
Submitted:
03 February 2025
Posted:
04 February 2025
You are already at the latest version
Abstract
Soil microorganisms play an important role in regulating the responses of plants to disease, but little is known regarding the role of the microbial community in rhizosphere soil in the response to diseases in papaya. To clarify how the abundance and composition of the microbial rhizosphere community of papaya change with viral infection, papaya plants resistant and susceptible to papaya ringspot virus (PRSV) were inoculated with PRSV in the field, and differences in rhizosphere soil microbial communities between healthy and susceptible papaya were analyzed. The physiochemical properties of the soil surrounding the root system were strongly affected by microbial taxa, but this did not have a significant effect on plant health. 16S rRNA and ITS sequencing revealed that disease stress increased the microbial diversity of bacteria and decreased the diversity of fungi. The analysis of soil microbial community shows that the abundance of Nitrospirae was significantly higher in HN soil (1.35 ± 0.41) than in NC soil (0.83 ± 0.13) and YK soil (0.79 ± 0.27); the abundance of Paenibacillus was higher in HN soil (0.83 ± 0.38) than in NC (0.35 ± 0.12)and YK soil (0.58 ± 0.19); the abundance of Acremonium was higher in HN soil (0.71 ± 0.37) than in NC soil (0.21 ± 0.18) and YK soil (0.48 ± 0.43). Nitrospira, Paenibacillus, and Acremonium were all beneficial microorganisms, and they were significantly higher in healthy papaya soil than in susceptible papaya soil may help viral resistance. We show for the first time that a non-soil-borne disease may influence the microbiomes of papaya rhizosphere soil. When resistant papaya was infected with PRSV, beneficial microorganisms could be recruited in the rhizosphere soil to resist viral infection. Although these results are only preliminary insights into the potential role of beneficial microorganisms in supporting plant health under viral stress, our research provides new insights with implications for the prevention and control of papaya ringspot disease, furthermore, microorganisms that promote viral resistance could expediate the development of biopharmaceuticals or green fertilizers.
Keywords:
microbial community
; microbiome
; rhizosphere soil
; papaya ringspot disease
; PRSV
1. Introduction
Papaya (Carica papaya Linn.) is one of the most economically important fruit crops in tropical and sub-tropical countries, and it has high nutritional value [1]. However, papaya viruses pose a significant threat to papaya production. Papaya ringspot disease caused by papaya ringspot virus (PRSV) is one of the main diseases of papaya plants [2,3]. PRSV infections in papaya farms can cause severe yield losses up to 40–100% [4]. PRSV is mainly transmitted through aphids in the field [5]. The application of insecticides can prevent the transmission of viruses from infected to healthy plants, but some aphids are resistant to commonly used insecticides [6]; the use of chemical pesticides can also have environmental and human health hazards [7]. Transgenic papaya could be used for the control of PRSV, and several genetically modified papayas and transgenic cultivars have been developed. However, these genetically modified products are not popular among consumers, and resistance to PRSV depends on the origin of the viral isolates [8]; differences between geographical isolates have precluded the development of transgenic plants with PRSV resistance [9]. The development of effective methods for controlling PRSV thus remains a major challenge [10], and exploration of novel methods of biological control could prove useful.
More and more research disclosed that plants are able to shape their rhizosphere microbiome and their microbial communities could play a central role upon pathogen or insect attack (Berendsen et al., 2012). Soil microorganisms play an important role in determining the responses of plants to biotic and abiotic stresses [11]. Plants can alter the soil microbial community around their roots; the rhizosphere microbial community can also affect the growth of plants [12]. Studies of the interactions between plant diseases and soil microbes have mostly focused on soil-borne diseases, such as Fusarium graminearum for barley [13], Rhizoctonia solani AG8 for wheat [14], bacterial wilt disease of tomato [15,16], and Fusarium wilt disease of bananas [17]. When plants are infected with pathogenic bacteria, the rhizosphere soil recruits beneficial microorganisms to directly inhibit the growth of pathogenic bacteria or indirectly induce host defense responses [18]. Many studies on rhizosphere soil microorganisms of various crops have shown that plant-related microorganisms play a major role in health and susceptible plants [11,19,20,21,22,23,24,25,26,27]. Rhizosphere soil microbes are more abundant in soil with healthy plants than in soil with diseased plants [28].
Papaya ringspot disease is spread by arboreal insects and is thus not a soil-borne disease. As host plants communicate and recruit microbial organisms to the rhizosphere, changes in the health of a host may alter signals released by the roots, especially under systemic viral infections. Thus, papaya rhizosphere microbiomes may be altered under PRSV infection. However, changes in papaya rhizosphere soil microbial communities under disease stress have not yet been reported, and little is known about the role of interactions between papaya and soil in the disease resistance of papaya. We identified individual papaya plants of one variety with natural PRSV tolerance in papaya fields. This variety is not cultivated commercially because of its undesirable horticultural characteristics. Here, we used this resistant papaya variety and two susceptible papaya varieties to study changes in rhizosphere soil microorganisms following inoculation with PRSV. We collected rhizosphere soils of healthy papaya plants and susceptible plants and performed metagenomic analyses. Our findings revealed a potential route for the biological control of PRSV, which will aid the development of green agricultural systems.
2. Results
2.1. Soil Physicochemical Properties
The content of soil organic matter (SOM) and total nitrogen (TN) was significantly higher in NC soil than in YK and HN soil (P < 0.05); however, no significant differences in SOM and TN were observed between HN and YK soil (Table S1). The other physicochemical properties (TP, TK, AN, pH, AP, and AK) did not significantly vary among NC, YK, and HN soil.
2.2. Sequence Data Analysis
After quality control filtering, 1,455,875 and 1,665,561 effective tags were obtained from the 16S rRNA and ITS data, respectively. A total of 16,086 bacterial OTUs and 2,353 fungal OTUs were obtained after OTU clustering at a 97% similarity threshold. Rarefaction analyses revealed that the curve of 16S genes plateaued at a sequencing depth of 52,857 reads (Figure S2a), and the curve of fungal ITS genes plateaued at a sequencing depth of 44,017 reads (Figure S2b). The classification of ASVs based on the different taxonomic levels is shown in Table 1; OTUs are shown in Table 2. The number of soil bacterial taxonomic groups was higher in HN soil than in NC and YK soil, and the number of soil fungal taxonomic groups was lower in HN soil than in NC and YK soil.
The number of soil bacterial taxonomic groups was higher in HN plants than in NC and YK plants; specifically, there were 1, 11, 15, and 816 more families, genera, species, and OTUs, respectively, in HN soil than in NC soil and 4, 8, 21, and 1,276 more families, genera, species, and OTUs, respectively, in HN soil than in YK soil. The number of fungal communities was lower in HN soil than in NC and YK soil; specifically, there were 11, 28, and 13 fewer families, genera, and species, respectively, in HN soil than in NC soil and 6, 20, and 6 fewer families, genera, and species, respectively, in HN soil than in YK soil. The results are shown in Table 2.
2.3. Microbial Richness and Diversity
The richness and diversity of the three groups of rhizosphere microbiomes were analyzed. The OTU counts and Chao1 indices revealed that bacterial diversity was higher and fungal diversity was lower in HN soil than in NC and YK soil (Table 3). However, no significant differences were observed among HN, NC, and YK soil in the diversity indices of bacteria and fungi (P > 0.05).
In the β-diversity analysis, the microflora of HN, NC, and YK soil was not completely clustered in the PCoA plot (Figure 1A, C). In the PLS-DA (Figure 1B, D), the rhizosphere microbiota was differentiated into three independent clusters, which revealed significant differences in the composition of the rhizosphere microbiota among HN, NC, and YK soil. The Bray-Curtis index for bacteria revealed significant differences in β-diversity between NC and YK soil (P = 0.003), HN and NC soil (P = 0.025), and HN and YK soil (P = 0.013). Similar results were obtained using the unweighted UniFrac distance index, which revealed significant differences in β-diversity between NC and YK soil (P = 0.004), HN and NC soil (P = 0.046), and HN and YK soil (P = 0.025). Fungal Bray-Curtis indices revealed significant differences in β-diversity between NC and YK soil (P = 0.004), HN and NC soil (P = 0.013), and HN and YK soil (P = 0.003) (Table 4). Similar results were obtained using the unweighted UniFrac distance index, which revealed significant differences in β-diversity between NC and YK soil (P = 0.021), HN and NC soil (P = 0.002), and HN and YK soil (P = 0.007) (Table 4).
2.4. Composition of the Soil Microbial Community
Genera and phyla comprising the microbiota with average proportions greater than 0.5% in all soil samples are shown in a bar plot (Figure 2). The dominant bacterial phyla were Proteobacteria, Actinobacteria, Firmicutes, and Acidobacteria, and the cumulative average proportion of these phyla was 81.41% (Figure 2A); 63.86% of bacterial genera were not classified. Among known taxonomic genera, the dominant genera were Bacillus and Streptomyces (Figure 2B). In the fungal community, the dominant phylum was Ascomycota, and its average relative abundance was 80.06% (Figure 2C); only 25.96% of fungal OTUs were not classified at the genus level. Among known genera, the dominant genera were Aspergillus, Myceliophothora, and Neocosmospora (Figure 2D). The dominant bacterial phyla and genera were the same in healthy and diseased papaya plant soil, but slight differences in their abundances were observed (Figure S3).
To clarify the composition of the soil microbiota in HN, NC, and YK soil, the relative richness of phyla and genera with relative abundance greater than 0.5% was analyzed (Table 5). The abundance of Nitrospirae was significantly higher in HN soil than in NC soil (P = 0.04) and YK soil (P = 0.01); the abundances of Gemmatimonadetes and Chloroflexi were higher and the abundance of TM7 was lower in HN soil than in NC and YK soil, but significant differences were only observed between HN and [YK] soil. The abundance of Nitrospira was higher in HN soil than in NC (P = 0.002) and YK (P = 0.001) soil, and the abundance of Paenibacillus was higher in HN soil than in NC and YK soil. The abundances of Rhodanobacter and Sphingomonas were lower in HN soil than in NC and YK soil; the differences between HN and YK soil were significant (P = 0.027 and 0.031, respectively) (Table 5).
The abundance of Glomeromycota was significantly higher in HN soil than in NC (P = 0.035) and YK (P = 0.035) soil. The abundance of Mortierellomycota was higher in HN soil than in NC and YK soil; however, significant differences in Mortierellomycota abundance were only observed between HN and NC soil (P = 0.023). The abundance of Basidiomycota was significantly higher in HN soil than in NC soil (P = 0.028), but lower in HN soil than in YK soil, albeit these differences were not significant. The abundances of Neocosmospora and Acremonium were higher and that of Polyschema was lower in HN soil than in NC and YK soil; the abundance of Trichoderma was significantly lower in HN soil than in NC soil (P = 0.013); Trichoderma abundance did not significantly differ in HN and YK soil (Table 6).
2.5. LEfSe Analysis Revealed Differences in Soil Microbial Genera
LEfSe analysis was performed to determine significant differences in microbial markers among HN, NC, and YK soil (LDA > 3, P < 0.05). The abundance of Nitrospirae, Chloroflexi, and Euryarchaeota had a significant effect on the rhizosphere soil microorganisms of healthy and diseased papaya plants; the abundance of Nitrospira, Paenibacillus, Rummeliibacillus, Oceanobacillus, Brevibacillus, Adhaeribacter, Filomicrobium, and Agromys was significantly higher in HN soil than in NC and YK soil (Figure 3A). LDA of fungal communities revealed that the abundance of Mortierellomycota and Glomeromycota had a significant effect on the rhizosphere soil microorganisms of healthy and diseased papaya plants; the abundance of Acremonium and Funneliformes was significantly higher in HN soil than in NC and YK soil (Figure 3B). The relative abundances of Nitrospira, Paenibacillus, and Acremonium were greater than or equal to 0.5%; the relative abundances of all other genera were less than 0.5%. These three genera were all enriched in the rhizosphere soil of healthy papaya plants.
2.6. Associations Between Soil Microbial Communities and Environmental Factors
Soil properties have a strong effect on the soil microbiota. Redundancy analysis (RDA) was used to evaluate the effects of environmental factors on the composition of rhizosphere soil microbial phyla. The first two RDA axes explained 58.08% and 21.09% of the total variation in soil bacterial community structure. The first axis was positively correlated with soil AK but negatively correlated with soil AN, AP, and SOM. The second axis was positively correlated with soil AK, AN, AP, and SOM and negatively correlated with soil pH (Figure 4A). We used Spearman correlations to evaluate the relationships between the abundance of phyla and soil physicochemical properties (Figure 4B). Bacteroidota, WS3, Gemmatimonadetes, Planctomycetes, Acidobacteria, TM6, Chlamydiae, Verrucomicrobia, and Crenarchaeota were significantly positively correlated with soil pH. These taxa were significantly negatively correlated with soil AK. Elusimicrobia was positively correlated with soil pH, and OP11 was positively correlated with AN and SOM. Proteobacteria was significantly negatively correlated with soil pH, and WS3 was significantly negatively correlated with soil AN; Euryarchaeota, Armatimonadetes, Nitrospirae, and OD1 were significantly negatively correlated with soil AK (P < 0.05) (Figure 4B).
The first two RDA axes explained 51.52% and 24.21% of the total variation in soil fungal community structure. The first axis was positively correlated with soil AK and negatively correlated with soil AN and SOM. The second axis was positively correlated with soil SOM, AN, and AK and negatively correlated with soil AP and pH (Figure 4C). Spearman correlation analysis between soil fungal communities and physicochemical properties showed that Basidiomycota was significantly positively correlated with soil AP, SOM, and AN (P < 0.05). Rozellomycota was positively correlated with soil AP and SOM (P < 0.05). There was a significant negative correlation of Mortierellomycota with soil AP and SOM (P < 0.05). Kickxellomycota and Blastocladiomycota were positively correlated with soil pH (P < 0.05) (Figure 4D).
3. Discussion
Papaya viral disease is the main disease of papaya. At present, research mainly focuses on genetic improvement, using genetic methods to obtain some resistant papaya varieties, such as Rainbow of Hawaii’s papaya, YK16-0-1 from papaya in Taiwan area, Huanan No.1 from the papaya in the South of China, and so on. However, these genetically modified varieties can only target specific viruses and lack broader resistance. Developing new varieties will takes a long time and it is urgent to search for new strategies to solve the practical problems in agricultural production. Currently, studying plant resistance from the perspective of soil microorganisms is a hot topic and had accomplished great success in some plant fields. Here, we compared the differences in soil microorganisms between resistant and susceptible papaya from the perspective of soil microorganisms. In this study, we only sampled once after inoculation health and susceptible plants with PRSV and we didn’t consider the dynamic effects from temperature and seasons so it is a preliminary study. The research objectives of this article mainly focused on exploring the differences in rhizosphere soil microorganisms between susceptible and healthy papaya, identifying the main dominant microbial species, and laying the foundation for exploring their applications. In future research, we will sample papaya soil at different growth temperatures and seasons, compared the differences of microbial populations between healthy and susceptible plants, and study the dynamic changes of microorganisms from papaya rhizosphere soil after virus infection.
Physicochemical properties significantly affect the structure of soil microbial communities [29]. RDA showed that organic matter, nitrogen, phosphorus, potassium, and pH were all strongly associated with soil microbial community structure. Thus, environmental factors had a major effect on the structure of bacterial and fungal communities of papaya rhizosphere soil. However, no significant differences were observed in all physicochemical properties between soils containing healthy and susceptible plants. SOM and TN were significantly higher in NC soil than in YK and HN soil, and no significant differences in SOM and TN were observed between YK soil and HN soil. Although soil physicochemical properties strongly affect the structure of soil microbial communities, no significant correlations have been observed between plant health status and soil physicochemical properties [30].
The health of papaya plants might be associated with the rhizosphere soil microbial community. The results of α-diversity analysis revealed no significant differences in the diversity index of bacteria and fungi in the rhizosphere soil of HN, NC, and YK papaya plants (P > 0.05). However, the OTUs and Chao1 index of soil bacteria were higher in HN soil than in NC and YK soil. The OTUs and Chao1 index for fungi were higher in HN soil than in NC and YK soil. This indicates that papaya infected with PRSV had increased bacterial diversity and decreased fungal diversity. The results of soil microbiome analysis revealed differences in microbial communities in soil with healthy papaya plants and susceptible papaya plants. Greater numbers of bacterial taxa were observed in HN soil than in NC and YK soil, and numbers of fungal taxa were lower in HN soil than in NC and YK soil. Previous studies have shown that changes in soil microbial communities can reduce the prevalence of diseases [17,31]. Abundances of soil bacteria and fungi decrease and increase, respectively, when crops are continuously cropped, which leads to imbalances in the structure of soil bacterial and fungal communities and reductions in the ability of crops to resist diseases [32]. In our study, bacterial and fungal communities were more and less abundant in HN soil than in NC and YK soil, and this may enhance the disease resistance of papaya. Therefore, the health of papaya plants is associated with different bacterial and fungal microbial communities.
Infection with viral agents can lead to alterations in the structure of microbial communities in rhizosphere soil. Changes in the structure of the rhizosphere microbiota contribute to plant disease resistance [11]. β-diversity analysis showed that the microbial communities of HN, NC, and YK soil were not completely clustered, indicating that the soil microbial community composition of the three papaya plants differed. Changes in soil microorganisms were induced by PRSV. Previous studies have revealed differences in the composition of soil bacterial phyla and genera in soils with high and low disease severity [33]. The abundance of Nitrospirae, Chloroflexi, and Paenibacillus was higher in HN soil than in NC and YK soil; the abundance of Glomeromycota, Mortierellomycota, and Neocosmospora was higher in HN soil than in NC and YK soil. The abundance of Nitrospira, Paenibacillus, and Acremonium was significantly higher in HN soil than in NC and YK soil. Nitrospira can oxidize ammonia to nitrate [34,35,36], Nitrospira is widespread in rhizosphere soils [37], and it affects the nitrogen fertilization efficiency of agricultural soils [38,39]. By promoting biological nitrogen fixation and helping meet the nitrogen nutrient requirements for crop growth and development [40], it promotes plant growth and is generally considered a potentially useful probiotic for enhancing nitrogen fixation in rhizosphere soil [41]. A previous study has shown that the abundance of Nitrospirillum is lower in diseased soil than in healthy soil; its relative abundance is significantly negatively correlated with the plant disease index [42]. Paenibacillus was a new genus proposed in a taxonomic study of the genus Bacillus [43]. Most members of the genus Paenibacillus occur in soil, and this genus currently comprises approximately 200 species [44]. Paenibacillus can induce resistance to plant diseases via the production of various antibacterial substances; many members can also trigger induced systemic resistance against pathogenic bacteria [45], fungi [46], nematodes [47], and viruses [48]. Due to its potential utility as a biological fertilizer or biocontrol agent [49], Paenibacillus has played an increasingly important role in sustainable agriculture [50]. Acremonium also plays an important role in the control of plant diseases, and a variety of compounds toxic to insects have been detected in extracts of Acremonium fungi [51], which could enhance resistance to aphids [52]. Previous studies have shown that plant–soil interactions affect the performance of aboveground herbivores that feed on this plant [53,54], and soil microbial communities could play an important role in shaping aboveground plant–insect interactions [55]. Plants affect the performance of aboveground aphids via species-specific effects on soil fungal communities [54], and the magnitude of these effects change over time [56]. Papaya ringspot disease is mainly transmitted by aphids; thus, the abundance of Acremonium was significantly higher in the rhizosphere soil of resistant papaya plants than in that of susceptible plants, which may affect the ability of aphids to transmit PRSV to papaya plants. The abundance of these bacteria genera was significantly higher in the rhizosphere soil of healthy papaya plants than in that of diseased papaya plants, indicating that resistant papaya plants can recruit beneficial microorganisms to mitigate the deleterious effects of PRSV; this might also affect the transmission of the virus via aboveground aphids by altering microbial communities and thus plant–soil interactions.
4. Materials and Methods
4.1. Experimental Design and Collection of Soil Samples
Field experiments were conducted at the experimental base of the Institute of Tropical Biotechnology, Chinese Academy of Tropical Agricultural Sciences, which is located in Wujitangxia Village, Wenchang City, Hainan Province, China. We established three experimental blocks containing (1) a papaya variety with PRSV resistance (HN; purchased from Guangzhou Fuqin Agriculture Co., Ltd.), (2) a papaya variety susceptible to PRSV (YK; provided by Hainan Academy of Agricultural Sciences); and (3) a local cultivated papaya variety susceptible to PRSV (NC; provided by Hainan Academy of Agricultural Sciences). Thirty papaya plants were planted in each block. Before planting, the experimental area was weeded, plowed, and leveled. After planting papaya, no pesticides or fertilizers were sprayed throughout the entire experimental period. To prevent weed growth, we covered the experimental area with plastic film.
One month after planting, PRSV was inoculated onto papaya plants using the friction inoculation method [6]. To ensure inoculation success, a second inoculation was performed one week later. Twenty-one days after the second inoculation, susceptible papaya varieties showed symptoms of infection with “green islands” on their leaves, and the resistant variety did not show symptoms of PRSV infection. Leaf ELISA detection was performed to confirm PRSV infection using a Papaya ringspot virus enzyme-linked immunosorbent assay kit (Jingmei, China). The content of the virus in the two diseased papaya plants was 414 ± 7.32 ng/L and 372.53 ± 9.64 ng/L, and that in the resistant papaya plants was 85.92 ± 10.04 ng/L (Fig. S1).
Rhizosphere soils of healthy and infected plants were collected. When sampling, weeds and leaves were removed from the surface (if any), then the surface soil was removeed, and finally, soil samples were collected at a 10–20 cm soil depth and the distance is10 cm away from the main stem. Eight replicate soil samples per plot were collected. The samples were placed into sterile plastic bags and transported to the laboratory immediately for further analysis. Each replicate was mixed evenly, sieved through a 2 mm sieve, and divided into two portions. One portion was stored at -80 °C for DNA extraction, and the remaining soil was air-dried at room temperature for subsequent analysis of physicochemical properties.
4.2. Determination of Physicochemical Properties
The physicochemical properties of soil samples were determined immediately after drying using a previously described method [57]. Soil pH was determined in slurries (soil:water 1:2.5) using an FE28 acidimeter (Mettler-Toledo International Inc., Shanghai, China). The soil organic matter (SOM) content was measured using the potassium dichromate method. First, the total soil organic C content was determined by wet digestion with H2SO4-K2Cr2O7; next, the TOC value was multiplied by 1.724 to obtain SOM. Total soil N (TN) was detected using a fully automatic Kjeldahl nitrogen analyzer (Kjeltec 8400, Foss, Denmark). Soil hydrolytic N (AN) was measured after treating soil with a 1.8 mol/L NaOH solution. Soil total phosphorus (TP) and potassium (TK) were detected using the acid leaching method. Soil available P and K were extracted using NH4F (0.2 mol/L) and CH3COONH4 (1 mol/L) solutions, respectively.
4.3. DNA Extraction, PCR Amplification, and Sequencing
Soil DNA was extracted using the CTAB method. DNA concentration and purity were monitored on 1% agarose gels; the DNA was then diluted to 1 ng/µL using sterile water. The V3-V4 region of the 16S rRNA gene was amplified to determine the soil bacterial communities using the 341F (5′-CCTAYGGGRBGCASCAG-3′) and 806R (5′-GGACTACNNGGGTATCTAAT-3′) primers [58]. The ITS gene was amplified to determine the soil fungal communities using the ITS1-1F-F (5′-CTTGGTCATTTAGAGGAAGTAA-3′) and ITS1-1F-R (5′-GCTGCGTTCTTCATCGATGC-3′) primers [59]. All PCR reactions were performed using 15 µL of Phusion® High-Fidelity PCR Master Mix (New England Biolabs, USA), 2 µM of forward and reverse primers, and approximately 10 ng of template DNA. The thermal cycling parameters were as follows: 98 ℃ for 1 min (initial denaturation); 30 cycles of 98 ℃ for 10 s (denaturation), 50 ℃ for 30 s (annealing), and 72 ℃ for 30 s (elongation); and 72 ℃ for 5 min (final extension).
The PCR products were purified using the Qiagen Gel Extraction Kit (Qiagen, Germany). Next, sequencing libraries were generated using the TruSeq® DNA PCR-Free Sample Preparation Kit (Illumina, CA, USA). The library was then sequenced on an Illumina NovaSeq platform, and 250 bp paired-end reads were generated at Microeco Technology Co., Ltd. (Shenzhen, China).
4.4. Bioinformatics Analysis
Bioinformatics analysis was conducted using the QIIME 2 software packages along with customized program scripts (https://docs.qiime2.org/2024.5/citation/). After the FASTQ files containing raw data were imported, the demultiplexed sequences from each sample were quality filtered, trimmed, denoised, and merged; the chimeric sequences were then identified and removed using the QIIME 2 dada2 plugin with default settings to obtain the feature table of amplicon sequence variants (ASVs) [60]. The QIIME 2 feature-classifier plugin was then used to align the ASV sequences to a pre-trained GREENGENES 13_8 99% database (16S) or the UNITE database 8.2 version (ITS) to generate a taxonomy table [61]. Any contaminating mitochondrial and chloroplast sequences were removed using the QIIME 2 feature-table plugin.
5. Conclusions
In summary, we conducted a preliminary study on the response of papaya rhizosphere soil to viral stress. To our knowledge, this is the first study to characterize changes in the abundances of microorganisms in papaya rhizosphere soil following PRSV infection. Our results indicate that effectively regulating the community of beneficial microbes in soil to promote plant growth and health may be a promising approach for the control of PRSV. Overall, our findings provide new insights that will help the control of PRSV. The differentially abundant microorganisms will promote the development of their applications as green fertilizers in management of papaya diseases. However, we only collected sample once after the plant showed PRSV symptoms. In future research, we will further investigate the dynamic changes of soil microorganisms in papaya at different growth stages and seasons in order to acquire more detailed data and support the green prevention and control of papaya diseases.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Yanhua Tan and Xiang Xie: Conceptualization, Writing-original draft, Writing-review & editing. Qixing Huang: Writing-review & editing, software. Yang Cao, Xia Zhou, Chunqiang Li, Jianbo Sun, and Hui Wang: validation. Guohua Yin, Hui Zhao, and Anping Guo: supervision, Writing-review & editing, project administration. All authors have read and agreed to the published version.
Funding
This study was supported by the Hainan Provincial Natural Science Foundation of China (No. 320RC716), the Hainan Major Science and Technology Project (No. ZDKJ202002), and Research Program of Qilu Institute of Technology (No. QIT23TP009).
Acknowledgments
We thank Kayla K. Pennerman for her valuable suggestions.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mukherjee, A.; Debnath, P.; Ghosh, S.K.; Medda, P.K. Biological control of papaya aphid (Aphis gossypii Glover) using entomopathogenic fungi. Vegetos 2020, 33, 1–10. [Google Scholar] [CrossRef]
- Maina, S.; Coutts, B.A.; Edwards, O.R.; de Almeida, L.; Ximenes, A.; Jones, R.A.C. Papaya ringspot virus populations from east timorese and northern Australian cucurbit crops: biological and molecular properties, and absence of genetic connectivity. Plant Dis. 2017, 101, 985–993. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, N.; Huang, Q.; Yin, G.; Guo, A.; Wang, X.; Xiong, Z.; Liu, Z. Complete genome of Hainan papaya ringspot virus using small RNA deep sequencing. Virus Genes 2014, 48, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Thirugnanavel, A.; Balamohan, T.N.; Karunakaran, G.; Manoranjitham, S.K. Effect of papaya ringspot virus on growth, yield and quality of papaya (Carica papaya) cultivars. Indian Journal of Agricultural Sciences 2015, 85, 1069–1073. [Google Scholar] [CrossRef]
- Hidayat, S.H.; Harmiyati, T.; Adnan, A.M. Insect vector and seedborne transmission of Papaya ringspot virus. Jurnal Fitopatologi Indonesia 2022, 18, 101–106. [Google Scholar] [CrossRef]
- Fan, H.; Yan, X.; Fu, M.; Liu, D.; Awan, A.W.; Chen, P.; Rasheed, S.M.; Gao, L.; Zhang, R. Interactive effect of biological agents chitosan, lentinan and ningnanmycin on Papaya Ringspot Virus resistance in papaya (Carica papaya L.). Molecules 2022, 27, 7474. [Google Scholar] [CrossRef]
- Kalleshwaraswamy, C.; Krishnakumar, N.; Chandrashekara, K.; Vani, A. Efficacy of insecticides and oils on feeding behaviour of Aphis gossypii Glover and transmission of papaya ringspot virus (PRSV). Karnataka Journal of Agricultural Sciences 2012, 25. [Google Scholar]
- Gonsalves, D. Control of papaya ringspot virus in papaya: a case study. Annu. Rev. Phytopathol. 1998, 36, 415–437. [Google Scholar] [CrossRef] [PubMed]
- Ruanjan, P.; Kertbundit, S.; Juříček, M. Post-transcriptional gene silencing is involved in resistance of transgenic papayas to papaya ringspot virus. Biologia Plantarum 2007, 51, 517–520. [Google Scholar] [CrossRef]
- Akhtar, N.; Perween, S.; Ansari, A.M. Resistance to Papaya Ringspot Virus a Review. Agriculture Association of Textile Chemical and Critical Reviews Journal 2023, 11, 68–78. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Vismans, G.; Yu, K.; Song, Y.; de Jonge, R.; Burgman, W.P.; Burmølle, M.; Herschend, J.; Bakker, P.A.H.M.; Pieterse, C.M.J. Disease-induced assemblage of a plant-beneficial bacterial consortium. The ISME journal 2018, 12, 1496–1507. [Google Scholar] [CrossRef]
- Van der Putten, W.H.; Bardgett, R.D.; Bever, J.D.; Bezemer, T.M.; Casper, B.B.; Fukami, T.; Kardol, P.; Klironomos, J.N.; Kulmatiski, A.; Schweitzer, J.A.; et al. Plant–soil feedbacks: the past, the present and future challenges. J. Ecol. 2013, 101, 265–276. [Google Scholar] [CrossRef]
- Dudenhöffer, J.H.; Scheu, S.; Jousset, A. Systemic enrichment of antifungal traits in the rhizosphere microbiome after pathogen attack. J. Ecol. 2016, 104, 1566–1575. [Google Scholar] [CrossRef]
- Yin, C.; Casa Vargas, J.M.; Schlatter, D.C.; Hagerty, C.H.; Hulbert, S.H.; Paulitz, T.C. Rhizosphere community selection reveals bacteria associated with reduced root disease. Microbiome 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Kong, H.G.; Song, G.C.; Ryu, C.M. Disruption of Firmicutes and Actinobacteria abundance in tomato rhizosphere causes the incidence of bacterial wilt disease. The ISME Journal 2021, 15, 330–347. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, J.; Liu, F.; Liang, J.; Zhao, P.; Tsui, C.K.M.; Cai, L. Cross-kingdom synthetic microbiota supports tomato suppression of Fusarium wilt disease. Nature Communications 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Zhou, Z.; Liu, M.; Shen, Z.; Wang, B.; Jousset, A.; Geisen, S.; Ravanbakhsh, M.; Kowalchuk, G.A.; Li, R.; et al. Intercropping with Trifolium repens contributes disease suppression of banana Fusarium wilt by reshaping soil protistan communities. Agric., Ecosyst. Environ. 2024, 361, 108797. [Google Scholar] [CrossRef]
- Van Loon, L.; Bakker, P.; Pieterse, C. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol. 1998, 36, 453–483. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Rybakova, D.; Grube, M.; Köberl, M. The plant microbiome explored: implications for experimental botany. J. Exp. Bot. 2016, 67, 995–1002. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Garrido-Oter, R.; Münch, P.C.; Weiman, A.; Dröge, J.; Pan, Y.; McHardy, A.C.; Schulze-Lefert, P. Structure and function of the bacterial root microbiota in wild and domesticated barley. Cell Host & Microbe 2015, 17, 392–403. [Google Scholar]
- Bulgarelli, D.; Rott, M.; Schlaeppi, K.; Ver Loren van Themaat, E.; Ahmadinejad, N.; Assenza, F.; Rauf, P.; Huettel, B.; Reinhardt, R.; Schmelzer, E. Revealing structure and assembly cues for Arabidopsis root-inhabiting bacterial microbiota. Nature 2012, 488, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Busby, P.E.; Soman, C.; Wagner, M.R.; Friesen, M.L.; Kremer, J.; Bennett, A.; Morsy, M.; Eisen, J.A.; Leach, J.E.; Dangl, J.L. Research priorities for harnessing plant microbiomes in sustainable agriculture. PLoS Biol. 2017, 15, e2001793. [Google Scholar] [CrossRef]
- Castrillo, G.; Teixeira, P.J.P.L.; Paredes, S.H.; Law, T.F.; De Lorenzo, L.; Feltcher, M.E.; Finkel, O.M.; Breakfield, N.W.; Mieczkowski, P.; Jones, C.D.; et al. Root microbiota drive direct integration of phosphate stress and immunity. Nature 2017, 543, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Leach, J.E.; Triplett, L.R.; Argueso, C.T.; Trivedi, P. Communication in the phytobiome. Cell 2017, 169, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, D.S.; Lebeis, S.L.; Paredes, S.H.; Yourstone, S.; Gehring, J.; Malfatti, S.; Tremblay, J.; Engelbrektson, A.; Kunin, V.; Rio, T.G.d.; et al. Defining the core Arabidopsis thaliana root microbiome. Nature 2012, 488, 86–90. [Google Scholar] [CrossRef]
- Müller, D.B.; Vogel, C.; Bai, Y.; Vorholt, J.A. The plant microbiota: systems-level insights and perspectives. Annu. Rev. Genet. 2016, 50, 211–234. [Google Scholar] [CrossRef]
- Walters, W.A.; Jin, Z.; Youngblut, N.; Wallace, J.G.; Sutter, J.; Zhang, W.; González-Peña, A.; Peiffer, J.; Koren, O.; Shi, Q.; et al. Large-scale replicated field study of maize rhizosphere identifies heritable microbes. Proceedings of the National Academy of Sciences 2018, 115, 7368–7373. [Google Scholar] [CrossRef]
- Huang, X.; Zhou, X.; Zhang, J.; Cai, Z. Highly connected taxa located in the microbial network are prevalent in the rhizosphere soil of healthy plant. Biol. Fertility Soils 2019, 55, 299–312. [Google Scholar] [CrossRef]
- Jin, L.; Jin, N.; Wang, S.; Li, J.; Meng, X.; Xie, Y.; Wu, Y.; Luo, S.; Lyu, J.; Yu, J. Changes in the microbial structure of the root soil and the yield of Chinese baby cabbage by chemical fertilizer reduction with bio-organic fertilizer application. Microbiology Spectrum 2022, 10, e01215–e01222. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Wang, X.; Hou, R.; Lu, C.; Fan, Z.; Li, J.; Wang, S.; Xu, Y.; Shen, Q.; Friman, V.-P.; et al. Rhizosphere phage communities drive soil suppressiveness to bacterial wilt disease. Microbiome 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Lin, M.; Rensing, C.; Qin, X.; Zhang, S.; Chen, J.; Wu, L.; Zhao, Y.; Lin, S.; Lin, W. Plant-mediated rhizospheric interactions in intraspecific intercropping alleviate the replanting disease of Radix pseudostellariae. Plant Soil 2020, 454, 411–430. [Google Scholar] [CrossRef]
- Kang, Y.; Jing, F.; Sun, W.; Liu, J.; Jiang, G. Soil microbial communities changed with a continuously monocropped processing tomato system. Acta Agriculturae Scandinavica, Section B — Soil & Plant Science 2018, 68, 149–160. [Google Scholar] [CrossRef]
- Saraiva, A.L.d.R.; Bhering, A.d.S.; do Carmo, M.G.; Andreote, F.D.; Dias, A.C.; Coelho, I.d.S. Bacterial composition in brassica-cultivated soils with low and high severity of clubroot. J. Phytopathol. 2020, 168, 613–619. [Google Scholar] [CrossRef]
- Daims, H.; Lebedeva, E.V.; Pjevac, P.; Han, P.; Herbold, C.; Albertsen, M.; Jehmlich, N.; Palatinszky, M.; Vierheilig, J.; Bulaev, A.; et al. Complete nitrification by Nitrospira bacteria. Nature 2015, 528, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; He, Z.-Y.; Hu, H.-W.; He, J.-Z. Niche specialization of comammox Nitrospira in terrestrial ecosystems: oligotrophic or copiotrophic? Crit. Rev. Environ. Sci. Technol. 2023, 53, 161–176. [Google Scholar] [CrossRef]
- Van Kessel, M.A.; Speth, D.R.; Albertsen, M.; Nielsen, P.H.; Op den Camp, H.J.; Kartal, B.; Jetten, M.S.; Lücker, S. Complete nitrification by a single microorganism. Nature 2015, 528, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, S.; Jiang, Y.; Zhou, J.; Han, C.; Zhu, G. Comammox bacterial abundance, activity, and contribution in agricultural rhizosphere soils. Sci. Total Environ. 2020, 727, 138563. [Google Scholar] [CrossRef]
- Ouyang, Y.; Evans, S.E.; Friesen, M.L.; Tiemann, L.K. Effect of nitrogen fertilization on the abundance of nitrogen cycling genes in agricultural soils: a meta-analysis of field studies. Soil Biol. Biochem. 2018, 127, 71–78. [Google Scholar] [CrossRef]
- Zhu, G.; Peng, Y.; Li, B.; Guo, J.; Yang, Q.; Wang, S. Biological removal of nitrogen from wastewater. Reviews of environmental contamination and toxicology 2008, 159–195. [Google Scholar]
- De Santis Sica, P.; Shirata, E.S.; Rios, F.A.; Brandao Filho, J.U.T.; Schwan Estrada, K.R.F.; Oliveira, R.S.d. Impact of N-fixing bacterium 'Nitrospirillum amazonense'on quality and quantitative parameters of sugarcane under field condition. Australian Journal of Crop Science 2020, 14, 1870–1875. [Google Scholar] [CrossRef]
- Chung, E.J.; Park, T.S.; Kim, K.H.; Jeon, C.O.; Lee, H.I.; Chang, W.S.; Aslam, Z.; Chung, Y.R. Nitrospirillum irinus sp. nov., a diazotrophic bacterium isolated from the rhizosphere soil of Iris and emended description of the genus Nitrospirillum. Antonie Van Leeuwenhoek 2015, 108, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Wang, T.; Huang, Q.; Guo, H.; Zhang, H.; Xu, Q.; Shen, Q.; Ling, N. Core species impact plant health by enhancing soil microbial cooperation and network complexity during community coalescence. Soil Biol. Biochem. 2024, 188, 109231. [Google Scholar] [CrossRef]
- Ash, C.; Priest, F.G.; Collins, M.D. Molecular identification of rRNA group 3 bacilli (Ash, Farrow, Wallbanks and Collins) using a PCR probe test: proposal for the creation of a new genus Paenibacillus. Antonie Van Leeuwenhoek 1993, 64, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Grady, E.N.; MacDonald, J.; Liu, L.; Richman, A.; Yuan, Z.C. Current knowledge and perspectives of Paenibacillus: a review. Microbial Cell Factories 2016, 15. [Google Scholar] [CrossRef]
- Phi, Q.-T.; Park, Y.-M.; Seul, K.-J.; Ryu, C.-M.; Park, S.-H.; Kim, J.-G.; Ghim, S.-Y. Assessment of root-associated Paenibacillus polymyxa groups on growth promotion and induced systemic resistance in pepper. Journal of Microbiology and Biotechnology 2010, 20, 1605–1613. [Google Scholar] [PubMed]
- Chen, B.; Han, H.; Hou, J.; Bao, F.; Tan, H.; Lou, X.; Wang, G.; Zhao, F. Control of maize sheath blight and elicit induced systemic resistance using Paenibacillus polymyxa strain SF05. Microorganisms 2022, 10, 1318. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Zhang, J.; Fu, Q.; Hao, G.; Liang, C.; Duan, F.; Zhao, H.; Song, W. Biocontrol efficacy and induced resistance of Paenibacillus polymyxa J2-4 against Meloidogyne incognita infection in cucumber. Phytopathology® 2024, 114, 538–548. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Al-Askar, A.A.; Elbeaino, T.; Moawad, H.; El-Gendi, H. Protective and curative activities of Paenibacillus polymyxa against Zucchini yellow mosaic virus infestation in squash plants. Biology 2022, 11, 1150. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; El Enshasy, H. Plant growth promoting rhizobacteria (PGPR) as green bioinoculants: recent developments, constraints, and prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Loni, P.P.; Patil, J.U.; Phugare, S.S.; Bajekal, S.S. Purification and characterization of alkaline chitinase from Paenibacillus pasadenensis NCIM 5434. J. Basic Microbiol. 2014, 54, 1080–1089. [Google Scholar] [CrossRef]
- Mousavi, K.; Rajabpour, A.; Parizipour, G.; Yarahmadi, F. Insecticidal bioactive compounds derived from Cladosporium cladosporioides (Fresen.) GA de Vries and Acremonium zeylanicum (Petch) W. Gams & HC Evans. Journal of Plant Diseases and Protection 2023, 130, 543–549. [Google Scholar] [CrossRef]
- Collinson, N.P.; Mann, R.C.; Giri, K.; Malipatil, M.; Kaur, J.; Spangenberg, G.; Valenzuela, I. Novel bioassay to assess antibiotic effects of fungal endophytes on aphids. PLoS One 2020, 15, e0228813. [Google Scholar] [CrossRef]
- Heinen, R.; van der Sluijs, M.; Biere, A.; Harvey, J.A.; Bezemer, T.M. Plant community composition but not plant traits determine the outcome of soil legacy effects on plants and insects. J. Ecol. 2018, 106, 1217–1229. [Google Scholar] [CrossRef]
- Kos, M.; Tuijl, M.A.B.; de Roo, J.; Mulder, P.P.J.; Bezemer, T.M. Species-specific plant–soil feedback effects on above-ground plant–insect interactions. J. Ecol. 2015, 103, 904–914. [Google Scholar] [CrossRef]
- Heinen, R.; Biere, A.; Bezemer, T.M. Plant traits shape soil legacy effects on individual plant–insect interactions. Oikos 2020, 129, 261–273. [Google Scholar] [CrossRef]
- Huberty, M.; Steinauer, K.; Heinen, R.; Jongen, R.; Hannula, S.E.; Choi, Y.H.; Bezemer, T.M. Temporal changes in plant–soil feedback effects on microbial networks, leaf metabolomics and plant–insect interactions. J. Ecol. 2022, 110, 1328–1343. [Google Scholar] [CrossRef]
- Lyu, J.; Jin, L.; Jin, N.; Xie, J.; Xiao, X.; Hu, L.; Tang, Z.; Wu, Y.; Niu, L.; Yu, J. Effects of different vegetable rotations on fungal community structure in continuous tomato cropping matrix in greenhouse. Frontiers in Microbiology 2020, 11, 829. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, L.; Li, Y.; Wang, P.; Wang, C. Background nutrients and bacterial community evolution determine 13C-17β-estradiol mineralization in lake sediment microcosms. Sci. Total Environ. 2019, 651, 2304–2311. [Google Scholar] [CrossRef]
- Li, Z.; Zu, C.; Wang, C.; Yang, J.; Yu, H.; Wu, H. Different responses of rhizosphere and non-rhizosphere soil microbial communities to consecutive Piper nigrum L. monoculture. Scientific Reports 2016, 6. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 1–17. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
β-diversity analysis of rhizosphere microbiota. (A) PCoA plots of the unweighted UniFrac distance among groups of bacteria. (B) PLS-DA of OTUs among groups of bacteria. (C) PCoA plots of the unweighted UniFrac distance among groups of fungi. (D) PLS-DA of OTUs among groups of fungi.
Figure 1.
β-diversity analysis of rhizosphere microbiota. (A) PCoA plots of the unweighted UniFrac distance among groups of bacteria. (B) PLS-DA of OTUs among groups of bacteria. (C) PCoA plots of the unweighted UniFrac distance among groups of fungi. (D) PLS-DA of OTUs among groups of fungi.
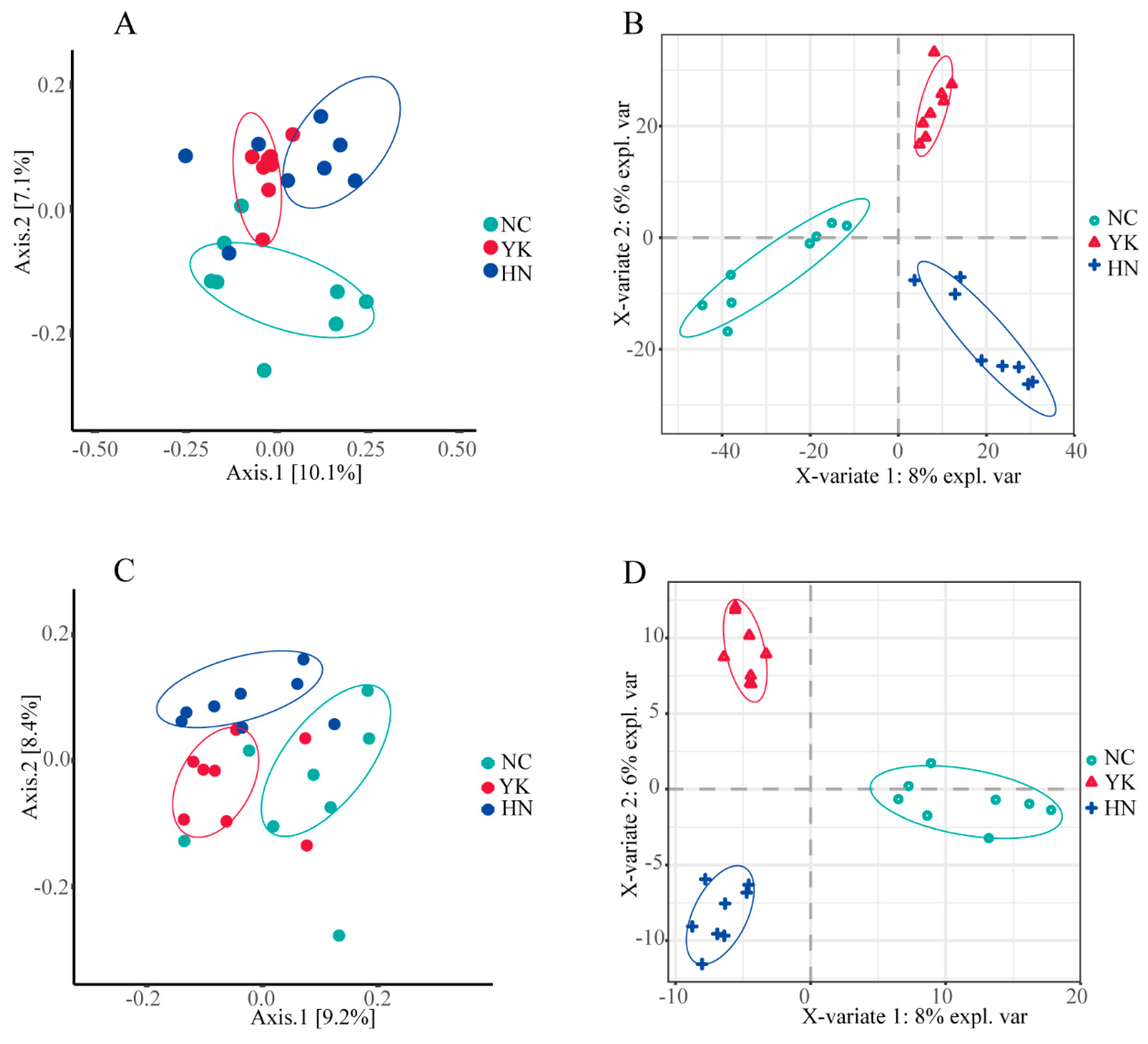
Figure 2.
The relative abundance of soil bacterial and fungal communities with average proportions greater than 0.5%.
Figure 2.
The relative abundance of soil bacterial and fungal communities with average proportions greater than 0.5%.
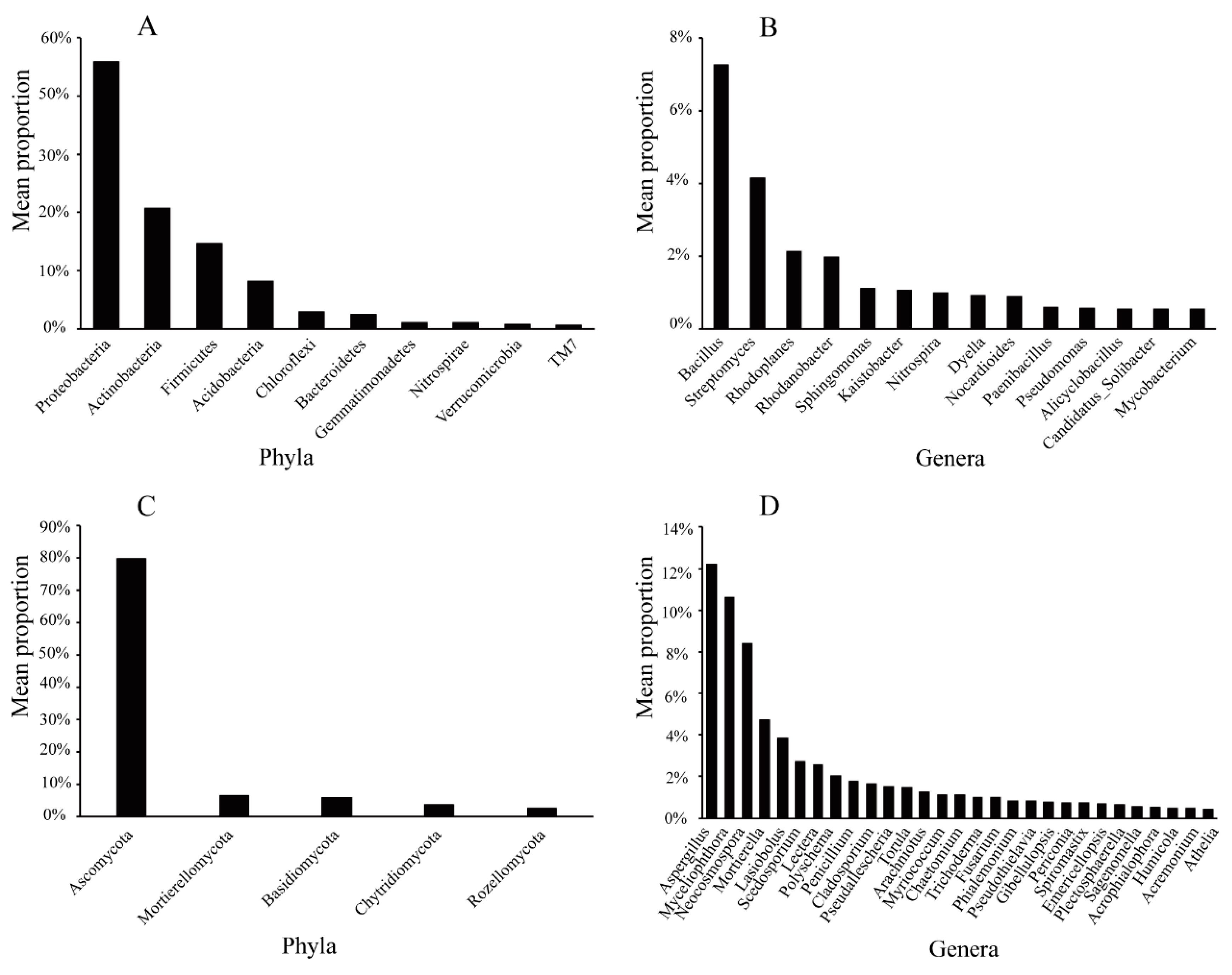
Figure 3.
LEfSe analysis of characteristic microbiota species among groups (LDA > 3, P < 0.05). (A) LEfSe showing differentially abundant bacterial taxa. (B) LEfSe showing differentially abundant fungal taxa. NC: susceptible papaya rhizosphere soil group; YK: susceptible papaya rhizosphere soil group; HN: Resistant papaya rhizosphere soil group.
Figure 3.
LEfSe analysis of characteristic microbiota species among groups (LDA > 3, P < 0.05). (A) LEfSe showing differentially abundant bacterial taxa. (B) LEfSe showing differentially abundant fungal taxa. NC: susceptible papaya rhizosphere soil group; YK: susceptible papaya rhizosphere soil group; HN: Resistant papaya rhizosphere soil group.
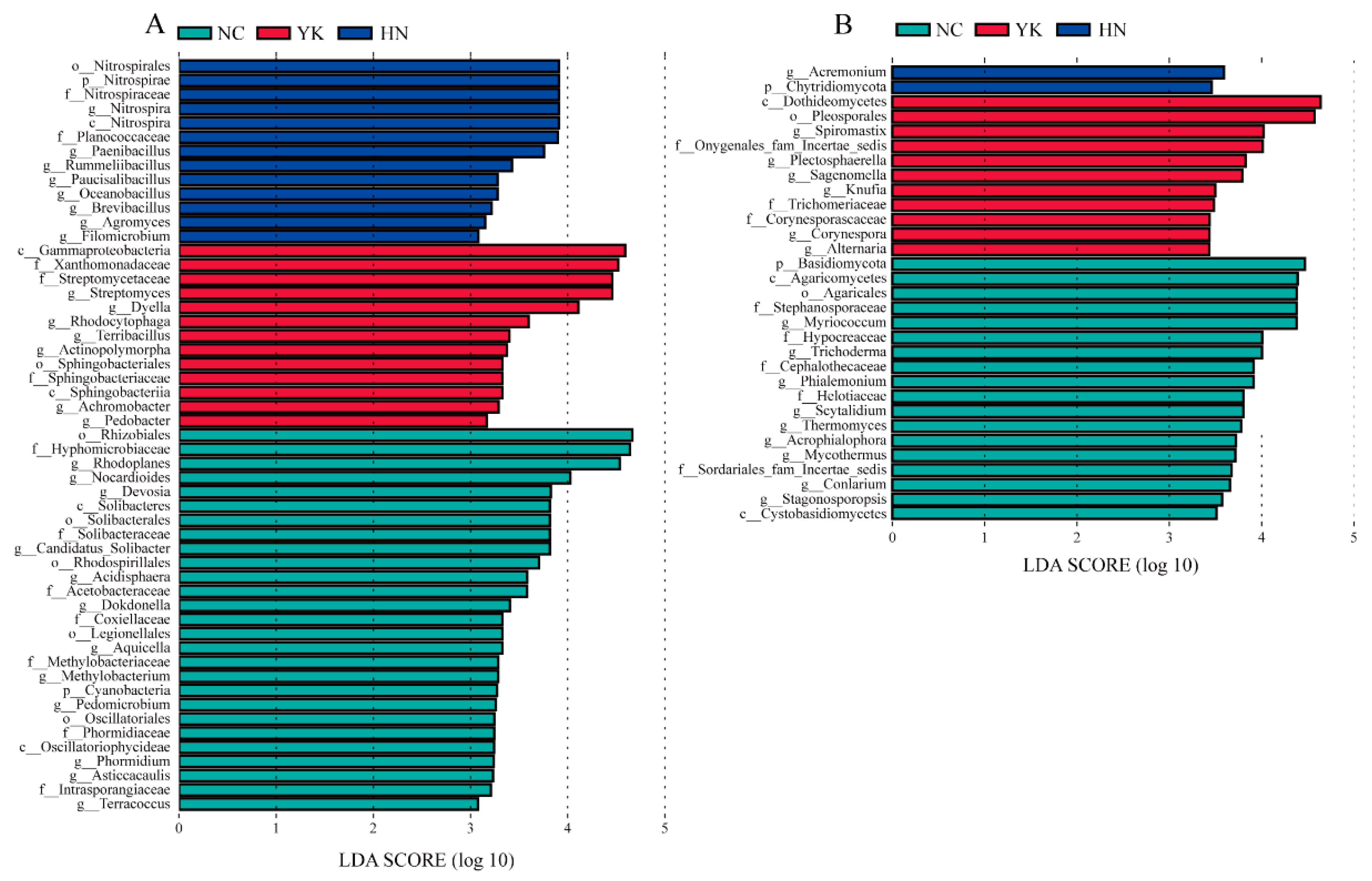
Figure 4.
Redundancy analysis (RDA) and Spearman’s rank correlation heatmap of phyla (P < 0.05 *, P < 0.01**). (A) and (B) show the RDA and heatmap of bacteria, respectively; (C) and (D) show the RDA and heatmap of fungi, respectively; AK, available potassium; AP, available phosphorus; AN, hydrolytic nitrogen; pH, soil pH; SOM, soil organic matter.
Figure 4.
Redundancy analysis (RDA) and Spearman’s rank correlation heatmap of phyla (P < 0.05 *, P < 0.01**). (A) and (B) show the RDA and heatmap of bacteria, respectively; (C) and (D) show the RDA and heatmap of fungi, respectively; AK, available potassium; AP, available phosphorus; AN, hydrolytic nitrogen; pH, soil pH; SOM, soil organic matter.
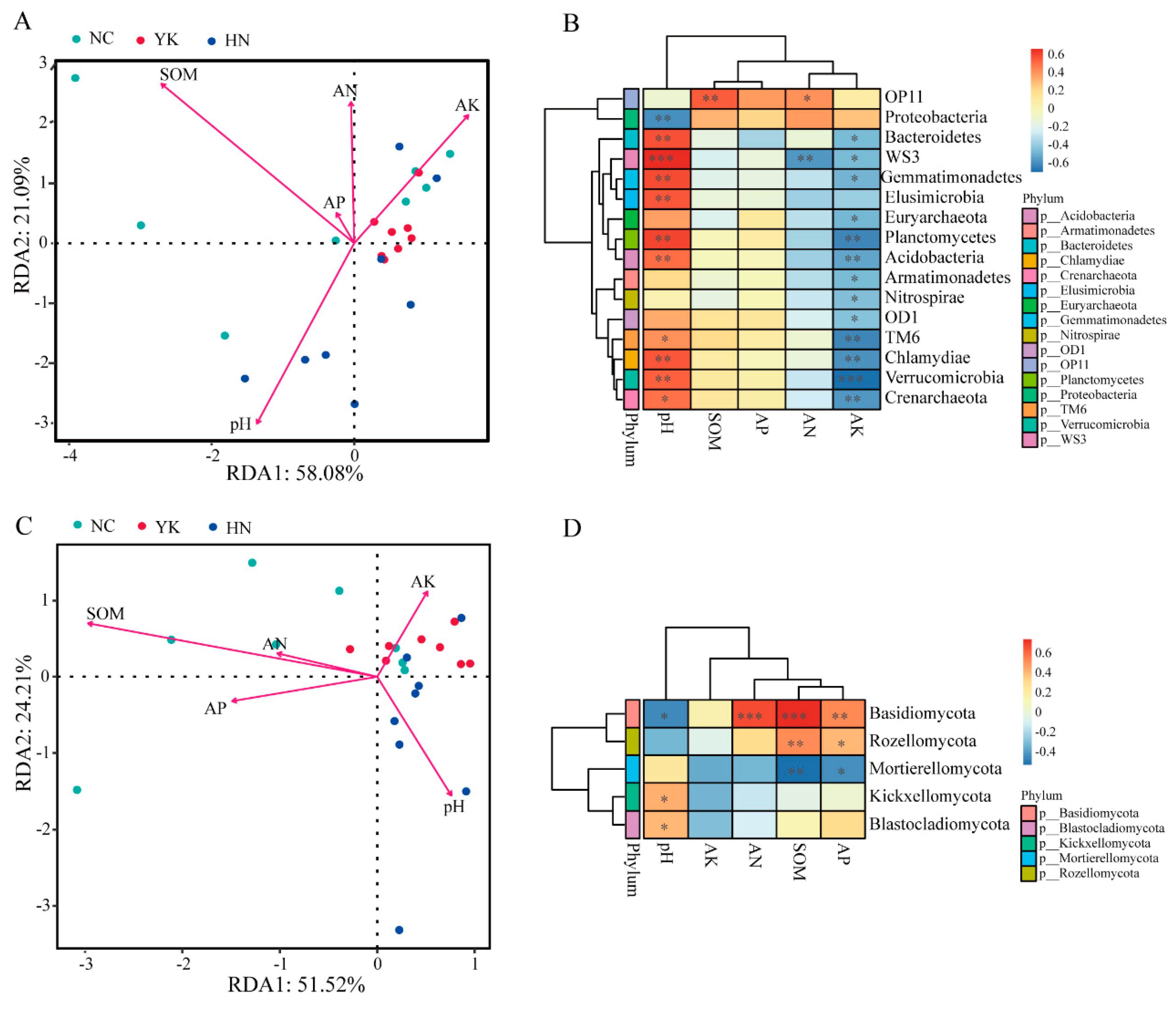

Table 1.
Classification of amplicon sequence variants.
| Taxonomic level | 16S | ITS | ||
|---|---|---|---|---|
| Number of unique features |
Percent of classified reads |
Number of unique features |
Percent of classified reads |
|
| Domain | 2 | 100.00 | 5 | 100 |
| Phylum | 41 | 99.84 | 11 | 93.56 |
| Class | 121 | 99.61 | 31 | 88.47 |
| Order | 192 | 95.43 | 75 | 88.20 |
| Family | 241 | 81.02 | 163 | 86.39 |
| Genus | 411 | 36.14 | 314 | 74.05 |
| Species | 238 | 11.87 | 424 | 43.95 |
Table 2.
Number of microbial taxa at different taxonomic ranks in the soil of healthy resistant plants and symptomatic susceptible plants.
Table 2.
Number of microbial taxa at different taxonomic ranks in the soil of healthy resistant plants and symptomatic susceptible plants.
| Soil sample | Phylum | Class | Order | Family | Genus | Species | OTU | |
|---|---|---|---|---|---|---|---|---|
| Fungi | NC (Diseased plant) | 11 | 26 | 66 | 135 | 235 | 273 | 1205 |
| YK (Diseased plant) | 10 | 27 | 61 | 130 | 227 | 266 | 1034 | |
| HN (Healthy plant) | 11 | 26 | 62 | 124 | 207 | 260 | 1105 | |
| Bacteria | NC (Diseased plant) | 40 | 109 | 173 | 216 | 306 | 160 | 7193 |
| YK (Diseased plant) | 35 | 103 | 161 | 213 | 309 | 154 | 6733 | |
| HN (Healthy plant) | 39 | 111 | 171 | 217 | 317 | 175 | 8009 |
Table 3.
Alpha diversity indices of microbial communities in rhizosphere soil with healthy resistant and diseased papaya plants.
Table 3.
Alpha diversity indices of microbial communities in rhizosphere soil with healthy resistant and diseased papaya plants.
| Taxon group | Soil Sample | OTUs | Chao 1 index | Shannon diversity | Simpson index |
|---|---|---|---|---|---|
| Bacteria | NC | 1652.88 ± 439.71a | 1661.62 ± 441.88 a | 8.80 ± 1.13 a | 0.99 ± 0.01 a |
| YK | 1524.5 ± 57.47 a | 1530.87 ± 56.96 a | 8.53 ± 0.31 a | 0.99 ± 0.05 a | |
| HN | 1768.5 ± 334.14 a | 1781.21 ± 336.21 a | 9.00 ± 0.81 a | 0.99 ± 0.01 a | |
| Fungi | NC | 276.63 ± 41.64 a | 276.63 ±41.64 a | 5.64 ± 0.64 a | 0.95 ± 0.34 a |
| YK | 262.38 ± 35.64 a | 262.38 ±35.64 a | 5.44 ± 0.44 a | 0.94 ± 0.04 a | |
| HN | 273.38 ±36.35 a | 273.38 ± 36.35 a | 5.35 ± 0.33 a | 0.94 ± 0.24 a |
Values are mean ± standard deviation. Different lowercase letters within a column for the same factor indicate significant differences according to a Tukey test (P < 0.05).
Table 4.
Analysis of β-diversity between pairs of soil samples (n = 16).
| Comparison groups | Bray-Curtis anosim-pairwise | Unweighted UniFrac anosim-pairwise | |||
|---|---|---|---|---|---|
| R | P | R | P | ||
|
16S |
NC-YK | 0.40 | 0.003 | 0.31 | 0.004 |
| NC-HN | 0.23 | 0.025 | 0.17 | 0.047 | |
| HN-YK | 0.22 | 0.013 | 0.17 | 0.025 | |
|
ITS |
NC-YK | 0.28 | 0.004 | 0.15 | 0.021 |
| NC-HN | 0.19 | 0.013 | 0.31 | 0.002 | |
| HN-YK | 0.32 | 0.003 | 0.20 | 0.007 | |
Table 5.
Relative abundance (%) of bacterial taxa among rhizosphere samples.
| Taxonomic level | Taxon | NC | YK | HN |
|---|---|---|---|---|
| Phylum | Nitrospirae | 0.10 ± 0.17** | 0.92 ± 0.28** | 1.55 ± 0.49 |
| Gemmatimonadetes | 0.97 ± 0.45* | 1.07 ± 0.31 | 1.48 ± 0.49 | |
| Chloroflexi | 3.00 ± 0.73* | 2.49 ± 0.54 | 3.55 ± 0.94 | |
| TM7 | 0.52 ± 0.45 | 1.07 ± 0.41** | 0.44 ± 0.34 | |
| Genus | Nitrospira | 0.83 ± 0.13** | 0.79 ± 0.27** | 1.35 ± 0.41 |
| Paenibacillus | 0.35 ± 0.12** | 0.58 ± 0.19 | 0.83 ± 0.38 | |
| Rhodanobacter | 1.44 ± 0.89 | 3.17 ± 2.38* | 1.34 ± 0.83 | |
| Sphingomonas | 0.86 ± 0.26 | 1.70 ± 1.28* | 0.78 ± 0.46 | |
| Dyella | 0.42 ± 0.11 | 1.68 ± 1.04** | 0.65 ± 0.52 |
Data are mean ± SEM (n= 8). *P < 0.05, **P < 0.01 compared with HN. t-test.
Table 6.
Relative abundance (%) of fungal taxa among rhizosphere samples.
| Taxonomic level | Taxon | NC | YK | HN |
|---|---|---|---|---|
| Phylum | Glomeromycota | 0.01 ± 0.19* | 0.01 ± 0.012* | 0.47 ± 0.71 |
| Mortierellomycota | 2.63 ± 1.60* | 6.45 ± 5.89 | 8.58 ± 5.77 | |
| Basidiomycota | 2.03 ± 2.29* | 3.29 ± 2.02 | 2.75 ± 3.01 | |
| Genus | Neocosmospora | 7.78 ± 4.82 | 6.13 ± 2.73* | 11.36 ± 4.18 |
| Trichoderma | 1.82 ± 1.34* | 0.62 ± 0.59 | 0.61 ± 0.47 | |
| Polyschema | 0.26 ± 0.54 | 4.85 ± 5.60* | 1.03 ± 1.84 | |
| Acremonium | 0.21 ± 0.18** | 0.48 ± 0.43 | 0.71 ± 0.37 |
Data are mean ± SEM (n = 8). *P < 0.05 and **P < 0.01 compared with HN, t-test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



