Submitted:
28 September 2025
Posted:
29 September 2025
You are already at the latest version
Abstract
Ocean acidification (OA) threatens marine microbial communities that underpin global biogeochemical cycles and marine food webs, however, a systematic synthesis of research progress in this area remains limited. This study presents the first comprehensive bibliometric analysis of ocean acidification impacts on microbial ecology, analyzing 495 Web of Science publications (2005-2025) using CiteSpace to characterize the field’s evolution and identify emerging frontiers. Global collaboration spans 53 countries, led primarily by China, the United States, and Germany, with the GEOMAR Helmholtz Centre for Ocean Research prominent within institutional networks. The research focus has shifted from basic chemical parameters to complex ecosystem processes, with “responses” identified as the most active contemporary research frontier. Overall, the field has matured into a highly internationalized, interdisciplinary domain. We outline four strategic directions for future work: (1) integrating advanceds molecular technologies, including multi-omics and single-cell approaches, to resolve mechanisms; (2) expanding temporal and spatial scales through global observatory networks; (3) quantifying multiple-stressor interactions, particularly with warming and deoxygenation; and (4) connecting molecular processes to biogeochemical cycles at the ecosystem level. These findings provide a data-driven roadmap for next-generation on OA–microbe interactions, essential for predicting marine ecosystem responses to accelerating environmental change and for informing evidence-based ocean conservation policy.
Keywords:
1. Introduction
2. Data and Methodology
2.1. Data Source and Retrieval Strategy
2.2. Bibliometric Analysis Framework
2.3. Quality Assessment and Validation
3. Results and Discussion
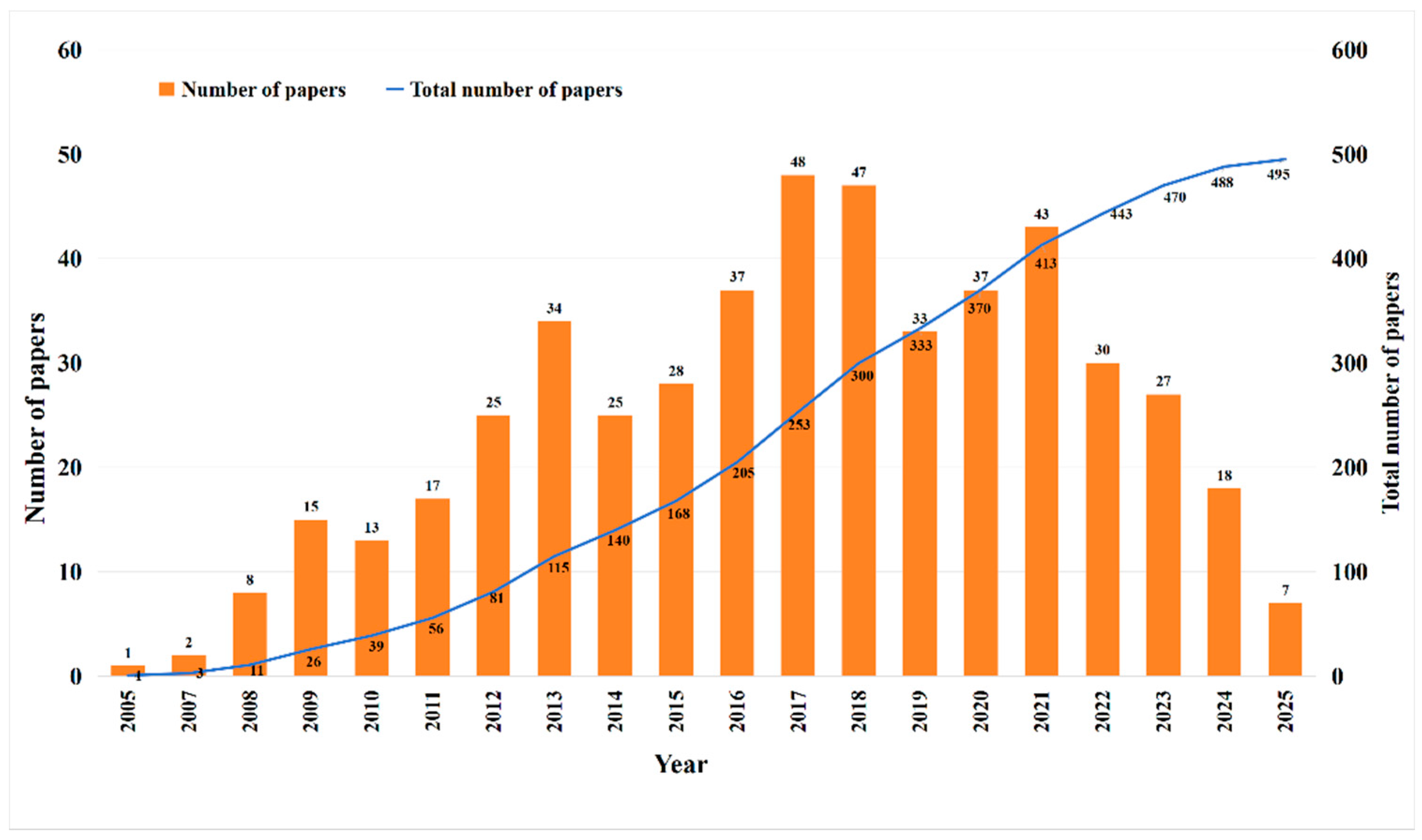
3.1. Publications and Annual Growth
3.2. The Interdisciplinary Character of the Field

3.3. Journal Co-citation Network Analysis

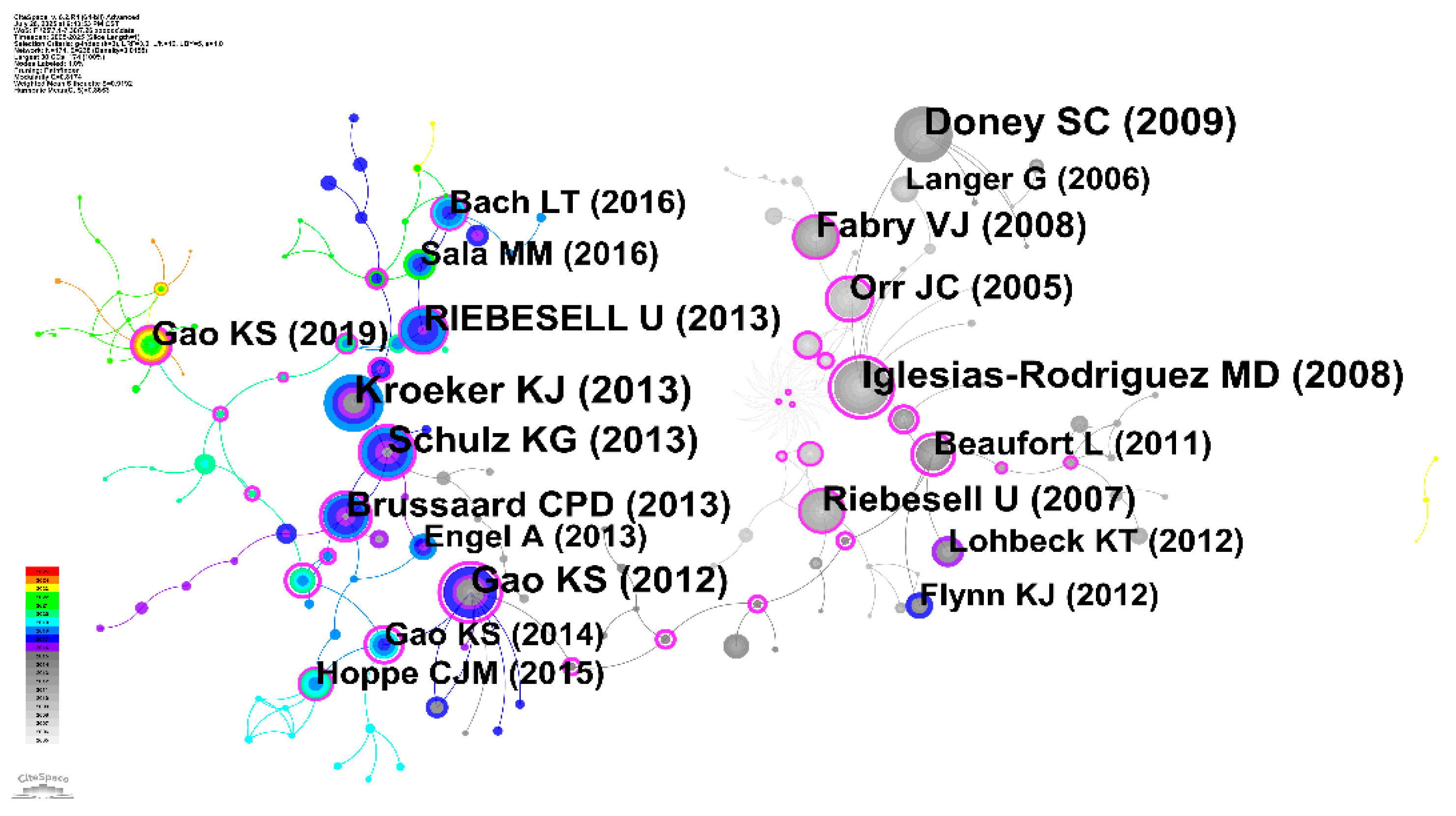
3.4 Literature Co-citation Network Analysis
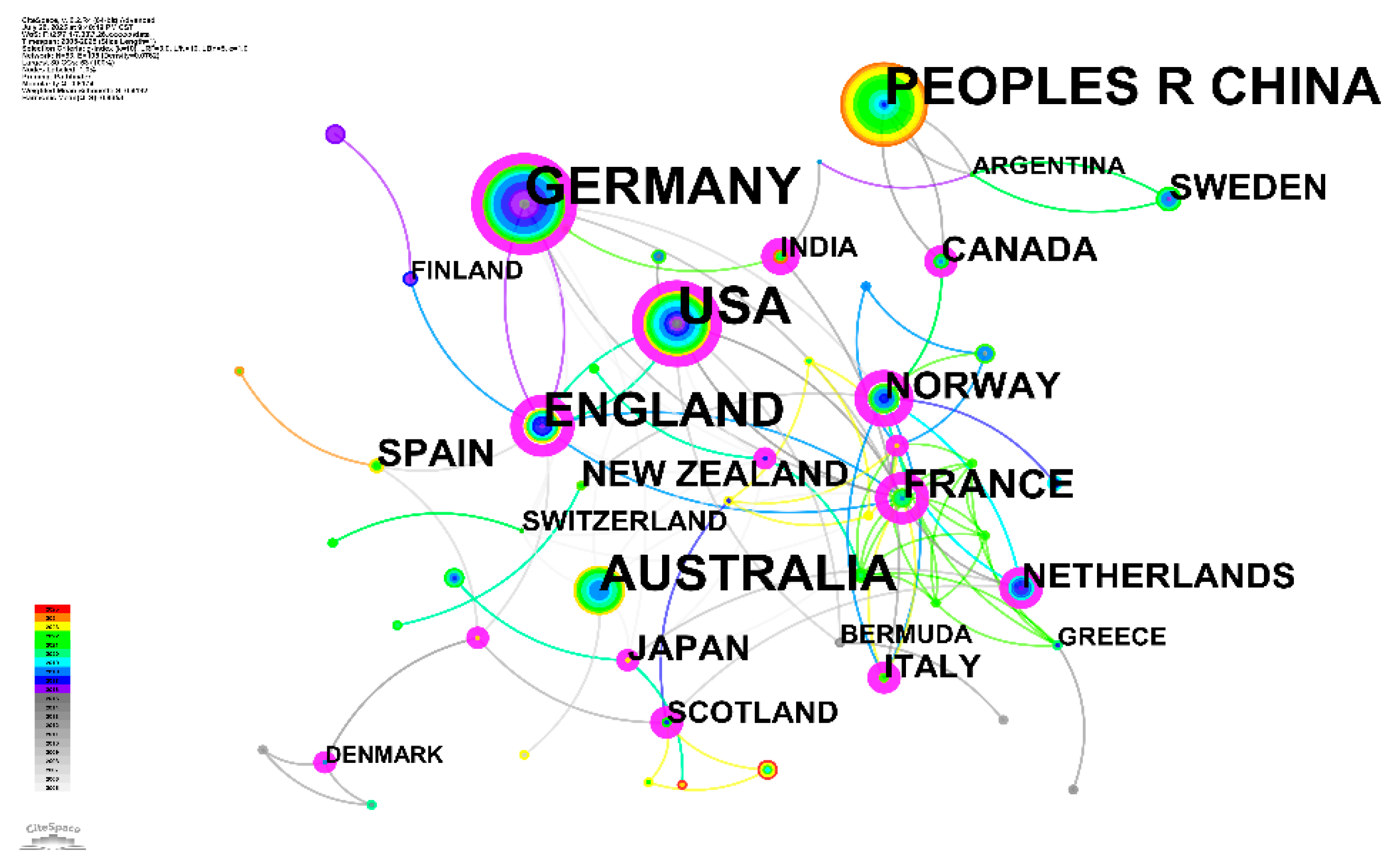
3.5. The Global Architecture of Scientific Production and Collaboration

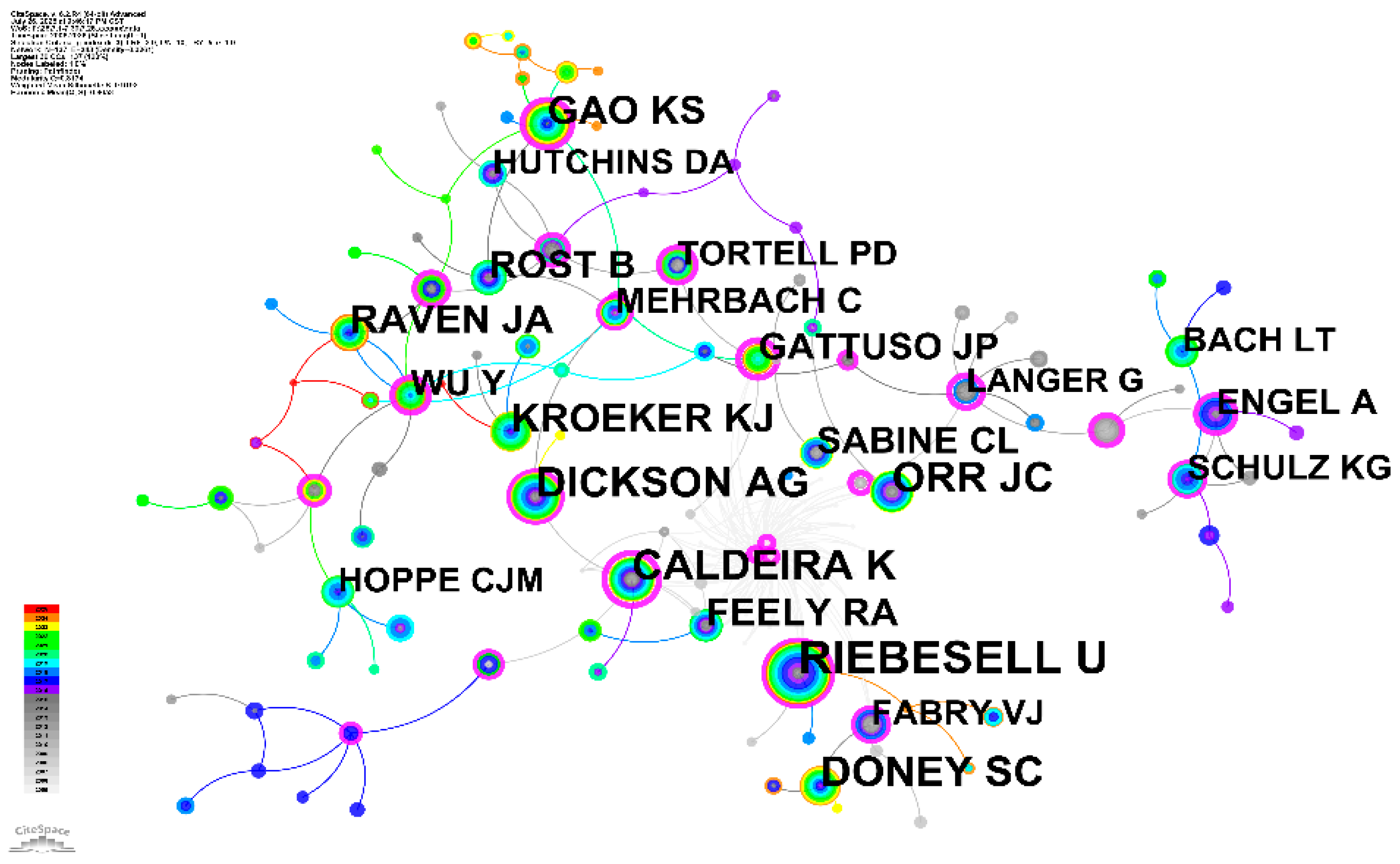
3.6 Influential Authors Network Analysis

3.7 Keyword Co-occurrence Network Analysis
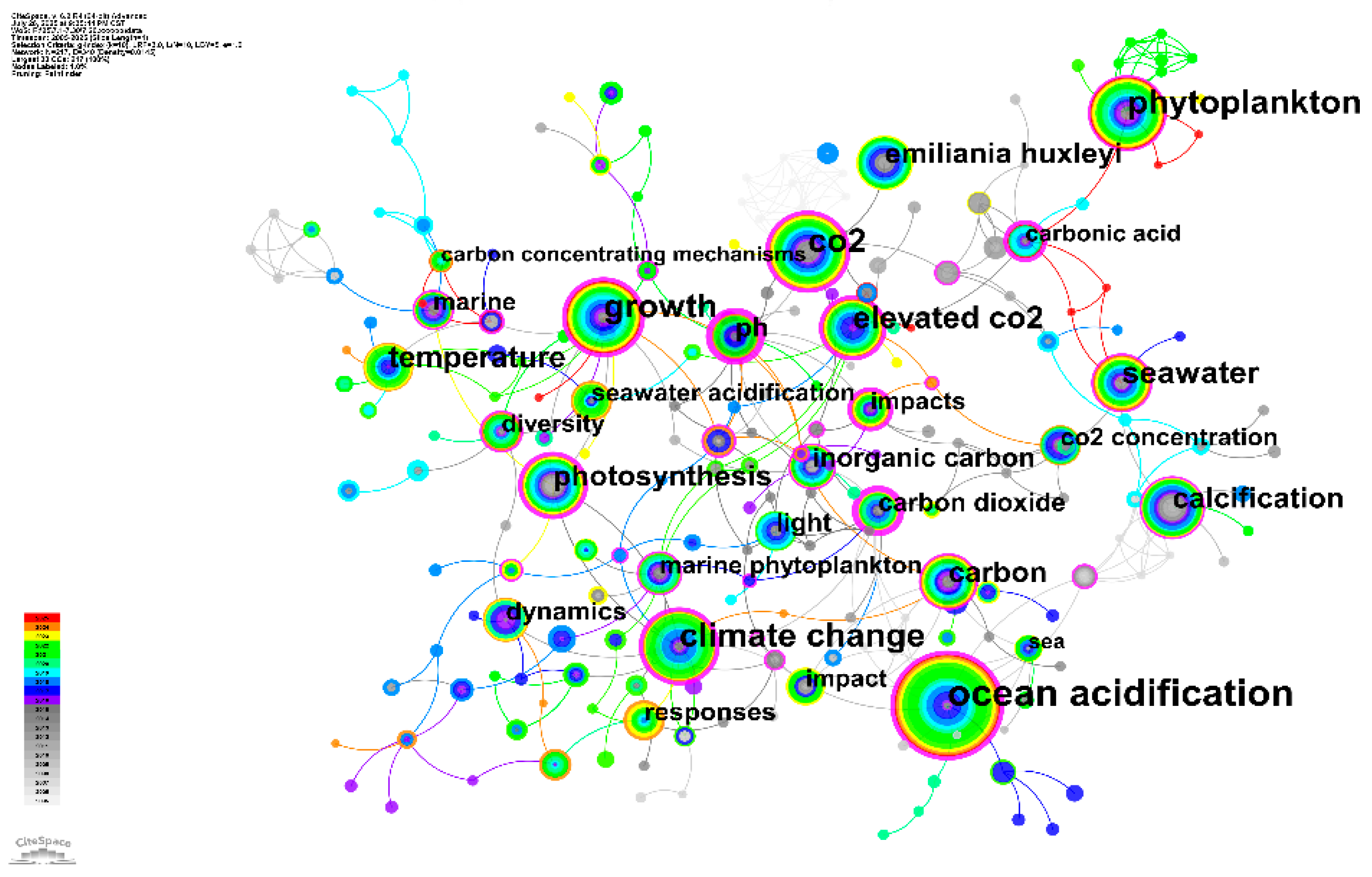
3.8 Keyword Clustering Map Analysis
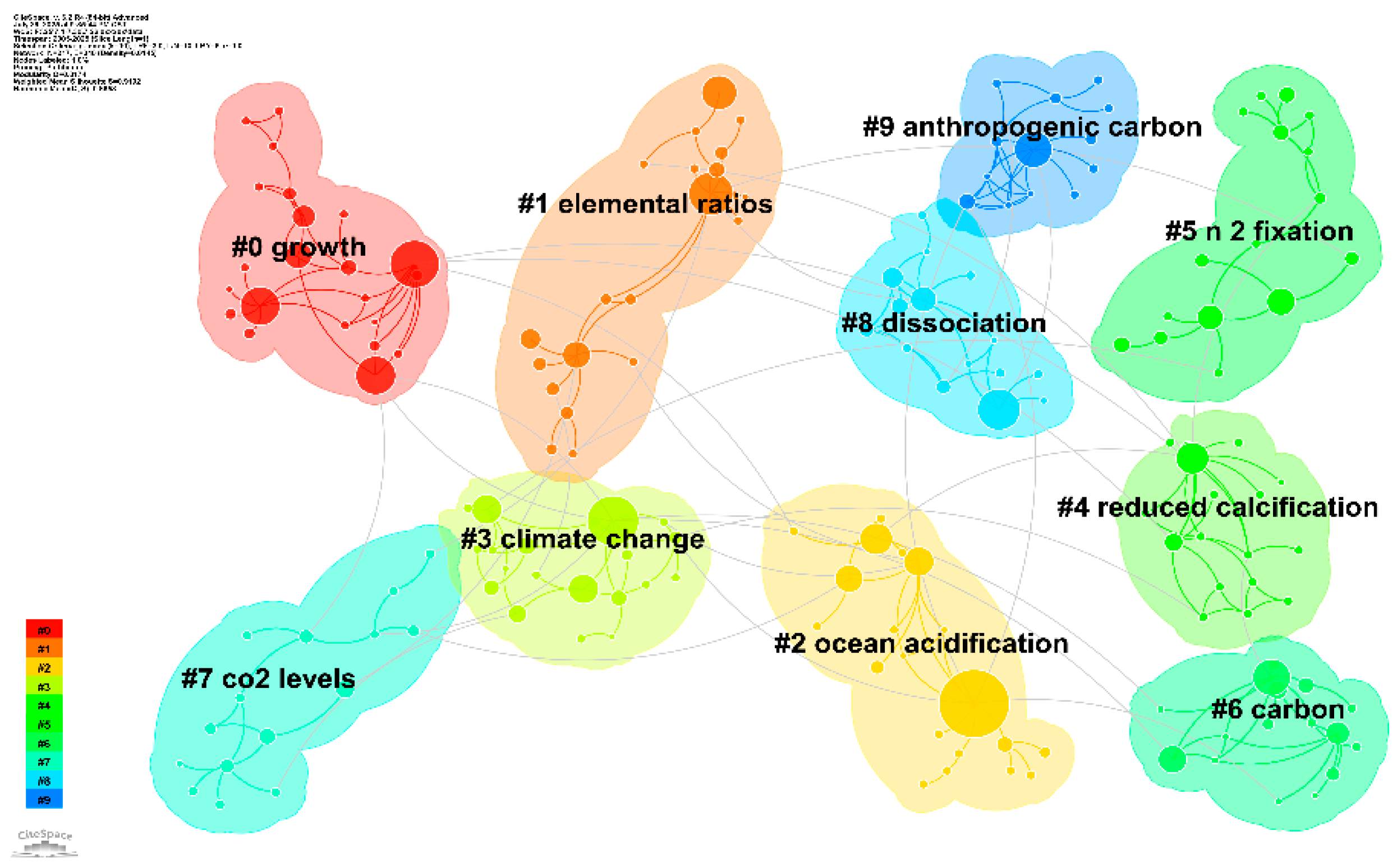
3.9 Temporal Evolution of Research Keyword
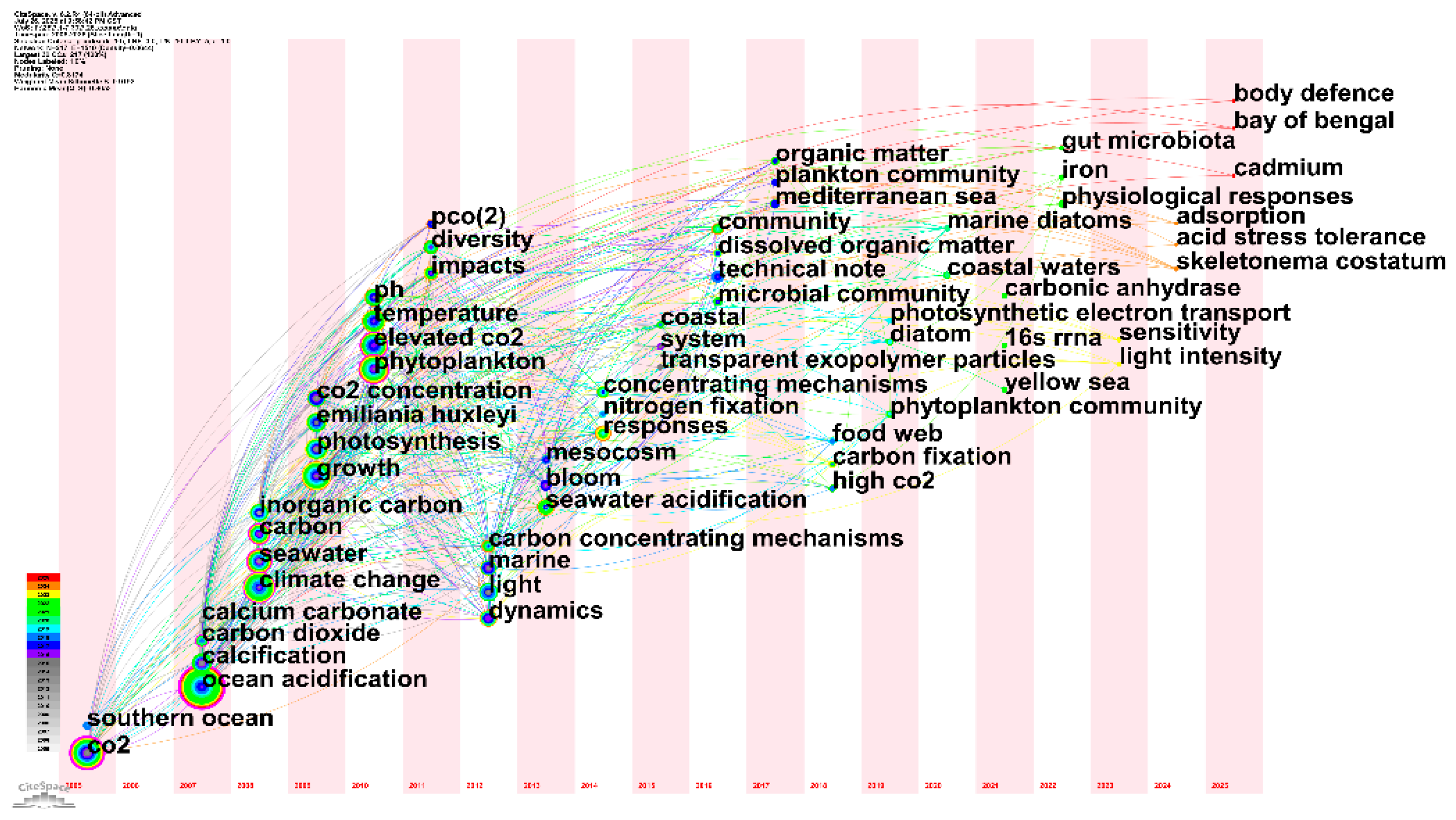
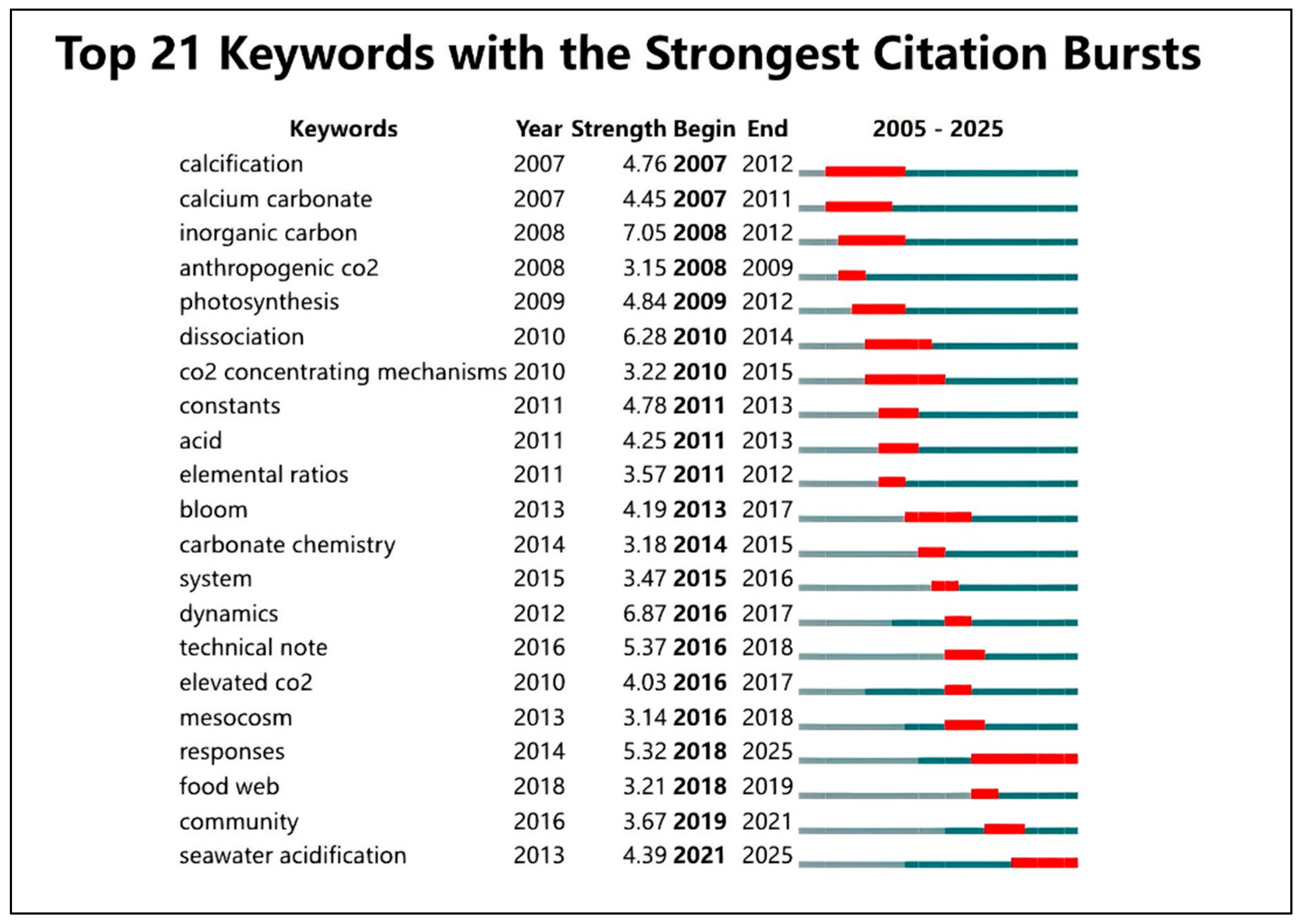
3.10 Citation Burst Analysis of Research Keywords
4. Prospects for OA and Microbial Ecology Research Field
4.1. Integration of Advanced Molecular Technologies and Systems Biology
4.2 Expansion of Temporal and Spatial Research Scales
4.3 Multiple Stressor Interaction and Climate Change Integration
4.4 Develop a New Generation of Experimental Systems
4.5 Computational Biology and Data Integration
4.6. Interdisciplinary Synthesis and Knowledge Integration
4.7. Policy-Science Interface Development
5. Limitations and Methodological Considerations
6. Conclusion
References
- Schönberg, C.H.L.; Fang, J.K.H.; Carreiro-Silva, M.; Tribollet, A.; Wisshak, M.; Norkko, J. Bioerosion: the other ocean acidification problem. ICES Journal of Marine Science 2017, 74, 895-925. [CrossRef]
- Gaylord, B.; Kroeker, K.J.; Sunday, J.M.; Anderson, K.M.; Barry, J.P.; Brown, N.E.; Connell, S.D.; Dupont, S.; Fabricius, K.E.; Hall-Spencer, J.M.; et al. Ocean acidification through the lens of ecological theory. Ecology 2015, 96, 3-15. [CrossRef]
- Doney, S.C.; Fabry, V.J.; Feely, R.A.; Kleypas, J.A. Ocean Acidification: The Other CO2Problem. Annual Review of Marine Science 2009, 1, 169-192. [CrossRef]
- Feely, R.A.; Sabine, C.L.; Lee, K.; Berelson, W.; Kleypas, J.; Fabry, V.J.; Millero, F.J. Impact of Anthropogenic CO2 on the CaCO3 System in the Oceans. Science 2004, 305, 362-366. [CrossRef]
- Orr, J.C.; Fabry, V.J.; Aumont, O.; Bopp, L.; Doney, S.C.; Feely, R.A.; Gnanadesikan, A.; Gruber, N.; Ishida, A.; Joos, F.; et al. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 2005, 437, 681-686. [CrossRef]
- Caldeira, K.; Wickett, M.E. Ocean model predictions of chemistry changes from carbon dioxide emissions to the atmosphere and ocean. Journal of Geophysical Research: Oceans 2005, 110. [CrossRef]
- Fabry, V.J.; Seibel, B.A.; Feely, R.A.; Orr, J.C. Impacts of ocean acidification on marine fauna and ecosystem processes. ICES Journal of Marine Science 2008, 65, 414-432. [CrossRef]
- Falkowski, P.G.; Fenchel, T.; Delong, E.F. The Microbial Engines That Drive Earth's Biogeochemical Cycles. Science 2008, 320, 1034-1039. [CrossRef]
- Arrigo, K.R. Marine microorganisms and global nutrient cycles. Nature 2004, 437, 349-355. [CrossRef]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary Production of the Biosphere: Integrating Terrestrial and Oceanic Components. Science 1998, 281, 237-240. [CrossRef]
- Madsen, E.L. Microorganisms and their roles in fundamental biogeochemical cycles. Current Opinion in Biotechnology 2011, 22, 456-464. [CrossRef]
- Azam, F.; Malfatti, F. Microbial structuring of marine ecosystems. Nature Reviews Microbiology 2007, 5, 782-791. [CrossRef]
- Wang, B.; Hua, L.; Mei, H.; Wu, X.; Kang, Y.; Zhao, N. Impact of Climate Change on the Dynamic Processes of Marine Environment and Feedback Mechanisms: An Overview. Archives of Computational Methods in Engineering 2024, 31, 3377-3408. [CrossRef]
- Hofmann, G.E.; Barry, J.P.; Edmunds, P.J.; Gates, R.D.; Hutchins, D.A.; Klinger, T.; Sewell, M.A. The Effect of Ocean Acidification on Calcifying Organisms in Marine Ecosystems: An Organism-to-Ecosystem Perspective. Annual Review of Ecology, Evolution, and Systematics 2010, 41, 127-147. [CrossRef]
- O'Brien, P.A.; Morrow, K.M.; Willis, B.L.; Bourne, D.G. Implications of Ocean Acidification for Marine Microorganisms from the Free-Living to the Host-Associated. Frontiers in Marine Science 2016, 3. [CrossRef]
- Liu, J.; Weinbauer, M.G.; Maier, C.; Dai, M.; Gattuso, J.P. Effect of ocean acidification on microbial diversity and on microbe-driven biogeochemistry and ecosystem functioning. Aquatic Microbial Ecology 2010, 61, 291-305. [CrossRef]
- Gao, K.; Gao, G.; Wang, Y.; Dupont, S. Impacts of ocean acidification under multiple stressors on typical organisms and ecological processes. Marine Life Science & Technology 2020, 2, 279-291. [CrossRef]
- Riebesell, U.; Zondervan, I.; Rost, B.; Tortell, P.D.; Zeebe, R.E.; Morel, F.M.M. Reduced calcification of marine plankton in response to increased atmospheric CO2. Nature 2000, 407, 364-367. [CrossRef]
- Meron, D.; Rodolfo-Metalpa, R.; Cunning, R.; Baker, A.C.; Fine, M.; Banin, E. Changes in coral microbial communities in response to a natural pH gradient. The ISME Journal 2012, 6, 1775-1785. [CrossRef]
- Piontek, J.; Lunau, M.; Händel, N.; Borchard, C.; Wurst, M.; Engel, A. Acidification increases microbial polysaccharide degradation in the ocean. Biogeosciences 2010, 7, 1615-1624. [CrossRef]
- Kuffner, I.B.; Andersson, A.J.; Jokiel, P.L.; Rodgers, K.u.S.; Mackenzie, F.T. Decreased abundance of crustose coralline algae due to ocean acidification. Nature Geoscience 2007, 1, 114-117. [CrossRef]
- Brussaard, C.P.D.; Noordeloos, A.A.M.; Witte, H.; Collenteur, M.C.J.; Schulz, K.; Ludwig, A.; Riebesell, U. Arctic microbial community dynamics influenced by elevated CO2levels. Biogeosciences 2013, 10, 719-731. [CrossRef]
- Nelson, K.S.; Baltar, F.; Lamare, M.D.; Morales, S.E. Ocean acidification affects microbial community and invertebrate settlement on biofilms. Scientific Reports 2020, 10. [CrossRef]
- Gunderson, A.R.; Armstrong, E.J.; Stillman, J.H. Multiple Stressors in a Changing World: The Need for an Improved Perspective on Physiological Responses to the Dynamic Marine Environment. Annual Review of Marine Science 2016, 8, 357-378. [CrossRef]
- Griffen, B.D.; Belgrad, B.A.; Cannizzo, Z.J.; Knotts, E.R.; Hancock, E.R. Rethinking our approach to multiple stressor studies in marine environments. Marine Ecology Progress Series 2016, 543, 273-281. [CrossRef]
- Riebesell, U.; Gattuso, J.-P. Lessons learned from ocean acidification research. Nature Climate Change 2014, 5, 12-14. [CrossRef]
- Wang, Y.; Zhang, R.; Yang, Y.; Tu, Q.; Zhou, J.; Jiao, N. Ocean acidification altered microbial functional potential in the Arctic Ocean. Limnology and Oceanography 2023, 68. [CrossRef]
- Ridgwell, A.; Schmidt, D.N.; Turley, C.; Brownlee, C.; Maldonado, M.T.; Tortell, P.; Young, J.R. From laboratory manipulations to Earth system models: scaling calcification impacts of ocean acidification. Biogeosciences 2009, 6, 2611-2623. [CrossRef]
- Hu, C.; Li, X.; He, M.; Jiang, P.; Long, A.; Xu, J. Effect of Ocean Acidification on Bacterial Metabolic Activity and Community Composition in Oligotrophic Oceans, Inferred From Short-Term Bioassays. Frontiers in Microbiology 2021, 12. [CrossRef]
- Witt, V.; Wild, C.; Anthony, K.R.N.; Diaz-Pulido, G.; Uthicke, S. Effects of ocean acidification on microbial community composition of, and oxygen fluxes through, biofilms from the Great Barrier Reef. Environmental Microbiology 2011, 13, 2976-2989. [CrossRef]
- Webster, N.S.; Negri, A.P.; Botté, E.S.; Laffy, P.W.; Flores, F.; Noonan, S.; Schmidt, C.; Uthicke, S. Host-associated coral reef microbes respond to the cumulative pressures of ocean warming and ocean acidification. Scientific Reports 2016, 6. [CrossRef]
- Zhang, X.; Chen, S.; Yao, P.; Han, J.; Jin, R. Knowledge Structure and Evolution of Wetland Plant Diversity Research: Visual Exploration Based on CiteSpace. Biology 2025, 14. [CrossRef]
- Chen, C. CiteSpace II: Detecting and visualizing emerging trends and transient patterns in scientific literature. Journal of the American Society for Information Science and Technology 2005, 57, 359-377. [CrossRef]
- Chen, C. Science Mapping: A Systematic Review of the Literature. Journal of Data and Information Science 2017, 2, 1-40. [CrossRef]
- Wang, B.; Pan, S.-Y.; Ke, R.-Y.; Wang, K.; Wei, Y.-M. An overview of climate change vulnerability: a bibliometric analysis based on Web of Science database. Natural Hazards 2014, 74, 1649-1666. [CrossRef]
- Wang, Q.; Wang, J.; Xue, M.; Zhang, X. Characteristics and Trends of Ocean Remote Sensing Research from 1990 to 2020: A Bibliometric Network Analysis and Its Implications. Journal of Marine Science and Engineering 2022, 10. [CrossRef]
- Wang, Z.; Zhao, Y.; Wang, B. A bibliometric analysis of climate change adaptation based on massive research literature data. Journal of Cleaner Production 2018, 199, 1072-1082. [CrossRef]
- Zhou, C.; Bi, R.; Su, C.; Liu, W.; Wang, T. The emerging issue of microplastics in marine environment: A bibliometric analysis from 2004 to 2020. Marine Pollution Bulletin 2022, 179. [CrossRef]
- Mao, N.; Zhu, Y.; Wang, J.; Shen, X.; Hou, J.; Miao, L. Adaptive Evolution of Marine Organisms: A Bibliometric Analysis Based on CiteSpace. Water 2023, 15. [CrossRef]
- Geng, Y.; Zhang, X.; Gao, J.; Yan, Y.; Chen, L. Bibliometric analysis of sustainable tourism using CiteSpace. Technological Forecasting and Social Change 2024, 202. [CrossRef]
- Jia, F.; Sun, D.; Looi, C.-k. Artificial Intelligence in Science Education (2013–2023): Research Trends in Ten Years. Journal of Science Education and Technology 2023, 33, 94-117. [CrossRef]
- Pranckutė, R. Web of Science (WoS) and Scopus: The Titans of Bibliographic Information in Today’s Academic World. Publications 2021, 9. [CrossRef]
- Chen, C. Searching for intellectual turning points: Progressive knowledge domain visualization. Proceedings of the National Academy of Sciences 2004, 101, 5303-5310. [CrossRef]
- Glanzel, W.; Chen, C.; Song, M. Visualizing a field of research: A methodology of systematic scientometric reviews. Plos One 2019, 14. [CrossRef]
- Wei, J.; Liang, G.; Alex, J.; Zhang, T.; Ma, C. Research Progress of Energy Utilization of Agricultural Waste in China: Bibliometric Analysis by Citespace. Sustainability 2020, 12. [CrossRef]
- Breitbarth, E.; Bellerby, R.J.; Neill, C.C.; Ardelan, M.V.; Meyerhöfer, M.; Zöllner, E.; Croot, P.L.; Riebesell, U. Ocean acidification affects iron speciation during a coastal seawater mesocosm experiment. Biogeosciences 2010, 7, 1065-1073. [CrossRef]
- Hancock, A.M.; King, C.K.; Stark, J.S.; McMinn, A.; Davidson, A.T. Effects of ocean acidification on Antarctic marine organisms: A meta-analysis. Ecology and Evolution 2020, 10, 4495-4514. [CrossRef]
- Wang, T.; Jin, P.; Wells, M.L.; Trick, C.G.; Gao, K. Insensitivities of a subtropical productive coastal plankton community and trophic transfer to ocean acidification: Results from a microcosm study. Marine Pollution Bulletin 2019, 141, 462-471. [CrossRef]
- Lin, S. Phosphate limitation and ocean acidification co-shape phytoplankton physiology and community structure. Nature Communications 2023, 14. [CrossRef]
- Cheriyan, E.; Kumar, B.S.K.; Gupta, G.V.M.; Rao, D.B. Implications of ocean acidification on micronutrient elements-iron, copper and zinc, and their primary biological impacts: A review. Marine Pollution Bulletin 2024, 199. [CrossRef]
- Salomon, A.K.; McKechnie, I. Insights Gained from Including People in Our Models of Nature and Modes of Science. Annual Review of Marine Science 2025, 17, 167-191. [CrossRef]
- Robinson, A.R.; McCarthy, J.J.; Rothschild, B.J. Interdisciplinary ocean science is evolving and a systems approach is essential. Journal of Marine Systems 1999, 22, 231-239. [CrossRef]
- Révelard, A.; Tintoré, J.; Verron, J.; Bahurel, P.; Barth, J.A.; Belbéoch, M.; Benveniste, J.; Bonnefond, P.; Chassignet, E.P.; Cravatte, S.; et al. Ocean Integration: The Needs and Challenges of Effective Coordination Within the Ocean Observing System. Frontiers in Marine Science 2022, 8. [CrossRef]
- Zeebe, R.E. History of Seawater Carbonate Chemistry, Atmospheric CO2, and Ocean Acidification. Annual Review of Earth and Planetary Sciences 2012, 40, 141-165. [CrossRef]
- Osborne, E.; Hu, X.; Hall, E.R.; Yates, K.; Vreeland-Dawson, J.; Shamberger, K.; Barbero, L.; Martin Hernandez-Ayon, J.; Gomez, F.A.; Hicks, T.; et al. Ocean acidification in the Gulf of Mexico: Drivers, impacts, and unknowns. Progress in Oceanography 2022, 209. [CrossRef]
- Wu, H.C.; Dissard, D.; Douville, E.; Blamart, D.; Bordier, L.; Tribollet, A.; Le Cornec, F.; Pons-Branchu, E.; Dapoigny, A.; Lazareth, C.E. Surface ocean pH variations since 1689 CE and recent ocean acidification in the tropical South Pacific. Nature Communications 2018, 9. [CrossRef]
- Johnson, M.D.; Rodriguez Bravo, L.M.; O’Connor, S.E.; Varley, N.F.; Altieri, A.H. pH Variability Exacerbates Effects of Ocean Acidification on a Caribbean Crustose Coralline Alga. Frontiers in Marine Science 2019, 6. [CrossRef]
- Flynn, K.J.; Blackford, J.C.; Baird, M.E.; Raven, J.A.; Clark, D.R.; Beardall, J.; Brownlee, C.; Fabian, H.; Wheeler, G.L. Changes in pH at the exterior surface of plankton with ocean acidification. Nature Climate Change 2012, 2, 510-513. [CrossRef]
- Kerrison, P.; Hall-Spencer, J.M.; Suggett, D.J.; Hepburn, L.J.; Steinke, M. Assessment of pH variability at a coastal CO2 vent for ocean acidification studies. Estuarine, Coastal and Shelf Science 2011, 94, 129-137. [CrossRef]
- Lowe, A.T.; Bos, J.; Ruesink, J. Ecosystem metabolism drives pH variability and modulates long-term ocean acidification in the Northeast Pacific coastal ocean. Scientific Reports 2019, 9. [CrossRef]
- Fennel, K.; Mattern, J.P.; Doney, S.C.; Bopp, L.; Moore, A.M.; Wang, B.; Yu, L. Ocean biogeochemical modelling. Nature Reviews Methods Primers 2022, 2. [CrossRef]
- Steffen, W.; Leinfelder, R.; Zalasiewicz, J.; Waters, C.N.; Williams, M.; Summerhayes, C.; Barnosky, A.D.; Cearreta, A.; Crutzen, P.; Edgeworth, M.; et al. Stratigraphic and Earth System approaches to defining the Anthropocene. Earth's Future 2016, 4, 324-345. [CrossRef]
- Zunino, S.; Libralato, S.; Melaku Canu, D.; Prato, G.; Solidoro, C. Impact of Ocean Acidification on Ecosystem Functioning and Services in Habitat-Forming Species and Marine Ecosystems. Ecosystems 2021, 24, 1561-1575. [CrossRef]
- Jin, P.; Hutchins, D.A.; Gao, K. The Impacts of Ocean Acidification on Marine Food Quality and Its Potential Food Chain Consequences. Frontiers in Marine Science 2020, 7. [CrossRef]
- Zhou, J.; Zheng, Y.; Hou, L.; Qi, L.; Mao, T.; Yin, G.; Liu, M. Nitrogen input modulates the effects of coastal acidification on nitrification and associated N2O emission. Water Research 2024, 261, 122041. [CrossRef]
- Hartin, C.A.; Bond-Lamberty, B.; Patel, P.; Mundra, A. Ocean acidification over the next three centuries using a simple global climate carbon-cycle model: projections and sensitivities. Biogeosciences 2016, 13, 4329-4342. [CrossRef]
- 4329-4342.
- DeVries, T. The Ocean Carbon Cycle. Annual Review of Environment and Resources 2022, 47, 317-341. [CrossRef]
- Ferderer, A.; Schulz, K.G.; Riebesell, U.; Baker, K.G.; Chase, Z.; Bach, L.T. Investigating the effect of silicate- and calcium-based ocean alkalinity enhancement on diatom silicification. Biogeosciences 2024, 21, 2777-2794. [CrossRef]
- Velthuis, M.; Keuskamp, J.A.; Bakker, E.S.; Boersma, M.; Sommer, U.; van Donk, E.; Van de Waal, D.B. Differential effects of elevated pCO2 and warming on marine phytoplankton stoichiometry. Limnology and Oceanography 2022, 67, 598-607. [CrossRef]
- Uchikawa, J.; Zeebe, R.E. Influence of terrestrial weathering on ocean acidification and the next glacial inception. Geophysical Research Letters 2008, 35. [CrossRef]
- Monteiro, T.; Batista, M.; Kerr, R. Spatial variability of marine carbonate system along the Drake Passage and northern Antarctic Peninsula during the austral summer. Marine Chemistry 2025, 271. [CrossRef]
- Qiu, L.; Esposito, M.; Martínez-Cabanas, M.; Achterberg, E.P.; Li, Q. Autonomous high-frequency time-series observations of total alkalinity in dynamic estuarine waters. Marine Chemistry 2023, 257. [CrossRef]
- Vodopivec, M. A numerical model sheds new light on the effects of ocean acidification on an indicator species. Global Change Biology 2024, 30. [CrossRef]
- Olson, E.M.; John, J.G.; Dunne, J.P.; Stock, C.A.; Drenkard, E.J. Potential for Regional Resilience to Ocean Warming and Acidification Extremes: Projected Vulnerability Under Contrasting Pathways and Thresholds. Global Change Biology 2025, 31. [CrossRef]
- Tagliabue, A.; Bopp, L.; Gehlen, M. The response of marine carbon and nutrient cycles to ocean acidification: Large uncertainties related to phytoplankton physiological assumptions. Global Biogeochemical Cycles 2011, 25, n/a-n/a. [CrossRef]
- Pritchard, H.D.; Ligtenberg, S.R.M.; Fricker, H.A.; Vaughan, D.G.; van den Broeke, M.R.; Padman, L. Antarctic ice-sheet loss driven by basal melting of ice shelves. Nature 2012, 484, 502-505. [CrossRef]
- Riebesell, U.; Schulz, K.G.; Bellerby, R.G.J.; Botros, M.; Fritsche, P.; Meyerhöfer, M.; Neill, C.; Nondal, G.; Oschlies, A.; Wohlers, J.; et al. Enhanced biological carbon consumption in a high CO2 ocean. Nature 2007, 450, 545-548. [CrossRef]
- Riebesell, U.; Czerny, J.; von Bröckel, K.; Boxhammer, T.; Büdenbender, J.; Deckelnick, M.; Fischer, M.; Hoffmann, D.; Krug, S.A.; Lentz, U.; et al. Technical Note: A mobile sea-going mesocosm system – new opportunities for ocean change research. Biogeosciences 2013, 10, 1835-1847. [CrossRef]
- Kroeker, K.J.; Kordas, R.L.; Crim, R.; Hendriks, I.E.; Ramajo, L.; Singh, G.S.; Duarte, C.M.; Gattuso, J.P. Impacts of ocean acidification on marine organisms: quantifying sensitivities and interaction with warming. Global Change Biology 2013, 19, 1884-1896. [CrossRef]
- Iglesias-Rodriguez, M.D.; Halloran, P.R.; Rickaby, R.E.M.; Hall, I.R.; Colmenero-Hidalgo, E.; Gittins, J.R.; Green, D.R.H.; Tyrrell, T.; Gibbs, S.J.; von Dassow, P.; et al. Phytoplankton Calcification in a High-CO2 World. Science 2008, 320, 336-340. [CrossRef]
- Beaufort, L.; Probert, I.; de Garidel-Thoron, T.; Bendif, E.M.; Ruiz-Pino, D.; Metzl, N.; Goyet, C.; Buchet, N.; Coupel, P.; Grelaud, M.; et al. Sensitivity of coccolithophores to carbonate chemistry and ocean acidification. Nature 2011, 476, 80-83. [CrossRef]
- Gao, K.; Xu, J.; Gao, G.; Li, Y.; Hutchins, D.A.; Huang, B.; Wang, L.; Zheng, Y.; Jin, P.; Cai, X.; et al. Rising CO2 and increased light exposure synergistically reduce marine primary productivity. Nature Climate Change 2012, 2, 519-523. [CrossRef]
- Gao, K.; Campbell, D.A. Photophysiological responses of marine diatoms to elevated CO2 and decreased pH: a review. Functional Plant Biology 2014, 41. [CrossRef]
- Anil, A.C.; Bach, L.T.; Taucher, J.; Boxhammer, T.; Ludwig, A.; Achterberg, E.P.; Algueró-Muñiz, M.; Anderson, L.G.; Bellworthy, J.; Büdenbender, J.; et al. Influence of Ocean Acidification on a Natural Winter-to-Summer Plankton Succession: First Insights from a Long-Term Mesocosm Study Draw Attention to Periods of Low Nutrient Concentrations. Plos One 2016, 11. [CrossRef]
- Dam, H.G.; Bach, L.T.; Alvarez-Fernandez, S.; Hornick, T.; Stuhr, A.; Riebesell, U. Simulated ocean acidification reveals winners and losers in coastal phytoplankton. Plos One 2017, 12. [CrossRef]
- Currie, A.R.; Tait, K.; Parry, H.; de Francisco-Mora, B.; Hicks, N.; Osborn, A.M.; Widdicombe, S.; Stahl, H. Marine Microbial Gene Abundance and Community Composition in Response to Ocean Acidification and Elevated Temperature in Two Contrasting Coastal Marine Sediments. Frontiers in Microbiology 2017, 8. [CrossRef]
- Nawaz, M.Z.; Subin Sasidharan, R.; Alghamdi, H.A.; Dang, H. Understanding Interaction Patterns within Deep-Sea Microbial Communities and Their Potential Applications. Marine Drugs 2022, 20. [CrossRef]
- Schulz, K.G.; Bellerby, R.G.J.; Brussaard, C.P.D.; Büdenbender, J.; Czerny, J.; Engel, A.; Fischer, M.; Koch-Klavsen, S.; Krug, S.A.; Lischka, S.; et al. Temporal biomass dynamics of an Arctic plankton bloom in response to increasing levels of atmospheric carbon dioxide. Biogeosciences 2013, 10, 161-180. [CrossRef]
- Ahme, A.; Von Jackowski, A.; McPherson, R.A.; Wolf, K.K.E.; Hoppmann, M.; Neuhaus, S.; John, U. Winners and Losers of Atlantification: The Degree of Ocean Warming Affects the Structure of Arctic Microbial Communities. Genes 2023, 14. [CrossRef]
- Jenrich, M.; Angelopoulos, M.; Liebner, S.; Treat, C.; Knoblauch, C.; Yang, S.; Grosse, G.; Giebeler, F.; Jongejans, Loeka L.; Grigoriev, M.; et al. Greenhouse Gas Production and Microbial Response During the Transition From Terrestrial Permafrost to a Marine Environment. Permafrost and Periglacial Processes 2024, 36, 63-82. [CrossRef]
- Rees, A.P.; Bange, H.W.; Arévalo-Martínez, D.L.; Artioli, Y.; Ashby, D.M.; Brown, I.; Campen, H.I.; Clark, D.R.; Kitidis, V.; Lessin, G.; et al. Nitrous oxide and methane in a changing Arctic Ocean. Ambio 2021, 51, 398-410. [CrossRef]
- Campen, H.I.; Arévalo-Martínez, D.L.; Artioli, Y.; Brown, I.J.; Kitidis, V.; Lessin, G.; Rees, A.P.; Bange, H.W. The role of a changing Arctic Ocean and climate for the biogeochemical cycling of dimethyl sulphide and carbon monoxide. Ambio 2021, 51, 411-422. [CrossRef]
- Trull, T.W.; Passmore, A.; Davies, D.M.; Smit, T.; Berry, K.; Tilbrook, B. Distribution of planktonic biogenic carbonate organisms in the Southern Ocean south of Australia: a baseline for ocean acidification impact assessment. Biogeosciences 2018, 15, 31-49. [CrossRef]
- Gao, K.; Helbling, E.W.; Häder, D.P.; Hutchins, D.A. Responses of marine primary producers to interactions between ocean acidification, solar radiation, and warming. Marine Ecology Progress Series 2012, 470, 167-189. [CrossRef]
- Hutchins, D.A.; Fu, F.-X.; Yang, S.-C.; John, S.G.; Romaniello, S.J.; Andrews, M.G.; Walworth, N.G. Responses of globally important phytoplankton species to olivine dissolution products and implications for carbon dioxide removal via ocean alkalinity enhancement. Biogeosciences 2023, 20, 4669-4682. [CrossRef]
- Rost, B.; Zondervan, I.; Wolf-Gladrow, D. Sensitivity of phytoplankton to future changes in ocean carbonate chemistry: current knowledge, contradictions and research directions. Marine Ecology Progress Series 2008, 373, 227-237. [CrossRef]
- Raven, J.A.; Crawfurd, K. Environmental controls on coccolithophore calcification. Marine Ecology Progress Series 2012, 470, 137-166. [CrossRef]
- Aberle, N.; Schulz, K.G.; Stuhr, A.; Malzahn, A.M.; Ludwig, A.; Riebesell, U. High tolerance of microzooplankton to ocean acidification in an Arctic coastal plankton community. Biogeosciences 2013, 10, 1471-1481. [CrossRef]
- Bach, L.T.; Riebesell, U.; Schulz, K.G. Distinguishing between the effects of ocean acidification and ocean carbonation in the coccolithophore Emiliania huxleyi. Limnology and Oceanography 2011, 56, 2040-2050. [CrossRef]
- Carter, B.R.; Sharp, J.D.; Dickson, A.G.; Álvarez, M.; Fong, M.B.; García-Ibáñez, M.I.; Woosley, R.J.; Takeshita, Y.; Barbero, L.; Byrne, R.H.; et al. Uncertainty sources for measurable ocean carbonate chemistry variables. Limnology and Oceanography 2023, 69, 1-21. [CrossRef]
- Dupont, S.; Rokitta, S.D.; John, U.; Rost, B. Ocean Acidification Affects Redox-Balance and Ion-Homeostasis in the Life-Cycle Stages of Emiliania huxleyi. PLoS ONE 2012, 7. [CrossRef]
- Goldenberg, S.U.; Nagelkerken, I.; Ferreira, C.M.; Ullah, H.; Connell, S.D. Boosted food web productivity through ocean acidification collapses under warming. Global Change Biology 2017, 23, 4177-4184. [CrossRef]
- Albright, R.; Caldeira, L.; Hosfelt, J.; Kwiatkowski, L.; Maclaren, J.K.; Mason, B.M.; Nebuchina, Y.; Ninokawa, A.; Pongratz, J.; Ricke, K.L.; et al. Reversal of ocean acidification enhances net coral reef calcification. Nature 2016, 531, 362-365. [CrossRef]
- Raven, J.A.; Giordano, M.; Beardall, J.; Maberly, S.C. Algal and aquatic plant carbon concentrating mechanisms in relation to environmental change. Photosynthesis Research 2011, 109, 281-296. [CrossRef]
- Chin, W.-C.; Kübler, J.E.; Dudgeon, S.R. Predicting Effects of Ocean Acidification and Warming on Algae Lacking Carbon Concentrating Mechanisms. Plos One 2015, 10. [CrossRef]
- Zhang, Y.; Beardall, J.; Gao, K. Combined effects of ocean acidification and warming on phytoplankton productivity and community structure in the coastal water of Southern East. Marine Environmental Research 2025, 210, 107352. [CrossRef]
- Simonetti, S.; Zupo, V.; Gambi, M.C.; Luckenbach, T.; Corsi, I. Unraveling cellular and molecular mechanisms of acid stress tolerance and resistance in marine species: New frontiers in the study of adaptation to ocean acidification. Marine Pollution Bulletin 2022, 185. [CrossRef]
- Liang, D.; Xiang, H.; Jin, P.; Xia, J. Response mechanism of harmful algae Phaeocystis globosa to ocean warming and acidification. Environmental Pollution 2023, 320. [CrossRef]
- Zhang, X.; Xi, T.; Wang, Y.; Fan, X.; Xu, D.; Zhang, P.; Sun, K.; Zhang, Y.; Ma, J.; Ye, N. Chemical interactions between kelp Macrocystis pyrifera and symbiotic bacteria under elevated CO2 condition. Marine Life Science & Technology 2024, 6, 700-712. [CrossRef]
- Iida, Y.; Takatani, Y.; Kojima, A.; Ishii, M. Global trends of ocean CO2 sink and ocean acidification: an observation-based reconstruction of surface ocean inorganic carbon variables. Journal of Oceanography 2020, 77, 323-358. [CrossRef]
- Hassenrück, C.; Fink, A.; Lichtschlag, A.; Tegetmeyer, H.E.; de Beer, D.; Ramette, A.; King, G. Quantification of the effects of ocean acidification on sediment microbial communities in the environment: the importance of ecosystem approaches. FEMS Microbiology Ecology 2016, 92. [CrossRef]
- Newbold, L.K.; Oliver, A.E.; Booth, T.; Tiwari, B.; DeSantis, T.; Maguire, M.; Andersen, G.; van der Gast, C.J.; Whiteley, A.S. The response of marine picoplankton to ocean acidification. Environmental Microbiology 2012, 14, 2293-2307. [CrossRef]
- Deppeler, S.; Petrou, K.; Schulz, K.G.; Westwood, K.; Pearce, I.; McKinlay, J.; Davidson, A. Ocean acidification of a coastal Antarctic marine microbial community reveals a critical threshold for CO2 tolerance in phytoplankton productivity. Biogeosciences 2018, 15, 209-231. [CrossRef]
- James, A.K.; Kelly, L.W.; Nelson, C.E.; Wilbanks, E.G.; Carlson, C.A. Elevated pCO2 alters marine heterotrophic bacterial community composition and metabolic potential in response to a pulse of phytoplankton organic matter. Environmental Microbiology 2019, 21, 541-556. [CrossRef]
- Spilling, K.; Schulz, K.G.; Paul, A.J.; Boxhammer, T.; Achterberg, E.P.; Hornick, T.; Lischka, S.; Stuhr, A.; Bermúdez, R.; Czerny, J.; et al. Effects of ocean acidification on pelagic carbon fluxes in a mesocosm experiment. Biogeosciences 2016, 13, 6081-6093. [CrossRef]
- Spisla, C.; Taucher, J.; Bach, L.T.; Haunost, M.; Boxhammer, T.; King, A.L.; Jenkins, B.D.; Wallace, J.R.; Ludwig, A.; Meyer, J.; et al. Extreme Levels of Ocean Acidification Restructure the Plankton Community and Biogeochemistry of a Temperate Coastal Ecosystem: A Mesocosm Study. Frontiers in Marine Science 2021, 7. [CrossRef]
- Taucher, J.; Boxhammer, T.; Bach, L.T.; Paul, A.J.; Schartau, M.; Stange, P.; Riebesell, U. Changing carbon-to-nitrogen ratios of organic-matter export under ocean acidification. Nature Climate Change 2020, 11, 52-57. [CrossRef]
- Stockdale, A.; Tipping, E.; Lofts, S.; Mortimer, R.J.G. Effect of Ocean Acidification on Organic and Inorganic Speciation of Trace Metals. Environmental Science & Technology 2016, 50, 1906-1913. [CrossRef]
- Cohen, A.L.; McCorkle, D.C.; de Putron, S.; Gaetani, G.A.; Rose, K.A. Morphological and compositional changes in the skeletons of new coral recruits reared in acidified seawater: Insights into the biomineralization response to ocean acidification. Geochemistry, Geophysics, Geosystems 2009, 10. [CrossRef]
- Simone, M.N.; Schulz, K.G.; Oakes, J.M.; Eyre, B.D. Warming and ocean acidification may decrease estuarine dissolved organic carbon export to the ocean. Biogeosciences 2021, 18, 1823-1838. [CrossRef]
- Heinze, C.; Meyer, S.; Goris, N.; Anderson, L.; Steinfeldt, R.; Chang, N.; Le Quéré, C.; Bakker, D.C.E. The ocean carbon sink – impacts, vulnerabilities and challenges. Earth System Dynamics 2015, 6, 327-358. [CrossRef]
- Dore, J.E.; Lukas, R.; Sadler, D.W.; Church, M.J.; Karl, D.M. Physical and biogeochemical modulation of ocean acidification in the central North Pacific. Proceedings of the National Academy of Sciences 2009, 106, 12235-12240. [CrossRef]
- Doney, S.C.; Balch, W.M.; Fabry, V.J.; Feely, R.A. OCEAN ACIDIFICATIONA CRITICAL EMERGING PROBLEM FOR THE OCEAN SCIENCES. Oceanography 2009, 22, 16-25.
- Lohbeck, K.T.; Riebesell, U.; Reusch, T.B.H. Adaptive evolution of a key phytoplankton species to ocean acidification. Nature Geoscience 2012, 5, 346-351. [CrossRef]
- Cornwall, C.E.; Hurd, C.L. Experimental design in ocean acidification research: problems and solutions. ICES Journal of Marine Science 2016, 73, 572-581. [CrossRef]
- Zark, M.; Riebesell, U.; Dittmar, T. Effects of ocean acidification on marine dissolved organic matter are not detectable over the succession of phytoplankton blooms. Science Advances 2015, 1. [CrossRef]
- Ianora, A.; Li, Y.; Zhuang, S.; Wu, Y.; Ren, H.; Chen, F.; Lin, X.; Wang, K.; Beardall, J.; Gao, K. Ocean acidification modulates expression of genes and physiological performance of a marine diatom. Plos One 2017, 12. [CrossRef]
- McNicholl, C.; Koch, M.S.; Hofmann, L.C. Photosynthesis and light-dependent proton pumps increase boundary layer pH in tropical macroalgae: A proposed mechanism to sustain calcification under ocean acidification. Journal of Experimental Marine Biology and Ecology 2019, 521. [CrossRef]
- Chakraborty, S.; Paidi, M.K.; Dhinakarasamy, I.; Sivakumar, M.; Clements, C.; Thirumurugan, N.K.; Sivakumar, L. Adaptive mechanism of the marine bacterium Pseudomonas sihuiensis-BFB-6S towards pCO2 variation: Insights into synthesis of extracellular polymeric substances and physiochemical modulation. International Journal of Biological Macromolecules 2024, 261, 129860. [CrossRef]
- Barreto, M.M.; Ziegler, M.; Venn, A.; Tambutté, E.; Zoccola, D.; Tambutté, S.; Allemand, D.; Antony, C.P.; Voolstra, C.R.; Aranda, M. Effects of Ocean Acidification on Resident and Active Microbial Communities of Stylophora pistillata. Frontiers in Microbiology 2021, 12. [CrossRef]
- Barrett, R.C.; Carter, B.R.; Fassbender, A.J.; Tilbrook, B.; Woosley, R.J.; Azetsu-Scott, K.; Feely, R.A.; Goyet, C.; Ishii, M.; Murata, A.; et al. Biological Responses to Ocean Acidification Are Changing the Global Ocean Carbon Cycle. Global Biogeochemical Cycles 2025, 39. [CrossRef]
- Kroeker, K.J.; Kordas, R.L.; Harley, C.D.G. Embracing interactions in ocean acidification research: confronting multiple stressor scenarios and context dependence. Biology Letters 2017, 13. [CrossRef]
- Rastelli, E.; Petani, B.; Corinaldesi, C.; Dell’Anno, A.; Lo Martire, M.; Cerrano, C.; Danovaro, R. A high biodiversity mitigates the impact of ocean acidification on hard-bottom ecosystems. Scientific Reports 2020, 10, 2948. [CrossRef]
- Ye, M.; Zhang, J.; Xiao, M.; Huang, J.; Zhou, Y.; Beardall, J.; Raven, J.A.; Gao, G.; Liang, X.; Wu, F.; et al. Multi-omics analyses reveal the signatures of metabolite transfers across trophic levels in a high-CO2 ocean. Limnology and Oceanography 2024, 69, 1667-1682. [CrossRef]
- Wang, Z.; Tsementzi, D.; Williams, T.C.; Juarez, D.L.; Blinebry, S.K.; Garcia, N.S.; Sienkiewicz, B.K.; Konstantinidis, K.T.; Johnson, Z.I.; Hunt, D.E. Environmental stability impacts the differential sensitivity of marine microbiomes to increases in temperature and acidity. The ISME Journal 2021, 15, 19-28. [CrossRef]
- Imdahl, F.; Saliba, A.-E. Advances and challenges in single-cell RNA-seq of microbial communities. Current Opinion in Microbiology 2020, 57, 102-110. [CrossRef]
- Raajaraam, L.; Raman, K. Modeling Microbial Communities: Perspective and Challenges. ACS Synthetic Biology 2024, 13, 2260-2270. [CrossRef]
- Dutta, A.; Goldman, T.; Keating, J.; Burke, E.; Williamson, N.; Dirmeier, R.; Bowman, J.S.; Denef, V.J. Machine Learning Predicts Biogeochemistry from Microbial Community Structure in a Complex Model System. Microbiology Spectrum 2022, 10. [CrossRef]
- Connors, E.; Dutta, A.; Trinh, R.; Erazo, N.; Dasarathy, S.; Ducklow, H.; Weissman, J.L.; Yeh, Y.-C.; Schofield, O.; Steinberg, D.; et al. Microbial community composition predicts bacterial production across ocean ecosystems. The ISME Journal 2024, 18. [CrossRef]
- Benway, H.M.; Lorenzoni, L.; White, A.E.; Fiedler, B.; Levine, N.M.; Nicholson, D.P.; DeGrandpre, M.D.; Sosik, H.M.; Church, M.J.; O’Brien, T.D.; et al. Ocean Time Series Observations of Changing Marine Ecosystems: An Era of Integration, Synthesis, and Societal Applications. Frontiers in Marine Science 2019, 6. [CrossRef]
- Godbold, J.A.; Solan, M. Long-term effects of warming and ocean acidification are modified by seasonal variation in species responses and environmental conditions. Philosophical Transactions of the Royal Society B: Biological Sciences 2013, 368. [CrossRef]
- Brown, M.V.; van de Kamp, J.; Ostrowski, M.; Seymour, J.R.; Ingleton, T.; Messer, L.F.; Jeffries, T.; Siboni, N.; Laverock, B.; Bibiloni-Isaksson, J.; et al. Systematic, continental scale temporal monitoring of marine pelagic microbiota by the Australian Marine Microbial Biodiversity Initiative. Scientific Data 2018, 5. [CrossRef]
- Sutton, A.J.; Battisti, R.; Carter, B.; Evans, W.; Newton, J.; Alin, S.; Bates, N.R.; Cai, W.-J.; Currie, K.; Feely, R.A.; et al. Advancing best practices for assessing trends of ocean acidification time series. Frontiers in Marine Science 2022, 9. [CrossRef]
- Leray, M.; Wilkins, L.G.E.; Apprill, A.; Bik, H.M.; Clever, F.; Connolly, S.R.; De León, M.E.; Duffy, J.E.; Ezzat, L.; Gignoux-Wolfsohn, S.; et al. Natural experiments and long-term monitoring are critical to understand and predict marine host–microbe ecology and evolution. PLOS Biology 2021, 19. [CrossRef]
- Krishna, S.; Lemmen, C.; Örey, S.; Rehren, J.; Pane, J.D.; Mathis, M.; Püts, M.; Hokamp, S.; Pradhan, H.K.; Hasenbein, M.; et al. Interactive effects of multiple stressors in coastal ecosystems. Frontiers in Marine Science 2025, 11. [CrossRef]
- Hu, N.; Bourdeau, P.E.; Hollander, J. Responses of marine trophic levels to the combined effects of ocean acidification and warming. Nature Communications 2024, 15. [CrossRef]
- Subramaniam, R.C.; Melbourne-Thomas, J.; Davidson, A.T.; Corney, S.P. Mechanisms driving Antarctic microbial community responses to ocean acidification: a network modelling approach. Polar Biology 2016, 40, 727-734. [CrossRef]
- Hakonen, A.; Anderson, L.G.; Engelbrektsson, J.; Hulth, S.; Karlson, B. A potential tool for high-resolution monitoring of ocean acidification. Analytica Chimica Acta 2013, 786, 1-7. [CrossRef]
- Sastri, A.R.; Christian, J.R.; Achterberg, E.P.; Atamanchuk, D.; Buck, J.J.H.; Bresnahan, P.; Duke, P.J.; Evans, W.; Gonski, S.F.; Johnson, B.; et al. Perspectives on in situ Sensors for Ocean Acidification Research. Frontiers in Marine Science 2019, 6. [CrossRef]
- Treusch, A.H.; Vergin, K.L.; Finlay, L.A.; Donatz, M.G.; Burton, R.M.; Carlson, C.A.; Giovannoni, S.J. Seasonality and vertical structure of microbial communities in an ocean gyre. The ISME Journal 2009, 3, 1148-1163. [CrossRef]
- Gao, K.; Ruan, Z.; Villafañe, V.E.; Gattuso, J.-P.; Helbling, E.W. Ocean acidification exacerbates the effect of UV radiation on the calcifying phytoplankter Emiliania huxleyi. Limnology and Oceanography 2009, 54, 1855-1862. [CrossRef]
- Zhang, C.L.; Xie, W.; Martin-Cuadrado, A.-B.; Rodriguez-Valera, F. Marine Group II Archaea, potentially important players in the global ocean carbon cycle. Frontiers in Microbiology 2015, 6. [CrossRef]
- Wannicke, N.; Frey, C.; Law, C.S.; Voss, M. The response of the marine nitrogen cycle to ocean acidification. Global Change Biology 2018, 24, 5031-5043. [CrossRef]
- Malits, A.; Boras, J.A.; Balagué, V.; Calvo, E.; Gasol, J.M.; Marrasé, C.; Pelejero, C.; Pinhassi, J.; Sala, M.M.; Vaqué, D. Viral-Mediated Microbe Mortality Modulated by Ocean Acidification and Eutrophication: Consequences for the Carbon Fluxes Through the Microbial Food Web. Frontiers in Microbiology 2021, 12. [CrossRef]
- Whitehead, A. Comparative genomics in ecological physiology: toward a more nuanced understanding of acclimation and adaptation. Journal of Experimental Biology 2012, 215, 884-891. [CrossRef]
- Vance, T.C.; Wengren, M.; Burger, E.; Hernandez, D.; Kearns, T.; Medina-Lopez, E.; Merati, N.; O’Brien, K.; O’Neil, J.; Potemra, J.T.; et al. From the Oceans to the Cloud: Opportunities and Challenges for Data, Models, Computation and Workflows. Frontiers in Marine Science 2019, 6. [CrossRef]
- Rubbens, P.; Brodie, S.; Cordier, T.; Destro Barcellos, D.; Devos, P.; Fernandes-Salvador, J.A.; Fincham, J.I.; Gomes, A.; Handegard, N.O.; Howell, K.; et al. Machine learning in marine ecology: an overview of techniques and applications. ICES Journal of Marine Science 2023, 80, 1829-1853. [CrossRef]
- Cui, S.; Gao, Y.; Huang, Y.; Shen, L.; Zhao, Q.; Pan, Y.; Zhuang, S. Advances and applications of machine learning and deep learning in environmental ecology and health. Environmental Pollution 2023, 335. [CrossRef]
- Canonico, G.; Buttigieg, P.L.; Montes, E.; Muller-Karger, F.E.; Stepien, C.; Wright, D.; Benson, A.; Helmuth, B.; Costello, M.; Sousa-Pinto, I.; et al. Global Observational Needs and Resources for Marine Biodiversity. Frontiers in Marine Science 2019, 6. [CrossRef]
- Thompson, C.L.; Alberti, M.; Barve, S.; Battistuzzi, F.U.; Drake, J.L.; Goncalves, G.C.; Govaert, L.; Partridge, C.; Yang, Y. Back to the Future: Reintegrating Biology to Understand How Past Eco-evolutionary Change Can Predict Future Outcomes. Integrative and Comparative Biology 2021, 61, 2218-2232. [CrossRef]
- Cooke, S.J.; Killen, S.S.; Metcalfe, J.D.; McKenzie, D.J.; Mouillot, D.; Jorgensen, C.; Peck, M.A. Conservation physiology across scales: insights from the marine realm. Conservation Physiology 2014, 2, cou024-cou024. [CrossRef]
- Roberts, J.M.; Devey, C.W.; Biastoch, A.; Carreiro-Silva, M.; Dohna, T.; Dorschel, B.; Gunn, V.; Huvenne, V.A.I.; Johnson, D.; Jollivet, D.; et al. A blueprint for integrating scientific approaches and international communities to assess basin-wide ocean ecosystem status. Communications Earth & Environment 2023, 4. [CrossRef]
- Wahl, M.; Saderne, V.; Sawall, Y. How good are we at assessing the impact of ocean acidification in coastal systems? Limitations, omissions and strengths of commonly used experimental approaches with special emphasis on the neglected role of fluctuations. Marine and Freshwater Research 2016, 67. [CrossRef]
- MÜller, M.N.; Antia, A.N.; LaRoche, J. Influence of cell cycle phase on calcification in the coccolithophore Emiliania huxleyi. Limnology and Oceanography 2008, 53, 506-512. [CrossRef]
- Merrie, A.; Dunn, D.C.; Metian, M.; Boustany, A.M.; Takei, Y.; Elferink, A.O.; Ota, Y.; Christensen, V.; Halpin, P.N.; Österblom, H. An ocean of surprises – Trends in human use, unexpected dynamics and governance challenges in areas beyond national jurisdiction. Global Environmental Change 2014, 27, 19-31. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



