
Submitted:
29 September 2025
Posted:
30 September 2025
You are already at the latest version
Abstract
Plant-parasitic nematodes (PPNs) cause big crop losses globally. Safe and reliable methods for their durable management strategies can astutely harness various beneficial relationships among plant immune system and related microbiomes. The molecular mechanisms basic to these relations reveal wide arrays of significant roles for plant-healthy growth. This review focuses on such relations to prime and immunize plants against PPNs. It highlights molecular issues facing PPN-resistant varieties with possible genetic breeding as solutions. Few resistant plant varieties, emergence of resistant pathotypes, fitness costs, and specific settings that may hinder the resistance are exemplified issues. These issues call for optimal uses of the microbiome to bridge the gap for boosting plant immunity. In contrast, widely spread microbiomes of plant growth-promoting rhizobacteria, biological control fungi, and arbuscular mycorrhizal fungi can immunize plants against PPNs. Related plant signaling hormones and transcription factors that regulate gene expression and modulate nematode-responsive genes to ease positive/negative adaptation are presented. Relevant genome editing and other molecular techniques are discussed to attract further attention for improved microbiome’s usage. They are promising, but only under specific biotic/abiotic settings. Hence, proper abiotic/biotic factors related to systemic acquired resistance (SAR) that impact plant–microbe interactions to immunize plants against PPNs are emphasized. For PPN control, the microbiomes can be added as inoculants and/or steering the indigenous rhizosphere ones. Consequently, SAR is mediated by the accumulation of the salicylic acid and subsequent expression of pathogenesis-related genes. To activate SAR, adequate priming and induction of plant defense against PPNs would rely on closely linked and crucial factors. They include the engaged microbiome species/strains, plant genotypes, existing fauna/flora, and compatibility with other involved biologicals as well as methods and rates/concentrations of the inoculants. Thus, it is suggested herein to test the related microbiomes on a case-by-case basis to avoid erratic results. A few reports focused on root-knot nematodes, but this review offered attractive examples for the related gains in immunizing plants to other key PPN species. The end in view is to optimize the related molecular plant defence and expand the use of the microbiomes against PPN species. Achieving such goals needs to raise growers’ awareness to harness advanced strategies tackled herein to boost the roles of SAR-inducing microbes. These microbes enjoy wide spectrum efficacy, low-fitness cost, and inheritance to next generations in sustainable agriculture.
Keywords:
acquired resistance
; biotic/abiotic factors
; molecular plant defence
; yield
1. Introduction
The cost for international development that can adequately mitigate the current localized food and war crises as well as global tariffs will finally get to levels that require close scrutiny to identify the most profitable investment goals. Apparently, agricultural development will top that list where integrated plant-parasitic nematode (PPN) management is increasingly demanding further optimization [1]. Obviously, the unmistakable demand to accelerate perfecting crop production without further unhealthy and polluting nematicidal usage is calling for realizing the reformed integrated PPN management plans [2]. Developing plant resistance/immunity against numerous abiotic and biotic stresses in many economically important crops is broadly being challenged. Activating and fortifying plant immune systems rank high as benign and reliable alternative approaches to long-used and unhealthy nematicides. The danger of these latter has led to banning prominently effective but unsafe related chemicals as aldicarb [3] and carbofuran [4].
In contrast, a considerable part of plant microbiomes can employ its well-defined rhizosphere habitat via distinguished physico-chemical attributes to safely offer benefits to their companion plants. Many microbial communities could be anthropogenically harnessed to boost and enforce plants-immunity response. In addition, plants under different types of stresses may deliberately seek and choose from their rhizospheres such useful communities/microbiomes [2]; e.g. arbuscular mycorrhizal fungi (AMF) and plant growth-promoting rhizobacteria (PGPR). Surely, the plants’ ability to perceive/react to biotic and abiotic pressures is crucial for their health and productivity. Yet, the related mechanisms are still far to be fully disclosed. The complex balance of plant–microbiome interactions is not limited to different beneficial/harmful nematode groups. It also engages abundance of other microbes, their cellular composition/exposure time, and soil traits in intimate contact with plant roots. They collectively govern the extent of the plant response.
Admittedly, PPNs are the most spreading and harming soil-borne pests. They mostly cause out of sight subterranean infections and non-specific symptoms [5]. So, the related interspecific interactions in the plant rhizosphere and crop losses are frequently unknown or largely underestimated. Therefore, this review initially clarifies molecular/cellular mechanisms of economically important plants to control PPNs. Drawbacks of PPN-resistant varieties and possible genetic breeding-based solutions are presented. This review addresses chemical nematicides-alternative approaches that strengthen plants-immune system via beneficial microorganisms. It helps to molecularly clarify the extent of microbiome ability and contribution in combating nematodes infecting plants. Direct/indirect mechanisms induced by microbiome activities for general improvement of this ability against PPNs are highlighted. Because of the limited review-space, it focuses on factors that boost molecular activities of microbiomes against only PPN species especially from the standpoint of expanding their related management. So, raising stakeholders’ awareness to make these microbiomes familiar and easy to use should enhance their wider application while supplying safe control tactics and enhanced crop yields.
2. Molecular Plant Mechanisms for Combating Nematodes Infecting Crops
It is well-established that plants possess several modes of action for protecting themselves. Initially, recognition of specific PPN-molecule signatures induces innate immune reactions in the attacked plants. The common term for naming this ordinary innate or basal immune system is the primary layer or first line of defense against PPNs [6]. It can detect nematode-associated molecular patterns (NAMPs) using pattern recognition receptors (PRRs) as special protein receptors (receptor-like kinases [RLKs] and receptor-like proteins) that can act to initiate this innate immunity, e.g., against root-knot nematodes (RKNs, Meloidogyne spp.) [7]. A striking example of an NAMP is ascaroside (Asc#18) that is secreted by PPNs; e.g. the cyst nematode (CN) Heterodera glycines Ichinohe (Tylenchida: Heteroderidae) [8]. On the other, the Arabidopsis leucine-rich repeat (LRR) receptor-like kinase NILR1 (nematode-induced LRR-RLK1) acts as the PRR for this ascaroside. Activating NAMP-triggered immunity (NTI) by ascr#18, induces distinct plant defences [9]. In order to offer plant protection, Klessig et al. [10] stated that NTI comprises consequent activation of mitogen-triggered protein kinases, salicylic acid- and jasmonic acid-interceded defence signalling pathways, and defence gene operation. Although Ascr#18 is a member in a family of PPN signalling molecules, its exogenous application in low nanomolar/micromolar concentrations could offer protection against a wide spectrum of pests/pathogens. These latter included not only PPN species but also viruses, fungi, bacteria, and oomycetes [10]. For PPN control, the NTI results in generating reactive oxygen species (ROS) and secondary metabolites, cell death encompassing the PPN-migratory tunnel, and/or strengthening plant-cell walls [11]. These plant responses may slow down the early stages of nematode infection. They contribute in tangible defenses in non-host plants. However, PPNs can defeat NTI by secreting effector proteins called effector-triggered suppression (ETS) to impede the innate immune reactions in nematode-susceptible plants. Such effectors as Ha18764, RHA1B, GrVAP1, and GrCEP12, are formed by the nematodes in their esophageal glands and transferred by PPN stylet into the plant roots regarding soil PPNs [8]. Having suppressed the basal immune responses, endoparasitic sedentary nematodes become able to establish feeding sites. These sites represented by giant cells for RKNs and syncytia for CNs are essential for RKNs and CNs to develop and reproduce on their susceptible hosts [9].
Additionally, a more sophisticated defense line is owned only by PPNs-resistant plants. It is widely assumed that the genetic constitution of such plants can encode for either single dominant resistance genes (R-genes) or quantitative trait loci (QTL) to demonstrate a specific defense response [8,9]. This assumption is supported as the R-gene may engage a small gene family with thoroughly homologous copies that are jointly clustered. The R-genes act using two main modes of PPN interaction [9]. First, these R-genes in plants can specifically interact with relevant avirulence (Avr) genes in the nematode resulting in an incompatible interaction. In other words, nematode genes responsible for effectors that could be recognized by the plant resistance-gene products are called Avr genes. Obviously, PPN-resistant crop cultivars with PPN-specific resistance carry R-genes within their intracellular immune receptors. The second mode, named the guard hypothesis, as its mechanism begins when nematode effectors trigger the plant-virulence factors (protein) which stimulates R-gene [12]. Overall, virulent and adapted PPNs can circumvent pathogen-triggered immunity (PTI) in nematode-susceptible plants by effectuating PPN-effector molecules directly into the plant cells. Conversely, specific PPN effectors can be recognized by nematode-resistant plants. These R-genes encode immune receptors that contain nucleotide-binding and LRR domains. Such proteins mediate the specific detection and recognition of PPN avirulence effectors to prompt defense responses. The implied reactions involve intracellular signaling pathways in order to validate the expression of the resistant response [9,13].
Among their many functions in supporting plant growth, three plant hormones are involved in inducing immunity against various stresses, e.g. attacking pests and pathogens. These are salicylic acid (SA), jasmonic acid (JA), and ethylene (ET). Their relevant operations engage the overexpression of defense genes such as the pathogenesis-related (PR) ones. Generally, effective plant defense for biotrophic pests/pathogens is mostly based on programmed cell death in the plant in addition to combined activation of defense reactions exerted by the SA-dependent pathway leading to the subsequent expression of PR genes. Conversely, necrotrophs are faced by another set of plant-defense responses operated by JA and ET signaling. Yet, more complexities/exceptions for these biotrophs and necrotrophs were reported [14]. What added to these complications is that both SA and JA signaling can commonly crosstalk antagonistically or synergistically, based on their combined concentrations [15]. So, it was stressed that the negative interaction between JA and SA signaling can be employed by pathogens to boost plant susceptibility [15].
Tackling families of transcription factors (TFs) can give valuable insights into the genetic and transcriptional dynamics associated with PPN invasion and development. Plant TFs regulate the expression of instructions contained in its genes in response to PPN attack and growth (Table 1). Different comparative gene expression analyses are progressing with functional reviews of plant TFs and their relevance in the plant-PPN interaction [16]. Identifying differentially expressed genes allows molecular biologists to grasp exact disease mechanisms and discover potential biomarkers. Interestingly, such differential expression may even be materialized by the same host plant, Oryza sativa L. (Poales: Poaceae) infected by two different PPN species, i.e. Meloidogyne graminicola Golden & Brichfield (Tylenchida: Heteroderidae) and Hirschmanniella oryzae (Van Breda de Haan) Luc & Goodey (Tylenchida: Pratylenchidae). While rice infection by M. graminicola suppresses the local defense pathways (e.g. of salicylic acid), H. oryzae invasion causes upregulation of biotic stress-related genes early in the nematode infection stage [17]. Fortunately, breeding for M. graminicola resistance could use CRISPR/Cas9 gene editing system to mutate the susceptibility gene OsHPP04 in rice, resulting in improved resistance to the nematode, without affecting desirable agronomic traits [18]. The immune responses in the ‘transgene-free’ homozygous mutants of rice plants triggered by flg22, involving reactive oxygen species burst, defence-related genes expression and callose deposition, were raised. Such responses showed potential in developing M. graminicola-resistant crops to contribute in durable agriculture and crop protection.
Trials are ongoing to aid in unravelling and grasping molecular complexities in microbiome-plant immunity interaction. Five tomato cultivars susceptible to RKN were treated prior to the nematode inoculation with a set of chemicals closely related to hormones/processes of plant defence. The chemicals comprised inhibitors of SA or JA-mediated defense pathways, hormone generators, compounds that interfere with calcium-mediated metabolism, ROS generators and scavengers, and inhibitors of ROS generation [24]. Before treatments, it was found that SA-responsive genes tested were down-regulated in response to infection by Meloidogyne incognita (Kofoid & White) Chitwood (Tylenchida: Heteroderidae). Generally, compounds that reduced SA and/or ROS levels enhanced M. incognita infection severity, but those that minimized JA/ET levels did not affect infection rates. Such reactions could demonstrate systemic acquired resistance (SAR) where the induced resistance is specifically mediated by the accumulation of SA and subsequent expression of PR genes [13,25]. The latter authors stated that SAR is activated by the attack of biotrophs, such as PPNs, or by treatment with chemical or natural materials. Parallel to this concept, the effect of M. incognita attack on the performance of the defense enzymes endoglucanase and endochitinase, encoded by PR-2 and PR-3, respectively, changed relying on the tested days post-inoculation (dpi) with RKN-second stage juveniles (J2s) [24]. Both enzymes were inhibited in inoculated plants compared to healthy controls at 5 dpi. Also, the genes encoding glutathione peroxidase and catalase (CAT), as components of the plant-antioxidant system, were highly overexpressed. In the infected roots, the activity of superoxide dismutase, CAT, and ascorbate peroxidase, as antioxidant enzymes, was elevated too. Within the context of SAR against biotic stresses, adding ROS generators could encourage resistance against M. incognita, but materials that silenced calcium signaling boosted infection symptoms. Such results [24] confirmed that SA-mediated plant immune reactions are consistently inhibited in the early stages of M. incognita infection to the susceptible cultivars. The inhibition was linked to the stimulation of the ROS-scavenging order. On the other, JA-responsive genes were unaffected by the RKN infection. This may indirectly agree with the induced systemic resistance (ISR) as JA-responsive genes are mostly induced by the necrotrophic pathogens [14]. Yet, some studies stated that ISR can also be induced via treating with chemical/natural compounds, attacking pathogens, and beneficial microbes like AMF and PGPR [14,25]. Finally, more exploration on exact recognition and signaling pathways linked to compounds/genes required for R operations are still needed.
3. Current Problems Linked to Using R-Genes with Possible Genetic Solutions
As the spread of PPNs increases and their related losses are worse, it becomes clearer that the number of nematode-resistant plant species/cultivars is insufficient, especially in key crops. Moreover, an individual R-gene in the cluster may be employed for resistance, but multi-gene background is typical for plant R-genes [9]. For example, Mi-1 contains a limited gene cohort with seven homologous copies jointly grouped on the small arm of chromosome 6 in the tomato-genetic constitution with RKN resistance [26]. Nonetheless, various Mi genes are exist which differs from Mi-1 in specificity, functional characteristics, and genetic locations [27,28]. In RKN-resistant tomatoes, out of ten R genes dedicated for the resistance, only seven genes can function at high temperatures, i.e., above 32 °C [7]. Recently, Devran et al. [29] found a tomato line, MT12, that can offer resistance to M. incognita at 32°C soil temperature. Mapping RRKN1 to chromosome 6 using Kompetitive Allele Specific PCR markers provides a genetic resource and open novel avenues for breeding tomato varieties with stable resistance under such conditions. On the other hand, a slow decline in the effectiveness of R-gene could sometimes be recorded. A striking example is the present issue in incorporating resistance derived from plant introduction accession 88788 into almost all H. glycines-resistant soybean varieties planted in the USA. Although this CN is a key pest of the soybean plant there, its relevant resistance - encoded by a high copy number of the rhg1-b allele -has already begun to decline. Therefore, incorporating Bacillus thuringiensis (Bt) Berliner (Bacillales: Bacillaceae) delta-endotoxin (Cry14Ab) into soybean plant could boost H. glycines resistance in the genetically engineered soybean [30]. It is still desirable to identify more resistance sources with broad-spectrum resistance to H. glycines and favorable agronomic traits. So, by assessing molecular marker haplotypes at the rhg1 and Rhg4 loci and testing resistance against multiple H. glycines races, 12 soybean cultivars were found to exhibit Peking-type resistance, effective against multiple nematode races [31]. Their detection offer valuable genetic resources for breeding H. glycines-resistant soybeans.
Another problem may be linked to the gene construct itself. Tomato plants with dual genes (PjCHI-1 and CeCPI) and synthetic promoters can produce transgenic lines that possess better resistance to RKN infection and multiplication than corresponding transgenic plants with a single gene [32]. Besides, most plant resistance genes utilized so far are operating against only the endoparasitic sedentary nematodes [9]. In contrast, other categories of PPNs are economically important, but efforts exerted to develop their resistant plant cultivars/varieties are scanty or fruitless. For instance, migratory endoparasitic nematodes such as the rice root nematode Hirschmanniella oryzae can cause significant yield losses. While H. oryzae does not induce a feeding structure, it can either completely enter the root making damaging tunnels or just embed its head into the root cortex to feed on root nutrients. An estimation of rice losses due to H. oryzae infestation in Egypt was 25% in infested fields [33]. Chorismate mutase was found to be secreted by H. oryzae to inhibit SA synthesis in order to suppress defense of rice plants against H. oryzae [34].
Resistance-breaking pathotypes have been widely recorded [7,28,35,36]. For such RKN pathotypes, their populations could be sourced from populations recovered from resistant tomatoes-cultivated fields, natural virulent populations (not from resistant tomatoes fields), and avirulent populations selected for virulence in the laboratory [37]. It is apparent that the genetic events leading to the acquisition of these pathotypes or virulent populations against the Mi-gene differ according to their source; i.e. selected versus natural populations. Moreover, Molinari [38] speculated that selection pressure for virulent population could also acquire another function that enabled the nematodes to escape the host reaction, e.g., by boosting activities of antioxidant enzymes. Unfortunately, such virulent populations are spreading [7]. There are no definite or clear reasons for their spread, but they are frequently found in monoculture systems that may back the events of their selection pressure. Other factors may contribute to their occurrence such as ecological complexes engaging changes in populations of multiple nematode species [39] and temperature [9]. Unfortunately, the resistance to RKNs has frequently failed due to the heat instability or high temperature sensitivity of the tomato-resistant gene in many regions worldwide [40]. In Florida (USA), threshold soil temperature is defined where gradual decreases in RKN resistance occur for each degree above 78°F (25.6 °C). So, at 91°F (32.8 °C), tomato plants with R-gene showed full susceptibility to RKNs [41]. The latter author stressed that Mi gene-virulent populations have been identified for different RKN species which are capable of damaging and reproducing on tomato plants. To make matters worse, many RKNs undergoes obligate mitotic parthenogenesis [i.e., M. incognita, M. javanica Treub (Tylenchida: Heteroderidae), and M. arenaria Neal] which would maintain virulent strains while others, e.g. M. hapla Chitwood, M. chitwoodi Golden, O'Bannon, Santo, & Finley, and M. fallax Karsen, mostly reproduce by facultative meiotic parthenogenesis [9]. For CN, their species with facultative multiplication typically have a limited host range. Yet, their sexual reproduction improves adaptability and heterogeneity among and within PPN populations [28]. Therefore, virulent isolates could be more inducible in sexually multiplied species too.
Resistance to PPN may also involve other direct plant–PPN interactions. These may engage structure and nature of PPN-feeding stylets, their secreted compounds, and consequent plant responses to the attacking nematodes. It was recorded that the PPNs-tube orifice of the stylet operates as a molecular sieve to absorb only definite molecules during feeding on tomato roots that manifest a nematicidal Bt crystal protein. The ultrastructure of the feeding tubes showed that RKNs, contrary to CNs, could ingest larger transgenic proteins [42,43]. So, transgenic 54 kDa Cry6A and Cry5B proteins were absorbed by and adversely affected M. incognita multiplication on tomato roots [43,44]. However, resistance to CNs in roots manifesting protein from B. thuringiensis was not attained. That is because the large 54 kDa Cry6A protein could not be absorbed by Heterodera schachtii A. Schmidt (Tylenchida: Heteroderidae) due to the small orifice of the tube [43]. This shackle severely traps the use of these transgenic proteins against H. schachtii.
Admittedly, more investigations with proper tools and sophisticated techniques should be directed to strengthen plant immunity/resistance. This is especially important as additional options seem to be costly and/or require extreme care as well as comprehensive knowledge. For instance, exogenously applied SA is generally known to strengthen plant immunity against the attacking nematodes [24]. However, its dosages that were lower than optimal cannot perform its assigned role, i.e. adversely affecting RKN infection to the plant hosts. On the contrary, fairly higher ones were toxic. Meanwhile, other Solanaceous species were stimulated differently against RKN by optimal SA rates [13].
4. Beneficial Roles of Plant-Associated Microorganisms
Having said the resistance genes-related issues and defects, it is necessary to search for a complementary framework to enhance plant immunity against PPNs. This does not negate the need for further research to solve relevant issues of the aforementioned plant–nematode interactions. In addition, the fruitful role of microbiome in plant immunity requires close scrutiny to identify their best exploitation. Strikingly, a standard type of accrued soil microbiome competitive tasks is assumed to function not only against PPNs. It can frequently act against a broad range of soil-borne diseases in addition to abiotic stresses [45,46]. Therefore, many firms do not sell numerous of their products containing biocontrol agents (BCAs) that can indirectly combat PPNs, e.g. formulated bacteria or fungi, as biological nematicides. That is because such beneficial soil-borne microorganisms can strengthen plant immunity in many different ways. Hence, they are preferably addressed herein under the following subtitles.
4.1. General Benefits of These Microorganisms
Many commercial microorganisms are sold as soil conditioners, plant growth promoters (biostimulants), or plant strengtheners. Examples are several products containing Trichoderma spp. and Agrinos’ proprietary High Yield TechnologyTM commodities [47]. Addressing progress in the commercialization of biological nematicides, the latter authors confirmed that these syntheses may improve plants’ ability to withstand PPN infections. Hence, applying microbial inoculants that can diminish stresses and enhance plant growth while raising plants’ ability to withstand PPN attacks should be considered for sustainable farming. Admittedly, such microbial inoculants may realize their final goals of boosting crop yields and growth parameters via direct and indirect mechanisms (Figure 1). Furthermore, PPN management by a single species or isolate of such biologicals can possibly give effective results [48]. Yet, their implications in multi-functionality and/or favorable synergism among two or more agricultural inputs should be exploited [49,50].
4.2. Categories of Plant Microbiomes That Activate Its Immunity
Although BCAs of PPNs comprise a wide array of beneficial soil-borne micro-organisms, only definite and related categories can activate plant immunity. For example, different species of trapping fungi and predatory nematodes [51] are used against PPNs without activating plant immunity; both groups are considered predacious microorganisms. In contrast, nematode parasitic Pasteuria spp. initiate their life-cycles when the endospores passively get attached to the outer cuticle layer of their specific host nematode popu lation/species, as the individuals move through the soil [52]. Once adhered, the bacterial endospore germinates, producing a germ tube which penetrates into the host, to infect the nematode body by multiplying bacterial cells. Interestingly, it was postulated that such microorganisms attaching to the infective stages of endoparasitic nematodes activate defence mechanisms in plants after they enter the roots [53]. Apart from such putative effects, BCAs that can colonize roots and stimulate plant immunity to PPNs could generally be divided into three main and widespread categories. These are AMF, biological control fungi (BCF), and PGPR [54]. As most PPN species are soil parasites, plant roots colonized by useful microorganisms of these categories have been recorded to generate SAR-like responses against the nematodes [13].
4.3. Molecular Mechanisms of the Microbiomes to Activate Plant Immunity
Kour et al. [55] presented the related role of AMF as remarkable example. They reported numerous ways that AMF can use to prime and immunize plants against biotic/abiotic stresses. Initially, significant plant priming could be obtained by improving one or more of the following: the stability of soil aggregates, its water holding capacity and root hydraulic conductivity, plant-water uptake and its root exudation, capacity of root surface area, and root development with consequent efficient uptake of nutrients/water. Then, AMF immunize plants against biotic stresses, such as PPN infection, via optimizing hormonal interplay and osmolyte augmentation, activates signaling transduction and phenol generation, and supports ionic homeostasis and impacts of PS-I and PS-II (two distinct protein complexes involved in Photosystem-I and Photosystem-II, respectively). These effects usually boost the plants’ ability to withstand biotic/abiotic stresses while improving its growth parameters and yields [55].
Although AMF are obligate root symbionts, they behave like pathogenic fungi at first. So, they begin with secreting defence suppressors to realize thriving colonization on roots. Thus, AMF are faced by SA-regulated defences, which show steady likeness between receptors and processes engaged in symbiosis and immunity molecular signalling [56,57]. Hence, SA-dependent PR-1 gene was upregulated only 3 days after AMF-treatment suggesting that the inoculated tomato plants had firstly recognized AMF as pathogens [58]. Thereafter, AMF could repress this reaction as a routine system so that AMF realize good colonization. Eventually, mycorrhiza-colonized plants are primed against different pathogens like PPNs. This status, named mycorrhiza-induced resistance (MIR), is well-documented for prompt and strong reaction to varying biotic attacks [13,59,60]. For PPNs, MIR could prime or immunize plants especially via upregulation of PR- and ACO-genes [58]. Such priming/immunizing of plants against RKNs was based on SAR [24]. Avowedly, RKNs not only lead a key group of PPNs that are dangerous to economic crops but also rank high among the endoparasitic sedentary nematodes. Nonetheless, even for other PPN groups with different infection cycle, MIR proved effectiveness. For instance, Vos et al. [61] found that AMF could systemically reduce infection by not only M. incognita but also the migratory endoparasitic nematode Pratylenchus penetrans (Cobb) Filipjev and Schuurmans Stekhoven (Tylenchida: Pratylenchidae). The overall reduction was 45% in the case of M. incognita and 87% for P. penetrans.
4.4. Examples of Microbiomes That Molecularly Activate Plant Immunity
Many fungal and bacterial nematicides that usually constitute collective communities are ranking high among microbiome groups for use in sustainable agriculture. They can function singly but to optimize their benefits, they can mostly be combined with other plant-beneficial microbes and/or other compatible inputs [46,47,62]. Their combination in integrated nematode management (INM) plans is intended to synergistically or additively act in integrated pest management (IPM) schemes [62]. Species of fungal genera in the rhizosphere soil or/and onto plant roots can share in setting promising microbial communities for PPN management; e.g. Trichoderma, Pochonia, Purpureocillium, Dactylellina, Catenaria, Actylellina, Arthrobotrys, Monacrosporium, Aspergillus, and Hirsutella [63]. Definitely, many endophytic fungi of the genera Trichoderma, Fusarium, Alternaria, Pochonia, Purpureocillium, and Acremonium are reported to colonize plant roots and boost plant defense against various stresses [64]. Ghahremani et al. [65] proved that Pochonia chlamydosporia (Goddard) Zare & W. Gams (Hypocreales: Clavicipitaceae) could induce plant-dependent systemic resistance to M. incognita while Könker et al. [66] very recently reported synergistic support by P. chlamydosporia to systemic defense of Phacelia tanacetifolia Benth. (Boraginales: Boraginaceae) against M. hapla Chitwood. Additionally, some Trichoderma spp. are well-known mycoparasites, but several species can antagonize PPNs and other plant pests, offering them as promising multifunctional BCAs [46,67]. In this context, PR-1, PR-5 and ACO genes were over-expressed 5 days after M. incognita inoculation into tomato plants pre-treated with T. harzianum Rifai (Hypocreales: Hypocreaceae) strains T908 and T908-5. However, the overexpression was not systemically transferred to leaves, but confined to roots [68]. In this case, it was assumed that the T. harzianum strains primed plants for SAR, yet, the entire pathway of SAR was not expressed until the subsequent M. incognita attack [13]. In contrast, inducing related SA-dependent defences was needed to restrict the first stages of M. incognita attack, when another T. harzianum strain (T-78) was utilized to colonize tomato roots [69]. Changes in the related hormones and enzymatic activities at dpi in these studies [68,69] and elsewhere [24] would largely answer open questions about how rapidly host plant–microbiome relationships can adapt for PPN management. Some studies [68,69] did not detect JA signalling for Trichoderma-induced resistance (TIR) against the early stages of M. incognita development, possibly because M. incognita is a biotroph (not a necrotroph). Also, JA-responsive genes were unaffected by the M. incognita infection [24]. Nonetheless, another study [69] stated that JA signalling was activated for this resistance at later stages of nematode infection. This claim was backed by the deficiency of TIR in mutants with imperfect JA biosynthesis (def-1) but a few weeks after M. incognita inoculation. Comparative analysis of the genomes from different Trichoderma spp. showed that their relevant impact was enforced by the existence of genes that enable effective competition and broad environmental opportunism [70]. Differently, endophytic non-pathogenic Fusarium oxysporum Schlecht. emend. Snyder & Hansen (Hypocreales: Nectriaceae) strains proved ability for controlling PPNs. These strains were effective against a wider range of individual PPN species infecting banana roots. Indeed, van Dessel et al. [71], Waweru et al. [72], and Mendoza and Sikora [73] proved such an effective ability against Helicotylenchus multicinctus (Cobb) Golden (Tylenchida: Hoplolaimidae), Pratylenchus goodeyi (Cobb) Sher & Allen (Tylenchida: Pratylenchidae), and Radopholus similis (Cobb) Thorne (Tylenchida: Pratylenchidae), respectively. Therefore, Kisaakye et al. [74] stressed the multiple benefit of F. oxysporum for PPN management, i.e. it can optimize host resistance to various pests/diseases and improve banana productivity. On the other hand, although Purpureocillium lilacinum (Thom) Luangsa-ard et al. (Hypocreales: Ophiocord-ycipitaceae), can directly parasitize PPNs, it has the ability to boost plant resistance. This could be achieved via activating defense-related enzymes such as peroxidase (POD), CAT, and phenylalanine ammonia-lyase (PAL); fortifying plant defense against pathogens. Thus, P. lilacinum can promote plant growth and upregulate SA biosynthesis genes necessary to build SAR in plants against PPNs [75].
Likewise, PGPR represent another group that can function as environment friendly alternative to unhealthy chemical nematicides. They can suppress diseases either directly via synthesizing pathogen-antagonizing compounds, or indirectly through triggering plant immune responses. Clearly, their general merit is to contribute in stimulating plant growth especially by inhibiting plant-pathogenic organisms. Jiao et al. [76] detailed underlying and diverse mechanisms of rhizobacteria for suppressing diseases. One or several mechanisms in combination can be associated with specific host plants. Their well-known direct mechanisms in biocontrol scenarios are suppressing pathogens via generating different anti-pathogen compounds, e.g. antibiotics, bacteriocins metabolites, antimicrobial peptides, toxins, and enzymes. Indirectly, PGPR can boost plant stress tolerance via synthesized microbe-plant signal compounds. Such compounds comprise phytohormones and specific signal chemicals for plant-microbe communications [77]. Also, bacterial species of genera like Pseudomonas and Bacillus are recorded as BCAs that trigger systemic resistance in the plants’ rhizosphere [78].
Consolidated usage of PGPR and other bio-nematicides/chemicals have been commercially and experimentally practiced. The plant activator acibenzolar-S-methyl (ASM) showed better persistence behavior in the tomato rhizosphere as well as more root uptake systemic translocation ability after its combination with certain Bacillus PGPR strains [79]. Furthermore, combined usage of the mutualistic endophyte F. oxysporum strain 162 and Bacillus firmus Bredemann & Werner (Bacillales: Bacillaceae) was the most effective treatment, among other combinations, in controlling R. similis on banana (86.2% reduction in nematode population density), followed by B. firmus alone (63.7%). Mendoza and Sikora [73] attributed such high efficacy to the compatibility of the two BCAs and the capacity of F. oxysporum to colonize banana roots. Also, they attributed the better R. similis control in banana to the combined usage of antagonists with different modes of action. Definitely, the combination enabled targeting different R. similis stages during the infection process [73]. Additionally, various microbial groups contained in a commercial formulation (Myco) were more effective in limiting RKN damage than a formulation containing the sole AMF [5]. For explanatory reconciliation, the exact effect of such beneficial biologicals on PPNs may depend on the involved species/strains of both the hosts and the microbes as well as on the related type of mechanism, compatibility of the engaged biologicals, methods and rates/concentrations of the inoculants [5,55,80].
The stable recognition that enzymes as CAT, PAL, polyphenol oxidase, superoxide dismutase, POD, proteinase, b-1,3-glucanase ascorbate, chitinase, and lipoxygenase participate in plant resistance mechanism by generating related chemicals as phytoalexins and phenolic compounds [78] is being stretched by further mechanism. Rhizobia are basically known for their mutualistic relation with plants where they offer the host legume a good service by fixing atmospheric nitrogen. Yet, another economic aid of rhizobia has been recently reported. Rhizobial invasion of host roots could also trigger an elevated systemic resistance in the host plant [81]. The related mechanism needs to be further explored as it contributes to plant health by inhibiting a broad range of pathogens. It can lead to developing biotechnological devices for sustainable crop production and optimizing the systemic protective impact provided by rhizobia.
5. Upgrading Microbiome Role against PPN Species
Beneficial roles of microbiomes against PPNs are not yet widely practiced in conventional agriculture, but recent results help to develop their optimum effectiveness. Advancing their efficacy will eventually lead toward their expanded field applications against not only PPNs but probably also other soil-borne pests. Ambitiously conceived paradigm, research efforts should provide sophisticated tools to unravel the related molecular mechanisms empowered by unbounded enthusiasm for relevant biotechnology. They should exploit the extraordinary potential biologicals offered as environmentally safe alternatives to chemicals. So, this is a timely opportunity for a review of our current knowledge, existing gaps, and the challenges ahead in order to upgrade microbiome role against PPN species. Beneficial microbes possess a wide array of strategies to target both motile and sedentary PPNs life stages [82] and should be fully exploited. However, these strategies are so diverse; their details are beyond the scope of the current review. Therefore, the focus herein is to exploit and improve emerging strategies related to the microbiome's ability to prime and boost plant defence against PPNs. Yet, as these microbiomes can finally act as plant health promoters and BCAs against serious PPNs, their role should be astutely expanded for large-scale application.
5.1. Grasp of Basic Factors Governing Composition and Activity of Microbiomes
The composition and activity of microbiomes are shaped by dynamic and multifarious interactions of ecological factors, host-derived constituents, and microbial interplay [83,84,85]. Environmental assessments especially those related to soil properties as soil type, temperature, pH, moisture, mulching/organic matter, and salinity are good indicators of microbiome impact and diversity [85]. Likewise, biotic factors governing composition and activity of microbiomes should be absorbed to manipulate their improved contribution in plant defence/health. Root bacterial profile was examined using sequencing of the bacterial 16S gene across different H. glycines-susceptible and resistant soybean genotypes. The change in the microbial community under H. glycines infection indicated that this profile of susceptible and resistant soybean roots and their soil samples encompassed different bacterial communities though slightly overlapped [83]. This study confirmed that the host genotype had significant effect on the diversity of the root microbiome under H. glycines pressure in the greenhouse. The authors speculated that such differences may potentially explain the impact of beneficial bacteria or/and secondary effects relevant to H. glycines resistance. Hanif et al. [85] gave examples to confirm that the microbiome composition varies notably due to varied plant species and/or native soils. Clearly, diverse microbial populations may exist even in closely located plants of phylogenetically distant species. Moreover, distinct microbial abundances have been linked to specific plant genotypes, e.g. chromosomal substitutions in wheat varieties could affect rhizosphere microflora [86]. Furthermore, the stage of plant development [87] and genetic modifications like introducing specific genes [88] could change microbial communities, as shown in Arabidopsis thaliana (L.) Heynh. (Brassicales: Brassicaceae). On the other hand, plant exudates like organic acids, phenolic compounds, amino acids, and sugars as well as volatile compounds significantly impact microbial colonization and diversity [85]. In contrast, microbial metabolites contain compounds that may enhance the benefits of applying biocontrol strategies [89,90,91]. To sum up, shifts in nematode population densities occur directly/indirectly by the impact of their hosts and/or enemies [1,78,92]. So, grasping the above-mentioned terms like NAMPs, PRRs, and RLKs in the context of these related basics for plant immunity can help to leverage microbiome community composition and activity. These fundamentals should act as backgrounds for the transformational technologies needed to adopt upgraded role for the microbiome in PPN control.
5.2. Manipulation of the Plant Microbiome Against PPNs
5.2.1. Consideration of Biotic and Abiotic Factors in the Settings
Having grasped fundamental factors that rule composition and activity of microbiomes, careful screening of microbial strains under controlled conditions should comply with such basics before microbial inoculation. Usually, microbial strains/species that do well in lab assays undergo greenhouse tests followed by field estimations. Contrary to lab and greenhouse tests, such field evaluations may show variable results with inconsistent microbial efficacy; indicating dire need to optimize settings (biotic/abiotic factors). That is because it is quite possible that competition of inoculants with native fauna or/and flora, suboptimal strain selection, and ecological variables can degrade inoculant performance against PPNs [93,94]. Therefore, microbial formulations of inoculants should consider both niche compatibility and cell viability for sustained colonization and perfect microbial function [84,92,94]. For instance, B. amyloliquefaciens strain FZB42 should be set but early in an adapted niche in order to trigger effective plant defenses [95].
Emerging techniques like encapsulation, surfactant addition, seed coating, gel, and nanotechnology can enhance adherence to root surfaces, reduce biotic/abiotic stresses, and/or be inexpensive [85,96]. Using 5% solution of Arabic gum could improve both adhesive capacity and distribution of Bacillus species on the outer parts of the treated strawberry roots just before transplanting [97]. Also, Ajijah et al. [98] stated that optimizing inoculant functions via proper biofilms on plant roots can generally help in thriving plants. They advocated that these biofilms can boost plant protection from not only PPNs but also other pathogens by improved root-colonization and competition with pathogenic microorganisms, stimulating SAR/ISR, and producing antimicrobials effectively. Consequent boosted activities may also include inducing signal substrate production, managing phyto-hormone levels, enhancing protein generation, and regulating gene expression [82,98]. Eventually, more research is needed to evaluate the reliability of these techniques in defending plants against PPNs at various agricultural scenarios under actual field conditions. That is presumably because SAR is not an inherent microbial feature by itself, but rather relies on the microbe’s interplay with the planted genotype, targeted PPN species/race, and other biotic/abiotic settings.
5.2.2. Examining the Exact Interactions for Components of the Microbial Consortia
Several advantages of using microbial consortia rather than single-strain inoculants can generally be offered to boost plant growth and defence against PPNs under various field settings [1]. These are: (1) multiple activities of the consortia along the course of the plant-growing season; (2) ability to function against more than one stage in the nematode life cycle; (3) diverse mechanisms to control PPN species/races; (4) various ecological niches of the used consortia may boost consistency in biocontrol activities over a wide profile of soil conditions; and (5) potential to encompass different microbial species/races that negatively affect more than one PPN species/race simultaneously, thus improving the spectrum of the used inoculants [99]. The latter authors gave numerous related practical examples. To this end, improving multi-functionality and stability of combined inoculants is better harnessed via integrating metabolic complementarity with other useful traits. For example, this integration can allow for functional variation, composition, and redundancy in addition to getting systematical inoculation impacts on beneficiary soils and/or plants [78,100]. Abd-Elgawad [1] stressed that microbial consumption of various resources in dynamic metabolic strategies is more likely to achieve not only merged but also harmonized combination. This is especially important as time-related variation in activities among microbial communities in/on the plant roots and the rhizosphere can bring about synergistic or additive effects. So, soil amendments combined with different Trichoderma spp. caused higher reduction in M. incognita population levels than those obtained by single ones [101]. So, several studies [83,84,85,86,87,88] stressed the importance of tailoring consortia to specific ecological conditions and plant genotypes/developmental stage to improve their efficacy. Ultimately, the beneficial and well-synthesized microbiome should be harnessed to reliably work in multiple useful directions and for a longer period of time.
On the contrary, antagonism between BCAs in the microbial consortia or product through competition and antibiosis may reduce their activities. Single treatments as seedling drenches by Burkholderia cepacia strains Bc-2 and Bc-F and of Trichoderma virens strain Gl-3 significantly reduced numbers of M. incognita eggs + J2 per gm bell pepper root. However, these nematode parameters - eggs + J2 – were not significantly affected from combining these treatments [102]. Furthermore, a single BCA species/strain may be more effective against PPNs than two other merged BCAs. A single fungal strain (P. lilacinum MR20) could outperform a mixture of two plant-growth-promoting bacterial strains (Pseudomonas fluorescens MR12 and Serratia marcescens MR25) in minimizing M. incognita population levels on tomato plants [103]. Also, efficacy of an individual species/strain in a microbial consortium may greatly vary even in some commercial products that are documented to act as a good plant-defense biostimulant against PPNs. For instance, a single component (B. subtilis) in a commercial product referred to as Myco increased M. incognita infection indicators at any tested cell density, however, the whole microbial consortium of Myco proved to activate plant immune response and prime tomato plants against RKNs [5]. The authors advocated that difference in microbial performance may depend on the used species/strains as well as methods and doses of the application. Therefore, Myco proved to be reliable for inducing resistance against RKNs but only in a dose-dependent manner [104].
On the opposite direction, specific microbiota naturally linked to PPNs may protect them against microbial antagonists [105]. Recently, Trejo-Meléndez and Contreras-Garduño [106] underscored the essential functions of some PPN-linked microbial communities in defeating ecological hurdles facing the nematodes. Their roles may include aiding in exploiting the plant by suppressing or evading plant defenses [106]. Such mixed results in the role of microbial communities should be considered in manipulating PPNs-infested soil. Microbial inoculants should be well-tailored to improve the synthesized microbiome for priming and defending plants against PPNs. Products containing such live organisms are handled carefully as they are more sensitive to biotic and abiotic stresses than those with only bioactive compounds. Such non-living products require less stringent handling; e.g. the product does not need refrigeration.
Ultimately, the exact interactions among inoculant components or with biotic/abiotic factors in intimate contact with the inocula are preferably done on a case-by-case basis to avoid negative or erratic outputs. For instance, the fitness impacts of numerous AMF symbionts remain context-dependent and can change by different biotic/abiotic factors, e.g. new host backgrounds or existing fauna/flora which intricate predictions of their long-term dynamics. Otherwise, an approach that explicitly incorporates context-dependent factors in SAR array is direly needed. It enhances the predictability of SAR induction under definite field conditions.
5.2.3. Leveraging Plant Microbiomes for SAR Induction in Advanced Strategies
Common practices as cover cropping, crop rotation, soil mulching, applying biologicals/their bioactive compounds, and organic amendments should be upgraded in parallel to recent advances in grasping the tri-trophic interactions. Such interactions (embracing host plants, PPN species, and natural enemies) would be leveraged to boost proliferation of useful microbial taxa and/or suppressing PPNs within comprehensive strategies [1,107]. Furthermore, techniques to exploit microbially induced SAR could be based on not only exogenously added inputs to the required crops but also steering the native rhizosphere microbiome within such strategies. Wang and Li [108] suggested certain crop rotation to build up positive soil legacies. These latter can steer soil microbiomes towards ones that benefit crop growth, e.g., by harbouring a large fraction of biota able to induce ISR/SAR. Such soil legacies could be set by previous crops via not only rotated crops but also cover crops [109]. Also, incorporating soil amendments can serve plant microbiomes in such comprehensive and sophisticated strategies. For example, added cow manure [containing 0.65% total nitrogen, 21.2 carbon to nitrogen (C/N) ratio] and poultry manure (0.72% total nitrogen, 20.1 C/N ratio) to two Egyptian farms of El-Ismailia governorate resulted in almost zero counts of RKNs in loamy sand soils whereas RKNs could thrive in ten other farms. Higher levels of recorded organic matter and microbial counts, e.g. recorded Trichoderma spp., in the two farms than in the other ones were likely responsible for RKN suppression [110]. Differently, using species within definite group of BCAs to simultaneously control pests/pathogens related to different taxa could be involved; e.g. entomopathogenic nematodes (EPNs) against both insect and nematode pests [111,112]. In such cases, advanced PCR-based techniques could differentiate closely related EPN species to distinguish their specific phenotypes; e.g. host specificity, persistence, and habitat adaptation [113]. Although cell-free supernatant generated by the EPN-symbiotic bacterial culture of few species showed inhibitory effects on RKNs [89], dynamics of such interactions remain largely elusive for many other species. So, the merits of such techniques should accurately uncover how molecular processes are involved in such nematode control and plant strengthening under various settings. The expected results would boost the chances of superior PPN control. Following the same line of thinking, Han et al. [114] proposed effective, eco-friendly crop protection strategies. The strategies implement multidimensional management of PPNs and other pests. They adjusted spatial dimensions for the interactive impacts of soil-crop-pest-natural enemy networks on pest control. Meanwhile, other related dimensions deal with time, i.e. pest interactions over the whole crop season. As microbiome-related enhancing plant immunity is engaged, adopting green IPM tactics like organic farming is fortified. They aimed to enhance microbial diversity and abundance to improve ecosystem resilience and plant health; gaining ecological benefits too.
On top of, integrating innovations to common practices can develop superior safe and reliable PPN control including relationships between plant microbiome and immunity. For example, useful top associations like root exudates or microbial metabolites, enhanced symbiosis, and plant-high responses to this symbiosis via plant breeding and targeted genome editing could be employed. They can selectively employ useful microbiomes to enhance favorable performance of the plant defence [85,109]. Thus, recent calls for designing new microbial systems based on safe microbiome engineering should be considered. The call based on setting synthetic microbial consortia that can prosper across various ecological conditions is known as Intraspecies Cross Environmental (ICE) system. It merges microbial strains with complementary traits in plant roots to optimize nutrient cycling and boost plant health/resistance [115,116]. Another Combinatorial CRISPR Array-Guided Engineering (CRAGE) system can employ Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) technology to engineer microbial genomes for definite tasks that immunize plants and back plant health. Surely, relevant gains in emerging biotechnologies as omics, bioinformatics, and high-throughput next-generation sequencing methods could incorporate and express endogenous and heterologous genes in plants and relevant microbiomes to upgrade their role in immunizing plants against PPNs. Ibrahim et al. [117] suggested to clone and overexpress the genes in charge of the PPN control task from the target and corresponding agents. They exemplified Paecilomyces javanicus (Friedrichs & Bally) A.H.S. Br. & G. Sm. (Eurotiales: Thermoascaceae) to benefit plant protection efforts via fortifying its immune response against PPN infection. Also, computer software named CRISPR-PN2 can help in preparing high-throughput gene editing at the given scale [118]. This editing may aid in boosting required traits in plants that do not have adequate PPN-resistance sources [119]. In other words, manipulating genes engaged in useful traits as PPN resistance can lead to microbial strains with optimized capabilities [120]. The methods can also facilitate the study of precise gene function in microbial communities. Absolutely, they pave the way for more reliable plant/related microbiome engineering strategies [120,121]. To cope with expected issues, it was recently affirmed that such techniques would be necessary devices for effective PPN control via crop genomics that comply with climate resilient agriculture [122].
5.3. Widening Microbiome Role Against Further Important PPN Species
Current researches on related microbiome activities are mostly focused against endoparasitic sedentary nematode species. Such activities should be stretched against other key PPN species. For instance, SA biosynthesis genes needed to induce SAR in plants against PPNs were recorded to be upregulated by the fungus P. lilacinum [75]. So, this fungus at 108 conidia ml−1 could consistently suppress populations of the rice white tip nematode, Aphelenchoides besseyi Christie (Tylenchida: Aphelenchoididae) [123], but further and related research against this and other PPN species is needed. Vicente et al. [124] speculated that a specific group of Ophiostomatales fungi (genera Leptographium, Ophiostoma, and Graphilbum) possess so adaptive reaction with the pinewood nematode Bursaphelenchus xylophilus Steiner & Buhrer (Nickle) (Aphelenchida: Parasitaphelenchidae) that they can significantly diminish the nematode-population levels. Their results for the functional role of fungal communities inhabiting B. xylophilus-infected trees can lead to identifying key points in the nematode life cycle to disrupt the disease cycle via more natural alternatives. Exploring molecular adaptation of such specific fungi and other bacterial endophytes [125] associated with Pinus species to optimize their biocontrol potential against B. xylophilus is suggested. Of special interest, Kim et al. [126] examined the potential of chosen pine endophytic bacteria to induce resistance against B. xylophilus. Their study could identify bacterial strains that triggered defense responses in the infected pine trees through their ability to induce the expression of the PR10 gene; a marker of plant defense. Out of 92 bacterial strains, only 15 ones were identified to significantly enhance PR10 expression in pine callus cultures. On the other, the interaction of Ditylenchus dipsaci Kuhn (Tylenchida: Anguinidae) and Fusarium oxysporum on garlic led to less severe disease in the infected bulb and lower D. dipsaci-population densities than when the plants were inoculated with the nematode alone [127]. The authors contemplated that suppression of D. dipsaci populations by the fungus may be attributed to the formations of physical barriers like a mycelial mat coating the basal plate, defence responses (e.g. cell wall thickening), or competition for available nutrients. Also, colonization by B. firmus I-1582 on roots of Arabidopsis thaliana could significantly suppress H. schachtii reproduction, pathogenicity, and development over two generations [128]. While I-1582 was attracted by A. thaliana root exudates and colonized the roots via a strictly pH-dependent development, it could protect the plant from infestation by Heterodera schachtii. Ahmed et al. [129] proved that B. cereus Frankland & Frankland (Bacillales: Bacillaceae) strain B48 significantly reduced H. avenae Wollenweber (Tylenchida: Heteroderidae)-white female development with enhanced wheat root length of infected plants relative to the untreated check. The authors confirmed the possible strain’s ability of inducing systemic resistance in plant system for protection from various soil-borne PPNs. In another study, the effect of AMF Glomus etunicatum (Glomerales: Glomeraceae) W.N. Becker & Gerd. on H. glycines resulted in a 28% reduction in nematode females in the soybean root system of mycorrhizal plants compared to untreated roots, suggesting that AMF immunized the host plant against H. glycines [130]. On testing biological induction of resistance to Globodera tabacum solanacearum Lownsbery & Lownsbery (Tylenchida: Heteroderidae) in flue-cured tobacco [Nicotiana tabacum L. (Solanales: Solanaceae)], Parkunan et al. [131] found that two Bacillus strains among four rhizobacterial combinations - had the most consistent effect in decreasing the nematode cysts under greenhouse and field conditions. Other bacterial species of the same genus showed also similar adverse effects on Rotylenchulus reniformis Linford & Oliveira (Tylenchida: Hoplolaimidae) infecting soybean plants [132]. Additional species like B. thuringiensis were reported to protect plants from Xiphinema index Thorne & Allen (Dorylaimida: Longidoridae) infecting grapevine [133]. For other PPN and bacterial genera, the endophytic bacterium Serratia ureilytica Bhadra et al. (Enterobacterales: Yersiniaceae) could negatively affect Nacobbus aberrans Thorne (Tylenchida: Pratylenchidae) on Chili plants. This bacterium significantly reduced the number of galls, egg masses, eggs, and reproduction factor of the nematode on the plant roots, compared to the control where S. ureilytica was not applied [134]. Mycorrhiza-induced resistance has been shown to be efficient against other important nematode species as Radopholus similis and Pratylenchus coffeae (Zimmermann) Filipjev & Schuurmans Stekhoven (Tylenchida: Pratylenchidae) infecting banana [135]. Therefore, AMF are being developed for commercial applications; e.g. Ectovit®, Symbivit®, and Rhodvit® [136].
Notwithstanding the utility of microbiomes against such key PPN species, the molecular – genetic/epigenetic - mechanisms by which the related plant immunity is triggered need further research to leverage and expand their usage. Priority research areas can merge plant microbiome profiling with breeding techniques to assemble superior microbiomes against PPNs. Other areas are to optimize advantageous effects of root exudates on the microbiomes. Also, research is needed to examine the impact of specific inoculants/treatments such as immune hormones especially SA, JA, and ET on the composition of microbiomes and how microbiomes shift the plant immune system. Eventually, understanding the related ecology, biology, mode of action, and interactions of these microbiomes with other agricultural inputs is still needed for future application against key PPN species in sustainable agriculture.
6. Microbiome Merits Serve to Raise Awareness and Increase Its Adoption
Currently, the consensus is that there is no single PPN control method that has all the relevant merits, that is, effective, inclusive, economical, and safe to non-targets [137]. Beneficial microbiome that raises plant immunity and offers defence priming is not an exception regarding this fact, i.e. the microbiome occasionally provides only partial plant protection. So, its integration with other PPN control method(s) is preferable. In contrast, the microbiome can contribute in immunizing plants for a long term and plant priming may persist even after removal of the priming agent [138]. That is because plants have a non-adaptive immune system that is set by molecular shifts. Epigenetic modification implied by induced resistance priming can enable plants to acquire memory of stressful conditions. Consequently, it leads to change in gene expression. Such changes are heritable to the plant progeny resulting in maintenance of priming up to the next-generation [139]. Tiwari et al. [138] assured that such a microbiome-merit in plant immunological memory should be added to other related ones. These include its wide-spectrum impact, low-fitness cost, and long-lasting durability which make it attractive for sustainable crop protection. On the contrary, plant (R) genes encode immune receptor proteins to induce defence responses that are mostly linked to a reduction in growth and yield, known as fitness costs [140].
Commonly, nematode suppression in soil can offer a quick and non-specific defensive line for other pathogens too. Yet, plant specific suppression of PPNs needs time to react and it possesses a memory to a specific invading pathogen [13,106]. Therefore, to get more benefits of soil microbiome and/or upgrade plant defence, activation of beneficial microbial community interfered with the specific PPN infection may be initiated or shortened by putting in the “microbial or material kick start”. This start could be via mulching the soil [107], adding organic amendments as supplementary food sources for the microbiome [141], applying useful endophytic microorganisms sole [142] or combined with a plant extract or other materials [1,58]. Moreover, pre-treating plant roots with favorable microbial communities or advantageous hormone generators could activate plant defence [24].
Limitations in expanding such processes and consequent merits are mostly stemmed from the lack of awareness of efficient implementation methods by stakeholders, especially resource-poor farmers. Also, limited knowledge of these beneficial microbiomes by extension personnel who advise growers usually aggravates the matter. To optimize household food security and concurrently improve their crop yields, related sustainable, low-input and environmentally-friendly PPN management strategies have to be simply applied and adapted to suit their needs. This concept has been discussed in the frame of conserving and enhancing biocontrol of PPNs [141] and theoretically presented to improve the adoption of green IPM tactics [114]. Nonetheless, thus far, it has not been comprehensively attempted in earnest. It could be achieved by means of innovative and applied research by appropriately trained nematologists and biologists. They can train, assist and guide extension officers and farmers to enhance the quality and quantity of their produce by minimizing the adverse effect of the PPNs in their crops. Therefore, relevant efforts for enlightening stakeholders will facilitate expanding the benefits of ISR and SAR-inducing microbes to extension agents and farmers. Moreover, improving commercial syntheses and formulations of related products and their application efficacy should be a continuous process. They should facilitate incorporating the beneficial microbiome into IPM plans. As complementary tools for PPN management, such plant microbiome and immunity interactions should be exploited for very efficient uses that maintain the environment, don’t pollute, and secure the quality/quantity of crops in an inexpensive manner. These activities will continually broaden the boundaries of sophisticated approaches and scalability of pathogen/pest management while boosting the crop health and yield.
7. Conclusions
Beneficial relationships between plant microbiome and molecular immunity can provide promising alternatives for safe and reliable PPN control strategies. Plants subjected to PPN--induced stresses usually seek and select from the environments beneficial microorganisms. However, steering the indigenous rhizosphere/soil microbiomes and/or adding relevant inoculants should be followed to optimize PPN management. Within this context, it is critical to properly activate ISR and/or SAR via the related factors. These latter comprise the involved strains of the microbiome, plant-genetic constitution, manipulating the existing fauna/flora, compatibility with the other engaged abiotic elements as well as techniques and doses/concentrations of the inoculants. Therefore, further research is required on such factors on a case-by-case basis to avoid unpredictable results. Sustainable IPM approaches should seek to boost the role of microbiome rhizosphere against various pests/pathogens. These would include immunizing plants against not only endoparasitic sedentary nematodes – RKNs and CNs - but also other key PPN species. Therefore, system-level agricultural production management is required to induce beneficial microbial shifts in the plant rhizosphere and soil for controlling PPNs. These would change the microbial processes occurring mostly at the microscales, molecular biology scales. However, harnessing such changes can have fruitful large-scale consequences on the crop yields at field and greenhouse levels. So, sufficient mechanistic grasping is needed to know how multifarious decisions at these large-scales can model these context-dependent molecular processes of beneficial microbiome activities in the long term. Research priorities are to genetically seek and upgrade favorable effects. They can include plant microbiome profiling-based genetic breeding techniques to synthesize superior safe and effective microbiomes for PPN control. Also, further research is needed to know how specific inoculants/treatments such as immune hormones can alter the composition of microbiomes, and how microbiome exactly shifts the plant immune system. Moreover, impact of plant-genetic constitution on shaping microbiome composition and relevant pathways adopted by the host plant should be explored. Gaining such knowledge will support the large-scale leverage of relevant microbiome merits. These latter involve its broad spectrum effectiveness, low-fitness cost, and inheritance to subsequent generations in sustainable agriculture. On the other hand, improved formulations and applications of products containing such microbiomes should be expanded. Meanwhile, agricultural extension workers should simplify relevant information for their familiar adoption and usage by stakeholders.
Funding
This research was funded by NRC In-House project No. 13050112 entitled: Pesticide alternatives against soil borne pathogens and pests attacking economically significant export crops.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
The author acknowledges the support in part of this study by NRC In-House project No. 13050112 entitled: Pesticide alternatives against soilborne pathogens and pests attacking economically significant export crops, and US-Egypt Project cycle 22 (No. 47725).
Conflicts of Interest
The author declares no conflicts of interest.
References
- Abd-Elgawad, M.M.M. Integrated nematode management strategies: Optimization of combined nematicidal and multi-functional inputs. Plants 2025, 14, 1004. [CrossRef]
- Sikora, R.A.; Desaeger, J.; Molendijk, L.P.G. (Eds.) Integrated Nematode Management: State-of-the-Art and Visions for the Future; CABI: Wallingford, UK, 2022; 498p.
- EPA. Aldicarb; Cancellation Order for Amendments to Terminate Usesfed; Regist; EPA: Washington, DC, USA, 2012.
- EPA. Carbofuran; Product Cancellation Orderfed; Regist; EPA: Washington, DC, USA, 2009.
- Molinari, S.; Leonetti, P. Resistance to plant parasites in tomato is induced by soil enrichment with specific bacterial and fungal rhizosphere microbiome. Int. J. Mol. Sci. 2023, 24, 15416. [CrossRef]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D. Hormone crosstalk in plant disease and defense: More than just jasmonate-salicylate antagonism. Ann. Rev. Phytopathol. 2011, 49, 317–343. [CrossRef]
- El-Sappah, A.H.; Islam, M.M.; El-Awady, H.H.; Yan, S.; Qi, S.; Liu, J.; Cheng, G.-T.; Liang, Y. Tomato natural resistance genes in controlling the root-knot nematode. Genes 2019, 10, 925. [CrossRef]
- Zheng, Q.; Putker, V.; Goverse, A. Molecular and cellular mechanisms involved in host-specific resistance to cyst nematodes in crops. Front. Plant Sci. 2021, 12, 641582. [CrossRef]
- Abd-Elgawad, M.M.M. Understanding molecular plant–nematode interactions to develop alternative approaches for nematode control. Plants 2022, 11, 2141. [CrossRef]
- Klessig, D.F.; Manohar, M.; Baby, S.; Koch, A.; Danquah, W.B.; Luna, E.; Park, H.J.; Kolkman, J.M.; Turgeon, B.G.; Nelson, R. 1009 Nematode ascaroside enhances resistance in a broad spectrum of plant–pathogen systems. J. Phytopathol. 2019, 167, 1010 265-272.
- Sato, K.; Kadota, Y.; Shirasu, K. Plant immune responses to parasitic nematodes. Front. Plant Sci. 2019, 10, 1165. [CrossRef]
- Dangl, J.L.; Jones, J.D. Plant pathogens and integrated defence responses to infection. Nature 2001, 411, 826–833.
- Molinari, S. Can the plant immune system be activated against animal parasites such as nematodes? Nematology 2020, 22, 481–492. [CrossRef]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205-227. [CrossRef]
- Mur, L.A.J.; Kenton, P.; Atzorn, R.; Miersch, O.; Wasternack, C. The outcomes of concentration-specific interactions between salicylate and jasmonate signaling include synergy, antagonism, and oxidative stress leading to cell death. Plant Physiol. 2006, 140, 249-262.
- Domínguez-Figueroa, J.; Gόmez-Rojas, A.; Escobar, C. Functional studies of plant transcription factors and their relevance in the plant root-knot nematode interaction. Front. Plant Sci. 2024, 15, 1370532. [CrossRef]
- Kyndt, T.; Denil, S.; Haegeman, A.; Trooskens, G.; Bauters, L.; Van Criekinge, W.; et al. Transcriptional reprogramming by root knot and migratory nematode infection in rice. New Phytol. 2012, 196, 887–900. [CrossRef]
- Huang, Q.; Lin, B.; Cao, Y.; Zhang, Y.; Song, H.; Huang, C.; Sun, T.; Long, C.; Liao, J.; Zhuo, K. CRISPR/Cas9-mediated mutagenesis of the susceptibility gene OsHPP04 in rice confers enhanced resistance to rice root-knot nematode. Front. Plant Sci. 2023, 14, 1134653. [CrossRef]
- Grunewald, W.; Karimi, M.; Wieczorek, K.; Van de Cappelle, E.; Wischnitzki, E.; Grundler, F.; Inzé, D.; Beeckman, T.; Gheysen, G. A role for AtWRKY23 in feeding site establishment of plant-parasitic nematodes. Plant Physiol. 2008, 148, 358–368. [CrossRef]
- Willig, J.J.; Guarneri, N.; van Loon, T.; Wahyuni, S..; Astudillo-Estévez, I.E.; Xu, L.; Willemsen, V.; Goverse, A.; Sterken, M.G.; Lozano-Torres, J.L.; Bakker, J.; Smant, G. Transcription factor WOX11 modulates tolerance to cyst nematodes via adventitious lateral root formation. Plant Physiol. 2024, 195, 799-811. [CrossRef]
- Zhao, J.; Huang, K.; Liu, R; Lai, Y.; Abad, P.; Favery, B.; Jian, H.; Ling, J.; Li, Y.; Yang, Y.; Xie, B.; Quentin, M.; Mao, Z. The root-knot nematode effector Mi2G02 hijacks a host plant trihelix transcription factor to promote nematode parasitism. Plant Commun. 2024, 5(2), 100723. [CrossRef]
- Chinnapandi, B.; Bucki, P.; Fitoussi, N.; Kolomiets, M.; Borrego, E.; Braun-Miyara, S. Tomato SlWRKY3 acts as a positive regulator for resistance against the root-knot nematode Meloidogyne javanica by activating lipids and hormone-mediated defense-signaling pathways. Plant Signaling Behav. 2019, 14, 1601951. [CrossRef]
- Cabrera, J.; Diaz-Manzano, F.E.; Saı́nchez, M.; Rosso, M.N.; Melillo, T.; Goh, T.; et al. A role for LATERAL ORGAN BOUNDARIES-DOMAIN 16 during the interaction Arabidopsis-Meloidogyne spp. provides a molecular link between lateral root and root-knot nematode feeding site development. New Phytol. 2014, 203, 632–645. [CrossRef]
- Molinari, S.; Farano, A.C.; Leonetti, P. Root-knot nematode early infection suppresses immune response and elicits the antioxidant system in tomato. Int. J. Mol. Sci. 2024, 25, 12602. [CrossRef]
- Llorens, E.; González-Hernández, A.I.; Scalschi, L.; Fernández-Crespo, E.; Camañes, G.; Vicedo, B.; García-Agustín, P. Priming mediated stress and cross-stress tolerance in plants: Concepts and opportunities. In Priming-mediated stress and cross-stress tolerance in crop plants; Hossain, M.A., Liu, F., Burritt, D. J., Fujita, M., Huang, B., Eds; Academic Press: Amsterdam, The Netherlands, 2020; pp. 1-20. [CrossRef]
- Seah, S.; Telleen, A.C.; Williamson, V.M. Introgressed and endogenous Mi-1 gene clusters in tomato differ by complex rearrangements in flanking sequences and show sequence exchange and diversifying selection among homologues. Theoret. Appl. Gen. 2007, 114, 1289–1302. [CrossRef]
- Ammiraju, J.S.S.; Veremis, J.C.; Huang, X.; Roberts, P.A.; Kaloshian, I. The heat-stable, root-knot nematode resistance gene Mi-9 from Lycopersicum peruvianum is localized on the short arm of chromosome 6. Theoret. Appl. Gen. 2003, 106, 478–484. [CrossRef]
- Molinari, S. Resistance and virulence in plant-nematode interactions. In Nematodes; Boeri, F., Chung, J.A., Eds.; Nova Science Publisher, Inc.: New York, NY, USA, 2012; pp. 59–82.
- Devran, Z.; Özalp, T.; Studholme, D.J.; Tör, M. Mapping of the gene in tomato conferring resistance to root-knot nematodes at high soil temperature. Front. Plant Sci. 2023, 14, 1267399. [CrossRef]
- Kahn, T.W.; Duck, N.B.; McCarville, M.T.A. Bacillus thuringiensis Cry protein controls soybean cyst nematode in transgenic soybean plants. Nat. Commun. 2021, 12, 3380. [CrossRef]
- Lian, Y.; Yuan, M.; Wei, H.; Li, J.; Ding, B.; Wang, J.; Lu, W.; Koch, G. Identification of resistant sources from Glycine max against soybean cyst nematode. Front. Plant Sci. 2023, 14, 1143676. [CrossRef]
- Chen, Y.L.; He, Y.; Hsiao, T.T.; Wang, C.J.; Tian, Z.; Yeh, K.W. Pyramiding taro cystatin and fungal chitinase genes driven by a synthetic promoter enhances resistance in tomato to root-knot nematode Meloidogyne incognita. Plant Sci. 2015, 231, 74–81. [CrossRef]
- Youssef, M.M.A.; Eissa, M.F.M. The rice root nematode, Hirschmanniella oryzae, its identification, economic importance and control measures in Egypt: a review. Arch. Phytopathol. Pl. Protect. 2014, 47, 2340–2351. [CrossRef]
- Mantelin, S.; Thorpe, P.; Jones, J.T. Suppression of plant defences by plant-parasitic nematodes. Adv. Botanical Res. 2015, 73, 325-337. [CrossRef]
- Janssen, R.; Bakker, J.; Gommers, F. Selection of virulent and avirulent lines of Globodera rostochiensis for the H1 resistance gene in Solanum tuberosum ssp. andigena CPC 1673. Rev. Nématol. 1990, 13, 265–268.
- Castagnone-Sereno, P. Genetic variability of nematodes: A threat to the durability of plant resistance genes? Euphytica 2002, 124, 193–199.
- Xu, J.; Narabu, T.; Mizukubo, T.; Hibi, T. A molecular marker correlated with selected virulence against the tomato resistance gene Mi in Meloidogyne incognita, M. javanica and M. arenaria. Phytopathology 2001, 91, 377–382.
- Molinari, S. Antioxidant enzymes in (a)virulent populations of root-knot nematodes. Nematology 2009, 11, 689–697. [CrossRef]
- Abd-Elgawad, M.M.M. Disease complexes involving multiple nematodes. In Nematode Disease Complexes in Agricultural Crops; Khan, M.R., Ed.; CABI, Wallingford, UK, 2025; pp. 219-242. [CrossRef]
- Abd-Elgawad, M.M.M. Optimizing biological control agents for controlling nematodes of tomato in Egypt. Egypt. J. Biol. Pest Cont. 2020, 30, 58. [CrossRef]
- Noling, J.W. Nematode management in tomatoes, peppers, and eggplant. University of Florida, IFAS, publication Series no. ENY-032, USA, p 16.
- Hussey, R.S.; Mims, C.W. Ultrastructure of esophageal glands and their secretory granules in the root-knot nematode Meloidogyne incognita. Protoplasma 1990, 156, 9-18. [CrossRef]
- Li, X.Q.; Wei, J.Z.; Tan, A.; Aroian, R.V. Resistance to root-knot nematode in tomato roots expressing a nematicidal Bacillus thuringiensis crystal protein. Plant Biotechnol. J. 2007, 5, 455-464. [CrossRef]
- Li, X.-Q.; Tan, A.; Voegtline, M.; Bekele, S.; Chen, C.-S.; Aroian, R.V. Expression of Cry5B protein from Bacillus thuringiensis in plant roots confers resistance to root-knot nematode. Biol. Control 2008, 47, 97-102. [CrossRef]
- Urwin, P.E.; Mcpherson, M.J.; Atkinson, H.J. Enhanced transgenic plant resistance to nematodes by dual proteinase inhibitor constructs. Planta 1998, 204, 472-479. [CrossRef]
- Guzmán-Guzmán, P.; Kumar, A.; de los Santos-Villalobos, S.; Parra-Cota, F.I.; Orozco-Mosqueda, M.d.C.; Fadiji, A.E.; Hyder, S.; Babalola, O.O.; Santoyo, G. Trichoderma species: Our best fungal allies in the biocontrol of plant diseases - A review. Plants 2023, 12, 432. [CrossRef]
- Wilson, M.J.; Jackson, T.A. Progress in the commercialization of bionematicides. BioControl 2013, 58, 715–722. [CrossRef]
- Abd-Elgawad, M.M.M. Upgrading strategies for managing nematode pests on profitable crops. Plants 2024, 13, 1558. [CrossRef]
- Saikia, D.; Bhagawati, B. Management of Meloidogyne incognita and Rhizoctonia solani complex on okra through integration of Trichoderma viride, Glomus fasciculatum and mustard cake. Indian J. Nematol. 2014, 44, 139–143.
- Youssef, M.M.A.; El-Nagdi, W.M.A.; Abd-El-Khair, H.; Elkelany, U.S.; Abd-Elgawad, M.M.M.; Dawood, M.G. Is use of two Trichoderma species sole or combined with a plant extract effective for the biocontrol Meloidogyne incognita on potato and soil microorganisms diversity? Pakistan J. Nematol. 2023, 41, 153–164. [CrossRef]
- Abd-Elgawad, M.M.M. Biological control agents of plant-parasitic nematodes: A review. Egypt. J. Biol. Pest Cont. 2016, 26(2), 423-429. https://www.cabidigitallibrary.org/doi/pdf/10.5555/20163278720.
- Abd-Elgawad, M.M.M. Pasteuria species for nematodes management in organic farms. In Sustainable Management of Nematodes in Agriculture, Vol.2: Role of Microbes-Assisted Strategies. Sustainability in Plant and Crop Protection, vol 19; Chaudhary, K.K., Meghvansi, M.K., Siddiqui, S., Eds; Springer International Publishing: Cham, Switzerland, 2024; pp. 265-296. [CrossRef]
- Topalović, O.; Heuer, H. Plant-nematode interactions assisted by microbes in the rhizosphere. Curr. Issues Mol. Biol. 2019, 30, 75-88. [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced systemic resistance by beneficial microbes. Ann. Rev. Phytopathol. 2014, 52, 347-375. [CrossRef]
- Kour, S.; Sharma, N.; Kumar, D.; Ali, M.; Sharma, R.; Parkirti, P.; Vikram, V.; Ohri, P. Role of beneficial microorganisms in vegetable crop production and stress tolerance. In Growth Regulation and Quality Improvement of Vegetable Crops; Ahammed, G.J., Zhou, J., Eds; Springer: Singapore. [CrossRef]
- Paszkowski, U. Mutualism and parasitism: the yin and yang of plant symbioses. Curr. Opin. Pl Biol. 2006, 8, 1-10. [CrossRef]
- Zipfel, C.; Oldoryd, G.E.D. Plant signalling in symbiosis and immunity. Nature 2017, 543, 328-336. [CrossRef]
- Molinari, S.; Leonetti, P. Bio-control agents activate plant immune response and prime susceptible tomato against root-knot nematodes. PLoS ONE 2019, 14, e0213230. [CrossRef]
- Pozo,M.J; Azcón-Aguilar, C. Unraveling mycorrhiza-induced resistance. Curr. Opin. Pl Biol. 2007, 10, 393-398. [CrossRef]
- Conrath, U.; Beckers, G.J.M.; Langenbach, C.J.G.; Jaskiewicz, M.R. Priming for enhanced defense. Ann. Rev. Phytopathol. 2015, 53, 97-119. [CrossRef]
- Vos, C.M.; Tesfahun, A.N.; Panis, B.; De Waele, D.; Elsen, A. Arbuscular mycorrhizal fungi induce systemic resistance in tomato against the sedentary nematode Meloidogyne incognita and the migratory nematode Pratylenchus penetrans. Appl. Soil Ecol. 2012, 61, 1-6. [CrossRef]
- Abd-Elgawad, M.M.M.; Askary, T.H. Fungal and bacterial nematicides in integrated nematode management strategies. Egypt. J. Biol. Pest Cont. 2018, 28, 74. [CrossRef]
- Abd-Elgawad, M.M.M. Optimizing safe approaches to manage plant-parasitic nematodes. Plants 2021, 10, 1911. [CrossRef]
- Schouten, A. Mechanisms involved in nematode control by endophytic fungi. Annu. Rev. Phytopathol. 2016, 54, 121–142. [CrossRef]
- Ghahremani, Z.; Escudero, N.; Saus, E.; Gabaldón, T.; Sorribas, F.J. Pochonia chlamydosporia induces plant-dependent systemic resistance to Meloidogyne incognita. Front. Plant Sci. 2019, 10, 945. [CrossRef]
- Könker, J.; Zenker, S.; Meierhenrich, A.; Patel, A.; Dietz, K-J. Pochonia chlamydosporia synergistically supports systemic plant defense response in Phacelia tanacetifolia against Meloidogyne hapla. Front. Plant Sci. 2025, 15, 1497575. [CrossRef]
- Elshahawy, I.; Saied, N.; Abd-El-Kareem, F.; Abd-Elgawad, M. Enhanced activity of Trichoderma asperellum introduced in solarized soil and its implications on the integrated control of strawberry-black root rot. Heliyon 2024, 10, 1016. [CrossRef]
- Leonetti, P.; Zonno, M.C.; Molinari, S.; Altomare, C. Induction of SA-signaling pathway and ethylene biosynthesis in Trichoderma harzianum-treated tomato plants after infection of the root-knot nematode Meloidogyne incognita. Pl. Cell Rep. 2017, 36, 621-631. [CrossRef]
- Martínez-Medina, A.; Fernandez, I.; Lok, G.B.; Pozo, M.J.; Pieterse, C.M.J.; Van Wees, S.C.M. Shifting from priming of salicylic acid- to jasmonic acid-regulated defences by Trichoderma protects tomato against the root knot nematode Meloidogyne incognita. New Phytol. 2017, 213, 1363-1377. [CrossRef]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Mukherjee, P.K.; Zeilinger, S.; Grigoriev, I.V.; Kubicek, C.P. Trichoderma: the genomics of opportunistic success. Nature Rev. 2011, 9, 749-759. [CrossRef]
- Van Dessel, P.; Coyne, D.; Dubois, T.; De Waele, D.; Franco, J. In vitro nematicidal effect of endophytic Fusarium oxysporum against Radopholus similis, Pratylenchus goodeyi and Helicotylenchus multicinctus. Nematropica 2011, 41, 154-160.
- Waweru, B.W.; Losenge, T.; Kahangi, E.M.; Dubois, T.; Coyne, D. Potential biological control of lesion nematodes on banana using Kenyan strains of endophytic Fusarium oxysporum. Nematology 2013, 15, 101–107. [CrossRef]
- Mendoza, A.R.; Sikora, R.A. Biological control of Radopholus similis in banana by combined application of the mutualistic endophyte Fusarium oxysporum strain 162, the egg pathogen Paecilomyces lilacinus strain 251 and the antagonistic bacteria Bacillus firmus. BioControl 2009, 54, 263–272. [CrossRef]
- Kisaakye, J.; Fourie, H.; Haukeland, S.; Kisitu, J.; Nakimera, S.; Cortada, L.; Subramanian, S.; Coyne, D. Endophytic non-pathogenic Fusarium oxysporum-derived dual benefit for nematode management and improved banana (Musa spp.) productivity. Agriculture 2022, 12, 125. [CrossRef]
- Rigobelo, E.C.; Nicodemo, D.; Babalola, O.O.; Desoignies, N. Purpureocillium lilacinum as an agent of nematode control and plant growth-promoting fungi. Agronomy 2024, 14, 1225. [CrossRef]
- Jiao, X.; Takishita, Y.; Zhou, G.; Smith, D.L. Plant associated rhizobacteria for biocontrol and plant growth enhancement. Front. Plant Sci. 2021, 12, 634796. [CrossRef]
- Lyu, D.; Backer, R.; Subramanian, S.; Smith, D. Phytomicrobiome coordination signals hold potential for climate change-resilient agriculture. Front. Plant Sci. 2020, 11, 634. [CrossRef]
- Yadav, S.P.; Sharma, C.; Pathak, P.; Kanaujia, A.; Saxena, M.J.; Kalra, A. Management of phyto-parasitic nematodes using bacteria and fungi and their consortia as biocontrol agents. Environ. Sci.: Adv. 2025, 4, 335–354. [CrossRef]
- Myresiotis, C.K.; Vryzas, Z.; Papadopoulu-Mourkidou, E. Enhanced root uptake of acibenzolar-S-methyl (ASM) by tomato plants inoculated with selected Bacillus plant growth-promoting rhizobacteria (PGPR). Appl. Soil Ecol. 2014, 77, 26–33. [CrossRef]
- Abd-Elgawad, M.M.M. Bacillus species: Prospects and applications for root-knot disease management. In Microbial biostimulants: Biorational pesticides for management of plant pathogens; Ahmad, F., Pandey, R., Eds.; Apple Academic Press: New Jersey, USA, 2025; pp. 79-105. https://appleacademicpress.com/microbial-biostimulants-biorational-pesticides-for-management-of-plant-pathogens/9781774916247.
- Tonelli, M.L.; Figueredo, M.S.; Rodríguez, J.; Fabra, A.; Ibañez, F. Induced systemic resistance-like responses elicited by rhizobia. Pl. Soil 2020, 448, 1–14. [CrossRef]
- Pires, D.; Vicente, C.S.L.; Menéndez, E.; Faria, J.M.S.; Rusinque, L.; Camacho, M.J.; Inácio, M.L. The fight against plant-parasitic nematodes: Current status of bacterial and fungal biocontrol agents. Pathogens 2022, 11, 1178. [CrossRef]
- Tran, D.T.; Mitchum, M.G.; Zhang, S.; Wallace, J.G. Li, Z. Soybean microbiome composition and the impact of host plant resistance. Front. Plant Sci. 2024, 14, 1326882. [CrossRef]
- Abd-Elgawad, M.M.M. Exploiting plant-phytonematode interactions to upgrade safe and effective nematode control. Life 2022, 12, 1916. [CrossRef]
- Hanif, M.S.; Tayyab, M.; Baillo, E.H.; Islam, M.M.; Islam, W.; Li, X. Plant microbiome technology for sustainable agriculture. Front. Microbiol. 2024, 15, 1500260. [CrossRef]
- Neal, J.L.; Atkinson, T.G.; Larson, R.I. Changes in the rhizosphere microflora of spring wheat induced by disomic substitution of a chromosome. Can. J. Microbiol. 1970, 16, 153–158. [CrossRef]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 2014, 8, 790–803. [CrossRef]
- Bressan, M.; Roncato, M.-A.; Bellvert, F.; Comte, G.; Haichar, F.E.Z.; Achouak, W.; et al. Exogenous glucosinolate produced by Arabidopsis thaliana has an impact on microbes in the rhizosphere and plant roots. ISME J. 2009, 3, 1243-1257. [CrossRef]
- Li, J.; Li, Y.; Wei, X.; Cui, Y.; Gu, X.; Li, X.; Yoshigan, T.; Abd-Elgawad, M.; Shapiro-Ilan, D.; Ruan, W.; Rasmann, S. Direct antagonistic effect of entomopathogenic nematodes and their symbiotic bacteria on root-knot nematodes migration toward tomato roots. Pl Soil 2023, 484, 441-455. [CrossRef]
- Abd-Elgawad M.M.M. Transformational technologies for more uptakes of entomopathogenic nematodes. Egypt. J. Biol. Pest Cont. 2025, 35:1. [CrossRef]
- Meidani, C.; Savvidis, A.; Lampropoulou, E.; Sagia, A.; Katsifas, E.; Monokrousos, N.; Hatzinikolaou, D.G.; Karagouni, A.D.; Giannoutsou, E.; Adamakis, I.-D.S. et al. The nematicidal potential of bioactive Streptomyces strains isolated from Greek rhizosphere soils tested on Arabidopsis plants of varying susceptibility to Meloidogyne spp. Plants 2020, 9, 699. [CrossRef]
- Abd-Elgawad, M.M.M. Expanding biological control spectra of entomopathogenic nematodes to tick pests. Pakistan J. Nematol. 2025, 43(1), 09-22. [CrossRef]
- Abd-Elgawad, M.M.M. Challenges in field application of biopesticides for successful crop pest management. In Pest management: Methods, applications and challenges; Askary, T., Ed.; Nova Science Publishers, Inc., New York, USA, 2022; pp. 331-366. [CrossRef]
- Abd-Elgawad, M.; El-Mougy, N.S.; El-Gamal, N.G.; Abdel-Kader, M.M.; Mohamed, M. Protective treatments against soilborne pathogens in citrus orchards. J. Pl. Protect. Res. 2010, 50(4), 512-519.
- Chowdhury, S.P.; Hartmann, A.; Gao, X.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42–a review. Front. Microbiol. 2015, 6, 137829. [CrossRef]
- Abd-Elgawad, M.M.M. Nanonematicides: production, mechanisms, efficacy, opportunities and challenges. Nematology 2024, 26(5), 479-489. [CrossRef]
- Abd-El-Kareem, F.; Elshahawy, I.E.; Abd-Elgawad, M.M.M. Application of Bacillus pumilus isolates for management of black rot disease in strawberry. Egypt. J. Biol. Pest Cont. 2021, 31, 25. [CrossRef]
- Ajijah, N.; Fiodor, A.; Pandey, A.K.; Rana, A.; Pranaw, K. Plant growth-promoting bacteria (PGPB) with biofilm-forming ability: A multifaceted agent for sustainable agriculture. Diversity 2023, 15, 112. [CrossRef]
- Meyer, S.L.F.; Roberts, D.P. Combinations of biocontrol agents for management of plant-parasitic nematodes and soilborne plant-pathogenic fungi. J. Nematol. 2002, 34, 1–8. PMID: 19265899; PMCID: PMC2620533.
- Liu, X.; Mei, S.; Salles, J.F. Inoculated microbial consortia perform better than single strains in living soil: A meta-analysis. Appl. Soil Ecol. 2023, 190, 105011. [CrossRef]
- El-Nagdi, W.M.A.; Youssef, M. M.A.; Abd-El-Khair, H.; Abd-Elgawad, M.M.M. Effect of certain organic amendments and Trichoderma species on the root-knot nematode, Meloidogyne incognita infecting pea (Pisum sativum L.) plants. Egypt. J. Biol. Pest Cont. 2019, 29, 75. [CrossRef]
- Meyer, S.L.F.; Roberts, D.P.; Chitwood, D.J.; Carta, L.K.; Lumsden, R.D.; Mao, W. Application of Burkholderia cepacia and Trichoderma virens, alone and in combinations, against Meloidogyne incognita on bell pepper. Nematropica 2001, 31, 75–86.
- Ali, A.A.I.; Mahgoub, S.A.; Ahmed, A.F.; Mosa, W.F.; El-Saadony, M.T.; Mohamed, M.D.; Alomran, M.M.; Al-Gheffari, H.K.; El-Tarabily, K.A.; AbuQamar, S.F.; et al. Utilizing endophytic plant growth-promoting bacteria and the nematophagous fungus Purpureocillium lilacinum as biocontrol agents against the root-knot nematode (Meloidogyne incognita) on tomato plants. Eur. J. Plant Pathol. 2024, 170, 417–436. [CrossRef]
- Molinari, S.; Akbarimotlagh, M.; Leonetti, P. Tomato root colonization by exogenously inoculated arbuscular mycorrhizal fungi induces resistance against root-knot nematodes in a dose-dependent manner. Int. J. Mol. Sci. 2022, 23, 8920. [CrossRef]
- Topalović, O.; Vestergård, M. Can microorganisms assist the survival and parasitism of plant-parasitic nematodes? Trends Parasitol. 2021, 37, 947–958. [CrossRef]
- Trejo-Meléndez, V.J.; Contreras-Garduño, J. Master of puppets: How microbiota drive the nematoda ecology and evolution?.” Ecol. Evol. 2025, 15, e71549. [CrossRef]
- Abd-Elgawad, M.M.M.; Duncan, L.W.; Hammam, M.M.A.; Elborai, F.; Shehata, I.E. Effect of mulching manures and use of Heterorhabditis bacteriophora on strawberry fruit yield, Temnorhynchus baal and Meloidogyne javanica under field conditions. Arab J. Plant Prot. 2024, 42, 306–317. [CrossRef]
- Wang, L.; Li, X. Steering soil microbiome to enhance soil system resilience. Crit. Rev. Microbiol. 2019, 45, 743–753. [CrossRef]
- Lee Díaz, A.S.; Macheda, D.; Saha, H.; Ploll, U.; Orine, D.; Biere, A. Tackling the context-dependency of microbial-induced resistance. Agronomy 2021, 11, 1293. [CrossRef]
- Hammam, M.M.A.; Abd-El-Khair, H.; El-Nagdi, W.M.A.; Abd-Elgawad, M.M.M. Can agricultural practices in strawberry fields induce plant–nematode interaction towards Meloidogyne-suppressive soils? Life 2022, 12, 1572. [CrossRef]
- Kepenekci, I.; Hazir, S.; Oksal, E.; Lewis, E.E. Application methods of Steinernema feltiae, Xenorhabdus bovienii and Purpureocillium lilacinum to control root-knot nematodes in greenhouse tomato systems. Crop Prot. 2018, 108, 31–38. [CrossRef]
- Hammam, M.M.A.; Shehata, I.E.; Abd-Elgawad, M.M.M. Biocontrol of plant-parasitic nematodes by entomopathogenic nematodes: Case studies for further optimization. Arab J. Plant Prot. 2025, 43, In Press.
- Dritsoulas, A.; El-Borai, F.E.; Shehata, I.E.; Hammam, M.M.; El-Ashry, R.M.; Mohamed, M.M.; Abd-Elgawad, M.M.; Duncan, L.W. Reclaimed desert habitats favor entomopathogenic nematode and microarthropod abundance compared to ancient farmlands in the Nile Basin. J. Nematol. 2021, 53, 1-13. [CrossRef]
- Han, P.; Rodriguez-Saona, C.; Zalucki, M.P.; Liu, S.S.; Desneux, N. A theoretical framework to improve the adoption of green integrated pest management tactics. Commun. Biol. 2024, 7, 337. [CrossRef]
- Uroz, S.; Courty, P.; Oger, P. Plant symbionts are engineers of the plant-associated microbiome. Trends Pl. Sci. 2019, 24, 905–916. [CrossRef]
- Afridi, M.S.; Ali, S.; Salam, A.K.; Terra, W.C.; Hafeez, A.; Sumaira, , et al. Plant microbiome engineering: hopes or hypes. Biology (Basel) 2022, 11, 1782. [CrossRef]
- Ibrahim, H.M.M.; Ahmad, E.M.; Martínez-Medina, A.; Aly, M.A.M. Effective approaches to study the plant-root knot nematode interaction. Pl. Physiol. Biochem. 2019, 141, 332–342. [CrossRef]
- O’Halloran, D.M. CRISPR-PN2: a flexible and genome-aware platform for diverse CRISPR experiments in parasitic nematodes. BioTechniques 2001, 71, 495–498. [CrossRef]
- Abd-Elgawad, M.M.M. Nematode spatial distribution in the service of biological pest control. Egypt. J. Biol. Pest Cont. 2024, 34, 3. [CrossRef]
- Dastogeer, G.; Kao-Kniffin, J.; Okazaki, S. Editorial: plant microbiome: diversity, functions, and applications. Front. Microbiol. 2022, 13, 1039212. [CrossRef]
- Malayejerdi, Z.; Pouresmaeil, O. Current applications of the microbiome engineering and its future: a brief review. J. Curr. Biomed. Rep. 2020, 1, 48–51. [CrossRef]
- Abd-Elgawad, M.M.M. Crop genomics for climate resilient agriculture. In Innovations in Climate Resilient Agriculture; Rajan, R., Ahmad, F., Pandey, K., Eds.; Springer, Cham, Switzerland, 2025; pp. 207-225. [CrossRef]
- Tülek, A.; Kepenekçi, ˙I.; Oksal, E.; Hazir, S. Comparative effects of entomopathogenic fungi and nematodes and bacterial supernatants against rice white tip nematode. Egypt. J. Biol. Pest. Cont. 2018, 28, 1–6. [CrossRef]
- Vicente, C.S.L.; Soares, M.; Faria, J.M.S.; Espada, M.; Mota, M.; Nóbrega, F.; Ramos, A.P.; Inácio, M.L. Fungal communities of the pine wilt disease complex: Studying the interaction of ophiostomatales with Bursaphelenchus xylophilus. Front. Plant Sci. 2022, 13, 1650. [CrossRef]
- Liu, Y.; Ponpandian, L.N.; Kim, H.; Jeon, J.; Hwang, B.S.; Lee, S.K.; Park, S.-C.; Bae, H. Distribution and diversity of bacterial endophytes from four Pinus species and their efficacy as biocontrol agents for devastating pine wood nematodes. Sci. Rep. 2019, 9, 1–12. [CrossRef]
- Kim, N.; Jeon, H.W.; Mannaa, M.; Jeong, S.I.; Kim, J.; Kim, J., et al. Induction of resistance against pine wilt disease caused by Bursaphelenchus xylophilus using selected pine endophytic bacteria. Pl. Pathol. 2019, 68, 434–444. [CrossRef]
- McDonald, M.R.; Ives, L.; Adusei-Fosu, K.; Jordan, K.S. Ditylenchus dipsaci and Fusarium oxysporum on garlic: One plus one does not equal two. Can. J. Plant Pathol. 2021, 43, 749–759. [CrossRef]
- Huang, M.; Bulut, A.; Shrestha, B.; Matera, C.; Grundler, F.M.W.; Schleker, A.S.S. Bacillus firmus I-1582 promotes plant growth and impairs infection and development of the cyst nematode Heterodera schachtii over two generations. Sci. Rep. 2021, 11, 1–15. [CrossRef]
- Ahmed, S.; Liu, Q.; Jian, H. Bacillus cereus a potential strain infested cereal cyst nematode (Heterodera avenae). Pakistan J. Nematol. 2019, 37, 53–61. [CrossRef]
- Benedetti, T.; Antoniolli, Z.I.; Sordi, E.; Carvalho, I.R.; Bortoluzzi, E.C. Use of the Glomus etunicatum as biocontrol agent of the soybean cyst nematode. Res. Soc. Dev. 2021, 10, e7310615132. [CrossRef]
- Parkunan, V.; Johnson, C.S.; Eisenback, J.D. Biological and chemical induction of resistance to the Globodera tabacum solanacearum in oriental and flue-cured tobacco (Nicotiana tabacum L.). J. Nematol. 2009, 41, 203–210. PMID: 22736815; PMCID: PMC3380491. [PubMed] [PubMed Central]
- Xiang, N.; Lawrence, K.S.; Kloepper, J.W.; Donald, P.A. Biological control of Rotylenchulus reniformis on soybean by plant growth-promoting rhizobacteria. Nematropica 2018, 48, 116–125.
- Aballay, E.; Prodan, S.; Correa, P.; Allende, J. Assessment of rhizobacterial consortia to manage plant parasitic nematodes of grapevine. Crop Prot. 2020, 131, 105103. [CrossRef]
- Wong-Villarreal, A.; Méndez-Santiago, E.W.; Gómez-Rodríguez, O.; Aguilar-Marcelino, L.; García, D.C.; García-Maldonado, J.Q.; Hernández-Velázquez, V.M.; Yañez-Ocampo, G.; Espinosa-Zaragoza, S.; Ramírez-González, S.I.; et al. Nematicidal activity of the endophyte Serratia ureilytica against Nacobbus aberrans in Chili plants (Capsicum annuum L.) and identification of genes related to biological control. Plants 2021, 10, 2655. [CrossRef]
- Elsen, A.; Gervacio, D.; Swennen, R.; de Waele, D. AMF-induced biocontrol against plant- parasitic nematodes in Musa sp.: a systemic effect. Mycorrhiza 2008, 18, 251–256. [CrossRef]
- Van Overbeek, L.S.; Leiss, K.A.; Bac-Molenaar, J.A.; Duhamel, M. Plant resilience - role of chemical and microbial elicitors on metabolome and microbiome. Available online: file:///C:/Users/Mahfouzz/Desktop/WPR-1043_LR_Plantresilience_metabolomeandmicrobiome.pdf (accessed on 30 August 2025).
- Abd-Elgawad, M.M.M. Plant-parasitic nematodes and their biocontrol agents: Current status and future vistas. In Management of Phytonematodes: Recent Advances and Future Challenges, 1st ed.; Ansari, R., Rizvi, R., Mahmood, I., Eds.; Springer, Singapore, 2020; pp. 171–203.
- Tiwari, M.; Pati, D.; Mohapatra, R.; Sahu, B.B.; Singh, P. Impact of microbes in plant immunity and priming induced inheritance: A sustainable approach for crop protection. Plant Stress 2022, 4, 100072. [CrossRef]
- Turgut-Kara, N.; Arikan, B.; Celik, H. Epigenetic memory and priming in plants. Genetica 2020, 148(2), 47–54. . [CrossRef]
- Richard, M.M.S.; Gratias, A.; Meyers, B.C.; Geffroy V. Molecular mechanisms that limit the costs of NLR-mediated resistance in plants. Mol. Plant Pathol. 2018, 19, 2516-2523. [CrossRef]
- Timper, P. Conserving and enhancing biological control of nematodes. J. Nematol. 2014, 46, 75–89. [PubMed].
- Osman, H.A.I.; Ameen, H.H.; Hammam, M.M.A.; El-Sayed, G.M.; Elkelany, U.S.; Abd-Elgawad, M.M.M. Antagonistic potential of an Egyptian entomopathogenic nematode, compost and two native endophytic bacteria isolates against the root-knot nematode (Meloidogyne incognita) infecting potato under field conditions. Egypt. J. Biol. Pest Cont. 2022, 32, 137. [CrossRef]
Figure 1.
Plant-parasitic nematodes and microbial communities: Various mechanisms induced by microbiome rhizosphere to boost plant growth and/or strengthen its molecular biology-based immunity response.
Figure 1.
Plant-parasitic nematodes and microbial communities: Various mechanisms induced by microbiome rhizosphere to boost plant growth and/or strengthen its molecular biology-based immunity response.
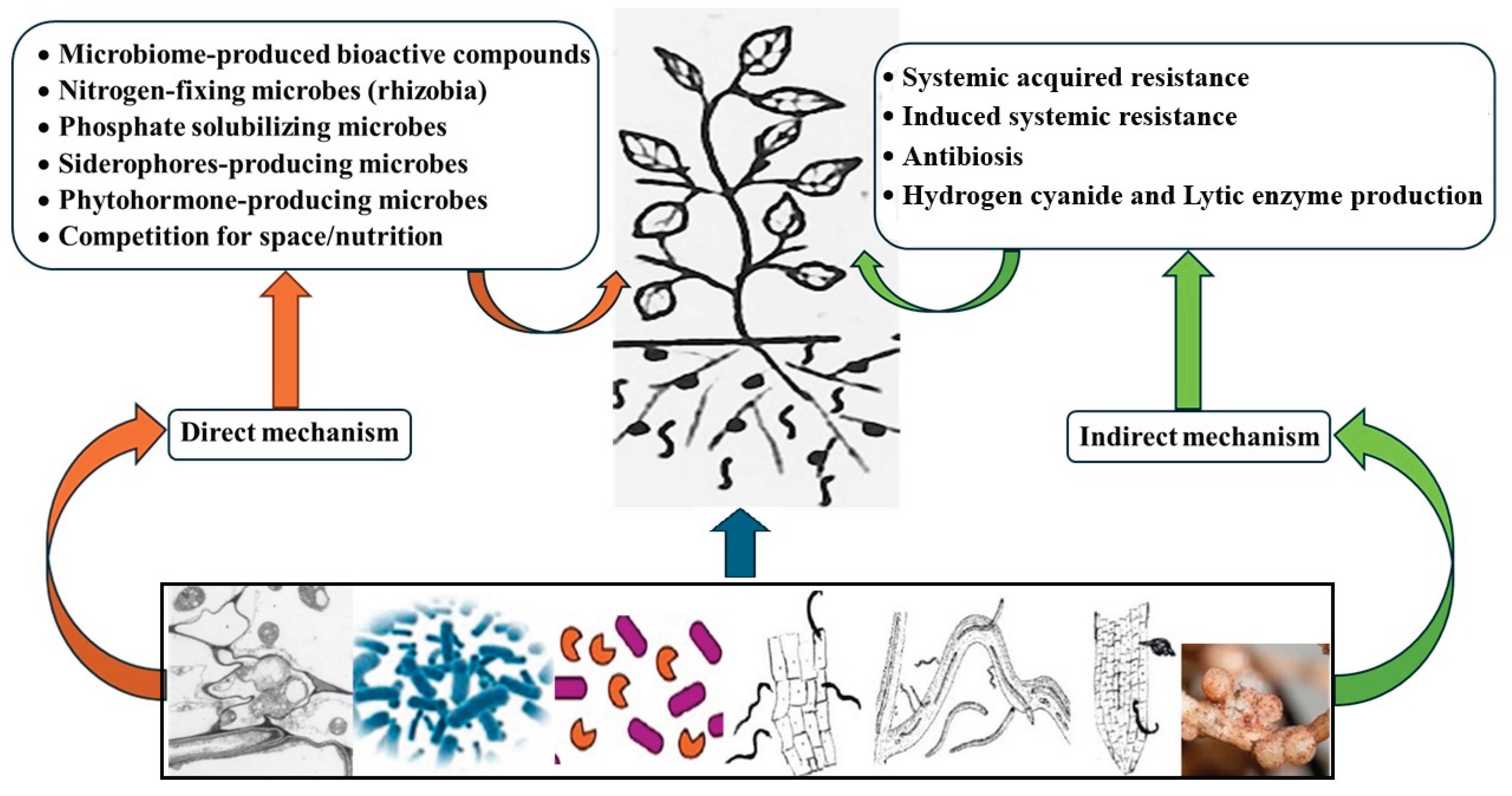

Table 1.
Molecular transcription factors (TF) engaged in plant responses to invasion and development of five key plant-parasitic nematode species.
Table 1.
Molecular transcription factors (TF) engaged in plant responses to invasion and development of five key plant-parasitic nematode species.
| TF | Plant | Gene/Genotype/TF | Function /nematode species | Ref. |
| WRKY | Arabidopsis thaliana | AtWRKY23 | Knocking down its expression lowered Heterodera schachtii infection | [19] |
| WOX11 | Arabidopsis thaliana | LBD16 and other WOX genes | Forming lateral roots to reduce the impact of H. schachtii infections | [20] |
| GT-3a (a trihelix TF) | Arabidopsis thaliana | TOZ and RAD23C | Its stabilization by Meloidogyne incognita effectors led to suppress related plant genes and ease the formation of the giant cells. | [21] |
| SlWRKY3 | Solanum lycopersicum | SlWRKY23 | Upregulated resistance to M. javanica via shikimate pathway activation | [22] |
| SLWRKY72a/SLWRKY72b | S. lycopersicum | Mi-1 induces effector-triggered immunity | Upregulating activator of immune response against M. incognita | [23] |
| WRKY | Oryza sativa | OsWRKY34, OsWRKY36, and OsWRKY62 | Their overexpression suppresses the defense-related genes, implying that they function as a negative regulator of innate immunity against Meloidogyne graminicola | [17] |
| WRKY | Oryza sativa | OsWRKY62,OsWRKY70 and OsWRKY11 | Their early up-regulation induced oxidative stress against Hirschmanniella oryzae, that is, up-regulated 3 Peroxidase precursors and 2 glutathione S-transferases | [17] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



