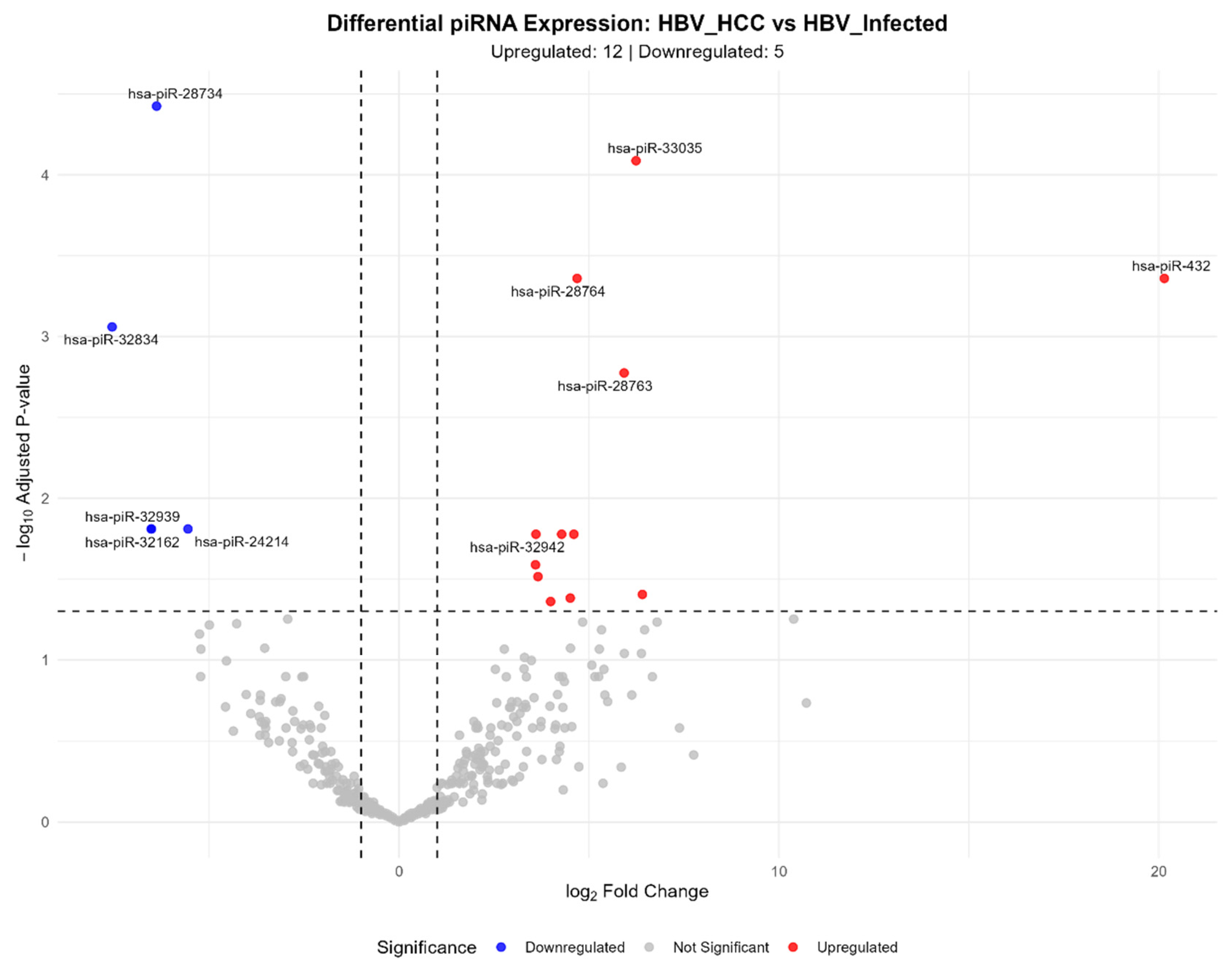
Figure 4.
Volcano Plot for HBV-infected vs. Normal Liver. A scatter plot visualizing differential piRNA expression between HBV-infected and Normal liver samples. Red dots indicate significantly up-regulated piRNAs, blue dots indicate significantly down-regulated piRNAs, and grey dots represent piRNAs that are not significant. The horizontal dashed line marks the significance threshold of padj = 0.05, and the vertical dashed lines indicate a log2-fold change of ±1. The top 10 most significant piRNAs are labeled.
Figure 4.
Volcano Plot for HBV-infected vs. Normal Liver. A scatter plot visualizing differential piRNA expression between HBV-infected and Normal liver samples. Red dots indicate significantly up-regulated piRNAs, blue dots indicate significantly down-regulated piRNAs, and grey dots represent piRNAs that are not significant. The horizontal dashed line marks the significance threshold of padj = 0.05, and the vertical dashed lines indicate a log2-fold change of ±1. The top 10 most significant piRNAs are labeled.

Table 1.
Top differentially expressed piRNAs in HBV-related HCC vs. Normal Liver.
| piRNA_ID | baseMean | log2FoldChange | p-value | padj | Sequence |
| hsa-piR-32885 | 247.62 | 9.91 | 1.11e-08 | 4.81e-06 | CACCAGTGTGAGTTCTACCATTGCCAAA |
| hsa-piR-32997 | 101.21 | 7.17 | 1.16e-07 | 2.51e-05 | TCACAGTGAATTCTACCAGTGCCATA |
| hsa-piR-25987 | 138.90 | 6.20 | 2.85e-06 | 3.08e-04 | TTTGGCAATGGTAGAACTCACACTGGTGAGGT |
| hsa-piR-32993 | 284.37 | 4.84 | 1.62e-05 | 1.40e-03 | TAGAGGAGCCTGTTCTGTAATCGATAAACC |
| hsa-piR-5969 | 57.80 | 4.18 | 4.04e-04 | 1.94e-02 | TCCGTAGTGTAGTGGTTATCACGTTCGCCT |
| hsa-piR-32872 | 109.14 | 3.67 | 3.69e-04 | 1.94e-02 | AGTCTCAGTTTCCTCTGCAAACAGTT |
| hsa-piR-33097 | 11.79 | 5.76 | 8.85e-04 | 3.82e-02 | CAGCTGATGATGATAATATTGCCTGAAGA |
| hsa-piR-32869 | 1458.61 | -2.97 | 1.15e-03 | 4.51e-02 | AGGGAGATGAAGAGGACAGTGACTGAGAGAC |
| hsa-piR-32886 | 2045.14 | -3.65 | 3.51e-04 | 1.94e-02 | CACCATGATGGAACTGAGGATCTGAGGAA |
| hsa-piR-28734 | 198.73 | -5.32 | 1.69e-06 | 2.44e-04 | GTTCACTGATGAGAGCATTGTTCTGAGCCA |
| hsa-piR-33093 | 9.71 | -6.11 | 3.69e-04 | 1.94e-02 | CAGCAAATGATGTGAGAGATTCTGCTGATA |
Table 2.
Top differentially expressed piRNAs in HBV-related HCC vs. HBV-infected Liver.
| piRNA_ID | baseMean | log2FoldChange | p-value | padj | Sequence |
| hsa-piR-432 | 58.19 | 20.14 | 4.22e-06 | 4.36e-04 | ACAGTAGCATTGGTGGTTCAGTGGTA |
| hsa-piR-1613 | 11.85 | 6.40 | 1.52e-03 | 3.94e-02 | ATAGGTTTGGTCCTAGCCTTTCTATT |
| hsa-piR-33035 | 123.76 | 6.24 | 4.18e-07 | 8.16e-05 | TGGTTCGTCCAAGTGCACTTTCCAGT |
| hsa-piR-28763 | 778.02 | 5.92 | 2.58e-05 | 1.68e-03 | GTTTAGACGGGCTCACATCACCCCATAAACA |
| hsa-piR-28764 | 718.99 | 4.68 | 4.48e-06 | 4.36e-04 | GTTTCCGTAGTGTAGTGGTCATCACGTTCGC |
| hsa-piR-23495 | 15.51 | 4.51 | 1.70e-03 | 4.15e-02 | CGTAGTGTAGTGGTCATCACGTTCGCCT |
| hsa-piR-32993 | 284.37 | 4.28 | 4.98e-04 | 1.67e-02 | TAGAGGAGCCTGTTCTGTAATCGATAAACC |
| hsa-piR-5969 | 57.80 | 4.60 | 5.09e-04 | 1.67e-02 | TCCGTAGTGTAGTGGTTATCACGTTCGCCT |
| hsa-piR-32885 | 247.62 | 5.70 | 1.04e-03 | 2.03e-02 | CACCAGTGTGAGTTCTACCATTGCCAAA |
| hsa-piR-28734 | 198.73 | -6.38 | 9.62e-08 | 3.75e-05 | GTTCACTGATGAGAGCATTGTTCTGAGCCA |
| hsa-piR-32162 | 19.55 | -6.52 | 2.88e-04 | 1.55e-02 | CACAATGCTGACACTCAAACTGCTGACA |
Table 3.
Top differentially expressed piRNAs in HBV-infected vs. Normal Liver.
| piRNA_ID | baseMean | log2FoldChange | p-value | padj | Sequence |
| hsa-piR-1900 | 1399.71 | 3.13 | 8.22e-04 | 1.78e-02 | ATTGGTGGTTCAGTGGTAGAATTCTCGCC |
| hsa-piR-26758 | 837.01 | 3.01 | 1.78e-03 | 3.16e-02 | GAGGAATGATGACAAGAAAAGGCCGAA |
| hsa-piR-5746 | 669.93 | 2.98 | 2.10e-03 | 3.42e-02 | TCCCTGGTGGTCTAGTGGTTAGGATA |
| hsa-piR-432 | 58.19 | -21.28 | 1.29e-07 | 5.02e-05 | ACAGTAGCATTGGTGGTTCAGTGGTA |
| hsa-piR-33001 | 428.37 | -12.75 | 7.06e-05 | 3.06e-03 | TCATGCTCTATCGACTGAGCTAGCCGGG |
| hsa-piR-32374 | 117.15 | -9.66 | 1.46e-06 | 2.86e-04 | GTCGATGATGATTGGTAAAAGGTCTGA |
| hsa-piR-32877 | 41.49 | -9.46 | 1.22e-05 | 1.19e-03 | ATCAATGATGAAACTAGCCAAATCTGAGC |
| hsa-piR-32888 | 38.81 | -7.85 | 4.20e-04 | 1.09e-02 | CAGAATGCTAACCATTACACGATGGAACC |
| hsa-piR-1087 | 9.69 | -7.55 | 1.45e-03 | 2.69e-02 | AGCCTATGATGGTTAGTTATCCCTGTCTGAAA |
| hsa-piR-27435 | 109176.10 | -7.41 | 2.73e-05 | 1.68e-03 | GCCCGGCTAGCTCAGTCGGTAGAGCATGAGAC |
| hsa-piR-32947 | 249.02 | -5.60 | 7.00e-04 | 1.71e-02 | GCTCAGAAAATACCTTTCAGTCACACATT |
| hsa-piR-32960 | 43.45 | -6.11 | 2.89e-05 | 1.68e-03 | GGTCAATGATGAATGGTAAAAGGTCTGAGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



