
Submitted:
01 October 2025
Posted:
02 October 2025
You are already at the latest version
Abstract
Most organisms are associated with microorganisms, which influence their behavior during their life cycles. Fungi are no exception; they interact with plants, viruses, and bacteria in various environments, forming complex communities. These associations can occur externally around the mycelia and internally within the hyphae. Fungi can harbor bacteria, mycoviruses, and other fungi within their hyphae. Some endofungal bacteria (EHB) have been shown to alter fungal host function, development, and interactions with other organisms. Most fungi that host EHB are plant-associated. Although members of the genus Metarhizium are among the most abundant fungi isolated from soils, their associated EHB have not been sufficiently studied. Endobacteria were recently detected in M. bibiondarum and M. anisopliae; however, the biological roles of these bacteria in the different Metarhizium life cycles remain unknown. In this study, Metarhizium strains were isolated from the rhizosphere and Phyllophaga spp. Bacillus subtilis was identified as an endobacterium, and its influence on Metarhizium’s physiology and entomopathogenic capacity was studied. Our analysis revealed that EHB have a negative impact on the virulence of Metarhizium against Galleria mellonella and Tenebrio molitor.
Keywords:
Metarhizium
; endofungal bacteria
; symbiosis
; fungal–bacteria relationship
; Bacillus
1. Introduction
Almost all organisms are associated with microorganisms during their life cycles, and some of these associations significantly influence their behavior. Symbiosis, the interaction between two or more species during development, is an example of this interaction. Based on the cost–benefit dynamics among the involved species, various symbiotic interactions occur, including parasitism, mutualism, and commensalism [1]. Fungi are not an exception; they interact with viruses and bacteria in various environments, forming complex communities that can influence their behavior. These associations can occur externally around the mycelia and internally within the hyphae [2]. Bacteria-like organisms living symbiotically inside the Endogone fungal hyphae have been reported since 1970 [3]. Filamentous fungi can harbor bacteria, mycoviruses, and other fungi within their hyphae [4]. Some endohyphal or endofungal bacteria (EHB) have been shown to alter fungal host function, development, and interactions with other organisms [2,4,5]. EHB have been identified in different phyla of fungi, including Mucormycota, Ascomycota, and Basidiomycota [6]. Potential bacterial associates were detected in 88% of the 702 fungal isolates, representing 366 genera [7]. Members of Proteobacteria, Actinobacteria, and Firmicutes were the most commonly associated with fungal isolates. The authors suggest that bacteria–fungi associations are the rule rather than the exception [4].
The presence of EHB in fungi holds significant implications for the biology of the host fungi; for instance, one of the earliest models of fungus–endobacteria symbiosis was described for the mycorrhizal fungus Gigaspora margarita and endobacteria belonging to the genus Burkholderia [8]. These EHB contain nitrogen-fixing genes [9]. An example of the importance of the fungus–endobacteria association is the plant pathogenic fungus Rhizopus microsporus, which causes rice seedling blight. In this fungus, the rhizoxin, a phytotoxin that binds the β-tubulin, inhibits mitosis and causes cell cycle arrest; it is biosynthesized by EHB in the genus Burkholderia [10]. In the same sense, R. microsporus contains the Ralstonia pickettii bacterial endosymbiont, which is required for virulence in zebrafish and mouse models and suppresses the growth of the amoeba Dictyostelium discoideum [11]. The endobacterium B. rhizoxinica is not only involved in R. microsporus virulence but also in spore formation, which is dependent on the presence of EHB. The reproduction of the fungus is dependent on the EHB, which, in turn, facilitates the formation of the toxin [12]. These EHB, previously classified as Burkholderia and then Paraburkholderia [13], now form the novel genus Mycetohabitans within Burkholderia sensu lato [14]. Other fungi also contain EHB; for example, in Ustilago maydis, the presence of a bacterial endosymbiont related to Bacillus pumilus with the capacity to fix nitrogen has been reported [15,16].
Most of the fungi that host EHB are plant-associated, including arbuscular and ectomycorrhizal fungi, plant pathogens, saprotrophic fungi, and endophytes within plant tissues [2]. These EHB may contribute to plant growth and disease resistance [5].
More than 90% of land plant species have symbiotic relationships with fungi [17]. Members of the genus Metarhizium are among the most abundant fungi isolated from soils [18,19]. Metarhizium has been isolated from soils and infected insects on all continents except Antarctica [20], and can colonize various environments, including forests, savannas, swamps, coastal zones, and deserts [21]. Metarhizium has a multifunctional lifestyle [22], being an insect-pathogenic fungus that can infect and kill more than 200 insect species, including those in at least seven orders [20]. Metarhizium plays a crucial ecological role and is closely associated with the plant rhizosphere. Metarhizium endophytically colonizes the roots of various plants, transferring insect-derived nitrogen to the plant host [23,24], and receives carbon that the plant fixes through photosynthesis [25]. This symbiotic interaction provides multiple benefits, promoting plant growth [26-29], improving salt tolerance [30,31], and protecting plants against foliar pests and plant-pathogenic fungi [32]. Metarhizium is also an antagonist of root pathogens, such as Fusarium [33], and promotes plant defense against invertebrate pests [34,35]. Metarhizium is not an obligate plant symbiont and can survive as a saprophyte [36].
Genomic analysis has shown that Metarhizium spp. is more closely related to endophytes and plant pathogens than to arthropod pathogens [22]. In this sense, the ancestors of entomopathogenic species were probably root-colonizing saprophytes that subsequently acquired genes for insect pathogenesis [37]. According to these antecedents, the Metarhizium's ability to switch between different lifestyles in soil could be influenced by bacteria present in the rhizosphere. Metarhizium–bacteria interactions have not been intensively studied, and bacterial interactions may influence Metarhizium behavior as an entomopathogen, plant growth promoter, or during plant endophytic colonization. Recently, the endobacteria Pelomonas paraquae were detected within the hyphae and conidia of M. bibiondarum and M. anisopliae [38]. However, the biological role of these bacteria in the different Metarhizium lifestyles remains unknown. In this study, strains of Metarhizium were isolated from the rhizosphere and Phyllophaga spp. Bacillus subtilis was identified as an endofungal bacterium, and its influence on the physiology and entomopathogenic capacity of Metarhizium was studied. Our analysis revealed that EHB have a negative impact on the virulence of Metarhizium against Galleria mellonella and Tenebrio molitor.
2. Materials and Methods
2.1. Fungal Isolates
Soil samples were collected in the "Las Siete Luminarias" nature reserve (20°18'46.17''N 101°12'10.41'' O) located in Valle de Santiago, Guanajuato, México, from a depth of approximately 10 cm, using a sterile hand shovel, and stored in polypropylene bags and transported to the laboratory. For each sample, 1 g of soil and 10 mL of 0.01% Triton™ X-100 (Sigma-Aldrich®) were mixed in a 15 mL Falcon tube using a vortex mixer. Serial dilutions from 10-1 to 10-7 were prepared for each sample; 100 μL of each dilution was inoculated in a non-selective medium (Potato Dextrose Agar, PDA (Bioxon®)) and incubated at 26 °C with a photoperiod of 16 h light/8 h darkness for 10–15 days. Single colonies that morphologically resembled Metarhizium were transferred to new PDA plates and incubated under the same conditions for 10 days, until conidiation occurred. Conidia were collected using a sterile toothpick and placed in an Eppendorf tube containing 1 mL of 0.01% Triton™ X-100 (Sigma-Aldrich®) and then counted using a Neubauer chamber (Hausser Scientific®). The concentration was adjusted to 1000 conidia/mL. A 50 μL volume of this dilution was inoculated on new PDA plates and incubated at 26 °C with a photoperiod of 16 h light/8 h darkness for ten days. Single colonies were selected and grown in minimal media, M-100 plates (containing 0.2% NH4NO3, 1% dextrose, 0.3% KH2PO4, and 2% 50× salt stock solution (25 g MgSO4, 0.09 g ZnSO4, 0.05 g FeSO4, 0.015 g MnSO4, and 0.02 g CuSO4)) incubated at 26 °C with photoperiod of 16 h light/8 h darkness for ten days until conidiation. Conidia were collected by scraping the colony, and the hyphae were removed by filtration through a sheet of sterile cheesecloth. The conidia were then washed three times by centrifugation and suspended in a sterile 0.01% Triton™ X-100 (Sigma-Aldrich®) solution. Additionally, larvae of Phyllophaga spp. were collected from soil obtained from "Puruagua," Jerécuaro, Guanajuato, México (20°04'45.0 "N 100°27'08.1" W). Each larva was collected manually and individually confined in a plastic container filled with sterilized soil. Plastic containers were incubated at room temperature. Dead larvae with visible Metarhizium colonization were collected, and conidia were recovered using the same protocol described for isolating Metarhizium from soil samples.
2.2. Identification of Metarhizium Isolates
Metarhizium isolates were initially identified based on morphological criteria and subsequently confirmed by molecular characterization using the gene sequence of the translation elongation factor 1-alpha (TEF1-α) [39]. The Metarhizium isolates were grown at 28 °C in Sabouraud Dextrose Medium (SDM) for 24 hours. Mycelia were recovered through filtration, and DNA was isolated using standard protocols [40]. A fragment of the TEF1-α gene was amplified by PCR using the primers EFT1 (5'-ATGGGTARGGAAGACAAGAC-3') and EFT2 (5'-GGAAGTACCAGTGATCATGTT- 3'). The PCR product was purified using the GenElute™ Gel Extraction kit (Sigma-Aldrich®), cloned into the pGEM®-T Easy Vector (Promega Corporation), and sequenced (Elim Biopharm, Inc., USA). The TEF1-α gene sequences were aligned against available representative sequences of the genus Metarhizium using MAFFT (https://www.ebi.ac.uk/Tools/msa/ mafft/).
2.3. Bacteria-free Metarhizium
To eliminate associated bacteria, conidia from the Metarhizium ES37 and PPH1 strains were successively inoculated in M-100 medium plates containing ciprofloxacin (50 µg.µL-1) and cefotaxime (100 µg.µL-1) antibiotics at 26 °C with a photoperiod of 16 h light/8 h darkness for ten days, until conidiation occurred. Conidia were collected by scraping the colony, the hyphae were removed by filtration through a sheet of sterile cheesecloth, and the conidia were washed three times by centrifugation and suspended in a sterile 0.01% Triton™ X-100 (Sigma-Aldrich®) solution. This procedure was repeated at least six times.
2.4. Endobacteria Isolation
Endobacteria were isolated following a previously described protocol [10], with some modifications. A stock solution of conidia from the Metarhizium ES37 and PPH1 strains containing 1x106 conidia.mL-1 was prepared. From this solution, 100 μL was inoculated in PDA Petri plates containing a cellophane film. The plates were incubated at 26 °C for 48 h in dark conditions to promote mycelial growth. Mycelia were recovered from the cellophane film and placed in a porcelain mortar containing 300 μL of glass beads and 1 mL of 1M NaCl. The mycelia were crushed with a porcelain pestle, and the disrupted cells were centrifuged at 19315 xg for 3 min. Subsequently, 75 μL of the supernatant was transferred to five different media. The Eppendorf tubes were again centrifuged at 19315 xg for 10 minutes, and 20 μL of the supernatant was transferred to five different media: 1. Nutrient agar (BD Bioxon®); 2. Yeast Mannitol Agar, YMA (yeast extract (BD Bioxon®), 1.5 g.L-1 mannitol (Sigma-Aldrich®), 10 g.L-1 MgSO4 (Karal®), 0.20 K2HPO4 g.L-1 (Karal®), 0.20 g.L-1 FeCl3 (Karal®), 1 mL-1 bacteriological agar (BD Bioxon®), and Congo red (Sigma-Aldrich®), pH 7, 10 mL.L-1); 3. YMA without Congo red; 4. Luria Bertani (BD Bioxon®); and 5. Actinomycete selective medium (20 g soybean flour, 5 g yeast extract (BD Bioxon®), and 20 g mannitol (Sigma-Aldrich®)). Samples were incubated at 30 °C for 24 hours. Bacterial isolates were then selected and purified by repeated subculturing and maintained in the corresponding media.
2.5. Identification of bacterial Isolates
Bacterial colonies were grown overnight at 30 °C in YMA medium without Congo red, and the cells were collected by centrifugation. Bacterial cells were identified morphologically by Gram staining. Bacterial cells were recovered and crushed in a porcelain mortar with liquid nitrogen. The obtained powder was resuspended in 400 mL of NTES (0.1 M NaCl, 10 mM Tris–HCl (pH 8.0), 1 mM EDTA, and 1% SDS), and DNA extraction was performed using the phenol–chloroform technique [40]. For molecular identification, the 16S rDNA was amplified using JumpStart™ Taq ReadyMix™ (Sigma-Aldrich®) and three pairs of universal primers: 1) 63F (CAGGCCTAACACATGCAAGTC) and M1387R (GGGCGGWGTGTACAAGRC); 2) 27F (AGAGTTTGATCMTGGCTCAG) and 1492R (TACGGYTACCTTGTTACGACTT); 3) 27F (AGAGTTTGATCMTGGCTCAG) and R1494 (CTACGGRTACCTTGTTGTTACGAC) [41,42]. PCR conditions were 95 °C for 1 min, 32 cycles at 95 °C for 20 s, 55 °C for 45 s, 72 °C for 2 min, and an additional cycle at 72 °C for 5 min. PCR products of the expected size (approximately 1.5 Kbp) were purified using the GenElute™ Gel Extraction kit (Sigma-Aldrich®), cloned into the pGEM™-T Easy Vector (Promega Corporation), and sequenced (Elim Biopharm, Inc., USA).
2.6. Bacterial DNA Extraction and Sequencing
To extract bacterial genomic DNA from the B. subtilis 1E strain, bacterial cultures were grown in liquid minimal medium (0.67% BD Yeast Nitrogen Base, 1% BD dextrose) to a concentration of 6 x107 cells.mL-1, and cells harvested by centrifugation in the early log phase of growth. Total bacterial DNA was extracted using the DNeasy Blood & Tissue Kit (Qiagen, cat. no. 69504) according to the manufacturer's instructions, which included a pretreatment step for Gram-positive bacteria. DNA integrity was verified by electrophoresis, and DNA concentration was measured using a NanoDrop 2000c spectrophotometer (Thermo Fischer Scientific). The bacterial genomic DNA from B. subtilis 1E was sequenced using the whole-genome sequencing and assembly of genomic DNA service offered by Plasmidsaurus (https://plasmidsaurus.com/bacteria_sequencing#%23bact_hybrid), employing the hybrid option. This option combines long-read sequencing using Oxford Nanopore Technology with Illumina bacterial genome sequencing to polish the Oxford Nanopore reads. The B. subtilis 1E sequencing results are presented in Table S1.
2.7. Comparative genomic Analysis
To investigate the pangenome structure of B. subtilis 1E, we performed pangenome analysis using PPanGGOLiN v1.1.126, using the default parameters [43]. The study included the genomes of B. subtilis 1E (NCBI accession number CP196825) and the reference B. subtilis strains BSP1 (GCF_000321395.1) and 168 (GCF_000009045.1), both retrieved from the NCBI RefSeq database. This approach allowed us to identify gene families shared by all strains, as well as those exclusive to either endosymbiotic or reference genomes, providing insights into their genomic diversities and potential functional differences. Following the comparative analysis, the singletons in B. subtilis 1E were annotated using eggNOG-mapper [44]. The annotation and function prediction were performed using different databases: GeneMarkS, Gene Ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG), and Clusters of Orthologous Groups (COG).
2.8. Localization of Endobacteria Using Microscopy
Endobacterial detection using microscopy was performed according to a previously described protocol [10], using the green-fluorescent nucleic acid stain SYTO®9 (Thermo Fisher Scientific). The mycelia of a growing culture of Metarhizium strains were transferred to an Eppendorf tube containing 0.5 mL of 0.85% NaCl. An aliquot (10 μL) was placed on a microscope slide and stained with 0.5 μL of SYTO® 9 (5 μM). The samples were incubated in the dark for 30 minutes and analyzed using a laser scanning confocal microscope (ZEISS LSM 700).
2.9. Conidial Yield
To determine the conidial yield, 1000 conidia from Metarhizium ES37 and PPH1 strains and their bacteria-free derivatives (Metarhizium ES37-2 and PPH1-1 strains) were inoculated in the center of Petri dishes containing M-100 media or Potato Dextrose Agar (PDA). The Petri plates were incubated at 28 °C for 10 days (M-100) or 15 days (PDA) with a photoperiod of 16 hours of light and 8 hours of darkness, or in complete darkness. After incubation, the colony diameter was measured, and the conidia were collected and resuspended in 1 mL of 0.01% Triton™ X-100 (Sigma-Aldrich®). Conidia concentration was determined using a Neubauer chamber under a microscope. The experiments were performed three times, with three replicates per experiment.
2.10. Conidia Germination Assay
To determine whether the presence of endobacteria in Metarhizium affects conidia germination, 1x103 conidia were inoculated in a sterile microscope glass slide containing 2.5 mL of M-100 or PDA medium. Slides were incubated at 28°C for 12, 18, and 24 h. At least 300 cells were counted under the microscope for each time point, and the percentage of germlings and ungerminated conidia was determined. A germling was defined as a germ tube equal in length to the width of the conidia. The experiments were performed three times, with three replicates per experiment.
2.11. Insect bioassays
Fresh conidia of Metarhizium strains with endobacteria (ES37 and PPH1) and endobacteria-free Metarhizium strains (ES37-2 and PPH1-1) were used in bioassays against Galleria mellonella larvae. Sixty larvae of each strain were inoculated by immersion with conidia suspended in 0.01% Triton™ X-100 (Sigma-Aldrich®) at a dilution of 1x107 conidia.mL-1. An additional 60 larvae were inoculated with 0.01% Triton™ X-100 (Sigma-Aldrich) and served as a control. Infection was monitored every 24 hours. Larvae were fed an artificial diet daily. The experiments were performed in four independent assays.
Fresh conidia of a Metarhizium strain with endobacteria (ES37) and an endobacteria-free Metarhizium strain (ES37-2) were used in bioassays against Tenebrio molitor larvae. All larvae were bred in plastic cages (47x26x12 cm). Wheat bran and cornmeal (1:1 ratio) were provided ad libitum as food, and apples and oranges were provided twice a week. A photoperiod of 12 hours of light and 12 hours of darkness, a temperature of 27 ± 2 °C, and a relative humidity of 45 ± 5% were maintained in the colony. Size was controlled (1.5–1.8 cm) in all experiments. A previously described experimental protocol was used [45]. Larvae (n = 300) were injected with a 1 μL suspension of PBS–Tween® 80 (0.01%) (Sigma-Aldrich®) containing three different conidia concentrations (5, 50, and 100 conidia) of the Metarhizium strain containing endobacteria (ES37) and endobacteria-free Metarhizium strain (ES37-2). PBS–Tween® 80 (0.01%) (Sigma-Aldrich®) was used as a control. Injections were performed through the pleural intersegmental membrane between the sixth and seventh abdominal segments using sterilized micro syringes (10 μL Hamilton syringe). Larvae were individually separated in plastic plates with 12 wells (Corning), and food was provided as described above. Survival was recorded every day for 10 days, and the cause of death was corroborated. The experiments were performed in four independent assays.
2.12. Phylogenetic analyses
Phylogenetic analyses were performed using MEGA-X v10.2.6, employing the statistical maximum likelihood method with the general time reversible (GTR) model. The phylogenetic tree was constructed using Bayesian inference with Mr. Bayes v3.2.6 software and the GTR model with an invariant gamma distribution range, with a bootstrap of 1000.
2.13. Statistical Analysis
Statistical analyses were performed using the GraphPad Prism 10.2 software. A p-value of less than 0.0001 was used. The results of the conidiation test were analyzed using the Kruskal–Wallis statistical test and one-way ANOVA. The unpaired multiple t-test statistic was used for the germination tests. For survival analyses, the Kaplan–Meier method was used to generate the survival curve, and the log-rank test (Mantel–Cox) was employed to compare the survival distributions between the groups.
3. Results
3.1. Associated endobacteria Can Be Identified in Native Metarhizium Strains
To investigate the presence of endobacteria in Metarhizium, strains from the rhizosphere and larvae of Phyllopagha spp. were isolated in two different regions of Guanajuato, México. The isolates were cultured in media without antibiotics and by successive reseeding. Isolates with morphological characteristics resembling those of the genus Metarhizium were selected.
We found associated Gram-positive bacteria in the periphery of the hyphae in some Metarhizium isolates; this interaction persisted even after consecutive reseedings. Conidia obtained from these isolates were decontaminated and washed extensively. These conidia, when observed under the microscope, did not exhibit any external bacterial associations; likewise, when these conidia were inoculated in Sabouraud dextrose medium, the Petri dishes did not show any bacterial colonies.
Two isolates, one from the rhizosphere (ES37) and one from Phyllopagha spp larvae (PPH1), were selected for further study. Molecular identification of both isolates was performed by amplifying and sequencing a fragment of the TEF-1α gene [39]. Sequence BLASTn analysis identified the isolated ES37 strain as M. robertsii, while PPH1 shared the highest similarity with M. pinghaense (Figure S1).
The presence of endobacteria in the M. robertsii and M. pinghaense strains was determined according to the protocol described by Partida-Martinez and Hertweck (2005) [10] using the SYTO9® green-fluorescent nucleic acid stain. After culturing for four days, the mycelia were stained with SYTO9®, and endobacteria appeared as fluorescent dots within the hyphae (Figure 1A, C). These fluorescent dots were consistently observed in both strains after several successive reseedings. Both strains' fluorescent dots remained visible after four years of cultivation. To obtain endobacteria-free Metarhizium, conidia from the Metarhizium ES37 and PPH1 strains were cultivated on M-100 medium plates containing the antibiotics ciprofloxacin and cefotaxime, which have previously been reported to isolate endobacteria-free fungi [10]. Endobacterial loss was confirmed when no amplification products were observed during 16S rDNA PCR amplification using 63F-M1387R and 27F-1492R primer pairs (Figure S2). These derivatives were designated ES37-2 and PPH1-1, respectively. Mycelia from these Metarhizium derivatives were stained with SYTO®9; the green fluorescent dots indicating the presence of endobacteria were not observed, despite overexposure of the samples under the microscope, which showed only nuclear fluorescence (Figure 1B, D).
3.2. Metarhizium Endobacteria Isolation and Identification
Mycelia from M. robertsii ES37 and M. pinghaense PPH1 were mechanically sheared and subjected to centrifugation. Aliquots of the supernatant were inoculated in five different media: Nutrient agar, Yeast Mannitol Agar (YMA), YMA without Congo red, Luria Bertani Agar (LBA), and an Actinomycete selective medium. Bacillus-like bacterial colonies were observed only in the YMA without Congo red medium. We isolated seven colonies from the M. robertsii ES37 and four colonies from the M. pinghaense PPH1. To identify the isolated bacterial species, the 16S rDNA gene sequences were amplified by PCR using three universal bacterial primer pairs: 63F–M1387R, 27F–1492R, and 27F–R1494 [41,42]. Amplification products were cloned and sequenced (accession numbers: 1B, PV770094; 1E, PV770095; 2B, PV770096; 4B, PV770097; 4D, PV770098; 4C, PV770099; 2C, PV770100; 4A, PV770101; I-2, PV770102; 1C, PV770103; I-4, PV770104). BLASTn analysis revealed that all colonies had the highest similarity with Bacillus subtilis (Table S2).
3.3. Endobacteria's Impact on Metarhizium Conidiation
Since both strains, M. robertsii ES37 and M. pinghaense PPH1, contain associated bacteria, their germination and conidiation capabilities were assessed to determine whether these endobacteria affected their morphology and growth. The obtained endobacteria-free M. robertsii ES37-2 and M. pinghaense PPH1-1 were used. A total of 1,000 conidia were inoculated in M-100 and PDA media. The strains were then incubated at 28 °C under a photoperiod or in total darkness. After ten days of incubation, the morphology of Metarhizium was compared between strains with endobacteria and bacteria-free strains (Figure 2). No significant difference was observed in the radial growth of colonies with endobacteria compared to the bacteria-free colonies. Furthermore, no other morphological changes were noted. The conidia produced by both endobacteria-containing strains and bacteria-free strains, cultured in M-100 and PDA media, were subsequently collected and quantified. Conidial production in M-100 medium in photoperiod showed no significant difference among strains (Figure 3A). However, under dark conditions, the conidiation of M. robertsii ES37 with endobacteria was higher than that of the bacteria-free strain (ES37-2). Conversely, the M. pinghaense strain with endobacteria (PPH1-1) exhibited a lower conidiation rate than the bacteria-free strain (PPH1) (Figure 3B). In the PDA medium, the conidiation rate of the bacteria-free M. pinghaense was higher than that of M. pinghaense with endobacteria (Figure 3C and D). No difference was found in the conidiation of the M. robertsii strains under photoperiod conditions (Figure 3C), and conidiation was absent under darkness (Figure 3D).
M. robertsii ES37 and M. guizhouuense PPH1 containing EHB and their derivatives, EHB-free M. robertsii ES37-2 and M. guizhouuense PPH1-1, grow in M-100 medium after 10 days of incubation, both under total darkness and a photoperiod of 16 h light/8 h darkness.
Conidia production by M. robertsii ES37, M. robertsii ES37-2, M. guizhouuense PPH1, and M. guizhouuense PPH1-1 was quantified after 10 days of incubation in M-100 medium (A, B) and after 15 days of incubation in PDA medium (C, D). Petri plates were incubated under photoperiods of 16 h light/8 h darkness (A, C) and total darkness (B, D). T-test statistical analysis yielded a p-value <0.005. The degree of significant difference is indicated with * in the range from one to three
The germination of conidia with endobacteria and bacteria-free conidia was assessed by inoculating 1000 conidia in M-100 and PDA media. The percentage germination was analyzed at 12, 18, and 24 hours. In the M-100 medium, the germination of bacteria-free strains exceeded that of strains with endobacteria (Figure 4). This difference was particularly pronounced in M. robertsii with endobacteria, which exhibited a percentage germination of 6.7% at 12 hours; in contrast, bacteria-free M. robertsii had a germination percentage of 19.2%. In PDA medium, all strains had surpassed 50% germination within 12 hours, except for M. robertsii with endobacteria, and by 18 hours, all had achieved 100% germination (Figure 4). The results show that the presence of endobacteria generally delays the germination of Metarhizium.
The percentage germination was quantified at 12, 18, and 24 hours in M-100 medium, and at 12 and 18 hours in PDA medium.
3.4. Associated Endobacteria Affected Metarhizium Virulence
Bioassays were conducted using Galleria mellonella larvae to investigate the influence of endobacteria on the virulence of Metarhizium. Larvae were inoculated by immersion in a 0.01% Triton™ X-100 conidia suspension, with a final concentration of 1x107 conidia. mL-1. As a control, larvae were immersed in 0.01% Triton™ X-100. Larval mortality was monitored every 24 hours. Bacteria-free M. robertsii ES37-2 exhibited a higher mortality rate, with over 50% mortality by the fifth day. Both M. robertsii ES37 and M. pinghaense PPH1 with endobacteria had significantly lower mortality rates compared with the bacteria-free strains (Figure 5). The presence of endobacteria in the two Metarhizium isolates resulted in decreased virulence.
Conidia (1x108 conidia.mL-1) from M. robertsii ES37, M. robertsii ES37-2, M. guizhouense PPH1, and M. guizhouense PPH1-1 were inoculated into G. mellonella larvae, and infection was monitored every 24 hours. Control treatments were carried out using 0.01% Triton X-100. The results show the mean percentage survival. The horizontal axis shows days after infection. Statistical analysis: Mantel–Cox (log-rank). X2 = 92.71 df = 4; p-value <0.001.
Virulence was measured by injecting T. molitor larvae instead of topical infection. Three different conidia concentrations of M. robertsii ES37 and ES37-2 strains were analyzed: 5, 50, and 100 (Figure 6). As observed with the topical infection, the presence of endobacteria in Metarhizium decreased the virulence, indicating that the presence of EHBA significantly increased survival.
3.5. Genomic Analysis of Associated Endobacteria
The previous experiments showed the influence of the endobacterium B. subtilis on the physiology and virulence of Metarhizium, as all the EHB isolated in this study were identified as B. subtilis. To analyze the genome differences between the B. subtilis strain isolated from Metarhizium and the B. subtilis reference strains, which may explain their presence in Metarhizium, the genome of the B. subtilis 1E strain, isolated from M. robertsii ES37, was sequenced (NCBI accession number CP196825). The genome is 4.1 Mb in size and contains 4242 genes (Table S1). The genome analysis predicted the presence of 86 tRNAs and 30 rRNAs (Table S3). The genome sequence was compared with that of the undomesticated strain B. subtilis BSP1 [46] and the reference strain B. subtilis 168 (Figure 7). The three genomes share 3551 genes, and the endobacterium B. subtilis 1E has 180 genes that are not present in the reference genomes BSP1 and 168 (Table S4). Classification of gene functions in the Database of Clustered Orthologous Groups (COGs) revealed that 101 correspond to an unassigned function, 23 have an unknown function, ten have a transcription function, eight have a replication and repair function, twelve have a cell wall/membrane/biogenesis function, and five have a defense mechanism function (Figure 8). The genes potentially encode proteins with transcription function, including the MerR family transcription factor regulator (OOBLJC_RS00805); members of this family respond to oxidative stress, cellular ion imbalance, toxins, endogenous metabolites, and antibiotics [47,48]. Members of the TetR/AcrT family transcriptional regulator (OOBLJC_RS08765) regulate genes involved in antibiotic production, osmotic stress, efflux pumps, multidrug resistance, metabolic modulation, and pathogenesis [49].
Among the twelve genes that have a cell wall/membrane/biogenesis function, we identified genes involved in the biosynthesis of wall teichoic acids (WTAs), essential components of the Gram-positive bacterial cell wall; the glycerol-3-phosphate cytidylyltransferase (OOBLJC_RS03400), which catalyzes the synthesis of CDP-glycerol; and glycerophosphotransferase (OOBLJC_RS03370; OOBLJC_RS03405; OOBLJC_RS03425), which transfers the glycerolphosphate unit to the growing teichoic acid polymer [50]. The tarQ gene (OOBLJC_RS03485) encodes the (poly)ribitol-phosphate teichoic acid beta-D-glucosyltransferase, which attaches b-glucose units to poly(RboP)-WTAs [51]. Analyzing 143 B. subtilis genomes revealed that the WTAs' gene cassettes exhibit a high level of variation, which may be attributed to one or more independent horizontal transfer events during the evolution of B. subtilis [52]. Of the 180 genes present in the B. subtilis 1E genome, we identified the putative holin-like toxin (OOBLJC_RS08630). One of these proteins was also identified in B. pumilus, indicating that it causes toxicity to the host cell by disrupting the membrane [53].
4. Discussion
Fungi are successful soil inhabitants due to their plasticity and capacity to respond to adverse or unfavorable conditions [54]. There is growing evidence of fungi, including yeast, and their associated EHB, as reported for Kluyvermyces marxianus [55] and Candida albicans [56,57]. As mentioned previously, more than 90% of land plant species have symbiotic relationships with fungi [17]. Potential bacterial associates have been detected in 88% of the 702 fungal isolates [7], and EHB have been identified in different phyla of fungi, including Mucormycota, Ascomycota, and Basidiomycota [6]. EHB have a distinct influence on fungal development [2,5,58,59]. The impact of EHB on fungal resistance to metals has been studied [60]. EHB influence fungal sexual reproduction [61] and enhance the production of indole-3-acetic acid [62]. EHB (Bacillus licheniformis, Achromobacter xylosoxidans, and Stenotrophomonas maltophilia) in the nematode-trapping fungus Arthrobotrys musiformis play an essential role in nitrogen cycling and nematode trap formation [63]. In fungal symbiosis with plants, EHB in the fungal host contribute to plant growth and disease resistance to microbial pathogens [5].
Since the Metarhizium genus is among the most abundant fungi isolated from soils [18,19], and it has been suggested that many fungi harbor their own microbiomes [4], we investigated the presence of associated endobacteria in native Metarhizium strains isolated from the rhizosphere and the insect Phyllophaga spp. We found associated Gram-positive bacteria in the periphery of the hyphae in some Metarhizium isolates; this interaction persisted even after consecutive reseedings. Conidia were obtained from these isolates, and after surface decontamination and extensive washing, EHB were detected using the SYTO®9 dye, revealing characteristic green dots, as reported previously in different fungi, including Metarhizium [10,38]. These signals persist despite continuous reseeding over four years. These signals were lost when the cells were cultured in the presence of antibiotics through successive reseeding. The absence of EHB was verified through PCR amplification of the 16S rDNA gene. It is essential to note that in some isolates, amplification of the 1.5 Kbp fragment corresponding to the 16S rDNA gene is positive despite the apparent lack of green signals produced by the SYTO®9 dye, indicating the presence of EHB. Only in two isolates, M. robertsii ES37 isolated from the rhizosphere and M. guizhouense PPH1 isolated from Phyllophaga spp., do we obtain strains with a correlation between the lack of green signals based on the SYTO®9 dye and the absence of amplification of the 1.5 Kbp 16S rDNA gene fragment.
To isolate the potential EHB, mycelia from M. robertsii ES37 and M. guizhouense PPH1 were crushed and analyzed in five different media. The bacteria grew only in the Yeast Mannitol Agar (YMA) medium, which is used to cultivate soil microorganisms. Bacterial colonies were isolated from the medium. Molecular analysis identified the colonies as B. subtilis. The intracellular occurrence of Bacillus has been described in other fungi. In U. maydis, B. pumilus has been detected in strains isolated from natural sources; these bacteria can fix nitrogen. Studying the interactions between fungi and bacteria in plant rhizoplanes revealed that Bacillus was associated as an exobacterium in 80% of the fungi and as a putative endobacterium in 15% of the fungi [64]. In the same study, 80% of the isolated fungi and endobacterial genera potentially involved in the nitrogen cycle were identified [64]. B. subtilis was identified as the endobacterium in Fusarium acuminatum isolated from the roots of Spiranthes sinesis (Orchidaceae) [65]. B. subtilis in Metarhizium isolates may be a more general phenomenon in soils and insects in different locations. These bacteria may have nitrogen-fixing functions, although they may also employ Metarhizium hyphae as a form of dispersal ("fungal highway"), as has been demonstrated in other systems [66,67].
In addition to the complexity of the soil, spatial heterogeneity, temporal dynamics, and land use [68], bacterial communities in soils from different locations worldwide have been analyzed. In China, the dominant bacterial groups identified in 13 saline soils were Proteobacteria, Bacteroidetes, Actinobacteria, Firmicutes, and Gemmatimonadetes [69]. Cyanobacteria, Proteobacteria, Firmicutes, and Bacteroidetes were the most abundant bacterial groups in grapevine habitats [70]. In the Mezquital Valley in México, Actinobacteria, Planctomycetes, Proteobacteria, Acidobacteria, Chloroflexi, Gemmatimonadetes, Verrucomicrobia, Bacteroidetes, Firmicutes, and Nitrospirae were identified in dryland soils [71].
Several bacterial strains have been isolated from the hemocoel of Phyllophaga spp., including Serratia marcescens [72] and Klebsiella [73]. In Anomala sp. (Coleoptera: Scarabaeidae), the distribution of gut bacteria revealed that Ochrobactrum spp. were the most dominant bacteria, followed by Bacillus, Citrobacter, Pseudomonas, Enterobacter, and Paenibacillus [74].
Since the Metarhizium strains ES37 and PPH1 were isolated from soil and Phyllophaga spp., respectively, we cannot dismiss the possibility that other EHB, besides B. subtilis, are present in different Metarhizium strains. Arbuscular mycorrhizal fungi (AMF) contain EHB in their hyphae and spores. EHB have also been identified in other fungi. The endobacteria Glomeribacter gigasporarum [75] and Moeniiplasma glomeromycotorum are the most common endobacteria in Gigaspora margarita [76]. In addition to G. gigasporarum and M. glomeromycotorum, bacteria belonging to ten other phyla, including Alphaproteobacteria (Rhizobiales, Rhodospirillales), Gammaproteobacteria (Escherichia, Aquicella, Pseudomonas), Actinobacteria, Acidobacteria, and Verrucomicrobia, were identified in spores of AMF isolated from different locations [77]. Bacillus sp., B. thuringiensis, and Paenibacillus rhizospherae were isolated as putative EHB from the spores of G. margarita [78].
The presence of endobacteria in M. robertsii ES37 and M. pinghaense PPH1 delays the germination of Metarhizium, influences conidia production, and decreases virulence against P. xylostella and T. molitor. The impact of bacteria on fungal development is well described; treatment with the B. subtilis strain HSY21 has been shown to inhibit the growth and expression of pathogenic genes in F. oxysporum [79]. Using the B. subtilis strain F62 against F. oxysporum showed that the inhibition of mycelial growth occurred primarily through diffusible compounds [80]. Soil isolates of B. subtilis exhibited significant fungistatic activity against M. robertsii [81]. Similarly, Aphis bacterial associates from different genera, including Acinetobacter, Bacillus, Pseudomonas, and Staphylococcus, inhibit the growth of Metarhizium mycelia [82].
The influence of Metarhizium in the rhizosphere microbiome has not been thoroughly investigated. The community profiles (bacteria and fungi) of the Phaseolus vulgaris rhizosphere have been examined following exposure to M. robertsii conidia in the presence and absence of G. mellonella larvae [83]. The addition of Metarhizium conidia increased the quantity of plant growth-promoting organisms, including Bradyrhizobium, Flavobacterium, Chaetomium, and Trichoderma. Similarly, the number of members of the genus Bacillus increased in the roots following the addition of Metarhizium conidia, with this increase being more pronounced in the presence of G. mellonella larvae [83]. Recent research suggests that bacteria can exploit fungi for shelter, thereby facilitating survival under unfavorable conditions, as internalization increases bacterial fitness when challenged by abiotic stresses [84].
The presence of B. subtilis as an endobacterium in Metarhizium suggests a close relationship that can facilitate gene exchange and horizontal gene transfer between the two organisms. It has been reported that M. robertsii has 18 genes involved in the infection process, which were potentially acquired through ancient horizontal gene transfer from a bacterium [85]. The authors suggested that the need to degrade insect cuticles served as a selective pressure to retain these genes, as twelve of them are upregulated during penetration, and six have a role in the penetration process [85].
Comparison of the B. subtilis 1E genome with those of B. subtilis reference strains 168 and BSP1 identified 180 genes that are absent in the reference genomes, 124 of which encode proteins with unassigned or unknown functions. Among the proteins with known function, ten are involved in transcription, eight in replication and repair, twelve in cell wall/membrane/envelope biogenesis, and five in defense mechanisms. A specific function that clearly explains the role of B. subtilis 1E in Metarhizium has not been identified.
The presence of extracellular bacterial communities or EHB in Metarhizium, including their influence on behavior in various Metarhizium lifestyles, has not been thoroughly studied. Our results demonstrate the inhibitory impact of bacteria on Metarhizium growth and virulence capacity; however, there may be mechanisms or conditions under which these bacterial communities provide advantages or benefits to Metarhizium, such as through plant interactions in low-nitrogen conditions, in which the presence of EHB may provide better survival conditions. For example, we identified 19 genes in the B. subtilis 1E genome that encode proteins involved in the nitrogen cycle or in nitrogen metabolism (Table S5), including the putative nitrogen fixation protein yutI (OOBLJC_05140), as well as narG (nitrate reductase, alpha subunit, OOBLJC_02520), narH (nitrate reductase, beta subunit, OOBLJC_02525), and narI (nitrate reductase, gamma subunit, OOBLJC_02535), belonging to the narGHJI locus, encoding the membrane-bound nitrate reductase, which is significantly induced when B. subtilis is undergoing anaerobic growth [86]. Other genes in the B. subtilis 1E genome that are involved in nitrate and nitrite respiration and are induced under anaerobic growth conditions include nark (nitrate transporter, OOBLJC_02500), nasD (Subunit of NADH-dependent nitrite reductase, OOBLJC_19895), and nasE (subunit of NADH-dependent nitrite reductase, OOBLJC_19900) [86].
This study presents several opportunities for exploring Metarhizium and its applications in agricultural pest control and plant growth promotion—areas that have not been thoroughly investigated to date. Several questions remain to be answered; for example, is the microbiome associated with Metarhizium dependent on where it is isolated? Is there a specific association with a particular bacterium? Is there a particular mechanism for bacterial entry, as analyses of lipopeptides have previously suggested? [84]. We plan to address these questions in future work.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/doi/s1, Figure S1: Phylogenetic analysis of Metarhizium strains; Figure S2: Gel electrophoresis of 16S rDNA fragments; Table S1: Statistics from B. subtilis 1E whole-genome sequence analysis; Table S2: Identification of endobacteria cultured from M. robertsii ES37 and M. pinghaense PPH1; Table S3: Comparative genomic features of B. subtilis 1E and the reference B. subtilis genomes; Table S4: Comparison of genes in B. subtilis 1E and the reference B. subtilis genomes; Table S5: Genes present in the B. subtilis 1E genome, encoding proteins involved in the nitrogen cycle or in nitrogen metabolism.
Author Contributions
Conceptualization, I.E.P.G. and J.C.T.G.; investigation, A.G.M.A., I.R.A., A.M.V., I.H.P.T., and A.D.C.; T. molitor bioassays, J.C.G.; DNA and sequence analysis, I.E.P.G. and Z.F.N-L; resources, G.A.G.H. and J.C.T.G.; writing—original draft preparation, A.G.M.A. and G.A.G.H.; writing—review and editing, J.C.T.G.; supervision, project administration, J.C.T.G.; funding acquisition, I.E.P.G., G.A.G.H., and J.C.T.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Secretaría de Ciencia, Humanidades, Tecnología e Innovación (grant number CBF2023-2024-1745) and Universidad de Guanajuato (grant numbers CIIC 027/2024 and CIIC 117/2025). A.G.M.A. and I.H.P.T. received scholarships from the Secretaría de Ciencia, Humanidades, Tecnología e Innovación.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
We thank Abraham Ashael Gasca-Venegas for his support with the microscopy observations.
Conflicts of Interest
The authors declare that they have no conflicts of interest. The funders had no role in the design of the study, in the collection, analysis, or interpretation of data, in the writing of the manuscript, or in the decision to publish the results.
Abbreviations
The following abbreviation is used in this manuscript:
| EHB | Endofungal bacteria |
References
- Moya, A.; Peretó, J.; Gil, R.; Latorre, A. Learning how to live together: genomic insights into prokaryote–animal symbioses. Nature Reviews Genetics 2008, 9, 218–229. [Google Scholar] [CrossRef]
- Araldi-Brondolo, S.J.; Spraker, J.; Shaffer, J.P.; Woytenko, E.H.; Baltrus, D.A.; Gallery, R.E.; Arnold, A.E. Bacterial Endosymbionts: Master Modulators of Fungal Phenotypes. Microbiology Spectrum 2017, 5, 10.1128. [Google Scholar] [CrossRef]
- Mosse, B. Honey-coloured, sessile Endogone spores: II. Changes in fine structure during spore development. Archiv für Mikrobiologie 1970, 74, 129–145. [Google Scholar] [CrossRef]
- Kelliher, J.M.; Robinson, A.J.; Longley, R.; Johnson, L.Y.D.; Hanson, B.T.; Morales, D.P.; Cailleau, G.; Junier, P.; Bonito, G.; Chain, P.S.G. The endohyphal microbiome: current progress and challenges for scaling down integrative multi-omic microbiome research. Microbiome 2023, 11, 192. [Google Scholar] [CrossRef]
- Alabid, I.; Glaeser, S.; Kogel, K.-H. Endofungal Bacteria Increase Fitness of their Host Fungi and Impact their Association with Crop Plants. Current issues in molecular biology 2018, 30, 59–74. [Google Scholar] [CrossRef]
- Partida-Martínez, L.P. The fungal holobiont: Evidence from early diverging fungi. Environ Microbiol 2017, 19, 2919–2923. [Google Scholar] [CrossRef]
- Robinson, A.J.; House, G.L.; Morales, D.P.; Kelliher, J.M.; Gallegos-Graves, L.V.; LeBrun, E.S.; Davenport, K.W.; Palmieri, F.; Lohberger, A.; Bregnard, D.; et al. Widespread bacterial diversity within the bacteriome of fungi. Communications Biology 2021, 4, 1168. [Google Scholar] [CrossRef]
- Bianciotto, V.; Bandi, C.; Minerdi, D.; Sironi, M.; Tichy, H.V.; Bonfante, P. An obligately endosymbiotic mycorrhizal fungus itself harbors obligately intracellular bacteria. Appl Environ Microbiol 1996, 62, 3005–3010. [Google Scholar] [CrossRef]
- Minerdi, D.; Fani, R.; Gallo, R.; Boarino, A.; Bonfante, P. Nitrogen Fixation Genes in an Endosymbiotic<i>Burkholderia</i> Strain. Applied and Environmental Microbiology 2001, 67, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Partida-Martinez, L.P.; Hertweck, C. Pathogenic fungus harbours endosymbiotic bacteria for toxin production. Nature 2005, 437, 884–888. [Google Scholar] [CrossRef] [PubMed]
- Itabangi, H.; Sephton-Clark, P.C.S.; Tamayo, D.P.; Zhou, X.; Starling, G.P.; Mahamoud, Z.; Insua, I.; Probert, M.; Correia, J.; Moynihan, P.J.; et al. A bacterial endosymbiont of the fungus Rhizopus microsporus drives phagocyte evasion and opportunistic virulence. Curr Biol 2022, 32, 1115–1130.e1116. [Google Scholar] [CrossRef]
- Partida-Martinez, L.P.; Groth, I.; Schmitt, I.; Richter, W.; Roth, M.; Hertweck, C. Burkholderia rhizoxinica sp. nov. and Burkholderia endofungorum sp. nov., bacterial endosymbionts of the plant-pathogenic fungus Rhizopus microsporus. Int J Syst Evol Microbiol 2007, 57, 2583–2590. [Google Scholar] [CrossRef] [PubMed]
- Sawana, A.; Adeolu, M.; Gupta, R.S. Molecular signatures and phylogenomic analysis of the genus Burkholderia: proposal for division of this genus into the emended genus Burkholderia containing pathogenic organisms and a new genus Paraburkholderia gen. nov. harboring environmental species. Front Genet 2014, 5, 429. [Google Scholar] [CrossRef] [PubMed]
- Partida-Martínez, L.P. Fungal holobionts as blueprints for synthetic endosymbiotic systems. PLoS biology 2024, 22, e3002587. [Google Scholar] [CrossRef]
- Ruiz-Herrera, J.; León-Ramírez, C.; Vera-Nuñez, A.; Sánchez-Arreguín, A.; Ruiz-Medrano, R.; Salgado-Lugo, H.; Sánchez-Segura, L.; Peña-Cabriales, J.J. A novel intracellular nitrogen-fixing symbiosis made by Ustilago maydis and Bacillus spp. New Phytologist 2015, 207, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rodríguez, F.; González-Prieto, J.M.; Vera-Núñez, J.A.; Ruiz-Medrano, R.; Peña-Cabriales, J.J.; Ruiz-Herrera, J. Wide distribution of the Ustilago maydis-bacterium endosymbiosis in naturally infected maize plants. Plant Signal Behav 2021, 16, 1855016. [Google Scholar] [CrossRef]
- Behie, S.W.; Padilla-Guerrero, I.E.; Bidochka, M.J. Nutrient transfer to plants by phylogenetically diverse fungi suggests convergent evolutionary strategies in rhizospheric symbionts. Commun Integr Biol 2013, 6, e22321. [Google Scholar] [CrossRef]
- Milner, R.; Lomer, C.; Prior, C. Biological control of locusts and grasshoppers. 1992.
- Nishi, O.; Sato, H. Isolation of Metarhizium spp. from rhizosphere soils of wild plants reflects fungal diversity in soil but not plant specificity. Mycology 2019, 10, 22–31. [Google Scholar] [CrossRef]
- Roberts, D.W.; St Leger, R.J. Metarhizium spp., cosmopolitan insect-pathogenic fungi: mycological aspects. Adv Appl Microbiol 2004, 54, 1–70. [Google Scholar] [CrossRef]
- Zimmermann, G. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Science and Technology 2007, 17, 553–596. [Google Scholar] [CrossRef]
- Stone, L.B.L.; Bidochka, M.J. The multifunctional lifestyles of Metarhizium: evolution and applications. Appl Microbiol Biotechnol 2020, 104, 9935–9945. [Google Scholar] [CrossRef] [PubMed]
- Behie, S.W.; Zelisko, P.M.; Bidochka, M.J. Endophytic insect-parasitic fungi translocate nitrogen directly from insects to plants. Science 2012, 336, 1576–1577. [Google Scholar] [CrossRef] [PubMed]
- Behie, S.W.; Bidochka, M.J. Ubiquity of insect-derived nitrogen transfer to plants by endophytic insect-pathogenic fungi: an additional branch of the soil nitrogen cycle. Appl Environ Microbiol 2014, 80, 1553–1560. [Google Scholar] [CrossRef]
- Behie, S.W.; Moreira, C.C.; Sementchoukova, I.; Barelli, L.; Zelisko, P.M.; Bidochka, M.J. Carbon translocation from a plant to an insect-pathogenic endophytic fungus. Nat Commun 2017, 8, 14245. [Google Scholar] [CrossRef]
- Garcia, J.; Elena; Posadas, J. ; Perticari, A.; Alejandro, L.; Roberto, E. Metarhizium anisopliae (Metschnikoff) Sorokin Promotes Growth and Has Endophytic Activity in Tomato Plants. Advances in Biological Research 2011, 5, 22–27. [Google Scholar]
- Liao, X.; O'Brien, T.R.; Fang, W.; St Leger, R.J. The plant beneficial effects of Metarhizium species correlate with their association with roots. Appl Microbiol Biotechnol 2014, 98, 7089–7096. [Google Scholar] [CrossRef] [PubMed]
- González-Pérez, E.; Ortega-Amaro, M.A.; Bautista, E.; Delgado-Sánchez, P.; Jiménez-Bremont, J.F. The entomopathogenic fungus Metarhizium anisopliae enhances Arabidopsis, tomato, and maize plant growth. Plant Physiol Biochem 2022, 176, 34–43. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, Y.; Wang, B. Entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae play roles of maize (Zea mays) growth promoter. Sci Rep 2022, 12, 15706. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Khan, S.A.; Kang, S.M.; Shinwari, Z.K.; Kamran, M.; Ur Rehman, S.; Kim, J.G.; Lee, I.J. Pure culture of Metarhizium anisopliae LHL07 reprograms soybean to higher growth and mitigates salt stress. World J Microbiol Biotechnol 2012, 28, 1483–1494. [Google Scholar] [CrossRef]
- Chowdhury, M.Z.H.; Mostofa, M.G.; Mim, M.F.; Haque, M.A.; Karim, M.A.; Sultana, R.; Rohman, M.M.; Bhuiyan, A.U.; Rupok, M.R.B.; Islam, S.M.N. The fungal endophyte Metarhizium anisopliae (MetA1) coordinates salt tolerance mechanisms of rice to enhance growth and yield. Plant Physiol Biochem 2024, 207, 108328. [Google Scholar] [CrossRef]
- Canassa, F.; Esteca, F.C.N.; Moral, R.A.; Meyling, N.V.; Klingen, I.; Delalibera, I. Root inoculation of strawberry with the entomopathogenic fungi Metarhizium robertsii and Beauveria bassiana reduces incidence of the twospotted spider mite and selected insect pests and plant diseases in the field. Journal of Pest Science 2020, 93, 261–274. [Google Scholar] [CrossRef]
- Sasan, R.; Bidochka, M. Antagonism of the endophytic insect pathogenic fungus Metarhizium robertsii against the bean plant pathogen Fusarium solani f. sp. phaseoli. Canadian Journal of Plant Pathology 2013, 35. [Google Scholar] [CrossRef]
- Cachapa, J.C.; Meyling, N.V.; Burow, M.; Hauser, T.P. Induction and Priming of Plant Defense by Root-Associated Insect-Pathogenic Fungi. Journal of Chemical Ecology 2021, 47, 112–122. [Google Scholar] [CrossRef]
- Gupta, R.; Keppanan, R.; Leibman-Markus, M.; Rav-David, D.; Elad, Y.; Ment, D.; Bar, M. The Entomopathogenic Fungi Metarhizium brunneum and Beauveria bassiana Promote Systemic Immunity and Confer Resistance to a Broad Range of Pests and Pathogens in Tomato. Phytopathology 2022, 112, 784–793. [Google Scholar] [CrossRef]
- St Leger, R.J.; Wang, J.B. Metarhizium: jack of all trades, master of many. Open Biol 2020, 10, 200307. [Google Scholar] [CrossRef] [PubMed]
- St Leger, R. The evolution of complex Metarhizium-insect-plant interactions. Fungal Biology 2024. [Google Scholar] [CrossRef]
- Ying, Y.; Liu, C.; He, R.; Wang, R.; Qu, L. Detection and Identification of Novel Intracellular Bacteria Hosted in Strains CBS 648.67 and CFCC 80795 of Biocontrol Fungi Metarhizium. Microbes Environ 2022, 37. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, J.F.; Rehner, S.A.; Humber, R.A. A multilocus phylogeny of the Metarhizium anisopliae lineage. Mycologia 2009, 101, 512–530. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W.; Irwin, C.A.; Janssen, K.A. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: 2001.
- Fredriksson, N.J.; Hermansson, M.; Wilén, B.M. The choice of PCR primers has great impact on assessments of bacterial community diversity and dynamics in a wastewater treatment plant. PLoS One 2013, 8, e76431. [Google Scholar] [CrossRef] [PubMed]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J Bacteriol 1991, 173, 697–703. [Google Scholar] [CrossRef]
- Gautreau, G.; Bazin, A.; Gachet, M.; Planel, R.; Burlot, L.; Dubois, M.; Perrin, A.; Médigue, C.; Calteau, A.; Cruveiller, S.; et al. PPanGGOLiN: Depicting microbial diversity via a partitioned pangenome graph. PLoS Comput Biol 2020, 16, e1007732. [Google Scholar] [CrossRef]
- Cantalapiedra, C.P.; Hernández-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. eggNOG-mapper v2: Functional Annotation, Orthology Assignments, and Domain Prediction at the Metagenomic Scale. Molecular Biology and Evolution 2021, 38, 5825–5829. [Google Scholar] [CrossRef]
- Castro-Vargas, C.; Linares-López, C.; López-Torres, A.; Wrobel, K.; Torres-Guzmán, J.C.; Hernández, G.A.; Lanz-Mendoza, H.; Contreras-Garduño, J. Methylation on RNA: A Potential Mechanism Related to Immune Priming within But Not across Generations. Front Microbiol 2017, 8, 473. [Google Scholar] [CrossRef]
- Schyns, G.; Serra, C.R.; Lapointe, T.; Pereira-Leal, J.B.; Potot, S.; Fickers, P.; Perkins, J.B.; Wyss, M.; Henriques, A.O. Genome of a Gut Strain of Bacillus subtilis. Genome Announc 2013, 1. [Google Scholar] [CrossRef]
- Brown, N.L.; Stoyanov, J.V.; Kidd, S.P.; Hobman, J.L. The MerR family of transcriptional regulators. FEMS Microbiol Rev 2003, 27, 145–163. [Google Scholar] [CrossRef] [PubMed]
- Tulin, G.; Figueroa, N.R.; Checa, S.K.; Soncini, F.C. The multifarious MerR family of transcriptional regulators. Mol Microbiol 2024, 121, 230–242. [Google Scholar] [CrossRef]
- Deng, W.; Li, C.; Xie, J. The underling mechanism of bacterial TetR/AcrR family transcriptional repressors. Cell Signal 2013, 25, 1608–1613. [Google Scholar] [CrossRef]
- Brown, S.; Meredith, T.; Swoboda, J.; Walker, S. Staphylococcus aureus and Bacillus subtilis W23 make polyribitol wall teichoic acids using different enzymatic pathways. Chem Biol 2010, 17, 1101–1110. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.; Santa Maria, J.P., Jr.; Walker, S. Wall teichoic acids of gram-positive bacteria. Annual review of microbiology 2013, 67, 313–336. [Google Scholar] [CrossRef]
- Sutton, G.; Fogel, G.B.; Abramson, B.; Brinkac, L.; Michael, T.; Liu, E.S.; Thomas, S. Horizontal transfer and evolution of wall teichoic acid gene cassettes in Bacillus subtilis. F1000Res 2021, 10, 354. [Google Scholar] [CrossRef] [PubMed]
- Aunpad, R.; Panbangred, W. Evidence for two putative holin-like peptides encoding genes of Bacillus pumilus strain WAPB4. Curr Microbiol 2012, 64, 343–348. [Google Scholar] [CrossRef]
- Frąc, M.; Hannula, S.E.; Bełka, M.; Jędryczka, M. Fungal Biodiversity and Their Role in Soil Health. Front Microbiol 2018, 9, 707. [Google Scholar] [CrossRef]
- Mares-Rodriguez, F.d.J.; Aréchiga-Carvajal, E.T.; Ruiz-Herrera Ŧ, J.; Moreno-Jiménez, M.R.; González-Herrera, S.M.; León-Ramírez, C.G.; Martínez-Roldán, A.d.J.; Rutiaga-Quiñones, O.M. A new bacterial endosymbiotic relationship in Kluyveromyces marxianus isolated from the mezcal fermentation process. Process Biochemistry 2023, 131, 133–143. [Google Scholar] [CrossRef]
- Sánchez-Alonzo, K.; Silva-Mieres, F.; Arellano-Arriagada, L.; Parra-Sepúlveda, C.; Bernasconi, H.; Smith, C.T.; Campos, V.L.; García-Cancino, A. Nutrient Deficiency Promotes the Entry of Helicobacter pylori Cells into Candida Yeast Cells. Biology (Basel) 2021, 10. [Google Scholar] [CrossRef]
- Tavakolian, A.; Heydari, S.; Siavoshi, F.; Brojeni, G.N.; Sarrafnejad, A.; Eftekhar, F.; Khormali, M. Localization of Staphylococcus inside the vacuole of Candida albicans by immunodetection and FISH. Infection, Genetics and Evolution 2019, 75, 104014. [Google Scholar] [CrossRef]
- Pawlowska, T.E.; Gaspar, M.L.; Lastovetsky, O.A.; Mondo, S.J.; Real-Ramirez, I.; Shakya, E.; Bonfante, P. Biology of Fungi and Their Bacterial Endosymbionts. Annu Rev Phytopathol 2018, 56, 289–309. [Google Scholar] [CrossRef] [PubMed]
- Arora, P.; Riyaz-Ul-Hassan, S. Endohyphal bacteria; the prokaryotic modulators of host fungal biology. Fungal Biology Reviews 2019, 33, 72–81. [Google Scholar] [CrossRef]
- Lupini, S.; Peña-Bahamonde, J.; Bonito, G.; Rodrigues, D.F. Effect of Endosymbiotic Bacteria on Fungal Resistance Toward Heavy Metals. Front Microbiol 2022, 13, 822541. [Google Scholar] [CrossRef] [PubMed]
- Partida-Martinez, L.P.; Monajembashi, S.; Greulich, K.O.; Hertweck, C. Endosymbiont-dependent host reproduction maintains bacterial-fungal mutualism. Curr Biol 2007, 17, 773–777. [Google Scholar] [CrossRef]
- Hoffman, M.T.; Gunatilaka, M.K.; Wijeratne, K.; Gunatilaka, L.; Arnold, A.E. Endohyphal bacterium enhances production of indole-3-acetic acid by a foliar fungal endophyte. PLoS One 2013, 8, e73132. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Chen, T.; Li, W.; Hong, J.; Xu, J.; Yu, Z. Endosymbiotic bacteria within the nematode-trapping fungus Arthrobotrys musiformis and their potential roles in nitrogen cycling. Front Microbiol 2024, 15, 1349447. [Google Scholar] [CrossRef]
- Lupini, S.; Nguyen, H.N.; Morales, D., 3rd; House, G.L.; Paudel, S.; Chain, P.S.G.; Rodrigues, D.F. Diversity of fungal microbiome obtained from plant rhizoplanes. Sci Total Environ 2023, 892, 164506. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Zheng, X.; Sun, Z.; Li, Y.; Deng, J.; Zhou, Y.I. Characterization of a Plant Growth-Promoting Endohyphal Bacillus subtilis in Fusarium acuminatum from Spiranthes sinensis. Pol J Microbiol 2023, 72, 29–37. [Google Scholar] [CrossRef]
- Kohlmeier, S.; Smits, T.H.; Ford, R.M.; Keel, C.; Harms, H.; Wick, L.Y. Taking the fungal highway: mobilization of pollutant-degrading bacteria by fungi. Environ Sci Technol 2005, 39, 4640–4646. [Google Scholar] [CrossRef]
- Berthold, T.; Centler, F.; Hübschmann, T.; Remer, R.; Thullner, M.; Harms, H.; Wick, L.Y. Mycelia as a focal point for horizontal gene transfer among soil bacteria. Scientific Reports 2016, 6, 36390. [Google Scholar] [CrossRef]
- Baldrian, P. The known and the unknown in soil microbial ecology. FEMS Microbiology Ecology 2019, 95. [Google Scholar] [CrossRef]
- Nan, L.; Guo, Q.; Cao, S.; Zhan, Z. Diversity of bacterium communities in saline-alkali soil in arid regions of Northwest China. BMC Microbiol 2022, 22, 11. [Google Scholar] [CrossRef] [PubMed]
- Zahid, M.S.; Hussain, M.; Song, Y.; Li, J.; Guo, D.; Li, X.; Song, S.; Wang, L.; Xu, W.; Wang, S. Root-Zone Restriction Regulates Soil Factors and Bacterial Community Assembly of Grapevine. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Lüneberg, K.; Schneider, D.; Siebe, C.; Daniel, R. Drylands soil bacterial community is affected by land use change and different irrigation practices in the Mezquital Valley, México. Scientific Reports 2018, 8, 1413. [Google Scholar] [CrossRef]
- Pineda-Castellanos, M.L.; Rodríguez-Segura, Z.; Villalobos, F.J.; Hernández, L.; Lina, L.; Nuñez-Valdez, M.E. Pathogenicity of Isolates of Serratia Marcescens towards Larvae of the Scarab Phyllophaga Blanchardi (Coleoptera). Pathogens 2015, 4, 210–228. [Google Scholar] [CrossRef]
- Enríquez, M.; Romero-López, A. Klebsiella Bacteria Isolated from the Genital Chamber of Phyllophaga obsoleta. Southwestern Entomologist 2017, 42. [Google Scholar] [CrossRef]
- M, H.; Sabtharishi, S.; Chandel, R.; Gandotra, S. Exploration of gut bacteria in white grub Anomala sp. (Coleoptera: Scarabaeidae), a major pest of vegetables and fruit trees in India. 2020.
- Desirò, A.; Salvioli, A.; Ngonkeu, E.L.; Mondo, S.J.; Epis, S.; Faccio, A.; Kaech, A.; Pawlowska, T.E.; Bonfante, P. Detection of a novel intracellular microbiome hosted in arbuscular mycorrhizal fungi. Isme j 2014, 8, 257–270. [Google Scholar] [CrossRef]
- Naito, M.; Desirò, A.; González, J.B.; Tao, G.; Morton, J.B.; Bonfante, P.; Pawlowska, T.E. ' Candidatus Moeniiplasma glomeromycotorum', an endobacterium of arbuscular mycorrhizal fungi. Int J Syst Evol Microbiol 2017, 67, 1177–1184. [Google Scholar] [CrossRef]
- Lastovetsky, O.A.; Caruso, T.; Brennan, F.P.; Wall, D.; Pylni, S.; Doyle, E. Spores of arbuscular mycorrhizal fungi host surprisingly diverse communities of endobacteria. New Phytologist 2024, 242, 1785–1797. [Google Scholar] [CrossRef]
- Cruz, A.F.; Ishii, T. Arbuscular mycorrhizal fungal spores host bacteria that affect nutrient biodynamics and biocontrol of soil-borne plant pathogens. Biol Open 2012, 1, 52–57. [Google Scholar] [CrossRef]
- Han, S.; Chen, J.; Zhao, Y.; Cai, H.; Guo, C. Bacillus subtilis HSY21 can reduce soybean root rot and inhibit the expression of genes related to the pathogenicity of Fusarium oxysporum. Pestic Biochem Physiol 2021, 178, 104916. [Google Scholar] [CrossRef] [PubMed]
- Russi, A.; Almança, M.A.K.; Schwambach, J. Bacillus subtilis strain F62 against Fusarium oxysporum and promoting plant growth in the grapevine rootstock SO4. An Acad Bras Cienc 2022, 94, e20210860. [Google Scholar] [CrossRef]
- Chertkova, E.; Kabilov, M.R.; Yaroslavtseva, O.; Polenogova, O.; Kosman, E.; Sidorenko, D.; Alikina, T.; Noskov, Y.; Krivopalov, A.; Glupov, V.V.; et al. Links between Soil Bacteriobiomes and Fungistasis toward Fungi Infecting the Colorado Potato Beetle. Microorganisms 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.P.S.; Keerthana, A.; Priya; Singh, S. K.; Rai, D.; Jaiswal, A.; Reddy, M.S.S. Exploration of culturable bacterial associates of aphids and their interactions with entomopathogens. Arch Microbiol 2024, 206, 96. [Google Scholar] [CrossRef]
- Barelli, L.; Waller, A.S.; Behie, S.W.; Bidochka, M.J. Plant microbiome analysis after Metarhizium amendment reveals increases in abundance of plant growth-promoting organisms and maintenance of disease-suppressive soil. PLoS One 2020, 15, e0231150. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, N.; Greco, C.; Drott, M.T.; Koss, M.J.; Ludwikoski, I.; Keller, N.M.; Keller, N.P. Bacterial hitchhikers derive benefits from fungal housing. Curr Biol 2022, 32, 1523–1533.e1526. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, X.; Xu, C.; Zhao, H.; Zhang, X.; Zeng, G.; Qian, Y.; Liu, R.; Guo, N.; Mi, W.; et al. Horizontal gene transfer allowed the emergence of broad host range entomopathogens. Proc Natl Acad Sci U S A 2019, 116, 7982–7989. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.W.; Tao, W.; Bedzyk, L.; Young, T.; Chen, M.; Li, L. Global gene expression profiles of Bacillus subtilis grown under anaerobic conditions. J Bacteriol 2000, 182, 4458–4465. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Confocal laser scanning micrographs of Metarhizium mycelia stained with SYTO® 9. M. robertsii ES37 (A), M. robertsii ES37-2 (B); M. guizhouuense PPH1 (C), and M. guizhouuense PPH1-1 (D). Samples B and D were overexposed to observe potential EHB. Mycelia samples were stained with SYTO® 9 and analyzed using a laser scanning confocal microscope (ZEISS LSM 700). SYTO®9, sample staining; BF, bright field; Merge, bright field and SYTO®9 fluorescence.
Figure 1.
Confocal laser scanning micrographs of Metarhizium mycelia stained with SYTO® 9. M. robertsii ES37 (A), M. robertsii ES37-2 (B); M. guizhouuense PPH1 (C), and M. guizhouuense PPH1-1 (D). Samples B and D were overexposed to observe potential EHB. Mycelia samples were stained with SYTO® 9 and analyzed using a laser scanning confocal microscope (ZEISS LSM 700). SYTO®9, sample staining; BF, bright field; Merge, bright field and SYTO®9 fluorescence.
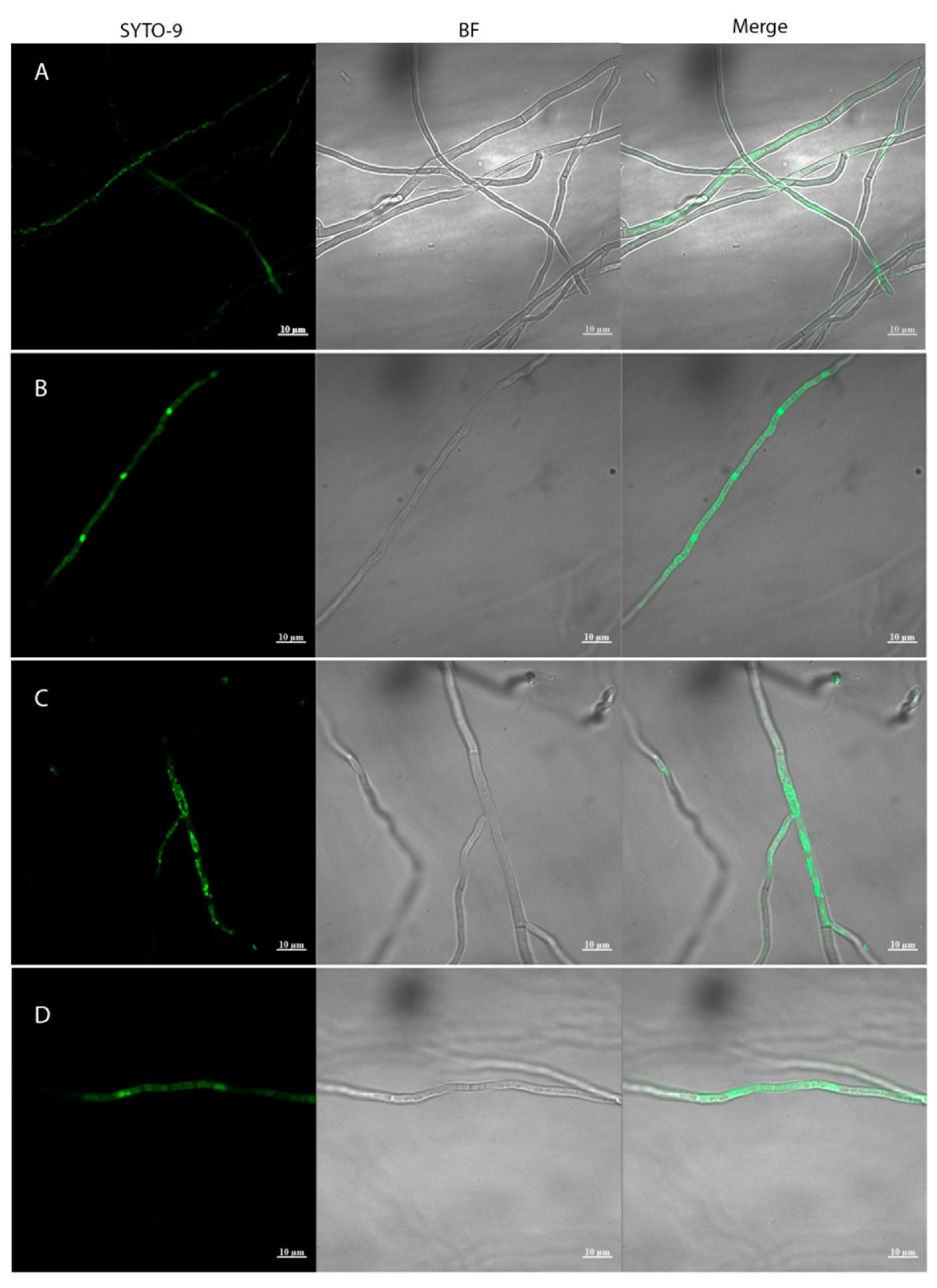
Figure 2.
The phenotypes of Metarhizium strains in minimal medium.
Figure 3.
Conidiation rates of Metarhizium strains grown in M-100 and PDA media.
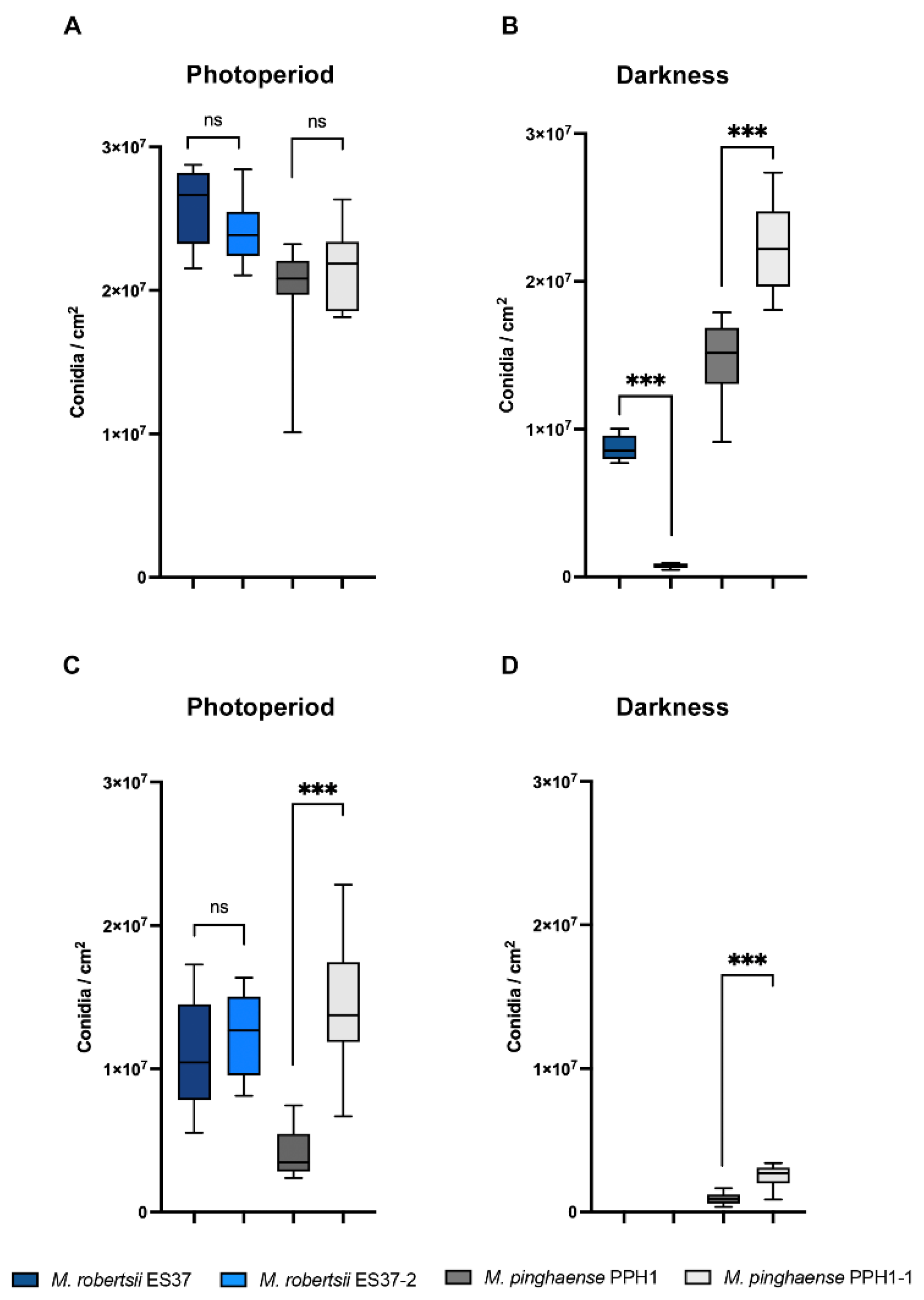
Figure 4.
Percentage germination of Metarhizium strains.
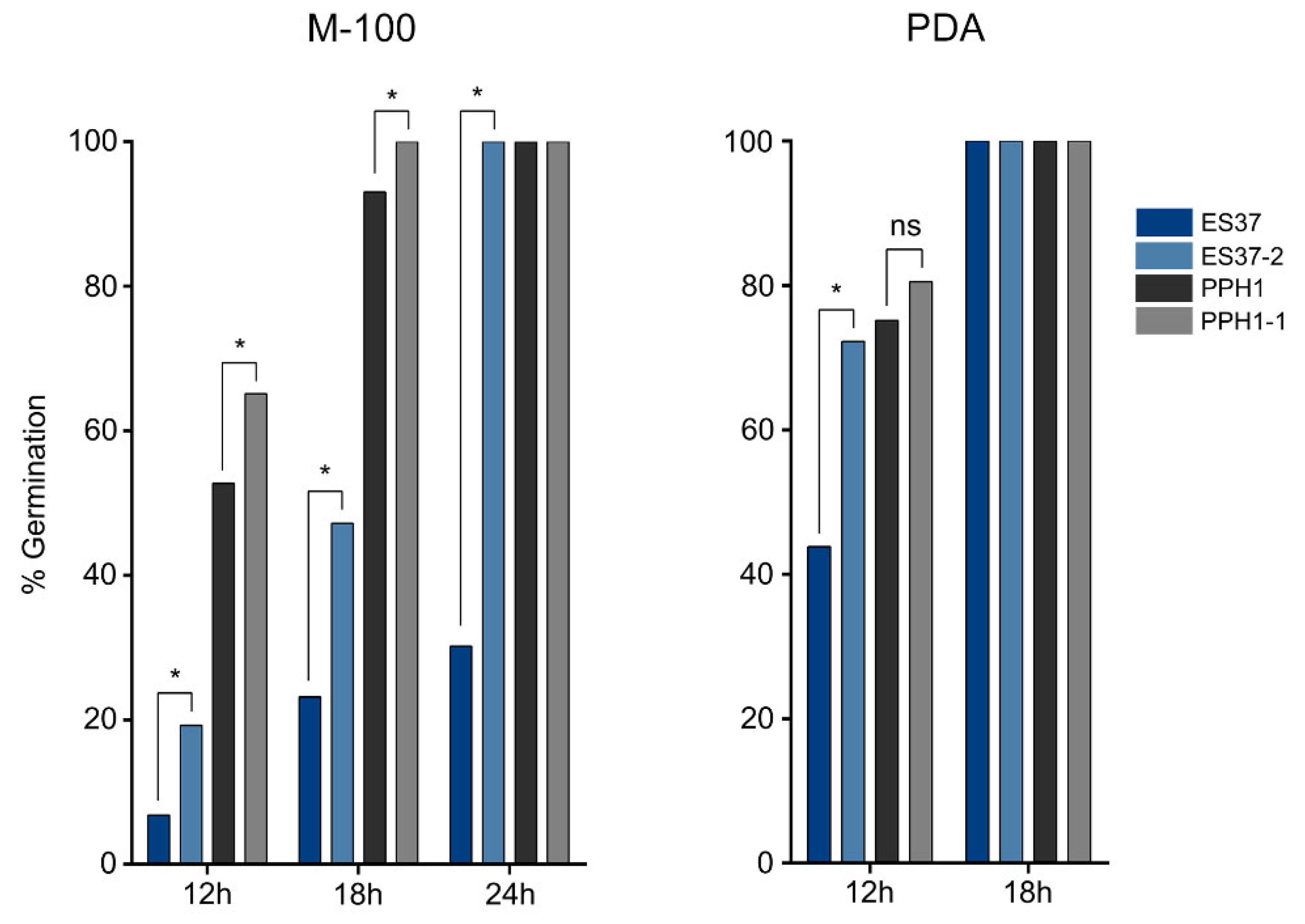
Figure 5.
Survival of G. mellonella larvae after the application of conidia from Metarhizium strains.
Figure 5.
Survival of G. mellonella larvae after the application of conidia from Metarhizium strains.
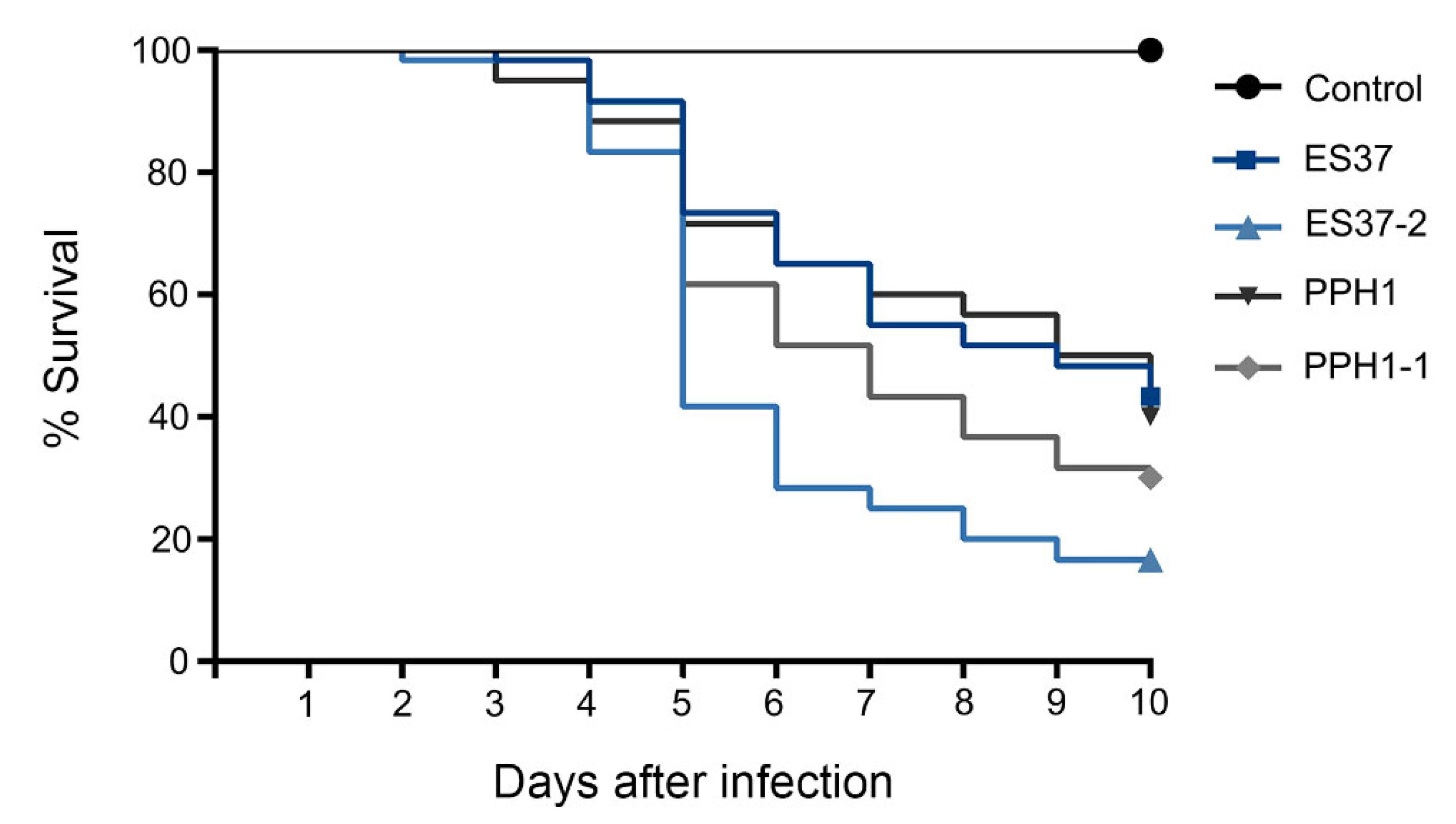
Figure 6.
Survival of T. molitor larvae after the application of different conidia concentrations of M. robertsii ES37 and M. robertsii ES37-2 strains. A) 5; B) 50; C) 100 conidia of M. robertsii ES37 and ES37-2 strains.
Figure 6.
Survival of T. molitor larvae after the application of different conidia concentrations of M. robertsii ES37 and M. robertsii ES37-2 strains. A) 5; B) 50; C) 100 conidia of M. robertsii ES37 and ES37-2 strains.
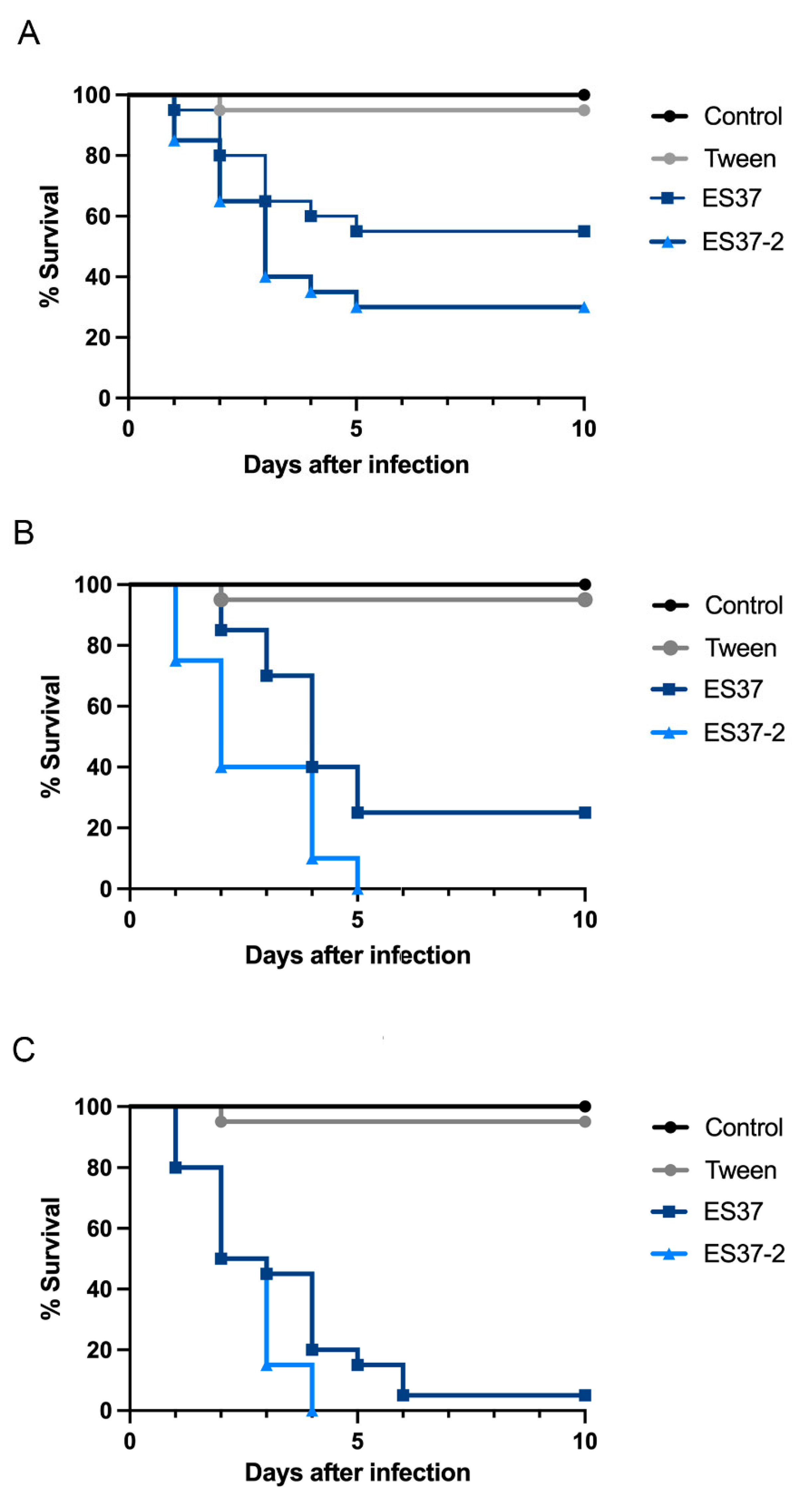
Figure 7.
Venn diagram showing the distribution of shared and unique gene clusters among Bacillus species: B. subtilis 1E, B. subtilis BSP1, and B. subtilis 168.
Figure 7.
Venn diagram showing the distribution of shared and unique gene clusters among Bacillus species: B. subtilis 1E, B. subtilis BSP1, and B. subtilis 168.
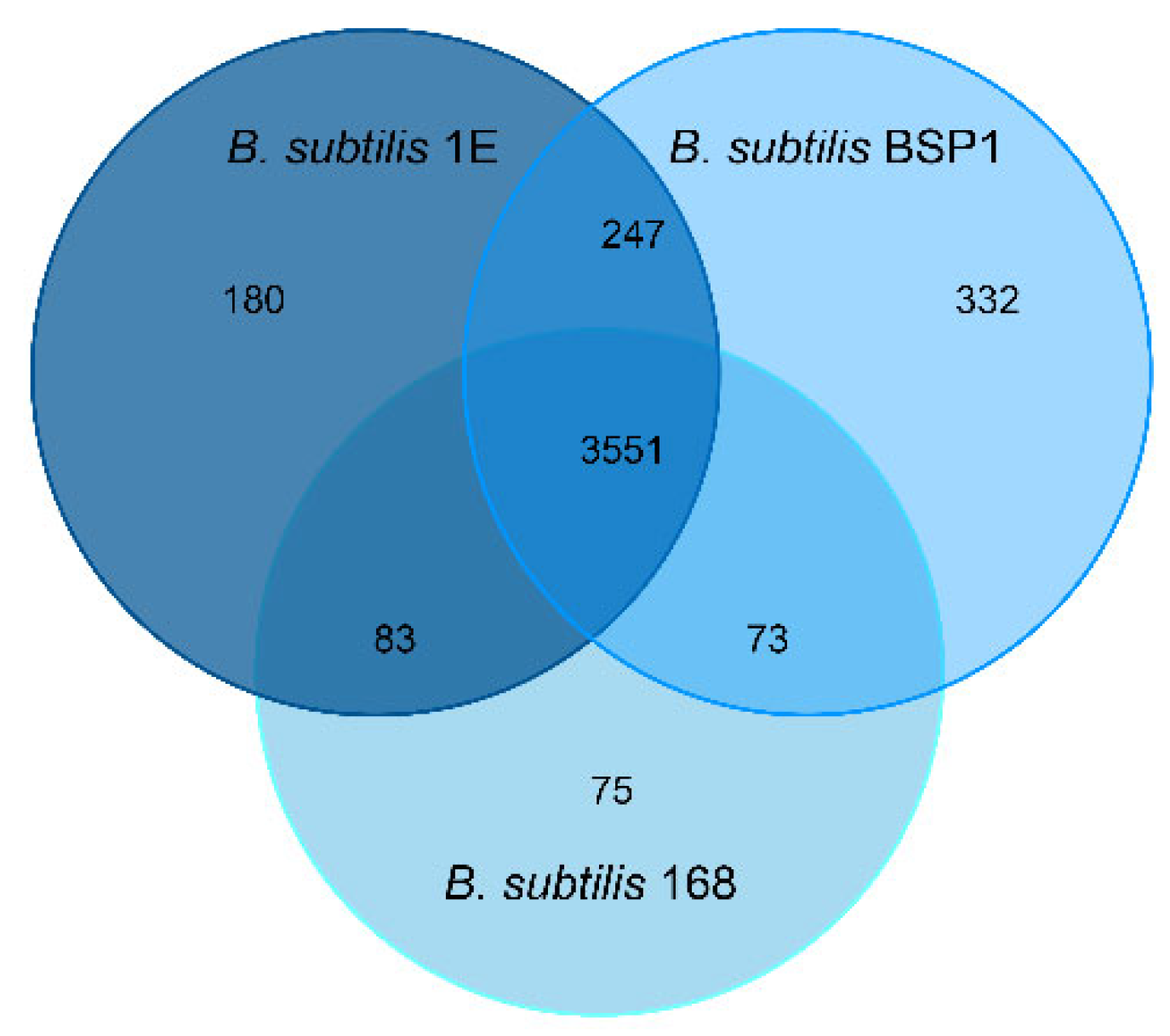
Figure 8.
Classification of annotated singleton (genes not found in other species) functions in the Clusters of Orthologous Groups (COG) of B. subtilis 1E.
Figure 8.
Classification of annotated singleton (genes not found in other species) functions in the Clusters of Orthologous Groups (COG) of B. subtilis 1E.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



