Submitted:
02 October 2025
Posted:
03 October 2025
You are already at the latest version
Abstract
Indoor cat allergens, particularly the major allergen Fel d 1 protein, represent significant environmental triggers for allergic rhinitis, asthma, and other immune-related disorders in humans. With the continuous global increase in pet ownership, cat allergen proteins are prevalent in diverse settings and can even be transmitted to pet-free locations via clothing and animal fur, thereby posing health risks to sensitized individuals. This review systematically provides the molecular characteristics, distribution patterns, and mechanisms of human sensitization to indoor cat allergen proteins. It focuses on a comparative analyses of the principles, sensitivity, and application of commonly used immunological methods (such as various modified ELISA, immunoblotting, and high-throughput multiplex detection technologies) alongside emerging real-time sensing platforms (including QCM, SAW, and LIF). Furthermore, this review summarizes key factors affecting indoor allergen concentrations, such as cat characteristics, architectural environments, human activities, and spatiotemporal variations. It also evaluates the efficacy and limitations of current allergy control strategies, covering source control (e.g., gene editing, immunomodulation), environmental management (e.g., air filtration), and medical treatments (e.g., allergen immunotherapy), and discusses future prospects. This review aims to offer a scientific foundation and systematic references for the detection, control, and public health protection related to indoor allergens.
Keywords:
allergy
; indoor cat allergen
; detection
; influencing factor
; treatment
1. Introduction
Allergic reactions are abnormal immune responses to typically environmental antigens, such as pollen, dust mites, microorganisms, and animal dander [1]. These conditions exhibit significant individual variability and include diverse clinical manifestations, such as asthma, allergic rhinoconjunctivitis, sinusitis, atopic dermatitis, angioedema, and urticaria. These symptoms may present in isolation or co-occur with other symptoms [2,3]. Asthma and atopic dermatitis, as classic allergic diseases, have seen rising global prevalence since the 1990s. Concurrently, the incidence of allergic rhinitis and asthma continues to rise [4]. In South Korea, for instance, allergic rhinitis affects approximately 37.6% of the population [5], while certain countries report asthma diagnosis rates as high as 15–20% [6]. According to the World Health Organization (WHO), the global number of asthma cases is expected to rise to 100 million by 2025 [7]. Modeling studies suggests that by 2050, over 15% of the population in key regions will be affected, amounting to an estimated 423 million cases globally [8,9,10,11].
In recent decades, allergies to furry animals have grown increasingly prevalent worldwide [7]. Epidemiological studies globally confirm that cat and dog allergens are major triggers of allergic sensitization and clinical symptoms [12]. In North America, sensitization rates to these animals can exceed 50% (Table 1). A nationwide allergy survey in China emphasized the critical role of assessing patients' specific antibody levels for accurate allergens identification, reporting that sensitization rates to cats and dogs rose from 1.33% and 0.83% in 2009 to 15.47% and 10.51% in 2021, respectively (Table 1) [13]. This increasing sensitization coincides with expanding pet ownership, indicating growing population exposure to these allergens. A large international survey across 22 countries involving over 27,000 participants revealed that 57% of households owned at least one pet, with dogs (33%) and cats (23%) being the most common [14]. In China, the number of dogs and cats kept urban households increased 99.15 million in 2019 to 120 million in 2023 [15,16]. This sustained growth in pet ownership suggests that environmental levels of cat and dog allergens will continue to increase. Pet owners can inadvertently transport allergens on clothing and hair [17], facilitating their transfer into various public spaces, such as schools, workplaces, or recreational facilities [18]. Even in households without pets, detectable levels of cat and dog allergens are often present, particularly in communities where pet ownerships is common [19].
Currently, convenient and effective control and intervention methods for managing health risks associated with indoor cat allergens remain insufficient. One report indicates that Fel d 1, the dominant cat allergen, can remain in indoor air for extended periods, up to 6–9 months, after the removal of pets from the environment [21]. Existing strategies can be categorized into four types. First, environmental management interventions aim to reduce airborne allergen levels through cleaning, ventilation, and high-efficiency filtration. However, their efficacy is often limited by the strong adhesive properties and aerodynamic behavior of allergens [22]. Second, animal-source interventions, such as administering products containing anti-Fel d 1 specific IgY, targeting the reduction of active allergen secretion from cat saliva and dander at the source [23]. Third, symptom-oriented drug therapy involves the use of antihistamines and nasal corticosteroids to alleviate bodily reactions after an allergic response occurs [24]. Fourth, allergen immunotherapy induces immune tolerance through prolonged, gradually increased exposure to allergens; however, its application is limited by extended treatment periods and heterogeneous patient responses [25,26].
In this study, a comprehensive literature review was performed on cat allergens, with systematic evaluation of their distribution characteristics, transmission pathways, and factors influencing exposure levels. Furthermore, we analyzed the principles, applicable scenarios, and utility of various detection technologies for allergen monitoring and control, aiming to propose improved strategies for the accurate detection and effective managing of indoor cat allergen proteins. Promising future research directions are also discussed.
2. Allergens from Cats
2.1. Molecular Insights of the Major Cat Allergen Fel d 1
Fel d 1 represents the dominant allergen in domestic cats [27]. At the molecular level, it is classified as a secretory protein belonging to the hemoglobin family, rather than the lipid carrier protein family [28,29]. This glycoprotein has a molecular weight of approximately 35–38 kDa [30]. Its quaternary structure consists of two identical homodimers that link noncovalently to form a tetramer, with each homodimer weighing approximately 18–19 kDa [31]. Although the biological function of Fel d 1 remains unclear, some studies reveal its structural homology with uterine hemoglobin, suggesting a potential role in skin barrier protection [32]. Alternatively, it may be involved in the transport of lipid molecules, such as steroids, hormones, or pheromones [33].
Regarding its role in sensitization, Fel d 1 is the major trigger of allergic responses to cats. Specific IgE antibodies against Fel d 1 are detectable in the serum of 80–95% of individuals allergic to cats [34], accounting for 60–90% of the total IgE response directed against cat allergens in sensitized patients [35,36]. The allergen is primarily produced in sebaceous glands and squamous epithelial cells, and its transferred to the fur through grooming and licking behaviors [37]. Skin biopsy studies further indicate that Fel d 1 is mainly secreted by sebaceous cells, with minor production in basal squamous epithelial cells, and is predominantly deposited in the epidermis and on the hair surface [38,39]. It is also found in salivary glands [40], perianal glands [41], and lacrimal glands, but is absent in urine or serum [42]. Notably, Fel d 1 levels are influenced by serval factors, including cat species [43], sex [44], sterilization status [45], and anatomical sites [46].
Figure 1.
Production, secretion, and transfer of allergen proteins (e.g. Fel d 1, Fel d 4, etc.) in cats.
Figure 1.
Production, secretion, and transfer of allergen proteins (e.g. Fel d 1, Fel d 4, etc.) in cats.
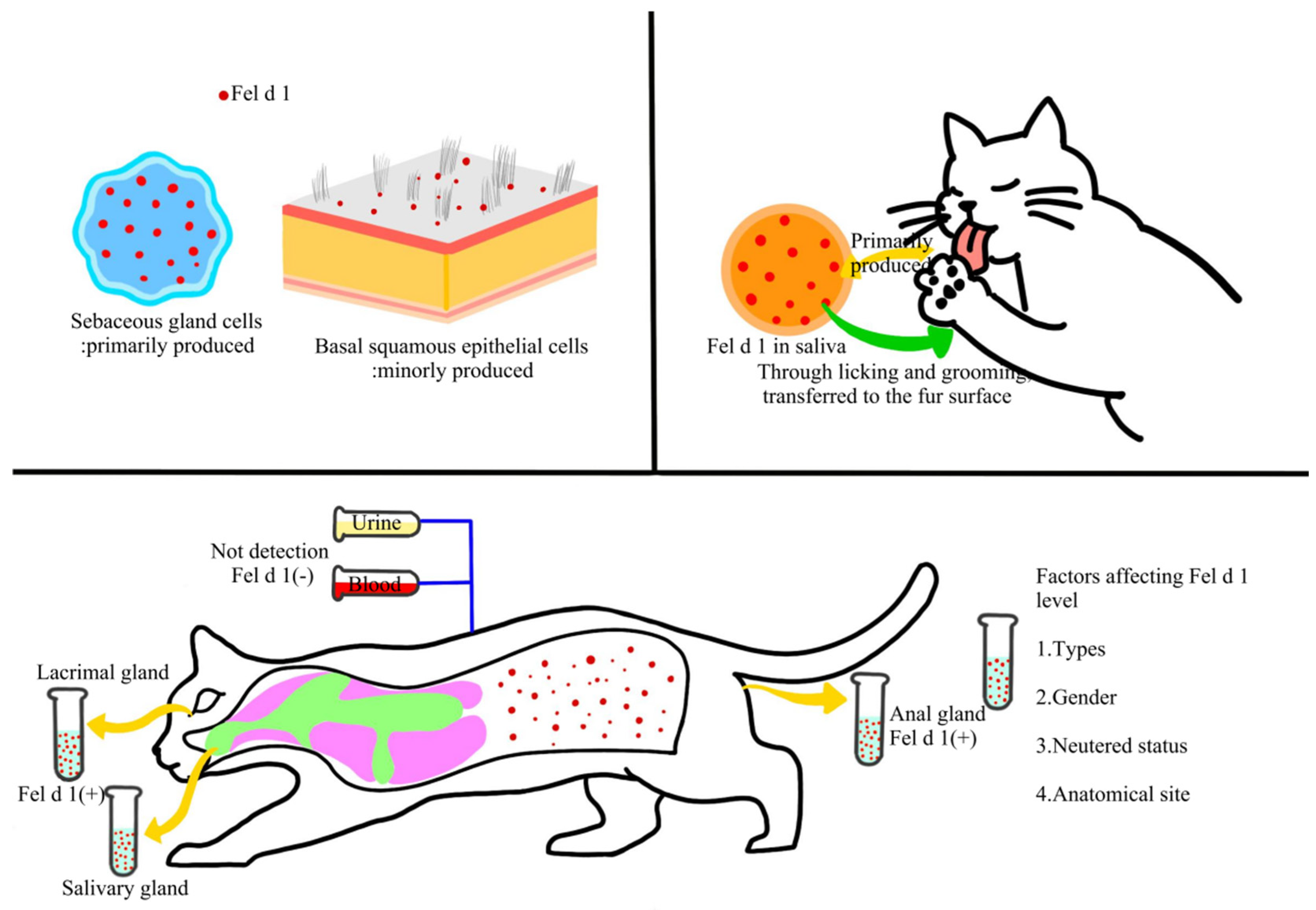
2.2. Secondary Cat Allergens and Their Cross-Reactivity Characteristics
Cats present a complex reservoir of allergen proteins. Besides the predominant allergen Fel d 1, other protein components contribute to sensitization [47]. According to the WHO/International Union of Immunological Societies (IUIS) has officially classified eight allergens from the domestic cats (Felis domesticus), designated as namely from Fel d 1 to Fel d 8 [48]. These proteins exhibit considerable diversity in both structure and biological function: Fel d 1 is a uteroglobin-like molecule, Fel d 2 is serum albumin, Fel d 3 is cystatin [49], Fel d 4 and Fel d 7 belong to the lipocalin family [50], Fel d 5 and Fel d 6 are immunoglobulins, and Fel d 8 is profiling [51]. Among these, Fel d 3, Fel d 4, and Fel d 7 are particularly notable, each capable of triggering IgE-mediated responses in more than 50% of allergic individuals to cats [52]. Notably, many of these minor allergens display high structural conservation with homologous proteins from other species, forming a molecular basis for the cross-reactivity [53,54]. For instance, Fel d 2 (serum albumin) share high sequence identity with Can f 3 from dogs [55], which can lead to cross-recognition of serum albumins from cats, dogs, and even other mammals (such as horses, mice, and cattle) [56]. Similarly, Fel d 4 and Fel d 7, as lipocalins, demonstrate functional and structural parallels with canine allergens Can f 1, Can f 2, and Can f 4 [57]; Fel d 4 shows high sequence similarity to Can f 6 from dogs [58]. These extensive cross-reactivities, combined with individual variations in sensitization, contribute to the heterogeneous clinical manifestations of allergy to cats and support the use of component-resolved diagnosis (CRD) for personalized treatment [59].
2.3. Distribution and Transmission of Cat Allergens in Indoor Environments
The particle size distribution of the major cat allergen Fel d 1 in indoor environments is influenced by multiple factors. Some studies indicate that Fel d 1 is predominantly carried on fine particles (≤5 μm) [60], which can remain suspended in the air for extended periods; however, other report a stronger association with larger particles (>4 μm) [61,62]. Despite these differences, a stable fraction of Fel d 1 bound to small particles has been detected across studies. Environmental disturbances such as air movement of human activity can resuspend large particles, thereby altering the overall size distribution [63]. The levels of Fel d 1 in residences with cats are significantly higher than those without cats [64]. Nevertheless, considerable levels of this allergen are still measurable in cat-free environments [13,19,65]. Cat allergens have also been detected in schools [66], offices [67], and public transportation systems [68], often at levels sufficient to trigger allergic symptoms in sensitized individuals. Primary sources of Fel d 1 in these settings include allergen-contaminated clothing [69] and the cross-environmental transport of cat hair. Clothing servers as a vehicle for transporting cat allergens into cat-free environments, where they are subsequently settle on surfaces and become suspended in the air [70]. Similarly, human hair represents another significant medium for the transfer and accumulation of cat allergens in public areas [71]. This mode of transmission explains why cat allergens can still be detected even in settings where strict avoidance measures are implemented [72].
2.4. Exposure Routes of Human to Cat Allergens and Sensitization Mechanisms
In humans, exposure to cat allergens (primarily Fel d 1) occurs predominantly through the respiratory tract [73], with secondary routes including conjunctival contact and limited skin exposure [74,75]. Inhalable particles, especially those smaller than 5 μm, can deliver Fel d 1 deep into the lower respiratory airways and represent the main route for asthma induction [76]. In contrast, larger particles are more likely to deposit in the upper respiratory tract and ocular mucosa, commonly leading to symptoms of allergic rhinitis and conjunctivitis [77]. In cases of compromised skin integrity, such as following a cat bite, Fel d 1 may also penetrate the skin and elicit an allergic reaction [78]. The underlying sensitization mechanism involves an exaggerated type 2 immune response [79,80]. Initial mucosal exposure to Fel d 1 leads to allergen capture by antigen-presenting cells (including dendritic cells), which then present the antigen via MHC-II molecules, resulting in T cell activation. Subsequently, activated Th2 cells release key cytokines such as interleukin-4 (IL-4), interleukin-5 (IL-5), interleukin-9 (IL-9), and interleukin-13 (IL-13), that promote the proliferation and differentiation of B cells into antibody-producing plasma cells. These plasma cells produce Fel d 1-specific IgE antibodies, which enter the circulation and bind to FcεRI receptors on the surface of mast cells and basophils, establishing a sensitized state. Upon re-exposure allergen-induced cross-linking of IgE triggers mast cell degranulation and the release of mediators such as histamine and leukotrienes, ultimately manifesting as clinical symptoms including rhinitis, conjunctivitis, and urticaria [79,81,82,83].
3. Indoor Cat Allergen Testing Methods
3.1. Immunological Assay
Several immunological methods are used to detect cat allergen proteins, with Fel d 1 detection technologies being particularly application-oriented (Table 2). Current technologies based on the enzyme-linked immunosorbent assay (ELISA) aim not only to improve sensitivity and specificity but also address key practical challenges—such as mitigating interference from Fel d 1 polymorphism in quantitative analysis and ensuring that measured results closely correlate with the protein’s in vivo allergenic activity. From rapid screening tools deployed indoor settings to high-precision quantitative standards used in manufacturing quality control [90], these approaches provide reliable technical support for accurate allergen management across diverse scenarios.
3.2. High-Throughput Technologies for the Multiplex Detection
To address the need for larger-scale screening of indoor cat allergens (e.g., Fel d 1), a variety of higher-throughput and efficient multiplex platforms have been developed beyond conventional ELISA. Fluorescence-encoded microsphere-based multiplex arrays [91] use spectrally distinct microspheres conjugated with allergen-specific antibodies. These are simultaneously analyzed via flow-cytometric detection to quantify up to six common indoor allergens [92], including Fel d 1 in parallel [93]. Molecular techniques integrating real-time quantitative PCR (qPCR) [94] with electrostatic dust collectors (EDC) can quantify cat-specific genomic DNA [95] from indoor surfaces, offering a means to estimate indirect exposure mediated by clothing or other pollutants. Furthermore, high-performance liquid chromatography-tandem mass spectrometry (HPLC-MS/MS) coupled with isobaric tagging allows not only high-throughput multiplex quantification of allergen extracts but also mechanistic analysis of dendritic-cell responses under allergen challenge [96]. Proteomic approaches have further advanced environmental allergen monitoring: studies such as those by Krutz [97] and López-Pedrouso [98] combined LC-MS/MS with bioinformatic pipelines to identify and semi-quantify allergen proteins within complex environmental samples, establishing a powerful tool for systematic multi-allergen profiling of indoor settings.
3.3. Real-Time Monitoring and Emerging Immunosensing Technologies
Bioaerosols consist of airborne particles, either dead or living airborne particles derived from plants, animals, and microorganisms [99]. These particles are extremely small (ranging from 0.001 to 100 μm in size) and lightweight, allowing them to remain suspended in the atmosphere for extended periods and travel over considerable distances, thereby posing a significant threat to human health [100]. A prominent constituent of bioaerosols is the cat allergen Fel d 1, which is often attached to particles ranging from 2 to 10 μm in size. This allergen can persist in suspension for extended durations and may be repeatedly re-aerosolized by air movement [61]. In recent years, significant advancements have been made in bioaerosol detection technologies, particularly in the fields of immunosensing and non-immunosensing methods [101]. In terms of immunosensing technology, the Quartz Crystal Microbalance (QCM) technique immobilizes Fel d 1-specific monoclonal antibodies via a cystamine self-assembled monolayer; by applying the Sauerbrey mass-frequency relationship, QCM enables label-free detection of airborne cat allergens with a sensitivity reaching the ng/cm² level [102]. Similarly, Surface Acoustic Wave (SAW) immunosensors enhance performance by tracking the slope of phase changes rather than conventional phase shift monitoring, significantly reducing response times and facilitating semi-continuous detection of allergens [103]. Within non-immunosensing technologies, Laser Particle Counters (LPC) utilize light scattering principles to perform real-time size-resolved particle counting (such as 2.5 μm, 5.0 μm, etc.), thereby allowing indirect estimation of Fel d 1 concentration and spatial distribution [104]. Ultraviolet Laser-Induced Fluorescence (UV-LIF) sensors use targeted UV wavelengths to excite endogenous fluorophores, such as proteins within bioaerosols, and analyzed the resulting fluorescence spectra for identification and quantification of biological particles. This makes UV-LIF particularly suitable for continuous, online monitoring of atmospheric bioaerosols [101]. Optical Particle Counters (OPC), also relying on light-scattering mechanisms, classify and count particles based on optical signals generated as they pass through a laser beam. OPC are widely used for cost-effective, long-term monitoring of indoor airborne particles [105]. The emergence of the Internet of Things (IoT) and smart home technologies has further accelerated the integration of multi-sensor networks into indoor environments. These systems are capable of real-time collection of diverse air quality parameters, including carbon monoxide, nitrogen dioxide, and biological contaminants, and transmit the data to central units for analysis. This integrated approach provides a basis for intelligent control of smart ventilation systems, thereby enhancing indoor environmental management [106].
4. Factors Influencing Indoor Cat Allergen Concentration
4.1. Cat-Related Factors
Individual cat characteristics significantly influence the production of Fel d 1. One study show that age is negatively correlated with skin Fel d 1 levels, with older cats exhibiting markedly lower concentrations than younger ones [107]. Pronounced anatomical variations also exist: facial skin demonstrates significantly higher allergen levels (mean 1,015.22 ± 821.6 ng cm⁻²) compared to the thoracic region (mean 115.2 ± 66.8 ng cm⁻²), a trend similarly reflected in in hair allergen concentration (face: 63.6 ± 34 µg g⁻¹; thorax: 29.6 ± 13.6 µg g⁻¹) [46]. Sex is another major factor; combined Fel d 1 concentrations across three trunk sites are higher in males (69.4 mU mL⁻¹) than in females (28.9 mU mL⁻¹, p < 0.05) [44]. This disparity is attributed to sex hormones: neutered males display significantly reduced skin Fel d 1 levels compared to intact males, whereas exogenous testosterone administration restores production to pre-neuter levels [45]. In contrast, among neutered cats, coat color, sex, and hair length exhibit no significant effect on Fel d 1 concentration [43,108].
4.2. Environmental and Architectural Factors
Environmental conditions and building characteristics play a critical role in regulating the accumulation and distribution of indoor cat allergen. In households without cats, Fel d 1 levels in settled dust demonstrate a negative correlation with relative humidity [109]. Upholstered furniture—particularly in public areas—acts as a major reservoir for allergens, with both fabric material and usage context markedly affecting Fel d 1 accumulation [67]. Within residential settings, mattress dust presents one of the dominant determinants of overall Fel d 1 concentration. Building age also shows a significant association; dwellings constructed in French after mattress Fel d 1 concentrations in constructed after 1948—particularly those built 1975–1989 and 1990–2000—are significantly lower Fel d 1 levels in mattresses compared to those built before 1948 (p < 0.05) [65].
4.3. Human Activity Factors
Human daily activities significantly influence the resuspension and removal of cat allergens. Car seats consistently contain Fel d 1 at concentrations exceeding established sensitization and symptom-provoking thresholds, irrespective of pet ownership status [110], demonstrating the role of human activity in the across-environmental transport of allergens. Additionally, vacuuming causes an immediate rise in airborne Fel d 1 levels; large particles (10.0–20.0 µm) settle below detection limits, while smaller particles (3.5–6.0 µm) remain suspended for prolonged periods [111]. Even when the vacuum cleaner is turned off, mere mechanical agitation (push/pull motions) from push/pull motions alone can resuspend large allergen-bearing particles [112]. Vacuum cleaners (VC) equipped with high-efficiency particulate air (HEPA) filters reduce indoor Fel d 1 levels significantly more effectively than non-HEPA models [113,114]. Furthermore, infrequent washing of bed linens (<1 × week⁻¹) and low bedroom ventilation rates (<1 × day⁻¹) promote allergen accumulation, reduce dispersal, and indirectly contribute to higher Fel d 1 concentrations [65].
4.4. Spatiotemporal Variation Patterns
Fel d 1 concentrations exhibit significant temporal and spatial variability. Seasonal fluctuations are observed, with levels peaking during winter and spring; levels in bed linens during spring can reach up 2.4-times those recorded in summer [115,116]. In public environments, Fel d 1 accumulates progressively over time, with high-traffic locations exhibiting accelerated accumulation rates. This pattern indicates the role of human foot traffic as a major factor in allergen transport and buildup [117].
5. Methods for Preventing and Improving Allergies
5.1. Source Control of Allergens
5.1.1. Gene Editing
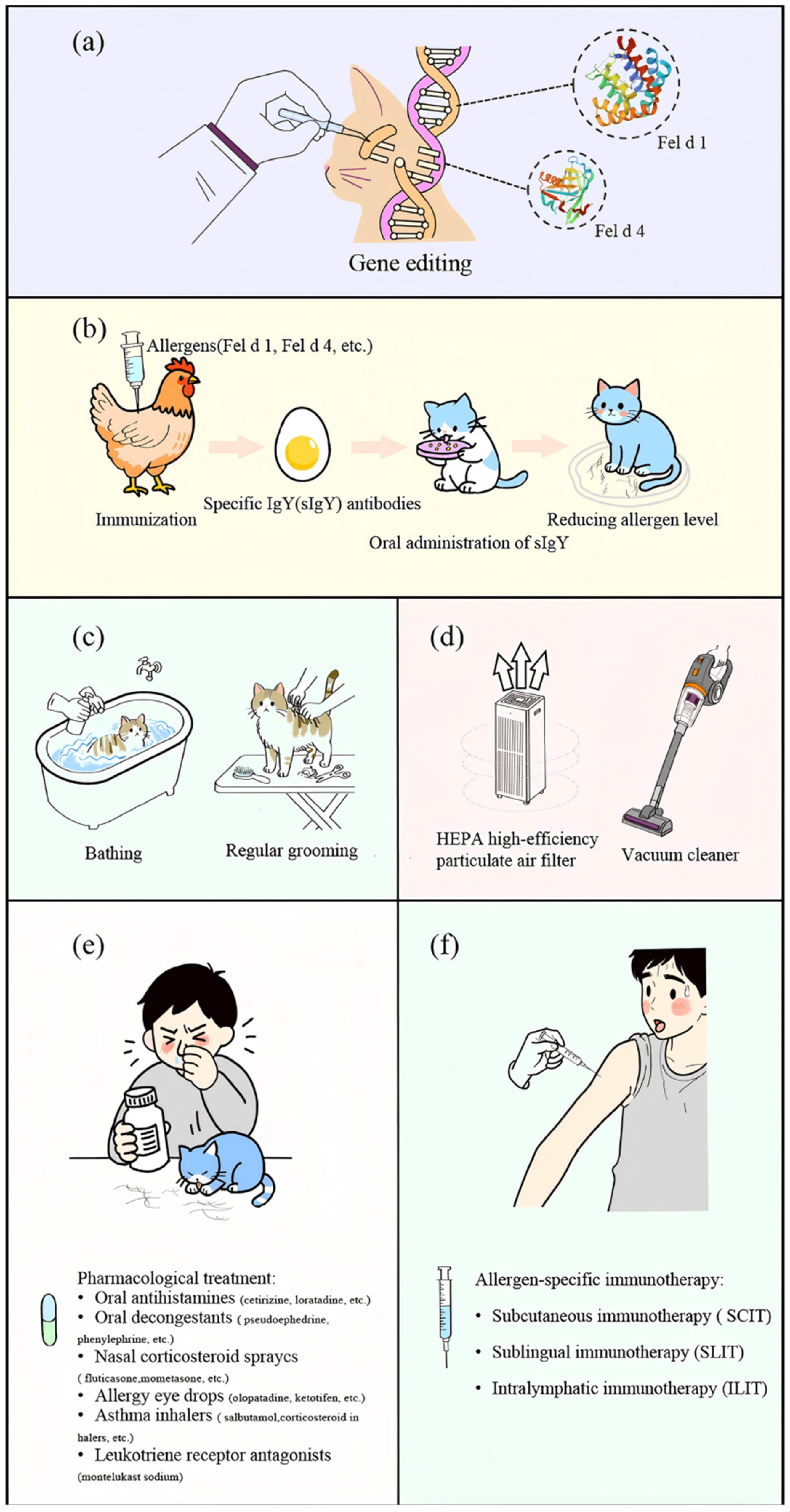
To achieve long-term allergen reduction, gene editing assays are being developed to generate hypoallergenic cat breeds [118]. Using CRISPR-Cas9, exons CH1 and CH2 was deleted in CRFK cells with an efficiency of 55% [119,120]. Lee et al. generated CH2⁻/⁻ kittens (“Alsik”) via cytoplasmic micro-injection, resulting in a 98.6% reduction in salivary and coat Fel d 1 levels, a trait that was stably maintained through cloning [121]. For immunotherapy, Zhu et al. engineered a human–cat Fcγ–Fel d 1 fusion protein that inhibited IgE-mediated histamine release by over 90% and alleviated airway responses in hFcεRIα mice [122]. Although gene editing presents a promising definitive strategy, potential adverse effects in subsequent generations require further investigation (Figure 2a).
5.1.2. Immunological Methods
Immunoglobulin Y (IgY), a polyclonal antibody produced by egg-laying animals, plays a key role in passive immunity through its ability to specifically bind and neutralize antigens. Following immunization of poultry with specific antigens, the resulting antibodies are transferred from the bloodstream to the ovary and accumulate within the egg yolk [123]. Due to phylogenetic divergence, IgY exhibits high affinity for mammalian proteins [124,125]. Satyaraj et al. demonstrated that chicken-derived IgY specifically targeting Fel d 1 to effectively inhibits its interaction with IgE [126]. In cat feeding experiments, oral administration of IgY antibodies led to a 20%–47% reduction in salivary Fel d 1 levels [127,128]. Long-term high-dose supplementation over 26 weeks resulted in no adverse changes in body weight, blood biochemical parameters, or toxicological indicators [129]. Under controlled conditions, allergic patients also showed significant improvement in symptoms [130]. Although this method is considered economical, environmentally friendly, and safe, further animal experiments and clinical evidence are required to confirm its efficacy (Figure 2b).
Another strategy involves active immunization of cats. For example, subcutaneous injection of a Fel d 1-CuMV virus-like particle vaccine elicits high titers of neutralizing IgG antibodies in immunized cats [131]. Post-vaccination of cats, serum antibody levels increased 300-fold, and the time to develop allergic symptoms in sensitized individuals was delayed 2-fold [132]. In addition, β-lactoglobulin (BLG), a whey milk protein, has been shown to regulate immune responses through the aryl hydrocarbon receptor (AHR) pathway and promote immune cell tolerance [133,134]. Clinical trials indicate that three month use of lozenges containing holographic BLG micronutrients enhances nasal tolerance to cat allergens in patients [135,136], suggesting that BLG-based intervention strategies may hold promise for preventing allergies not only to cats but also to other furry animals.
5.1.3. Physical Methods
Removing the cat from the household is an effective way to reduce allergen exposure. Studies indicate that indoor airborne Fel d 1 levels decrease by several hundred-fold within 9 to 43 weeks after the removal of the animal [137]. Treatment with sodium hypochlorite solution has been shown to disrupt the structure of recombinant Fel d 1 protein, eliminate its antibody-binding capacity, and inhibit histamine release. However, its practical application is limited due to interference from contaminating proteins, residual toxicity, and corrosive properties [138]. Bathing cats for three minutes using warm water and pet-specific shampoo can reduce airborne allergen levels by 40% to 79% [139,140], thought the effect is transient—allergen levels rebound within 3 h and return to baseline within 24 h. Regular grooming, such as brushing fur and trimming nails, reduces shedding and dander from your cat (Figure 2c). Moreover, frequent bathing is often impractical for both feline welfare and owner compliance.
5.2. Environmental Interventions
Air filtration can markedly reduce indoor levels of cat allergens (Figure 2 (d) [141]. Following the use of an air purifier, the Fel d 1 concentration decreased by 76% [142]. However, measurements from nasal-samplers indicated a 3- to 5-fold increase in the amount of cat allergen inhaled by occupants. High-efficiency particulate air (HEPA)-filtered vacuum cleaners (VCs) demonstrate a more substantial effect on controlling cat allergen levels [113], although the vacuuming process temporarily resuspends allergens into the air. In a murine model, photo-electrochemical oxidation (PECO) technology effectively degraded Fel d 1 peptides and improved markers of allergic airway inflammation. Nevertheless, this approach remains confined to animal studies and requires further validation [143].
5.3. Personal Protection and Medical Intervention
5.3.1. Chemical Drug Therapy
Chemical drug therapy serves as the cornerstone of the symptomatic management for allergic diseases, utilizing various classes of pharmaceuticals that target different pathways of the allergic response (Figure 2e). First-line treatments commonly involve oral second-generation antihistamines, such as cetirizine and loratadine, which alleviate symptoms including rhinorrhea, and pruritus by antagonizing histamine release [144,145]. For nasal congestion, oral decongestants (e.g., pseudoephedrine and phenylephrine) induce vasoconstriction to provide symptomatic relief [146,147]. Intranasal corticosteroid sprays (such as fluticasone[148] and mometasone[149]) are widely recognized as the most effective treatment for allergic rhinitis due to their potent anti-inflammatory effects [150]. Ocular symptoms are managed with topical agents like olopatadine [151] and ketotifen [152]. In allergic asthma, disease management depends on inhaled corticosteroids for sustained control and short-acting beta-agonists [153] (e.g., salbutamol [154]) for acute symptom relief. Furthermore, leukotriene receptor antagonists (e.g., montelukast [155,156]) provide an alternative oral maintenance option by inhibiting inflammatory mediators derived from arachidonic acid metabolism [157,158].
5.3.2. Immunotherapy
Allergen-specific immunotherapy (ASIT) represents an effective treatment for IgE-mediated allergies, including subcutaneous immunotherapy (SCIT), sublingual immunotherapy (SLIT), and intralymphatic immunotherapy (ILIT) (Figure 2f). Through repeated administration, ASIT promotes antigen-specific immune tolerance [159,160]. The monoclonal antibody regimen REGN1908/1909 demonstrated significant symptom relief and durable efficacy in a phase Ib clinical trial [161]. Omalizumab at dose of 150–300 mg showed significant clinical benefits and safety in Chinese adult patients with anti-histamine refractory chronic spontaneous urticaria [162]. Another candidate vaccine, PreS-Cat 1-5, is a recombinant fusion protein that uses hepatitis B virus PreS as a carrier for hypoallergenic peptides derived from Fel d 1, Fel d 4 and Fel d 7. This construct elicits potent IgE-blocking antibodies and has shown a favorable safety profile [163]. Nevertheless, ASIT is associated with certain risks: 41% of cat-allergic patients receiving subcutaneous cat allergen extracts experienced severe systemic reactions [164]. Thus, developing novel antigen formulations with high immunogenicity and reduce allergenic toxicity remains crucial for enhancing the safety of ASIT. Additionally, the potential protective effects of early-life pet exposure against asthma-like symptoms appears to be closely linked to family (especially maternal) history of asthma [165,166].
6. Prospects for Future Managing Indoor Cat Allergens
Currently, artificial intelligence (AI) is revolutionizing healthcare by enhancing diagnostics, treatment personalization, and operational efficiency [167]. The integration of immunoinformatics and AI is fundamentally reshaping the development of strategies to prevent and treat human allergy to cat allergens [168,169]. By harnessing advanced computational power to analyze complex biological data—including allergen structures, T-cell and B-cell epitopes, and immune signaling pathways—these technologies enable the precise identification of key therapeutic targets. AI-driven algorithms can predict allergenic epitopes with high accuracy and design hypoallergenic derivatives or multi-epitope vaccines that minimize IgE reactivity while promoting protective immune responses [170]. Furthermore, machine learning models can analyze large-scale clinical and omics datasets to uncover novel biomarkers, stratify patient populations, and guide the creation of personalized immunotherapy regimens [170]. As these in silico predictions are progressively validated through experimental and clinical trials, the synergy of immunoinformatics and AI promises to accelerate the translation of foundational research into safe, effective, and targeted interventions, shifting the paradigm from symptomatic management toward curative immune tolerance.
7. Conclusions
This review summarizes the molecular characteristics of major cat allergens, along with current approaches for their detection and management in indoor cat allergens. It highlights that key factors such as cat-related characteristics, environmental conditions, and spatiotemporal dynamics significantly regulate allergen expression levels. Allergens bind to air particles, enabling prolonged airborne suspension and human-mediated dispersal to cat-free settings environments. Detection relies on conventional immunoassays (e.g., ELISA) for sensitivity, advanced molecular methods (e.g. qPCR) for multiple profiling, and real-time sensors (e.g., QCM, SAW) for dynamic monitoring. Control strategies include source-directed approaches (e.g. genetic editing, vaccination), environmental measures (e.g. HEPA filtration), and medical treatments, though challenges remain in safety, efficacy, and practicality. Effective management requires integrating precise detection, environmental control, and personalized treatment, with future efforts directed toward AI-based immunomodulatory therapies.
Author Contributions
Literature search and Writing—original draft preparation, JYX, and TXY; writing—review and editing, WXM and WZL; funding acquisition, WJQ. All authors have read and agreed to the published version of the manuscript.;
Funding
This work has received funding support from the Science and Technology Innovation Engineering Fund of Institute of Feed Research of Chinese Academy of Agricultural Sciences (CAAS-ASTIP-2023-IFR-14).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
WHO, World Health Organization
IUIS, International Union of Immunological Societies
CRD, component-resolved diagnosis
IL-4, interleukin-4
IL-5, interleukin-5
IL-9, interleukin-9
IL-13, interleukin-13
ELISA, the enzyme-linked immunosorbent assay
CARD, catalytic reported deposition
qPCR, quantitative PCR
EDC, electrostatic dust collectors
HPLC-MS/MS, high-performance liquid chromatography-tandem mass spectrometry
QCM, Quartz Crystal Microbalance
SAW, Surface Acoustic Wave
LPC, Laser Particle Counters
OPC, Optical Particle Counters
IoT, Internet of Things
VC, Vacuum cleaners
HEPA, high-efficiency particulate air
IgY, Immunoglobulin Y
BLG, β-lactoglobulin
AHR, aryl hydrocarbon receptor
HEPA, High-efficiency particulate air
PECO, photo-electrochemical oxidation
ASIT, allergen-specific immunotherapy
SCIT, encompassing subcutaneous immunotherapy
SLIT, sublingual immunotherapy
ILIT, intralymphatic immunotherapy
AI, Artificial intelligence
References
- Dramburg S, Hilger C, Santos A F, et al. EAACI molecular allergology user's guide 2.0. Egypt. J. Pediatr. Alle. 2023, 34, e13854.
- Holgate S T, Polosa R. Treatment strategies for allergy and asthma. Nat. Rev. Immunol. 2008, 8, 218-230.
- Kay A B. Allergy and allergic diseases. N. Engl. J. Med. 2001, 344, 30-37.
- Asher M I, Montefort S,Björkstén B, et al. Worldwide time trends in the prevalence of symptoms of asthma, allergic rhinoconjunctivitis, and eczema in childhood: ISAAC phases one and three repeat multicountry cross-sectional surveys. Lancet 2006, 368, 733-743.
- Park H J, Kim E, Yoon D, et al. Prevalence of self-reported allergic diseases and IgE levels: a 2010 KNHANES analysis. Allergy Asthma Immun. 2017, 9, 329.
- Edwards-Salmon S E, Padmanabhan S L, Kuruvilla M, et al. Increasing prevalence of allergic disease and its impact on current practice. Curr. Otorhinol. Rep. 2022, 10, 278-284.
- Merhej T, Zein J G. Epidemiology of asthma: prevalence and burden of disease. Adv. Exp. Med. Biol. 2023, 1426, 3-23.
- Oh J, Kim S, Kim M S, et al. Global, regional, and national burden of asthma and atopic dermatitis, 1990--2021, and projections to 2050: a systematic analysis of the global burden of disease study 2021. Lancet Resp. Med. 2025, 13, 425-446.
- Shin Y H, Hwang J, Kwon R, et al. Global, regional, and national burden of allergic disorders and their risk factors in 204 countries and territories, from 1990 to 2019: a systematic analysis for the global burden of disease study 2019. Allergy 2023, 78, 2232-2254.
- Melén E, Garcia-Aymerich J. Predicting the future global burden of asthma and atopic dermatitis: identifying strategies for prevention. Lancet Resp. Med. 2025, 13, 376-378.
- Takkouche B, González-Barcala F-J, Etminan M, et al. Exposure to furry pets and the risk of asthma and allergic rhinitis: a meta-analysis. Allergy 2008, 63, 857-864.
- Chan S K, Leung D Y. Dog and cat allergies: current state of diagnostic approaches and challenges. Allergy Asthma Immun. 2018, 10, 97.
- Yin W, Xiaoli Z, Wenjin D, et al. Sensitization profiles of aeroallergens among allergic rhinitis patients in China: a 13-year multicenter retrospective study. Allergy 2023, 79, 1329-1332.
- Schoos A M, Nwaru B I, Borres M P. Component-resolved diagnostics in pet allergy: current perspectives and future directions. J. Allergy Clin. Immun. 2021, 147, 1164-1173.
- Liang H, Ouyang Z, Liu J, et al. The development trend analysis of pet industry, 2025. IEEE.
- Shen R, Shen F, Zhao Y. Pet industry analysis based on multiple linear regression and aeima models, 2025. IEEE.
- Zahradnik E, Raulf M. Animal allergens and their presence in the environment. Front. Immunol. 2014, 5, 76.
- Konradsen J R, Fujisawa T, Van Hage M, et al. Allergy to furry animals: new insights, diagnostic approaches, and challenges. Allergy Clin. Immun. 2015, 135, 616-625.
- Arbes Jr S J, Cohn R D, Yin M, et al. Dog allergen (Can f 1) and cat allergen (Fel d 1) in US homes: Results from the national survey of lead and allergens in housing. Ann. Allerg. Asthma Im. 2008, 101, 517.
- An W, Li T, Tian X, et al. Allergies to allergens from cats and dogs: a review and update on sources, pathogenesis, and strategies. Int. J. Mol. Sci. 2024, 25, 10520.
- Lei D K, Grammer L C. An overview of allergens. Allergy Asthma Proc. 2019, 40, 362-365.
- Chapman M D, Wood R A. The role and remediation of animal allergens in allergic diseases. Allergy Clin. Immun. 2001, 107, S414-S421.
- Nadeau K C. Allergen-specific IgG antibodies for cat allergy? Ann. Am. Thorac. Soc. 2021, 204, 1-2.
- Brożek J L, Bousquet J, Agache I, et al. Allergic rhinitis and its impact on asthma (ARIA) guidelines—2016 revision. Allergy Clin. Immun. 2017, 140, 950-958.
- Dávila I, Dom I Nguez-Ortega J, Navarro-Pulido A, et al. Consensus document on dog and cat allergy. Allergy 2018, 73, 1206-1222.
- Sahiner U M, Giovannini M, Escribese M M, et al. Mechanisms of allergen immunotherapy and potential biomarkers for clinical evaluation. J. Pers. Medi. 2023, 13, 845.
- Ohman Jr J L, Lowell F C, Bloch K J. Allergens of mammalian origin: III. Properties of a major feline allergen. J. Immunol. 1974, 113, 1668-1677.
- Klug J, Beier H M, Bernard A, et al. Uteroglobin/Clara cell 10-kDa family of proteins: nomenclature committee report. Ann. N. Y. Acad. Sci. 2000, 923, 348-354.
- Kaiser L, Grönlund H, Sandalova T, et al. The crystal structure of the major cat allergen Fel d 1, a member of the secretoglobin family. J. Biol. Chem. 2003, 278, 37730-37735.
- Duffort O, Carreira J E, Lombardero M. Monoclonal antibodies against Fel d I and other clinically relevant cat allergens. Immunol. Let. 1988, 17, 71-77.
- Kaiser L, Velickovic T C, Badia-Martinez D, et al. Structural characterization of the tetrameric form of the major cat allergen Fel d 1. J. Mol. Biol. 2007, 370, 714-727.
- Karn R C. The mouse salivary androgen-binding protein (ABP) alpha subunit closely resembles chain 1 of the cat allergen Fel dI. Biochem. Genet. 1994, 32, 271-277.
- Bonnet B, Messaoudi K, Jacomet F, et al. An update on molecular cat allergens: Fel d 1 and what else? Chapter 1: Fel d 1, the major cat allergen. Allergy Asthma Cl. Im. 2018, 14, 14.
- van Ree R, van Leeuwen W A, Bulder I, et al. Purified natural and recombinant Fel d 1 and cat albumin in in vitro diagnostics for cat allergy. J. Allergy Clin. Imm. 1999, 104, 1223-1230.
- Cleveland III C W, Davis B W, Khatri K, et al. Genetic diversity of the major cat allergen, Fel d 1. PNAS Nexus 2024, 3, pgae447.
- Thomas W R. The advent of recombinant allergens and allergen cloning. J. Allergy Clin. Imm. 2011, 127, 855-859.
- Charpin C, Mata P, Charpin D, et al. Fel d I allergen distribution in cat fur and skin. J. Allergy Clin. Imm. 1991, 88, 77-82.
- Dabrowski A J, Van der Brempt X, Soler M, et al. Cat skin as an important source of Fel d I allergen. J. Allergy Clin. Imm. 1990, 86, 462-465.
- Bartholome K, Kissler W, Baer H, et al. Where does cat allergen 1 come from? J. Allergy Clin. Imm. 1985, 76, 503-506.
- Brown P R, Leitermann K, Ohman Jr J L. Distribution of cat allergen 1 in cat tissues and fluids. Int. Arch. Allergy Imm. 1984, 74, 67-70.
- De Andrade A D, Birnbaum J, Magalon C, et al. Fel d I levels in cat anal glands. Clin. Exp. Allergy 1996, 26, 178-180.
- Anderson M C, Baer H, Ohman Jr J L. A comparative study of the allergens of cat urine, serum, saliva, and pelt. J. Allergy Clin Imm. 1985, 76, 563-569.
- Siebers R, Healy B, Holt S, et al. Fel d 1 levels in domestic living rooms are not related to cat color or hair length. 2001.
- Jalil-Colome J E, de Andrade A E L D, Birnbaum J E L, et al. Sex difference in Fel d 1 allergen production. J. Allergy Clin Imm. 1996, 98, 165-168.
- Zielonka T M, Charpin D, Berbis P H, et al. Effects of castration and testosterone on Fel d I production by sebaceous glands of male cats: I—immunological assessment. Revue Francaise D'allergologie et D'immunologie Clinique, 1995, 35, 359-363.
- Carayol N, Birnbaum J, Magnan A, et al. Fel d 1 production in the cat skin varies according to anatomical sites. Allergy 2000, 55, 570-573.
- Trifonova D, Curin M, Riabova K, et al. Allergenic activity of individual cat allergen molecules. Int. J. Mol. Sci. 2023, 24, 16729.
- Popescu F, Ganea C S, Panaitescu C, et al. Molecular diagnosis in cat allergy. World J. Methodol. 2021, 11, 46-60.
- Ichikawa K, Vailes L D, Pomes A, et al. Molecular cloning, expression and modelling of cat allergen, cystatin (Fel d 3), a cysteine protease inhibitor. Clin. Exp. Allergy 2001, 31, 1279-1286.
- Smith W, Butler A, Hazell L A, et al. Fel d 4, a cat lipocalin allergen. Clin. Exp. Allergy 2004, 34, 1732-1738.
- Smith W, O Neil S E, Hales B J, et al. Two newly identified cat allergens: the von Ebner gland protein Fel d 7 and the latherin-like protein Fel d 8. Int. Arch. Allergy Imm. 2011, 156, 159-170.
- Riabova K, Karsonova A V, van Hage M, et al. Molecular allergen-specific IgE recognition profiles and cumulative specific IgE levels associated with phenotypes of cat allergy. Int. J. Mol. Sci. 2022, 23, 6984.
- Caraballo L, Valenta R, Puerta L, et al. The allergenic activity and clinical impact of individual IgE-antibody binding molecules from indoor allergen sources. World Allergy Organ. 2020, 13, 100118.
- Lu J, Zhu H, Yang Q, et al. Associations of protein classes with cross-reactivity and cross-sensitization in furry animal allergens: A component-resolved diagnostics study. J. Asthma Allergy 2025, 363-375.
- Saarelainen S, Rytk O Nen-Nissinen M, Rouvinen J, et al. Animal-derived lipocalin allergens exhibit immunoglobulin E cross-reactivity. Clin. Exp. Allergy 2008, 38, 374-381.
- Popescu F, Vieru M. Precision medicine allergy immunoassay methods for assessing immunoglobulin E sensitization to aeroallergen molecules. World J. Methodol. 2018, 8, 17-36.
- Hilger C, Swiontek K, Arumugam K, et al. Identification of a new major dog allergen highly cross-reactive with Fel d 4 in a population of cat-and dog-sensitized patients. J. Allergy Clin. Immun. 2012, 129, 1149-1151.
- Yamamoto K, Ishibashi O, Sugiura K, et al. Crystal structure of the dog allergen Can f 6 and structure-based implications of its cross-reactivity with the cat allergen Fel d 4. Sci. Rep. 2019, 9, 1503.
- Liu Z, Trifonova D, Tulaeva I, et al. Albumins represent highly cross-reactive animal allergens. Front. Immunol. 2023, 14, 1241518.
- Luczynska C M, Li Y, Chapman M D, et al. Airborne concentrations and particle size distribution of allergen derived from domestic cats (Felis domesticus). Am. Rev. Respir. Dis. 1990, 141, 361-367.
- Wood R A, Laheri A N, Eggleston P A. The aerodynamic characteristics of cat allergen. Clin. Exp. Allergy 1993, 23, 733-739.
- Swanson M C, Agarwal M K, Reed C E. An immunochemical approach to indoor aeroallergen quantitation with a new volumetric air sampler: studies with mite, roach, cat, mouse, and guinea pig antigens. J. Allergy Clin. Immun. 1985, 76, 724-729.
- De Blay F, Heymann P W, Chapman M D, et al. Airborne dust mite allergens: comparison of group II allergens with group I mite allergen and cat-allergen Fel d I. J. Allergy Clin. Immun. 1991, 88, 919-926.
- Wood R A, Eggleston P A, Lind P, et al. Antigenic analysis of household dust samples1-4. Am. Rev. Respir. Dis. 1988, 137, 358-363.
- Dallongeville A, Le Cann P, Zmirou-Navier D, et al. Concentration and determinants of molds and allergens in indoor air and house dust of French dwelling. Sci. Total Environ. 2015, 536, 964-972.
- Ritz B R, Hoelscher B, Frye C, et al. Allergic sensitization owing to ‘second-hand’cat exposure in schools. Allergy 2002, 57, 357-361.
- Custovic A, Green R, Taggart S, et al. Domestic allergens in public places II: dog (Can f 1) and cockroach (Bla g 2) allergens in dust and mite, cat, dog and cockroach allergens in the air in public buildings. Clin. Exp. Allergy 1996, 26, 1246-1252.
- Partti-Pellinen K, Marttila O, M A Kinen-Kiljunen S, et al. Occurrence of dog, cat, and mite allergens in public transport vehicles. Allergy 2000, 55, 65-68.
- Enberg R N, Shamie S M, McCullough J, et al. Ubiquitous presence of cat allergen in cat-free buildings: probable dispersal from human clothing. Ann. Allergy 1993, 70, 471-474.
- De Lucca S D, O Meara T J, Tovey E R. Exposure to mite and cat allergens on a range of clothing items at home and the transfer of cat allergen in the workplace. J. Allergy Clin. Immun. 2000, 106, 874-879.
- Karlsson A, Renstr O M A. Human hair is a potential source of cat allergen contamination of ambient air. Allergy 2005, 60, 961-964.
- Karlsson A, Renstr O M A, Hedren M, et al. Allergen avoidance does not alter airborne cat allergen levels in classrooms. Allergy, 2004, 59, 661-667.
- van Hage M, Käck U, Asarnoj A, et al. An update on the prevalence and diagnosis of cat and dog allergy--emphasizing the role of molecular allergy diagnostics. Mol. Immunol. 2023, 157, 1-7.
- Maeda Y, Akiyama K. Anaphylaxis after a cat bite. Allergol. Int. 2012, 61, 511-512.
- Leung D Y, Berdyshev E, Goleva E. Cutaneous barrier dysfunction in allergic diseases. J. Allergy Clin. Immun. 2020, 145, 1485-1497.
- Reinero C R. Advances in the understanding of pathogenesis, and diagnostics and therapeutics for feline allergic asthma. Vet. J. 2011, 190, 28-33.
- Platts-Mills T A, Woodfolk J A, Erwin E A, et al. Mechanisms of tolerance to inhalant allergens: the relevance of a modified Th2 response to allergens from domestic animals. Springer Semin. Immunopathol. 2004, 25, 271-279.
- Leo G, Incorvaia C, Arasi S. Could a bite trigger the onset of cat allergy? Pediat. Aller. Immunol. 2022, 33, e13841.
- Jones M. Understanding of the molecular mechanisms of allergy. Methods Mol. Med. 2008, 138, 1-15.
- Han X, Krempski J W, Nadeau K. Advances and novel developments in mechanisms of allergic inflammation. Allergy 2020,75(12),3100-3111.
- Shamji M H, Valenta R, Jardetzky T, et al. The role of allergen-specific IgE, IgG and IgA in allergic disease. Allergy 2021,76(12),3627-3641.
- Nwaru B I, Suzuki S, Ekerljung L, et al. Furry animal allergen component sensitization and clinical outcomes in adult asthma and rhinitis. J. Aller. Cl. Imm-Pract. 2019, 7, 1230-1238.
- Özuygur Ermis S S, Norouzi A, Borres M P, et al. Sensitization patterns to cat molecular allergens in subjects with allergic sensitization to cat dander. Clin. Transl. Allergy, 2023, 13, e12294.
- Bollinger M E, Wood R A, Chen P, et al. Measurement of cat allergen levels in the home by use of an amplified ELISA. J. Allergy Clin. Immun. 1998, 101, 124-125.
- Tasaniyananda N, Tungtrongchitr A, Seesuay W, et al. Quantification of Fel d 1 in house dust samples of cat allergic patients by using monoclonal antibody specific to a novel IgE-binding epitope. Asian Pac. J. Allergy, 2018, 36, 8-15.
- Lau S, Schulz G, Sommerfeld C, et al. Comparison of quantitative ELISA and semiquantitative dustscreen™ for determination of Der p 1, Der f 1, and Fel d 1 in domestic dust samples. Allergy, 2001,56,993-995.
- Kelly S M, Karsh J, Marcelo J, et al. Fel d 1 and Fel d 4 levels in cat fur, saliva, and urine. J. Allergy Clin. Immun. 2018, 142, 1990-1992.
- Huynh S, Menzies S, Khurana T, et al. Radial Immunodiffusion (RID) to sandwich ELISA for the quantitation of Fel d 1 and Amb a 1 in cat and short ragweed pollen allergenic extracts. J. Allergy Clin. Immun. 2012, 129, AB88.
- Rabin R L, Croote D, Chen A, et al. A human monoclonal antibody based immunoenzymetric assay to measure Fel d 1 concentrations in cat hair and pelt allergenic extracts. Frontiers in Allergy, 2024,5,1417879.
- van Ree F. Analytic aspects of the standardization of allergenic extracts. Allergy, 1997,52.
- King E M, Filep S, Smith B, et al. A multi-center ring trial of allergen analysis using fluorescent multiplex array technology. J. Immunol. Methods, 2013, 387, 89-95.
- Filep S C, Black K R, Smith B R, et al. Simultaneous quantification of specific food allergen proteins using a fluorescent multiplex array. Food Chem. 2022, 389, 132986.
- Earle C D, King E M, Tsay A, et al. High-throughput fluorescent multiplex array for indoor allergen exposure assessment. J. Allergy Clin Immun. 2007, 119, 428-433.
- Earle C D, King E M, Tsay A, et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clinical Chemistry, 2009,55,4611-4622.
- Scherer E, Valot B, Vacheyrou M, et al. Assessment of pets (cats and dogs) in homes using electrostatic dust collectors and QPCR: new tools to evaluate exposure and risk of allergies. Int. J. Environ. Heal. R. 2016, 26, 589-599.
- Bustin S A, Benes V, Garson J A, et al. Unbiased quantitative proteomics reveals a crucial role of the allergen context for the activation of human dendritic cells. Sci. Rep-UK. 2017, 7, 16638.
- Krutz N L, Kimber I, Winget J, et al. Identification and semi-quantification of protein allergens in complex mixtures using proteomic and AllerCatPro 2.0 bioinformatic analyses: a proof-of-concept investigation. J. Immunotoxicol. 2024, 21, 2305452.
- López-Pedrouso M, Lorenzo J M, Dios Alché J D, et al. Advanced proteomic and bioinformatic tools for predictive analysis of allergens in novel foods. Biology, 2023, 12, 714.
- Wéry N, Galès A, Brunet Y. Bioaerosol sources. Microbiology of Aerosols, 2017,115-135.
- Haig C W, Mackay W G, Walker J T, et al. Bioaerosol sampling: sampling mechanisms, bioefficiency and field studies. J. Hosp. Infect. 2016, 93, 242.
- Kabir E, Azzouz A, Raza N, et al. Recent advances in monitoring, sampling, and sensing techniques for bioaerosols in the atmosphere. ACS sensors, 2020, 5, 1254-1267.
- Morris D R, Fatisson J, Olsson A L, et al. Real-time monitoring of airborne cat allergen using a QCM-based immunosensor. Sensor. Actuat. B-Chem. 2014, 190, 851-857.
- Toma K, Horibe M, Kishikawa C, et al. Rapid and repetitive immunoassay with a surface acoustic wave device for monitoring of dust mite allergens. Sensor. Actuat. B-Chem. 2017, 248, 924-929.
- Hossenbaccus L, Walker T, Ellis A K. Technical validation of controlled exposure to cat dander in the specialized particulate control environmental exposure unit (SPaC-EEU). Allergy Asthma Cl. Im. 2025, 21, 6.
- Salimifard P, Rim D, Freihaut J D. Evaluation of low-cost optical particle counters for monitoring individual indoor aerosol sources. Aerosol Sci. Tech. 2020, 54, 217-231.
- Fong A, Fong B. Indoor air quality control for asthma patients using smart home technology, 2011. IEEE.
- Bastien B C, Gardner C, Satyaraj E. Influence of time and phenotype on salivary Fel d1 in domestic shorthair cats. J. Feline Med. Surg. 2019, 21, 867-874.
- Bienboire-Frosini C E C, Lebrun R E G, Vervloet D, et al. Distribution of core fragments from the major cat allergen Fel d 1 is maintained among the main anatomical sites of production. Int. Arch. Allergy Imm 2010, 152, 197-206.
- Peterson E L, Ownby D R, Johnson C C. The relationship of housing and household characteristics to the indoor concentrations of Der f 1, Der p 1, and Fel d 1 measured in dust and air samples. Ann. Allergy Asthma Im. 2003, 90, 564-571.
- Neal J S, Arlian L G, Morgan M S. Relationship among house-dust mites, Der 1, Fel d 1, and Can f 1 on clothing and automobile seats with respect to densities in houses. Ann. Allergy Asthma Im. 2002, 88, 410-415.
- Montoya L D, Hildemann L M. Evolution of the mass distribution of resuspended cat allergen (Fel d 1) indoors following a disturbance. Atmos. Environ. 2001, 35, 859-866.
- Corsi R L, Siegel J A, Chiang C. Particle resuspension during the use of vacuum cleaners on residential carpet. J. Occup. Environ. Hyg. 2008, 5, 232-238.
- De Blay F, Spirlet F, Gries P, et al. Effects of various vacuum cleaners on the airborne content of major cat allergen (Fel d 1). Allergy, 1998, 53, 411-414.
- Hegarty J M, Rouhbakhsh S, Warner J A, et al. A comparison of the effect of conventional and filter vacuum cleaners on airborne house dust mite allergen. Resp. Med. 1995, 89, 279-284.
- Chew G L, Higgins K M, Gold D R, et al. Monthly measurements of indoor allergens and the influence of housing type in a northeastern US city. Allergy, 1999, 54, 1058-1066.
- Chew G L, Saha S. Impacts of climate change on indoor allergens. Impacts of climate change on allergens and allergic diseases. Cambridge University Press Cambridge, 2016,119.
- Egmar A, Almqvist C, Emenius G, et al. Deposition of cat (Fel d 1), dog (Can f 1), and horse allergen over time in public environments--a model of dispersion. Allergy, 1998, 53, 957-961.
- Lee S R, Lee K, Song S, et al. Generation and cloning of CH2 knockout cats by CRISPR-Cas9 system: Hypoallergenic cats. Authorea Preprints, 2023.
- Brackett N F, Davis B W, Adli M, et al. Evolutionary biology and gene editing of cat allergen, Fel d 1. CRISPR J. 2022, 5, 213-223.
- Brackett N, Riedy J, Adli M, et al. CRISPR gene editing of the major cat allergen, Fel d 1. Allergy, 2020,75,59-60.
- Lee S R, Lee K, Song S, et al. Generation of Fel d 1 chain 2 genome-edited cats by CRISPR-Cas9 system. Scientific Rep-UK. 2024, 14, 4987.
- Zhu D, Kepley C L, Zhang K, et al. A chimeric human-cat fusion protein blocks cat-induced allergy. Nat. Med. 2005, 11, 446-449.
- Wang H, Zhong Q, Lin J. Egg yolk antibody for passive immunization: status, challenges, and prospects. J. Agr. Food Chem. 2023, 71, 5053-5061.
- Zhang X, Calvert R A, Sutton B J, et al. IgY: a key isotype in antibody evolution. Biol. Rev. 2017, 92, 2144-2156.
- Kovacs-Nolan J, Mine Y. Egg yolk antibodies for passive immunity. Annu. Rev. Food Sci. T. 2012, 3, 163-182.
- Satyaraj E, Li Q, Sun P, et al. Anti-Fel d1 immunoglobulin Y antibody-containing egg ingredient lowers allergen levels in cat saliva. J. Feline Med. Surg. 2019, 21, 875-881.
- Satyaraj E, Wedner H J, Bousquet J. Keep the cat, change the care pathway: a transformational approach to managing Fel d 1, the major cat allergen. Allergy, 2019, 74, 5-17.
- Matulka R A, Thompson L, Corley D. Multi-level safety studies of anti Fel d 1 IgY ingredient in cat food. Front. Vet. Sci. 2020, 6, 477.
- Hedrick E D, Matulka R A, Conboy-Schmidt L, et al. Evaluation of anti-Fel d 1 IgY ingredient for pet food on growth performance in kittens. Front. Vet. Sci. 2024, 11, 1355390.
- Satyaraj E, Gardner C, Filipi I, et al. Reduction of active Fel d1 from cats using an antiFel d1 egg IgY antibody. Immun. Inflamm. Dis. 2019, 7, 68-73.
- Thoms F, Jennings G T, Maudrich M, et al. Immunization of cats to induce neutralizing antibodies against Fel d 1, the major feline allergen in human subjects. J. Allergy Clin. Immun. 2019, 144, 193-203.
- Thoms F, Haas S, Erhart A, et al. Immunization of cats against Fel d 1 results in reduced allergic symptoms of owners. Viruses, 2020, 12, 288.
- Bergmann K, Graessel A, Raab J, et al. Targeted micronutrition via holo-BLG based on the farm effect in house dust mite allergic rhinoconjunctivitis patients—first evaluation in a standardized allergen exposure chamber. Allergo Journal International, 2021,30(4),141-149.
- Bergmann K, Raab J, Krause L, et al. Long-term benefits of targeted micronutrition with the holoBLG lozenge in house dust mite allergic patients. Allergo Journal International, 2022, 31, 161-171.
- Jensen-Jarolim E, Jensen S A, Bergmann K. Allergy to the cat—from diagnosis to management. Allergo Journal International, 2023,32(5),130-137.
- Bergmann K, Raab J, Graessel A, et al. The holo beta-lactoglobulin lozenge reduces symptoms in cat allergy—evaluation in an allergen exposure chamber and by titrated nasal allergen challenge. Clin. Transl. Allergy. 2023, 13, e12274.
- Wood R A, Chapman M D, Adkinson Jr N F, et al. The effect of cat removal on allergen content in household-dust samples. J. Allergy Clin. Immun. 1989, 83, 730-734.
- Matsui E, Kagey-Sobotka A, Chichester K, et al. Allergic potency of recombinant Fel d 1 is reduced by low concentrations of chlorine bleach. J. Allergy Clin. Immun. 2003, 111, 396-401.
- Avner D B, Perzanowski M S, Platts-Mills T A, et al. Evaluation of different techniques for washing cats: quantitation of allergen removed from the cat and the effect on airborne Fel d 1. J. Allergy Clin. Immun. 1997, 100, 307-312.
- Nageotte C, Park M, Havstad S, et al. Duration of airborne Fel d 1 reduction after cat washing. J. Allergy Clin. Immun. 2006, 118, 521-522.
- Maya-Manzano J E M I, Pusch G, Ebner Von Eschenbach C, et al. Effect of air filtration on house dust mite, cat and dog allergens and particulate matter in homes. Clin. Transl. Allergy, 2022, 12, e12137.
- Gherasim A, de Blay F E D E. Does air filtration work for cat allergen exposure? Curr. Allergy Asthm. R. 2020, 20, 18.
- Devadoss D, Surbaugh K, Manevski M, et al. Indoor-air purification by photoelectrochemical oxidation mitigates allergic airway responses to aerosolized cat dander in a murine model. Sci. Rep-UK, 2023, 13, 10980.
- Golightly L K, Greos L S. Second-generation antihistamines: actions and efficacy in the management of allergic disorders. Drugs, 2005, 65, 341-384.
- Linton S, Hossenbaccus L, Ellis A K. Evidence-based use of antihistamines for treatment of allergic conditions. Ann. Allergy Asthma Im. 2023, 131, 412-420.
- Desjardins P J, Berlin R G. Efficacy of phenylephrine. Brit. J. Clin. Pharmaco. 2007, 64, 555.
- Krause H F. Antihistamines and decongestants. Otolaryngolog. Head Neck , 1992, 107, 835-840.
- Paggiaro P L, Dahle R, Bakran I, et al. Multicentre randomised placebo-controlled trial of inhaled fluticasone propionate in patients with chronic obstructive pulmonary disease. The Lancet, 1998, 351, 773-780.
- Penagos M, Compalati E, Tarantini F, et al. Efficacy of mometasone furoate nasal spray in the treatment of allergic rhinitis. Meta-analysis of randomized, double-blind, placebo-controlled, clinical trials. Allergy, 2008, 63, 1280-1291.
- Hoang M P, Chitsuthipakorn W, Seresirikachorn K, et al. As-needed intranasal corticosteroid spray for allergic rhinitis: a systematic review and meta-analysis. Rhinology, 2022, 60, 242-251.
- Abelson M B. A review of olopatadine for the treatment of ocular allergy. Expert Opin Pharmaco. 2004, 5, 1979-1994.
- Grant S M, Goa K L, Fitton A, et al. Ketotifen: a review of its pharmacodynamic and pharmacokinetic properties, and therapeutic use in asthma and allergic disorders. Drugs, 1990, 40, 412-448.
- Barnes P J, Pedersen S O R, Busse W W. Efficacy and safety of inhaled corticosteroids: new developments. Am. J. Crit. Care .1998, 157, S1-S53.
- Marques L, Vale N. Salbutamol in the management of asthma: A review. Int. J. Mol. Sci. 2022, 23, 14207.
- Nayak A, Langdon R B. Montelukast in the treatment of allergic rhinitis: an evidence-based review. Drugs, 2007, 67, 887-901.
- Noonan M J, Chervinsky P, Brandon M, et al. Montelukast, a potent leukotriene receptor antagonist, causes dose-related improvements in chronic asthma. Eu. Respir. J. 1998, 11, 1232-1239.
- Lipworth B J. Leukotriene-receptor antagonists. The Lancet, 1999, 353, 57-62.
- Montuschi P, Peters-Golden M L. Leukotriene modifiers for asthma treatment. Clin. Exp. Allergy, 2010, 40, 1732-1741.
- Arshad S H. An update on allergen immunotherapy. Clin. Med. Res. 2016, 16, 584-587.
- Adlany Y K, V S O V S I C L, Senti G, et al. Quality of life in allergic rhinitis patients treated with intralymphatic immunotherapy (ILIT): A 19-year follow-up. J. Allergy Clin. Immun. 2023, 2, 43-50.
- Orengo J M, Radin A R, Kamat V, et al. Treating cat allergy with monoclonal IgG antibodies that bind allergen and prevent IgE engagement. Nat. Commun. 2018, 9,1 421.
- Yuan W, Hu S, Li M, et al. Efficacy and safety of omalizumab in Chinese patients with anti-histamine refractory chronic spontaneous urticaria. Dermatol. Ther. 2022, 35, e15303.
- Trifonova D, Curin M, Focke-Tejkl M, et al. Recombinant hypoallergenic cat allergy vaccines. Allergy 2025, 80, 2622-2635.
- De Filippo M, Votto M, Caminiti L, et al. Safety of allergen-specific immunotherapy in children. Pediat. Allerg. Imm-UK. 2022, 33, 27-30.
- Remes S T, Castro-Rodriguez J A, Holberg C J, et al. Dog exposure in infancy decreases the subsequent risk of frequent wheeze but not of atopy. J. Allergy Clin. Immun. 2001, 108, 509-515.
- Celed O N J C, Litonjua A A, Ryan L, et al. Exposure to cat allergen, maternal history of asthma, and wheezing in first 5 years of life. The Lancet, 2002, 360, 781-782.
- Dias R, Torkamani A. Artificial intelligence in clinical and genomic diagnostics. Genome Med., 2019, 11, 70.
- MacMath D, Chen M, Khoury P. Artificial intelligence: exploring the future of innovation in allergy immunology. Curr. Allergy Asthm. R. 2023, 23, 351-362.
- Alanazi H H. Role of artificial intelligence in advancing immunology. Immunol. Res. 2025, 73, 76.
- Indolfi C, Klain A, Dinardo G, et al. Artificial intelligence in the transition of allergy: A valuable tool from childhood to adulthood. Front. Med-PRC. 2024, 11, 1469161.
Figure 2.
Control of cat allergens. (a) Gene editing. (b) Immunological methods. (c) Physical methods. (d) Environmental interventions. (e) Chemical drug therapy. (f) Immunotherapy.
Figure 2.
Control of cat allergens. (a) Gene editing. (b) Immunological methods. (c) Physical methods. (d) Environmental interventions. (e) Chemical drug therapy. (f) Immunotherapy.

| Countries | Methods | Total | Cats | Dogs |
| China | Blood testing | 16,664 | 15.47% | 10.50% |
| Russia | Blood testing | 513 | 24.10% | 21.40% |
| South Korea | Skin prick test | 7504 | 20.60% | 15.20% |
| Germany | Blood testing | 356 | 34.80% | 31.70% |
| Japan | Blood testing | 12,205,097 | 18.20% | 18.90% |
| America | Blood testing | 478 | 54.40% | 64.70% |
| Canada | Skin prick test | 623 | 53.10% | 17.30% |
| Qatar | Skin prick test | 473 | 6.18% | 0.50% |
| Lebanon | Skin prick test | 919 | 29.90% | 21.90% |
| Thailand | Skin prick test | 1516 | 12.90% | 10.00% |
| Nepal | Skin prick test | 170 | 15.30% | 14.10% |
| Mexico | Skin prick test | 761 | 26.70% | 33.90% |
Table 2.
Comparison analysis of detection methods for Fel d1 allergen in indoor environments.
| Methods | Technical Features | Application Scenarios | Sensitivity | References |
| Signal amplification ELISA | Employing catalytic reported deposition (CARD) technology for the cascade amplification of enzyme-substrate reactions | Ultra-low concentration air samples | 156 pg Fel d 1 / mL | [84] |
| Double-point sandwich ELISA | MAbC48 targets conserved IgE epitopes, avoiding interference from Fel d 1 polymorphisms | Indoor dust samples | 390 pg Fel d 1 / mL | [85] |
| Immunodot assay (Dustscreen™) | Semi-quantitative, 4h results, capable of multiple allergen testing | Rapid screening for allergens in public areas | 100 pg Fel d 1/mL | [86] |
| Double antibody sandwich ELISA | Quantified using manufacturer standard curves, between plates CV<10% | For detecting allergen content in clinical or research settings | 80,000 pg Fel d 1 | [87] |
| scFv-Sandwich ELISA | The 96-well high-throughput format replaces traditional radial diffusion, eliminating the need for subjective interpretation | Specific for the potency verification of commercially available standardized allergen extracts | 500 pg Fel d 1/mL | [88] |
| Human IgG4-tweezed sandwich ELISA | Using high-affinity humanized monoclonal antibodies, between plates CV<9% | Suitable for manufacturers calibration of cat allergen concentrations | 250 pg Fel d 1/mL | [89] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



