
Submitted:
07 October 2025
Posted:
08 October 2025
You are already at the latest version
Abstract
Phosphorus (P) is an essential nutrient that is required for plant growth and for the productivity and sustainability of global agriculture. Phosphorus deficiency is a major constraint for crop production and efficient phosphorus management is important to address food security efforts worldwide. Advances in plant genetics and breeding have led to promising avenues to enhance P-use efficiency (PUE), enabling crops to better acquire, mobilize, and utilize soil phosphorus. Strategies such as optimizing root architecture, fostering beneficial mycorrhizal associations, and targeting genes involved in P transport and remobilization have potential to sustainably increase crop productivity while minimizing external inputs. These improvements can support resilient farming systems adaptable to diverse environmental conditions.However, much of the existing research on genetic improvement in PUE has been conducted under controlled environments and in pot trials that may not be representative of realistic agricultural systems. Evaluation of the performance of genetic methods for improving PUE must be conducted using a range of diverse production practices under realistic field conditions. Breeding programs must balance P-related traits with other priorities such as yield, disease resistance, drought tolerance, and seed quality, which often extend the timeline for development. Benefits of these genetic improvements are region-specific and dependent on local soils, climate, and farming practices.While progress is encouraging, many genetic innovations are still in development and require translation into practical field applications. The most promising genetic approaches, including root system modification and seed phytate reduction, offer significant potential when integrated with optimal nutrient management strategies. Moving forward, combining advanced breeding tools, genomic technologies, and region-sensitive agronomic practices presents an excellent opportunity to achieve more sustainable and efficient phosphorus utilization in agriculture.
Keywords:
Phosphorus use efficiency (PUE)
; phosphorus uptake
; plant breeding
; sustainable agriculture
; nutrient management
; crop productivity
Section 1. Phosphorus in Plant Nutrition
Key Points
- Phosphorus is an essential plant nutrient required for energy transfers, photosynthesis, and cell division
- Plants have developed genetic strategies to access and efficiently utilize P
- Metabolically active P is a small proportion of the total plant P if P is not deficient
- Plant breeding may be able to improve phosphorus use efficiency by developing plants that are better able to access and utilize P
1.1. Introduction
Phosphorus is one of the 17 essential plant nutrients that are critical for a plant to grow and reproduce. It is one of the nutrients most commonly limiting for plant growth and reproduction in natural ecosystems. Therefore, plants have developed natural strategies to efficiently access and utilize P. These strategies are under genetic control and allow the plant to optimize P uptake and allocation to satisfy critical metabolic functions.
The concentration of P in plant tissue varies with crop type, part and maturity, but typically is in the range of 0.1 to 0.5% P. Phosphorus is essential for photosynthesis and for the energy reactions in the plant because it is a component of adenosine triphosphate (ATP), adenosine diphosphate (ADP), and nicotinamide adenine dinucleotide phosphate (NADP, NADPH, NADP+) (Glass et al. 1980; Hopkins 2015; Sultenfuss and Doyle 1999; Raven et al. 2005). During photosynthesis, light energy is absorbed by chlorophyll and stored in the high energy bonds in ATP. The NADPH and ATP formed using the energy captured during the light reactions are used to reduce carbon dioxide to carbohydrates during the dark reactions of photosynthesis.

In addition to its role in photosynthesis, ATP is also the primary molecule for energy storage and transport in the plant. When the phosphate is transferred from the ATP to another molecule by hydrolytic enzymes, the high energy bond in the ATP is broken, and the chemical energy is released to drive energy-requiring reactions of plant metabolism.
Phosphorus also plays an important structural role in the plant. It is a critical component of the phospholipid bilayer of cell membranes and is essential for cellular reproduction and plant growth. Under P stress, plants may be able to substitute some of the P-containing phospholipids with galactolipids and sulfolipids that do not contain P, particularly in older tissue, to free up inorganic P (Pi) for other metabolic uses (Poirier et al. 2022; Navea et al. 2024). However, phospholipids are normally needed in young tissue during leaf growth and development to maintain membrane integrity during cell division, elongation, and expansion.
Phosphorus is also a structural component of the nucleic acids, including deoxyribonucleic acid (DNA) and ribonucleic acid (RNA), that make up genes and chromosomes, and a large proportion of the organic P in the plant is in the form of ribosomal RNA (Poirier et al. 2022; Veneklaas et al. 2012). These nucleic acids are required for the transfer of genetic information during cell division and reproduction and to enable protein synthesis. Phosphorus is a component of many coenzymes and phosphoproteins and of phosphate compounds that are intermediate metabolites in a wide range of metabolic processes (Raven et al. 2005). The level of inorganic P present in the cell plays a role in enzyme regulation and in the control of starch synthesis (Mills and Jones 1996). Dissociation of phosphoric acid is involved in the buffering of cellular pH and the maintenance of homeostasis (Mills and Jones 1996).
The importance of P in photosynthesis, all energy transfers, cell division and a wide range of metabolic processes means that P is required from the initial reactions in the germinating seed, throughout plant growth, to maturity. Each time a cell divides, P is required to provide energy for reactions, to replicate the genetic material that is passed to the new cell, to form the phospholipids of the cell membranes, and to produce a wide range of critical enzymes and other P-containing cellular components.
1.2. Accumulation of P by the Plant
Seeds contain a reserve of P stored primarily as phytic acid (Sparvoli and Cominelli 2015). For the first few days of growth, a plant may rely on its seed reserves to supply the P that it requires, but as the reserves are depleted, the plant must access P from the soil to maintain growth (Nadeem et al. 2011). An adequate supply of P is needed by the crop throughout its life cycle, but it is particularly important that the plant can access sufficient P during early growth to ensure optimum crop yield (Grant et al. 2001). If the plant can access more P than its current requirement, it will accumulate surplus P. The “luxury consumption” can be stored as inorganic phosphate in plant vacuoles as a reserve that can be mobilized to supply P if external supply becomes restricted (Yang et al. 2017; El Mazlouzi et al. 2020a; El Mazlouzi et al. 2020b).
Phosphorus in the grain will come from a combination of external sources or internal remobilization of P from plant organs, with 75 to 80% of the total plant P being present in the grain at maturity (El Mazlouzi et al. 2020a; El Mazlouzi et al. 2020b; Mohamed and Marshall 1979). Plants are very efficient at remobilizing the P from vegetative tissue to the seed, especially under P stress (Veneklaas et al. 2012) . Using 32P, it was found that durum wheat plants with low P status remobilized most of their stored P, resulting in elevated concentrations of P in the vegetative tissue (El Mazlouzi et al. 2020a; El Mazlouzi et al. 2020b). About 80% of the grain P came from remobilization of P in low P plants, while 65% came from remobilization in high P plants, with the remaining 20 to 35% coming from absorption between anthesis and maturity. Plant uptake of P can continue into the ripening phase, but the supply in the early growth stages strongly affects plant response (Malhi et al. 2006, 2007b, a). Therefore, early season limitations in P availability can result in restrictions in crop growth from which the plant will not recover, even when P supply is increased to adequate levels later in the season (Grant et al. 2001).
1.3. Phosphorus Deficiency Symptoms
Phosphorus deficiency symptoms are often subtle, and mild deficiencies may not produce obvious symptoms. Plants experiencing P deficiency may increase root production at the expense of the above-ground plant in an attempt to access more P from the soil (Malhotra et al. 2018; Hinsinger 2001). Phosphorus deficient plants may also increase the secretion of organic acids in an attempt to mobilize P. As P becomes more limiting, deficiency may interfere with metabolic pathways and cause carbohydrates or anthocyanins to accumulate, producing dark green or purple discoloration of the leaves and stems (Hopkins 2015; Hoppo et al. 1999; Close and Beadle 2003). Protein synthesis may also be impaired, so soluble N compounds accumulate in the tissue. The lack of proteins interferes with metabolic processes, reducing cell growth and leading to shorter plants, delayed leaf emergence, delayed development, reduced tillering and secondary root development, decreased dry matter yield, and reduced seed production (Elliott et al. 1997; Glass et al. 1980; Grant et al. 2001; Hoppo et al. 1999; Konesky et al. 1989). Plant maturity is delayed, and crop yield declines.
In response to P deficiency, plants attempt to increase the probability of producing at least some viable seed with the limited P supply. For example, in cereal crops under P stress, seed number will be reduced because the number of fertile tillers and the number of grains per tiller decrease, but seed size will be maintained (Hoppo et al. 1999; El Mazlouzi et al. 2020b). Similarly, in soybeans, P deficiency reduces the number of pods and seeds, but seed size is not affected, increasing the likelihood that the remaining seeds will be viable. The decrease in seed number leads to a decrease in final crop yield.
Sufficiency thresholds for P concentration in crop tissue can be used to determine if plant P supply is adequate. Deficiency thresholds will vary with crop, growth stage and tissue type, but plant tissue concentrations of P below 0.2% often indicate deficiency (Mills and Jones 1996). As the plant ages, the proportion of metabolically active tissue declines, and the amount of low-P structural tissue increases (Bélanger and Richards 1999; Elliott et al. 1997a; Racz et al. 1965), so the critical P concentration required in the tissue for optimum growth decreases as plants age (Elliott et al. 1997a; Elliott et al. 1997b; Tomasiewicz 2000).
As mentioned previously, P is generally mobile in the crop and will re-translocate from vegetative tissue to storage organs such as seeds (Veneklaas et al. 2012). As a result, most of the P taken up by the crop will be removed in the harvested material. Replacement of P removed in the harvest crop is important to maintain long-term soil productivity.
1.4. Summary
Phosphorus plays a critical role in crop physiology from the initial processes in the germinating seed, throughout plant growth, to maturity. If plants cannot access sufficient P, yield will decline. Because of the importance of P in physiology, plants have evolved strategies to increase their ability to access and efficiently utilize P for plant growth and reproduction. These strategies are under genetic control and may be factors that can be manipulated through plant breeding to improve phosphorus use efficiency in agricultural systems.
Section 2. Phosphorus Use Efficiency
Key Points
- Phosphorus use efficiency (PUE) can be measured through different methods that consider the short- and long-term use of P in the cropping system
- Selection of the most appropriate method of assessing phosphorus use efficiency depends on the goal
- Short-term assessments of PUE may miss legacy benefits from P applications
- Factors other than P supply that limit yield potential will decrease PUE
- Assessment of both fertility and physiologically based PUE can be important tools that plant breeders can use to improve the PUE of new cultivars
2.1. What is Phosphorus Use Efficiency?
Phosphorus use efficiency (PUE) refers to how well phosphorus is used by a crop. It is a critical aspect of P management as it influences both profitability for the producer and the potential for negative environmental impacts. One can consider PUE in terms of the ability of the plant to take up P from the soil or in terms of the ability of the plant to utilize the P that it takes up, also referred to as external and internal PUE, respectively (van de Wiel et al. 2016). However, there are different methods of viewing both external and internal PUE with different implications for productivity, economics of production and environmental impact.
2.2. Fertility Based Measurements of Phosphorus Use Efficiency
Crop recovery of added fertilizer P can be measured directly by using 32 P, but this is effective primarily for short-term studies due to the limited half-life of the isotope. More commonly, crop recovery efficiency (RE) of P is measured by the difference method, where the P taken up by the unfertilized crop (U0) is subtracted from the P taken up by the fertilized crop (UP) and divided by the amount of fertilizer P applied (FP), then expressed as a percentage (Roberts and Johnston 2015).
RE = (Up− U0 / Fp) x 100
Values for RE in the first growing season after applying P fertilizers are often very low, in the 10-15% range, meaning that most of the P that is present in the crop comes from soil P reserves or legacy P that has accumulated from previous P inputs. Values for RE will tend to be low if soil P reserves are high.
In addition to being used directly, fertilizer P can also be used by the crop after it cycles through the soil reserves. Therefore, much of the fertilizer applied in a particular year serves to replenish the reserves that are being used. The RE measures only the short-term benefit of the fertilizer and does not consider the legacy benefit of P fertilizer. As such, it can underestimate the benefits derived from fertilization.
An alternate method of assessing P use efficiency that considers the use of this “legacy P” is the partial nutrient balance (PNB) method, or the balance method (Syers et al. 2008).
PNB=Up/FP
The PNB method is the ratio of the P removed by the crop (Up) relative to the P fertilizer input (Fp). If more P is removed than applied, the P in the available or sparingly available pools will be depleted over time. If more P is applied than removed, the P in the pools will increase over time. The PNB is often in the range of 50-70%. If the PNB is very low over the long-term, the P is being used inefficiently, and the management practices being used should be reviewed to determine if efficiency can be improved. To be meaningful, PNB should be assessed over several years to determine long-term trends.
If P input and offtake are nearly balanced, plant-available P as measured by a soil test will be relatively stable over time, and the efficiency of P use will be relatively high. One fertilizer management strategy, therefore, would be to regularly assess the soil test levels. The critical level for crop growth is often viewed as the soil test level above which applications of P fertilizer do not provide a yield response (Johnston et al. 2014; Roberts and Johnston 2015). If soil test levels are substantially below the critical level, P should be added at higher than removal levels to build soil test P and improve the soil P status. If the P level is above the critical level (as may occur on manured soils), P should be applied at less than removal levels to draw down the soil P. If the P in the soil is near an acceptable moderate level, P should be applied at approximately removal values to keep soil P in the desired range.
For a producer, the impact that a fertilizer application has on crop yield and profitability is often the most relevant consideration. This is measured by Partial Factor Productivity (PFP) which refers to the units of crop yield (Y) produced per unit of nutrient applied (Fp) (Roberts and Johnston 2015; Fixen et al. 2015). The PFP will be strongly affected by the overall productivity of the system, so factors that increase crop yield will tend to increase measured PFP. On the other hand, if crop yield is limited by factors other than P, the PFP will tend to be low.
PFP=Y/Fp
Agronomic efficiency (AE) addresses the question of how much agronomic benefit was gained by applying the fertilizer. It is calculated as the difference in yield between the fertilized (Yp) and unfertilized crop (Y0), divided by the fertilizer applied (F).
AE=(Yp-Y0)/F
If soils have high levels of available P, approaching or exceeding the critical soil test value, crop demand for P may be satisfied from the soil reserves. Therefore, at high soil test levels, yield response to P and the AE will be both be low. Similarly, if yield is limited by factors other than P supply, for example by drought or frost, the yield response to fertilizer application will be low and AE will decline.
Return on investment, (ROI) measures the economic benefit of fertilizer application. It can be calculated as the difference in yield of the fertilized crop (Yp) and the unfertilized crop (Y0) multiplied by the price of the crop divided by all costs associated with applying the fertilizer.
ROI=(Yp-Y0)*crop price/fertilizer cost
The return on investment is often of greatest importance to a producer. The ROI can be considered on an annual basis, but the legacy benefits of P application are only included if the ROI is calculated over the long term.
2.3. Physiologically Based Measurement of Phosphorus Use Efficiency
Internal Utilization Efficiency (IUE) describes the ability of the crop to convert the nutrients that it accumulates into yield. It is calculated as yield (Y) divided by nutrient uptake (U). This provides information on the genetic ability of a crop to convert P into marketable yield, but will also be affected by environmental conditions that restrict yield potential.
IEF=Y/U
Physiological efficiency (PE) describes the ability of the plant to transform nutrients supplied by the fertilizer into yield. It is calculated as the difference between yield in the fertilized (Yp) and unfertilized (Y0) crop divided by the P uptake in the fertilized (Up) and unfertilized (U0) crop.
PE=(Yp-Y0)/(Up-U0)
As with the IEF, this provides information on the genetic capacity of a plant to respond to fertilizer application but is also affected by environmental conditions.
Photosynthetic phosphorus-use efficiency (PPUEmax) is the instantaneous light-saturated rate of leaf photosynthesis per unit leaf P (Veneklaas et al. 2012; Hidaka and Kitayama 2013; Lambers 2022). It is calculated as the ratio of mass-based maximum photosynthetic carbon assimilation rate (Amass) per concentration of total foliar P, or as the ratio of area-based maximum photosynthetic carbon assimilation rate (Aarea) per area-based content of total foliar P. The PPUEmax is an indication of the ability of the plant to sustain photosynthetic activity with limited P supply.
PPUEmax=Amass/Pfoliar
2.4. Interpretation of Phosphorus Use Efficiency
The selection of the most appropriate method of assessing phosphorus use efficiency depends on the goal. The RE provides a short-term assessment of P fertilizer recovery in the year of application. It is useful for comparing relative efficiencies of various fertilizer products in the year of application, but does not consider long-term benefits or effects on residual soil P. The PNB considers the P that is taken up by the crop from both the soil reserves and the applied fertilizer. Long-term measurement of PNB provides information on long-term fertilizer use efficiency and on the potential depletion or accumulation of legacy P reserves in the soil.
The PFP assesses the yield produced per unit of P applied and is useful as an indicator of the productivity of the system. It will decrease if factors other than P supply are limiting to crop yield. Agronomic efficiency and return on investment provide an assessment of the short-term effect of fertilizer applications on crop productivity and economic return. This would be useful for producers to determine the short-term benefits of fertilization, especially in years where financial margins are tight or if the land tenure was short-term. However, if P application is lower than P removal, the short-term ROI may be high but decreases in long-term soil productivity due to P depletion may decrease future economics of production.
From a plant breeding perspective, both fertility-based and physiological meaures of PUE may be very important. Fertility-based PUE assessment values can provide a broad assessment of the performance of specific breeding lines to respond to soil P supply, while assessement of PE, IEF or PPUEmax values can identify the genetic capacity of breeding lines to convert available P or fertilizer P into yield. Understanding the physiological and genetic basis for higher PUE can help the breeder determine the feasibilty of incorporating specific PUE traits into the selection program.
2.5. Summary
Phosphorus use efficiency can be measured through different methods that consider the short- and long-term use in the cropping system. If crop productivity is limited by factors other than P supply, PUE will decline. Improvements in phosphorus use efficiency can contribute to long-term sustainability of agricultural production. In a breeding program, both fertility-based and physiologically-based assessments of PUE can be useful.
Section 3. Description of Genetic Techniques in Plant Breeding
Key Points
- Plant breeding has improved crop yield, agronomic characteristics, disease resistance and nutritional quality
- Traditional plant breeding selected for superior physical characteristics that were present in the natural population
- The ability to rapidly assess the genetic composition of plants, to quickly measure physical traits and to analyse large amounts of data has improved the ability to breed for complex characteristics
- New molecular techniques can generate genetic diversity by moving genes between unrelated species or by precisely editing genes.
- Molecular techniques can allow the breeder to make selections early in the breeding process at the molecular, cellular, or tissue level
- Modern breeding techniques can shorten the time and reduce the costs for developing improved cultivars and breed for characteristics that are not normally found in the population
3.1. Introduction
People have been using plant breeding techniques to improve the crops they grow since agriculture began. Initially, plants were selected from natural populations for their desirable physical traits, such as yield, seed size or lodging resistance. Over the years, simple selection based on physical observation provided great improvements in crop yield and quality. The discovery of basic genetic principles of inheritance by Gregor Mendel in the 1800s allowed the application of science to plant breeding. The combination of genetic improvements and targeted agronomic practices during the “Green Revolution” greatly increased crop yield and provided a more stable food supply worldwide. More recently, improvements in the ability to quickly read the genetic code in plants, measure the physical characteristics of plants, and statistically interpret the huge amounts of data generated have greatly expanded the identification of specific genetic areas associated with desirable traits. These new techniques have the potential to create cultivars with greater phosphorus use efficiency.
3.2. Traditional Plant Breeding
Traditional plant breeding takes advantage of the natural variability in a plant population to produce new cultivars (cultivated varieties) containing desired characteristics. The techniques used will depend on whether a plant is cross-pollinated or self-pollinated (https://www.britannica.com/science/plant-breeding/Evaluation-of-plants, accessed on January 4, 2025).
A seed is produced when male pollen fertilizes a female egg cell. A self-pollinated plant is fertilized when pollen comes from any flower of the same plant, and a cross-pollinated crop is fertilized with pollen that comes from a flower on a different plant. Because a self-pollinated plant only has one parent, it produces a more uniform population of plants that are genetically similar or homozygous, meaning that they contain identical alleles on paired chromosomes for most traits. In contrast, a cross-pollinated plant has two parents that are likely to differ in many genes so cross-pollination will produce a more diverse population of plants that are heterozygous meaning that they contain different alleles on paired chromosomes for many traits.
Many cultivated plant species reproduce by self-pollination, including some of the most important cereal crops such as wheat, rice, barley and oat, many legume crops such as pea, peanut, chickpea, soybean and lentil, as well as other crops such as linseed, potatoes, lettuce and tomato (Akhtar et al. 2023). Cross-pollinated crops include maize, rye, sunflower, alfalfa, sugarcane, and many vegetable and tree crops. Some plants are considered partially or often cross-pollinated and these include sorghum, cotton, faba bean, tobacco, some Brassica species and safflower. Partially cross-pollinated crops are not completely self-pollinated, and their cross-pollination often exceeds 5% and may reach 50%. Their genetic architecture is between self-pollinated and cross-pollinated species.
3.2.1. Traditional Breeding in Self-Pollinated Crops
In self-pollinated crops, traditional plant breeding can use several techniques. These include: (1) mass selection, (2) pure-line selection, (3) hybridization, and (4) development of hybrid varieties (Akhtar et al. 2023).
Mass selection is likely the oldest plant breeding method used. Seeds are collected from a large population of plants with desirable traits, and their seeds are pooled together and sown. Plants with desirable traits are moved to the next generation, and/or plants with undesirable traits are destroyed. The selection is based on the plant appearance (i.e. phenotype) and is repeated over multiple generations to develop a population with the desired characteristics. Because the plants being selected are self-fertilized, the resulting offspring will be homozygous.
With pure-line selection, numerous superior plants are selected from a diverse population. The seeds of the individual plants are grown for several generations, and the most desirable progeny is selected each cycle and grown the following year. The process of growth and selection is repeated for several years. When the line appears to be stable and uniform, the plants are evaluated in field trials to see if they are superior to existing varieties.
Since the 20th century, hybridization has been the dominant method of plant breeding. Hybridization combines desirable genes by crossing two or more carefully selected parents to produce pure-breeding progeny that are superior to the parental lines. The plant breeder can use the pedigree, bulk, or the backcross methods to handle the selection process.
In the pedigree method, superior types are selected in successive generations, and a record of parent–progeny relationships is maintained. Pedigree breeding begins withcrossing of two parent lines with complementary characteristics desired for a new variety. This produces a heterozygous hybrid F1(Filial1) generation. For the F2 generation, the breeder may either cross two of the F1 progeny or may cross F1 progeny with a third parent line to bring in other important traits that were not present in the initial two parent lines. From the F2 generation onward, undesirable plants are eliminated from the population, and the most desirable plants are moved on to the next generation. From the F3 generation on, plants are self-pollinated and become increasingly homozygous. By F5, relatively homozygous families have segregated, and the most promising families are selected. By F7 or F8, a manageable number of homozygous lines have been selected, and the most desirable move is to conduct several years of field testing for yield, agronomic characteristics and quality.
In the bulk-population method, the F2 generation is sown at normal commercial planting rates in a large plot and harvested in bulk. The seeds are propagated in bulk for the next generation without tracking the ancestry. Natural selection eliminates plants with poor survival value, while directed selection is also undertaken to destroy plants that carry undesirable major genes. Other mass selection techniques may be used, such as harvesting early for early maturity, or inoculation with disease organisms to select for resistance. Single plant selections are then made and evaluated in the same way as in the pedigree breeding method. The bulk selection population method allows the breeder to handle very large numbers of plants more inexpensively than with the pedigree method.
Back-crossing is a more targeted method of pedigree selection. An outstanding cultivar may possess many desirable traits that are genetically complex, but may be lacking a specific required characteristic, such as disease resistance. In that case, a cross can be made between the outstanding cultivar and a donor plant that carries the required characteristic. Then, the progeny can be repeatedly crossed back to the outstanding parent and offspring selected at each stage that have retained the target characteristic from the donor parent. After five or six backcrosses, the progeny will be hybrid for the character being transferred but will be like the superior parent for all other genes. The progeny is then self-crossed, and the next generations selected for the desired characteristics to give progeny pure breeding for the genes being transferred. Back-crossing is relatively rapid, requires a relatively small number of plants, and is more predictable than less targeted methods.
The development of commercial hybrids attempts to take advantage of hybrid vigour, or heterosis, where heterozygous plants are stronger and more productive than the homozygous parent plants. With hybrid plants, the heterozygous F1 hybrid plants are the commercial outcome rather than a homozygous pure-breeding population.
Production of hybrid seed requires a male sterile parent that does not produce functional pollen. Male sterility can be genetic, cytoplasmic or cytoplasmic-genetic. A single recessive gene governs genetic male sterility and may be maintained by crossing it to heterozygous male fertile plants. Cytoplasmic male sterility is carried in the cytoplasm, which comes from the egg cell, so the progeny of such male sterile plants is always male sterile. It may be maintained in a variety by using that variety as a pollinator in six to seven successive generations of a backcrossing program. Cytoplasmic-genetic male sterility is a cytoplasmic male sterility where a dominant nuclear restorer gene is used to restore fertility in the male sterile line.
Hybrid seed is produced by interplanting a male sterile version of a pure line in isolation with another pure line that will produce pollen. All the seeds produced on the male sterile plant will be F1 hybrids of the two strains, and the F1 hybrid is the commercial cultivar. The breeder must develop the homozygous sterile and fertile breeding lines to use as parent lines for hybrid production.
3.2.2. Traditional Breeding in Cross-Pollinated Crops
Cross-pollinated crops may be hybrids, open-pollinated crops or synthetic varieties. Cross-pollinated hybrids consist of one genotype while open-pollinated and synthetic varieties will contain many genotypes. As opposed to the homozygous self-pollinated plants, cross-pollinated plants are highly heterozygous, and their vigour and yield may decline with inbreeding. The breeding methods used need to preserve or restore heterozygosity in the final variety to avoid inbreeding depression.
Breeding of cross-pollinated species can use mass selection, development of hybrid varieties, and development of synthetic varieties.
Mass selection in cross-pollinated species follows the same pattern as with self-pollinated species. A large population of superior plants are selected and harvested in bulk, and the resulting seed is used to produce the next generation. Mass selection has been used for centuries to breed cross-pollinated species and is still used for economically less important species.
Production of hybrid varieties in cross-pollinated crops follows the same path as with self-pollinated crops. Superior plants are selected and selfed for several generations to produce inbred lines that are uniform and homozygous. To ensure selfing in cross-pollinating plants, flowers must be protected from foreign pollen to avoid contamination. Selected parent inbred lines are chosen that are genetically contrasting and carry the desired traits. Inbreeding tends to decrease the vigour of the lines substantially, but crossing the unrelated inbred at the final stage leads to hybrid vigour, with the resulting F1 hybrids generally being superior to those of open-pollinated cultivars. Because the inbred lines are homozygous, the hybrid progeny of two inbreds will always be the same. However, if hybrid plants reproduce, the progeny will differ from the parent hybrid, and the hybrid vigour will be lost in the next generation. Therefore, seed produced from hybrid varieties is not normally retained to be used for planting. This contrasts with open pollinated cultivars, where if the plant self-pollinates or is pollinated by another plant of the same variety, the resulting progeny will be very similar to the parents.
Maize, or corn, is an ideal model of a cross-pollinated hybrid crop. Maize is wind-pollinated, so controlled cross-pollination in the field can be accomplished by interplanting two or three rows of the seed parent inbred with one row of the pollinator inbred and detasselling the seed parent before it sheds pollen. Cytoplasmic male sterility can also be used to avoid the need to detassel the seed parent and reduce the costs of production.
A synthetic variety is developed by intercrossing various genotypes that are known to display good combining ability, which means that they show good performance when crossed in all combinations. The commercial variety is a mixture of different lines rather than a genetically uniform population. Synthetic varieties demonstrate hybrid vigour but can produce usable seed for succeeding seasons. Synthetic varieties are often used for forage crops where the development and use of hybrid varieties would be too costly.
3.3. Molecular Plant Breeding Techniques
Successful plant breeding relies on the ability to select superior genetics from a diverse population. Historically, breeding was limited to recombining closely related plant material by selection of superior physical characteristics (phenotypes). Now molecular techniques are available that can generate genetic diversity by moving genes between unrelated species or by precisely editing genes. Molecular techniques can also allow the breeder to make selections early in the breeding process at the molecular, cellular, or tissue level. These techniques can shorten the time and reduce the costs for developing improved cultivars and allow for the insertion of characteristics into a species that are not normally found in the population.
3.3.1. Marker Assisted Selection
Marker-assisted selection exploits linkage disequilibrium (LD) between markers and desirable traits being targeted in a breeding program (Yadav et al. 2021). Linkage disequilibrium refers to a non-random segregation of alleles at two or more loci so that they occur together either more or less often than they would if their occurrence were random (Slatkin 2008). If the LD shows that the alleles occur together more often than expected by chance, it may indicate that the alleles are located close together on the same chromosome, although other factors may also cause LD. The LD identifies sites that may be used as markers for desired traits in a breeding program.
A molecular or DNA marker is a difference in a DNA nucleotide sequence that is tightly linked to a target gene that expresses a trait. The target gene and its expressed trait or biological function, and the molecular marker are inherited together (Amiteye 2021). Marker assisted selection uses these genetic markers to identify the presence of a specific gene that carries a given trait and track its presence into succeeding generations. Selections can be made using markers without growing the plant out to physically observe the desired characteristic.
Markers rely on the existence of polymorphisms, or positions in the genes where the alleles that code for a trait can vary in their nucleotide composition, giving more than one possible form in the population. The most common polymorphisms are single nucleotide polymorphisms (SNPs), where a single nucleotide base (adenine, thymine, guanine or cytosine) variant occurs at a particular position within the target sequence. Insertion/deletion (INDEL) polymorphisms are slightly less common and occur when a small number of bases have been inserted or deleted from a DNA sequence.
A quantitative trait locus (QTL) is a section of the DNA associated with a quantitative phenotypic trait. The QTLs are mapped by identifying which molecular markers correlate with an observed trait. Many QTLs may be associated with a single trait.
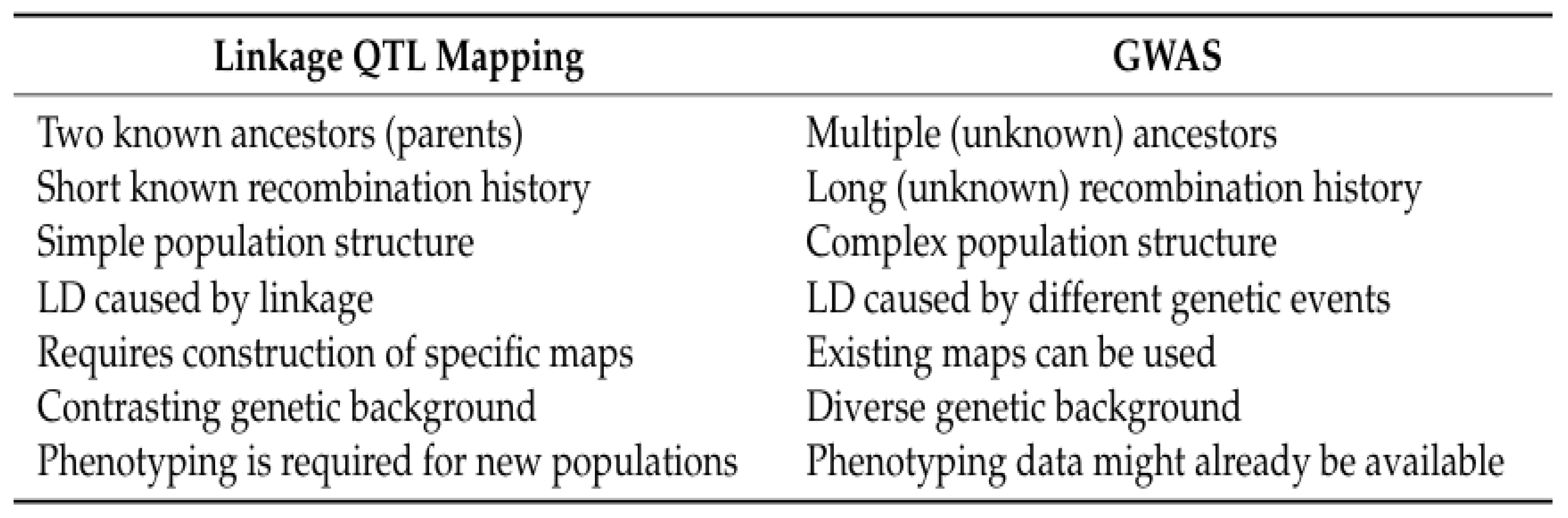
Innovations in DNA sequencing technology have allowed crops to be genotyped at the whole genome level (Shen et al. 2022). High-quality reference genomes will allow the determination of genetic patterns behind complex traits by using techniques such as quantitative trait locus (QTL) mapping and genome-wide association study (GWAS) (Table 3.1).
GWAS is a method that involves surveying the genomes of a population, looking for genetic variants that are associated with a specific trait and comparing them to those without the trait (https://www.genome.gov/genetics-glossary/Genome-Wide-Association-Studies-GWAS, accessed on January 7, 2025). GWAS will typically focus on associations between SNPs and target traits. GWAS is particularly useful to identify genetic variants that are associated with complex traits.

Table 3.1.
Comparison between biparental QTL mapping and genome-wide association study
(GWAS)(Colasuonno et al. 2021).
Table 3.1.
Comparison between biparental QTL mapping and genome-wide association study
(GWAS)(Colasuonno et al. 2021).
 |
Linkage maps show the position of molecular markers along the chromosomes based on the recombination events between individuals (Colasuonno et al. 2021). Genetic mapping is used to determine the linkage between genes affecting desired agronomic traits and the SNP or INDEL markers. The markers are then used to make selections from the segregating population. The marker may be the gene that codes for the desired trait, or more commonly, it is a segment that is located close to the gene of interest on the chromosome. A number of genetic marker systems are available, including restriction fragment length polymorphism (RFLP) markers, simple sequence repeat (SSR), Random Amplified Polymorphic DNA (RAPD) and amplified fragment length polymorphism (AFLP) markers (Guo-Liang 2013). Selection for the markers rather than directly for the desired trait can have advantages. Marker analyses can be more economical than phenotypic selection, but the major benefit is that the trait may be selected for before it expresses phenotypically. The trait may even be detected in a portion of the seed to allow selection prior to seeding, effectively reducing the need to grow and evaluate undesirable genotypes.
3.3.2. Genomic Selection
Genomic selection (GS) applies the genetic information to the breeding program. It involves using genomic information, mainly in the form of genome-wide SNP markers, to predict the genetic potential of individuals using statistical models without needing to phenotypically evaluate the plants for the desired traits (Griffin et al. 2025; Colasuonno et al. 2021). Genomic selection creates a training population using available genomic and phenotypic data and develops GS models that are fit to the training data. These statistical algorithms are used to characterize the relationship between genome-wide markers and the target trait, with weights being assigned to the markers. The best model is then used to analyse the genotype data of the prediction population to compute genomic-estimated breeding values (GEBVs). Genomic selection can predict the genetic value of individuals for use in a breeding program, allow selection of desired plants early in the breeding cycle, and is effective for multiple traits and traits with low heritability.
3.3.3. Doubled Haploids
In genetics, ploidy refers to the number of copies of the genome (n) that a cell carries. For example, humans are diploid (2n) with our cells carrying two copies of the genome, one from the mother and one from the father. The reproductive cells, the egg and sperm, are haploid (n) so that when the sperm (n) fertilizes the egg (n), the resulting embryo is diploid (2n).
In plant breeding, haploids and doubled haploids are used to facilitate the creation of homozygous lines (Murovec and Bohanec 2012). Haploid cells can be found in the reproductive cells of the ovaries and the anthers. The haploid pollen or egg cells are grown in vitro and treated, often with colchicine, to force the chromosomes to double, forming doubled haploids, with two identical chromosomes. Haploid cells can also be derived from wide crossing of distantly related species, where only one set of chromosomes ends up surviving. Radiation treatment can also be used to destroy one set of chromosomes and allow parthenogenic reproduction to create a haploid.
With some crop species, the genetics is a bit more complicated because instead of two copies of each chromosome, they may have multiple copies. For example, bread wheat is hexaploid, containing 6 copies of each chromosome in the mature cells (6x) while durum wheat and cotton are both tetraploids, containing four copies of each chromosome (4x). When using doubled haploid techniques, if the original plant was diploid (2x), the haploid cells contain one set of chromosomes (1x), while haploid cells derived from tetraploids will have two sets of chromosomes (2x), and those derived from hexaploids will have three sets of chromosomes (3x). So, a haploid cell in bread wheat is 1n and 3x, and the doubled haploid will be 2n and 6x.
Haploids are valuable tools in plant breeding. When haploid cells (n) are doubled, the resulting embryo/plant will be 2n for their chromosome number and be 100% homozygous. This means that desired traits are fixed within one generation, and the plant breeder can avoid the numerous cycles of inbreeding needed to produce homozygosity by conventional methods. Doubled haploid lines are also valuable for genetic mapping and trait analysis, allowing the identification of genotypes with improved disease resistance and yield.
3.3.4. Plant Tissue Culture
Tissue culture is the in vitro growth of cells, tissues, organs or whole plants to produce clones of plants (Argaye 2021). Tissue culture is an important step in doubled haploid production and large-scale plant multiplication. A single plant can be multiplied into several thousand plants in a relatively short time.
There are several different forms of tissue culture used in plant breeding. Embryo culture is used to grow embryos from seeds and ovules in a nutrient medium, with the plant developing directly from the embryo or indirectly through the formation of callus tissue followed by the formation of shoots and roots. In somatic embryogenesis, somatic cells or tissues develop into differentiated embryos and then into whole plants without undergoing fertilization. Embryo rescue involves excising plant embryos and growing them in media culture, often to create interspecific and intergeneric crosses where the seed would be aborted under normal circumstances. This allows wide hybridization, increasing the genetic pool for breeding.
3.3.5. Speed Breeding
Speed breeding can be used to help in the development of new mapping populations and to advance the first generations in the breeding programs (Colasuonno et al. 2021). Speed breeding techniques involve using extended photoperiods in a controlled environment to allow generations to be grown more rapidly. Some studies have produced up to six generations in a year (Watson et al. 2018). Speed breeding with the use of molecular markers can reduce the time needed to select improved lines, saving time for breeders.
3.4. Development of Variability in the Breeding Population
Plant breeding involves selecting for superior traits, but variability must exist in the population for the selection to be possible. Historically, variability arose naturally in the population due to accidental intrusion of genes from external sources or natural mutations that created new alleles or recombined existing or newly mutated alleles.
Mutations can also be artificially induced in plants by physical and chemical methods. Physical mutation occurs when radiation causes changes in the chromosome structure and number or changes in the base sequence of the DNA. Plants can also be treated with different chemicals that induce mutations. With both physically and chemically induced mutations, the changes are random and often detrimental. Screening and selection are required to identify beneficial changes and move them into the breeding program.
Many natural mutation occurs through the activity of Transposons (Lisch 2013). Transposable elements or Transposons are short strands of DNA that move or insert themselves at a new location in the genome. This genetic transposition can change the structural and functional characteristics of a gene by changing its position in the genome. Transposable elements cause mutation by both insertion and deletion. The mutations are most commonly either neutral or detrimental, but are important for the creation of a pool of genetic diversity that can be a platform for natural or targeted selection.
Newer methods of creating diversity can be more targeted to insert specific genes coding for desired characteristics. Genetically modified organisms (GMOs) have had their genome altered, usually by introducing genes from unrelated species, to supply traits that are not normally found in the population. Cisgenesis and transgenesis are distinct approaches used in GM technology. Cisgenesis refers to introducing genes from the same species or a close relative, while transgenesis refers to introducing foreign genes from an unrelated organism (Hamdan and Tan 2024). The transgene carrying a desired trait, such as improved nutritional content or resistance to herbicides or disease, is inserted into a vector, typically a plasmid, that can be used to introduce the trait into the plant cell. The insertion can occur using particle bombardment to fire the vector into the plant cell. Transfer DNA insertion (T-DNA) uses the tumor-inducing plasmid of some bacteria, such as Agrobacterium tumefaciens, to insert desired genes, often accompanied by a selection marker, into the plant genome. Insertion of the foreign gene into the genome creates a transgenic plant carrying the desired gene. After the trait is incorporated into the plant genome, the plant is used as a parent line in the breeding program to move the desired characteristic into an acceptable cultivar. GM technology has been widely used to develop herbicide tolerant crops, improve disease resistance, and improve nutritional quality. However, the transfer of genetic material across species lines has led to widespread consumer concern. In response, there is strict regulation of GM crops in many countries, ranging from outright bans to strict labeling requirements.
Gene editing uses engineered nucleases that contain a piece that binds to a specific DNA sequence, plus a nuclease that cuts the DNA strand (Joung and Sander 2013). The breaks are then repaired by mechanisms that produce a targeted change in the genetic sequence at the site where the cut was made. There are several gene editing methods used for plant breeding.
Zinc-finger nucleases (ZFNs) are engineered site-directed nucleases (SDNs) where a zinc-finger is the DNA-binding domain that is linked to the nuclease (Davies et al. 2017). Each zinc-finger recognizes a specific three base pair DNA sequence, and usually four to six individual zinc-fingers are linked together to recognize a unique 12-18 base pair DNA sequence. Two ZF proteins bind above and below the cut site which increases the specificity of the complete ZFN to 24–36 nucleotides. Targeted double-stranded breaks are made at the designated locations in the genome to produce precise deletion, addition, and editing of genes. This produces new genetic variation by deleting or editing gene sequences.
TALENs are similar to ZFNs, being engineered SDNs in which a transcription activator-like effector (TALE) domain is linked to an endonuclease (Joung and Sander 2013). Like ZFNs, TALENs cut the DNA and allow targeted changes to the gene.
The CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) system is another method of finding and editing a specific piece of DNA within a cell (Akhtar et al. 2023; Tao et al. 2023). Bacteria use CRISPR-associated (Cas) proteins to destroy viral DNA, and the discovery of this natural mechanism led to its application in gene editing. The Cas9 enzymes linked to the CRISPR sequences (CRISPR-Cas) can be used to edit genes within living organisms. The Cas9 enzyme uses the CRISPR sequences as a guide to recognize and open the DNA strands that are complementary to the target CRISPR sequence. A piece of RNA is used to program the Cas9 protein to guide it to find and bind to a specific target sequence. The Cas9 protein will hook up with the guide RNA and move along the strands of DNA until it finds and binds to a sequence of 20 DNA bases that match part of the guide RNA sequence. The Cas9 protein then cuts the DNA at the target site. Mutations are introduced as the cut is repaired, editing the gene and usually disabling it.
Other Cas proteins may not cut the DNA, but rather just turn the genes on (CRISPRa for gene activation) or off (CRISPRi for gene expression interference). In the CRISPRa system, enzymatically deficient Cas9 (dCas9), acting with transcriptional activators, leads to gene expression upregulation, while in CRISPRi, dCas9 and transcriptional repressor(s) repress gene expression (https://www.origene.com/products/gene-expression/crispr-cas9/crispra-crispri, accessed on January 7, 2025). The dCas9 contains mutations in two active endonuclease domains that make it lose its ability to cut DNA, but it can still target a specific DNA location guided by the RNA sequence.
There are also CRISPR systems called base editors that can specifically edit the DNA by changing one letter of the DNA code to another (Tao et al. 2023). DNA base editors may be more precise tools compared with the original CRISPR-Cas nuclease system.
Application of these gene editing tools can allow the “re-writing” of the genetic code for specific traits to introduce desirable characteristics into the breeding program.
3.5. Analytical Techniques
Plant breeding is a numbers game where vast quantities of genotypic and phenotypic data are generated. Modern breeding programs use high-throughput phenotyping techniques to collect and analyse digital data with the aid of drones, sensors, data collection and computation, and computer software. The ability to capture and quantify the physical characteristics of the plant to be targeted in the breeding program is as essential as the ability to characterize the genetic makeup. Advances in technology have allowed the collection of information in various aspects of “omics”, including genomics, proteomics, transcriptomics, metabolomics, and physiognomics. An understanding of how these factors interact to determine the characteristics of an organism is critical to successfully use the information in a breeding program (Shen et al. 2022). Statistical analysis of the collected data is needed to determine the relationship between physiological traits and the markers that identify the genetic locations that influence the traits.
Machine learning (ML) is a form of artificial intelligence (AI), where breeders can analyse and interpret huge data sets to link the phenotype with the genotype and guide their decision. AI and machine learning will allow the plant breeding program to dissect complex traits using omics-based interdisciplinary approaches and use this information to more efficiently develop improved cultivars. Machine learning can be used to identify trait-related genes and apply marker-assisted selection and/or genome editing to develop improved plant cultivars (Yan and Wang 2023). Machine learning is an important tool for genomic selection.
3.6. Summary
Plant breeding has played an important role in the improvement of crop yield, agronomic characteristics, disease resistance and nutritional quality. Traditional plant breeding relied on the selection of superior physical characteristics that were present in the natural population. New molecular techniques can generate genetic diversity by moving genes between unrelated species or by precisely editing genes. By being able to rapidly assess both genetic and phenotypic characteristics in a plant and to statistically determine the relationship between genetic regions and desired traits plant breeders can more accurately select for improved crops. Molecular techniques can also allow the breeder to make selections early in the breeding process at the molecular, cellular, or tissue level. These techniques can shorten the time and reduce the costs for developing improved cultivars and allow for the insertion of characteristics into a species that are not normally found in the population.
Section 4. Phosphorus Reactions in the Soil that Affect Phosphorus Availability
Key Points
- Plants take up P from the soil solution as the inorganic orthophosphate ion (Pi)
- Phosphorus concentration in the soil solution is very low and must be replenished from other soil pools to meet plant demand
- Supply of Pi to the plant roots will be affected by the concentration of Pi at the root surface and the speed that the concentration can be replenished
- Water-soluble P fertilizer will undergo a series of adsorption and precipitation reactions that move it from solution into less soluble labile and non-labile pools of P in the soil. These reactions are reversible and respond to the concentration gradient
- Plants respond to P deficiency with strategies to increase Pi in the soil solution, the root area available for P uptake and the ability to take up and move P into and throughout the plant
- Plant breeding can select for genetic traits such as root growth and root exudation that address restrictions in soil Pi to improve phosphorus use efficiency
4.1. Uptake of P from the Soil Solution
Phosphorus is taken up by the plant from the soil solution as the inorganic orthophosphate ion (Pi). Phosphorus uptake occurs mainly in the area of actively growing cells just behind the root cap, where root hair density is high (Hopkins 2015). Phosphorus uptake by the plant is driven by the concentration of Pi in the soil solution at the root surface and the area of absorbing surface in contact with the solution (Barber 1995). The concentration of Pi in the soil solution is normally very low, ranging from 10-8 M in low fertility tropical soil, to 10-6 M in deficient soils and as high as 10-4 M in some high P soils (Pierre and Parker 1927; Plaxton and Lambers 2015; Syers et al. 2008). The concentration of Pi in the soil is as much as one thousand times lower than that in the plant, so it must move from the soil solution into the plant against a steep concentration gradient, requiring an input of energy (Schachtman et al. 1998; Vance et al. 2003). Both the movement of Pi across cell membranes into the plant root and the distribution of Pi from the root throughout the plant are active processes using transporter proteins to move the Pi through the symplasm, xylem and across cell and organelle membranes to the regions of the cell cytoplasm and organelles where it is required (Schroeder et al. 2013).
An actively growing crop can require between 0.3 and 0.5 kg P ha− 1 each day (Johnston et al. 2014). The Pi in the soil solution around the root hairs is depleted quickly, leading to a zone of low Pi concentration at the root surface (Bagshaw et al. 1972). Roots can access some P as they grow into parts of the soil that have not been depleted, but only a very small proportion of the soil is explored by roots in any one growing season. Therefore, the P accessed by the plant through direct interception is less than 1% of the crop requirement (Barber 1995). Most of the Pi that the plant requires is supplied by replenishment of the Pi in the rhizosphere surrounding the plant root through mass flow and diffusion (Barber 1980; Barber et al. 1963).
Mass flow is the movement of dissolved nutrients with water as the water moves to the roots to meet the plant’s transpirational demand, while diffusion is the movement of nutrients through the soil solution from an area of high concentration to an area of low concentration (Glossary of Soil Science Terms | Soil Science Society of America, accessed on December 5, 2024). The movement of Pi by mass flow is low because of the low concentration of Pi present in the soil solution; so mass flow provides only about 2-3% of the amount required for optimum crop growth (Johnston et al. 2014). Most Pi moves to the root surface by diffusion down the concentration gradient created by the active uptake of Pi by plant roots (Barber 1995). Phosphorus diffusion in soil is affected by the water-filled pore space through which the ions can move and by retention reactions that impede ion movement. The path of movement through the soil moisture films around the soil particles is long and tortuous, and its movement along the path is slow, so the net movement of Pi through the soil is low, in the range of 0.13 mm per day (Johnston et al. 2014).
4.2. Reactions of P in the Soil
The Pi in the soil solution is present as orthophosphate ions, usually H2PO4– and HPO42–, with the dominant form in solution depending on the soil pH (Figure 1). Within the normal range of soil pH, HPO4-2 dominates if pH is greater than 7.2, and H2PO4- dominates at pH levels below 7.2 (Pierzynski and McDowell 2005). The availability of P for plant uptake tends to be the greatest in a pH range of 5-6, where the monovalent form of P dominates (Schachtman et al. 1998).
Plants take up the P ions from the soil solution, but the soluble Pi in solution generally represents less than 1% of P used by crops over a growing season and less than 1% of the total amount of P present in the soil. Most soil P is present in organic and inorganic forms that can be viewed as being “pools” of P that vary in availability. Labile P is the pool that moves in and out of the soil solution in the short-term, while non-labile P is more stable, slowly retaining and releasing P over the long-term (Johnston et al. 2014; Syers et al. 2008) (Figure 2).
If P is added or removed from the soil solution, P can move from pool to pool along the resulting concentration gradients (Johnston et al. 2014; Syers et al. 2008). For example, plant uptake reduces the Pi concentration of the soil solution at the root surface, creating a concentration gradient that drives the diffusion of P to the root surface and the mobilization of Pi into solution from the labile pools. Conversely, adding soluble forms of P, such as fertilizers, increases the soil solution Pi concentration and shifts the equilibria towards movement of P into the less available pools (Morel and Plenchette 1994; Morel et al. 2000; Schneider and Morel 2000).
The intensity factor (I) refers to the concentration of Pi in the soil solution while the quantity factor (Q) or the P buffering capacity describes the ability of the soil to replenish the Pi in the soil solution from other pools of P (Morel et al. 2000). During periods of peak P demand, the P in the soil solution at the root hair surface must be replenished at least 10 to 20 times per day (Syers et al. 2008). Therefore, plant-available P is affected both by the concentration of P in the soil solution (I) and the amount and rate of release of P from other soil pools (Q) (Morel et al. 2000).
When a fertilizer granule is added to the soil, the P source must dissolve and enter the soil solution before it becomes available for soil reactions or plant uptake. Phosphate fertilizers are hygroscopic, so will be dissolved by water vapour from the soil air-filled space and soil porewater that moves towards the fertilizer through mass flow and capillary flow (McLaughlin et al. 2011; Hettiarachchi et al. 2006). As the granule or droplet dissolves, the highly concentrated P solution created from the dissolution of the fertilizer granule begins to diffuse away from the application point, with the P concentration decreasing with increasing distance from the origin. The initial dissolution and movement of P out of the granule occurs rapidly, within a few days (Lombi et al. 2004; Hedley and McLaughlin 2005; McLaughlin et al. 2011). The residual granule or droplet contains insoluble P compounds from the original fertilizer as well as the compounds that precipitated from the reaction of the highly concentrated fertilizer solution with reacting cations (Hedley and McLaughlin 2005; Kar et al. 2012). Beyond this is a zone of soil surrounding the granule where the capacity of the soil to adsorb P has been saturated, and precipitates have formed from the reaction of the fertilizer solution with the metal ions and organic matter released from the soil. As the distance from the application point increases, the solution becomes more dilute, and the soil will be able to adsorb the P without being saturated.
Only a small portion of the P from the fertilizer remains in solution because it will equilibrate with the less soluble labile and non-labile pools of P in the soil through a series of adsorption and precipitation reactions (McLaughlin et al. 2011; Sample et al. 1980; Hedley and McLaughlin 2005). These reactions include adsorption on the surface of the soil particles, diffusion (absorption) of the adsorbed P from the outer surface to the inner surfaces of the particles, where it is less available, and precipitation of a new sparingly soluble solid phase P (Johnston et al. 2014; Ajiboye et al. 2007; Ajiboye et al. 2008). The reactions that occur depend on the concentration of the phosphate and reacting ions in the soil solution.
The soil pH affects the formation of phosphate compounds. In high pH soils, phosphate ions react with Ca and Mg ions to produce sparingly soluble Ca and Mg phosphate compounds.
e.g., dicalcium phosphate dihydrate (DCPD):
CaHPO4 ∙ 2H2O + H+ ↔ H2PO4− + Ca2+ + 2H2O
In a high pH soil, H+ is an ingredient in the dissolution reactions for calcium and magnesium phosphates, so decreasing pH increases dissolution. In contrast, Ca and Mg and phosphate are reactants in the precipitation side of the reaction, so increasing their concentration will increase precipitation. Hydroxyapatite is generally the most stable Ca-P form in soils, but a range of metastable forms, such as dicalcium phosphate dihydrate or octacalcium phosphate, can exist (Lombi et al. 2006; Fixen et al. 1983; Sample et al. 1980)
In acid soils, phosphate reacts with Fe or Al to form increasingly less available compounds.
e.g., for strengite:
FePO4 ∙ 2H2O + H2O ↔ H2PO4− + H+ + Fe(OH)3
Since H+ is a product of the reactions for Fe or Al phosphates, low pH and increasing concentrations of Fe, Al or phosphate will increase precipitation. There is little evidence of bulk precipitation of P with Al or Fe in soils, but phosphate may be adsorbed onto Fe and Al (hydr)oxides, form complexes with these oxides, or become occluded within Fe and Al oxide-rich minerals such as hematite, goethite, and gibbsite (Mabagala and Mng'ong'o 2022; Penn and Camberato 2019; Hedley and McLaughlin 2005).
The formation of compounds in the residual granule and the surrounding fertilizer-soil interface is affected primarily by the type of solution formed by the applied fertilizer and the available moisture from the soil (Hedley and McLaughlin 2005; Sample et al. 1980). Mass flow of reacting cations in the soil water moving towards the granule may increase P precipitation, limiting the movement of P away from the granule and reducing the volume of the fertilizer reaction zone. In some soils, when P fertilizer is applied as a solution rather than as a granule there is less movement of water carrying reacting ions towards the fertilizer, so precipitation is reduced, and P will move further away from the site of application, increasing its availability (Bertrand et al. 2006; Holloway et al. 2001; McBeath et al. 2005; Lombi et al. 2004, 2005).
Blending soluble salts such as ammonium nitrate, ammonium sulphate, potassium nitrate, potassium chloride or potassium sulphate with the phosphate fertilizer can produce relatively soluble reaction products such as Ca2KH7(PO4)4.2H2O or Ca2NH4H7(PO4)4.2H2O and increase phosphate movement away from the granule (Sample et al. 1980; Hedley and McLaughlin 2005). Creating a larger reaction zone with more soluble reaction products can improve plant availability of the P.
These retention reactions reduce the immediate availability of P fertilizers, but the process is reversible, and the retained P forms can become available over time in response to P removal from the soil solution (Syers et al. 2008). Therefore, effects of soil reactions on P fertilizer availability need to be considered in both in the short-term and over time.
4.3. Plant-Soil Interactions
Availability of P for crop uptake is influenced by the concentration of Pi in the soil solution at the root surface and by the root absorption area. When P is scarce, plants can act to increase their access to P by various methods that are under genetic control. These methods include:
- 1.
- Increasing root production and altering root architecture to increase the volume of soil explored and the root surface area available for P uptake.
- 2.
- Excretion of various organic acids and enzymes that can increase the amount of Pi that moves into the soil solution.
- 3.
- Increased activity of transporter proteins to more efficiently take up P from the soil solution, depleting the soil solution concentration of Pi and encouraging more movement of Pi into the soil solution and towards the root surface.
- 4.
- Enhanced mycorrhizal associations to further increase soil exploration for P in certain plant species.
Understanding the physiological and genetic factors controlling these plant responses to limited P may help to identify breeding targets that can be used to select for plants with a higher phosphorus use efficiency.
4.4. Summary
Plants take up P from the soil solution as the inorganic orthophosphate ion Pi. The concentration of Pi in the soil solution is very low and is depleted quickly by plant uptake. The Pi in the soil solution must be replenished from other soil pools to meet plant demand. Uptake of Pi by the plant is affected by the concentration of Pi at the root surface and the speed at which the concentration can be replenished. Phosphorus fertilizer is used to increase the supply of P to the plant when soil supplies are insufficient. Phosphorus fertilizer will undergo a series of adsorption and precipitation reactions that move it from solution into less soluble labile and non-labile pools of P in the soil. These reactions are reversible and respond to the concentration gradient. Plants have developed strategies to increase Pi in the soil solution, increase the volume of soil explored for Pi and enhance the uptake of Pi to maintain Pi supply when its availability is limited. Breeding programs may be able to target genetic factors that influence the ability of the plant to mobilize, access and utilize soil P, thus increasing the phosphorus use efficiency of plants.
Section 5. Phosphorus Accumulation and Utilization in Crops
Key Points
- Plants require P for cell growth from the earliest stages of germination
- After seed reserves of P are depleted, plants rely on inorganic P from the soil solution to support growth
- Uptake of P from the soil solution is a function of the absorbing area of the root and the concentration of Pi in the soil solution at the root surface
- Root growth and architecture respond to P availability and are under genetic control
- Most plant species form mycorrhizal associations to improve access to soil P
- Plants secrete low molecular weight amino acids that increase P solubility and mobility, thus increasing its availability
- Uptake of Pi from the soil solution and its distribution throughout the plant is facilitated by transporter proteins whose production and function are affected by plant genetics
- Surplus Pi is stored in the vacuole and can be mobilized to maintain cytoplasmic Pi concentration when Pi supply is low
- The ability of the plant to mobilize and adjust the distribution of P among the various P forms present in the plant to maintain homeostasis is under genetic control and may be a target for breeding
- Reduction of phytate concentration in seeds could reduce P concentration in manure and sewage, improving both environmental sustainability and phosphorus use efficiency
5.1. Introduction
Phosphorus is essential for all energy and growth reactions in the plant, and so must be present in the cells in adequate amounts from the earliest stages of germination (Grant et al. 2001). For the first few days to weeks of life, the plant relies on P stored in the seed (Veneklaas et al. 2012). How long the seed reserves will last is a function of the seed size, the concentration of P in the seed, the concentration of P in the soil and the P demand from the growing seedling. The seed only contributes a small amount of the total P needed by the plant, but it is very important in determining early growth and vigour that allows the young seedling to access moisture, nutrients and light and to compete with weeds. As the seed reserves decline, the plant begins to access Pi from the soil solution.
Plants take up inorganic Pi from the soil solution. The inorganic Pi in the soil solution is present as orthophosphate P ions, usually H2PO4– and HPO42–, with most Pi being present as H2PO4- if the pH is below 6. Uptake rates of Pi by plants tend to be greatest between pH of 5 and 6, indicating that Pi is primarily taken up by the plant in the monovalent form, H2PO4- (Schachtman et al. 1998). Uptake of Pi by the plant is a function of the concentration of P ions in the soil solution at the root surface and the area of absorbing surface in contact with the solution (Barber 1995).
5.2. Root Exploration of the Soil
An actively growing crop can require between 0.3 and 0.5 kg P ha− 1 each day and will quickly deplete the Pi in the soil solution around the root hairs, leading to a zone of low Pi concentration at the root surface (Bagshaw et al. 1972; Johnston et al. 2014). Roots can access additional P as they grow into parts of the soil that have not been depleted, but only a very small proportion of the soil is explored, and direct interception of Pi by the root is less than 1% of the crop requirement (Barber 1995). Most of the Pi that the plant requires is supplied by replenishment of the P within the rhizosphere (Barber 1980; Barber et al. 1963; Johnston et al. 2014).
The Pi in solution at the root surface will be replenished through mass flow, diffusion and by mobilization of P from various soil P pools. Due to the low concentration of Pi present in the soil solution, mass flow can provide only about 2-3% of the amount required for optimum crop growth (Johnston et al. 2014). Most Pi moves to the root surface by diffusion down the concentration gradient created by the active uptake of Pi (Barber 1995). The path of movement through the soil moisture films around the soil particles is long and tortuous, and its movement along the path is slow, so the net movement of Pi through the soil is also low, in the range of 0.13 mm per day (Johnston et al. 2014). The low mobility of P in the soil means that mobilization of Pi from P pools in the soil (Quantity factor, Q, or the P buffering capacity) is also important in replenishing the Pi in solution around the root (Morel et al. 2000). The low concentration of P in the soil solution and its relative immobility create challenges for plant access to the P that they require.
The importance of P for plant growth has led plants to develop strategies to improve their ability to access P when deficiencies occur. These strategies are under genetic control and could be targeted in plant breeding efforts to increase phosphorus use efficiency. One strategy used by plants to increase access to P is enlargement of the root surface area and alteration of the root system architecture. Under P deficiency, total root length and biomass may decline, but the plant modifies its growth characteristics to minimize the negative effects on P acquisition (Lopez et al. 2022). When P supply is limited, plants will retain P in the root to support root growth and improve the ability of the root system to access soil P (Loneragan and Asher 1967; Schjørring and Jensén 1984; Sutton et al. 1983). With P deficiency, many plants will also allocate more carbohydrate to roots than shoots (Marschner et al. 1996), increasing the root:shoot ratio (Brenchley 1929; Schjørring and Jensén 1984; Gaume et al. 2001). Under low-P conditions, plants develop highly branched roots, numerous and long root hairs, and a greater root length per unit mass (Barber 1977; Ozanne 1980; Hodge 2004; Brenchley 1929; Schjørring and Jensén 1984; Tomasiewicz 2000; Vance et al. 2003). Having finer, longer, and more abundant roots and root hairs allows the plant to explore more soil and to expose a greater surface area of root for P uptake. Distribution of the roots through the soil profile can also affect the ability to access P. Phosphorus tends to concentrate in the soil surface because of deposition from decomposing plant residue (biocycling). In fertilized systems, stratification is increased further because fertilizer P applications are normally placed in the upper soil layers (Grant and Lafond 1994). Since the concentration of P is usually greater in the surface soil layers, increasing the proportion of roots near the soil surface can increase P uptake (Lynch 2007b; Lynch and Brown 2001).
In addition, many plants, such as canola and buckwheat, will preferentially increase root density when they encounter a region of high P concentration, such as a fertilizer reaction zone, increasing the ability of the plant to extract P from that area (Strong and Soper 1974a, 1974b; Drew and Saker 1978; Foehse and Jungk 1983). Uptake of P by roots is proportional to both the concentration of the P at the root surface and the area of absorbing root surface that contacts the P, so root proliferation in a zone of high P concentration increases the ability of the plant to take up P. Other plants, such as flax and soybean, are less able to proliferate roots in a high-P zone and are less effective at using fertilizer P.
Many plants secrete low molecular weight organic acids that acidify the rhizosphere, compete with phosphate ions for sorption sites, and complex cations bound to phosphate ions, thus increasing P availability (Jungk et al. 1993; Hoffland et al. 1989; Hinsinger 1998, 2001; Hinsinger and Gilkes 1995; Gaume et al. 2001). Some plants can also excrete phosphatases into the rhizosphere to release P from organic forms (Lefebvre and Glass 1982; Hinsinger 2001; Ashworth and Mrazek 1995). Plants can also produce compounds that encourage the proliferation of P-solubilizing microorganisms in the rhizosphere (Kucey et al. 1989; Richardson 2001; Richardson et al. 2009).
Another strategy used by most plant species to improve access to soil P is to form mycorrhizal associations (Bolan 1991; Grant et al. 2005; Hamel and Strullu 2006; McGonigle et al. 2011; Miller 2000; Monreal et al. 2011; Ryan and Graham 2002; Ryan et al. 2000; Jakobsen 1986; Smith et al. 2011). In the mycorrhizal symbiosis, the host plant provides fixed carbon to the fungi and in return, the fungi capture P, water and some other nutrients from the soil and transfer them to the plant. The mycorrhizal hyphae are finer than plant root hairs and will extend further from the root and into finer soil pores, greatly increasing the effective volume of soil that can be accessed by the mycorrhizal association. Plants vary substantially in their reliance on mycorrhizal colonization for P access, with some crops, such as flax and maize, relying heavily on mycorrhizal associations and other crops, such as canola and sugar beet, having no mycorrhizal associations. Mycorrhizal associations are effective at low to moderate soil P concentrations but are less valuable where P concentration is either very low or very high. If the nutrient level in the soil is high, the plant may be able to access sufficient P to optimize crop yield, rendering the mycorrhizae unnecessary. On the other hand, if the P level in the soil is very low, the fungi may compete with the plant for the limited amount of P, reducing the P that is available for crop assimilation (Ryan and Graham 2002).
5.3. Transport of P into the root
The growing end of the root consists of the root cap and the root tip (Figure 3). The root cap protects the root tip, an area of high meristematic activity where root cells are produced (Kumpf and Nowack 2015). The root cap also senses conditions in the environment and triggers tropic responses, guiding the root toward or away from specific soil conditions (Kumpf and Nowack 2015). In addition, the root cap has a role in sensing P deficiency and initiating root growth responses to increase the plant’s ability to access P from the soil.
The root tip is divided into three main parts: an area of cell division, an area of cell elongation and an area of cell maturation and differentiation (Figure 3). New cells are produced in the area of cell division, and the undifferentiated cells expand in the area of cell elongation. In the area of cell maturation and differentiation, the cells become specialized for absorption, transportation or protection. Root hairs are formed at the beginning of the area of maturation. Root hairs are specialized epidermal cells that are very long and extend into the soil. Their large surface area increases their absorption capacity, providing a very high ability for P uptake relative to the resources invested. The major location for P uptake by plants, therefore, is in the area of actively growing cells just behind the root cap, where root hair density is high (Hopkins 2015). The area of differentiation is also where endodermis forms the Casparian strip and a suberin layer that acts as a barrier to the passive movement of ions and water (Chen et al. 2022). This restriction of the passive movement of water and nutrients in and out of the symplast is critical for the control of nutrient uptake.
Under P-sufficient conditions, the concentration of Pi in the cytosol is in the millimolar range, while the Pi concentration in the soil solution is as much as 1000-fold lower than that in the plant (Schachtman et al. 1998; Vance et al. 2003; Wang et al. 2021). Passive movement of Pi in response to the concentration gradient would be out of the root and into the soil solution. Therefore, roots must take up Pi from the soil solution against a steep concentration gradient using an active process that requires energy.
Movement of Pi across cell membranes relies on root phosphate transporters (PHTs). Various PHT families exist, with the PHT1 family being present on the plasma membrane and involved in the uptake of Pi from the soil solution (Wang et al. 2021). A suite of transporter proteins exists, with both low- and high-affinity transporters being involved in phosphate uptake (Schroeder et al. 2013). The high affinity system tends to function when Pi concentrations are low, improving the plant’s ability to transport the orthophosphate across the root membranes when P supply is restricted. Since P is transported as an anion, co-transport requires a counterion. Uptake of P increases the pH of the external solution, indicating that Pi uptake involves transport of H+ as a counterion, with two to four H+ ions being expelled per H2PO4− moved (Wang et al. 2021; Vance et al. 2003).
Plants can implement physiological changes in response to P deficiency to enhance the ability of the root to absorb P from the solution (Hodge 2004). The uptake rate per unit of root can be increased in P-deficient plants (Hodge 2004; Drew et al. 1984; Green et al. 1973; Jungk and Barber 1974; Anghinoni and Barber 1980; Borkert and Barber 1983). These changes can involve alterations in the number and activity of the various phosphate transporters and are under genetic control. The internal concentration of Pi in the plant is a trigger for the degree of PHT1 expression. Expression of PHT1 is low at high internal P concentrations while expression is higher when P concentration in the plant declines (Wang et al. 2021). This pattern of expression may indicate upregulation in response to deficiency or downregulation when P is sufficient (Lambers 2022).
5.4. Movement of P from the Root Throughout the Plant
After Pi is taken up and transported across the plasma membranes of the root cells, it moves in the symplasm from the root surface to the xylem. The Pi then travels in the xylem from the root and is distributed throughout the plant, again by passing through the membranes of other cells and organelles (Schachtman et al. 1998; Rouached et al. 2010). Movement from the xylem to the cell cytoplasm is also against a steep concentration gradient, so it requires active transport. Therefore, cross-membrane transport is needed not only to acquire Pi from the soil solution but also to move it within the plant’s inside cells, between cells, and between plant organs and organelles. Phosphate transporters function in the distribution of Pi throughout the plant and in regulating phosphate homeostasis. The PHT2/3/4 families are involved in transporting Pi into or out of subcellular organelles, such as mitochondria, plastids or the Golgi apparatus, while the PTH5 family is involved in the movement of Pi in and out of the vacuole (Wang et al. 2021). Again, the synthesis and activity of these transporters are under genetic control and are responsive to the Pi status of the plant.
5.5. Phosphorus Utilization in the Plant
Phosphorus is present in the plant in four major forms: inorganic P, metabolic P or P-esters, P in phospholipids and P in nucleic acids, as well as in a poorly defined residual fraction that may include phosphorylated proteins (Lambers 2022; Veneklaas et al. 2012). If P is not highly deficient, most P is present in the vacuole, while the size of organic P pools usually decreases in the order RNA-P > lipid-P > P-ester > DNA-P. The distribution of P among the main organic P pools differs slightly among species, but it is affected largely by the P status of the plant (Figure 4). Most of the P in the plant is present as inorganic phosphate, with only a small portion being metabolically active. This small amount of active P remains relatively constant with changes in P supply, while the concentrations of inorganic P may vary substantially, being stored or mobilized as external supplies increase or decrease. Since P is a major component of critical cellular components, including nucleic acids, membrane lipids, and ATP, cellular P homeostasis is essential. The ability of the plant to mobilize and adjust the distribution of P among the various P forms present in the plant is under genetic control and again may be a target for breeding efforts to improve phosphorus use efficiency.
The young seedling initially relies on seed reserves to supply the P needed for growth. As the vegetative growth increases, P uptake by roots is by far the most important P source as root system expands and explores the soil. During the late vegetative and early reproductive stages growth rate declines, the older leaves start senescing, and root expansion and P uptake by root systems decreases. During these later growth stages, remobilization of P from plant tissue can provide significant amounts of P for new growth and seed development (Veneklaas et al. 2012). Remobilized P is particularly important in P deficient plants. Under P deficiency, plants develop adaptive responses not only to efficiently access and take up Pi by altering root architecture and function, but also to efficiently utilize the Pi that is present in the cell by adjusting internal Pi recycling, restricting P consumption, and reallocating P from old tissues to young, actively growing tissues. Low inorganic P is the prime signaling mechanism in the plant indicating deficiency and inducing genetic responses to deal with low P status.
When plants have a sufficient supply of P, surplus Pi will be stored in the vacuole, where it can be a reserve for future deficiencies. Phosphorus deficiency can trigger an active movement of Pi from the vacuole to the cytoplasm to meet the metabolic requirements of the plant (Yang et al. 2017). Under P stress, P concentration in the vacuole will decrease to very low levels, while the decrease in P concentration in the cytoplasm will be much smaller due to the buffering from the vacuole (Veneklaas et al. 2012). Although 85% to 95% of the cellular Pi is present in the vacuole, movement of Pi from the vacuole may not be enough to maintain P concentration in the cytosol during severe Pi limitations.
The coordinated activity of Pi transporters controls movement of P within the plant. These transporters are under genetic control and are induced by P deficiency. Under Pi stress, plants will preferentially allocate Pi to photosynthetic tissues to maintain photosynthesis and carbon metabolism (Han et al. 2021). In a dense crop canopy, the older leaves become shaded and their photosynthetic ability decreases (Veneklaas et al. 2012). The shaded leaves will begin to senesce and plants will mobilize Pi from older tissue and translocate it to young, actively growing tissue to maintain photosynthetic capacity (Han et al. 2021).
Additional Pi can be transported from the Golgi luminal space, recycling Pi released from glycosylation (Shen et al. 2011). The Pi can also be recycled from other P organic sources, such as phosphomonoesters and nucleic acids. The low molecular weight P esters including adenosine diphosphate, adenosine triphosphate and sugar phosphates generally comprise 15–21 % of total P in most plants (Han et al. 2021; Veneklaas et al. 2012). Under P deficient conditions and during leaf senescence, these esters can be scavenged for Pi.
The nucleic acid pool is usually the largest pool of organic P and will contain between 40 and 60% of the organic P pool in a plant (Veneklaas et al. 2012). At least 85% of the nucleic acid is RNA and the rest is DNA. Most of the RNA in the plant is ribosomal RNA (rRNA), with a very small amount present as messenger RNA (mRNA) and transfer RNA (tRNA). The RNA supply is associated with the ability of the plant to synthesize protein. The RNA content of the leaves increases as the leaves develop until full expansion to support synthesis of the proteins needed for photosynthesis. After full leaf expansion, RNA supply decreases as senescence proceeds. Remobilization of the P present in RNA is a significant source of Pi that can be moved from older to younger plant parts and from vegetative to reproductive tissue.
When the plant enters the reproductive stage, Pi must be mobilized from the vegetative tissue and moved to the seed. Most of the P in the seed originates from the remobilization of Pi from vegetative tissue, with the proportion of remobilized P increasing with P deficiency (Shen et al. 2011; Han et al. 2021). Phytate is the main storage form for P in the seed and is an antinutritional component for human and monogastric livestock and poultry diets. The high concentration of P in seeds means that a large amount of P is transported from the area of production to the area of consumption, where it ultimately enters the waste stream or manure. Reduction of the amount of phytate in seeds would be desirable if it could be accomplished without reducing seedling vigour (Rose et al. 2013). Therefore, the reduction of seed phytate could be a useful target for breeding to improve phosphorus use efficiency.
Phosphorus requirements may also be reduced by decreasing phospholipid and increasing non-P lipids in the membranes (Essigmann et al. 1998). In addition, plants may also be able to use alternative cellular respiratory pathways in place of the adenylate and Pi reactions of respiration to allow respiration to proceed with reduced Pi demand under P starvation (Theodorou and Plaxton 1993).
5.6. Summary
Phosphorus is essential for all energy and growth reactions in the plant and so must be present in the cells in adequate amounts from the earliest stages of germination. For the first few days to weeks of life, the plant relies on Pi stored in the seed, but as the seed reserves are depleted, plants rely on Pi from the soil solution to support growth. Plants take up P as orthophosphate ions against a concentration gradient in a process facilitated by transporter proteins. Uptake of P from the soil solution is a function of the absorbing area of the root and the concentration of Pi in the soil solution at the root surface. When P is deficient, root growth is increased and root architecture changes to increase root surface area and soil exploration for P. Plants may also form mycorrhizal associations to increase the volume of soil accessed. Phosphorus is present in the plant as inorganic P, metabolic P or P-esters, P in phospholipids and P in nucleic acids. Surplus Pi is stored in the vacuole and can be mobilized to maintain cytoplasmic P concentration when Pi supply is low. The major storage form of P in seeds is phytic acid, which can cause environmental issues when it produces high P levels in manures and sewage. Reduction of phytate concentration in seeds could lower the removal of P from the soil and reduce P concentration in manure and sewage, improving both environmental sustainability and phosphorus use efficiency. Factors that influence P uptake, distribution and homeostasis are under genetic control, and plant breeding may be able to target these physiological processes to improve phosphorus use efficiency without negatively impacting crop yield.
Section 6. Genetic Improvement of Phosphorus Access From the Soil
Key Points
- Plants have developed strategies to improve their ability to access P from the soil
- Genetic modification of the size and shape of the root system may increase the ability of the plant to access and absorb P from the soil solution
- Genetic modification of the ability of a plant to form a mycorrhizal association is possible, but may not provide a consistent enough benefit to warrant inclusion in a breeding program
- Plant root exudates may mobilize P into the soil solution and influence mycorrhizal colonization and/or rhizosphere; however, effects may not be persistent enough to justify breeding to modify exudation
6.1. Introduction
Crops take up Pi from the soil solution through the root system. Uptake of Pi by the crop is a function of the root surface area and the concentration of Pi in the soil solution (Barber 1995). Therefore, the ability of the crop to take up Pi will be affected by the size and surface area of the root system and its distribution in the soil. The critical nature of P for plant growth has led plants to develop strategies to improve Pi uptake from the soil. Mechanisms that increase root uptake of Pi may include changes in the root system, interactions with microorganisms and chemical modifications of the rhizosphere (Magalhaes et al. 2017). These mechanisms are genetically based and respond to changes in P status in the soil and plant. Because these characteristics are genetically based, it should be possible to breed for plants that have a greater ability to access and take up P from the soil by modifying the root system, relations with microorganisms and the ability of the plant to modify P solubility in the rhizosphere.
6.2. Breeding for an Improved Root System
Uptake of Pi by the plant is a function of the root surface area and the concentration of Pi in the soil solution at the root surface. The size, structure and distribution of the root system will influence both the total root surface area available for P uptake and the regions of the soil that the root system explores. Although roots explore only a very small proportion of the soil and direct interception of Pi by the root is less than 1% of crop requirement, Pi diffuses to the root over very short distances, so root growth and architecture still have a major impact on the ability of the crop to access and accumulate P (Barber 1995).
Root system size and architecture are influenced by both genetic and environmental factors. The type of root system varies with crop species. Monocots have a fibrous root system consisting of embryonic seminal roots, crown roots or adventitious roots, lateral roots and root hairs. Adventitious roots form in both monocots and dicots at the junction between the root and stem. These crown or nodal roots help to anchor the plant and to better explore the soil surface where P is concentrated (Magalhaes et al. 2017). Dicots have a taproot system with an embryonic primary root, lateral roots, and root hairs. In dicots, the primary root typically becomes a dominant central taproot. The taproot gives rise to the lateral roots that develop to form a complex root system. The primary root normally continues to function throughout the plant’s life (Lopez et al. 2022). The lateral roots usually grow out of the primary root at a specific root growth angle that influences the pattern and distribution of the root system. Root hairs form from specific epidermal cells on the roots and greatly increase the surface area available for nutrient and water absorption (Magalhaes et al. 2017).
An important plant response to P stress is to modify the root system to increase soil exploration for P and the root surface area available for P absorption. While root length and biomass generally decrease with P deficiency, the plant attempts to reduce the impact on P uptake. With limited P, plants will retain P in the root and allocate more carbohydrate to roots than shoots to support root growth, increase the root:shoot ratio, and improve the ability of the root system to access soil P (Gaume et al. 2001). Root system architecture changes under P stress, developing highly branched roots, numerous and long root hairs, and a greater root length per unit mass (Barber 1977; Ozanne 1980; Hodge 2004; Brenchley 1929; Schjørring and Jensén 1984; Tomasiewicz 2000; Vance et al. 2003; Lynch 2022; Lynch 2007a, b; Lynch and Brown 2001), allowing the plant to explore more soil and increase the surface area of roots for P uptake (Bernardino et al. 2019). The highest concentration of P in the soil is usually in the upper 10 cm, so increasing the concentration of roots in the soil surface increases the ability of the plant to take up P (Carkner et al. 2023; Grant and Bailey 1994; Lynch 2022; Lynch and Brown 2001). In studies with maize, wheat and beans, cultivars with a shallow rooting system were more P-efficient than cultivars with a deeper rooting system (Zhu et al. 2005; Richardson et al. 2011). Therefore, increasing root density with roots growing at a shallower angle helps in scavenging of P from the upper soil profile, where concentration is higher than at depth (Fradgley et al. 2020; Lynch and Brown 2001).
Root hairs are also extremely important in the uptake of relatively insoluble nutrients such as P. Root hairs have a very high surface area in proportion to their weight. Increasing root hair length and density will increase the surface area available for P uptake and the ability of the plant to access P (Richardson et al. 2011; Heuer et al. 2017) .
Formation of dense “cluster roots” can occur in plants, such as Proteaceae and the Cyperaceae, that are adapted to extreme Pi scarcity (Lambers 2022; Lambers et al. 2003). They can also be found in white lupin and some native Australian legumes (Richardson et al. 2011). These are shallow clusters of short, lateral roots that grow near the soil surface where P concentration is higher than in the deeper soil layers. Higher root surface area in soil regions where the P concentration is highest increases the ability of the plant to access P, especially where overall P supply in the soil is low. High root surface area relative to biomass also increases the efficiency of P uptake per unit investment in dry matter (Lynch 2022; Lynch 2007b; Lynch and Brown 2001). These structures may also secrete organic acids to solubilize P from the soil (Lambers 2022; Lambers et al. 2003).
Many plants, such as canola and buckwheat, will also preferentially increase root density when they encounter a region of high P concentration, such as a fertilizer reaction zone (Strong and Soper 1974a, 1974b; Drew and Saker 1978; Foehse and Jungk 1983). Uptake of P by roots is proportional to both the concentration of the P at the root surface and the area of absorbing root surface that contacts the P, so root proliferation in a zone of high P concentration increases the ability of the plant to take up P (Barber 1995).
Genetic differences exist in root growth and architecture, and in the ability of the plant to modify root development in response to P limitations (Bayuelo-Jiménez and Ochoa-Cadavid 2014). Therefore, it should be possible to selectively breed for desired characteristics to improve the efficiency of the plant root system to access P (Li et al. 2024; Pan et al. 2008). However, historically, breeders have selected new cultivars based on above-ground traits, and there appears to have been passive selection against larger root systems over time (Fradgley et al. 2020; McGrail et al. 2023). Older varieties generally had more crown and nodal roots and narrower root systems than modern cultivars. Recently, with the recognition of the importance of root traits for crop resiliency, there has been an increase in efforts to breed for desired root characteristics.
Selection for specific traits requires the ability to identify or measure variation in the characteristic or phenotype and to select for the genotype that controls or contributes to producing the trait. Breeding specifically for root characteristics has been challenging in part because determining the phenotype of roots is difficult since they grow below ground. Also, root growth is dynamic and can change in response to water and nutrient distribution. In the past, phenotyping methods have focused mainly on root traits in young plants under controlled conditions that do not necessarily reflect soil conditions in the field (Fradgley et al. 2020; Griffin et al. 2025; Mikwa et al. 2024; Atkinson et al. 2019; Atkinson et al. 2015). However, recent advances have led to improved methods for collecting, imaging and handling more accurate and realistic root information. Soil-filled rhizotrons consisting of boxes with transparent front panels can be used to assess root growth over time. Roots can also be observed by growing plants in transparent tubes or in semi-hydroponic systems (Li et al. 2024). Crop root systems can also be measured in 3D in pots with the help of computed tomography (CT) scanning, magnetic resonance imaging (MRI) and positron emission tomography (PET) (Atkinson et al. 2019). In Arabidopsis, including a luminescence trait in a GLO-Root system facilitated imaging and analysis of thin roots (Deja-Muylle et al. 2020).
While assessing root growth under controlled conditions can provide important information, the artificial environment can alter root growth characteristics. Measurement under field conditions will produce data that more closely represents the real-life situation. Phenotyping methods in the field are time-consuming, labor-intensive and highly variable but progress is being made to improve root phenotyping under field conditions (Fradgley et al. 2020; Atkinson et al. 2019). Use of minirhizotrons can help visualize the root system non-destructively (Griffin et al. 2025). Digging up the roots can destroy their architecture, but information on the root architecture can be retained during root excavation by using “Shovelomics”. This involves digging a column around the plant in the field, keeping the roots intact. The root column is carefully washed, again keeping the root system as intact as possible. Imaging techniques are used to score the roots for traits such as root angle (RA), crown root number (CRN), nodal root number (NRN) and seminal root number (SRN) (Fradgley et al. 2020). Convolutional neural networks (CNNs), that are deep learning neural networks that allow a computer to understand and interpret a visual image, can be used for rapid analysis and processing of large numbers of root images.
Once the desired root traits have been identified they must be selected for in the breeding program. Traditionally, breeding for cultivars with the desired root traits would involve making crosses and selecting for the plants that possessed the desired traits. Breeding is a numbers game, and success requires making many crosses and selections. Since phenotyping the plants for root traits is time-consuming, slow and costly, even with new techniques, selecting root characteristics based solely on physical characteristics is problematic (Dinh et al. 2023). Technological advances have made it possible to rapidly and inexpensively determine gene sequences, leading to the application of molecular breeding techniques that allow selection of improved root traits based on genotype assessment (Maharajan et al. 2018). Use of molecular markers to select for plants carrying genes for a desired trait would be more efficient than selecting based on measurement of root traits. This requires the ability to identify specific genetic markers associated with the desired trait.
Determining the genetic factors behind root system development and P uptake efficiency is complex. Root growth is very plastic, changing in response to environmental conditions and changes in sugars, auxins and ethylene (Magalhaes et al. 2017). Root system architecture is influenced by multiple interactions among tens to hundreds of genes, making it difficult to identify specific genes that control it (Wachsman et al. 2015; Magalhaes et al. 2017). However, progress in the collection and analysis of large amounts of genotypic and phenotypic data has improved the ability to deal with this complexity. Using regression models to link genotype with phenotype information can allow the breeder to determine the statistical association between the target trait and single-nucleotide polymorphisms (SNPs) or QTLs and identify candidate genetic loci to use as genetic markers (Wachsman et al. 2015). Single-trait QTL mapping and multi-trait QTL mapping can be used to identify QTLs associated with desired traits using contrasting parental lines (Bernardino et al. 2019). Genome-wide association studies (GWASs) can use lines from different geographical areas that vary widely in their genetic composition to identify significant QTLs and markers associated with traits of interest (Rajamanickam et al. 2024; Deja-Muylle et al. 2020). These techniques can be applied to complex traits that are affected by interactions among genes and environmental factors (Deja-Muylle et al. 2020).
Multi-trait QTL mapping, interval mapping and GWAS techniques have been used to identify potential markers for RSA in a wide range of crop species, including wheat, sorghum, maize, soybean and rice (Dinh et al. 2023; Yuan et al. 2023; Ribeiro et al. 2023; Kaeppler et al. 2000). Several genes controlling lateral root development have been identified in Arabidopsis and rice, while at least six root QTLs have been identified in maize (Bovill et al. 2013). QTLs have been found for early season root growth in wheat seedlings, which is associated with nutrient uptake (Atkinson et al. 2015). Multi-trait QTL mapping identified QTLs that were associated with fine root area, root diameter and grain yield in sorghum (Bernardino et al. 2019). The QTLs found in the sorghum were analogous to those found in maize. GWAS was used to identify QTLs associated with PUE traits in various crops under various P levels (Mikwa et al. 2024). For example, GWAS identified a P-responsive QTL that affected the root system and could be used to select for better P efficiency (Mikwa et al. 2024). The evaluated genes were shown to serve as positive/negative regulators for root branching in the spring. Using GWAS, SNPs and underlying genes were discovered that could be used for marker-assisted breeding programs of high-efficiency root systems in maize, as well as supporting the breeding of high-yielding maize varieties (Moussa et al. 2021).
Root hairs play a pivotal role in the phosphate uptake process as they increase the root uptake surface. Root hairs could be a good target for breeding because there is large genetic variation in root hair architecture (Richardson et al. 2011). Some genetic factors affecting root hair formation in barley have also been identified (Chmielewska et al. 2014). Several major QTLs that affect root hair length and density in maize and common bean have been determined, providing potential for marker-assisted selection (Richardson et al. 2011). GWAS was used to identify genes associated with root hair density and surface in Arabidopsis thaliana under high and low P conditions (Stetter et al. 2015). Loss-of-function mutants showed increased root hair densities, corroborating the role of these genes in the process. Loss-of-function mutants were used to show the specific genes that were involved in the development of root hair density and length (Deja-Muylle et al. 2020; Stetter et al. 2015).
The identification of markers associated with specific characteristics creates the potential for their use in a breeding program. Marker-assisted selection (MAS) has usually focused on genetic markers associated with QTLs that have large effects on a trait. But, rooting traits tend to be quantitative traits controlled by many small-effect QTLs (Sharma et al. 2021). A new method of MAS, genomic prediction (GP), uses thousands of molecular markers that span the entire genome. Machine learning methods are required to handle the dimensional data, which uses markers saturating the whole genome. A multi-QTL model may be sufficient for less genetically complex traits controlled by fewer loci with large additive effects, but for traits controlled by many QTLs with small individual effects, a GP model may be needed for accurate predictions.
Gene editing using CRISP/Cas or TALENS may be used to confirm the role of identified genes and assess the impact that they have on plant function (Ojeda-Rivera et al. 2022). For example, CRISPR/Cas9 was used to alter specific genes and determine their influence on root system architecture and plant response to P deficiency in rice (Lee et al. 2019). Specific editing of genes may be used in the future to create lines with the desired genetic characteristic in a controlled fashion without the product being classified as a genetically modified organism.
6.3. Breeding for Enhanced Mycorrhizal Association
Another strategy used by most plant species to improve access to soil P is to form mycorrhizal associations where the host plant provides fixed carbon to the fungi, and in return the fungi capture P, water and some other nutrients from the soil and transfer them to the plant (Bolan 1991; Grant et al. 2005; Hamel and Strullu 2006; McGonigle et al. 2011; Miller 2000; Monreal et al. 2011; Ryan and Graham 2002; Ryan et al. 2000; Jakobsen 1986; Smith et al. 2011). The fine mycorrhizal hyphae extend far from the root into small soil pores, greatly increasing the effective volume of soil accessed for nutrients and water. Mycorrhizal plants may be able to take up P directly by root epidermal cells and root hairs, and also by the AM fungi that deliver P into the root cortex (Glassop et al. 2005). Considerable variation exists among species and among cultivars in the ability to form mycorrhizal associations (Veršulienė et al. 2024; Grant et al. 2005; Hamel and Strullu 2006; Kirk et al. 2011). It is interesting to note that past plant breeding efforts did not seem to have selected against mycorrhizal colonization, as older cultivars did not have less colonization than modern cultivars (De Vita et al. 2018; Kirk et al. 2011).
Since genetic variability exists in mycorrhizal colonization, selection for this trait within a breeding program is possible (McGrail et al. 2023; De Vita et al. 2018; Kaeppler et al. 2000). In maize, QTLs were identified that influenced mycorrhizal responsiveness and colonization (Kaeppler et al. 2000). GWAS was used in a panel of durum wheat lines to identify QTLs associated with mycorrhizal association (De Vita et al. 2018). Mycorrhizal-specific Pi transporters expressed in cells containing arbuscules have been identified in several species and appear to increase scavenging of P from the cortical apoplast surrounding mycorrhizal structures (Glassop et al. 2005).
While the capability may exist to select for enhanced mycorrhizal colonization, this will not necessarily provide an advantage to the crop. Mycorrhizal association in a plant may lead to an improvement in yield from the interaction or compensation for poor performance when the mycorrhizal association is absent (Maharajan et al. 2018). However, the benefit of mycorrhizal colonization in increasing P uptake and growth over non-mycorrhizal plants is not consistent. Under very low or relatively high P conditions, there may be little to no advantage of colonization and growth rates may even be lower with mycorrhizal association (Ryan and Graham 2002; Ryan and Kirkegaard 2012; Grant et al. 2005; Richardson et al. 2011). At soil P levels needed to optimize crop yield, there would often be little advantage of mycorrhizal colonization. The lack of consistent benefit of mycorrhizal colonization would discourage its use as a selection parameter in a breeding program.
The composition of the root biome may also be affected by the plant and its genetics (Salas-González et al. 2021). Root exudates can influence the rhizosphere microbiome (Dakora and Phillips 2002). Plants can produce compounds that encourage the proliferation of P-solubilizing microorganisms in the rhizosphere (Kucey et al. 1989; Richardson 2001; Richardson et al. 2009). This effect is not as specific as the mycorrhizal interaction, but changes in root exudates may influence the form and activity of the microbial community in the rhizosphere. Changes in root exudation are discussed in section 6.4.
6.4. Rhizosphere Modification
Another strategy that plants may have to increase P access is to increase the concentration of plant-available P at the root surface by increasing the solubility of P in the soil solution. Most Pi that the plant requires is supplied by replenishment of the P in the rhizosphere through mass flow, diffusion and by mobilization of P from various soil P pools (Barber 1980; Barber et al. 1963; Johnston et al. 2014). Increasing the amount of P mobilized from the sparingly soluble P pools can increase P supply to the plant.
Many plants will secrete low molecular weight amino acids that acidify the rhizosphere, compete with phosphate ions for sorption sites, and complex cations bound to phosphate ions, thus increasing P availability (Jungk et al. 1993; Hoffland et al. 1989; Hinsinger 1998, 2001; Hinsinger and Gilkes 1995; Wang et al. 2024; Wang et al. 2020; Wang and Lambers 2020). Production of organic acids, such as malate, citrate, succinate and tartrate, can vary in response to P concentration in the soil solution (McGrail et al. 2023; Lipton et al. 1987; Wang et al. 2016; Wang et al. 2020; Wang and Lambers 2020). There are genetic differences among crop species and cultivars in their ability to produce organic acids or exude protons to enhance P availability (McGrail et al. 2023). Cluster roots that occur in plants like Proteaceae and the Cyperaceae that are adapted to very low Pi conditions tend to secrete large amounts of carboxylates, mainly citrate, to solubilize both inorganic and organic P from the soil (Lambers 2022; Lambers et al. 2003).
Root exudates can also influence the formation of mycorrhizal associations and influence the rhizosphere microbiome (Dakora and Phillips 2002). Plants can produce compounds that encourage the proliferation of P-solubilizing microorganisms in the rhizosphere (Kucey et al. 1989; Richardson 2001; Richardson et al. 2009).
The secretion of root exudates will be affected by their biosynthesis and transport into the rhizosphere (Richardson et al. 2011). Overexpression of genes that are involved in organic anion synthesis in roots is a possible way to increase the concentrations of exudates in the rhizosphere. Tobacco plants modified to express a citrate synthase gene from Pseudomonas aeruginosa were able to mobilise P from an alkaline soil and, under restricted P nutrition, yielded more seed than a control (López-Bucio et al. 2000). Unfortunately, other studies were unable to achieve the same results (Richardson et al. 2011). Manipulation of genes that code for the proteins that transport organic acids might also be an avenue for breeding; however, overexpression of these genes that led to higher efflux did not show a clear benefit for P nutrition (Richardson et al. 2011). The root endodermis and exodermis act as control points for the movement of substances in and out of the root (Salas-González et al. 2021). Genes that affect the root diffusion barriers can influence organic acid efflux and the rhizosphere microbiota.
It may be difficult to increase organic acid exudation to levels that would have a significant effect on P availability. In addition, organic anions are utilized by rhizosphere microbes, and the increased concentration in the rhizosphere may be temporary (Bovill et al. 2013). More information on the integrated effects of root exudates on the rhizosphere biome and P status of crops is needed.
A large proportion of the total soil P can be in organic forms, so increasing its mineralization could potentially increase the P supply for the growing plant. Some plants can excrete enzymes, such as phytase, acid phosphomonoesterase, and acid phosphatase, into the rhizosphere to release P from organic forms (Lefebvre and Glass 1982; Hinsinger 2001; Ashworth and Mrazek 1995), with differences in the type and amount of exudation occurring among plant genotypes (McGrail et al. 2023; Wang et al. 2016). Phosphatases catalyze the hydrolysis of organic P to release free orthophosphate, so increasing the exudation of phosphatases could allow plants to increase P availability. Under P stress, plants may increase the excretion of APase to increase the amount of Pi in the rhizosphere (Wang and Liu 2018). Arabidopsis Purple Acid Phosphatase10 (AtPAP10) is an intracellular acid phosphatase found mainly in the plant root. It is induced upon phosphate starvation and acts against various organophosphate substrates to scavenge Pi to maintain phosphate homeostasis (Francis et al. 2023). Its expression is induced according to the Pi limitations up to the post-transcriptional level. Identification and validation of QTLs for traits like APase could provide a method of breeding for improved availability of soluble P to the plants by the release of Pi from organic pools. Root-associated phosphatase activity was found to vary substantially among wheat genotypes, but did not affect the P nutrition of plants grown in soil (George et al. 2008). Therefore, breeding for enhanced phosphatase activity may be possible, but may not provide enough benefit to make it an attractive breeding factor.
6.5. Summary
For a characteristic to be used in crop breeding, there must be genetic variation and a feasible way of selecting for the trait. There also needs to be a demonstrated advantage for the characteristic that would encourage the breeding program to pursue it in conjunction with the many other characteristics sought for in the breeding program. Advances in phenotyping, genotyping and methods of data handling have made selection for root characteristics feasible. While there has not been a lot of success in this area so far, there appears to be substantial potential for breeding for improved root system architecture and root hair development that could lead to improved ability for the plant to access P from the soil. Breeding for mycorrhizal associations and root exudation is likely possible, but may not have sufficient consistent benefit to include in a breeding program.
Section 7. Genetic Improvement of Root Uptake of Phosphorus
Key Points
- P must move from the soil solution into the plant against a concentration gradient using transporters that actively move Pi across the cell membrane
- Cross-membrane transport is also needed to move P within the plant inside cells, between cells, and between plant organs and organelles to support cellular functions and plant growth
- The genetic regulation of the P transporters involves response at the transcriptional, post-transcriptional, translational, and post-translational levels
- A broader understanding of how genetic factors controlling P transport systems interact in representative environments is needed to be able to effectively manipulate their performance through plant breeding to improve phosphorus use efficiency
7.1. Introduction
The cytoplasm is the metabolic portion of the plant. The concentration of P in the cytoplasm must be closely regulated because both excess and insufficient P concentrations are harmful. Therefore, P homeostasis is critical for plant health. The plant accesses the P that it requires from the soil solution and moves it through the symplasm in the root to the xylem and the aerial parts of the plant. The destination for P taken by the plant is determined by the plant's requirements. The P may be used to synthesize P-containing substances, such as phospholipids and nucleic acids; it may be moved to mitochondria or plastids, where it is used in metabolic processes; it may be moved to other cells where the demand is high, or it may be stored in vacuoles (Jha et al. 2023). The plant controls uptake of P by the root and distribution among cells, tissues, organelles and the vacuole to try to optimize metabolic functions.
Phosphorus in the soil solution at the root surface is at a much lower concentration than Pi in the cell. Therefore, Pi must pass into the cell against a concentration gradient using transporters that actively move Pi across the cell membrane (Ojeda-Rivera et al. 2022). Movement from the xylem to the cell cytoplasm is also against a steep concentration gradient, so it also requires active transport. Therefore, cross-membrane transport is needed not only to acquire Pi from the soil solution but also to move it within the plant inside cells, between cells, and between plant organs to ensure that P is available where needed to support cellular functions and plant growth.
7.2. Regulation of Phosphorus Uptake and Distribution
Phosphorus uptake and distribution within the plant is complex and highly regulated, and while many aspects have been identified, the understanding of the process is still developing. Plants can implement physiological changes in response to P deficiency to enhance the ability of the root to absorb P from the solution (Hodge 2004; Jeong et al. 2015). For example, phosphorus-deficient plants can increase their uptake rate per unit of root (Hodge 2004; Drew et al. 1984; Green et al. 1973; Jungk and Barber 1974; Anghinoni and Barber 1980; Borkert and Barber 1983). These changes can involve changes in the number and activity of the various phosphate transporters.
A suite of phosphorus transporter proteins exists to move P across cell membranes, with both low- and high-affinity transporters being involved in phosphate uptake (Schroeder et al. 2013). Phosphate transporters function in the uptake of Pi from the soil and the distribution of Pi throughout the plant, and in regulating phosphate homeostasis. The high-affinity system tends to function when Pi concentrations are low, improving the plant’s ability to transport the orthophosphate across the root membranes when P supply is restricted. The high-affinity Pi transporters are encoded by PHOSPHATE TRANSPORTER (PHT) genes (Ojeda-Rivera et al. 2022; Wang et al. 2021). They are classified into different families depending on their location and function. The PHT1 family is present on the plasma membrane and involved in the uptake of Pi from the soil solution and/or Pi translocation between cells or tissues (Wang et al. 2021). The PHT2/3/4 families are involved in transporting Pi into or out of subcellular organelles, such as mitochondria, plastids or the Golgi apparatus, while the PTH5 family is involved in the movement of Pi in and out of the vacuole (Wang et al. 2021). As a result, the PHT1-5 Pi transporter proteins help maintain Pi homeostasis by mediating Pi uptake and remobilization of internal Pi among different tissues or organelles (Ojeda-Rivera et al. 2022).
A domain is a compact, distinct functional or structural unit in a protein that contributes to its ability to complete its role. A domain can be associated with a corresponding segment of DNA. A major facilitator superfamily (MFS) domain is found in many transporters. The SYG1/PHO81/XPR1 (SPX) domain-containing proteins are involved in the regulation of Pi signaling and transport. Other identified transporter families include the SPX-EXS subfamily PHO1, which is involved in Pi transport from the root to the shoot and the SPX-MFS subfamily that is involved in vacuolar Pi transport (Wang et al. 2018; Wang et al. 2021; Ojeda-Rivera et al. 2022). The SPX-MFS proteins are considered to be members of the PHOSPHATE TRANSPORTER 5 family (PHT5) and function as vacuolar Pi transporters helping to regulate cytoplasmic Pi homeostasis (Liu et al. 2016). This is particularly important because most of the Pi in the cell is stored in the vacuole, while only 1–5% is present in the cytoplasm. The movement of Pi into and out of vacuoles is very important to buffer cytoplasmic Pi concentrations against fluctuations in external Pi. The PHT5 transporter appears to be involved in Pi movement into the vacuole, while the VPE (Vacuolar phosphate efflux) transporters are involved in the movement of Pi outwards across the tonoplast (Guo et al. 2025). In the study by Guo et al (2025), overexpression of the VPEs increased Pi efflux and movement of Pi from senescing tissue in older leaves to younger leaves, while loss of function decreased remobilization. Loss-of-function mutants for PHT5 accumulate less Pi and have a lower vacuolar-to-cytoplasmic Pi ratio than controls, while overexpression of PHT5 increases Pi sequestration into vacuoles and alters the regulation of Pi starvation-responsive genes. However, the specifics of its coordination to maintain homeostasis need more investigation (Wang et al. 2021).
Mycorrhizal host plants have specific Pi transporters that increase expression when plants are colonised by AM fungi (Glassop et al. 2005). In rice, specific arbuscular mycorrhiza-induced PHT1 are required for the development of the symbiosis (Chang et al. 2019; Yang et al. 2012). Some Pi transporters have evolved that are specifically expressed in infected cells (Jeong et al. 2015).
The production, distribution, activation and degradation of phosphorus transporters in plants are influenced by the P status of the system. Plants respond to both the external and the internal concentration of Pi. Sensing and signaling systems identify and communicate the internal and external P status to the plant (Pan et al. 2022). Local signaling relies on external Pi concentration, while systematic or long-distance signaling depends on the Pi state of the whole plant. Local signaling initiates changes in root system architecture to enhance Pi acquisition, while systemic signaling influences Pi uptake, mobilization, and redistribution. signaling molecules such as sugars, miRNA, and hormones are produced in response to the changes in Pi concentrations (Pan et al. 2022).
When Pi is scarce, plants initiate Phosphate Starvation Responses (PSRs) to increase their ability to access and utilize P efficiently. In relation to the P transporters, these responses influence the production, distribution and activity of the transport proteins. Phosphate Starvation Response 1 (PHR1) serves as a central regulator for Pi- signaling and Pi-homeostasis, and its expression is triggered by the internal concentration of Pi in the plant. Expression of PHT1 is low at high internal P concentrations, while expression is higher when P concentration in the plant declines (Wang et al. 2021). This may indicate upregulation in response to deficiency or downregulation when P is sufficient (Lambers 2022).
The “reading” of the genetic code and use of the information to affect cellular function takes several steps, and the plant can regulate gene functions throughout this process. Gene expression involves transcription and translation to produce the protein that is coded by the gene. The mRNA is the nucleotide sequence that serves as the blueprint for protein production. The mRNA is produced from the DNA of specific genes in the nucleus of the cell in a process known as transcription. The mRNA that comes off from the DNA is like a first draft and must undergo some transformations or editing that can influence its function. In this way, one gene may code for several different forms of a protein. The edited mRNA then moves into the cytoplasm to the ribosomes, where it guides the placement of the amino acids during protein construction, which is known as translation. Transcriptional regulation is the control of the levels of RNA produced during transcription. Post-transcriptional regulation is the control of gene expression at the mRNA level, between the transcription and translation steps. It can involve the maturation of the RNA to an active phase, its transport to the cytoplasm, or its stability. Translational regulation is the control of the levels of protein synthesized from the mRNA. Post-translational regulation is the control of the levels of active protein by things such as phosphorylation of the protein. These various regulation methods interact to influence the concentration and activity of the target proteins. The regulation of the P transporters involves response at the transcriptional, post-transcriptional, translational, and post-translational levels (Pan et al. 2022).
Under P deficiency, plants begin a P starvation response (PHSR1) that regulates the transcription and production of the PHT transporters (Gu et al. 2016). Under low Pi conditions, most phosphate transporters are induced through upregulation of transcription to increase the ability of the plant to access P (Wang et al. 2021; Ojeda-Rivera et al. 2022). Regulation also appears to occur at the post-transcriptional level and post-translational level. The movement of PHT1 proteins to the plasmalemma and their stability are regulated post-translationally by the Pi supply (Poirier et al. 2022). If P is in short supply, movement of P from the endoplasmic reticulum to the plasma membrane is increased through the upregulation of a transporter gene (Wang et al. 2018). The VPE (vacuolar phosphate efflux) transporters are involved in the export of Pi from the vacuole, while the PHT5 transporters are involved in its import (Poirier et al. 2022). The VPE genes are transcriptionally induced by Pi deficiency, but the PHT5 genes are not and may be regulated at the post-translational level. There is also regulation of homeostasis by genes that influence the degradation of Pi-transporter proteins PHT1-4. Proteins have a finite life span, ranging from several minutes to a few hours. The rates of protein production and degradation must be balanced to maintain the proper levels of transporter activity.
MiRNAs (MicroRNAs ) are non-coding small-molecule RNAs that influence the activity of target genes at the post-transcriptional or translational level (Du et al. 2023). Studies in rice, wheat, rapeseed and soybean have shown that miR399 can be specifically induced by Pi deficiency and that it inhibits the breakdown of the Pi transporter protein PHO1 as well as the PHT1 family. The acquisition, translocation, and xylem loading of Pi during P deficiency are enhanced, helping plants to accumulate sufficient Pi under low P conditions.
While it is apparent that plants have evolved complex networks to use the activity of P transporters to regulate P status, the understanding of the system is still evolving (Poirier et al. 2022). Specifically, research is continuing to better define the regulation of transporter function and the potential for genetic manipulation to improve PUE.
7.3. Genetic Manipulation of Transporter Systems
Plant breeders have traditionally taken advantage of natural or induced genetic variation to produce improved cultivars. While there have been large improvements in yield potential, there has not been great progress in improving tolerance to low P or decreasing the need for P fertilizer inputs to maintain crop productivity. The increased understanding of the genetic base for P uptake and its regulation in plants may help to identify specific breeding targets, but currently there is more potential than progress.
Natural variation exists in phosphorus use efficiency (Ouma et al. 2012) and advances in the phenotypic and molecular characterization of germplasm lines can be used to identify many quantitative trait loci associated with low P tolerance (Yang et al. 2024). Genomic techniques such as genome-wide association studies can identify natural genetic variations that influence Pi uptake and homeostasis and could eventually be used to breeding Pi efficient crops (Wang et al. 2018). For example, genome-wide association studies of soybean germplasms pinpointed a gene variant that contributes to P acquisition (Yang et al. 2024). Genome-wide association studies of soybean germplasm also identified a variant in soybean PHF1 as a key determinant for P acquisition.
Mutant and transgenic plants have been used to confirm the role of many of the PHT1 genes, but further information is needed to determine the mechanisms for their regulation and response to Pi deficiency (Wang et al. 2021). Researchers have tried to improve phosphorus use efficiency by transforming Pi-related transcription factors, phosphate transporters, and their interacting proteins, with varying success. Transgenic rice plants have been produced that overexpress several of the transport facilitator genes that influence the movement of the P transporter proteins to the plasmalemma, leading to both increases and decreases in plant Pi uptake and yield. Other transgenic rice plants with a modified form of the OsPHT1;8 gene showed greater Pi uptake. So, it may be possible to improve PUE and crop yield when Pi is restricted by targeting Pi transporters (Wang et al. 2021). The use of genome editing techniques such as CRISPR/Cas9 to modify Pi transporters and regulatory genes in crop plants may also be possible.
Manipulation of transporter proteins has often led to side effects associated with Pi toxicity. Overexpression or mutation of phosphorus transport-related genes can give rise to excessive Pi accumulation, especially when P supply is not deficient, leading to toxicity and yield decline (Wang et al. 2021). Problems may also occur if the targeted genes have other functions unrelated to their role in P transport that may be compromised by manipulation (Gu et al. 2016). Therefore, while manipulation of the genes influencing Pi transport holds potential, a better understanding of their function and potential unintended consequences is needed to effectively apply this in a practical breeding program.
7.4. Summary
There has been a huge increase in the delineation of phosphorus uptake and transport in plants and the genetic factors controlling it. However, there are still limitations in the understanding of how these genes operate to regulate P homeostasis in the plant. Uptake and distribution of Pi must be coordinated with that of other nutrients that are required for optimizing plant growth and development. Therefore, the impact of genetic manipulation on the interaction between P and other nutrients must be considered when attempting to manipulate the genetics of phosphorus use efficiency.
Much of the work defining the system has been done using model plants such as Arabidopsis grown under highly artificial conditions. More work needs to be done evaluating traits in crop plants under field conditions to effectively integrate the knowledge. A broader understanding of how genetic factors controlling P transport systems interact in representative environments is needed to be able to effectively manipulate their performance through plant breeding to improve phosphorus use efficiency.
Section 8. Improvements in Physiological Efficiency of Phosphorus Use
Key Points
- Plant strategies to improve physiological phosphorus use efficiency are under genetic control
- Most P in a well-nourished plant is stored in the vacuole, and remobilization of inorganic P (Pi) from the vacuole can mitigate short-term Pi deprivation and prevent Pi starvation
- Organic P in the plant is present as nucleic acids (mainly RNA) > phospholipids > P-esters > DNA > phosphorylated proteins
- Plants sense P supply locally at the root tip or systemically by the concentration of substances, such as inositol pyrophosphates and microRNA 399
- Some plants can replace the P-containing phospholipids with lipids that do not contain P to free up Pi for other metabolic uses
- Substituting enzymes that use inorganic pyrophosphate rather than ATP and Pi to catalyze critical reactions or by-passing Pi- or ATP-demanding enzymes and/or metabolic pathways could conserve Pi when it is limiting
- Pi can be mobilized and recycled from older to younger vegetative tissues to support growth and improved photosynthetic capacity
- RNA is a large reserve of Pi that can potentially be remobilized.
- Reducing phytate concentration in the seed can reduce P removal from the system and potentially reduce P moving into waste streams
- A greater understanding of physiological regulation and interactions of P in the plant and of genetic by environment interactions under field conditions is needed
8.1. Introduction
When the external supply of Pi is limited and uptake is restricted, plants initiate various strategies to improve the internal phosphorus use efficiency. Internal P use efficiency is the ability of the plant to produce biomass with the limited P that it has available. Phosphorus-efficient cultivars can produce more biomass at a lower tissue concentration of P than less efficient cultivars. Only a small proportion of the P in a well-nourished plant is metabolically active, with most of the P being stored in the vacuole. Organic P in the plant is present as nucleic acids (mainly RNA) > phospholipids > P-esters > DNA > phosphorylated proteins (Veneklaas et al. 2012; Dissanayaka et al. 2018). Improving internal P use efficiency involves diverse strategies to direct available P to support priority metabolic functions that can maintain growth and reproduction. Such strategies may include reducing the concentrations of inorganic P in vacuoles, enhancing mobilisation of P from metabolically inactive to active tissue, reducing the investment of Pi in plant components, such as ribosomal RNA or phospholipids, or decreasing the concentration of Pi present as phytic acid in the seed.
8.2. Sensing of P Status
For a plant to respond to Pi deficiency, it must have a way to sense the Pi status of the system. The Pi status of the system can be sensed locally or systemically (Yang et al. 2024). Local sensing of Pi status occurs at the root tip, where the root cap protects the root apical meristem (Abel 2017). The concentration of Pi in the soil solution at the root tip serves as a trigger to influence root growth and architecture, restricting primary root growth and encouraging root hair development. Phosphate availability influences the root system architecture by adjusting meristem activity. When Pi is deficient, plants over-accumulate Fe in the roots and shoots. The full process not fully understood, but it is known that LOW PHOSPHATE ROOT 1 (LPR1), LPR2, and PHOSPHATE DEFICIENCY RESPONSE 2 (PDR2) are involved in root Pi sensing. When Pi is limited, the activity of the two interacting genes LPR1 and PDR2 leads to Fe and callose deposition in the meristem and elongation zone of primary roots. Antagonistic interactions of Pi and Fe availability will restrict primary root growth by promoting callose formation in the meristems, where cell reproduction would otherwise occur (Müller et al. 2015).
Several different systems have been identified in systemic sensing of Pi status. Inositol polyphosphates (InsPs) are Pi storage and signaling molecules that play a role in hormonal signaling, energy sensing, and responses to biotic and abiotic stresses (Pan et al. 2022). InsPs are highly phosphorylated and can be further phosphorylated to generate inositol pyrophosphates (PP-InsPs), which contain one or more high-energy pyrophosphate groups. The binding of PP-InsP to proteins that contain the SPX domain is an important step in cell Pi sensing and activation of Pi deficiency responses (Poirier et al. 2022; Pan et al. 2022). The enzymes involved in PP-InsP synthesis are regulated by the cell ATP/ADP ratio, which depends on the cell Pi status. The enzymes have two functions, firstly in kinase and ADP phosphotransferase reactions where a phosphate group is transferred from ATP to a protein and secondly in phosphatase activities where the phosphate is removed from a protein (Poirier et al. 2022). If Pi is high, the high ATP/ADP ratio stimulates the kinase reaction, leading to high InsP levels, while the low ATP/ADP ratio that occurs when Pi is low favors the dephosphorylation reaction, leading to low InsP. The InsPs bind to the SPX proteins involved in the Pi starvation responses. If Pi levels are sufficient, there is a large amount of PP-InsP in the system that binds to the SPX proteins and blocks the binding of PHR to its target promoter. If Pi is low, the SPX protein cannot bind PHR because of low PP-InsP levels, and the genes involved in the P deficiency response are activated at the transcriptional level. There are also other pathways where the PP-InsP levels that are affected by Pi status may interact directly or indirectly with proteins to influence P acquisition and efficiency, and a full understanding of the system is still developing.
Another key signaling system involves microRNA 399 (MiR399). Under Pi deficiency, synthesis of miR399 in the shoot is upregulated under control of the PHR1 transcription factor (Poirier et al. 2022). The miR399 is transported from the shoot to the root through the phloem, where it targets the plant’s miRNA pathway to suppress the transcription of the PHO2 enzyme that is involved in PHO1 and PHT1 protein degradation in roots (Du et al. 2023; Poirier et al. 2022). When miR399 suppresses PHO2, it prevents the degradation of these transporter proteins. Therefore, increasing miR399 increases transporter protein activity and enhances Pi acquisition, translocation, and xylem Pi loading. This in turn increases phosphate uptake and transport from the roots to the shoots, contributing to Pi homeostasis.
It may be possible to modify some of the key genes involved in P signaling processes by using traditional mutagenesis techniques or molecular tools, such as transgenesis or CRISPR/Cas9, editing to modify the plant’s ability to sense and respond to Pi deficiency. Influencing signaling could enhance the plant’s P starvation response in terms of root development, P acquisition and internal physiological P efficiency. However, progress so far has been limited. There is a relatively good understanding of which genes are involved with P uptake, but a less complete understanding of the interactive signaling and control between and within cells. Knowledge and understanding of the physiological and genetic factors affecting the P deficiency response is increasing rapidly; however, the ability to actively influence this in a breeding program is becoming more likely.
8.3. Mobilization from the Vacuole
The vacuole is the main storage organ of the plant. When Pi levels are high, the plant can store surplus Pi in the vacuole as a reserve. When Pi levels are sufficient, as much as 80% of the total cellular Pi may be present in the vacuole. If the Pi that the plant can access declines, Pi can be mobilized from the vacuole to maintain cytoplasmic Pi concentration (Poirier et al. 2022). Therefore, movement of Pi across the vacuolar membrane (tonoplast) is critical in maintaining Pi homeostasis. The PHT5 transporters are involved in the import of Pi to the vacuole, while the vacuolar phosphorus efflux (VPE) transporters are involved in the movement of Pi outwards across the tonoplast. The VPE genes are transcriptionally induced by Pi deficiency, while the PHT5 genes are not and may be regulated at the post-translational level (Poirier et al. 2022). Overexpression of the VPEs increases Pi efflux from the vacuole and movement of Pi from senescing tissue in older leaves to younger leaves, while loss of function decreases remobilization (Guo et al. 2025).
Remobilizing Pi from the vacuole can mitigate short-term Pi deprivation and prevent Pi starvation (Dissanayaka et al. 2018). If Pi deficiency persists, the Pi pool in the vacuole can become depleted, and remobilization will no longer be sufficient to support Pi homeostasis, so that the cytoplasmic Pi levels will decline. The plant will respond by modifying its metabolism and mobilizing Pi from different Po pools to redistribute P to support plant growth and development (Veneklaas et al., 2012).
8.4. Modification of Phospholipids
A major portion of the P in the plant is used in the manufacture of the phospholipids that compose the cell membranes. Phospholipids represent about 25% of the organic P in a mature leaf (Veneklaas et al. 2012). Remobilization of this pool from senescing leaves would provide a significant reserve of Pi.
Under P stress, some plants can respond to Pi deficiency by replacing some of their P-containing phospholipids with galactolipids and sulfolipids that do not contain P (Navea et al. 2024; Lambers et al. 2012). This substitution frees up Pi for other metabolic uses and may allow the plant to sustain photosynthesis when P supply is restricted (Poirier et al. 2022; Lambers 2022). It appears that replacement of phospholipids occurs more in older tissue than in younger tissue in a number of plants, including Proteaceae, soybean and rice (Dissanayaka et al. 2018). It may be that a higher proportion of phospholipid is needed in young tissue during leaf growth and development to maintain membrane integrity for cell division, elongation, and expansion.
The genes that code for the phospholipases and lipid biosynthetic enzymes involved in this substitution are strongly upregulated at the transcriptional level when Pi is deficient (Dissanayaka et al. 2018; Navea et al. 2024). This response may be a potential target for genetic improvement in internal P use efficiency.
8.5. Energy Reactions
Phosphorus is a component of ATP, the critical energy storage molecule in the plant. As discussed in section 8.2, the cell’s ATP/ADP ratio, which depends on the cell’s Pi status, plays an important role in sensing and responding to Pi deficiency.
Under long-term P deficiency, the levels of Pi and ATP in the cell decrease, but the inorganic pyrophosphate (PPi) levels remain fairly constant (Poirier et al. 2022). Internal phosphorus use efficiency could potentially be increased by substituting enzymes that use PPi rather than ATP and Pi to catalyze critical reactions (Poirier et al. 2022).
Plants may also try to bypass Pi- or ATP-demanding enzymes and/or metabolic pathways to conserve Pi when it is limiting (van de Wiel et al. 2016). For example, in carbohydrate metabolism, several phosphorylated intermediates such as triose-P (glyceraldehyde-3-phosphate) are produced during carbon fixation (van de Wiel et al. 2016). In sugar beet, under low Pi, starch in the plastids and sucrose in the cytosol increase as the triose-P levels decline. The potential for genetic modification of these processes to improve PUE has not been explored to any great extent.
8.6. Recycling of P
Phosphorus recycling within the plant is an important method of optimizing physiological PUE (Veneklaas et al. 2012). Older leaves are often shaded and are less photosynthetically active than younger tissue. Pi can be mobilized and recycled from older to younger vegetative tissues to support growth and improved photosynthetic capacity. Under Pi deficiency, there is initially greater retention of Pi in the root to encourage the production of a larger, finer root system that can increase the ability of the plant to access Pi from the soil (See Section 6). As the plant ages, it begins to rely more on internal remobilization of Pi and less on Pi newly accessed from the soil. The Pi present in the root systems, well as in older leaves may then act as a reserve to be used to support Pi homeostasis and growth in the above-ground tissue. As the plant matures and moves into reproductive stages, Pi can be remobilized again, moving from vegetative tissue to the seed to optimize reproductive efficiency. This remobilization of material from the vegetative tissue to seeds is critical for the creation of yield in cereal, oilseed and legume crops.
Plants contain intracellular hydrolytic enzymes that scavenge and recycle intracellular organic P to recover Pi from cells that are losing their usefulness and move it to actively growing tissue where Pi would be more effective (Veneklaas et al. 2012). As plant tissue senesces, the scavenging activity increases to move the Pi to sustain the growth of active issue and storage or reproductive tissue so that the Pi is not lost from the plant when the leaves drop. Phosphorus remobilization becomes particularly important when Pi is deficient.
A greater understanding of the control of Pi reallocation from senescing to actively growing tissue could identify areas where plant breeding could be effective. For example, preferential movement of Pi to photosynthetic mesophyll cells rather than the epidermal cells may be a reason for the high photosynthetic activity of Proteaceae under very low leaf Pi concentrations (Poirier et al. 2022).
About 40% of the organic P pool in the plant is contained in nucleic acids, with a large part of that in ribosomal RNA (Poirier et al. 2022; Veneklaas et al. 2012). Therefore, it serves as a large reserve of Pi that can potentially be remobilized. The RNA plays a key role in protein synthesis, and restrictions in rRNA content and therefore a reduction in ribosome number and protein synthesis capacity can lead to a reduction in growth rate (Dissanayaka et al. 2018; Veneklaas et al. 2012). However, remobilization of Pi from RNA during senescence is an important source of Pi. Purple APases (PAPs) and RNases function in the recycling of Pi from senescing leaves to growing organs and seeds (Dissanayaka et al. 2018). For rice, genes related to RNA degradation are upregulated in the flag leaf during the grain-filling stage, leading to the release of and transport of Pi to developing grains.
The nucleases and phosphatases involved in breaking down ribosomal RNA are induced by both Pi deficiency and senescence. Many phosphatases that are involved in the degradation of mitochondrial and plastidial DNA are strongly upregulated under Pi-deficient conditions and during senescence. In rice, a ribonuclease (RNS) gene family is involved in Pi recycling through RNAse activity. Under Pi deficiency, overall RNA content in both shoot and root tissues was reduced while RNase activity was increased (Navea et al. 2024). In response to Pi starvation, plants typically secrete PAPs that hydrolyze Pi from a wide range of Pi monoesters (Navea et al. 2024).
The genes influencing P recycling may be a potential target for breeding for improved P-efficient genotypes. A greater understanding of the functional activity of the genes involved in remobilization and their broader influence on plant function is needed. Field validation of the agronomic performance of modified plants in P-deficient situations is also needed to fully understand their performance in a realistic growth environment.
8.7. Regulation in Other Organelles
Other transporters exist that move Pi across the membranes of organelles, such as the Golgi apparatus, mitochondria, and thylakoids (Poirier et al. 2022). Coordinated transport of Pi across various tissues and organelles is important in the internal regulation of Pi, but knowledge of the regulation of the Pi transporters involved in these processes is limited. More research targeted at understanding transporter function at the organelle level is needed before this becomes a realistic target for breeding activities.
8.8. Phytate Level in the Seed
Seedlings depend on seed stores for Pi during early growth, so an adequate level of seed P is important for early-season seed vigor (White and Veneklaas 2012). Seed P in cereal crops is primarily stored as phytate, which is poorly metabolized by humans and monogastric animals (Veneklaas et al. 2012). The plant needs some Pi to be stored as phytate to supply the P required for germination and seedling establishment, but seed P concentration often exceeds these requirements, particularly if the plants were grown under P-sufficient conditions (Veneklaas et al. 2012; White and Veneklaas 2012). Much of the P in the diet of livestock and humans is excreted, and the high P content of manures and sewage can lead to environmental problems if not carefully managed. It may be possible to decrease the amount of P stored in the seed without negatively affecting either seed vigor or nutritional quality (Rose et al. 2013). Concentration of P in the seed may be less important in promoting seedling vigor in agronomic systems where P fertilizer is placed near the seed-row.
An evaluation of P efficiency in maize lines suggested that P-efficient accessions were able to produce high grain yield with low grain P uptake (Bayuelo-Jiménez and Ochoa-Cadavid 2014). Seed P concentrations have already been observed to have decreased with breeding efforts to increase harvest index (HI) in grains, but strategies aimed at improving PUE by drastically bringing down seed phytate levels often affect seed vigour. A low phytate barley cultivar registered in Canada had phytic concentrations 60-65% lower than conventional varieties, but its yield was substantially lower than that of other cultivars (Rossnagel et al. 2008). The low phytate characteristic was related to the presence of a specific lpa3-1 gene. Evaluation of a range of low phytate lines produced in maize and barley showed that in slightly more than half of pairs, the low phytate trait produced lower grain yield (Raboy 2002). Since some of the lines with reduced phytate levels produced adequate grain yield, there appears to be significant potential for the production of low phytate lines without reducing crop yield and quality.
In rice, SULTR-like Pi DISTRIBUTION TRANSPORTER (SPDT) controls Pi allocation to the grain (Ojeda-Rivera et al. 2022). Knock-out lines reduced phytate content in grain without reducing yield and seed germination, while knocking out kinase-coding genes in several crops, including maize, rice, and wheat, also produced plants that accumulated less phytic acid in the seed. Both total P and phytate in the brown de-husked rice were reduced by 20–30% in the knockout lines of SPDT, while yield, seed germination, and seedling vigour were not compromised (Dissanayaka et al. 2018; Yamaji et al. 2017). CRISPR/Cas 9 has also been used to alter an allele in barley and produce barley lines that were low in phytate (Vlčko and Ohnoutková 2020). Therefore, there appears to be good potential to breed for lower phytate crops without necessarily having a negative effect on seedling vigor, crop yield or nutritional quality. Field proofing would be necessary to ensure that the agronomic performance of the cultivar was not compromised.
8.9. Use of Phosphite
Phosphite (Phi or PO33−) is a reduced form of P that can be taken up by plants through Pi transporters (Manna et al. 2016). It is a secondary manufacturing product derived from industries, such as electronics, pharmaceuticals, and automotive, that use sodium hypophosphite as a reducing agent in activities, such as nickel plating and polishing (Navea et al. 2024). Plants are normally unable to metabolize Phi, and its uptake can lead to severe growth inhibition and death. It is sometimes used as a selective herbicide and may suppress some plant diseases (Lim et al. 2013). Phi is thought to disrupt Pi metabolism by interfering with enzymes involved in the Pi starvation response. Some bacteria can metabolize Phi by using an enzyme known as Phi dehydrogenase to convert Phi into Pi. Transgenic rice, maize and cotton were produced that contained the bacterial gene for Phi breakdown and were able to maintain normal yield when fertilized with phosphite (Nahampun et al. 2016; Ojeda-Rivera et al. 2022; Manna et al. 2016; Pandeya et al. 2018). Phosphite fertilization may be able to support the growth of the transgenic plants while also acting as a herbicide, since the phosphite can suppress growth of weeds and diseases (Manna et al. 2016; Pandeya et al. 2018). This technology may be promising, but there may be concerns about the ecological impact of adding phosphite to soils (Ojeda-Rivera et al. 2022).
8.10. Summary
There are many areas where alteration in the genetics of internal P physiology may be targeted to improve P use efficiency. However, much of the information available comes from controlled environment experiments with plants grown in soils that are severely P-deficient. Understanding of how these characteristics may interact under more realistic field conditions is limited. Gene targets are not clearly defined because our understanding of the physiological underpinnings of phosphorus use efficiency and its interaction with other physiological processes is not complete. Many factors could influence PUE via complex interactions among different metabolic processes. Many of the genes that may be targeted are involved in a range of important growth functions within the plant, and their manipulation may have unintended consequences. While studies are increasing knowledge of P physiology in the plant, there are still large gaps in the understanding of its regulation and interaction with other plant functions. Little data has been collected under field conditions, and more information is needed on genotype by environment interactions under varying conditions and differing P supplies.
Despite the current limitations, the rapid advancement in molecular and genomic techniques is providing an opportunity to greatly increase understanding of the genetic basis of physiological PUE and identify target genes for breeding programs. There is likely to be considerable progress in this area in the near future.
Section 9. Fitting Genetic Technology into a Sustainable Phosphorus Management System
Key Points
- The 4Rs of source, rate, time and place interact with one another, with other agronomic management practices, and with economic, environmental and social goals
- Phosphorus supply should be balanced with phosphorus removal over the long term to avoid excess depletion or accumulation
- Matching root system architecture with P distribution in the soil and fertilizer management systems may improve system efficiency
- Reduction in phytic acid concentration in the seed could reduce P removal and the risk of damage from excess P in manures and wastes
9.1. Sustainable Phosphorus Management
Agricultural production must support food security and economic production for an expanding world population, while addressing climate change and degradation of land, air and water resources. An ideal production system would optimize yield potential as determined by the fundamental limiting factors of light, water and length of growing season while avoiding environmental degradation.
Sustainable nutrient management is one part of this process. The fertilizer industry has adopted the 4R nutrient management approach as a method of communicating requirements for sustainable nutrient management. This approach refers to the use of the right rate, source, timing and placement of nutrients to meet economic, environmental and social objectives. The goal of 4R management is to provide the optimum amount of nutrients to the growing crop at the time it is required, in the most cost-efficient manner, with the least environmental risk (IPNI 2016 ). From a P management viewpoint, sufficient P should be available for crop uptake at the proper time to optimize yield potential with P inputs and off-take balanced to avoid either excess accumulation or depletion.
An effective 4R management program for P fertilization will deliver maximum overall benefits only if all the “pieces” in the management system “puzzle” are optimized to fit properly with each other (Figure 5). The overall production system, including agronomic management practices and crop rotation, needs to be optimized for the specific conditions of each individual field. This requires identifying the most limiting factors in the system and managing the system to use them most efficiently.
9.2. Role of Plant Genetics in Sustainable P Management
Plant genetics is one tool that can be used to improve the sustainability of a crop production system. Plants have evolved over millennia to interact with their environment for growth and survival. Part of that adaptation was the development of strategies to improve PUE to optimize the chances for survival and reproduction in a P limited environment. Agricultural systems can utilize additional genetic adaptations to further improve productivity and efficiency.
Different agricultural systems have different limitations. A major consideration is the available P present in the soil. There are many locations where P has accumulated in the soil over time, either from high rates of P fertilization or from excess application of manures and municipal biosolids. Growth of plants that can extract this P from the soil in a timely fashion would be beneficial from an environmental perspective. Other soils may have low P fertility, and producers may have limited access to P fertilizer to augment the supply. In some cases, environmental concerns about water quality may restrict producers’ ability to increase soil P fertility. Plants with the ability to produce high biomass yields despite low P fertility would be helpful in these situations. Therefore, the genetics of the crop sown should match the environmental constraints, the management practices and the 4R fertilizer management strategy.
The genetic characteristics of the plant must address the goals and constraints of the production system for which they are targeted. Genetic manipulation can increase phosphorus use efficiency by targeting plant rooting and root architecture. The ideal rooting system will vary depending on the constraints of the system. In soils with an accumulation of P at the surface, shallower root angles and surface root proliferation may be beneficial to encourage efficient topsoil foraging (Oo et al. 2024; Fujii 2024; Lynch 2022; Lynch 2007b; Lynch and Brown 2001). In contrast, in soils where P is more uniformly distributed, such as in puddled rice or in non-fertilized systems, deeper rooting may allow uptake of P from throughout the soil profile (Oo et al. 2024; Manske et al. 2000; Lynch 2013). Similarly, in arid regions where the soil surface dries out, deeper rooting may be beneficial to allow for continued P uptake from nutrient reserves deeper in the soil.
Plant growth may be constrained on soils not because there is inadequate P present in the soil, but because the plant's ability to access sufficient P early in the season is limited. Phosphorus uptake by the plant may be restricted by slow root growth that limits uptake or by dry or cool conditions that limit P mobility. Genetic improvements that encourage early-season root growth and proliferation can enhance early-season P uptake.
Plant root architecture should be selected to complement fertilizer placement strategies. Band placement of fertilizer P near the seed can allow the plant to contact the P early in the growing season. The ability of the plant to proliferate its roots in the fertilizer reaction zone would improve plant uptake of P from the band and improve PUE. Plants with the ability for rapid germination and early-season root development would be better able to access available water and nutrients in environments that dry out over the season. Under dry conditions, deep placement of P combined with selection of a cultivar with deeper rooting could allow P uptake to continue from moist soil deep in the profile throughout the growing season.
The ability of the plant to form mycorrhizal associations or encourage the proliferation of specific P solubilizing organisms in the rhizosphere can also be affected genetically. Mycorrhizae or P-solubilizing microorganisms could theoretically aid in accessing P from soils that contain a moderate supply of P. These strategies are attractive for organic production systems, which are frequently limited by P deficiency and where applications of soluble inorganic P fertilizers are not permitted. However, the benefits of such associations may be limited where the soil is high in P and able to supply the plant adequately without microbial interactions. Benefits may also be limited on soils where P is low, and the microorganisms may compete with the plant for available P. Microbial interactions only increase uptake of the P that is present in the soil, and increased soil uptake will speed soil depletion and reduce soil fertility in the long-term when P removal is more than P addition.
A more sustainable production strategy could be to develop plants that are more physiologically efficient in accessing and utilizing P for crop production with higher biomass production per unit of P in the system. Mobilization of P from older tissue and its movement to younger photosynthetically active tissue and from there to reproductive tissue would reduce the total amount of P required to optimize crop yield. A “tighter” P utilization system would improve the sustainability of production.
Off-take of P from the system could be reduced by decreasing the phytate concentration in the seed. This would be beneficial both because the amount of P required to offset crop removal would be reduced and the amount of P moving into waste streams would be reduced as well.
Regardless of what strategies are implemented to improve phosphorus use efficiency, balancing long-term phosphorus uptake with off-take is required to avoid excess accumulation or depletion of soil P.
9.2. Summary
Efficient phosphorus management is important to help agriculture meet the demands of a growing world population, as well as food security in the face of climate change and degradation of natural resources. Plant genetic characteristics should be used to complement nutrient management practices and site-specific field characteristics to optimize phosphorus use efficiency and agricultural productivity.
Section 10. Summary and Need for Future Work
Key Points
- Modern breeding techniques can shorten the time and reduce the costs for developing improved cultivars, and allow breeders to select for characteristics that are not normally found in the population
- Root growth and architecture are key targets for breeding to improve PUE
- Uptake of P from the soil solution and its distribution across cell membranes throughout the plant against a concentration gradient is facilitated by transporter proteins whose production and function may be improved through plant breeding
- The ability of the plant to mobilize and adjust the distribution of P among the various P forms present in the plant to maintain homeostasis is under genetic control and may be a target for breeding
- Reduction of phytate concentration in seeds could reduce P removal from the field and decrease P concentration in manure and sewage, improving both environmental sustainability and phosphorus use efficiency
- Mycorrhizal associations and plant root exudates could be modified by plant breeding; however, benefits may not be consistent enough to justify inclusion in a breeding program
- A broader understanding of how genetic factors controlling PUE interact in representative environments is needed to be able to effectively manipulate their performance through plant breeding
10.1. Summary
Plant breeding has played an important role in the improvement of crop yield, agronomic characteristics, disease resistance and nutritional quality. Traditional plant breeding relied on the selection of superior physical characteristics that were present in the natural population. For a characteristic to be used in crop breeding, there must be genetic variation and a feasible way of selecting for the trait. There also needs to be a demonstrated advantage for the characteristic that would encourage the breeder to pursue it in conjunction with the many other characteristics sought for in the breeding program.
Advances in phenotyping, genotyping and methods of data handling have expanded the potential characteristics that can be successfully manipulated through plant breeding to improve PUE. New molecular techniques can generate genetic diversity by moving genes between unrelated species or precisely editing genes. Rapid assessment of genetic and phenotypic characteristics in a plant and the ability to statistically determine the relationship between genetic regions and desired traits can help plant breeders more quickly and accurately select for improved crops. Molecular techniques can also allow the breeder to make selections early in the breeding process at the molecular, cellular, or tissue level. These techniques can shorten the time, reduce the costs of developing improved cultivars and allow for the insertion of characteristics into a species that are not normally found in the population.
Breeding programs may be able to target genetic factors that influence the ability of the plant to mobilize, access and utilize soil P and thus increase the PUE of plants. Uptake of P from the soil solution is a function of the absorbing area of the root and the concentration of Pi in the soil solution at the root surface. When P is deficient, root architecture changes to increase root surface area and soil exploration for P. Improvements in the ability to measure root growth and root system architecture, combined with molecular breeding techniques that can identify the genes that control rooting, have made it more feasible to breed for root characteristics that increase PUE. While there has not been a lot of success in this area so far, there appears to be substantial potential for breeding for improved root system architecture and root hair development that could lead to improved ability for the plant to access P from the soil.
Plants may form mycorrhizal associations to increase the volume of soil explored and so improve the ability of the plant to access soil P. While mycorrhizal association is under genetic control, increased mycorrhizal colonization may not always provide a benefit to the plant (Ryan and Graham 2002; Ryan and Kirkegaard 2012). Similarly, increased root exudation of substances that enhance Pi concentration in the rhizosphere is under genetic control, but the transient nature of the exudates and their variable performance may limit their usefulness as a breeding target. Therefore, while breeding for mycorrhizal associations and root exudation is possible, these characteristics may not have sufficient consistent benefit to include in a breeding program.
Plants take up P as orthophosphate ions at the root surface against a concentration gradient in a process facilitated by transporter proteins. Transporter proteins also function in the distribution of Pi throughout the plant to the sites where it is required to support plant metabolism. There has been great progress in defining the process of phosphorus uptake and transport in plants and the genetic factors involved. However, there are still limitations in the understanding of how these genes operate to regulate P homeostasis in the plant. Mutant and transgenic plants have been used to confirm the role of many of the genes controlling transporter production and function, but further information is needed to determine the mechanisms for their regulation and response to Pi deficiency. It may be possible to improve PUE and crop yield when Pi is restricted by targeting Pi transporters (Wang et al. 2021). The use of genome editing techniques, such as CRISPR/Cas9, to modify Pi transporters and regulatory genes in crop plants may also be possible. However, the manipulation of transporter proteins has often led to side effects associated with Pi toxicity. Overexpression or mutation of phosphorus transport related genes can give rise to excessive Pi accumulation, especially when P supply is not deficient, leading to toxicity and yield decline (Wang et al. 2021). Problems may also occur if the targeted genes have other functions unrelated to their role in P transport that may be compromised by manipulation (Gu et al. 2016). Uptake and distribution of Pi must be coordinated with that of other nutrients that are required for optimizing plant growth and development, so the impact of genetic manipulation on the interaction between P and other nutrients must be considered. Therefore, while manipulation of the genes influencing Pi transport holds potential, a better understanding of their function and potential unintended consequences is needed to effectively apply this in a practical breeding program. A broader understanding of how genetic factors controlling P transport systems interact in representative field environments is also needed to be able to effectively manipulate their performance through plant breeding to improve PUE.
Phosphorus is present in the plant as inorganic P, metabolic P or P-esters, P in phospholipids and P in nucleic acids. Surplus Pi is stored in the vacuole and can be mobilized to maintain cytoplasmic Pi concentration when Pi supply is low. Genetic manipulation of factors controlling the mobilization of P from the vacuole could increase physiological PUE. Mobilization of P from organic P forms present in aging plant tissue may be another breeding target. As plant tissue ages, it becomes less physiologically efficient. Plants contain intracellular hydrolytic enzymes that scavenge and recycle intracellular organic P to recover Pi from cells that are losing their usefulness and move it to actively growing tissue where Pi would be more effective (Veneklaas et al. 2012). Plant breeding could target genes that control Pi reallocation from senescing to actively growing tissue to increase the efficiency of P remobilization. However, a greater understanding of the functional activity of the genes involved in remobilization and their broader influence on plant function is needed.
The major storage form of Pi in seeds is phytic acid, and excess phytic acid in the diet can cause environmental issues when it produces high P levels in manures and sewage. Reduction of phytate concentration in seeds could reduce P concentration in manure and sewage, improving both environmental sustainability and phosphorus use efficiency. While low phytate cultivars are often associated with decreases in seedling vigour and crop yield, there seems to be potential to develop low phytate crops without these drawbacks. Therefore, breeding for low phytate crops appears to be a priority goal.
There are many areas where alteration in the genetics of internal P physiology may be targeted to improve P use efficiency. However, gene targets are not always clearly defined because the physiological underpinnings of phosphorus use efficiency and its interaction with other physiological processes are not entirely understood. Many factors could influence PUE via complex interactions among different metabolic processes. Many of the genes that may be targeted are involved in a range of important growth functions within the plant, and their manipulation may have unintended consequences. While studies are increasing knowledge of P physiology in the plant, there are still large gaps in the understanding of its regulation and interaction with other plant functions. Another issue is that much of the work defining the system comes from controlled environment experiments with model plants, such as Arabidopsis, grown under highly artificial conditions in soils that are severely P-deficient. Response of plants may vary substantially between growth chamber and field conditions (Gunes et al. 2006). There are limited data collected under realistic field conditions, and more information is needed on the genetic and environmental interactions under a range of agronomic systems. More work needs to be done to evaluate traits in crop plants under field conditions to effectively integrate the knowledge.
Despite the current limitations, the rapid advancement in molecular and genomic techniques is providing an opportunity to greatly increase understanding of the genetic basis of physiological PUE and identify target genes for breeding programs. There is likely to be considerable progress in this area in the near future. However, field validation of the agronomic performance of modified plants in P-deficient situations is critical to fully understand their performance in a realistic growth environment.
References
- Abel S (2017) Phosphate scouting by root tips. Current Opinion in Plant Biology 39:168-177. [CrossRef]
- Ajiboye B, Akinremi OO, Hu Y, Flaten DN (2007) Phosphorus speciation of sequential extracts of organic amendments using nuclear magnetic resonance and x-ray absorption near-edge structure spectroscopies. J Environ Qual 36 (6):1563-1576. [CrossRef]
- Ajiboye B, Akinremi OO, Hu Y, Jürgensen A (2008) XANES speciation of phosphorus in organically amended and fertilized Vertisol and Mollisol. Soil Sci Soc Am J 72 (5):1256-1262. [CrossRef]
- Akhtar S, Rao E, Uike A, Saatu M (2023) Plant Breeding Strategies: Traditional and Modern Approaches. In: Genetic Revolution in Agriculture: Unleashing the Power of Plant Genetics vol 1. Elite Publishing House, Delhi, pp 21-43.
- Amiteye S (2021) Basic concepts and methodologies of DNA marker systems in plant molecular breeding. Heliyon 7 (10):e08093. [CrossRef]
- Anghinoni Ia, Barber S (1980) Phosphorus influx and growth characteristics of corn roots as influenced by phosphorus supply. Agron J 72 (4):685-688.
- Argaye S (2021) Development and the Role of Tissue Culture in Plant Breeding: A review. International Journal of Research Studies in Agricultural Sciences.
- Ashworth J, Mrazek K (1995) “Modified Kelowna” test for available phosphorus and potassium in soil. Commun Soil Sci Plant Anal 26 (5-6):731-739. [CrossRef]
- Atkinson JA, Pound MP, Bennett MJ, Wells DM (2019) Uncovering the hidden half of plants using new advances in root phenotyping. Curr Opin Biotech 55:1-8. [CrossRef]
- Atkinson JA, Wingen LU, Griffiths M, Pound MP, Gaju O, Foulkes MJ, Le Gouis J, Griffiths S, Bennett MJ, King J, Wells DM (2015) Phenotyping pipeline reveals major seedling root growth QTL in hexaploid wheat. J Exp Bot 66 (8):2283-2292. [CrossRef]
- Bagshaw R, Vaidyanathan LV, Nye PH (1972) The supply of nutrient ions by diffusion to plant roots in soil - V. Direct determination of labile phosphate concentration gradients in a sandy soil induced by plant uptake. Plant Soil 37 (3):617-626.
- Barber S (1995) Soil Nutrient Availability. A Mechanistic Approach. 2nd edn. Wiley, New York.
- Barber SA (1977) Application of phosphate fertilizers: Methods, rates and time of application in relation to the phosphorus status of soils. Phosphorus Agric 70:109-115.
- Barber SA (1980) Soil-plant interactions in the phosphorus nutrition of plants. In: Khasawneh FE, Sample EC, Kamprath EJ (eds) The Role of Phosphorus in Agriculture. ASA, CSSA, and SSSA Books. ASA, CSSA, and SSSA Madison, WI, pp 591-615. [CrossRef]
- Barber SA, Walker JM, Vasey EH (1963) Mechanisms for the movement of plant nutrients from the soil and fertilizer to the plant root. J Agric Food Chem 11 (3):204-207.
- Bayuelo-Jiménez JS, Ochoa-Cadavid I (2014) Phosphorus acquisition and internal utilization efficiency among maize landraces from the central Mexican highlands. Field Crop Res 156:123-134. [CrossRef]
- Bernardino KC, Pastina MM, Menezes CB, de Sousa SM, Maciel LS, Jr GC, Guimarães CT, Barros BA, da Costa e Silva L, Carneiro PCS, Schaffert RE, Kochian LV, Magalhaes JV (2019) The genetic architecture of phosphorus efficiency in sorghum involves pleiotropic QTL for root morphology and grain yield under low phosphorus availability in the soil. BMC Plant Biol 19 (1):87. [CrossRef]
- Bertrand I, McLaughlin MJ, Holloway RE, Armstrong RD, McBeath T (2006) Changes in P bioavailability induced by the application of liquid and powder sources of P, N and Zn fertilizers in alkaline soils. Nutr Cycling Agroecosyst 74 (1):27-40.
- Bolan NS (1991) Critical review on the role of mycorrhizal fungi in the uptake of phosphorus by plants. Plant Soil 141:1-11.
- Borkert C, Barber S (1983) Effect of supplying P to a portion of the soybean root system on root growth and P uptake kinetics. J Plant Nutr 6 (10):895-910.
- Bovill W, Huang C-Y, McDonald G (2013) Genetic approaches to enhancing phosphorus-use efficiency (PUE) in crops: Challenges and directions. Crop Pasture Sci 64:179. [CrossRef]
- Brenchley WE (1929) The phosphate requirement of barley at different periods of growth. Ann Bot 43:89-112.
- Carkner MK, Gao X, Entz MH (2023) Ideotype breeding for crop adaptation to low phosphorus availability on extensive organic farms. Front Plant Sci 14:1225174. [CrossRef]
- Chang MX, Gu M, Xia YW, Dai XL, Dai CR, Zhang J, Wang SC, Qu HY, Yamaji N, Feng Ma J, Xu GH (2019) OsPHT1;3 mediates uptake, translocation, and remobilization of phosphate under extremely low phosphate regimes. Plant Physiol 179 (2):656-670. [CrossRef]
- Chen A, Liu T, Wang Z, Chen X (2022) Plant root suberin: A layer of defence against biotic and abiotic stresses. Front Plant Sci 13:1056008. [CrossRef]
- Chmielewska B, Janiak A, Karcz J, Guzy-Wrobelska J, Forster BP, Nawrot M, Rusek A, Smyda P, Kedziorski P, Maluszynski M, Szarejko I (2014) Morphological, genetic and molecular characteristics of barley root hair mutants. J Appl Genet 55 (4):433-447. [CrossRef]
- Close DC, Beadle CL (2003) The ecophysiology of foliar anthocyanin. The Botanical Review 69 (2):149-161.
- Colasuonno P, Marcotuli I, Gadaleta A, Soriano JM (2021) From genetic maps to QTL cloning: an overview for durum wheat. Plants (Basel) 10 (2). [CrossRef]
- Dakora FD, Phillips DA (2002) Root exudates as mediators of mineral acquisition in low-nutrient environments. Plant Soil 245 (1):35-47. [CrossRef]
- Davies JP, Kumar S, Sastry-Dent L (2017) Chapter Three - Use of Zinc-Finger Nucleases for Crop Improvement. In: Weeks DP, Yang B (eds) Progress in Molecular Biology and Translational Science, vol 149. Academic Press, pp 47-63. [CrossRef]
- De Vita P, Avio L, Sbrana C, Laidò G, Marone D, Mastrangelo AM, Cattivelli L, Giovannetti M (2018) Genetic markers associated to arbuscular mycorrhizal colonization in durum wheat. Scientific Reports 8 (1):10612. [CrossRef]
- Deja-Muylle A, Parizot B, Motte H, Beeckman T (2020) Exploiting natural variation in root system architecture via genome-wide association studies. J Exp Bot 71 (8):2379-2389. [CrossRef]
- Dinh LT, Ueda Y, Gonzalez D, Tanaka JP, Takanashi H, Wissuwa M (2023) Novel QTL for lateral root density and length improve phosphorus uptake in rice (Oryza sativa L.). Rice 16 (1):37. [CrossRef]
- Dissanayaka DMSB, Plaxton WC, Lambers H, Siebers M, Marambe B, Wasaki J (2018) Molecular mechanisms underpinning phosphorus-use efficiency in rice. Plant, Cell & Environment 41 (7):1483-1496. [CrossRef]
- Drew M, Saker L (1978) Nutrient supply and the growth of the seminal root system in barley: III. Compensatory increases in growth of lateral roots, and in rates of phosphate uptake, in response to a localized supply of phosphate. J Exp Bot 29 (2):435-451.
- Drew MC, Saker LR, Barber SA, Jenkins W (1984) Changes in the kinetics of phosphate and potassium absorption in nutrient-deficient barley roots measured by a solution-depletion technique. Planta 160 (6):490-499. [CrossRef]
- Du K, Yang Y, Li J, Wang M, Jiang J, Wu J, Fang Y, Xiang Y, Wang Y (2023) Functional analysis of Bna-miR399c-PHO2 regulatory module involved in phosphorus stress in Brassica napus. Life 13 (2):310.
- El Mazlouzi M, Morel C, Chesseron C, Robert T, Mollier A (2020a) Contribution of external and internal phosphorus sources to grain P loading in durum wheat (Triticum durum L.) grown under contrasting P levels. Frontiers in Plant Science Volume 11 - 2020. [CrossRef]
- El Mazlouzi M, Morel C, Robert T, Yan B, Mollier A (2020b) Phosphorus uptake and partitioning in two durum wheat cultivars with contrasting biomass allocation as affected by different P supply during grain filling. Plant and Soil 449 (1):179-192. [CrossRef]
- Elliott DE, Reuter DJ, Reddy GD, Abbott RJ (1997) Phosphorus nutrition of spring wheat (Triticum aestivum L.). 2. Distribution of phosphorus in glasshouse-grown wheat and the diagnosis of phosphorus deficiency by plant analysis. Aust J Agr Res 48 (6):869-881.
- Essigmann B, Güler S, Narang RA, Linke D, Benning C (1998) Phosphate availability affects the thylakoid lipid composition and the expression of SQD1, a gene required for sulfolipid biosynthesis in Arabidopsis thaliana. Proc Natl Acad Sci U S A 95 (4):1950-1955. [CrossRef]
- Fixen P, Brentrup F, Bruulsema T, Garcia F, Norton R, Zingore S (2015) Nutrient/fertilizer use efficiency: measurement, current situation and trends. In: Drechsel P, Heffer P, Magen H, Mikkelsen R, Wichelns D (eds) Managing water and fertilizer for sustainable agricultural intensification, vol 8. International Fertilizer Industry Association (IFA), International Water Management Institute (IWMI), International Plant Nutrition Institute (IPNI), and International Potash Institute (IPI). , Paris, France,.
- Fixen P, Ludwick A, Olsen S (1983) Phosphorus and potassium fertilization of irrigated alfalfa on calcareous soils: II. Soil phosphorus solubility relationships 1. Soil Sci Soc Am J 47 (1):112-117.
- Foehse D, Jungk A (1983) Influence of phosphate and nitrate supply on root hair formation of rape, spinach and tomato plants. Plant Soil 74 (3):359-368.
- Fradgley N, Evans G, Biernaskie JM, Cockram J, Marr EC, Oliver AG, Ober E, Jones H (2020) Effects of breeding history and crop management on the root architecture of wheat. Plant Soil 452 (1):587-600. [CrossRef]
- Francis B, Aravindakumar CT, Brewer PB, Simon S (2023) Plant nutrient stress adaptation: A prospect for fertilizer limited agriculture. Environ Exp Bot 213:105431. [CrossRef]
- Fujii K (2024) Plant strategy of root system architecture and exudates for acquiring soil nutrients. Ecological Research 39 (5):623-633. [CrossRef]
- Gaume A, Mächler F, De León C, Narro L, Frossard E (2001) Low-P tolerance by maize (Zea mays L.) genotypes: Significance of root growth, and organic acids and acid phosphatase root exudation. Plant Soil 228 (2):253-264. [CrossRef]
- George TS, Gregory PJ, Hocking P, Richardson AE (2008) Variation in root-associated phosphatase activities in wheat contributes to the utilization of organic P substrates in vitro, but does not explain differences in the P-nutrition of plants when grown in soils. Environ Exp Bot 64 (3):239-249. [CrossRef]
- Glass ADM, Beaton JD, Bomke A (1980) Role of P in plant nutrition. Proceedings of the Western Canada Phosphate Symposium:357-368.
- Glassop D, Smith S, Smith F (2005) Cereal phosphate transporters associated with the mycorrhizal pathway of phosphate uptake into roots. Planta 222:688-698. [CrossRef]
- Grant C, Bittman S, Montreal M, Plenchette C, Morel C (2005) Soil and fertilizer phosphorus: Effects on plant P supply and mycorrhizal development. Can J Plant Sci 85 (1):3-14.
- Grant CA, Bailey LD (1994) The effect of tillage and KCl addition on pH, conductance, NO3-N, P, K and Cl distribution in the soil profile. Can J Soil Sci 74 (3):307-314.
- Grant CA, Flaten DN, Tomasiewicz DJ, Sheppard SC (2001) The importance of early season phosphorus nutrition. Can J Plant Sci 81 (2):211-224.
- Grant CA, Lafond GP (1994) The effects of tillage systems and crop rotations on soil chemical properties of a Black Chernozemic soil. Can J Soil Sci 74 (3):301-306.
- Green DG, Ferguson WS, Warder FG (1973) Accumulation of toxic levels of phosphorus in the leaves of phosphorus-deficient barley. Can J Plant Sci 53:241-246.
- Griffin AJ, Jungers JM, Bajgain P (2025) Root phenotyping and plant breeding of crops for enhanced ecosystem services. Crop Sci 65 (1):e21315. [CrossRef]
- Gu M, Chen A, Sun S, Xu G (2016) Complex regulation of plant phosphate transporters and the gap between molecular mechanisms and practical application: what is missing? Molecular Plant 9 (3):396-416. [CrossRef]
- Gunes A, Ali I, Mehmet A, and Cakmak I (2006) Genotypic variation in phosphorus efficiency between wheat cultivars grown under greenhouse and field conditions. Soil Sci Plant Nutr 52 (4):470-478. [CrossRef]
- Guo-Liang J (2013) Molecular Markers and Marker-Assisted Breeding in Plants. In: Sven Bode A (ed) Plant Breeding from Laboratories to Fields. IntechOpen, Rijeka, p Ch. 3. [CrossRef]
- Guo Z, Zhang C, Zhao H, Liu Y, Chen X, Zhao H, Chen L, Ruan W, Chen Y, Yuan L, Yi K, Xu L, Zhang J (2025) Vacuolar phosphate efflux transporter ZmVPEs mediate phosphate homeostasis and remobilization in maize leaves. Journal of Integrative Plant Biology 67 (2):311-326. [CrossRef]
- Hamdan MF, Tan BC (2024) Genetic modification techniques in plant breeding: A comparative review of CRISPR/Cas and GM technologies. Horticultural Plant Journal. [CrossRef]
- Hamel C, Strullu D-G (2006) Arbuscular mycorrhizal fungi in field crop production: potential and new direction. Can J Plant Sci 86 (4):941-950.
- Han Y, White PJ, Cheng L (2021) Mechanisms for improving phosphorus utilization efficiency in plants. Ann Bot 129 (3):247-258. [CrossRef]
- Havlin JL, Tisdale SL, Nelson WL, Beaton JD (2014) Soil Fertility and Fertilizers: An Introduction to Nutrient Management. 8th edn. Pearson, Inc., Upper Saddle River, NJ, USA.
- Hedley M, McLaughlin M (2005) Reactions of phosphate fertilizers and by-products in soils. In: Sims JT, Sharpley AN (eds) Phosphorus: Agriculture and the Environment. vol 46. American Society of Agronomy, Crop Science Society of America, Soil Science Society of America, Madison, WI, pp 181-252.
- Hettiarachchi GM, Lombi E, McLaughlin MJ, Chittleborough D, Self P (2006) Density changes around phosphorus granules and fluid bands in a calcareous soil. Soil Sci Soc Am J 70 (3):960-966.
- Heuer S, Gaxiola R, Schilling R, Herrera-Estrella L, López-Arredondo D, Wissuwa M, Delhaize E, Rouached H (2017) Improving phosphorus use efficiency: a complex trait with emerging opportunities. The Plant Journal 90 (5):868-885. [CrossRef]
- Hidaka A, Kitayama K (2013) Relationship between photosynthetic phosphorus-use efficiency and foliar phosphorus fractions in tropical tree species. Ecology and Evolution 3 (15):4872-4880. [CrossRef]
- Hinsinger P (1998) How do plant roots acquire mineral nutrients? Chemical processes involved in the rhizosphere. Adv Agron 64:225-265.
- Hinsinger P (2001) Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: A review. Plant Soil 237 (2):173-195.
- Hinsinger P, Gilkes R (1995) Root-induced dissolution of phosphate rock in the rhizosphere of lupins grown in alkaline soil. Soil Res 33 (3):477-489. [CrossRef]
- Hodge A (2004) The plastic plant: root responses to heterogeneous supplies of nutrients. New Phytol 162 (1):9-24. [CrossRef]
- Hoffland E, Findenegg GR, Nelemans JA (1989) Solubilization of rock phosphate by rape - II. Local root exudation of organic acids as a response to P-starvation. Plant Soil 113 (2):161-165.
- Holloway RE, Bertrand I, Frischke AJ, Brace DM, McLaughlin MJ, Shepperd W (2001) Improving fertiliser efficiency on calcareous and alkaline soils with fluid sources of P, N and Zn. Plant Soil 236 (2):209-219.
- Hopkins BG (2015) Phosphorus. In: Barker AV, Pilbeam DJ (eds) Handbook of Plant Nutrition. second edn. CRC press, Boca Ratan, FL, pp 65 -126.
- Hoppo SD, Elliott DE, Reuter DJ (1999) Plant tests for diagnosing phosphorus deficiency in barley (Hordeum vulgare L.). Aust J Exp Agr 39 (7):857-872.
- IPNI (2016 ) 4R Plant Nutrition Manual: A Manual for Improving the Management of Plant Nutrition. In: Bruulsema TW, Fixen PE, Sulewski GD (eds). International Plant Nutrition Institute, Peachtree Corners, GA, USA,.
- Jakobsen I (1986) Vesicular-arbuscular mycorrhiza in field-grown crops. III. Mycorrhizal infection and rates of phosphorus inflow in pea plants. New Phytol 104:573-581.
- Jeong K, Mattes N, Catausan S, Chin JH, Paszkowski U, Heuer S (2015) Genetic diversity for mycorrhizal symbiosis and phosphate transporters in rice. Journal of Integrative Plant Biology 57 (11):969-979. [CrossRef]
- Jha UC, Nayyar H, Parida SK, Beena R, Pang J, Siddique KHM (2023) Breeding and genomics approaches for improving phosphorus-use efficiency in grain legumes. Environ Exp Bot 205:105120. [CrossRef]
- Johnston AE, Poulton PR, Fixen PE, Curtin D (2014) Phosphorus: its efficient use in agriculture. In: Advances in Agronomy, vol 123. Elsevier, pp 177-228.
- Joung JK, Sander JD (2013) TALENs: a widely applicable technology for targeted genome editing. Nat Rev Mol Cell Biol 14 (1):49-55. [CrossRef]
- Jungk A, Barber SA (1974) Phosphate uptake rate of corn roots as related to the proportion of the roots exposed to phosphate. Agron J 66:554-557.
- Jungk A, Seeling B, Gerke J (1993) Mobilization of different phosphate fractions in the rhizosphere. Plant Soil 155-156 (1):91-94.
- Kaeppler SM, Parke JL, Mueller SM, Senior L, Stuber C, Tracy WF (2000) Variation among maize inbred lines and detection of quantitative trait loci for growth at low phosphorus and responsiveness to arbuscular mycorrhizal fungi. Crop Sci 40 (2):358-364. [CrossRef]
- Kar G, Peak D, Schoenau JJ (2012) Spatial distribution and chemical speciation of soil phosphorus in a band application. Soil Sci Soc Am J 76 (6):2297-2306.
- Kirk A, Entz M, Fox S, Tenuta M (2011) Mycorrhizal colonization, P uptake and yield of older and modern wheats under organic management. Can J Plant Sci 91 (4):663-667. [CrossRef]
- Konesky D, Siddiqi M, Glass A, Hsiao A (1989) Wild oat and barley interactions: varietal differences in competitiveness in relation to phosphorus supply. Can J Bot 67 (11):3366-3371.
- Kucey R, Janzen H, Leggett M (1989) Microbially mediated increases in plant-available phosphorus. In: Advances in agronomy, vol 42. Elsevier, pp 199-228.
- Kumpf RP, Nowack MK (2015) The root cap: a short story of life and death. J Exp Bot 66 (19):5651-5662. [CrossRef]
- Lambers H (2022) Phosphorus acquisition and utilization in plants. Annu Rev Plant Biol 73 (Volume 73, 2022):17-42. [CrossRef]
- Lambers H, Cawthray GR, Giavalisco P, Kuo J, Laliberté E, Pearse SJ, Scheible W-R, Stitt M, Teste F, Teste F, Turner BL (2012) Proteaceae from severely phosphorus-impoverished soils extensively replace phospholipids with galactolipids and sulfolipids during leaf development to achieve a high photosynthetic phosphorus-use-efficiency. The New phytologist 196 (4):1098-1108. [CrossRef]
- Lambers H, Cramer MD, Shane MW, Wouterlood M, Poot P, Veneklaas EJ (2003) Structure and functioning of cluster roots and plant responses to phosphate deficiency. Plant Soil 248 (1-2):ix-xix.
- Lee HY, Chen Z, Zhang C, Yoon GM (2019) Editing of the OsACS locus alters phosphate deficiency-induced adaptive responses in rice seedlings. J Exp Bot 70 (6):1927-1940. [CrossRef]
- Lefebvre DD, Glass ADM (1982) Regulation of phosphate influx in barley roots: Effects of phosphate deprivation and reduction of influx with provision of orthophosphate. Physiol Plant 54:199-206.
- Li X, Chen Y, Xu Y, Sun H, Gao Y, Yan P, Song Q, Li S, Zhan A (2024) Genotypic variability in root morphology in a diverse wheat genotypes under drought and low phosphorus stress. Plants 13.
- Lim S, Borza T, Peters RD, Coffin RH, Al-Mughrabi KI, Pinto DM, Wang-Pruski G (2013) Proteomics analysis suggests broad functional changes in potato leaves triggered by phosphites and a complex indirect mode of action against Phytophthora infestans. Journal of proteomics 93:207-223.
- Lipton DS, Blanchar RW, Blevins DG (1987) Citrate, malate, and succinate concentration in exudates from P-sufficient and P-stressed Medicago sativa L. seedlings 1. Plant Physiol 85 (2):315-317. [CrossRef]
- Lisch D (2013) How important are transposons for plant evolution? Nature Reviews Genetics 14 (1):49-61.
- Liu T-Y, Huang T-K, Yang S-Y, Hong Y-T, Huang S-M, Wang F-N, Chiang S-F, Tsai S-Y, Lu W-C, Chiou T-J (2016) Identification of plant vacuolar transporters mediating phosphate storage. Nature Communications 7 (1):11095. [CrossRef]
- Lombi E, McLaughlin MJ, Johnston C, Armstrong RD, Holloway RE (2004) Mobility and lability of phosphorus from granular and fluid monoammonium phosphate differs in a calcareous soil. Soil Sci Soc Am J 68 (2):682-689.
- Lombi E, McLaughlin MJ, Johnston C, Armstrong RD, Holloway RE (2005) Mobility, solubility and lability of fluid and granular forms of P fertiliser in calcareous and non-calcareous soils under laboratory conditions. Plant Soil 269 (1-2):25-34.
- Lombi E, Scheckel KG, Armstrong RD, Forrester S, Cutler JN, Paterson D (2006) Speciation and Distribution of Phosphorus in a Fertilized Soil. Soil Sci Soc Am J 70 (6):2038-2048. [CrossRef]
- Loneragan JF, Asher CJ (1967) Response of plants to phosphate concentration in solution culture: II. Rate of phosphate absorption and its relation to growth. Soil Sci 103 (5):311-318.
- López-Bucio J, de la Vega OM, Guevara-García A, Herrera-Estrella L (2000) Enhanced phosphorus uptake in transgenic tobacco plants that overproduce citrate. Nat Biotechnol 18 (4):450-453. [CrossRef]
- Lopez G, Ahmadi SH, Amelung W, Athmann M, Ewert F, Gaiser T, Gocke MI, Kautz T, Postma J, Rachmilevitch S, Schaaf G, Schnepf A, Stoschus A, Watt M, Yu P, Seidel SJ (2022) Nutrient deficiency effects on root architecture and root-to-shoot ratio in arable crops. Front Plant Sci 13:1067498. [CrossRef]
- Lynch JP (2007a) Rhizoeconomics: the roots of shoot growth limitations. HortScience 42 (5):1107-1109.
- Lynch JP (2007b) Roots of the second green revolution. Australian Journal of Botany 55 (5):493-512.
- Lynch JP (2013) Steep, cheap and deep: an ideotype to optimize water and N acquisition by maize root systems. Ann Bot 112 (2):347-357.
- Lynch JP (2022) Harnessing root architecture to address global challenges. The Plant Journal 109 (2):415-431. [CrossRef]
- Lynch JP, Brown KM (2001) Topsoil foraging–an architectural adaptation of plants to low phosphorus availability. Plant Soil 237:225-237.
- Mabagala FS, Mng'ong'o ME (2022) On the tropical soils; The influence of organic matter (OM) on phosphate bioavailability. Saudi Journal of Biological Sciences 29 (5):3635-3641. [CrossRef]
- Magalhaes JV, de Sousa SM, Guimaraes CT, Kochian LV (2017) Chapter 7 - The role of root morphology and architecture in phosphorus acquisition: physiological, genetic, and molecular basis. In: Hossain MA, Kamiya T, Burritt DJ, Tran L-SP, Fujiwara T (eds) Plant Macronutrient Use Efficiency. Academic Press, pp 123-147. [CrossRef]
- Maharajan T, Ceasar SA, Ajeesh krishna TP, Ramakrishnan M, Duraipandiyan V, Naif Abdulla AD, Ignacimuthu S (2018) Utilization of molecular markers for improving the phosphorus efficiency in crop plants. Plant Breed 137 (1):10-26. [CrossRef]
- Malhi SS, Johnston AM, Schoenau JJ, Wang ZH, Vera CL (2006) Seasonal biomass accumulation and nutrient uptake of wheat, barley and oat on a Black Chernozem soil in Saskatchewan. Can J Plant Sci 86 (4):1005-1014.
- Malhi SS, Johnston AM, Schoenau JJ, Wang ZH, Vera CL (2007a) Seasonal biomass accumulation and nutrient uptake of canola, mustard, and flax on a Black Chernozem soil in Saskatchewan. J Plant Nutr 30 (4):641-658.
- Malhi SS, Johnston AM, Schoenau JJ, Wang ZH, Vera CL (2007b) Seasonal biomass accumulation and nutrient uptake of pea and lentil on a Black Chernozem soil in Saskatchewan. J Plant Nutr 30 (5):721-737.
- Malhotra H, Vandana, Sharma S, Pandey R (2018) Phosphorus Nutrition: Plant Growth in Response to Deficiency and Excess. Plant Nutrients and Abiotic Stress Tolerance. Springer, Singapore. [CrossRef]
- Manna M, Achary VMM, Islam T, Agrawal PK, Reddy MK (2016) The development of a phosphite-mediated fertilization and weed control system for rice. Scientific Reports 6 (1):24941. [CrossRef]
- Manske GGB, Ortiz-Monasterio JI, Van Ginkel M, González RM, Rajaram S, Molina E, Vlek PLG (2000) Traits associated with improved P-uptake efficiency in CIMMYT's semidwarf spring bread wheat grown on an acid Andisol in Mexico. Plant Soil 221 (2):189-204. [CrossRef]
- Marschner H, Kirkby EA, Cakmak I (1996) Effect of mineral nutritional status on shoot-root partitioning of photoassimilates and cycling of mineral nutrients. J Exp Bot 47 (SPEC. ISS.):1255-1263.
- McBeath TM, Armstrong RD, Lombi E, McLaughlin MJ, Holloway RE (2005) Responsiveness of wheat (Triticum aestivum) to liquid and granular phosphorus fertilisers in southern Australian soils. Aust J Soil Res 43 (2):203-212.
- McGonigle TP, Hutton M, Greenley A, Karamanos R (2011) Role of mycorrhiza in a wheat–flax versus canola–flax rotation: A case study. Commun Soil Sci Plant Anal 42 (17):2134-2142. [CrossRef]
- McGrail RK, Van Sanford DA, McNear DH (2023) Breeding milestones correspond with changes to wheat rhizosphere biogeochemistry that affect P acquisition. Agronomy 13 (3):813.
- McLaughlin MJ, McBeath TM, Smernik R, Stacey SP, Ajiboye B, Guppy C (2011) The chemical nature of P accumulation in agricultural soils—implications for fertiliser management and design: an Australian perspective. Plant Soil 349 (1-2):69-87.
- Mikwa EO, Wittkop B, Windpassinger SM, Weber SE, Ehrhardt D, Snowdon RJ (2024) Early exposure to phosphorus starvation induces genetically determined responses in Sorghum bicolor roots. Theor Appl Genet 137 (10). [CrossRef]
- Miller MH (2000) Arbuscular mycorrhizae and the phosphorus nutrition of maize: A review of Guelph studies. Can J Plant Sci 80 (1):47-52.
- Mills HA, Jones JB, Jr. (1996) Plant Analysis Handbook II. MicroMacro Publishing, Inc., Jefferson City, MO.
- Mohamed GES, Marshall C (1979) The pattern of distribution of phosphorus and dry matter with time in spring wheat. Ann Bot 44 (6):721-730.
- Monreal MA, Grant CA, Irvine RB, Mohr RM, McLaren DL, Khakbazan M (2011) Crop management effect on arbuscular mycorrhizae and root growth of flax. Can J Plant Sci 91 (2):315-324. [CrossRef]
- Morel C, Plenchette C (1994) Is the isotopically exchangeable phosphate of a loamy soil the plant-available P? Plant Soil 158 (2):287-297.
- Morel C, Tunney H, Plenet D, Pellerin S (2000) Transfer of phosphate ions between soil and solution: Perspectives in soil testing. J Environ Qual 29 (1):50-59.
- Moussa AA, Mandozai A, Jin Y, Qu J, Zhang Q, Zhao H, Anwari G, Khalifa MAS, Lamboro A, Noman M, Bakasso Y, Zhang M, Guan S, Wang P (2021) Genome-wide association screening and verification of potential genes associated with root architectural traits in maize (Zea mays L.) at multiple seedling stages. BMC Genomics 22 (1):558. [CrossRef]
- Müller J, Toev T, Heisters M, Teller J, Moore Katie L, Hause G, Dinesh Dhurvas C, Bürstenbinder K, Abel S (2015) Iron-dependent callose deposition adjusts root meristem maintenance to phosphate availability. Developmental Cell 33 (2):216-230. [CrossRef]
- Murovec J, Bohanec B (2012) Haploids and Doubled Haploids in Plant Breeding. In: Abdurakhmonov IY (ed) Plant Breeding. IntecOpen, pp 87-106.
- Nadeem M, Mollier A, Morel C, Vives A, Prud’homme L, Pellerin S (2011) Relative contribution of seed phosphorus reserves and exogenous phosphorus uptake to maize (Zea mays L.) nutrition during early growth stages. Plant Soil 346 (1):231-244. [CrossRef]
- Nahampun HN, López-Arredondo D, Xu X, Herrera-Estrella L, Wang K (2016) Assessment of ptxD gene as an alternative selectable marker for Agrobacterium-mediated maize transformation. Plant Cell Rep 35 (5):1121-1132. [CrossRef]
- Navea IP, Yang S, Tolangi P, Sumabat RM, Zhang W, Chin JH (2024) Enhancement of rice traits for the maintenance of the phosphorus balance between rice plants and the soil. Current Plant Biology 38:100332. [CrossRef]
- Ojeda-Rivera JO, Alejo-Jacuinde G, Nájera-González H-R, López-Arredondo D (2022) Prospects of genetics and breeding for low-phosphate tolerance: an integrated approach from soil to cell. Theor Appl Genet 135 (11):4125-4150. [CrossRef]
- Oo AZ, Tsujimoto Y, Mukai M, Nishigaki T, Takai T, Uga Y (2024) Significant interaction between root system architecture and stratified phosphorus availability for the initial growth of rice in a flooded soil culture. Rhizosphere 31:100947. [CrossRef]
- Ouma E, Dickson L, Matonyei T, Were B, Joyce A, Too E, Augustino O, Gudu S, Peter K, John O, Caleb O (2012) Development of maize single cross hybrids for tolerance to low phosphorus. African Journal of plant science 6:394-402. [CrossRef]
- Ozanne PG (1980) Phosphate nutrition of plants - A general treatise. The Role of Phosphorus in Agriculture:559-589.
- Pan X-w, Li W-b, Zhang Q-y, Li Y-h, Liu M-s (2008) Assessment on phosphorus efficiency characteristics of soybean genotypes in phosphorus-deficient soils. Agr Sci China 7 (8):958-969. [CrossRef]
- Pan Y, Song Y, Zhao L, Chen P, Bu C, Liu P, Zhang D (2022) The genetic basis of phosphorus utilization efficiency in plants provide new insight into woody perennial plants improvement. Int J Mol Sci 23 (4). [CrossRef]
- Pandeya D, López-Arredondo DL, Janga MR, Campbell LM, Estrella-Hernández P, Bagavathiannan MV, Herrera-Estrella L, Rathore KS (2018) Selective fertilization with phosphite allows unhindered growth of cotton plants expressing the ptxD gene while suppressing weeds. Proceedings of the National Academy of Sciences 115 (29):E6946-E6955. doi:doi:10.1073/pnas.1804862115.
- Penn CJ, Camberato JJ (2019) A critical review on soil chemical rocesses that control how soil pH affects phosphorus availability to plants. Agriculture 9 (6):120.
- Pierre W, Parker F (1927) Soil phosphorus studies: II. The concentration of organic and inorganic phosphorus in the soil solution and soil extracts and the availibility of the organic phosphorus to plants. Soil Sci 24 (2):119-128.
- Pierzynski GM, McDowell RW (2005) Chemistry, cycling, and potential movement of inorganic phosphorus in soils. Phosphorus: agriculture and the environment (phosphorusagric):53-86.
- Plaxton W, Lambers H (2015) Phosphorus: back to the roots. Annual plant reviews 48:3-15.
- Poirier Y, Jaskolowski A, Clúa J (2022) Phosphate acquisition and metabolism in plants. Current Biology 32 (12):R623-R629. [CrossRef]
- Raboy V (2002) Progress in breeding low phytate crops. J Nutr 132 (3):503S-505S. [CrossRef]
- Rajamanickam V, Sevanthi AM, Swarbreck SM, Gudi S, Singh N, Singh VK, Wright TIC, Bentley AR, Muthamilarasan M, Das A, Chinnusamy V, Pandey R (2024) High-throughput root phenotyping and association analysis identified potential genomic regions for phosphorus use efficiency in wheat (Triticum aestivum L.). Planta 260 (6):142. [CrossRef]
- Raven PH, Evert RF, Eichhorn SE (2005) Biology of Plants. Seventh edn. W. H. Freeman and Company, New York.
- Ribeiro CAG, de Sousa Tinoco SM, de Souza VF, Negri BF, Gault CM, Pastina MM, Magalhaes JV, Guimarães LJM, de Barros EG, Buckler ES, Guimaraes CT (2023) Genome-wide association Sstudy for root morphology and phosphorus acquisition efficiency in diverse maize panels. International Journal of Molecular Sciences 24 (7):6233.
- Richardson AE (2001) Prospects for using soil microorganisms to improve the acquisition of phosphorus by plants. Funct Plant Biol 28 (9):897-906. [CrossRef]
- Richardson AE, Hocking PJ, Simpson RJ, George TS (2009) Plant mechanisms to optimise access to soil phosphorus. Crop Pasture Sci 60 (2):124-143.
- Richardson AE, Lynch JP, Ryan PR, Delhaize E, Smith FA, Smith SE, Harvey PR, Ryan MH, Veneklaas EJ, Lambers H, Oberson A, Culvenor RA, Simpson RJ (2011) Plant and microbial strategies to improve the phosphorus efficiency of agriculture. Plant Soil 349 (1):121-156. [CrossRef]
- Roberts TL, Johnston AE (2015) Phosphorus use efficiency and management in agriculture. Resources, Conservation and Recycling 105:275-281. [CrossRef]
- Rose T, Liu L, Wissuwa M (2013) Improving phosphorus efficiency in cereal crops: Is breeding for reduced grain phosphorus concentration part of the solution? Frontiers in Plant Science 4. [CrossRef]
- Rossnagel BG, Zatorski T, Arganosa G, Beattie AD (2008) Registration of ‘CDC Lophy-I’ Barley. Journal of Plant Registrations 2 (3):169-173. [CrossRef]
- Rouached H, Arpat AB, Poirier Y (2010) Regulation of phosphate starvation responses in plants: signaling players and cross-talks. Molecular Plant 3 (2):288-299.
- Ryan MH, Graham JH (2002) Is there a role for arbuscular mycorrhizal fungi in production agriculture? Plant Soil 244 (1-2):263-271.
- Ryan MH, Kirkegaard JA (2012) The agronomic relevance of arbuscular mycorrhizas in the fertility of Australian extensive cropping systems. Agr Ecosyst Environ 163:37-53. [CrossRef]
- Ryan MH, Small DR, Ash JE (2000) Phosphorus controls the level of colonisation by arbuscular mycorrhizal fungi in conventional and biodynamic irrigated dairy pastures. Aust J Exp Agr 40 (5):663-670.
- Salas-González I, Reyt G, Flis P, Custódio V, Gopaulchan D, Bakhoum N, Dew TP, Suresh K, Franke RB, Dangl JL (2021) Coordination between microbiota and root endodermis supports plant mineral nutrient homeostasis. Science 371 (6525):eabd0695.
- Sample EC, Soper RJ, Racz GJ (1980) Reaction of phosphate fertilizers in soils. In: Khasawneh FE, Sample EC, Kamprath EJ (eds) The Role of Phosphorus in Agriculture. ASA, CSSA, and SSSA, Madision, , WI, pp 262-310. [CrossRef]
- Schachtman DP, Reid RJ, Ayling SM (1998) Phosphorus uptake by plants: from soil to cell. Plant Physiol 116 (2):447-453. [CrossRef]
- Schjørring JK, Jensén P (1984) Phosphorus nutrition of barley, buckwheat and rape seedlings. I. Influence of seed-borne P and external P levels on growth, P content and P/P-fractionation in shoots and roots32 31. Physiol Plant 61:577-583.
- Schneider A, Morel C (2000) Relationship between the isotopically exchangeable and resin-extractable phosphate of deficient to heavily fertilized soil. Eur J Soil Sci 51 (4):709-715.
- Schroeder JI, Delhaize E, Frommer WB, Guerinot ML, Harrison MJ, Herrera-Estrella L, Horie T, Kochian LV, Munns R, Nishizawa NK (2013) Using membrane transporters to improve crops for sustainable food production. Nature 497 (7447):60.
- Sharma S, Pinson SRM, Gealy DR, Edwards JD (2021) Genomic prediction and QTL mapping of root system architecture and above-ground agronomic traits in rice (Oryza sativa L.) with a multitrait index and Bayesian networks. G3 Genes|Genomes|Genetics 11 (10). [CrossRef]
- Shen J, Yuan L, Zhang J, Li H, Bai Z, Chen X, Zhang W, Zhang F (2011) Phosphorus dynamics: from soil to plant. Plant Physiol 156 (3):997-1005. [CrossRef]
- Shen Y, Zhou G, Liang C, Tian Z (2022) Omics-based interdisciplinarity is accelerating plant breeding. Current Opinion in Plant Biology 66:102167. [CrossRef]
- Slatkin M (2008) Linkage disequilibrium--understanding the evolutionary past and mapping the medical future. Nat Rev Genet 9 (6):477-485. [CrossRef]
- Smith SE, Jakobsen I, Grønlund M, Smith FA (2011) Roles of arbuscular mychorrhizas in plant phosphorus nutrition: Interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition. Plant Physiol 156 (3):1050-1057. [CrossRef]
- Sparvoli F, Cominelli E (2015) Seed biofortification and phytic acid reduction: a conflict of interest for the plant? Plants (Basel) 4 (4):728-755. [CrossRef]
- Stetter MG, Schmid K, Ludewig U (2015) Uncovering genes and ploidy involved in the high diversity in root hair density, length and response to local scarce phosphate in Arabidopsis thaliana. PLoS One 10 (3):e0120604. [CrossRef]
- Strong WM, Soper RJ (1974a) Phosphorus utilization by flax, wheat, rape, and buckwheat from a band or pellet-like application. I. Reaction zone proliferation. Agron J 66:597-601.
- Strong WM, Soper RJ (1974b) Phosphorus utilization by flax, wheat, rape, and buckwheat from a band or pellet-like application. II. Influence of reaction zone phosphorus concentration and soil phosphorus supply. Agron J 66:601-605.
- Sultenfuss J, Doyle W (1999) Functions of phosphorus in plants. Better Crops 83 (1):6-7.
- Sutton PJ, Peterson GA, Sander DH (1983) Dry matter production in tops and roots of winter wheat as affected by phosphorus availability during various growth stages. Agron J 75:657-663.
- Syers J, Johnston A, Curtin D (2008) Efficiency of soil and fertilizer phosphorus use., FAO Fertilizer and Plant Nutrition Bulletin No. 18.(FAO: Rome).
- Tao J, Bauer DE, Chiarle R (2023) Assessing and advancing the safety of CRISPR-Cas tools: from DNA to RNA editing. Nature Communications 14 (1):212. [CrossRef]
- Theodorou ME, Plaxton WC (1993) Metabolic adaptations of plant respiration to nutritional phosphate deprivation. Plant Physiol 101 (2):339-344. [CrossRef]
- Tomasiewicz DJ (2000) Advancing the understanding and interpretation of plant and soil tests for phosphorus in Manitoba. University of Manitoba, Winnipeg, MB.
- van de Wiel CC, van der Linden CG, Scholten OE (2016) Improving phosphorus use efficiency in agriculture: opportunities for breeding. Euphytica 207:1-22.
- Vance CP, Uhde-Stone C, Allan DL (2003) Phosphorus acquisition and use: critical adaptations by plants for securing a nonrenewable resource. New Phytol 157 (3):423-447. [CrossRef]
- Veneklaas EJ, Lambers H, Bragg J, Finnegan PM, Lovelock CE, Plaxton WC, Price CA, Scheible W-R, Shane MW, White PJ, Raven JA (2012) Opportunities for improving phosphorus-use efficiency in crop plants. New Phytol 195 (2):306-320. [CrossRef]
- Veršulienė A, Hirte J, Ciulla F, Camenzind M, Don A, Durand-Maniclas F, Heinemann H, Herrera JM, Hund A, Seidel F, da Silva-Lopes M, Toleikienė M, Visse-Mansiaux M, Yu K, Bender SF (2024) Wheat varieties show consistent differences in root colonization by mycorrhiza across a European pedoclimatic gradient. Eur J Soil Sci 75 (4):e13543. [CrossRef]
- Vlčko T, Ohnoutková L (2020) Allelic variants of CRISPR/Cas9 induced mutation in an inositol trisphosphate 5/6 kinase gene manifest different phenotypes in barley. Plants 9 (2):195.
- Wachsman G, Sparks EE, Benfey PN (2015) Genes and networks regulating root anatomy and architecture. New Phytol 208 (1):26-38. [CrossRef]
- Wang F, Deng M, Xu J, Zhu X, Mao C (2018) Molecular mechanisms of phosphate transport and signaling in higher plants. Seminars in Cell & Developmental Biology 74:114-122. [CrossRef]
- Wang L, Liu D (2018) Functions and regulation of phosphate starvation-induced secreted acid phosphatases in higher plants. Plant Sci 271:108-116. [CrossRef]
- Wang X, Cheng L, Xiong C, Whalley WR, Miller AJ, Rengel Z, Zhang F, Shen J (2024) Understanding plant–soil interactions underpins enhanced sustainability of crop production. Trends Plant Sci 29 (11):1181-1190. [CrossRef]
- Wang X, Whalley WR, Miller AJ, White PJ, Zhang F, Shen J (2020) Sustainable cropping requires adaptation to a heterogeneous rhizosphere. Trends Plant Sci 25 (12):1194-1202. [CrossRef]
- Wang Y, Krogstad T, Clarke JL, Hallama M, Øgaard AF, Eich-Greatorex S, Kandeler E, Clarke N (2016) Rhizosphere organic anions play a minor role in improving crop species' ability to take up residual phosphorus (P) in agricultural soils low in P availability. Front Plant Sci 7:1664. [CrossRef]
- Wang Y, Lambers H (2020) Root-released organic anions in response to low phosphorus availability: recent progress, challenges and future perspectives. Plant Soil 447 (1):135-156. [CrossRef]
- Wang Y, Wang F, Lu H, Liu Y, Mao C (2021) Phosphate uptake and transport in plants: An elaborate regulatory system. Plant Cell Physiol 62 (4):564-572. [CrossRef]
- Watson A, Ghosh S, Williams MJ, Cuddy WS, Simmonds J, Rey MD, Asyraf Md Hatta M, Hinchliffe A, Steed A, Reynolds D, Adamski NM, Breakspear A, Korolev A, Rayner T, Dixon LE, Riaz A, Martin W, Ryan M, Edwards D, Batley J, Raman H, Carter J, Rogers C, Domoney C, Moore G, Harwood W, Nicholson P, Dieters MJ, DeLacy IH, Zhou J, Uauy C, Boden SA, Park RF, Wulff BBH, Hickey LT (2018) Speed breeding is a powerful tool to accelerate crop research and breeding. Nat Plants 4 (1):23-29. [CrossRef]
- White PJ, Veneklaas EJ (2012) Nature and nurture: the importance of seed phosphorus content. Plant Soil 357 (1):1-8. [CrossRef]
- Yadav S, Ross EM, Aitken KS, Hickey LT, Powell O, Wei X, Voss-Fels KP, Hayes BJ (2021) A linkage disequilibrium-based approach to position unmapped SNPs in crop species. BMC Genomics 22 (1):773. [CrossRef]
- Yamaji N, Takemoto Y, Miyaji T, Mitani-Ueno N, Yoshida KT, Ma JF (2017) Reducing phosphorus accumulation in rice grains with an impaired transporter in the node. Nature 541 (7635):92-95. [CrossRef]
- Yan J, Wang X (2023) Machine learning bridges omics sciences and plant breeding. Trends Plant Sci 28 (2):199-210. [CrossRef]
- Yang S-Y, Grønlund M, Jakobsen I, Grotemeyer MS, Rentsch D, Miyao A, Hirochika H, Kumar CS, Sundaresan V, Salamin N, Catausan S, Mattes N, Heuer S, Paszkowski U (2012) Nonredundant regulation of rice arbuscular mycorrhizal symbiosis by two members of the PHOSPHATE TRANSPORTER1 gene family. The Plant Cell 24 (10):4236-4251. [CrossRef]
- Yang S-Y, Huang T-K, Kuo H-F, Chiou T-J (2017) Role of vacuoles in phosphorus storage and remobilization. J Exp Bot 68 (12):3045-3055.
- Yang S-Y, Lin W-Y, Hsiao Y-M, Chiou T-J (2024) Milestones in understanding transport, sensing, and signaling of the plant nutrient phosphorus. The Plant Cell 36 (5):1504-1523. [CrossRef]
- Yuan P, Liu H, Wang X, Hammond JP, Shi L (2023) Genome-wide association study reveals candidate genes controlling root system architecture under low phosphorus supply at seedling stage in Brassica napus. Mol Breed 43 (8). [CrossRef]
- Zhu J, Kaeppler SM, Lynch JP (2005) Topsoil foraging and phosphorus acquisition efficiency in maize (Zea mays). Funct Plant Biol 32 (8):749-762. [CrossRef]
Figure 1.
Influence of pH on the distribution of orthophosphate forms in solution (Havlin et al. 2014).
Figure 1.
Influence of pH on the distribution of orthophosphate forms in solution (Havlin et al. 2014).
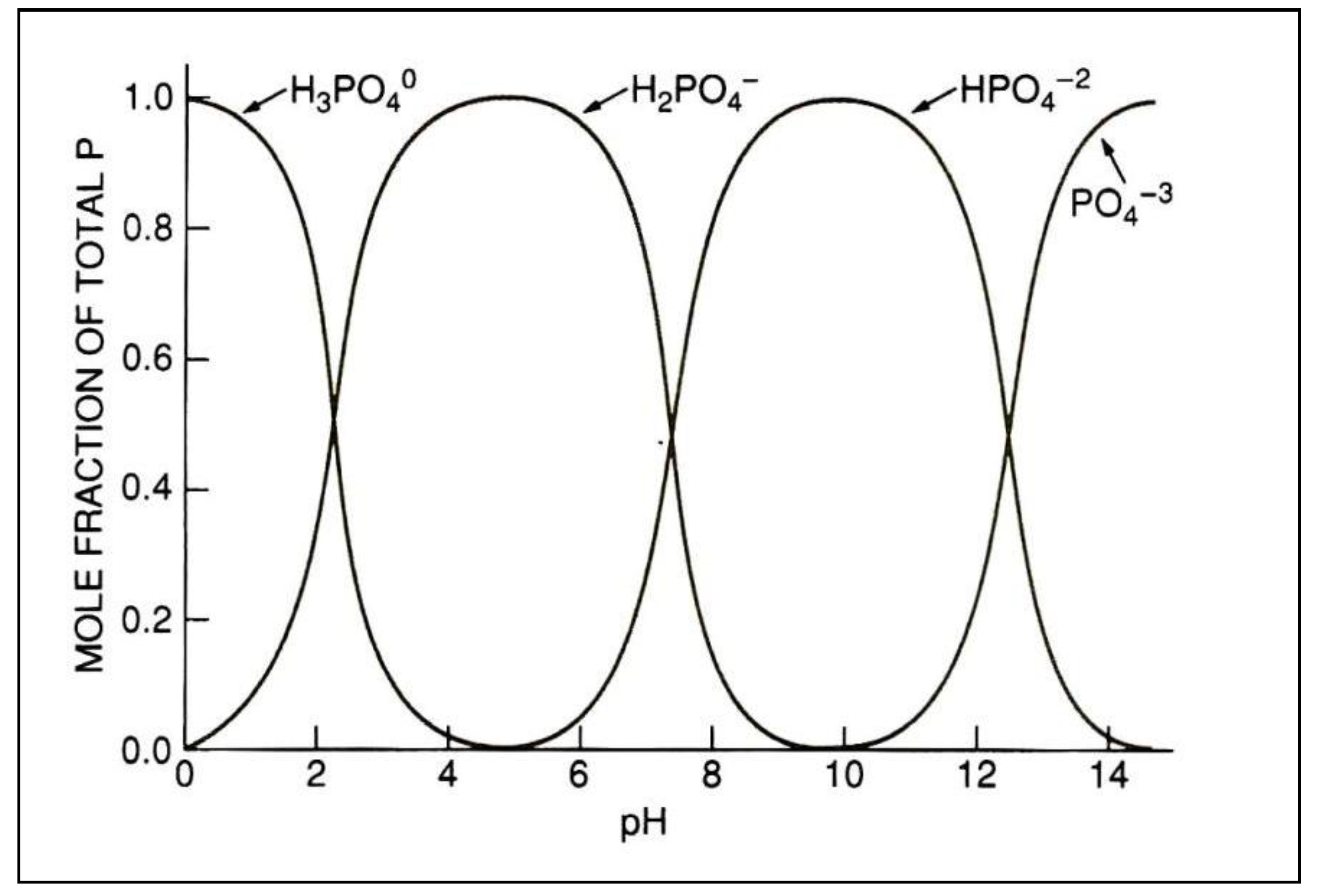
Figure 2.
Conceptual diagram for the forms of inorganic P in soils categorized in terms of accessibility, extractability and plant availability (Johnston et al. 2014).
Figure 2.
Conceptual diagram for the forms of inorganic P in soils categorized in terms of accessibility, extractability and plant availability (Johnston et al. 2014).
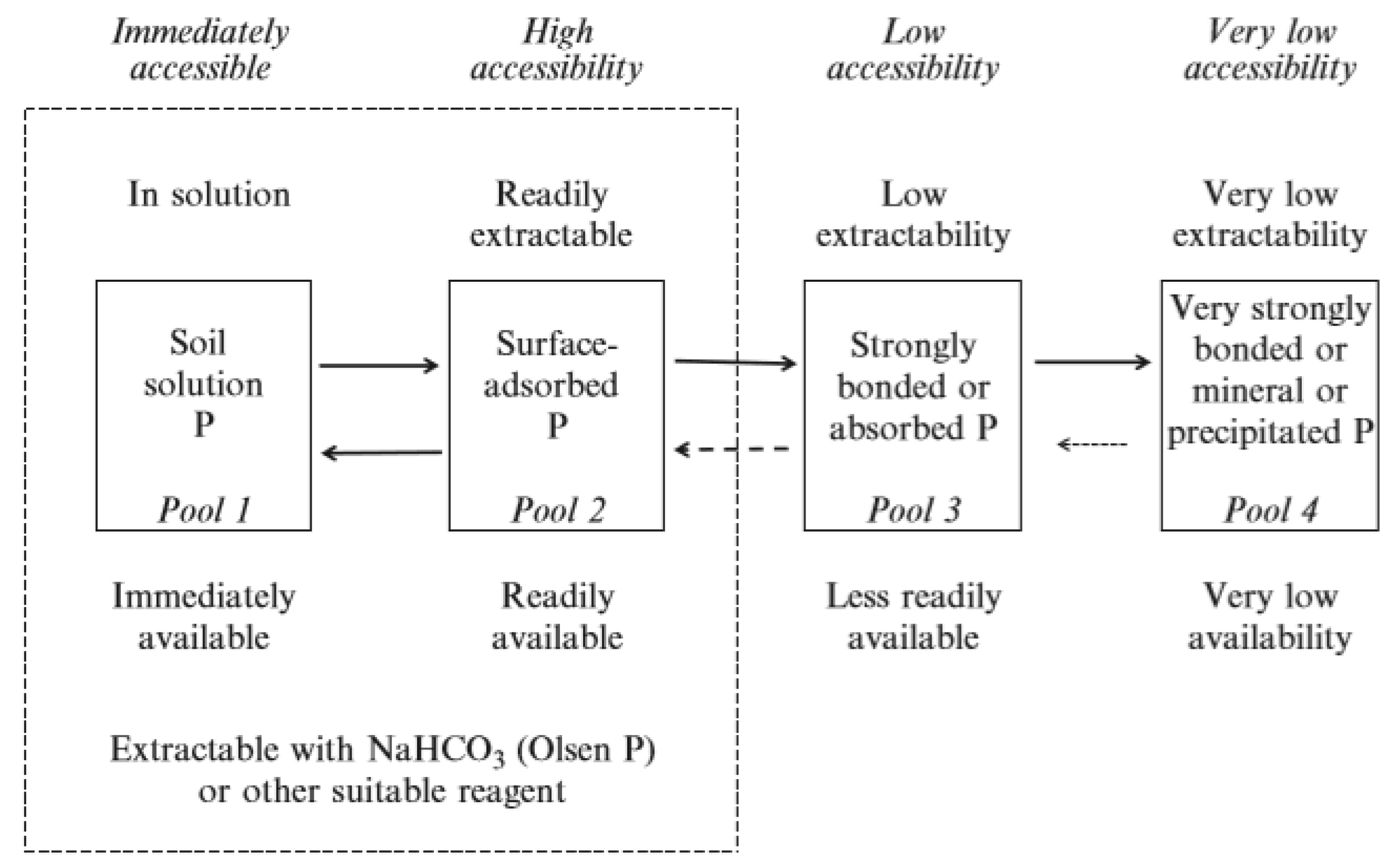
Figure 3.
Structure of the root tip (https://s3-us-west-2.amazonaws.com/courses-images/wp-content/uploads/sites/1842/2017/05/26232216/figure-30-03-02.jpeg accessed on March 21, 2025).
Figure 3.
Structure of the root tip (https://s3-us-west-2.amazonaws.com/courses-images/wp-content/uploads/sites/1842/2017/05/26232216/figure-30-03-02.jpeg accessed on March 21, 2025).
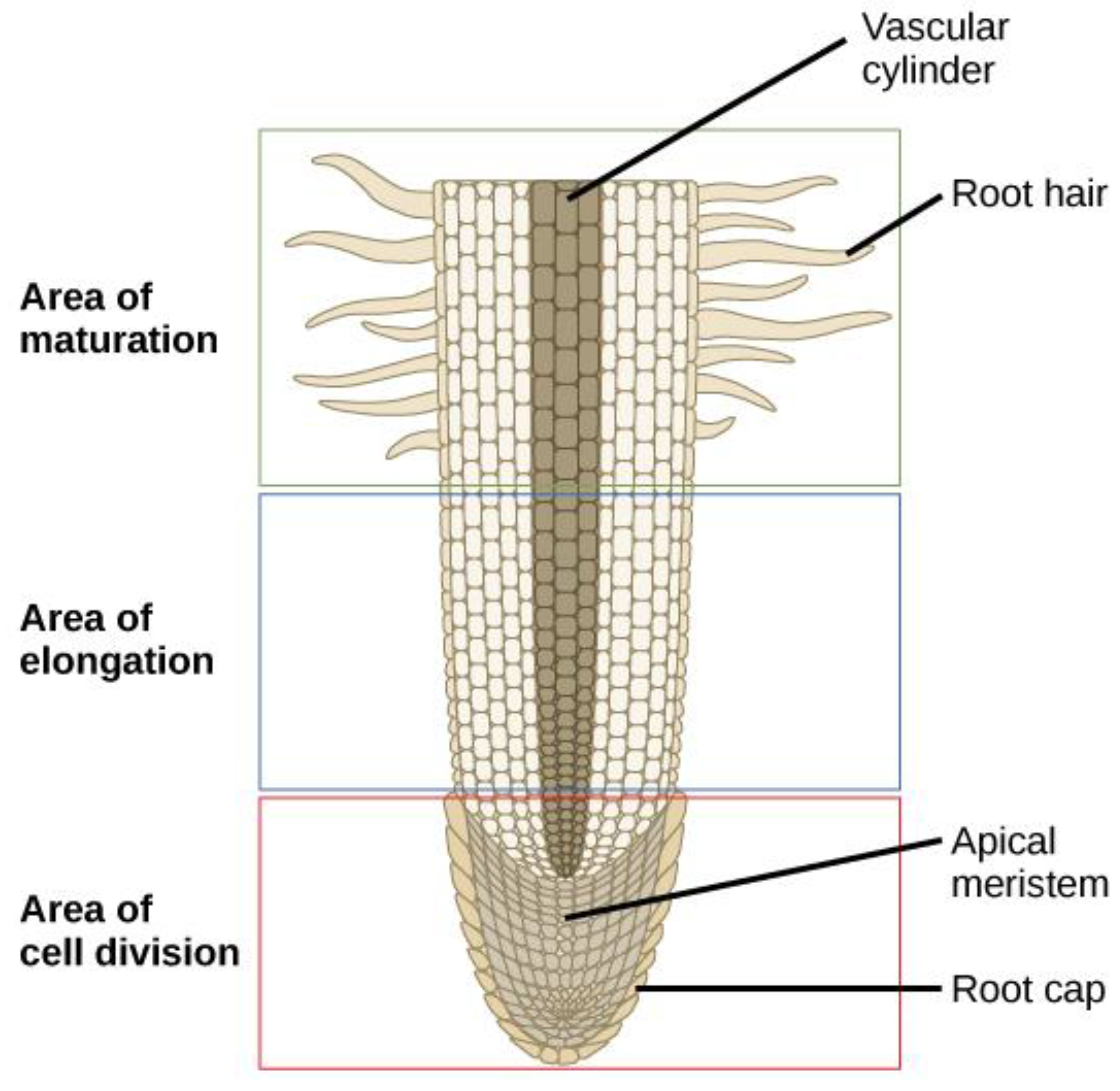
Figure 4.
Phosphorus (P) pools in photosynthetic tissues of a wide range of plants grown at different P supplies. The inset summarizes relative proportions of different P pools as the total P concentration increases; the colours of the bars correspond with the colours in the main figure (Veneklaas et al. 2012).
Figure 4.
Phosphorus (P) pools in photosynthetic tissues of a wide range of plants grown at different P supplies. The inset summarizes relative proportions of different P pools as the total P concentration increases; the colours of the bars correspond with the colours in the main figure (Veneklaas et al. 2012).
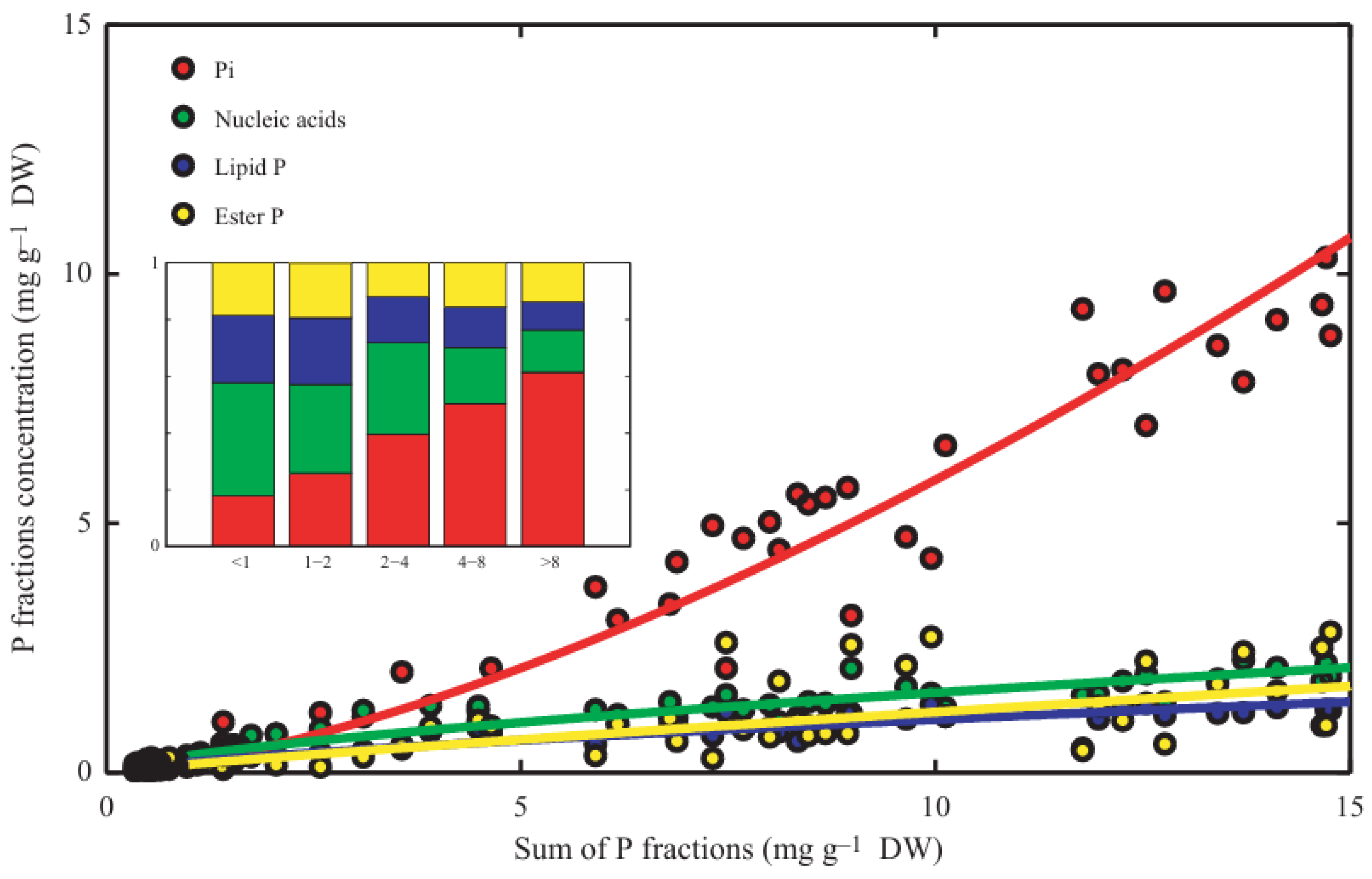
Figure 5.
The 4R nutrient stewardship concept defines the right source, rate, time, and place for plant nutrient application as those producing the economic, social, and environmental outcomes desired by all stakeholders to the soil-plant ecosystem (IPNI 2016 ).
Figure 5.
The 4R nutrient stewardship concept defines the right source, rate, time, and place for plant nutrient application as those producing the economic, social, and environmental outcomes desired by all stakeholders to the soil-plant ecosystem (IPNI 2016 ).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



