Submitted:
14 October 2025
Posted:
14 October 2025
You are already at the latest version
Abstract
We investigated how tannin supplementation influences the behavior and performance of beef cattle grazing a meadow bromegrass monoculture. Twenty-four Angus cow–calf pairs were allocated to six 3.6-ha paddocks (four pairs/paddock), randomly assigned to Control (Ctrl; n = 3) or Tannin treatment (TT; n = 3). Animals received 1 kg/cow/day of DDGs, with TT receiving an added 0.4% tannins (2:1 condensed:hydrolyzable). Grazing occurred during four 15-day periods (July–September) across two years. Data were analyzed with mixed-effects models. Tannins did not alter biomass removal or cow weight loss (p > 0.05). However, TT cows exhibited longer evening grazing (2.9 vs. 2.1 h), fewer standing-to-lying transitions (5.7% vs. 7.3%), and more even grazing distribution (CV = 1.86 vs. 2.13; p < 0.05), and greater water consumption (147 vs. 121 L/day; p < 0.01). Average daily gain of calves was numerically greater in TT compared to Ctrl (1.03 vs. 0.93 kg/day), although this difference was not significant (p = 0.2699). Findings indicate that tannin supplementation extended evening grazing, reduced rumination, and promoted even distribution, indicative of increased forage selectivity. Fewer standing-to-lying transitions indicated reduced restlessness, potentially improving welfare. Increased water intake highlights a physiological adjustment that may require management in water-limited rangelands.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Animal and Grazing Protocol
2.3. Measurements of the Impact of Tannin Supplementation
2.3.1. Herbage Availability
2.3.2. Foraging Behavior
2.3.3. Grazing Distribution
2.3.4. Activity Level Estimates
2.3.5. Water Consumption Estimates
2.3.6. Animal Performance
2.4. Chemical Analyses
2.5. Statistical Analysis
3. Results
3.1. Herbage Availability, Water Consumption, and Animal Performance
3.2. Nutritional Composition
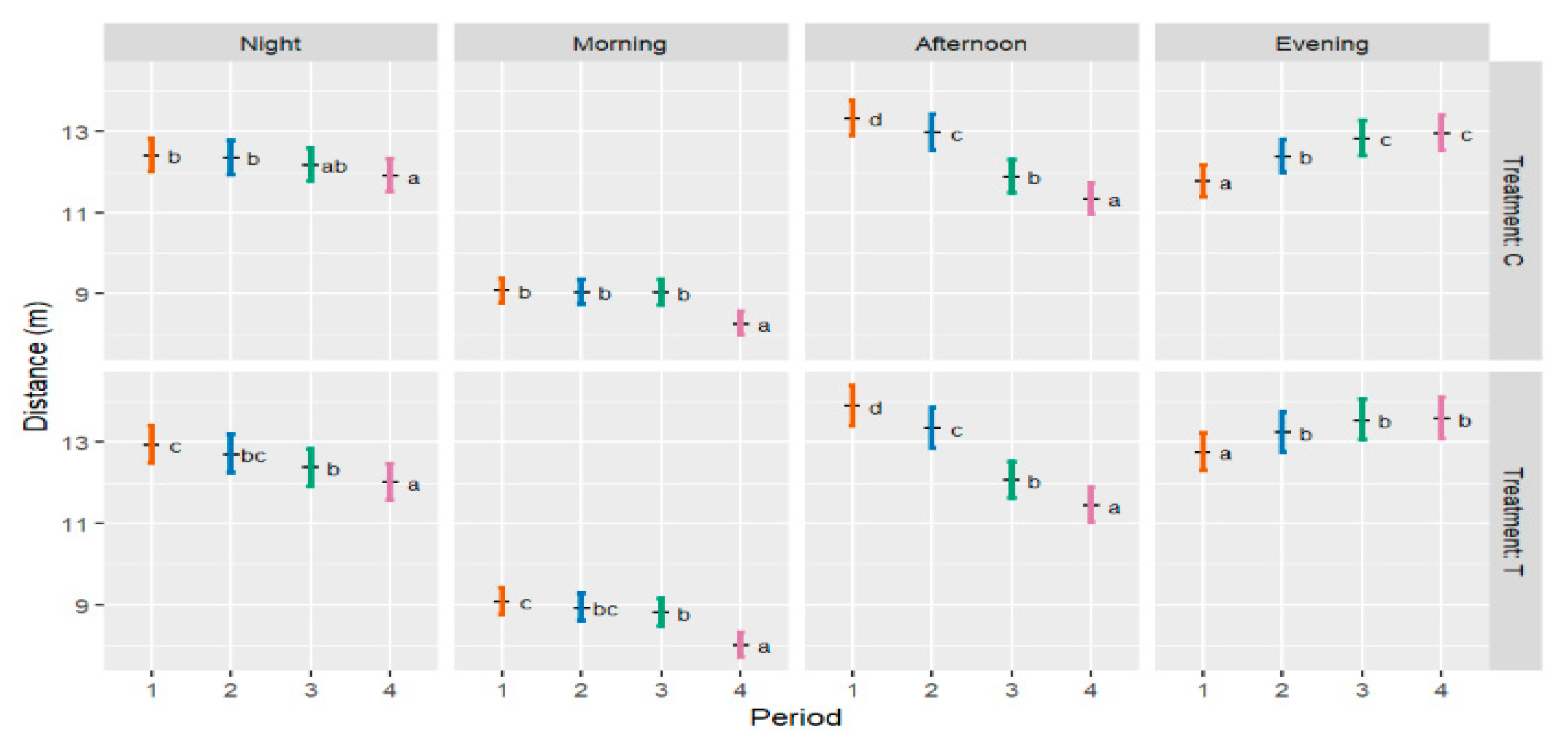
3.3. Distance Travelled
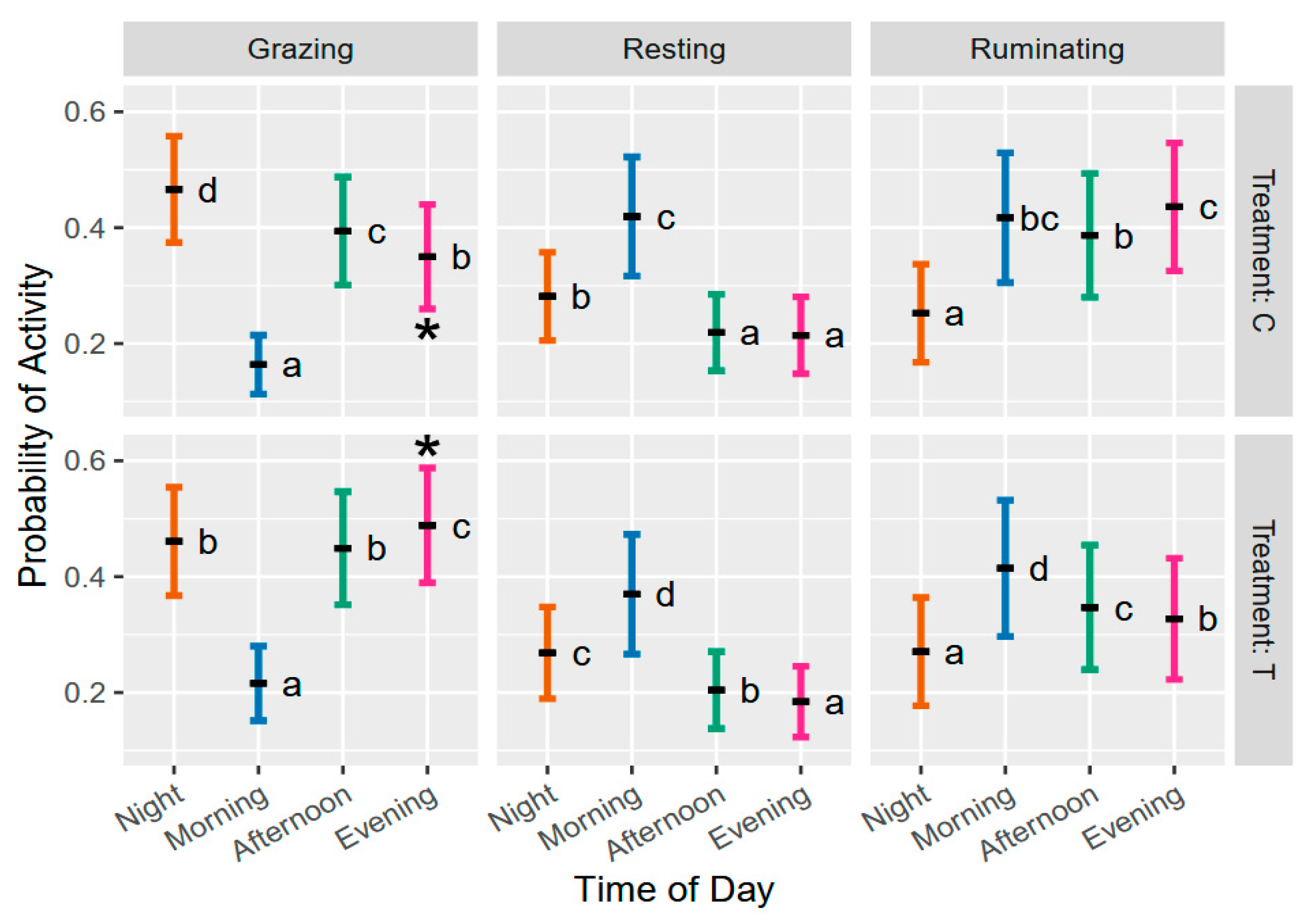
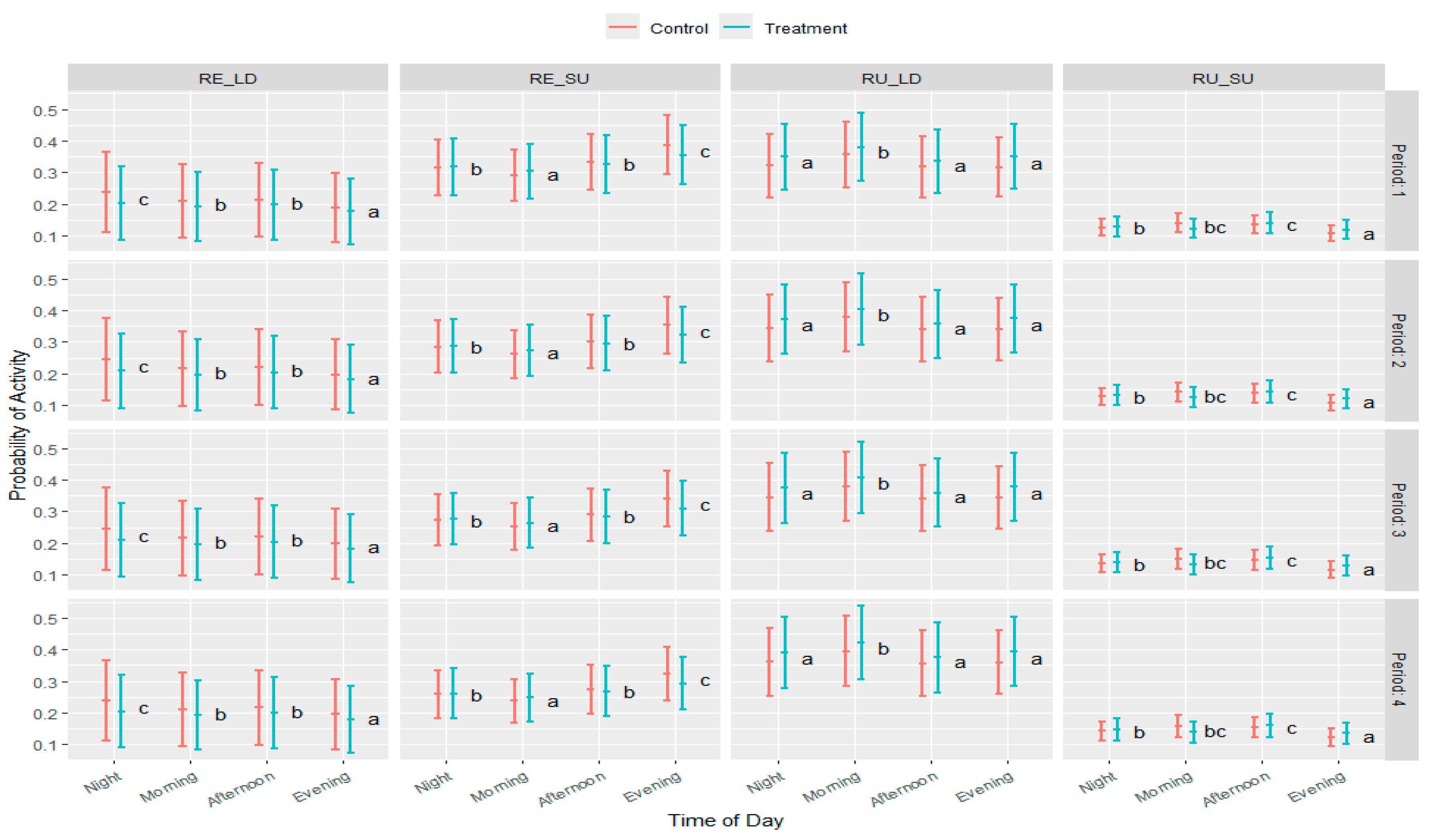
3.4. Impact of Tannin on States and Foraging Activities
3.5. Impact of Tannins on Foraging Behavior When Discriminating Static Activities
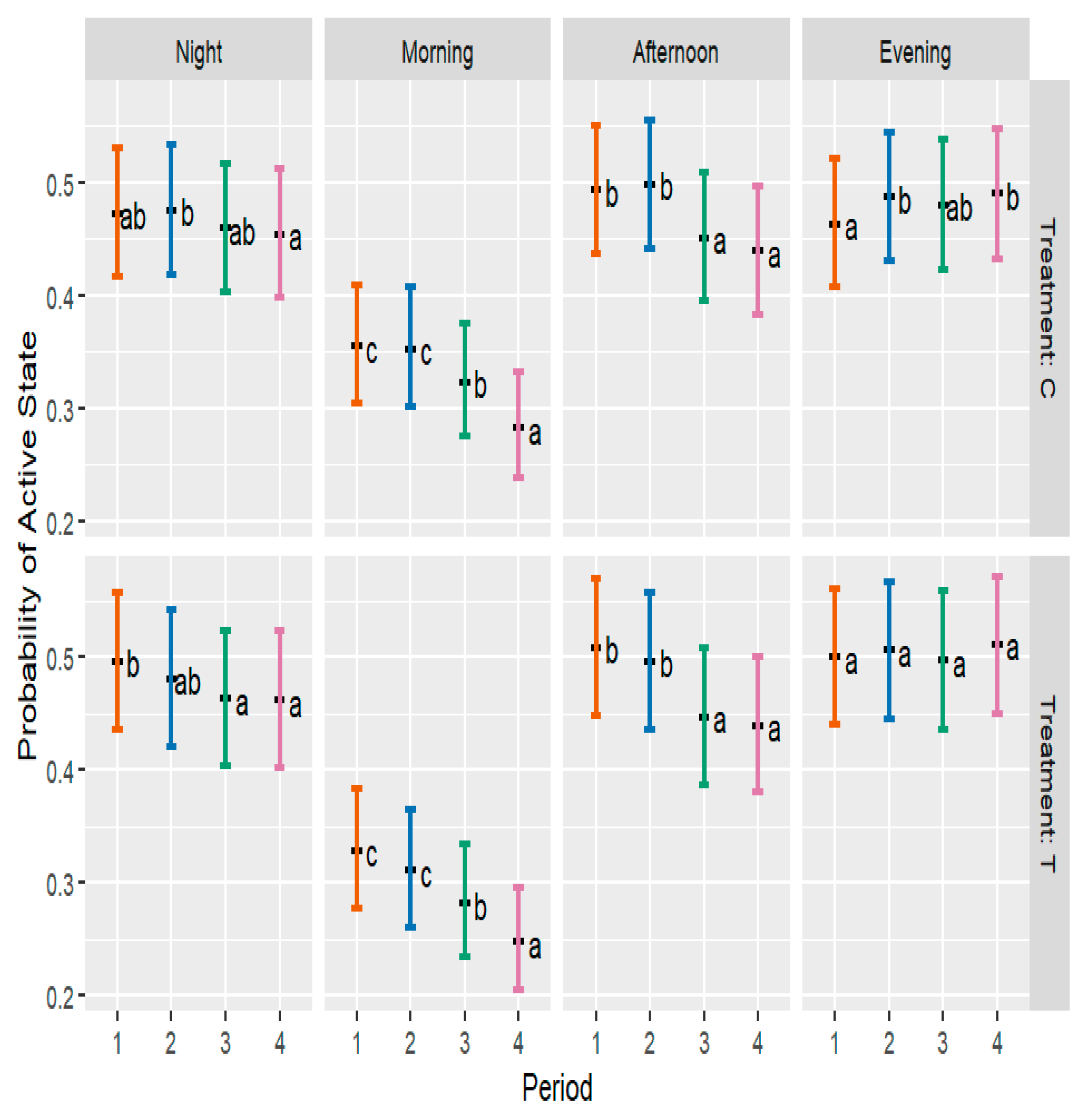
3.6. Behavioral Levels of Activity
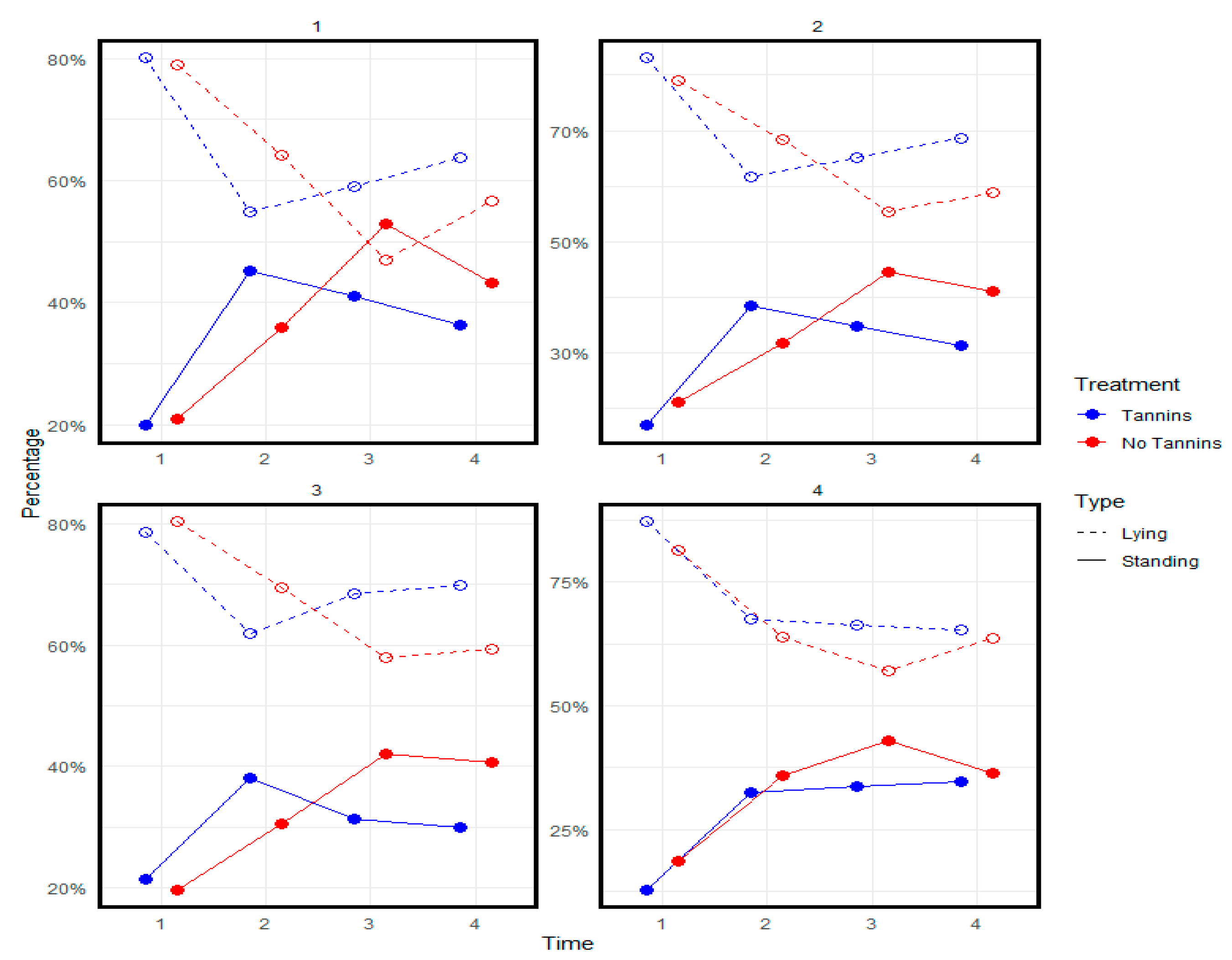
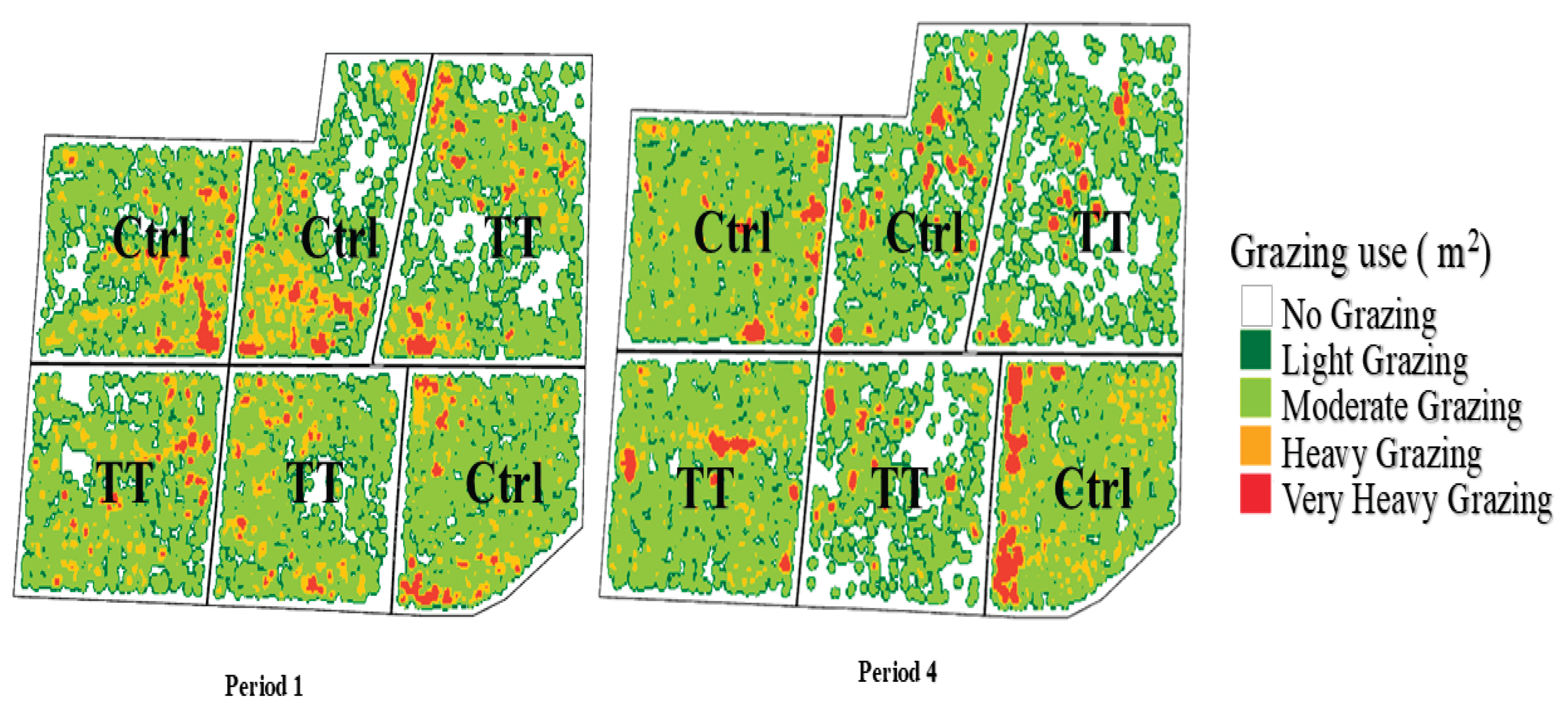
3.7. Animal Distribution
4. Discussion
4.1. Forage Biomass and Nutritional Value
4.2. Supplementation and Forage Disappearance
4.3. Impact of Tannin on Water Consumption and Performance
4.4. Impact of Tannin on Animal Foraging Behaviors
4.5. Impact of Tannin on Animal Grazing Distribution
4.6. Impact of Tannins on Animal Activity Levels
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADG | Average Daily Gain |
| ADL | Acid Detergent Lignin |
| ADF | Acid Detergent Fiber |
| AOAC | Association of Official Analytical Chemists |
| BUN | Blood Urea Nitrogen |
| BW | Body Weight |
| CI | Confidence Interval |
| CP | Crude Protein |
| Ctrl | Control Treatment |
| CV | Coefficient of Variation |
| 5FCV | five-fold cross-validation |
| DDGs | Distillers Dried Grains with Solubles |
| DM | Dry Matter |
| GR | Grazing |
| GPS | Global Positioning System |
| HTs | Hydrolyzable Tannins |
| IACUC | Institutional Animal Care and Use Committee |
| LD | Lying Down (posture classification) |
| MBG | Meadow Bromegrass (Bromus inermis) |
| MALDI-TOF | Matrix-Assisted Laser Desorption/Ionization-Time of Flight (mass spectrometry) |
| ML | Machine Learning |
| N | Nitrogen |
| NDF | Neutral Detergent Fiber |
| NASS | National Agricultural Statistics Service |
| NZ | New Zealand |
| P | Probability value (p-value in statistics) |
| PD | Prediction Dataset |
| PFB | Predicted foraging behaviors |
| PSC / PSCs | Plant Secondary Compounds |
| PSM / PSMs | Plant Secondary Metabolites |
| RE | Resting |
| RE_LD | Resting while Lying Down |
| RE_SU | Resting while Standing Up |
| RF | Random Forest |
| RTS | Random Test-Split |
| RU | Rumination |
| RU_LD | Rumination while Lying Down |
| RU_SU | Rumination while Standing Up |
| SE | Standard Error |
| SEM | Standard Error of the Mean |
| STD | Standard Deviation |
| SU | Standing Up (posture classification) |
| TOD | Time of Day |
| TT | Tannin Treatment |
| UTM | Universal Transverse Mercator (coordinate system) |
| WGS84 | World Geodetic System 1984 |
Appendix A

| Periods | Start Date | End Date | Number of Days |
| Period 1 | July 13 | July 27 | 15 |
| Period 2 | July 28 | Aug 11 | 15 |
| Period 3 | Aug 12 | Aug 26 | 15 |
| Period 4 | Aug 27 | Sept 10 | 15 |
| Predictors | Definition |
| x | left-right cows’ head movement |
| y | forward-backward cow’s head movement |
| z | up-down cow’s head movement |
| Actindex | The cows' activity levels, where higher values indicate more active behaviors and lower values suggest rest periods |
| Distance | Distance traveled between two consecutive GPS points. |
| Speed | Distance the cows traveled over time during the observation period. |
| Time of Day Interval | Hour Range |
| 1 | 4:00 am–10:00 am |
| 2 | 10:00 am–4:00 pm |
| 3 | 4:00 pm–10:00 pm |
| 4 | 10:00 pm–4:00 am |
References
- United States Cattle Inventory Report. Available online: https://www.nass.usda.gov/Newsroom/2025/07-25-2025.php#skipnav (accessed on 1 October 2025).
- ritzler, L.D. Grazing patterns, diet quality, and performance of cow-calf pairs grazing short grass prairie using continuous or high stocking density grazing strategies. Master’s thesis, West Texas A&M University, Canyon, Texas, August 2020. 20 August.
- Putnam, D.H.; DelCurto, T. Forage Systems for Arid Areas. Forages: The Science of Grassland Agriculture: Volume 2, 7th Edition 2020, 2, 433–451. [Google Scholar] [CrossRef]
- Mccollum, F.T.; Horn, G.W. Protein Supplementation of Grazing Livestock: A Review. Prof Anim Sci 1990, 6, 1–16. [Google Scholar] [CrossRef]
- Cappellozza, B.I.; Cooke, R.F.; Guarnieri Filho, T.A.; Bohnert, D.W. Supplementation Based on Protein or Energy Ingredients to Beef Cattle Consuming Low-Quality Cool-Season Forages: I. Forage Disappearance Parameters in Rumen-Fistulated Steers and Physiological Responses in Pregnant Heifers. J Anim Sci 2014, 92, 2716–2724. [Google Scholar] [CrossRef]
- Keller, M.; Kreuzer, M.; Reidy, B.; Scheurer, A.; Liesegang, A.; Giller, K. Methane Emission, Nitrogen and Energy Utilisation of Beef Cattle When Replacing or Omitting Soybean Meal in a Forage-Based Diet. Anim Feed Sci Technol 2022, 290, 115362. [Google Scholar] [CrossRef]
- Barry, T.N.; Duncan, S.J. The Role of Condensed Tannins in the Nutritional Value of Lotus Pedunculatus for Sheep: 1. Voluntary Intake. British Journal of Nutrition 1984, 51, 485–491. [Google Scholar] [CrossRef]
- Kadigi, J.H.; Muzzo, B.I.; Schreiber, S. Potential Benefits of Tannins on Ruminant Health, Production and Environmental Sustainability. Eur J Nutr Food Saf 2024, 16, 13–24. [Google Scholar] [CrossRef]
- Lagrange, S.; Villalba, J.J. Tannin-Containing Legumes and Forage Diversity Influence Foraging Behavior, Diet Digestibility, and Nitrogen Excretion by Lambs 1,2 3995 Tannin-Containing Legumes and Forage Intake. J. Anim. Sci 2019, 97, 3994–4009. [Google Scholar] [CrossRef] [PubMed]
- Molle, G.; Decandia, M.; Sölter, U.; Greef, J.M.; Rochon, J.J.; Sitzia, M.; Hopkins, A.; Rook, A.J. The effect of different legume-based swards on intake and performance of grazing ruminants under Mediterranean and cool temperate conditions. Grass Forage Sci. 2008, 63, 513–530. [Google Scholar] [CrossRef]
- Ebert, P.J.; Bailey, E.A.; Shreck, A.L.; Jennings, J.S.; Cole, N.A. Effect of Condensed Tannin Extract Supplementation on Growth Performance, Nitrogen Balance, Gas Emissions, and Energetic Losses of Beef Steers. J Anim Sci 2017, 95, 1345–1355. [Google Scholar] [CrossRef] [PubMed]
- Orzuna-Orzuna, J.F.; Dorantes-Iturbide, G.; Lara-Bueno, A.; Mendoza-Martínez, G.D.; Miranda-Romero, L.A.; Hernández-García, P.A. Effects of Dietary Tannins’ Supplementation on Growth Performance, Rumen Fermentation, and Enteric Methane Emissions in Beef Cattle: A Meta-Analysis. Sustainability (Switzerland) 2021, 13, 7410. [Google Scholar] [CrossRef]
- Galyean, M.L.; Gunter, S.A. Predicting forage intake in extensive grazing systems. J. Anim. Sci. 2016, 94 (Suppl. 6), 26–43. [Google Scholar] [CrossRef]
- Dearing, M.D.; Mangione, A.M.; Karasov, W.H. Plant Secondary Compounds as Diuretics: An Overlooked Consequence. Integr Comp Biol 2001, 41, 890–901. [Google Scholar] [CrossRef]
- Dearing, M.D.; Mangione, A.; Karasov, W.H. Ingestion of Plant Secondary Compounds Causes Diuresis in Desert Herbivores. 2002. [CrossRef]
- Beale, P.K.; Foley, W.J.; Moore, B.D.; Marsh, K.J. Warmer Ambient Temperatures Reduce Protein Intake by a Mammalian Folivore. 2023. [CrossRef]
- Marsh, K.J.; Wallis, I.R.; Andrew, R.L.; Foley, W.J. The Detoxification Limitation Hypothesis: Where Did It Come from and Where Is It Going? J Chem Ecol 2006, 32, 1247–1266. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Rong, H.; Liu, H.; Wang, X.; Gao, Y.; Deng, R.; Liu, R.; Liu, Y.; Zhang, D. Detoxification Mechanisms, Defense Responses, and Toxicity Threshold in the Earthworm Eisenia Foetida Exposed to Ciprofloxacin-Polluted Soils. Science of The Total Environment 2018, 612, 442–449. [Google Scholar] [CrossRef]
- Provenza, F.D.; Villalba, J.J.; Dziba, L.E.; Atwood, S.B.; Banner, R.E. Linking Herbivore Experience, Varied Diets, and Plant Biochemical Diversity. Small Ruminant Research 2003, 49, 257–274. [Google Scholar] [CrossRef]
- Distel, R.A.; Arroquy, J.I.; Lagrange, S.; Villalba, J.J. Designing Diverse Agricultural Pastures for Improving Ruminant Production Systems. Front Sustain Food Syst 2020, 4. [Google Scholar] [CrossRef]
- Tedeschi, L.O.; Muir, J.P.; Naumann, H.D.; Norris, A.B.; Ramírez-Restrepo, C.A.; Mertens-Talcott, S.U. Nutritional aspects of ecologically relevant phytochemicals in ruminant production. Front. Vet. Sci. 2021, 8, 628445. [Google Scholar] [CrossRef]
- Mlambo, V.; Marume, U.; Gajana, C.S. Utility of the Browser’s Behavioural and Physiological Strategies in Coping with Dietary Tannins: Are Exogenous Tannin-Inactivating Treatments Necessary? S Afr J Anim Sci 2015, 45, 441–451. [Google Scholar] [CrossRef]
- Derix, J. The effect of high tannin concentrations in feed on protein digestion: grazers versus intermediate browsers. Master’s dissertation, Ghent University, Ghent, Belgium, 2017.
- Climate United States - Monthly Averages. Available online: https://www.usclimatedata.com/ (accessed on 1 October 2025).
- Decker, T. Targeted-Grazing as a Fuels Reduction Treatment: Evaluation of Vegetation Dynamics and Utilization Levels. Master’s thesis, Utah State University, Logan, Utah, May 2018.
- Pizzi, A.; Pasch, H.; Rode, K.; Giovando, S. Polymer Structure of Commercial Hydrolyzable Tannins by Matrix-Assisted Laser Desorption/Ionization-Time-of-Flight Mass Spectrometry. J Appl Polym Sci 2009, 113, 3847–3859. [Google Scholar] [CrossRef]
- Lagrange, S.; Beauchemin, K.A.; MacAdam, J.; Villalba, J.J. Grazing Diverse Combinations of Tanniferous and Non-Tanniferous Legumes: Implications for Beef Cattle Performance and Environmental Impact. Science of the Total Environment 2020, 746. [Google Scholar] [CrossRef] [PubMed]
- Šárová, R.; Špinka, M.; Panamá, J.L.A.; Šimeček, P. Graded Leadership by Dominant Animals in a Herd of Female Beef Cattle on Pasture. Anim Behav 2010, 79, 1037–1045. [Google Scholar] [CrossRef]
- Muzzo, B.I.; Bladen, K.; Perea, A.; Nyamuryekung’e, S.; Villalba, J.J. Multi-Sensor Integration and Machine Learning for High-Resolution Classification of Herbivore Foraging Behavior. Animals 2025, Vol. 15, Page 913 2025, 15, 913. [Google Scholar] [CrossRef]
- Paz-Kagan, T.; Chang, J.G.; Shoshany, M.; Sternberg, M.; Karnieli, A. Assessment of Plant Species Distribution and Diversity along a Climatic Gradient from Mediterranean Woodlands to Semi-Arid Shrublands. GIsci Remote Sens 2021, 58, 929–953. [Google Scholar] [CrossRef]
- Finney, G.; Gordon, A.; Scoley, G.; Morrison, S.J. Validating the IceRobotics IceQube Tri-Axial Accelerometer for Measuring Daily Lying Duration in Dairy Calves. Livest Sci 2018, 214, 83–87. [Google Scholar] [CrossRef]
- Robertson, J.B. and Van Soest, P.J. (1981) The Detergent System of Analysis. In James, W.P.T. and Theander, O. (Eds.), The Analysis of Dietary Fibre in Food, Chapter 9, Marcel Dekker, New York, 123-158. - References - Scientific Research Publishing. Available online: https://www.scirp.org/reference/referencespapers?referenceid=1881877 (accessed on 2 October 2025).
- Hartig, F. Residual Diagnostics for Hierarchical (Multi-Level / Mixed) Regression Models [R Package DHARMa Version 0.4.7]. CRAN: Contributed Packages 2024. [CrossRef]
- Box, G.E.P.; Cox, D.R. An Analysis of Transformations. J R Stat Soc Series B Stat Methodol 1964, 26, 211–243. [Google Scholar] [CrossRef]
- Villalba, J.J.; Provenza, F.D. Foraging in Chemically Diverse Environments: Energy, Protein, and Alternative Foods Influence Ingestion of Plant Secondary Metabolites by Lambs. J Chem Ecol 2005, 31, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Waghorn, G. Beneficial and Detrimental Effects of Dietary Condensed Tannins for Sustainable Sheep and Goat Production—Progress and Challenges. Anim Feed Sci Technol 2008, 147, 116–139. [Google Scholar] [CrossRef]
- Patra, A.K.; Saxena, J. Exploitation of Dietary Tannins to Improve Rumen Metabolism and Ruminant Nutrition. J Sci Food Agric 2011, 91, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xu, X.; Cao, Z.; Wang, Y.; Yang, H.; Azarfar, A.; Li, S. Effect of Different Tannin Sources on Nutrient Intake, Digestibility, Performance, Nitrogen Utilization, and Blood Parameters in Dairy Cows. Animals 2019, Vol. 9, Page 507 2019, 9, 507. [Google Scholar] [CrossRef] [PubMed]
- Naumann, H.D.; Tedeschi, L.O.; Zeller, W.E.; Huntley, N.F. The Role of Condensed Tannins in Ruminant Animal Production: Advances, Limitations and Future Directions. Revista Brasileira de Zootecnia 2017, 46, 929–949. [Google Scholar] [CrossRef]
- Kelln, B.M.; Penner, G.B.; Acharya, S.N.; McAllister, T.A.; Lardner, H.A. Impact of condensed tannin-containing legumes on ruminal fermentation, nutrition, and performance in ruminants: A review. Can. J. Anim. Sci. 2020, 101, 210–223. [Google Scholar] [CrossRef]
- Cao, F.; Li, W.; Jiang, Y.; Gan, X.; Zhao, C.; Ma, J. Effects of Grazing on Grassland Biomass and Biodiversity: A Global Synthesis. Field Crops Res 2024, 306. [Google Scholar] [CrossRef]
- Teague, R.; Provenza, F.D.; Norton, B.; Steffens, T.; Barnes, M.; Kothmann, M.; Roath, R. Benefits of Multi-Paddock Grazing Management on Rangelands: Limitations of Experimental Grazing Research and Knowledge Gaps.
- Tavakoli, H.; Saghi, D.A.; Kazemi, M. Response of Russian Brome, Crested Wheatgrass and Tall Wheatgrass to Annual Precipitation and Grazing Management in a Semi-Arid Area;
- Osborne, C.; Yorty, D. Northern Utah Report 2023-24.
- Izaurralde, R.C.; Thomson, A.M.; Morgan, J.A.; Fay, P.A.; Polley, H.W.; Hatfield, J.L. Climate Impacts on Agriculture: Implications for Forage and Rangeland Production. Agron J 2011, 103, 371–381. [Google Scholar] [CrossRef]
- Van Soest, P.J. Nutritional ecology of the ruminant. Cornell university press 1994, ix–xii.
- Jensen, K.B.; Robins, J.G.; Rigby, C.; Waldron, B.L. Comparative Trends in Forage Nutritional Quality across the Growing Season in 13 Grasses. Canadian Journal of Plant Science 2016, 97, 72–82. [Google Scholar] [CrossRef]
- Gillette, K. Yield and quality of cool-season perennial grasses for forage and biomass feedstocks in northeast Colorado. Master’s thesis, Colorado State University, Fort Collins, Colorado, Spring 2011.
- Ritz, K.E.; Heins, B.J.; Moon, R.; Sheaffer, C.; Weyers, S.L. Forage Yield and Nutritive Value of Cool-Season and Warm-Season Forages for Grazing Organic Dairy Cattle. Agronomy 2020, 10. [Google Scholar] [CrossRef]
- DelCurto, T.; Cochran, R.C.; Harmon, D.L.; Beharka, A.A.; Jacques, K.A.; Towne, G.; Vanzant, E.S. Supplementation of Dormant Tallgrass-Prairie Forage: I. Influence of Varying Supplemental Protein and(or) Energy Levels on Forage Utilization Characteristics of Beef Steers in Confinement. J Anim Sci 1990, 68, 515–531. [Google Scholar] [CrossRef] [PubMed]
- Köster, H.H.; Woods, B.C.; Cochran, R.C.; Vanzant, E.S.; Titgemeyer, E.C.; Grieger, D.M.; Olson, K.C.; Stokka, G. Effect of Increasing Proportion of Supplemental N from Urea in Prepartum Supplements on Range Beef Cow Performance and on Forage Intake and Digestibility by Steers Fed Low-Quality Forage. J Anim Sci 2002, 80, 1652–1662. [Google Scholar] [CrossRef]
- Fonseca, N.V.B.; Cardoso, A. da S.; Granja-Salcedo, Y.T.; Siniscalchi, D.; Camargo, K.D.V.; Dornellas, I.A.; Silva, M.L.C.; Vecchio, L. dos S. Del; Grizotto, R.K.; Reis, R.A. Effects of Condensed Tannin-Enriched Alternative Energy Feedstuff Supplementation on Performance, Nitrogen Utilization, and Rumen Microbial Diversity in Grazing Beef Cattle. Livest Sci 2024, 287. [Google Scholar] [CrossRef]
- Muzzo, I.; Blanchard, H.; Cabral, C.; Villalba, J.J. PSXIII-5 Behavior and Blood Urea Nitrogen in Cattle Grazing Meadow Bromegrass and Supplemented with Condensed and Hydrolizable Tannins. J Anim Sci 2024, 102, 614–615. [Google Scholar] [CrossRef]
- Blanchard, H.P. Supplementing Tannins to Increase Performance and Reduce Methane Emissions in Cows Grazing on Western Grasslands. Master’s thesis, Utah State University, Logan, Utah, August 2025. 20 August.
- Mueller-Harvey, I. Unravelling the Conundrum of Tannins in Animal Nutrition and Health. J Sci Food Agric 2006, 86, 2010–2037. [Google Scholar] [CrossRef]
- Matabane, D.M.; Ng’ambi, J.W.; Mabelebele, M.; Gunya, B.; Manyelo, T.G.; Matabane, D.M.; Ng’ambi, J.W.; Mabelebele, M.; Gunya, B.; Manyelo, T.G. The Role of Secondary Metabolites on Methane Reduction in Small Ruminants. 2024. [CrossRef]
- Villalba, J.J.; Provenza, F.D.; Catanese, F.; Distel, R.A. Understanding and Manipulating Diet Choice in Grazing Animals. Anim Prod Sci 2015, 55, 261–271. [Google Scholar] [CrossRef]
- Provenza, F.D.; Villalba, J.J. Foraging in Domestic Herbivores: Linking the Internal and External Milieux. Feeding in Domestic Vertebrates 2006, 210–240. [Google Scholar] [CrossRef]
- Villalba, J.J.; Mereu, A.; Ipharraguerre, I.R. Influence of Dietary Flavours on Sheep Feeding Behaviour and Nutrient Digestibility. Anim Prod Sci 2014, 55, 634–638. [Google Scholar] [CrossRef]
- Bach, A.; Villalba, J.J.; Ipharraguerre, I.R. Interactions between Mild Nutrient Imbalance and Taste Preferences in Young Ruminants, J Anim Sci 2012, 90, 1015–1025. [Google Scholar] [CrossRef]
- Waghorn, G.C.; McNabb, W.C. Consequences of Plant Phenolic Compounds for Productivity and Health of Ruminants. Proceedings of the Nutrition Society 2003, 62, 383–392. [Google Scholar] [CrossRef]
- Min, B.R.; Solaiman, S. Comparative Aspects of Plant Tannins on Digestive Physiology, Nutrition and Microbial Community Changes in Sheep and Goats: A Review. J Anim Physiol Anim Nutr (Berl) 2018, 102, 1181–1193. [Google Scholar] [CrossRef] [PubMed]
- da Silva Aguiar, F.; Bezerra, L.R.; Cordão, M.A.; Cavalcante, I.T.R.; de Oliveira, J.P.F.; do Nascimento, R.R.; de Souza, B.B.; Oliveira, R.L.; Pereira, E.S.; Pereira Filho, J.M. Effects of Increasing Levels of Total Tannins on Intake, Digestibility, and Balance of Nitrogen, Water, and Energy in Hair Lambs. Animals 2023, 13, 2497. [Google Scholar] [CrossRef] [PubMed]
- Besharati, M.; Maggiolino, A.; Palangi, V.; Kaya, A.; Jabbar, M.; Eseceli, H.; De Palo, P.; Lorenzo, J.M. Tannin in Ruminant Nutrition: Review. Molecules 2022, Vol. 27, Page 8273 2022, 27, 8273. [Google Scholar] [CrossRef]
- Makkar, H.P.S. Effects and Fate of Tannins in Ruminant Animals, Adaptation to Tannins, and Strategies to Overcome Detrimental Effects of Feeding Tannin-Rich Feeds. Small Ruminant Research 2003, 49, 241–256. [Google Scholar] [CrossRef]
- Carter, R.R.; Grovum, W.L. A review of the physiological significance of hypertonic body fluids on feed intake and ruminal function: Salivation, motility and microbes. J. Anim. Sci. 1990, 68, 2811–2832. [Google Scholar] [CrossRef] [PubMed]
- Hoste, H.; Jackson, F.; Athanasiadou, S.; Thamsborg, S.M.; Hoskin, S.O. The Effects of Tannin-Rich Plants on Parasitic Nematodes in Ruminants. Trends Parasitol 2006, 22, 253–261. [Google Scholar] [CrossRef]
- Silanikove, N. The Physiological Basis of Adaptation in Goats to Harsh Environments. Small Ruminant Research 2000, 35, 181–193. [Google Scholar] [CrossRef]
- Gedir, J. V.; Cain, J.W.; Krausman, P.R.; Allen, J.D.; Duff, G.C.; Morgart, J.R. Potential Foraging Decisions by a Desert Ungulate to Balance Water and Nutrient Intake in a Water-Stressed Environment. PLoS One 2016, 11. [Google Scholar] [CrossRef]
- Li, C.; Zhang, F.; Leong Tan, M.; Pan Chun, K.; Iddy Muzzo, B.; Douglas Ramsey, R.; Villalba, J.J. Changes in Climate and Their Implications for Cattle Nutrition and Management. Climate 2025, Vol. 13, Page 1 2024, 13, 1. [Google Scholar] [CrossRef]
- Schlecht, E.; Dickhöfer, U.; Predotova, M.; Buerkert, A. The Importance of Semi-Arid Natural Mountain Pastures for Feed Intake and Recycling of Nutrients by Traditionally Managed Goats on the Arabian Peninsula. J Arid Environ 2011, 75, 1136–1146. [Google Scholar] [CrossRef]
- Burgos, M.S.; Senn, M.; Sutter, F.; Kreuzer, M.; Langhans, W. Effect of Water Restriction on Feeding and Metabolism in Dairy Cows. Am J Physiol Regul Integr Comp Physiol 2001, 280. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.H.; Mc Geough, E.J.; Acharya, S.; McAllister, T.A.; McGinn, S.M.; Harstad, O.M.; Beauchemin, K.A. Enteric Methane Emission, Diet Digestibility, and Nitrogen Excretion from Beef Heifers Fed Sainfoin or Alfalfa. J Anim Sci 2013, 91, 4861–4874. [Google Scholar] [CrossRef]
- Ruckle, M.E.; Meier, M.A.; Frey, L.; Eicke, S.; Kölliker, R.; Zeeman, S.C.; Studer, B. Diurnal leaf starch content: An orphan trait in forage legumes. Agronomy 2017, 7, 16. [Google Scholar] [CrossRef]
- Gregorini, P.; Pas, †; Tamminga, S.; Gunter, S.A. REVI EW: Behavior and Daily Grazing Patterns of Cattle. Prof Anim Sci 2006, 22, 201–209. [CrossRef]
- Bryant, J.P.; Stuart Chapin, F.; Klein Bryant, D.R.; Bryant, J.P.; Chapin, F.S. Herbivore-Plant Interactions at Northern Latitudes. Source: Oikos 40, 357–368.
- Min, B.R.; Barry, T.N.; Attwood, G.T.; McNabb, W.C. The Effect of Condensed Tannins on the Nutrition and Health of Ruminants Fed Fresh Temperate Forages: A Review. Anim Feed Sci Technol 2003, 106, 3–19. [Google Scholar] [CrossRef]
- Barry, T.N.; Mcnabb, W.C. The Implications of Condensed Tannins on the Nutritive Value of Temperate Forages Fed to Ruminants. Br. J. Nutr. 1999, 81, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Gargallo, S.; Ferret, A.; Calsamiglia, S. Ruminal Microbial Degradation of Individual Amino Acids from Heat-Treated Soyabean Meal and Corn Gluten Meal in Continuous Culture. Animals 2022, Vol. 12, Page 688 2022, 12, 688. [Google Scholar] [CrossRef]
- Herremans, S.; Vanwindekens, F.; Decruyenaere, V.; Beckers, Y.; Froidmont, E. Effect of dietary tannins on milk yield and composition, nitrogen partitioning and nitrogen use efficiency of lactating dairy cows: A meta-analysis. J. Anim. Physiol. Anim. Nutr. 2020, 104(5), 1209–1218. [Google Scholar] [CrossRef]
- Dschaak, C.M.; Williams, C.M.; Holt, M.S.; Eun, J.S.; Young, A.J.; Min, B.R. Effects of Supplementing Condensed Tannin Extract on Intake, Digestion, Ruminal Fermentation, and Milk Production of Lactating Dairy Cows. J Dairy Sci 2011, 94, 2508–2519. [Google Scholar] [CrossRef] [PubMed]
- López-Andrés, P.; Luciano, G.; Vasta, V.; Gibson, T.M.; Biondi, L.; Priolo, A.; Mueller-Harvey, I. Dietary Quebracho Tannins Are Not Absorbed, but Increase the Antioxidant Capacity of Liver and Plasma in Sheep. British Journal of Nutrition 2013, 110, 632–639. [Google Scholar] [CrossRef]
- The Effect of Wattle Tannin on Intake, Digestibility, Nitrogen Retention and Growth Performance of Goats in South Africa. Available online: https://www.lrrd.cipav.org.co/lrrd19/4/beng19050.htm (accessed on 2 October 2025).
- Min, B.R.; Solaiman, S.; Terrill, T.; Ramsay, A.; Mueller-Harvey, I. The Effects of Tannins-Containing Ground Pine Bark Diet upon Nutrient Digestion, Nitrogen Balance, and Mineral Retention in Meat Goats. J Anim Sci Biotechnol 2015, 6, 1–8. [Google Scholar] [CrossRef]
- Tshabalala, T.; Sikosana, J.L.N.; Chivandi, E. Nutrient Intake, Digestibility and Nitrogen Retention in Indigenous Goats Fed on Acacia Nilotica Fruits Treated for Condensed Tannins. S Afr J Anim Sci 2013, 43, 457–463. [Google Scholar] [CrossRef]
- Tucker, C.B.; Rogers, A.R.; Schütz, K.E. Effect of Solar Radiation on Dairy Cattle Behaviour, Use of Shade and Body Temperature in a Pasture-Based System. Appl Anim Behav Sci 2008, 109, 141–154. [Google Scholar] [CrossRef]
- Schütz, K.E.; Rogers, A.R.; Poulouin, Y.A.; Cox, N.R.; Tucker, C.B. The Amount of Shade Influences the Behavior and Physiology of Dairy Cattle. J Dairy Sci 2010, 93, 125–133. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Kreuzer, M.; O’Mara, F.; McAllister, T.A. Nutritional Management for Enteric Methane Abatement: A Review. Aust J Exp Agric 2008, 48, 21–27. [Google Scholar] [CrossRef]
- Jayanegara, A.; Leiber, F.; Kreuzer, M. Meta-Analysis of the Relationship between Dietary Tannin Level and Methane Formation in Ruminants from in Vivo and in Vitro Experiments. J Anim Physiol Anim Nutr (Berl) 2012, 96, 365–375. [Google Scholar] [CrossRef]
- Gregorini, P. Diurnal Grazing Pattern: Its Physiological Basis and Strategic Management. Anim Prod Sci 2012, 52, 416–430. [Google Scholar] [CrossRef]
- Mayland, H.; Mertens, D.; Taylor, T.; Burns, J.; Fisher, D.; Gregorini, P.; Ciavarella, T.; Smith, K.; Shewmaker, G.; Griggs, T. Diurnal changes in forage quality and their effects on animal preference, intake, and performance. In Proceedings of the 35th California Alfalfa and Forage Symposium, Visalia, CA, USA, 12–14 December 2005; University of, California, Ed.; University of California: Davis, CA, USA, 2005; pp. 223–230. [Google Scholar]
- Bailey, D.W.; Stephenson, M.B.; Pittarello, M. Effect of Terrain Heterogeneity on Feeding Site Selection and Livestock Movement Patterns. Anim Prod Sci 2015, 55, 298–308. [Google Scholar] [CrossRef]
- Masters, D.G.; Blache, D.; Lockwood, A.L.; Maloney, S.K.; Norman, H.C.; Refshauge, G.; Hancock, S.N. Shelter and Shade for Grazing Sheep: Implications for Animal Welfare and Production and for Landscape Health. Anim Prod Sci 2023, 63, 623–644. [Google Scholar] [CrossRef]
- Xiao, X.; Zhang, T.; Angerer, J.P.; Hou, F. Grazing Seasons and Stocking Rates Affects the Relationship between Herbage Traits of Alpine Meadow and Grazing Behaviors of Tibetan Sheep in the Qinghai–Tibetan Plateau. Animals 2020, Vol. 10, Page 488 2020, 10, 488. [Google Scholar] [CrossRef] [PubMed]
- van Beest, F.M.; Van Moorter, B.; Milner, J.M. Temperature-Mediated Habitat Use and Selection by a Heat-Sensitive Northern Ungulate. Anim Behav 2012, 84, 723–735. [Google Scholar] [CrossRef]
- Owen-Smith, N.; Fryxell, J.M.; Merrill, E.H. Foraging Theory Upscaled: The Behavioural Ecology of Herbivore Movement. Philosophical Transactions of the Royal Society B: Biological Sciences 2010, 365, 2267–2278. [Google Scholar] [CrossRef]
- Launchbaugh, K.L.; Howery, L.D. Understanding Landscape Use Patterns of Livestock as a Consequence of Foraging Behavior. Rangel Ecol Manag 2005, 58, 99–108. [Google Scholar] [CrossRef]
- Sellick, J.; Yarwood, R. Placing Livestock in Landscape Studies: Pastures New or Out to Graze? Landsc Res 2013, 38, 404–420. [Google Scholar] [CrossRef]
- Bailey, D.W. Management Strategies for Optimal Grazing Distribution and Use of Arid Rangelands, J Anim Sci 2004, 82, E147–E153. [Google Scholar]
- De Weerd, N.; Van Langevelde, F.; Van Oeveren, H.; Nolet, B.A.; Kölzsch, A.; Prins, H.H.T.; De Boer, W.F. Deriving Animal Behaviour from High-Frequency GPS: Tracking Cows in Open and Forested Habitat. PLoS One 2015, 10, e0129030. [Google Scholar] [CrossRef]
- Holbrook, J.D.; Olson, L.E.; DeCesare, N.J.; Hebblewhite, M.; Squires, J.R.; Steenweg, R. Functional Responses in Habitat Selection: Clarifying Hypotheses and Interpretations. Ecological Applications 2019, 29, e01852. [Google Scholar] [CrossRef] [PubMed]
- Bailey, D.W.; Provenza, F.D. Mechanisms Determining Large-Herbivore Distribution. Resource Ecology 2008, 7–28. [Google Scholar] [CrossRef]
- Provenza, F.D.; Papachristou, T.G. Behavior-Based Management of Ecosystems.
- Jin, Z.; Guo, L.; Shu, H.; Qi, J.; Li, Y.; Xu, B.; Zhang, W.; Wang, K.; Wang, W. Behavior Classification and Analysis of Grazing Sheep on Pasture with Different Sward Surface Heights Using Machine Learning. Animals 2022, Vol. 12, Page 1744 2022, 12, 1744. [Google Scholar] [CrossRef]
- Teague, R.; Kreuter, U. Managing Grazing to Restore Soil Health, Ecosystem Function, and Ecosystem Services. Front Sustain Food Syst 2020, 4, 534187. [Google Scholar] [CrossRef]
- Bailey, D.W.; Brown, J.R. Rotational Grazing Systems and Livestock Grazing Behavior in Shrub-Dominated Semi-Arid and Arid Rangelands. Rangel Ecol Manag 2011, 64, 1–9. [Google Scholar] [CrossRef]
- Zilverberg, C.J.; Angerer, J.; Williams, J.; Metz, L.J.; Harmoney, K. Sensitivity of Diet Choices and Environmental Outcomes to a Selective Grazing Algorithm. Ecol Modell 2018, 390, 10–22. [Google Scholar] [CrossRef]
- Lagrange, S.P.; Macadam, J.W.; Villalba, J.J. The Use of Temperate Tannin Containing Forage Legumes to Improve Sustainability in Forage-Livestock Production. 2021. [CrossRef]
- Tan, P.; Liu, H.; Zhao, J.; Gu, X.; Wei, X.; Zhang, X.; Ma, N.; Johnston, L.J.; Bai, Y.; Zhang, W.; et al. Amino Acids Metabolism by Rumen Microorganisms: Nutrition and Ecology Strategies to Reduce Nitrogen Emissions from the inside to the Outside. Science of The Total Environment 2021, 800, 149596. [Google Scholar] [CrossRef]
- Cézilly, F. Stephens, D.W. and Krebs, J.R. — Foraging Theory. Princeton University Press, Princeton N.J., Monographs in Behavior and Ecology, Vol. 4, 1986. Revue d’Écologie (La Terre et La Vie) 1988, 43, 327–327. [Google Scholar]
- Baumont, R.; Prache, S.; Meuret, M.; Morand-Fehr, P. How Forage Characteristics Influence Behaviour and Intake in Small Ruminants: A Review. Livest Prod Sci 2000, 64, 15–28. [Google Scholar] [CrossRef]
- Hall-McMaster, S.; Dayan, P.; Schuck, N.W. Control over Patch Encounters Changes Foraging Behavior. iScience 2021, 24, 103005. [Google Scholar] [CrossRef]
- Villalba, J.J.; Provenza, F.D. Preference for Flavored Wheat Straw by Lambs Conditioned with Intraruminal Infusions of Acetate and Propionate. J Anim Sci 1997, 75, 2905–2914. [Google Scholar] [CrossRef]
- Villalba, J.J.; Provenza, F.D. Preference for Flavoured Foods by Lambs Conditioned with Intraruminal Administration of Nitrogen. British Journal of Nutrition 1997, 78, 545–561. [Google Scholar] [CrossRef]
- Bailey, D.W.; Trotter, M.G.; Tobin, C.; Thomas, M.G. Opportunities to Apply Precision Livestock Management on Rangelands. Front Sustain Food Syst 2021, 5, 611915. [Google Scholar] [CrossRef]
- Lagrange, S.; Macadam, J.W.; Stegelmeier, B.; Villalba, J.J. Grazing Diverse Combinations of Tanniferous and Nontanniferous Legumes: Implications for Foraging Behavior, Performance, and Hair Cortisol in Beef Cattle. J Anim Sci 2021, 99, 1–12. [Google Scholar] [CrossRef]
- Villalba, J.J.; Costes-Thiré, M.; Ginane, C. Phytochemicals in Animal Health: Diet Selection and Trade-Offs between Costs and Benefits. Proceedings of the Nutrition Society 2017, 76, 113–121. [Google Scholar] [CrossRef]
- Boissy, A.; Aubert, A.; Désiré, L.; Greiveldinger, L.; Delval, E.; Veissier, I. Cognitive Sciences to Relate Ear Postures to Emotions in Sheep. Animal Welfare 2011, 20, 47–56. [Google Scholar] [CrossRef]
- Nalon, E.; Contiero, B.; Gottardo, F.; Cozzi, G. The Welfare of Beef Cattle in the Scientific Literature From 1990 to 2019: A Text Mining Approach. Front Vet Sci 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Langbein, C.; Coulon, J.; Gabor, M.; Oesterwind, V.; Benz-Schwarzburg, S.; Borell, J. Farm Animal Cognition-Linking Behavior, Welfare and Ethics. Frontiers in Veterinary Science. 2019, 6, 24. [Google Scholar] [CrossRef]
- Villalba, J.J.; Provenza, F.D.; Shaw, R. Initial Conditions and Temporal Delays Influence Preference for Foods High in Tannins and for Foraging Locations with and without Foods High in Tannins by Sheep. Appl Anim Behav Sci 2006, 97, 190–205. [Google Scholar] [CrossRef]
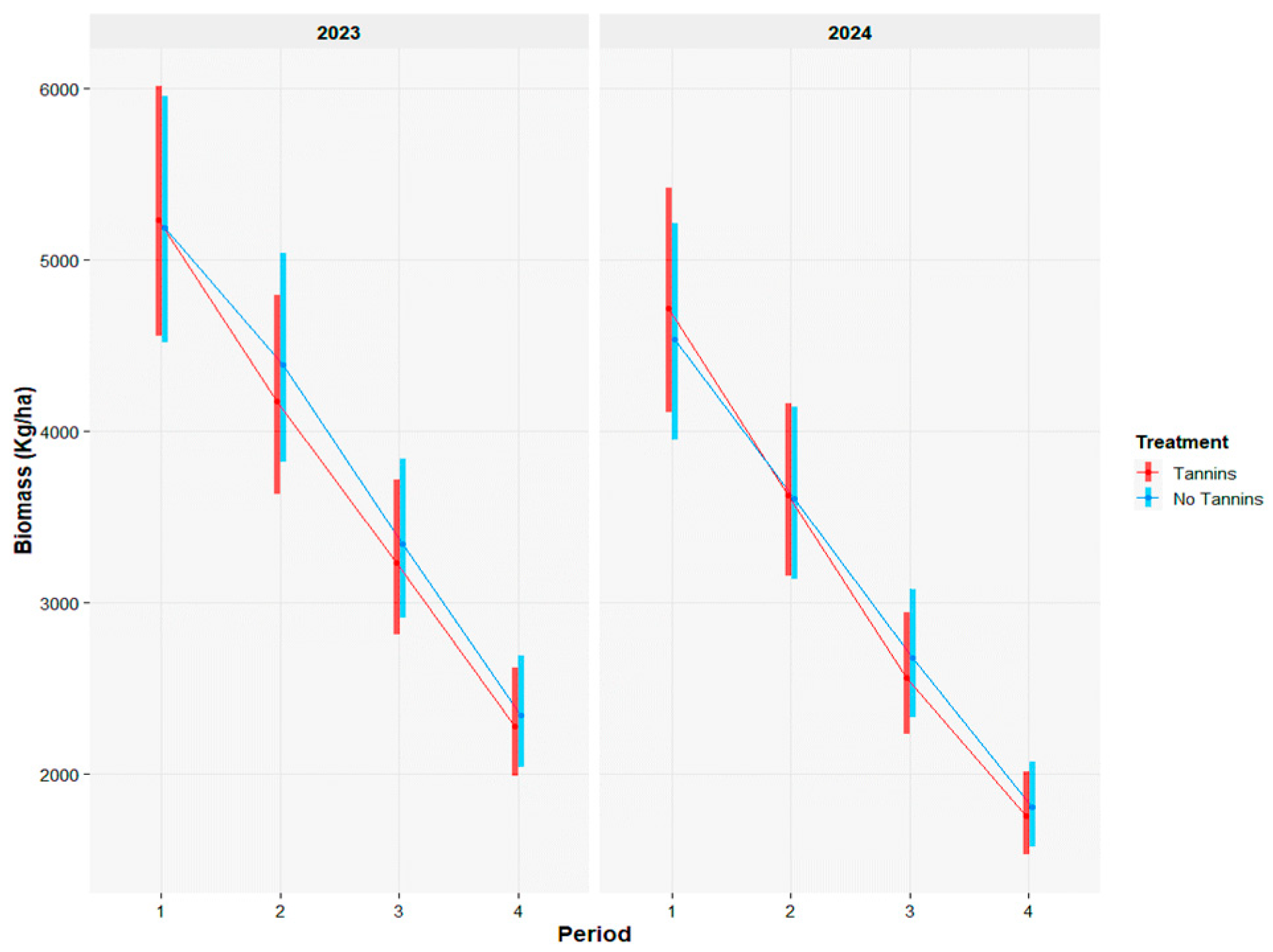
| 2023 | Tannins | No Tannin | SE | ||||||
| Periods | |||||||||
| 1 | 2 | 3 | 4 | 1 | 2 | 3 | |||
| DM (%) | 95.50ᵃ | 96.87ᵇ | 97.47ᶜ | 97.77ᶜ | 95.87ᵃᵇ | 96.13ᵃᵇ | 97.33ᶜ | 97.37ᶜ | ± 0.233 |
| CP (%) | 7.03ᶜ | 5.30ᵇ | 4.77ᵃ | 3.90ᵃ | 6.47ᶜ | 5.13ᵇ | 4.60ᵃ | 3.83ᵃ | ± 0.194 |
| ADF (%) | 38.8ᵃ | 38.6ᵃ | 41.9ᵇ | 43.6ᵇ | 38.5ᵃ | 39.5ᵃ | 42.5ᵇ | 44.2ᵇ | ± 0.477 |
| NDF (%) | 63.6ᵃᵇ | 63.4ᵃᵇ | 64.6ᵃᵇ | 64.8ᵃᵇ | 62.2ᵃ | 63.8ᵃᵇ | 64.7ᵃᵇ | 65.4ᵇ | ± 0.692 |
| 2024 | |||||||||
| DM (%) | 95.60ᵃ | 96.37ᵃᵇ | 96.83ᵇ | 98.00ᶜ | 95.53ᵃ | 96.07ᵃᵇ | 96.73ᵇ | 98.20ᶜ | ± 0.184 |
| CP (%) | 7.07ᶜ | 3.93ᵇ | 3.40ᵇ | 2.47ᵃ | 7.00ᶜ | 4.07ᵇ | 3.43ᵇ | 2.43ᵃ | ± 0.161 |
| ADF (%) | 37.7ᵃ | 41.5ᵇᶜ | 41.9ᵇ | 43.9ᵈ | 37.8ᵃ | 41.7ᵇᶜ | 40.8ᵇ | 43.0ᶜᵈ | ± 0.386 |
| NDF (%) | 59.3ᵃᵇ | 63.0ᶜ | 64.0ᶜ | 68.4ᵈ | 57.8ᵃ | 63.1ᵇᶜ | 64.4ᶜ | 65.5ᶜᵈ | ± 0.77 |
| P values | ||||
| Parameters | % Standing Time | Number of Steps | Motion Index | % Transition Up |
| Treatment | 0.388 | 0.8909 | 0.8986 | 0.0115 |
| Time of the day | 0.0001 | 0.0001 | 0.0001 | 0.058 |
| Period | 0.0077 | 0.0005 | <0.0001 | 0.5806 |
| Year | 0.6178 | 0.1498 | 0.0621 | 0.5258 |
| Treatment × Time of the day | 0.815 | 0.1633 | 0.7370 | 0.1178 |
| Treatment × Period | 0.1417 | 0.3537 | 0.094 | 0.2645 |
| Treatment × Year | 0.5468 | 0.1502 | 0.5468 | 0.2546 |
| Time of the day × Period | 0.0522 | 0.3902 | 0.0522 | 0.3902 |
| Time of the day × Year | 0.0001 | 0.0001 | 0.0001 | 0.0001 |
| Period × Year | 0.0001 | 0.0546 | 0.0001 | 0.0546 |
| Treatment × Time of the day × Period | 0.4013 | 0.7989 | 0.4012 | 0.2645 |
| Treatment × Time of the day × Year | 0.1933 | 0.1124 | 0.1933 | 0.1124 |
| Time of the day × Period × Year | 0.0036 | 0.4401 | 0.0004 | 0.0005 |
| Treatment × Time of the day × Period × Year | 0.3763 | 0.8053 | 0.3763 | 0.8053 |
| Period | Treatment | Min | Max | Mean | STD | Coeff. Var | Count |
| P1 | Tannin | 0.00010 | 1.24642 | 0.07001 | 0.08057 | 1.15084 | 254184 |
| P1 | Control | 0.00010 | 1.78293 | 0.08721 | 0.10641 | 1.22024 | 244770 |
| P4 | Tannin | 0.00010 | 2.59267 | 0.06810 | 0.12674 | 1.86097 | 254184 |
| P4 | Control | 0.00010 | 4.65824 | 0.09744 | 0.20753 | 2.12974 | 244770 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



