
Submitted:
02 October 2025
Posted:
03 October 2025
You are already at the latest version
Abstract
The attachment-invasion locus protein Ail of pathogenic Yersinia strains is an important virulence factor for both invasion of eucaryotic cells as well as serum resistance. In other Yersinia strains, e.g. those belonging to biotype (BT) 1A of Yersinia enterocolitica, ail has only occasionally been described. Sequence analysis of 370 BT 1A isolates in our laboratory revealed 41 (11.1%) being ail-positive. Most of these isolates were recovered from minced meat and wild boars and belong to 17 MLST allele profiles. A closer look at DNA sequences surrounding ail disclosed that the gene of most isolates is embedded in DNA regions encoding phage proteins. The genomes of four prophages belonging to four different phylogenetic clusters were determined and analysed by in silico studies. They have sizes of 34.9 and 50.7 kb and are closely related to each other, but not to known phages. Unlike other regions of the prophages, the integrases and attachment sites of some of them diverge leading to different integration sites in the isolates. In a fifth cluster, ail is relocated at a different position on the Y. enterocolitica chromosome, but surrounded by prophage-related sequences. In addition, highly pathogenic 1B/O:8 strains contain a DNA segment including ail that is similar to the prophage sequences determined in this study.
Keywords:
ail
; virulence
; Yersinia enterocolitica
; prophage
; horizontal gene transfer
Dr. Stefan Hertwig, Consiliary Laboratory for Yersinia, Department of Biological Safety, German Federal Institute for Risk Assessment, Diedersdorfer Weg 1, 12277 Berlin, Germany, telephone: +49 30 18412 74502
1. Introduction
Yersiniosis is an infectious disease of the gastrointestinal tract caused by Yersinia (Y.) enterocolitica and, to a lesser extent, by Y. pseudotuberculosis [1,2]. It is the third most common bacterial enteritis in Europe. Infections are mainly caused by the consumption of raw or undercooked pork [3,4,5,6,7,8,9]. Y. enterocolitica comprises six biotypes (BT) and up to 70 serotypes. While the BTs 1B, 2, 3, 4 and 5 possess the 70 kb virulence plasmid pYV encoding a type III secretion system and effector proteins (Yersinia outer proteins = YOPs) with in part toxic activity for eucaryotic cells, BT 1A strains are generally devoid of pYV [10]. Moreover, some important chromosomally encoded virulence factors of pathogenic biotypes are only rarely or not existing in BT 1A [11,12]. Two of those are the enterotoxin YstA and the attachment-invasion locus protein (Ail), which is involved in both the invasion of eucaryotic cells as well as in serum resistance [13,14,15]. The ail gene has yet only been detected in some BT 1A strains [16,17]. Because of the lack of these virulence factors, BT 1A strains have been considered to be non-pathogenic for a long time. However, during the last years, there is an increasing number of reports describing BT 1A isolates from clinical cases [18,19,20,21,22,23,24,25,26]. This biotype obviously comprises at least two phylogenetic lineages, each with different virulence factors, some of which are toxins [17,27]. The question arises, whether horizontal gene transfer may be involved in the heterogeneity of this biotype. In this study, we sequenced 370 BT 1A isolates from different sources, of which 41 contain the virulence gene ail. A closer analysis of this gene revealed that unlike in the strictly pathogenic biotypes, ail of most BT 1A isolates is located on the genome of prophages, which were characterized by in silico analyses.
2. Results
2.1. Forty-One Out of 370 BT 1A Genomes Contain a Prophage-Associated Ail Gene.
Sequence analyses of 370 Y. enterocolitica BT 1A isolates revealed that they harbor the gene ail. They were isolated between 2013 and 2025 from different sources (mainly minced meat and wild boars) and belong to 17 different MLST allele profiles (Figure 1A). All isolates possess the enterotoxin gene ystB, 30, 10 and 3 of them additionally the virulence-associated genes hreP, myfA and tccC, respectively (Table S1). For almost all isolates, plasmid-borne sequences of approximately 3 to 106 kb were predicted.
A closer look at the ail gene of these isolates showed that it is up to 99% identical to ail of pathogenic biotypes, e.g. the bio/serotype 1B/O:8 strain 8081. In addition, the upstream sequence containing the promoter and ribosome binding site of ail are similarly related to their counterpart in other biotypes suggesting that ail may be active in BT 1A.
However, the regions encompassing ail diverge significantly in the various biotypes. In most BT 1A isolates like in 24-YE00064 studied here, the gene is surrounded by partitioning genes and genes for cell lysis (lysin and holin) and DNA packaging (small and large terminase) typically associated with phage (Table 1). Moreover, genes for phage assembly (capsid and tail), the genetic switch as well as an integrase and excisionase were also identified in most chromosomes suggesting that ail is part of a prophage (Table S2).
2.2. Comparison of the Prophages Indicates Relationships Between Them.
The 41 prophage sequences identified in the investigated isolates by comparison with the vB_Yen-24-YE00064 prophage form five major clusters (Figure 1A), of which the prophages in the clusters C2 to C5 harbor the ail gene, whereas prophages in cluster C1 are devoid of ail, since here, the gene is located at a different position on the chromosome (see below). An alignment disclosed that most regions of the prophages, particularly those encoding structural proteins, are closely related, whereas e.g. the integrase genes show major differences (Figure 1B). Short read sequencing allowed the prediction of four whole prophage genomes (vB_Yen_16-YE00051, vB_Yen_20-YE00187, vB_Yen_24-YE00064 and vB_Yen_25-YE00027) belonging to four clusters (Figure 1A). They have genome sizes of 34,918 to 50,744 bp and are composed of 55 to 78 Open Reading Frames (ORFs, Table S1). The overall genome organization of the prophages is similar (Figure 2A). As with other temperate phages, ORFs for repressor proteins, cell lysis, DNA packaging, capsid and tail assembly are clustered, even though some ORFs, particularly those encoding structural proteins, are missing in vB_Yen_16-YE00051. The prophages showed no identities to other phages and only some relatedness to two Y. enterocolitica BT 1A chromosomes (Y201, CP124238.1 and Y115, CP124259.1).
Three prophages (vB_Yen_16-YE00051, vB_Yen_20-YE00187 and vB_Yen_25-YE00027) have an identical attachment site att of 40 bp (Figure 2B). Their integrases are 100% identical (Figure 2C). By contrast, the prophage vB_Yen_24-YE00064 has an att site of only 27 bp (Figure 2B). The integrase of this prophage is only approximately 35% identical to those of the other ones (Figure 2C). Thus, it is not surprising that the two groups have different integration sites on the bacterial chromosome. While the prophages vB_Yen_16-YE00051, vB_Yen_20-YE00187 and vB_Yen_25-YE00027 are integrated between two genes for hypothetical proteins, vB_Yen_24-YE00064 is integrated between a gene for an integrase and a YebY family protein.
2.3. Relocation of ail in Cluster C1 and Analysis of 1B/O:8 Strains.
Sequence analysis of cluster C1 revealed similar prophage sequences as in isolate 24-YE00064. However, at the position of ail in vB_Yen_24-YE00064, there is a gap in the C1 prophage genomes (Figure 3A). In this cluster, ail is located approximately 600 kb apart from the vB_Yen_24-YE00064-related prophage sequences. Interestingly enough, the gene and its adjacent sequences are surrounded by DNA segments, which are similarly present in the corresponding prophages, as shown for the prophage vB_Yen_23-YE00044.2 (Figure 3B). The homologous upstream and downstream sequences of ail have a length of 144 bp and 925 bp, respectively, but are not related to each other. Therefore, it remains open how ail was relocated in isolates belonging to cluster C1. Nevertheless, it is noteworthy that even in the highly pathogenic Y. enterocolitica 1B/O:8 strains 8081 (AM286415.1), WA (CP009367.1) and Billups-1803-68 (CP173224.1), ail is associated with phage genes. Indeed, a stretch of approximately 20 kb of strain 8081 containing ail is similar to vB_Yen_24-YE00064 (Figure 3C). This stretch essentially corresponds to ØYE200 identified in 8081 (Thomson et. al, 2006), which, however, has been determined as a smaller prophage (15.5 kb) without ail. Besides ail, the 20 kb prophage of strain 8081 comprises genes for e.g. an integrase, lysis proteins (holin and lysin) and the terminase large subunit. Moreover, the fact that this DNA segment also contains the 27 bp attachment site of vB_Yen_24-YE00064 upstream of the integrase gene and that is linked to tRNA genes suggests that ail was once associated with a similar prophage. It is conspicuous that the DNA segment in strain 8081 harbors several transposase genes which are lacking in vB_Yen_24-YE00064 and which might have been involved in genetic reassortments.
3. Discussion
The attachment-invasion locus protein Ail of Y. enterocolitica is an important virulence factor, which is produced by all pathogenic biotypes of this species, as well as by Y. pseudotuberculosis and Y. pestis [28]. It has yet only rarely been described in BT 1A strains of Y. enterocolitica and in Y. enterocolitica-like species, e.g. Y. kristensenii, some of which have been reported to contain an additional ail-related gene which may be associated with plasmids or phages [11,29,30]. For that reason, ail is routinely used as target for the detection of pathogenic Y. enterocolitica and Y. pseudotuberculosis by RT-PCR (ISO TS 18867:2015). Though, sequencing of 370 BT 1A isolates showed that ail is more commonly present in this biotype as expected. We identified the gene in 41 (11.1%) out of 370 isolates recovered from food and wild boars in the last 12 years. The ail-positive isolates represent a broad range of MLST alleles profiles, even though some types (ST304, ST428, ST832) were prevailing. The question arises, how BT 1A strains may acquire ail. This study suggests that it may occur by lysogenic conversion via temperate phages. Indeed, the analysis of ail-positive BT 1A isolates showed that in most of them the gene is located on a prophage. The prophages are related to each other, but form five different clusters. Up to now, four prophage genomes containing ail could be analysed in detail. The analysis suggests that three of them (vB_Yen_20-YE00187, vB_Yen_24-YE00064 and vB_Yen_25-YE00027) may be active, since all elements required for the formation of a phage particle are obviously present, whereas the prophage vB_Yen_16-YE00051 is presumably defective, because some essential ORFs for capsid and tail proteins are lacking. The fact that genes for structural proteins of the complete prophages are very similar indicates that the corresponding phage particles may have the same morphology. A striking difference, however, pertains to the integration site of them on the Y. enterocolitica chromosome. Unlike some other parts of their genomes, genes for the integrase and the attachment sites are in part highly diverse. As a consequence, the prophages are not integrated at the same position in BT 1A strains. Whether these sites also exist in other Y. enterocolitica biotypes or even other Yersinia species and whether those strains may also acquire ail by phage-mediated transfer has still to be studied. It has to be taken into account that for lysogenic conversion, the host range of a temperate phage is of major importance. Yersinia enterocolitica BT 1A strains belong to various serotypes, which may determine the host specificity of a phage. It appears, however, that ail-prophages are subjected to genetic recombinations. This can be clearly seen in cluster C1 comprising related prophages, in which ail including adjacent sequences has been relocated. Similarly, highly pathogenic 1B strains like 8081 contain remnants of ail-prophages suggesting recombination events or even a horizontal gene transfer in the past. Regrettably, information on temperate Y. enterocolitica phages and their potential to exchange genes is scarce [31,32]. We will therefore now determine the inducibility of the identified ail-prophages and phenotypic properties of possibly produced phage particles.
4. Materials and Methods
4.1. Typing of Y. enterocolitica Strains.
Isolates were initially cultivated on Columbia agar supplemented with 5% sheep blood (bioMérieux Deutschland GmbH, Nürtingen, Germany) at 28°C for 16-20 hours for whole-cell matrix-assisted laser-desorption/ionization time-of-flight mass spectrometrical identification (MALDI-TOF MS) using the direct transfer method with HCCA matrix on a Biotyper (Bruker Daltonics GmbH & Co. KG, Bremen, Germany). In addition, physiological and biochemical tests using classical tube and plate procedures for species confirmation and biochemical differentiation were conducted as previously described [33]. Unless otherwise indicated, YP cultivation was conducted at aerobic conditions at 28°C for 18-24 hours using lysogeny broth (LB)-based media. For solid media preparation, LB medium was supplemented with 1.8% bacto agar no. 1 (Oxoid Deutschland GmbH, Wesel, Germany) [34,35].
4.2. Genome Sequencing and Bioinformatics Analysis
Whole-genome sequencing (WGS) of Y. enterocolitica BT 1A isolates was performed by short-read, paired-end sequencing (2 x150 cycles) on a NextSeq500 benchtop device (Illumina Inc., San Diego, CA, USA). Bacterial genomic DNA was extracted from liquid cultures grown at 37°C for 20-24 hours using the PureLink Genomic DNA Mini Kit (Invitrogen, Ebersberg, Germany) according to the recommendation of the manufacturers. DNA sequencing libraries were prepared using the Nextera XT DNA Sample Preparation Kit (Illumina Inc.) [31,36,37,38]. Raw sequencing data were subjected to the Aquamis pipeline (Deneke et al., 2021) for quality evaluation, demultiplexing and trimming, while general in silico typing purposes were conducted using BakCharak (https://gitlab.com/bfr_bioinformatics/bakcharak; access: Jul-2025) [39]. Prophage detection was conducted using the Phastest tool for initial screening, while manual curation using Accelrys DS Gene (v2.5; Accelrys Inc., San Diego, CA, USA) was performed to determine the complete prophage sequence from attachment sites of the bacteria (attB) and prophages (attP). Genome annotation of the prophage genomes was conducted using the Bacterial and Viral Bioinformatics Resource Center (BV-BRC). Initial functional prediction of open reading frames (ORFs) was manually curated according to predicted functions of closely related protein sequences derived from blastp searches at NCBI (National Center for Biotechnology Information) [31,36,37,38]. Phylogenetic relationship of vB_Yen_24-YE00064 (reference) to Y. enterocolitica genome datasets encoding Ail were conducted using CSI phylogeny (v1.4; default settings; https://cge.food.dtu.dk/services/CSIPhylogeny/; access: Sep-2025)
4.3. Genome Accession Numbers
Deposition of prophage genomes at NCBI Genbank was conducted using BankIt for the prophages vB_Yen_24-YE00064 (PV779719), vB_Yen_16-YE00051 (SUBMITTED), vB_Yen_20-YE00187 (PX109664) and vB_Yen_25-YE00027 (PX109665).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: in-silico typing features of Y. enterocolitica genomes analyzed in this study; Table S2: Annotation of the prophage genomes vB_Yen_24-YE00064, vB_Yen_25-YE00027, vB_Yen_20-YE00187 and vB_Yen_16-YE00051.
Author Contributions
Conceptualization, Stefan Hertwig and Jens Hammerl; Data curation, Jens Hammerl; Formal analysis, Stefan Hertwig and Jens Hammerl; Funding acquisition, Stefan Hertwig and Jens Hammerl; Investigation, Stefan Hertwig and Jens Hammerl; Methodology, Stefan Hertwig and Jens Hammerl; Project administration, Stefan Hertwig; Resources, Stefan Hertwig; Software, Jens Hammerl; Validation, Stefan Hertwig and Jens Hammerl; Visualization, Jens Hammerl; Writing – original draft, Stefan Hertwig and Jens Hammerl; Writing – review & editing, Jens Hammerl.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original data presented in the study are openly available in Genbank under accession numbers PV779719 (vB_Yen_24-YE00064), SUBMITTED (vB_Yen_16-YE00051), PX109664 (vB_Yen_20-YE00187) and PX109665 (vB_Yen_25-YE00027).
Acknowledgments
The authors thank the Bundesinstitut für Risikobewertung for financial support of this study (grant no. 45-003 & 45-002).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Carniel, E. 2001. The Yersinia high-pathogenicity island: an iron-uptake island. Microbes Infect 3:561-9.
- panel) EFSAB. 2007. Monitoring and identification of human enteropathogenic Yersinia spp. ESFA Journal (2007) 595:1-30.
- Boqvist S, Pettersson H, Svensson A, Andersson Y. 2009. Sources of sporadic Yersinia enterocolitica infection in children in Sweden, 2004: a case-control study. Epidemiol Infect 137:897-905.
- Centorame P, Sulli N, De Fanis C, Colangelo OV, De Massis F, Conte A, Persiani T, Marfoglia C, Scattolini S, Pomilio F, Tonelli A, Prencipe VA. 2017. Identification and characterization of Yersinia enterocolitica strains isolated from pig tonsils at slaughterhouse in Central Italy. Vet Ital 53:331-344.
- Fredriksson-Ahomaa M, Korte T, Korkeala H. 2000. Contamination of carcasses, offals, and the environment with yadA-positive Yersinia enterocolitica in a pig slaughterhouse. J Food Prot 63:31-5.
- Fredriksson-Ahomaa M, Korte T, Korkeala H. 2001. Transmission of Yersinia enterocolitica 4/O:3 to pets via contaminated pork. Lett Appl Microbiol 32:375-8.
- Fredriksson-Ahomaa M, Stolle A, Korkeala H. 2006. Molecular epidemiology of Yersinia enterocolitica infections. FEMS Immunol Med Microbiol 47:315-29.
- Ostroff SM, Kapperud G, Hutwagner LC, Nesbakken T, Bean NH, Lassen J, Tauxe RV. 1994. Sources of sporadic Yersinia enterocolitica infections in Norway: a prospective case-control study. Epidemiol Infect 112:133-41.
- Tauxe RV, Vandepitte J, Wauters G, Martin SM, Goossens V, De Mol P, Van Noyen R, Thiers G. 1987. Yersinia enterocolitica, 1: and pork: the missing link. Lancet 1, 1129.
- Platt-Samoraj A, Syczylo K, Szczerba-Turek A, Bancerz-Kisiel A, Jablonski A, Labuc S, Pajdak J, Oshakbaeva N, Szweda W. 2017. Presence of ail and ystB genes in Yersinia enterocolitica biotype 1A isolates from game animals in Poland. Vet J 221:11-13.
- Joutsen S, Johansson P, Laukkanen-Ninios R, Bjorkroth J, Fredriksson-Ahomaa M. 2020. Two copies of the ail gene found in Yersinia enterocolitica and Yersinia kristensenii. Vet Microbiol 247:108798.
- Le Guern AS, Martin L, Savin C, Carniel E. 2016. Yersiniosis in France: overview and potential sources of infection. Int J Infect Dis 46:1-7.
- Kirjavainen V, Jarva H, Biedzka-Sarek M, Blom AM, Skurnik M, Meri S. 2008. Yersinia enterocolitica, e: proteins YadA and ail bind the complement regulator C4b-binding protein. PLoS Pathog 4, 1000.
- Thomson JJ, Plecha SC, Krukonis ES. 2019. Ail provides multiple mechanisms of serum resistance to Yersinia pestis. Mol Microbiol 111:82-95.
- Miller VL, Beer KB, Heusipp G, Young BM, Wachtel MR. 2001. Identification of regions of Ail required for the invasion and serum resistance phenotypes. Mol Microbiol 41:1053-62.
- Kraushaar B, Dieckmann R, Wittwer M, Knabner D, Konietzny A, Made D, Strauch E. 2011. Characterization of a Yersinia enterocolitica biotype 1A strain harbouring an ail gene. J Appl Microbiol 111:997-1005.
- Sihvonen LM, Jalkanen K, Huovinen E, Toivonen S, Corander J, Kuusi M, Skurnik M, Siitonen A, Haukka K. 2012. Clinical isolates of Yersinia enterocolitica biotype 1A represent two phylogenetic lineages with differing pathogenicity-related properties. BMC Microbiol 12:208.
- Hunter E, Greig DR, Schaefer U, Wright MJ, Dallman TJ, McNally A, Jenkins C. 2019. Identification and typing of Yersinia enterocolitica and Yersinia pseudotuberculosis isolated from human clinical specimens in England between 2004 and 2018. J Med Microbiol 68:538-548.
- McNally A, Cheasty T, Fearnley C, Dalziel RW, Paiba GA, Manning G, Newell DG. 2004. Comparison of the biotypes of Yersinia enterocolitica isolated from pigs, cattle and sheep at slaughter and from humans with yersiniosis in Great Britain during 1999-2000. Lett Appl Microbiol 39:103-8.
- McNally A, Dalton T, Ragione RM, Stapleton K, Manning G, Newell DG. 2006. Yersinia enterocolitica, 1: of differing biotypes from humans and animals are adherent, invasive and persist in macrophages, but differ in cytokine secretion profiles in vitro. J Med Microbiol 55, 1725.
- Rivas L, Horn B, Armstrong B, Wright J, Strydom H, Wang J, Paine S, Thom K, Orton A, Robson B, Lin S, Wong J, Brunton C, Smith D, Cooper J, Mangalasseril L, Thornley C, Gilpin B. 2024. A case-control study and molecular epidemiology of yersiniosis in Aotearoa New Zealand. J Clin Microbiol 62:e0075424.
- Rivas L, Strydom H, Paine S, Wang J, Wright J. 2021. Yersiniosis in New Zealand. Pathogens 10.
- Stephan R, Joutsen S, Hofer E, Sade E, Bjorkroth J, Ziegler D, Fredriksson-Ahomaa M. 2013. Characteristics of Yersinia enterocolitica biotype 1A strains isolated from patients and asymptomatic carriers. Eur J Clin Microbiol Infect Dis 32:869-75.
- Clarke M, Dabke G, Strakova L, Jenkins C, Saavedra-Campos M, McManus O, Paranthaman K. 2020. Introduction of PCR testing reveals a previously unrecognized burden of yersiniosis in Hampshire, UK. J Med Microbiol 69:419-426.
- Colbran C, May F, Alexander K, Hunter I, Stafford R, Bell R, Cowdry A, Vosti F, Jurd S, Graham T, Micalizzi G, Graham R, Slinko V. 2024. Yersiniosis outbreaks in Gold Coast residential aged care facilities linked to nutritionally-supplemented milkshakes, January-23. Commun Dis Intell (2018) 48. 20 April.
- Horn BP, I.; Cressey, P.; Armstrong, B.; Lopez, L. 2023. Annual report concerning Foodborne Diseases in New Zealand 2022. New Zealand Food Safety Technical Paper No: 2023/17.
- Platt-Samoraj, A. 2022. Toxigenic Properties of Yersinia enterocolitica Biotype 1A. Toxins (Basel) 14.
- Kolodziejek AM, Hovde CJ, Minnich SA. 2012. Yersinia pestis, 1: roles of a single protein. Front Cell Infect Microbiol 2.
- Stevens MJA, Horlbog JA, Diethelm A, Stephan R, Nuesch-Inderbinen M. 2024. Characteristics and comparative genome analysis of Yersinia enterocolitica and related species associated with human infections in Switzerland 2019-2023. Infect Genet Evol 123:105652.
- Sihvonen LM, Hallanvuo S, Haukka K, Skurnik M, Siitonen A. 2011. The ail gene is present in some Yersinia enterocolitica biotype 1A strains. Foodborne Pathog Dis 8:455-7.
- Hammerl JA, Barac A, Jackel C, Fuhrmann J, Gadicherla A, Hertwig S. 2022. Phage vB_YenS_P400, a Novel Virulent Siphovirus of Yersinia enterocolitica Isolated from Deer. Microorganisms 10.
- Popp A, Hertwig S, Lurz R, Appel B. 2000. Comparative study of temperate bacteriophages isolated from Yersinia. Syst Appl Microbiol 23:469-78.
- Wauters G, Kandolo K, Janssens M. 1987. Revised biogrouping scheme of Yersinia enterocolitica. Contrib Microbiol Immunol 9:14-21.
- Hammerl JA, Klein I, Lanka E, Appel B, Hertwig S. 2008. Genetic and functional properties of the self-transmissible Yersinia enterocolitica plasmid pYE854, which mobilizes the virulence plasmid pYV. J Bacteriol 190:991-1010.
- Hammerl JA, Rivas L, Cornelius A, Manta D, Hertwig S. 2025. The beta-glucosidase gene for esculin hydrolysis of Yersinia enterocolitica is a suitable target for the detection of biotype 1A strains by polymerase chain reaction. J Appl Microbiol 136.
- Brauer JA, Hammerl JA, El-Mustapha S, Fuhrmann J, Barac A, Hertwig S. 2023. The Novel Yersinia enterocolitica Telomere Phage vB_YenS_P840 Is Closely Related to PY54, but Reveals Some Striking Differences. Viruses 15.
- Hammerl JA, Barac A, Erben P, Fuhrmann J, Gadicherla A, Kumsteller F, Lauckner A, Muller F, Hertwig S. 2021. Properties of Two Broad Host Range Phages of Yersinia enterocolitica Isolated from Wild Animals. Int J Mol Sci 22.
- Hammerl JA, El-Mustapha S, Bolcke M, Trampert H, Barac A, Jackel C, Gadicherla AK, Hertwig S. 2022. Host Range, Morphology and Sequence Analysis of Ten Temperate Phages Isolated from Pathogenic Yersinia enterocolitica Strains. Int J Mol Sci 23.
- Deneke C, Brendebach H, Uelze L, Borowiak M, Malorny B, Tausch SH. 2021. Species-Specific Quality Control, Assembly and Contamination Detection in Microbial Isolate Sequences with AQUAMIS. Genes (Basel) 12.
Figure 1.
Relationship of prophages containing an ail gene. A. Clusters of similar prophage genomes in Y. enterocolitica BT 1A isolates, obtained by comparison with the vB_Yen_24-YE00064 prophage (R) of cluster 3. The ST type of their hosts as well as the source and year of isolation are stated. Red stars indicate isolates in which the whole ail prophage sequences were determined. B. Conserved DNA regions of the prophage genomes. The plot gives the numbers of related phage sequences identified in the compared WGS dataset of the isolates. The map shows the genome of the prophage vB_Yen_24-YE00064 used as reference for sequence comparison (Table 1).
Figure 1.
Relationship of prophages containing an ail gene. A. Clusters of similar prophage genomes in Y. enterocolitica BT 1A isolates, obtained by comparison with the vB_Yen_24-YE00064 prophage (R) of cluster 3. The ST type of their hosts as well as the source and year of isolation are stated. Red stars indicate isolates in which the whole ail prophage sequences were determined. B. Conserved DNA regions of the prophage genomes. The plot gives the numbers of related phage sequences identified in the compared WGS dataset of the isolates. The map shows the genome of the prophage vB_Yen_24-YE00064 used as reference for sequence comparison (Table 1).
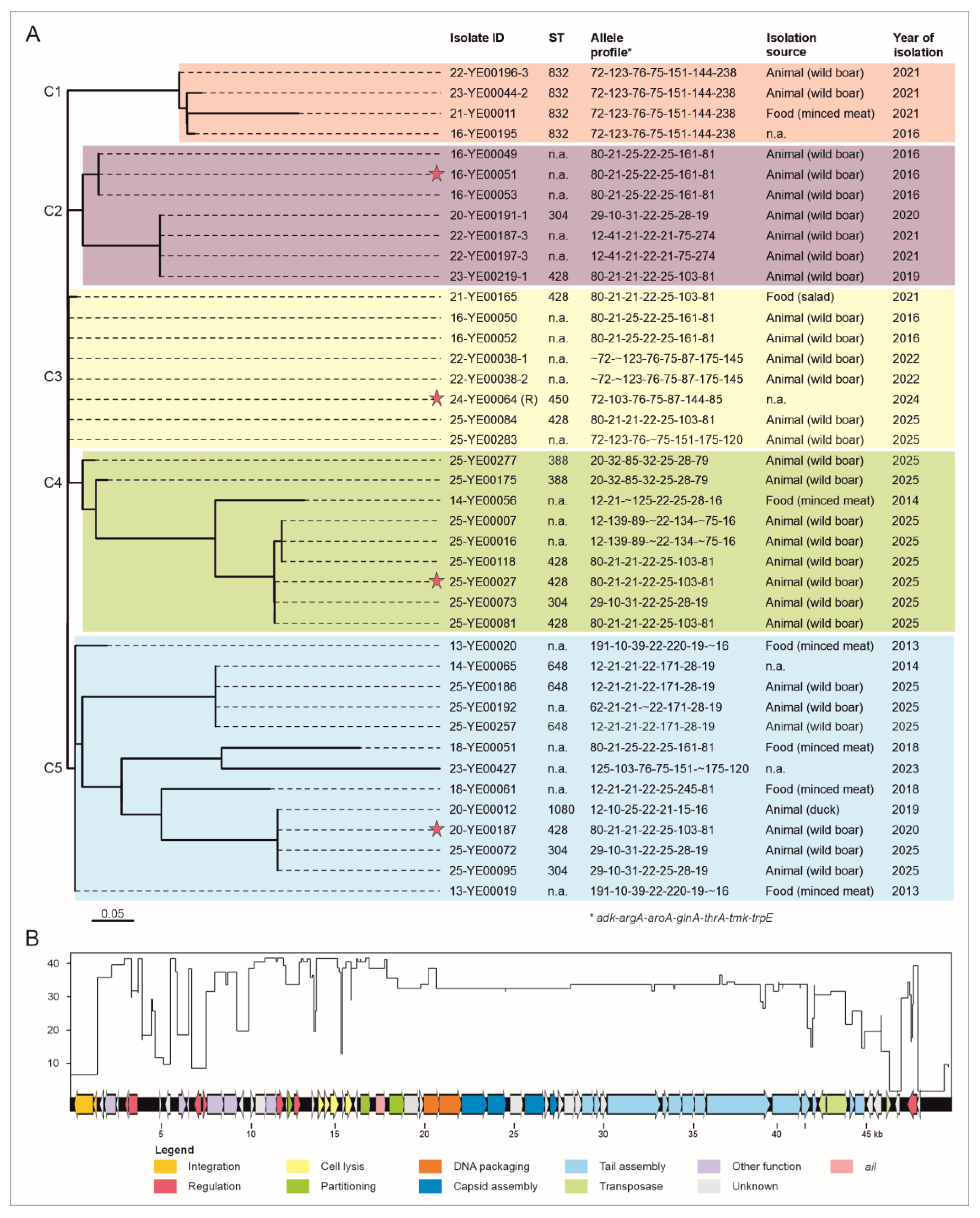
Figure 2.
Genome organization and similarities of the integrases and attachment sites of the four analysed prophages. A. Genome maps of the prophages. Sizes of the genomes and predicted functions of assigned ORF are indicated. B. Sequences of the attachment sites of the prophages. C. Similarities of their integrases.
Figure 2.
Genome organization and similarities of the integrases and attachment sites of the four analysed prophages. A. Genome maps of the prophages. Sizes of the genomes and predicted functions of assigned ORF are indicated. B. Sequences of the attachment sites of the prophages. C. Similarities of their integrases.

Figure 3.
Dot plots of the prophages vB_Yen_24-YE00064, vB_Yen_23-YE0044.2 and the ØYE200 region of strain 8081. (A) The vB_Yen_24-YE00064 ail-prophage is similar to the cluster C1 prophage vB_Yen_23-YE0044.2 lacking ail. (B) In isolate 23-YE00044.2, ail is located on a different contig (C46) than the prophage, but surrounded by prophage-related sequences. The dot plot shows only the small part of vB_Yen_23-YE0044.2, which is similar to the ail region. (C) vB_Yen_24-YE00064 is related to the ØYE200 prophage region of strain 8081, which also includes ail and an abridged terminase large subunit gene.
Figure 3.
Dot plots of the prophages vB_Yen_24-YE00064, vB_Yen_23-YE0044.2 and the ØYE200 region of strain 8081. (A) The vB_Yen_24-YE00064 ail-prophage is similar to the cluster C1 prophage vB_Yen_23-YE0044.2 lacking ail. (B) In isolate 23-YE00044.2, ail is located on a different contig (C46) than the prophage, but surrounded by prophage-related sequences. The dot plot shows only the small part of vB_Yen_23-YE0044.2, which is similar to the ail region. (C) vB_Yen_24-YE00064 is related to the ØYE200 prophage region of strain 8081, which also includes ail and an abridged terminase large subunit gene.
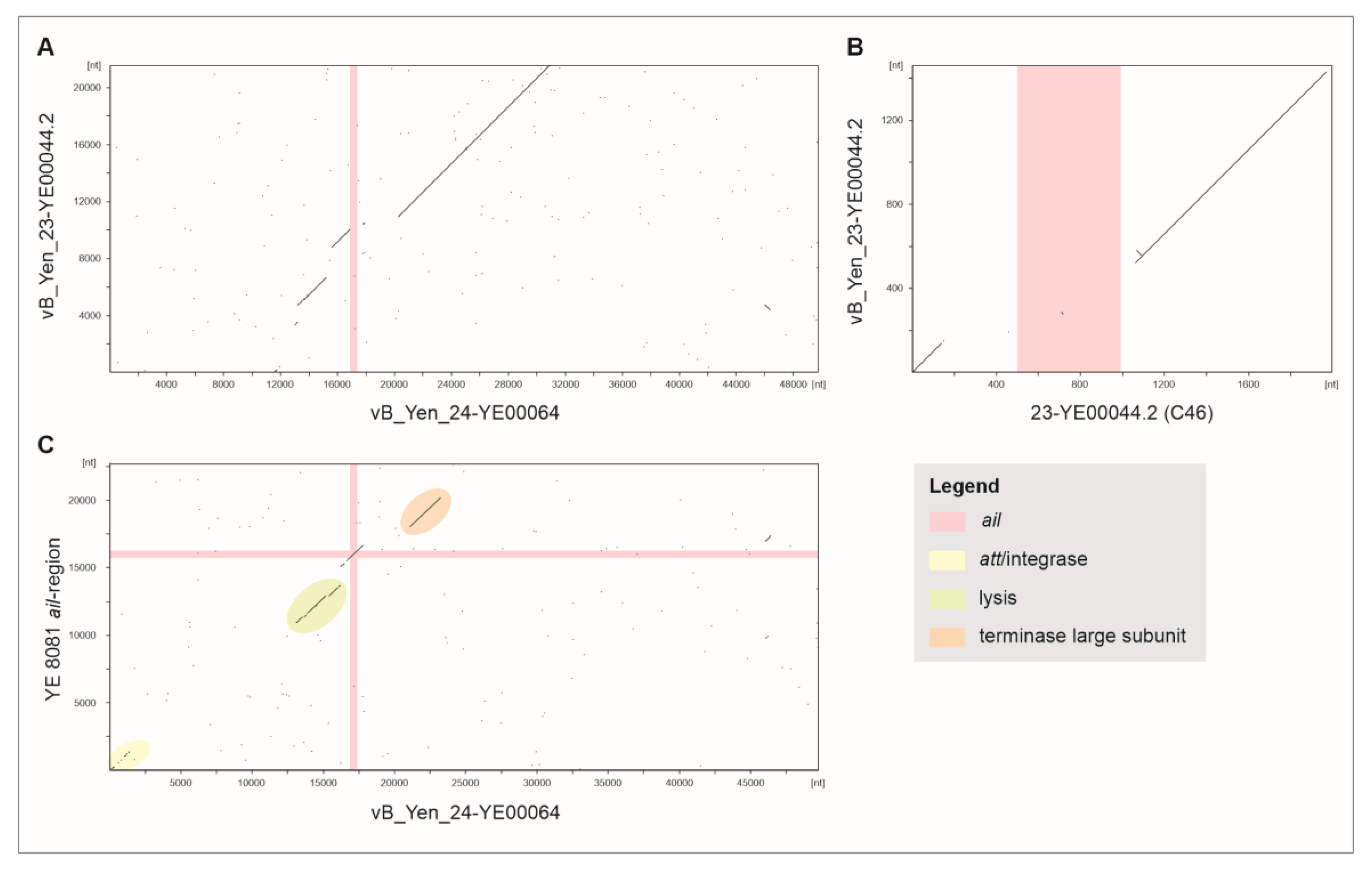

Table 1.
ORF analysis of the prophage vB_Yen_24-YE00064.
| Element | Start | Stop | Strand | Predicted function | Accession | E-value |
| ORF01 | 1229 | 147 | - | Phage integrase | WP_339099454 | 0 |
| ORF02 | 1473 | 1204 | - | Phage excisionase | WP_032819545 | 9,65E-58 |
| ORF03 | 1850 | 1548 | - | Unknown | WP_050123456 | 9,16E-67 |
| ORF04 | 2496 | 1873 | - | Single-stranded DNA-binding protein | WP_050123460 | 8,2E-151 |
| ORF05 | 2676 | 2506 | - | Unknown | WP_219644695 | 8,92E-32 |
| ORF06 | 3206 | 3033 | - | Phage CIII repressor | CNH66013 | 3,38E-33 |
| ORF07 | 3745 | 3230 | - | Phage AntA/AntB antirepressor | EKN4711341 | 3,1E-123 |
| ORF08 | 4959 | 5075 | + | Unknown | - | - |
| ORF09 | 5588 | 5280 | - | Unknown | WP_219644503 | 9,54E-66 |
| ORF10 | 6039 | 6506 | + | Phage super-infection exclusion protein B | EKN5118889 | 1,6E-107 |
| ORF11 | 6667 | 6509 | - | Unknown | WP_400175511 | 2,44E-28 |
| ORF12 | 7338 | 6916 | - | Phage CI-like repressor | WP_219649054 | 6,25E-98 |
| ORF13 | 7442 | 7699 | + | Phage Cro/Cl family transcriptional regulator | WP_151432101 | 1,28E-56 |
| ORF14 | 7686 | 8636 | + | DNA-binding protein | WP_258018631 | 0 |
| ORF15 | 8626 | 9372 | + | Replisome organizer | EKN4711333 | 0 |
| ORF16 | 9741 | 9454 | - | Unknown | WP_400175508 | 1,3E-63 |
| ORF17 | 10129 | 10335 | + | Unknown | WP_050123484 | 5,88E-40 |
| ORF18 | 10421 | 11020 | + | Unknown | EKN4711330 | 1,3E-143 |
| ORF19 | 11020 | 11592 | + | Phage NinG rap recombination | WP_400175506 | 3,3E-138 |
| ORF20 | 11592 | 12005 | + | Phage antitermination protein Q | WP_219648280 | 6,23E-97 |
| ORF21 | 12185 | 12090 | - | Unknown | EHB0983027 | 0,008902 |
| ORF22 | 12226 | 12510 | + | Type II toxin-antitoxin system (RelE/ParE family) | EKN5956923 | 3,7E-59 |
| ORF23 | 12583 | 12957 | + | Transcriptional regulator | WP_050162972 | 1,09E-84 |
| tRNA01 | 13325 | 13400 | + | tRNA-Thr-CGT | ||
| tRNA02 | 13402 | 13476 | + | tRNA-Gly-TCC | ||
| ORF24 | 13653 | 13772 | + | Unknown | WP_144405165 | 9,81E-16 |
| ORF25 | 13925 | 14320 | + | Phage holin | WP_050123331 | 3,02E-88 |
| ORF26 | 14320 | 14616 | + | Phage holin family protein | WP_219647003 | 1,4E-62 |
| ORF27 | 14603 | 15145 | + | Phage lysozyme (N-acetylmuramidase) family | HDL7801217 | 1,6E-125 |
| ORF28 | 15305 | 15418 | + | Unknown | WP_219644478 | 1,2E-15 |
| ORF29 | 15474 | 15869 | + | Phage endopeptidase Rz | EKN4799071 | 3,51E-87 |
| ORF30 | 16181 | 15999 | - | Unknown | SRY18578 | 1,42E-06 |
| ORF31 | 16338 | 16895 | + | KilA-N domain-containing protein | WP_219644727 | 9,9E-132 |
| ORF32 | 17202 | 17738 | + | Attachment invasion locus protein Ail | WP_219647006 | 4E-126 |
| ORF33 | 17951 | 18820 | + | Chromosome (plasmid) partitioning protein ParB | CNF12705 | 0 |
| ORF34 | 18813 | 19685 | + | Unknown | WP_050130101 | 0 |
| ORF35 | 19670 | 19918 | + | Unknown | WP_050130103 | 9,51E-50 |
| ORF36 | 19922 | 20788 | + | Phage terminase, small subunit | WP_258018632 | 0 |
| ORF37 | 20766 | 22070 | + | Phage terminase, large subunit | WP_050123349 | 0 |
| ORF38 | 22075 | 23487 | + | DNA-binding protein | WP_050123375 | 0 |
| ORF39 | 23492 | 24604 | + | Phage head morphogenesis protein | ELI7924874 | 0 |
| ORF40 | 24795 | 25547 | + | Unknown | WP_151431638 | 9,3E-180 |
| ORF41 | 25602 | 26747 | + | Phage major capsid protein | WP_242365527 | 0 |
| ORF42 | 26814 | 27002 | + | Unknown | WP_219647076 | 1,39E-34 |
| ORF43 | 27014 | 27496 | + | DnaT-like ssDNA-binding protein | WP_050123385 | 1,4E-114 |
| ORF44 | 27500 | 27853 | + | Unknown | WP_050123386 | 6,52E-79 |
| ORF45 | 27856 | 28446 | + | Unknown | WP_151431635 | 1E-141 |
| ORF46 | 28443 | 28862 | + | Unknown | WP_050123389 | 4,56E-98 |
| ORF47 | 28880 | 29542 | + | Phage tail protein | WP_050123390 | 1,4E-159 |
| ORF48 | 29565 | 29921 | + | Phage tail assembly chaperone | WP_050123392 | 1,56E-80 |
| ORF49 | 29924 | 30235 | + | Unknown | WP_373368631 | 6,9E-70 |
| ORF50 | 30232 | 33339 | + | Phage tail, tail length tape-measure protein H | WP_219651824 | 0 |
| ORF51 | 33412 | 33753 | + | Phage tail tip, assembly protein M | WP_050123394 | 1,7E-77 |
| ORF52 | 33762 | 34514 | + | Phage tail tip, assembly protein L | WP_219654916 | 0 |
| ORF53 | 34517 | 35233 | + | Phage tail tip, assembly protein K | MFM1259745 | 8,9E-178 |
| ORF54 | 35233 | 35838 | + | Phage tail tip, assembly protein I | MFM1259744 | 9,4E-141 |
| ORF55 | 35851 | 39552 | + | Phage tail tip, host specificity protein J | MFM1259743 | 0 |
| ORF56 | 39619 | 41256 | + | Phage tail fiber protein | WP_289823745 | 0 |
| ORF57 | 41256 | 41783 | + | Phage tail fiber assembly protein | MFM1259741 | 8,3E-124 |
| ORF58 | 41881 | 42189 | + | Phage tail fiber protein | WP_400175531 | 8,95E-65 |
| ORF59 | 42650 | 42225 | - | Transposase | WP_050132355 | 3,3E-98 |
| ORF60 | 42708 | 43871 | + | Transposase | WP_400175835 | 0 |
| ORF61 | 43997 | 44314 | + | Phage tail fiber protein | MFJ1219555 | 1,36E-65 |
| ORF62 | 44321 | 44884 | + | Phage tail fiber assembly protein | WP_050162967 | 1,7E-135 |
| ORF63 | 45339 | 44956 | - | Unknown | WP_032820973 | 1,07E-84 |
| ORF64 | 45809 | 45339 | - | Unknown | WP_004392760 | 6,9E-111 |
| ORF65 | 46078 | 46362 | + | Transposase, IS3/IS911 family | AJI84358 | 1,41E-55 |
| ORF66 | 46856 | 46572 | - | Unknown | WP_050123427 | 9,42E-62 |
| ORF67 | 47840 | 47244 | - | Phage antirepressor protein | WP_339099468 | 7,8E-142 |
| ORF68 | 48028 | 47837 | - | Unknown | WP_032820969 | 3,09E-37 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



