Submitted:
14 October 2025
Posted:
14 October 2025
You are already at the latest version
Abstract
E-cigarettes (E-cigs) are increasing in popularity and are considered as a potentially safer alternative to traditional cigarettes. However, prior studies have demonstrated that inhalation of nicotine-containing e-cigs can cause substantial pathophysiologic changes, and “vaping” of some substances has led to severe lung damage. Our group recently described the role of cortactin (CTTN), a cytoskeletal actin-binding regulatory protein, in mediating cigarette smoke (CS) and E-cig-induced lung endothelial apoptosis and mitochondrial dysfunction. In the current study, we advance this work by characterizing the effects of E-cig on lung endothelial nanomechanical properties and barrier function. Lung EC exposure to E-cig extract (50 µg/ml) resulted in disruption of endothelial barrier properties as assessed by Electric Cell Substrate Impedance (ECIS). Since excess mitochondrial ROS (mitoROS) is an important marker of mitochondrial dysfunction, we next assessed the effect of Mito-TEMPO (10 µM, 3 h), a cell-permeable antioxidant, on E-cig-induced endothelial permeability. Pretreatment with Mito-TEMPO provided EC barrier protection after E-cig challenge, suggesting a key role of mitoROS in E-cig-induced EC permeability. E-cig exposure induces cytoskeleton rearrangement leading to gap formation in lung EC and significantly alters EC elastic properties as assessed by atomic force microscopy (AFM). Reduction in CTTN expression by siRNA further augmented the injurious effects of E-cig on EC permeability and elastic properties. This is the first study to explore the role of CTTN in evaluating the effect of E-cigarette exposure on lung endothelium using AFM and provides novel mitochondrial and biophysical characterization of the effects of E-cig exposure on human lung EC. This work advances our understanding of the pathophysiologic effects of E-cig exposure.
Keywords:
mitochondrial dysfunction
; permeability
; elastic properties
; atomic force microscopy
1. Introduction
Electronic cigarettes (E-cig) are gaining popularity, especially among youth, and have been considered as a potentially safer alternative to traditional cigarettes [1,2]. E-cigarettes (E-cig) first entered the US market in 2006, while pod mode forms such as “JUUL” were introduced in 2015. In 2019 the Centers for Disease Control and Prevention (CDC) and Food and Drug Administration (FDA) declared an “E-cigarette epidemic” due to a national outbreak of E-cig-induced acute lung injury (EVALI) cases [3]. E-cig inhalation causes lung tissue damage thereby increasing the risk of chronic lung disease [4,5,6] including COPD. The underlying mechanisms by which E-cigs contribute to lung disease are incompletely understood; however, recent reports suggest that E-cigs cause inflammation, apoptosis, and tissue damage [7,8].
Despite the growing interest in their potential negative effects, it is unknown how E-cig exposure may alter the biomechanical properties of lung endothelial cells (EC), which play a critical role in maintaining vascular barrier integrity [9]. EC dysfunction is a hallmark of various pulmonary diseases, including those associated with cigarette and E-cig exposure [1,9,10]. Cytoskeletal changes are central to EC responses to injurious stimuli such as cigarette smoke (CS) [11]. CS exposure directly modulates EC barrier permeability, necrosis, and apoptosis through multiple signaling pathways involving RhoA, FAK, and p38, which all participate in cytoskeletal rearrangements [12,13,14]. Our group has identified cortactin (CTTN), a central regulator of the actin cytoskeleton, as an important modulator of lung EC function and lung injury responses [15,16]. Understanding the potential detrimental effects of E-cigs on EC cytoskeletal organization and associated biomechanical properties may provide novel insights into EVALI pathophysiology.
In prior work, we have shown that atomic force microscopy (AFM) is an invaluable tool to characterize nanoscale mechanical changes in living cells including endothelium9,10. AFM offers the sensitivity to detect early or subtle cytoskeletal disruptions that may precede overt morphological damage. AFM is a high-resolution scanning probe technique designed to investigate interatomic forces and surface topography at the nanoscale [10]. Using force-indentation curves and contact mechanics models such as Hertz or Sneddon, AFM enables the precise measurement of local elastic responses from cell membranes and underlying cytoskeletal structures [9,10,17]. For most cell types, Young’s modulus typically ranges between 1 and 100 kPa [18,19], with stiffness largely governed by the integrity of the actin cytoskeleton and internal mechanical forces. These nanomechanical insights are critical, as alterations in cell stiffness are closely linked to changes in cellular function and are often associated with pathological processes. For example, malignant cancer cells may appear morphologically similar to benign ones, they can often be differentiated by their lower elastic modulus—reflecting cytoskeletal remodeling that facilitates invasion and metastasis [20]. This softening is often driven by upregulation of actin-regulating proteins such as Arp2/3, cortactin, and cofilin, promoting the formation of invasive structures like podosomes and invadopodia [20,21,22,23]. These findings highlight AFM’s sensitivity in detecting biomechanical signatures of disease, particularly those driven by cytoskeletal reorganization. However, accurate interpretation of AFM-derived mechanical data depends on the probe geometry, indentation models, applied forces, and cell type, along with calibration for reproducibility.
In this study, we apply AFM to evaluate the nanomechanical properties of lung endothelial cells exposed to E-cigarette extract. We characterize cell stiffness through spatial force mapping and analyze elastic modulus, providing insight into how E-cigarette exposure disrupts endothelial mechanical integrity. This work provides novel insight into the mechanical underpinnings of E-cig-induced endothelial dysfunction and further advances our understanding of the pathophysiological effects of E-cigs on the lung vasculature.
2. Materials and Methods
2.1. Cell Culture
Human pulmonary artery endothelial cells (HPAECs) were purchased from Lonza (Walkersville, MD) and cultured in Endothelial Cell Growth Medium-2 (EGM-2) (Lonza) supplemented with 10% fetal bovine serum (FBS) (Sigma, St Louis, MO). Cells were maintained at 37 °C in a 5% CO2 incubator and used at passages 6–8 for all experiments. Cells were starved for 2 hours in 2% FBS media prior to treatments.
2.2. E-Cigarette Preparation
“JUUL” E-cigarette was obtained commercially. The 5.0% nicotine, unflavored pod was used for our experiments and contains nicotine, propylene glycol, glycerin, and benzoic acid. Cells were treated with 50 μg/ml of E-cig liquid for 24 hours.
2.3. siRNA Transfection
HPAECs were transfected with scrambled RNA or CTTN siRNA (100 nM) using the DharmaFECT 1 transfection reagent. 48 hours after transfection, EC were challenged with E-cig. Transfection efficiency was determined by western blotting.
2.4. Immunofluorescence Microscopy
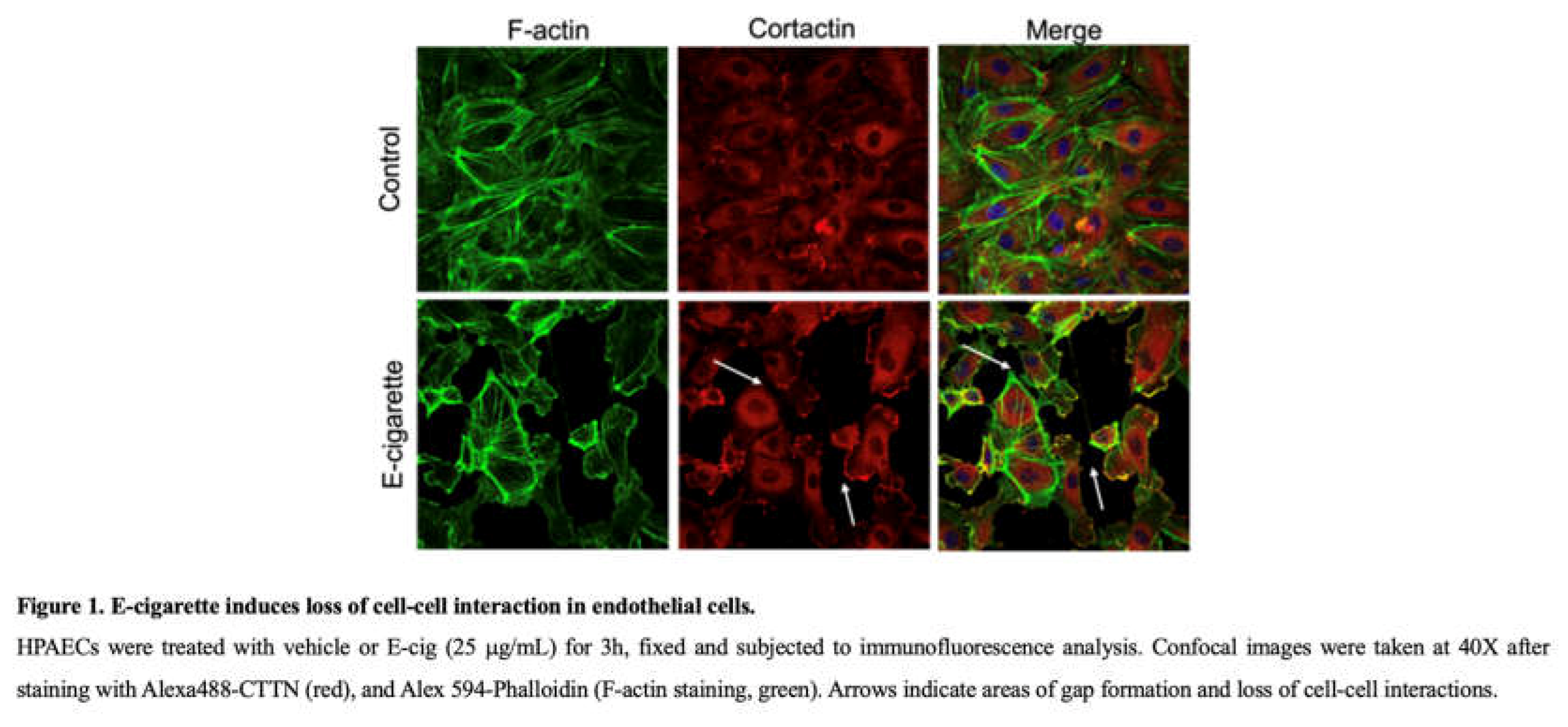
HPAECs were grown on 8 well glass chamber slide to 80-90% confluence in EGM-2 medium. After indicated treatments, cells were fixed with 3.7% paraformaldehyde for 10 min followed by three washes with PBS. The cells were then permeabilized with 0.25% Triton X 100 for 5 min and rinsed with PBS for 5 mins followed by incubation in blocking buffer (1% BSA-PBS) for 1 hour. Cells were then incubated with cortactin antibody for 1 hour, washed with PBS and then incubated with secondary antibody-Alexa Fluor 488 and Alexa 594-Phalloidin (F-actin staining) for 1 hour. After washing at least four times, the coverslips were mounted with profound gold DAPI (Invitrogen). Images were taken using a Zeiss confocal microscope at 40x magnification.
2.5. Reagents
Horseradish Peroxidase (HRP)-linked anti-mouse and anti-rabbit secondary antibodies, anti-β actin antibodies and MitoTEMPO were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). SiRNA (control and CTTN) and DharmaFECT1 transfection reagent were purchased from Dharmacon (Horizon Inspired Cell Solutions). Anti-CTTN antibody was purchased from Sigma Aldrich.
2.6. Trans-Endothelial Monolayer Electrical Resistance (TEER) Measurements
Electrical cell impedance sensing (ECIS) (Applied Biophysics, Troy, NY) was used to evaluate the integrity of the EC monolayer, as described in our previous work [24]. Briefly, HPAEC were grown to confluence on gold microelectrodes, and the cells were subjected to a weak electric current for the continuous measurement of TER. After 1 h of measurement (to establish a stable baseline resistance), the indicated stimuli were added to the wells and measurement continued for 20 h. The data were analyzed using custom-designed Epool software by normalizing each resistance value to the starting resistance for that electrode as we have previously described [24].
2.7. AFM Imaging
Asylum MFP-3D-Bio atomic microscope (Oxford Instruments Asylum research, Santa Barbara, CA) and PNP-TR-Au-1 triangular silicon nitride cantilevers (with tip height 3.5 µm and radius of curvature 10 nm) were purchased from Asylum Oxford Research Group. Imaging for cells in liquid was performed in contact mode. Cantilevers with spring constant of k=0.08N/m or 0.17N/m were used for imaging and elasticity measurements. Mechanical properties of the cell cytoskeleton were acquired in tapping force mode. The tip velocity was set at 1.99 µm/s. The cell indentation part of the force curves were fitted to the Hertz model [25] and yield the Young’s elastic modulus as a measure of cell stiffness, E.
2.8. Statistical Significance and Data Analysis
All data are expressed as mean ± SEM from at least three independent experiments. Statistical analysis was performed using the GraphPad Prism 8 software. Student’s t-test or two-way ANOVA (Tukey’s or Dunnett’s post hoc tests) were used to compare two or more groups respectively. Values of *p < 0.05 were considered statistically significant.
3. Results
3.1. E-Cigarette Exposure Induces Cytoskeletal Rearrangement and Gap Formation in Lung ECs
We first evaluated the effects of E-cig on the structure of critical cytoskeletal proteins cortactin (CTTN) and actin in cultured human lung EC [15]. As revealed by immunofluorescence imaging, E-cig stimulation of HPAEC causes redistribution of actin and CTTN that decreases their expression at the cell periphery (Figure 1). This pattern is similar to that we previously reported in response to CS exposure and is associated with decreased cell-cell interaction, gap formation between cells, and increased EC permeability [11,26,27].
3.2. Trans-Endothelial Resistance Is Decreased by E-Cigarette Exposure in a Dose-Dependent Manner in Lung ECs
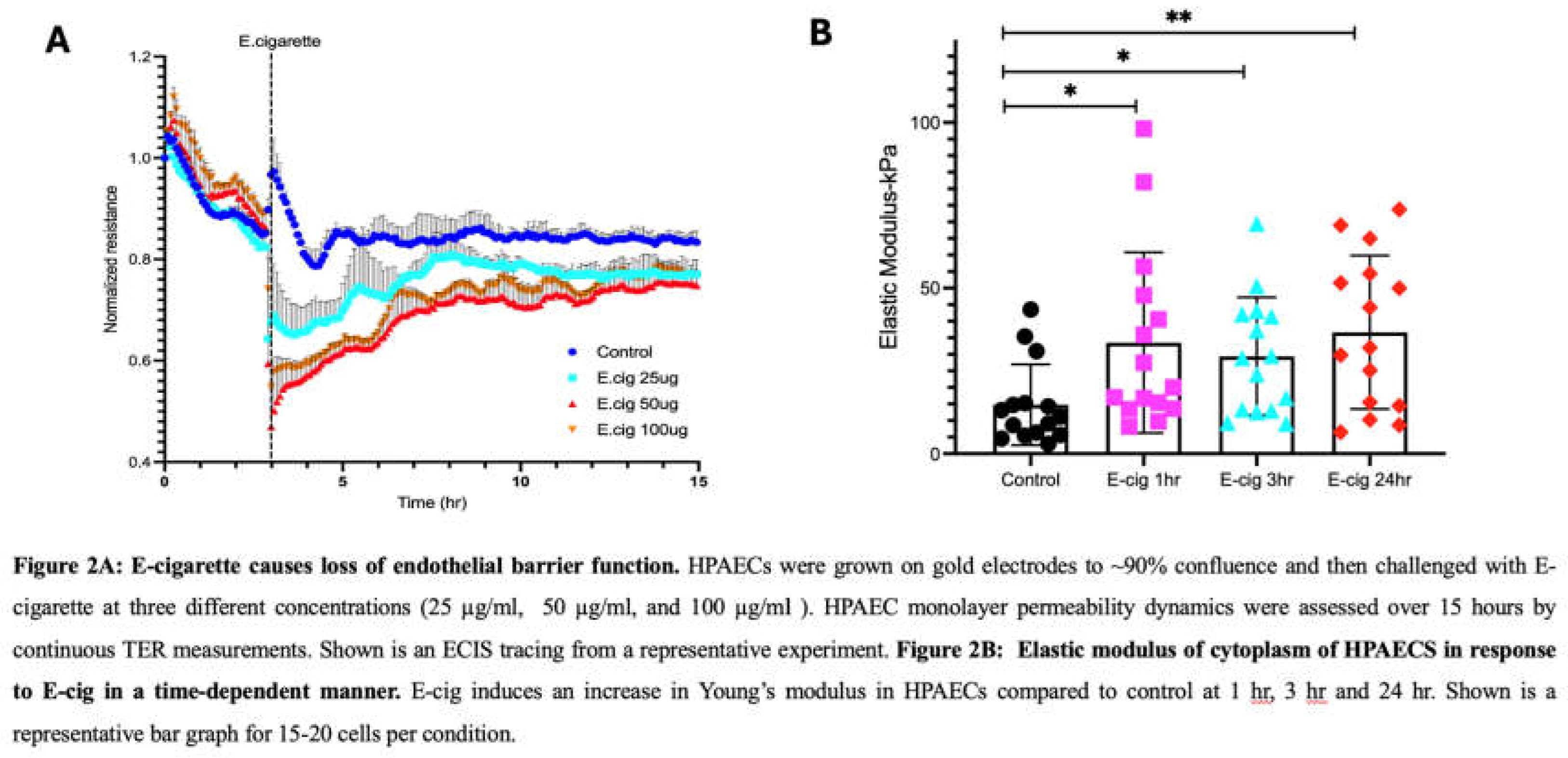
Trans-endothelial electrical resistance (TER) measurements were obtained to investigate the effects of E-cig on EC barrier function, a primary physiologic role for the endothelium [17,28]. Dose-dependent E-cig exposures resulted in prolonged disruption of EC barrier function with exposure to 25 µg/ml or higher of E-cig (Figure 2A).
3.3. Elastic Modulus Magnitude Is Increased in Lung ECs by E-Cig Exposure
We next characterized the mechanical properties of EC via AFM. Sub-confluent EC were first imaged using AFM contact mode, and then force curves were subsequently generated. AFM analyses of untreated cells were used as controls and compared to EC exposed to E-cig in a time-dependent manner (1 hr, 3 hr, and 24 hr). AFM measurements show that E-cig exposure significantly increased cell stiffness of ECs as compared to controls without E-cig exposure \(Figure 2B). For the maximum loads applied during our experiments (~2nN), indentation made on ECs by the AFM tip was typically ~200-600 nm.
3.4. Cortactin Expression Modulates the Lung Barrier Effects of E-Cigarettes
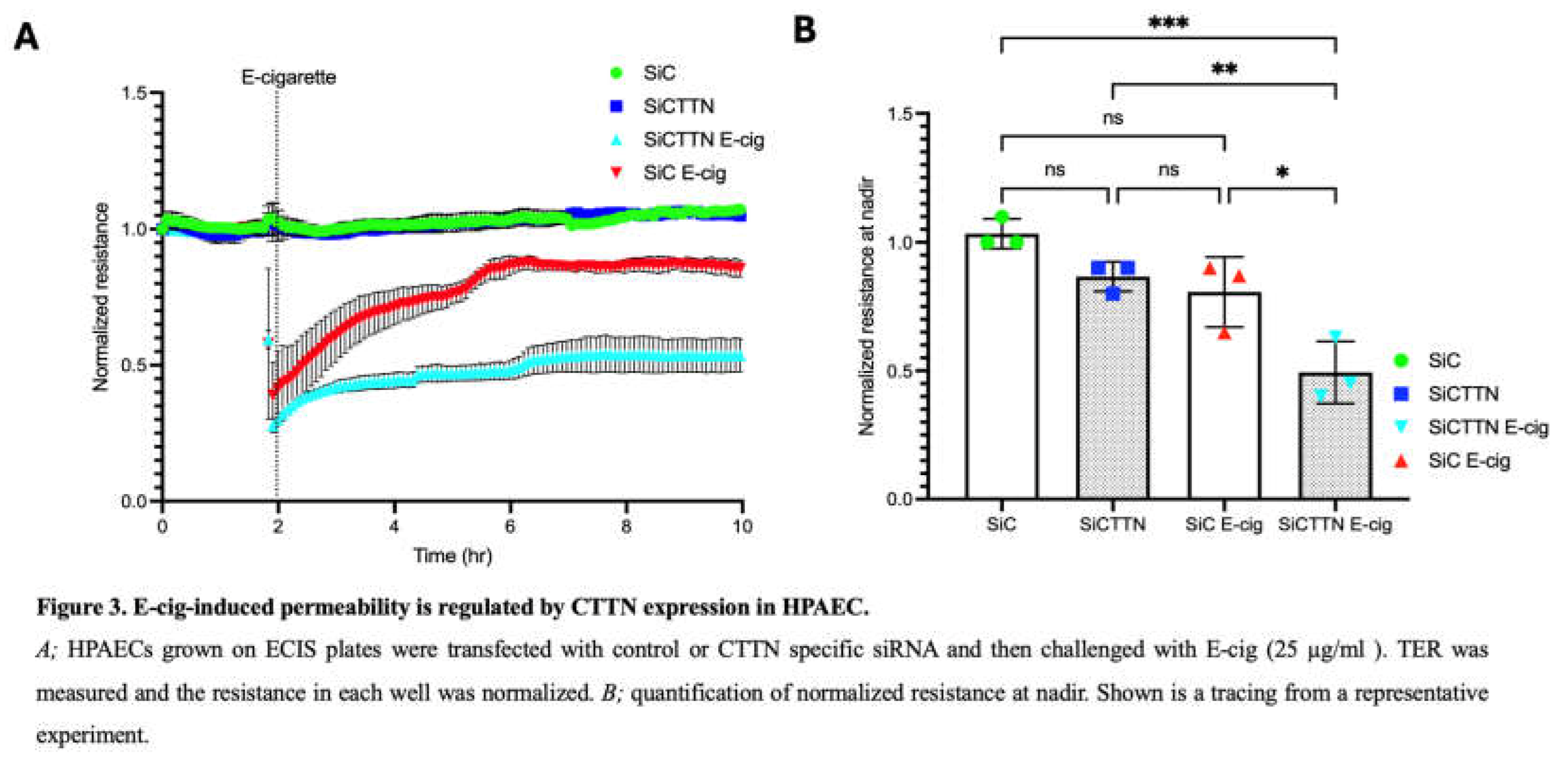
Lung EC permeability is regulated by CTTN expression and function [15,27,29,30,31]. To investigate its role in lung EC barrier integrity during E-cig exposure, ECs were transfected with control or CTTN siRNA for 48 hours and then seeded on TEER plates. Western blotting confirmed significant reduction in protein expression of CTTN via siRNA (data not shown). TEER measurements demonstrated that E-cig-induced barrier disruption is significantly increased in ECs treated with CTTN siRNA (Figure 3). These data support an important role for CTTN expression in attenuating EC barrier disruption caused by E-cig.
3.5. Role of MitoROS in E-Cig-Induced Lung EC Permeability
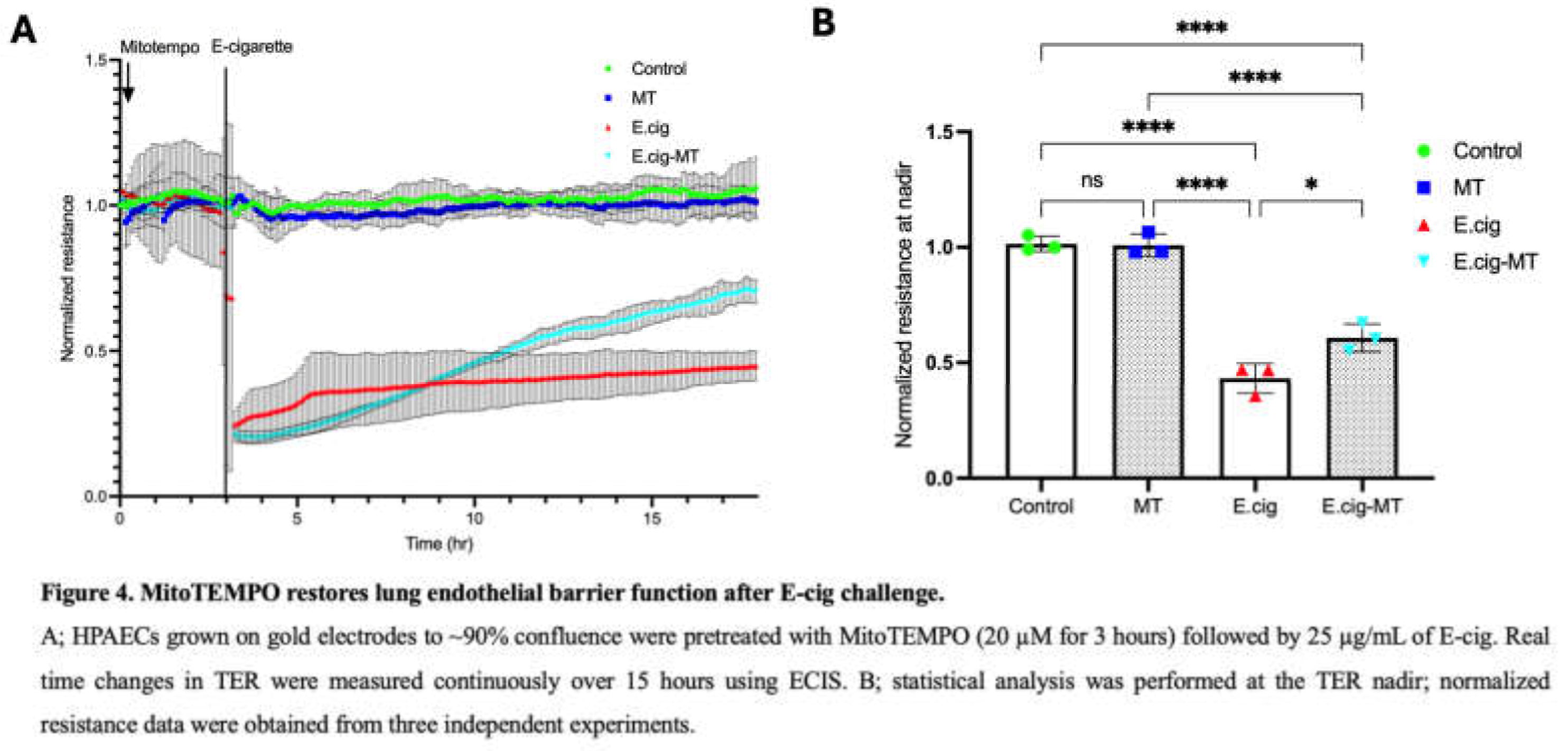
Our recent report demonstrated a novel role for CTTN in mitoROS generation and lung endothelial apoptosis [11]. MitoTEMPO is a scavenger of mitoROS and an effective tool for assessing its functional significance [32]. Here we investigated the role of mitoROS in E-cig-induced lung EC permeability by pretreating ECs with MitoTEMPO (20 µM for 3 hrs) prior to E-cig challenge (25 µg/ml). TER measurements demonstrated partial attenuation of E-cig-induced barrier disruption in EC pretreated with MitoTEMPO, confirming an important functional role for mitoROS in E-cig-induced barrier disruption (Figure 4).
3.6. Mitochondrial ROS Participates in E-Cig-Induced Elastic Modulus Changes in Lung ECs
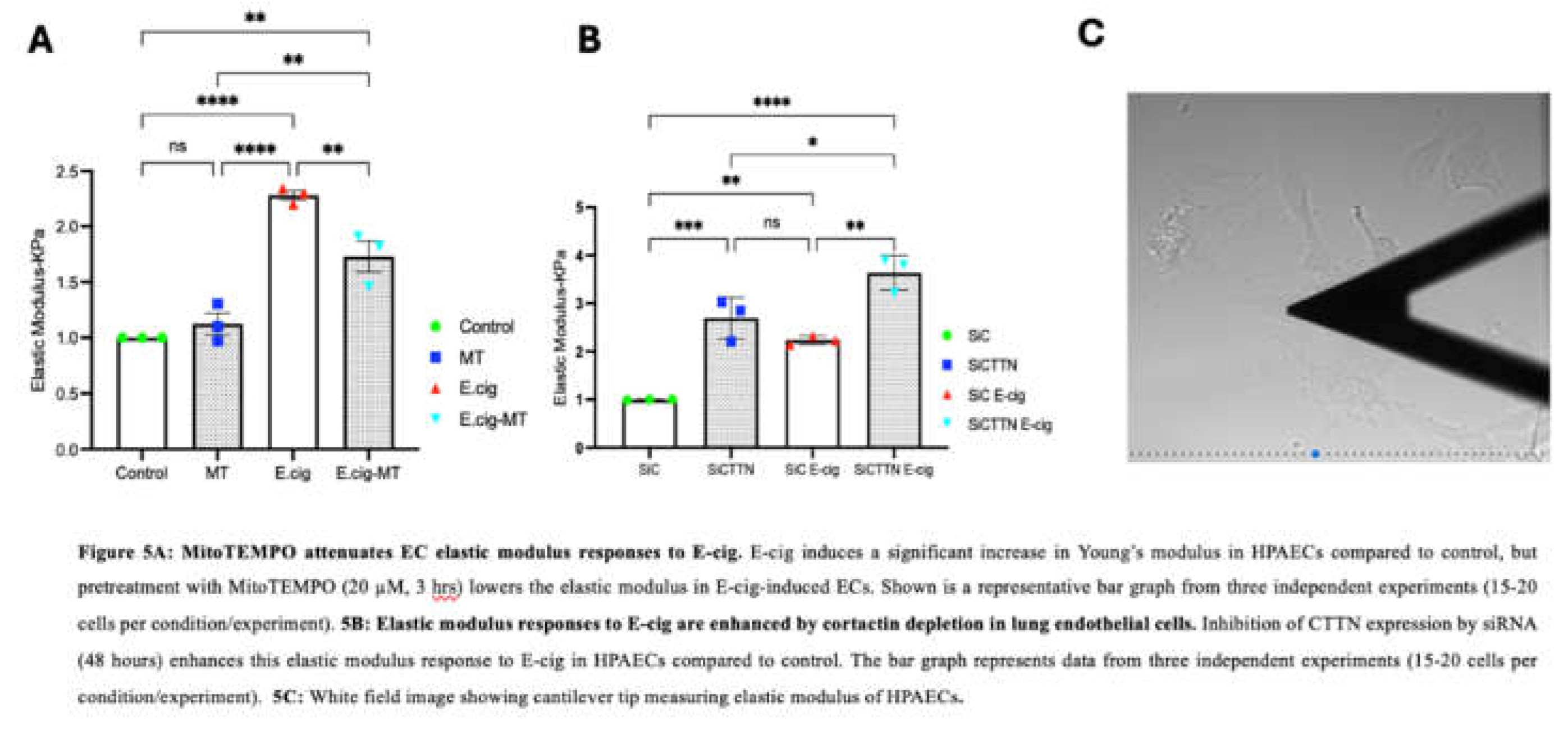
We next characterized the role of mitochondrial ROS in EC nanomechanical responses to E-cig using AFM. Sub-confluent EC were first imaged using contact mode, and then force-distance maps were subsequently generated. AFM analyses of untreated cells were used as controls and compared to EC pretreated with MitoTEMPO (20 µm, 3 h) and then challenged with E-cig for 1hr. AFM measurements were used to generate elastic maps to determine Youngs modulus. For maximum loads applied during our experiments (~2nN), deformation of the ECs by the AFM tip was typically ~200-600 nM. EC elastic modulus was significantly increased by E-cig exposure, but this increase was significantly attenuated in EC pretreated with MitoTEMPO, supporting an important functional role for mitochondrial ROS in E-cig-induced EC nanomechanical responses (Figure 5A).
3.7. CTTN Expression Regulates E-Cig-Induced Elastic Modulus Responses in Lung Endothelial Cells
We further explored the role of CTTN in E-cig-induced elastic modulus responses using AFM. EC were transfected with control or CTTN-specific siRNA for 48 h and then were challenged with E-cig for 1 h. Youngs modulus was then determined as above. Elastic modulus was again increased upon E-cig challenge. This increase was significantly enhanced in siCTTN cells, further demonstrating an important role for CTTN in regulating E-cig-induced EC nanomechanical responses (Figure 5B).
4. Discussion
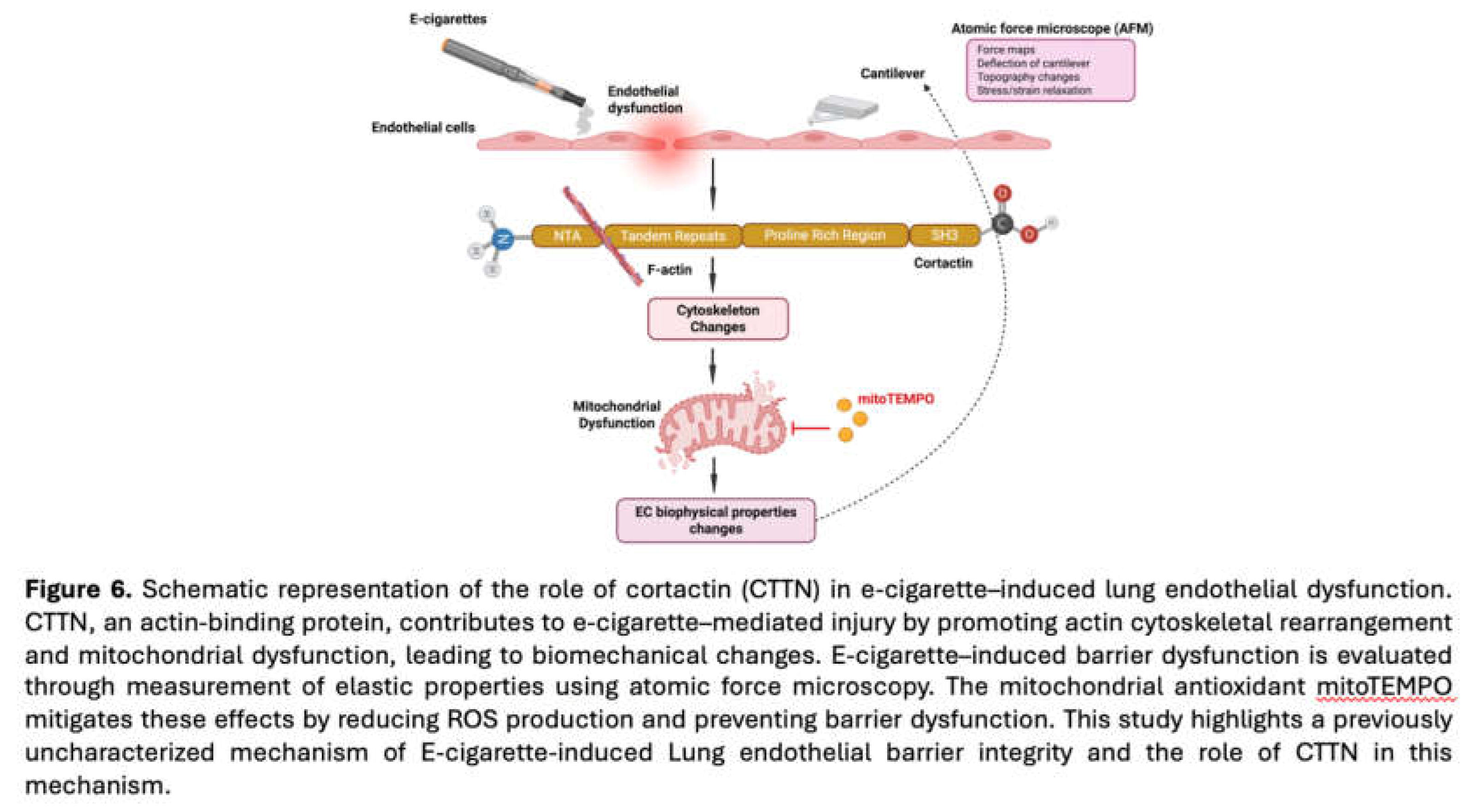
In 2019 the CDC/FDA declared an E-cigarette epidemic due to a national outbreak of E-cigarette-induced acute lung injury cases [E-cig or Vaping product use-associated acute lung injury (EVALI)] [33]. The underlying mechanisms by which E-cig contribute to lung disease remain incompletely understood, but it is likely that pathophysiologic effects on lung endothelial function are mechanistically involved. However, the effects of E-cig on lung ECs are only now beginning to be explored. Cortactin (CTTN) is an actin-binding protein that regulates cytoskeletal dynamics such as cell migration, adhesion, and apoptosis [11,13,15,34]. It plays a critical role in vascular integrity and has been studied in various diseases such as cancer and both acute and chronic lung disorders [21]. The potential mechanistic association between CTTN and E-cig-induced endothelial permeability is of particular interest given the essential barrier role of the lung endothelium. In our previous work, we demonstrated that CTTN plays a critical role in regulating lung endothelial apoptosis and mitochondrial ROS. Specifically, silencing CTTN or blocking its SH3 domain significantly increased endothelial cell apoptosis, providing mechanistic insights into COPD pathogenesis [11,15]. Building on these findings, the current study integrates atomic force microscopy (AFM), transendothelial electrical resistance (TEER) permeability assays, and immunofluorescence imaging to examine how E-cigarette exposure alters cytoskeletal organization, cell–cell interactions, and the biomechanical properties of endothelial cells (Figure 6). The major findings of this study are i) E-cig induces loss of cell-cell interaction causing gap formation in ECs, ii) E-cig causes loss of endothelial barrier function and increases elastic modulus (Y), indicative of increased cellular stiffness, iii) EC “Y” is attenuated by pretreatment with MitoTEMPO (MT), iv) EC “Y” is enhanced in CTTN-silenced EC during E-cig challenge, v) Pretreatment with MT protects the endothelial barrier during E-cig challenge, and vi) CTTN reduction by siRNA increases barrier disruption after E-cig challenge.
Mechanical properties regulate cell development and function, and the relationship between force generation and cell architecture is an emerging area for providing fundamental insights [35,36]. The current study focuses on exploring the role of CTTN structure/function (Figure 3 and Figure 5B) in modulating nanomechanical and rheological responses during E-cig-induced lung EC dysfunction [24,34]. External mechanical forces impact EC permeability through cytoskeleton rearrangement, junctional complex disassembly, and altered actomyosin contractility [17]. Here we identify that E-cig exposure leads to loss of cell-cell interaction and increases in EC permeability (Figure 1 and Figure 2). To further investigate the role of CTTN and mitoROS in E-cig-induced lung EC dysfunction, AFM was utilized to characterize biomechanical properties in lung EC after manipulation of CTTN expression/function (via siRNA) and ROS inhibitor (MitoTEMPO). The silicon nitride cantilever used for these AFM measurements assesses the rheological property of EC in a minimally-invasive manner [37]. We employed AFM contact mode to generate force maps to calculate Young’s modulus and determine topographical changes in lung EC upon E-cig or CSC challenge. We hypothesized that reducing CTTN by siRNA, or inhibition of mitoROS generation with MitoTEMPO, alters E-cig responses in lung EC in a manner that contributes to our understanding of their pathogenetic effects.
Our data indicate E-cig exposure increases the elastic modulus in lung ECs (as estimated with the Hertzian model) compared to vehicle controls. Force-distance mapping assesses the deflection of the AFM cantilever as it approaches the sample, demonstrating that cantilever deflection is lower in E-cig compared to vehicle (Figure 5A–C). Reduction of CTTN by siRNA results in an increase in the elastic modulus, and this effect is accentuated further upon E-cig exposure (Figure 5B). In contrast, scavenging mitoROS with MitoTEMPO ameliorates this E-cig effect on elastic modulus levels, suggesting a functional role in E-cig induced lung endothelial effects (Figure 5A). Elasticity is defined as a substance’s ability to resist deformation. To complement our AFM data, we measured trans-endothelial resistance (TEER) using Electric Cell-substrate Impedance Sensing (ECIS) to demonstrate that E-cig exposure reduces EC barrier function in a dose-dependent fashion when compared to vehicle (Figure 2). Reduction in CTTN expression by siRNA enhanced this pathophysiologic disruption of EC permeability by E-cig (Figure 3), while MitoTEMPO inhibition of mitoROS resulted in some EC barrier protection [32,38,39] (Figure 4).
In summary, these data indicate that E-cig exposure induces a series of pathophysiologic events in lung EC, including increased elastic modulus, loss of intercellular interaction, decreased resistance, and increased EC permeability (Figure 6). Mechanistically, these responses are modulated in part by CTTN expression and mitoROS generation. These observations add to our previous work describing CSC- and E-cig-induced apoptotic changes [11] and further support an important functional role for CTTN in mediating these responses. In addition, prior reports have demonstrated spatially-specific alterations of the EC cytoskeleton in response to various agonists, with differential effects at the cell periphery, cytoplasm and nucleus [9]. Further studies are therefore needed to measure the E-cig-induced deformations at different subcellular regions such as nucleus, cytoplasm, and periphery to better understand the nanomechanical properties within the cell. Next steps also may include CTTN overexpression studies to determine if this approach is protective against E-cig-induced EC dysfunction. In vivo correlation is likewise needed and may include the use of genetically modified CTTN mice and AFM measurements of lung tissue after E-cigarette exposure. Future work may focus on assessing other EC biomechanical properties with AFM such as height and other morphometric changes after E-cig exposure. These assessments can determine EC shrinking and loss of cell-cell interaction. Direct comparisons between the effects of CSC and E-cig also can be made. Finally, the potential signaling connection between CTTN structure/function and mitoROS needs further characterization.
Author Contributions
Conceptualization: SMD, JGN, MB. Methodology: MB, XG, JL. Data acquisition and investigation: MB. Data analysis: MB. Writing—original draft preparation: MB. Writing—review and editing: MB, SMD., JGN, JL. Reagents: SMD, JGN. Supervision: SMD; JL. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
This research was funded by American Heart Association 829814 (M.B.), NIH P01 HL126609 (J.G.N.G.), and NIH R01 HL133059 (S.M.D.).
References
- Dockrell, M.; Morrison, R.; Bauld, L.; McNeill, A. E-Cigarettes: Prevalence and Attitudes in Great Britain. Nicotine Tob. Res. Off. J. Soc. Res. Nicotine Tob. 2013, 15, 1737–1744. [Google Scholar] [CrossRef] [PubMed]
- Hajek, P.; Etter, J.-F.; Benowitz, N.; Eissenberg, T.; McRobbie, H. Electronic Cigarettes: Review of Use, Content, Safety, Effects on Smokers and Potential for Harm and Benefit. Addict. Abingdon Engl. 2014, 109, 1801–1810. [Google Scholar] [CrossRef]
- Bizoń, M.; Maciejewski, D.; Kolonko, J. E-Cigarette or Vaping Product Use-Associated Acute Lung Injury (EVALI) as a Therapeutic Problem in Anaesthesiology and Intensive Care Departments. Anaesthesiol. Intensive Ther. 2020, 52, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Morjaria, J.B.; Mondati, E.; Polosa, R. E-Cigarettes in Patients with COPD: Current Perspectives. Int. J. Chron. Obstruct. Pulmon. Dis. 2017, 12, 3203–3210. [Google Scholar] [CrossRef]
- Kalininskiy, A.; Bach, C.T.; Nacca, N.E.; Ginsberg, G.; Marraffa, J.; Navarette, K.A.; McGraw, M.D.; Croft, D.P. E-Cigarette, or Vaping, Product Use Associated Lung Injury (EVALI): Case Series and Diagnostic Approach. Lancet Respir. Med. 2019, 7, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Chen, W.; Moshensky, A.; Shakir, Z.; Khan, R.; Crotty Alexander, L.E.; Ware, L.B.; Aldaz, C.M.; Jacobson, J.R.; Dudek, S.M.; Natarajan, V.; Machado, R.F.; Singla, S. Cigarette Smoke and Nicotine-Containing Electronic-Cigarette Vapor Downregulate Lung WWOX Expression, Which Is Associated with Increased Severity of Murine Acute Respiratory Distress Syndrome. Am. J. Respir. Cell Mol. Biol. 2021, 64, 89–99. [Google Scholar] [CrossRef]
- Khalil, C.; Chahine, J.B.; Haykal, T.; Al Hageh, C.; Rizk, S.; Khnayzer, R.S. E-Cigarette Aerosol Induced Cytotoxicity, DNA Damages and Late Apoptosis in Dynamically Exposed A549 Cells. Chemosphere 2021, 263, 127874. [Google Scholar] [CrossRef]
- Anderson, C.; Majeste, A.; Hanus, J.; Wang, S. E-Cigarette Aerosol Exposure Induces Reactive Oxygen Species, DNA Damage, and Cell Death in Vascular Endothelial Cells. Toxicol. Sci. Off. J. Soc. Toxicol. 2016, 154, 332–340. [Google Scholar] [CrossRef]
- Wang, X.; Bleher, R.; Brown, M.E.; Garcia, J.G.N.; Dudek, S.M.; Shekhawat, G.S.; Dravid, V.P. Nano-Biomechanical Study of Spatio-Temporal Cytoskeleton Rearrangements That Determine Subcellular Mechanical Properties and Endothelial Permeability. Sci. Rep. 2015, 5, 11097. [Google Scholar] [CrossRef]
- Birukova, A.A.; Arce, F.T.; Moldobaeva, N.; Dudek, S.M.; Garcia, J.G.N.; Lal, R.; Birukov, K.G. Endothelial Permeability Is Controlled by Spatially Defined Cytoskeletal Mechanics: Atomic Force Microscopy Force Mapping of Pulmonary Endothelial Monolayer. Nanomedicine Nanotechnol. Biol. Med. 2009, 5, 30–41. [Google Scholar] [CrossRef]
- Bandela, M.; Letsiou, E.; Natarajan, V.; Ware, L.B.; Garcia, J.G.N.; Singla, S.; Dudek, S.M. Cortactin Modulates Lung Endothelial Apoptosis Induced by Cigarette Smoke. Cells 2021, 10, 2869. [Google Scholar] [CrossRef]
- Lu, Q.; Gottlieb, E.; Rounds, S. Effects of Cigarette Smoke on Pulmonary Endothelial Cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 314, L743–L756. [Google Scholar] [CrossRef]
- Jacobson, J.R.; Dudek, S.M.; Singleton, P.A.; Kolosova, I.A.; Verin, A.D.; Garcia, J.G.N. Endothelial Cell Barrier Enhancement by ATP Is Mediated by the Small GTPase Rac and Cortactin. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 291, L289–L295. [Google Scholar] [CrossRef]
- Tomar, A.; Lawson, C.; Ghassemian, M.; Schlaepfer, D.D. Cortactin as a Target for FAK in the Regulation of Focal Adhesion Dynamics. PloS One 2012, 7, e44041. [Google Scholar] [CrossRef]
- Bandela, M.; Belvitch, P.; Garcia, J.G.N.; Dudek, S.M. Cortactin in Lung Cell Function and Disease. Int. J. Mol. Sci. 2022, 23, 4606. [Google Scholar] [CrossRef]
- Belvitch, P.; Htwe, Y.M.; Brown, M.E.; Dudek, S. Cortical Actin Dynamics in Endothelial Permeability. Curr. Top. Membr. 2018, 82, 141–195. [Google Scholar] [CrossRef]
- Arce, F.T.; Younger, S.; Gaber, A.A.; Mascarenhas, J.B.; Rodriguez, M.; Dudek, S.M.; Garcia, J.G.N. Lamellipodia Dynamics and Microrheology in Endothelial Cell Paracellular Gap Closure. Biophys. J. 2023, 122, 4730–4747. [Google Scholar] [CrossRef]
- Gavara, N.; Chadwick, R.S. Determination of the Elastic Moduli of Thin Samples and Adherent Cells Using Conical Atomic Force Microscope Tips. Nat. Nanotechnol. 2012, 7, 733–736. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Xia, Y.; Sandig, M.; Yang, J. Characterization of Cell Elasticity Correlated with Cell Morphology by Atomic Force Microscope. J. Biomech. 2012, 45, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Cross, S.E.; Jin, Y.-S.; Rao, J.; Gimzewski, J.K. Nanomechanical Analysis of Cells from Cancer Patients. Nat. Nanotechnol. 2007, 2, 780–783. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, H.; Condeelis, J. Regulation of the Actin Cytoskeleton in Cancer Cell Migration and Invasion. Biochim. Biophys. Acta 2007, 1773, 642–652. [Google Scholar] [CrossRef]
- Lekka, M.; Gil, D.; Pogoda, K.; Dulińska-Litewka, J.; Jach, R.; Gostek, J.; Klymenko, O.; Prauzner-Bechcicki, S.; Stachura, Z.; Wiltowska-Zuber, J.; Okoń, K.; Laidler, P. Cancer Cell Detection in Tissue Sections Using AFM. Arch. Biochem. Biophys. 2012, 518, 151–156. [Google Scholar] [CrossRef]
- Iyer, S.; Gaikwad, R.M.; Subba-Rao, V.; Woodworth, C.D.; Sokolov, I. Atomic Force Microscopy Detects Differences in the Surface Brush of Normal and Cancerous Cells. Nat. Nanotechnol. 2009, 4, 389–393. [Google Scholar] [CrossRef]
- Htwe, Y.M.; Wang, H.; Belvitch, P.; Meliton, L.; Bandela, M.; Letsiou, E.; Dudek, S.M. Group V Phospholipase A2 Mediates Endothelial Dysfunction and Acute Lung Injury Caused by Methicillin-Resistant Staphylococcus Aureus. Cells 2021, 10, 1731. [Google Scholar] [CrossRef] [PubMed]
- Le Master, E.; Paul, A.; Lazarko, D.; Aguilar, V.; Ahn, S.J.; Lee, J.C.; Minshall, R.D.; Levitan, I. Caveolin-1 Is a Primary Determinant of Endothelial Stiffening Associated with Dyslipidemia, Disturbed Flow, and Ageing. Sci. Rep. 2022, 12, 17822. [Google Scholar] [CrossRef]
- Sun, X.; Sun, B.; Sammani, S.; Dudek, S.M.; Belvitch, P.; Camp, S.M.; Zhang, D.; Bime, C.; Garcia, J.G.N. Genetic and Epigenetic Regulation of Cortactin (CTTN) by Inflammatory Factors and Mechanical Stress in Human Lung Endothelial Cells. Biosci. Rep. 2024, 44, BSR20231934. [Google Scholar] [CrossRef] [PubMed]
- Belvitch, P.; Dudek, S.M. Role of FAK in S1P-Regulated Endothelial Permeability. Microvasc. Res. 2012, 83, 22–30. [Google Scholar] [CrossRef]
- Wang, L.; Dudek, S.M. Regulation of Vascular Permeability by Sphingosine 1-Phosphate. Microvasc. Res. 2009, 77, 39–45. [Google Scholar] [CrossRef]
- Rizzo, A.N.; Belvitch, P.; Demeritte, R.; Garcia, J.G.N.; Letsiou, E.; Dudek, S.M. Arg Mediates LPS-Induced Disruption of the Pulmonary Endothelial Barrier. Vascul. Pharmacol. 2020, 128–129, 106677. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Letsiou, E.; Wang, H.; Belvitch, P.; Meliton, L.N.; Brown, M.E.; Bandela, M.; Chen, J.; Garcia, J.G.N.; Dudek, S.M. MRSA-Induced Endothelial Permeability and Acute Lung Injury Are Attenuated by FTY720 S-Phosphonate. Am. J. Physiol. Lung Cell. Mol. Physiol. 2022, 322, L149–L161. [Google Scholar] [CrossRef]
- Mitra, S.; Epshtein, Y.; Sammani, S.; Quijada, H.; Chen, W.; Bandela, M.; Desai, A.A.; Garcia, J.G.N.; Jacobson, J.R. UCHL1, a Deubiquitinating Enzyme, Regulates Lung Endothelial Cell Permeability in Vitro and in Vivo. Am. J. Physiol. Lung Cell. Mol. Physiol. 2021, 320, L497–L507. [Google Scholar] [CrossRef]
- Suryadevara, V.; Huang, L.; Kim, S.-J.; Cheresh, P.; Shaaya, M.; Bandela, M.; Fu, P.; Feghali-Bostwick, C.; Di Paolo, G.; Kamp, D.W.; Natarajan, V. Role of Phospholipase D in Bleomycin-Induced Mitochondrial Reactive Oxygen Species Generation, Mitochondrial DNA Damage, and Pulmonary Fibrosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 317, L175–L187. [Google Scholar] [CrossRef] [PubMed]
- Layden, J.E.; Ghinai, I.; Pray, I.; Kimball, A.; Layer, M.; Tenforde, M.W.; Navon, L.; Hoots, B.; Salvatore, P.P.; Elderbrook, M.; Haupt, T.; Kanne, J.; Patel, M.T.; Saathoff-Huber, L.; King, B.A.; Schier, J.G.; Mikosz, C.A.; Meiman, J. Pulmonary Illness Related to E-Cigarette Use in Illinois and Wisconsin - Final Report. N. Engl. J. Med. 2020, 382, 903–916. [Google Scholar] [CrossRef] [PubMed]
- Birukov, K.G.; Birukova, A.A.; Dudek, S.M.; Verin, A.D.; Crow, M.T.; Zhan, X.; DePaola, N.; Garcia, J.G.N. Shear Stress-Mediated Cytoskeletal Remodeling and Cortactin Translocation in Pulmonary Endothelial Cells. Am. J. Respir. Cell Mol. Biol. 2002, 26, 453–464. [Google Scholar] [CrossRef]
- Malek, A.M.; Izumo, S. Mechanism of Endothelial Cell Shape Change and Cytoskeletal Remodeling in Response to Fluid Shear Stress. J. Cell Sci. 1996, 109 Pt 4, 713–726. [Google Scholar] [CrossRef]
- Janmey, P.A.; Miller, R.T. Mechanisms of Mechanical Signaling in Development and Disease. J. Cell Sci. 2011, 124 Pt 1, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Kihara, T.; Nakamura, C.; Suzuki, M.; Han, S.-W.; Fukazawa, K.; Ishihara, K.; Miyake, J. Development of a Method to Evaluate Caspase-3 Activity in a Single Cell Using a Nanoneedle and a Fluorescent Probe. Biosens. Bioelectron. 2009, 25, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Bandela, M.; Suryadevara, V.; Fu, P.; Reddy, S.P.; Bikkavilli, K.; Huang, L.S.; Dhavamani, S.; Subbaiah, P.V.; Singla, S.; Dudek, S.M.; Ware, L.B.; Ramchandran, R.; Natarajan, V. Role of Lysocardiolipin Acyltransferase in Cigarette Smoke-Induced Lung Epithelial Cell Mitochondrial ROS, Mitochondrial Dynamics, and Apoptosis. Cell Biochem. Biophys. 2022, 80, 203–216. [Google Scholar] [CrossRef]
- Huang, L.S.; Kotha, S.R.; Avasarala, S.; VanScoyk, M.; Winn, R.A.; Pennathur, A.; Yashaswini, P.S.; Bandela, M.; Salgia, R.; Tyurina, Y.Y.; Kagan, V.E.; Zhu, X.; Reddy, S.P.; Sudhadevi, T.; Punathil-Kannan, P.-K.; Harijith, A.; Ramchandran, R.; Bikkavilli, R.K.; Natarajan, V. Lysocardiolipin Acyltransferase Regulates NSCLC Cell Proliferation and Migration by Modulating Mitochondrial Dynamics. J. Biol. Chem. 2020, 295, 13393–13406. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



