
Submitted:
27 September 2025
Posted:
28 September 2025
You are already at the latest version
Abstract
The anatomy of the vegetative organs of Vanilla pompona subsp. grandiflora, an endemic species of the humid forests of northeastern Peru and of great ecological and economic importance, is described through histological sections and histochemical tests. The leaves showed homogeneous mesophyll and epidermis with slightly depressed anomocytic and tetracytic stomata. The stems showed a continuous ring of perivascular sclerenchyma and vascular bundles with sclerenchyma reinforcement, while the roots showed uniseriate velamen, thickened exodermis, parenchymatic cortex and a vascular cylinder with large-diameter xylem elements. Histochemical assays revealed the presence of quinones in the leaf mesophyll, lipids in the shoot sclerenchyma, lignin in the xylem, and starches in the root. These traits provide diagnostic characteristics for subspecies recognition and constitute a basis for a comprehensive morphological characterization, complemented by preliminary histochemical observations, which will provide input for future anatomical and taxonomic studies of Amazonian floodplain plants.
Keywords:
histochemistry
; histology
; plant anatomy
; vegetative organs
1. Introduction
The Orchidaceae family has a pantropical distribution, also reaching temperate, boreal, and subantarctic zones, although its greatest diversity is concentrated in the tropics [1]. It is one of the two largest families of angiosperms, along with Asteraceae, and its modern classification recognizes five subfamilies: Apostasioideae, Cypripedioideae, Vanilloideae, Orchidoideae, and Epidendroideae, organized into 21 tribes [2]. With nearly 28,000 species worldwide, Peru has 212 genera and around 2,206 species, with an estimated 2,500 to 3,500 species [3,4].
The subfamily Vanilloideae comprises 15 genera and 180 species [5], predominantly found in South America [6]. The most representative genus is Vanilla Mill., which is economically important and consists of hemiepiphytic lianas with fleshy stems, adventitious roots, and showy flowers, distributed in pantropical regions [7,8]. Mexico is the center of origin and domestication of Vanilla planifolia [9], the world's best-known vanilla species, whose mature pods are highly prized and in high demand on the international market [10] in both the perfume and food and gastronomy industries [11]. Among its South American representatives, Vanilla pompona subsp. grandiflora (Lindl.) Soto Arenas, endemic to the permanently flooded forests of northeastern Peru, where it is part of aguajal ecosystems and aguajal-renacal transition territories [8,12].
In Peru, documentation of the genus Vanilla has been limited. The earliest records date back to the manuscripts of Ruiz & Pavón in the 18th century, followed by Humboldt's observations [13,14] in Cajamarca. In the 20th century, Kraenzlin [15] described V. weberbaueriana based on specimens from Junín, while Schlechter [16] and Schweinfurth [17] expanded the lists for the country. Recent research has incorporated new records and species, with 17 taxa currently recognized in Peruvian territory, several of them with restricted distribution [18,19,20,21]. In this regard, recent studies on Vanilla in the San Martín region record attempts at vegetative propagation with contrasting results. It has been reported that the type of substrate significantly influences the number and length of V. pompona roots, as well as sprouting and shoot diameter, with leaf litter being the most favorable substrate, thus providing patterns that suggest that vegetative behavior depends on the interaction between substrate and time, and that limitations in propagation may be related to anatomical characteristics not yet described [22].
Comparative anatomy provides useful diagnostic characters for differentiating morphologically similar species [23]. Furthermore, in orchids, anatomy reflects adaptations to humid and epiphytic environments, in which structures such as storage tissues, vascular system organization, and epidermal morphology are associated with water tolerance, mechanical support, and photosynthetic efficiency [24,25].
Anatomical studies on the genus Vanilla have been scarce and mainly focus on cultivated species such as V. planifolia [25], while wild species remain largely unexplored, despite their ecological and cultural relevance to local communities [26]. Thus, the lack of anatomical information represents an obstacle both to understanding the biology of the species and to designing cultivation and conservation strategies [27,28]. In this context, the histological study of V. pompona subsp. grandiflora is particularly relevant, considering its restricted distribution, the pressure from deforestation, and the growing interest in its productive value and in situ conservation [28].
This study describes the anatomy of the vegetative organs of V. pompona subsp. grandiflora in wild populations of the Alto Mayo Valley (VAM), with the aim of providing diagnostic characteristics to differentiate it from other related species and providing information on its degree of adaptation to flood-prone environments.
2. Materials and Methods
2.1. Study Area and Plant Material
Five adult wild individuals of Vanilla pompona subsp. grandiflora (Lindl.) Soto Arenas were selected from the province and district of Moyobamba, San Martín department, Peru (05°54′17.9″ S, 77°07′07.5″ W).
Samples were collected, including aerial roots (approximately 60 mm), stems at the internode level (approximately 140 mm), and leaves (approximately 170 mm). These were temporarily stored in 70% ethanol at 15 °C until processing. Voucher specimens were deposited in the Herbarium San Marcos (USM), Universidad Nacional Mayor de San Marcos, under collection numbers V040 (USM 347166) and V151 (USM 347167). Collections were conducted under authorization RD Nº D000022-2024-MIDAGRI-SERFOR-DGGSPFFS-DGSPF.
2.2. Anatomical Study
Fresh material was hand-sectioned to obtain transverse cuts of stems and roots, and transverse and surface cuts of leaves, mounted in water for preliminary observation. In fixed material, transverse and surface sections were cleared with sodium hypochlorite (50%), bleached with chloral hydrate (5%), and stained with toluidine blue (1%). Permanent and semi-permanent slides were prepared in synthetic resin (temporal) or glycerin gelatin.
Histochemical tests were applied on hand sections: sodium hydroxide 2.5% for diaphanization, ethanol boiling at 96%, clearing with sodium hypochlorite (50%), bleaching with chloral hydrate, and staining with toluidine blue, following venation pattern evaluation according to Hickey [40] and Ash et al. [41]. For ultrastructural analysis and high-quality microphotographs, modified paraffin embedding was used, samples were dehydrated in ascending ethanol concentrations (70%, 80%, 96%, 100% I–III), passed through ethanol-xylene (I–III) and xylene (I–II), and infiltrated in paraffin (I–III). Tissues were embedded in paraffin blocks on a warm plate and sectioned with a Leica SM 2010R rotary microtome. Sections were mounted on slides coated with Mayer’s albumin, deparaffinized, and rehydrated through a descending ethanol series (100%–50%). Staining was carried out with toluidine blue. Finally, samples were dehydrated, cleared in xylene (III–IV), and mounted in Entellan, then air-dried before microscopic observation. [42].
For metabolite recognition in fresh sections, histochemical reagents were applied: phloroglucinol in acidic medium (lignin), Sudan III (suberin), Lugol’s solution (starch), ferric chloride 1% (tannins), Dragendorff’s reagent (alkaloids), and sodium hydroxide 5% (quinones), following the methods modified by Locke [43] and D’Ambrogio de Argüeso [42].
Microphotographs were taken with a Leica ICC50 microscope and LAS EZ software; additional measurements were made with ImageJ [44]. Panoramic images (40× and 100×) and detailed images (400×) were captured in three fields per sample. Microscopic measurements were made in triplicate per individual: leaf anatomy, surface view (3 fields, 10 measurements each) and transverse view (3 cuts, 10 measurements each); stem anatomy, transverse view (3 cuts, 10 measurements each); root anatomy, transverse view (3 cuts, 10 measurements each).
The qualitative and quantitative parameters to be evaluated for the comparative anatomical study of Vanilla pompona subsp. grandiflora (Lindl.) Soto Arenas were specific to each organ. The criteria followed the method modified from Evans [45]and Ash et al. [41]:

Table 1.
Qualitative and quantitative anatomical parameters of Vanilla pompona subsp. grandiflora evaluated in this study.
Table 1.
Qualitative and quantitative anatomical parameters of Vanilla pompona subsp. grandiflora evaluated in this study.
| Plant organ | Parameters | |
| Qualitative | Quantitative | |
| Leaf | Type of mesophyll, presence or absence of trichomes or glands, presence or absence of crystals, stomatal types, stomatal position and vascular bundle distribution. | Stomatal index and density, mesophyll thickness, cuticle thickness. |
| Stem | Type of epidermis, type of cuticle, presence or absence of trichomes, tissue organization, vascular bundle organization, and fiber classification. | Cuticle thickness, epidermal thickness, vessel diameter. |
| Root | Type of xylem, presence of exodermis and endodermis, sclerenchyma fiber bundles and vascular bundle organization. | Root diameter, periderm thickness (if present), xylem vessel diameter. |
3. Results
3.1. Leaf Anatomy
The leaf of Vanilla pompona subsp. grandiflora is thick and broad. The adaxial epidermis (Figure 1A, 1E) consists of periclinal cells with a thickened outer wall, whereas the abaxial epidermis (Figure 1A, 1F) is composed of isodiametric or periclinal cells. Both leaf surfaces (Figure 1A, 1B) are covered by a cuticle with an average thickness of 4.50 ± 0.37 µm (Table 2). Rectangular crystals were observed scattered throughout the mesophyll, as evidenced in micrographs at 1000× magnification (Figure 1C). The mesophyll (Figure 1A) is homogeneous, consisting of compact, thin-walled cells, with an average thickness of 1144.88 ± 54.17 µm (Table 2). The vascular bundles (Figure 1C, 1D) are accompanied on the phloem side by sclerenchyma bands that reinforce mechanical support. The leaf is hypostomatic and lacks trichomes, and the stomata, restricted to the abaxial epidermis, are superficial, anomocytic and tetracytic (Figure 1F), slightly sunken below the surface, with a density of 4.5 ± 0.85 stomata/mm² and a stomatal index of 7.92 ± 1.32% (Table 2).
3.2. Stem Anatomy
In the stem of Vanilla pompona subsp. grandiflora, a primary structure is present, characterized by a uniseriate epidermis (Figure 2A, B) measuring 34.09 ± 5.92 µm in thickness and a cuticle of 5.26 ± 1.90 µm (Table 2). A continuous ring of perivascular sclerenchyma, one to four cell layers thick, provides mechanical support to the structure (Figure 2A, D). The vascular bundles are well defined and surrounded by sclerenchyma fibers, reinforcing the anatomical identity of the tissue and its adaptation to a climbing, epiphytic habit. The pith parenchyma occupies the central region of the stem in cross-section (Figure 2C), while in the xylem, vessels with an average diameter of 35.52 ± 4.27 µm (Table 2) are observed, along with tracheary elements in longitudinal section that display annular, spiral, scalariform, and reticulate thickenings (Figure 2E, F). Notably, trichomes are absent on the stem surface.
3.3. Root Anatomy
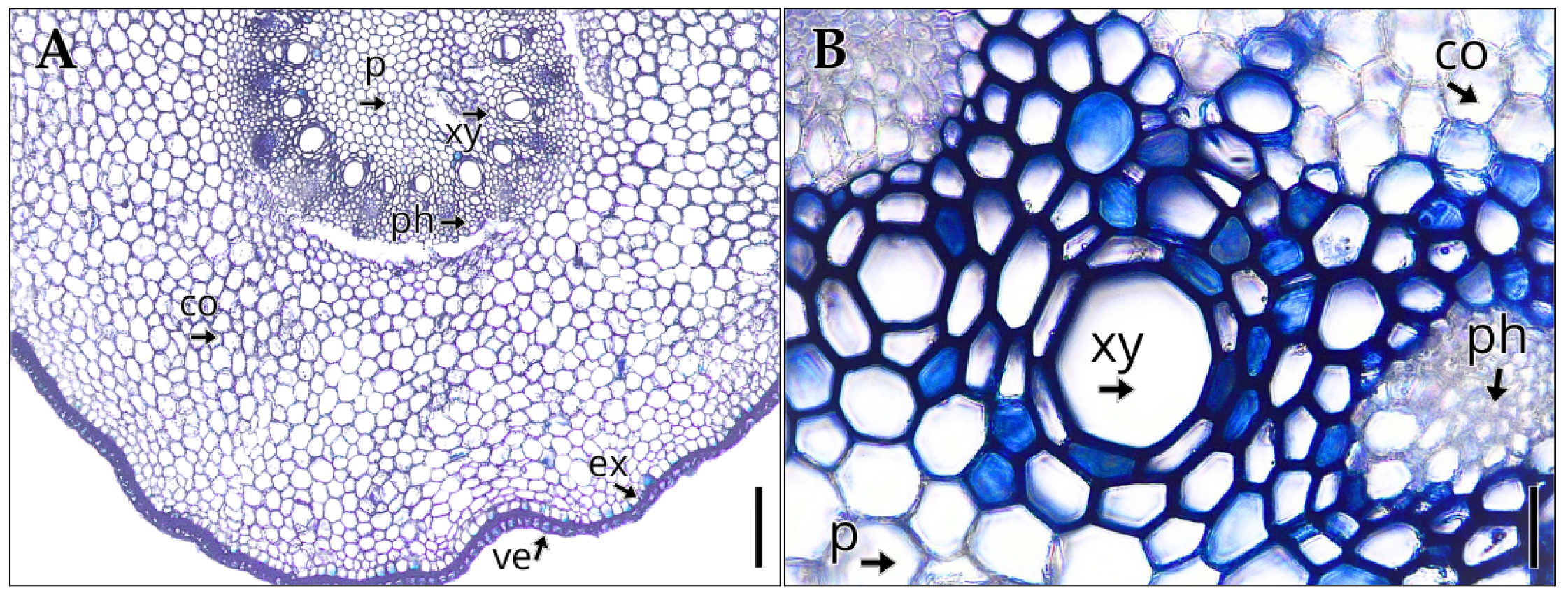
The root of Vanilla pompona subsp. grandiflora, with an average diameter of 1482.55 ± 10.01 µm (Table 2), exhibits in cross-section a primary polyarch structure (Figure 3A), with a uniseriate velamen whose anticlinal cells show thickenings on their inner and lateral walls. Beneath this layer lies the exodermis, followed by a broad cortical parenchyma composed of isodiametric, thin-walled cells, in which root hairs are absent. The vascular cylinder (Figure 3B) displays a radial arrangement, with the phloem organized in narrow strands and the xylem formed by wide-diameter vascular elements. In the central region, an abundant pith parenchyma is present, consisting of rounded, thin-walled cells.
Figure 3.
Internal structure of the leaf of Vanilla pompona subsp. grandiflora. (A) Mesophyll with aquiferous parenchyma. (B) Transverse section showing cuticle and epidermis in detail. (C) Crystals in the mesophyll (D) Detail of vascular bundle. (E) Adaxial epidermis. (F) Abaxial epidermis. Abbreviations: abe, abaxial epidermis; ade, adaxial epidermis; ap, aquiferous parenchyma; bs, bundle sheath; cu, cuticle; lv, leaf vessels; me, mesophyll; vb, vascular bundle; sta, anomocytic stoma; stt, tetracytic stoma. Scale bar: (A) 200 µm; (B) 16 µm; (C) 80 µm; (D–F) 40 µm.
Figure 3.
Internal structure of the leaf of Vanilla pompona subsp. grandiflora. (A) Mesophyll with aquiferous parenchyma. (B) Transverse section showing cuticle and epidermis in detail. (C) Crystals in the mesophyll (D) Detail of vascular bundle. (E) Adaxial epidermis. (F) Abaxial epidermis. Abbreviations: abe, abaxial epidermis; ade, adaxial epidermis; ap, aquiferous parenchyma; bs, bundle sheath; cu, cuticle; lv, leaf vessels; me, mesophyll; vb, vascular bundle; sta, anomocytic stoma; stt, tetracytic stoma. Scale bar: (A) 200 µm; (B) 16 µm; (C) 80 µm; (D–F) 40 µm.
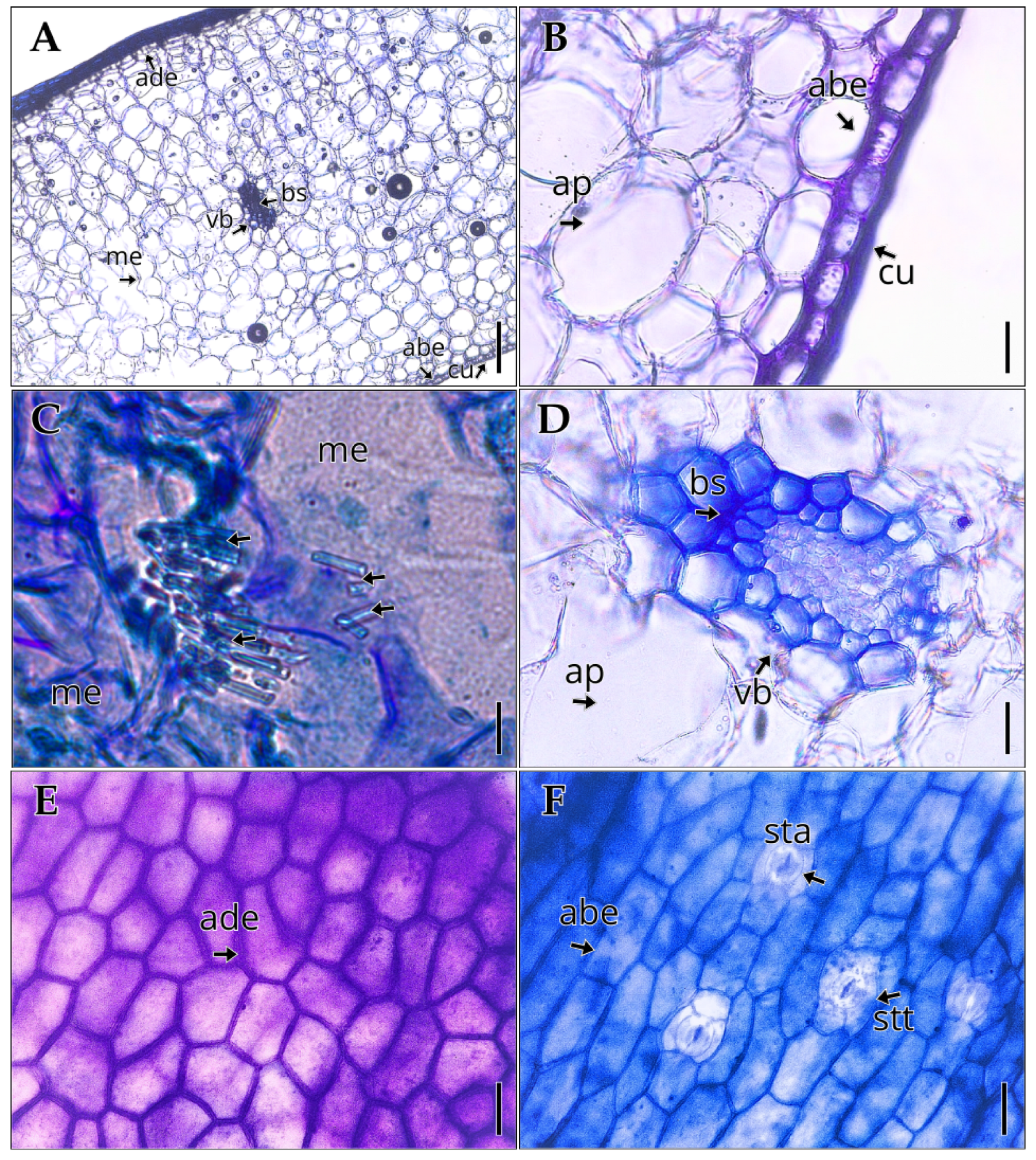
Figure 4.
Internal structure of the stem of Vanilla pompona subsp. grandiflora. (A) Cortex. (B) Epidermis and cuticle. (C) Pith parenchyma. (D) Vascular bundle. (E) Tracheids. (F) Vessels. Abbreviations: sf, sclerenchyma fibers; c, cortex; cu, cuticle; ep, epidermis; pa, parenchyma; ph, phloem; scl, sclerenchyma; vb, vascular bundle; xf, xylary fibers. Scale bar: (A) 200 µm; (B, D) 40 µm; (C, E, F) 80 µm.
Figure 4.
Internal structure of the stem of Vanilla pompona subsp. grandiflora. (A) Cortex. (B) Epidermis and cuticle. (C) Pith parenchyma. (D) Vascular bundle. (E) Tracheids. (F) Vessels. Abbreviations: sf, sclerenchyma fibers; c, cortex; cu, cuticle; ep, epidermis; pa, parenchyma; ph, phloem; scl, sclerenchyma; vb, vascular bundle; xf, xylary fibers. Scale bar: (A) 200 µm; (B, D) 40 µm; (C, E, F) 80 µm.
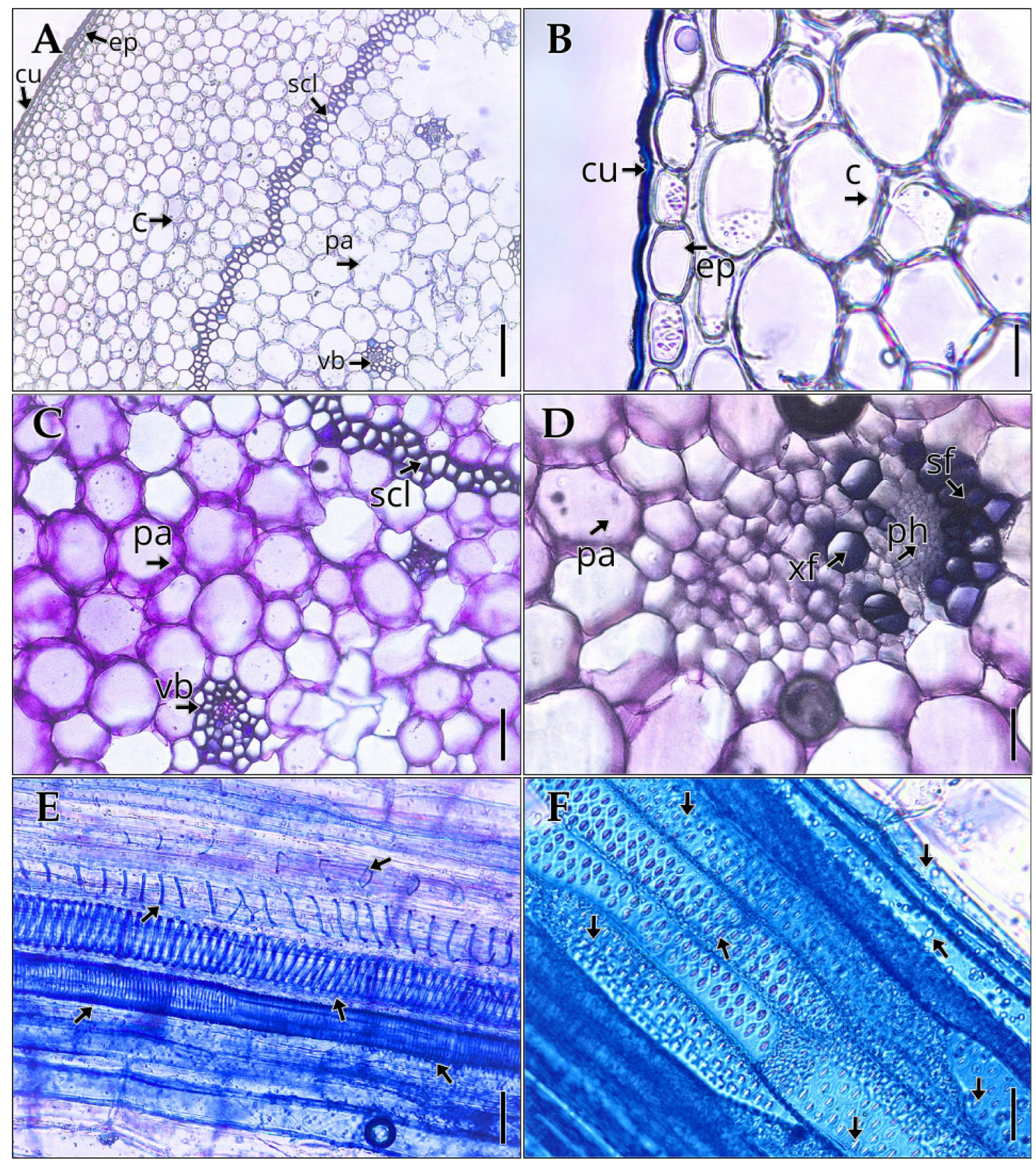
Figure 5.
Internal structure of the root of Vanilla pompona subsp. grandiflora. (A) Transverse root section. (B) Vascular bundle. Abbreviations: co, cortical parenchyma; ex, exodermis; ph, phloem; p, pith parenchyma; xy, xylem; ve, velamen. Scale bar: (A) 200 µm; (B) 80 µm.
Figure 5.
Internal structure of the root of Vanilla pompona subsp. grandiflora. (A) Transverse root section. (B) Vascular bundle. Abbreviations: co, cortical parenchyma; ex, exodermis; ph, phloem; p, pith parenchyma; xy, xylem; ve, velamen. Scale bar: (A) 200 µm; (B) 80 µm.
3.4. Histochemistry
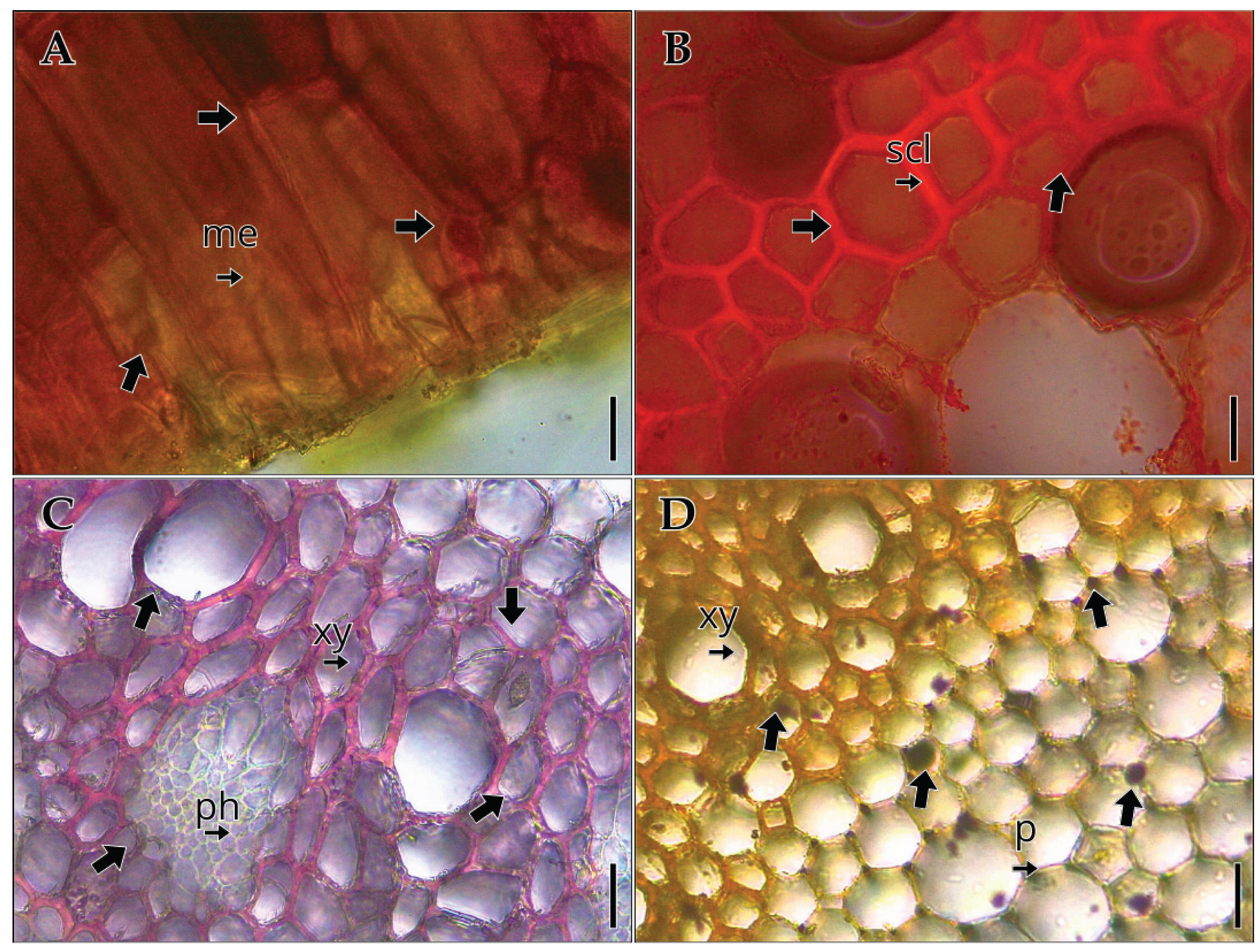
Histochemical tests on the vegetative organs of Vanilla pompona subsp. grandiflora revealed the presence of different metabolite groups. In the leaf (A), a positive reaction to quinones with 5% sodium hydroxide was observed mainly in the mesophyll. In the stem (B), lipid detection using Sudan III showed positive staining in the sclerenchyma bands. In the root (C), the floroglucinol test confirmed lignin deposition in the xylem elements. Finally, in the root (D), Lugol’s iodine test revealed the presence of starch grains in the pith.
Figure 6.
Histochemistry of the vegetative organs of Vanilla pompona subsp. grandiflora. (A) quinones in mesophyll, (B) lipids in stem sclerenchyma, (C) lignin in root xylem, (D) starch in root pith. Abbreviations: me, mesophyll; ph, phloem; p, pith parenchyma; scl, sclerenchyma; xy, xylem. Scale bar: (A–C) 40 µm; (D) 80 µm.
Figure 6.
Histochemistry of the vegetative organs of Vanilla pompona subsp. grandiflora. (A) quinones in mesophyll, (B) lipids in stem sclerenchyma, (C) lignin in root xylem, (D) starch in root pith. Abbreviations: me, mesophyll; ph, phloem; p, pith parenchyma; scl, sclerenchyma; xy, xylem. Scale bar: (A–C) 40 µm; (D) 80 µm.
4. Discussion
The anatomy of the vegetative organs observed in Vanilla pompona subsp. grandiflora is consistent with that documented for the genus, while also providing precise metrics that enrich its ecofunctional interpretation [29].
In the leaf, the mesophyll is homogeneous and compact (1144.88 ± 54.17 µm) and the arrangement of collateral bundles with predominantly phloematic sclerenchymatic reinforcement is consistent with that described for homogeneous laminae with sclerenchymatic inclusions and sheaths in Vanilla [23]. The occurrence of rectangular crystals throughout the mesophyll is consistent with prior reports for the genus [26] and may serve as an additional anatomical feature useful for comparative and taxonomic purposes. In addition to their taxonomic significance, calcium oxalate crystals have been suggested to play diverse physiological roles in plants, such as regulating tissue calcium and ionic balance, providing structural support, protecting against herbivory, and contributing to the detoxification of heavy metals; they may also influence light capture and reflection [30].The epidermis with thickened periclinal cells on the adaxial side and isodiametric cells on the abaxial side is consistent with the epidermal variability of the group, in line with what has been described for V. planifolia, where epidermal morphology and stomatal diversity have shown phylogenetic relevance [24]. Although Raffi et al. [23] report tetracytic predominance with the presence of anisocytic and anomocytic complexes, in this subspecies the tetracytic and anomocytic condition stands out, in slight depression and superficial, with a density of 4.5 ± 0.85 stomata/mm² and an index of 7.92 ± 1.32%, within the range of variation of the genus. The thickness of the cuticle (4.50 ± 0.37 µm) adds a value that is poorly documented for the subspecies and useful for future comparisons [23], showing similarities with what has been described in leaves of species of the Orchidaceae family such as Cattleya caulescens and C. endsfeldzii, which also have thickened cuticles with suprastomatal chambers and uniseriate epidermis [31], as well as with what has been reported in Epidendrum radicans, where a 3-4 µm cuticle, uniseriate epidermis, and paracytic stomata on the adaxial surface are observed [32]. The tetracytic-anomocytic predominance and low stomatal index suggest optimization of gas exchange under high humidity conditions, while the cuticle thickness (4.5 µm) offers a diagnostic character to differentiate this subspecies from other taxa of the pompona complex. Along these lines, the results of Barreda-Castillo et al. [33] provide complementary evidence, showing that V. pompona and its hybrids with V. planifolia exhibit a differential response to induced water stress, with a lower reduction in stomatal conductance, leaf area, and water content compared to V. planifolia. These responses are consistent with the anatomy documented here, particularly with sparse stomata, thickened cuticle, and compact mesophyll, which promote efficient water use in humid environments while also offering resilience under conditions of water variation.
In the stem, the continuous ring of perivascular sclerenchyma (1-4 layers), the well-defined bundles with fascicular sheaths, and the glabrescence support a functional interpretation of mechanical support and resistance in a climbing and/or epiphytic habit, consistent with classic descriptions of peripheral reinforcement and water storage in Vanilla [23,26]. The xylem architecture (vessels of 35.52 ± 4.27 µm) with annular, spiral, and reticulate thickenings, as well as areolate and scalariform pores, suggests efficient conduction with mechanical flexibility in young organs [23,26]. In turn, these characteristics find parallels in pseudobulbs and rhizomes of other orchids, such as Cattleya and Oncidium, where collateral bundles appear surrounded by fibers and associated with reserve cells [31,34], and in Epidendrum radicans, whose stems show a uniseriate epidermis, a 2-6 µm cuticle, and collateral bundles covered by sclerenchymatic reinforcement [32]. The thicknesses of the epidermis (34.09 ± 5.92 µm) and cuticle (5.26 ± 1.90 µm) provide functional metrics for exploring tolerance to desiccation and insolation, aspects that are rarely quantified. The coexistence of peripheral sclerenchymatic reinforcement and medium-diameter vessels reflects mechanical support and hydraulic efficiency in a climbing-epiphytic habit, providing metrics that could be used as differential criteria compared to related species.
At the root, the combination of a uniseriate velamen, exodermis with thickened walls, thin-walled parenchymatic cortex, abundant pith, and a vascular cylinder with collateral bundles and large-diameter xylem elements matches the aerial root model of the genus [26,35]. More recent observations show that in V. phaeantha the structure varies between free aerial roots, roots attached to the phorophyte, and buried roots, with differences in cell wall epitopes reflecting adaptive plasticity [36], and in V. pompona, the sequence of adhesion stages has been documented with variations in velamen development, the presence of raphides, and the abundance of root hairs depending on the attachment substrate [37].
In the histochemical results, the positive reaction to quinones in the leaf mesophyll supports the presence of phenolic compounds with defensive and oxidative functions, consistent with the inclusions reported for the genus [23]. The lipid signal in cauline sclerenchyma bands suggests cutinized and suberized impregnations associated with waterproofing and mechanical reinforcement in epiphytes [26]. In roots, positive phloroglucinol confirms xylem lignification, while marked staining for starch emphasizes the reserve role of the cortex and pith in epiphytic environments with high water variability [35,38], similar to that observed in Saoncella et al. [34], where lignification in the exodermis and the presence of crystals and flavonoids in the cortex of epiphytic roots are reported. In line with these observations, recent studies show that Vanilla pompona has a different chemical profile from V. planifolia, characterized by the presence of benzyl alcohol, ethyl phenyl alcohol, and benzaldehyde in the early stages of refining, along with a lower relative content of vanillin [39]. This chemical differentiation is consistent with the abundance of starchy reserves and phenolic compounds detected in the vegetative organs of V. pompona subsp. grandiflora, suggesting that anatomical specialization in storage and defense structures is also reflected in its diversity of secondary metabolites. Thus, the anatomical features and chemical profiles reported coincide in highlighting the subspecies' ability to respond to conditions of high humidity and environmental variability, while providing differential criteria compared to V. planifolia.
5. Conclusions
The anatomy of Vanilla pompona subsp. grandiflora reveals adaptations to high humidity and a hemiepiphytic climbing habit. These adaptations are evident in support and reserve structures, including cauline sclerenchyma, root velamen, and abundant starch in the roots. Notably, among the quantitative traits not previously analyzed in the genus Vanilla are cuticular thickness (4.5 µm), low stomatal index (7.9%), and large root diameter (1.48 mm). These traits strengthen the interpretation and provide comparative characteristics with Neotropical congeners. These results expand upon anatomical information, now complemented by a basic histochemical assessment, and go beyond what had previously been reported from cultivated materials. They also offer diagnostic criteria for taxonomic delimitation of the group. Together, these results highlight the importance of integrating anatomical knowledge into biodiversity research, as they contribute baseline data for in situ conservation, sustainable management, and the preservation of genetic resources of Amazonian orchids.
Author Contributions
M.P.-M. and S.T.-A. conceived and designed the study; M.Q.-S. and M.M.B. conducted the histological and histochemical analyses; B.M.S. contributed to the anatomical interpretation and provided critical revisions; D.G.-T. coordinated the project and supervised the laboratory work; Y.Q.-G. and J.M.-B. performed data processing, analyses, and contributed to the ecological interpretation; M.P.-M., S.T.-A., and M.Q.-S. prepared the first draft of the manuscript; all authors contributed to the review and approval of the final version of the manuscript.
Acknowledgments
We thank our friends at ADECARAM Tingana and Finca Agroecológica Don Pepito for their valuable collaboration in carrying out the study and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Taylor, A.; Zotz, G.; Weigelt, P.; Cai, L.; Karger, D.N.; König, C.; Kreft, H. Vascular epiphytes contribute disproportionately to global centres of plant diversity. Glob. Ecol. Biogeogr. 2021, 30, 2584–2598. [Google Scholar] [CrossRef]
- Chase, M.W.; Cameron, K.M.; Freudenstein, J.V.; Pridgeon, A.M.; Salazar, G.; van den Berg, C.; Schuiteman, A. An updated classification of Orchidaceae. Bot. J. Linn. Soc. 2015, 177, 151–174. [Google Scholar] [CrossRef]
- Ministerio del Ambiente. Orquídeas del Perú y herramientas para su identificación. Ministerio del Ambiente, 2017. Available online: https://www.minam.gob.pe/diversidadbiologica/wp-content/uploads/sites/131/2018/12/LIBRO-ORQUIDEAS-2017_.pdf (accessed on 15 September 2025).
- Zotz, G.; Weigelt, P.; Kessler, M.; Kreft, H.; Taylor, A. EpiList 1.0: A global checklist of vascular epiphytes. Ecology 2021, 102, e03326. [Google Scholar] [CrossRef]
- Gastelbondo, M.; Nicholls, U.; Chen, S.; Chambers, A.; Wu, X. First gynogenesis of Vanilla planifolia for haploid production and ploidy verification protocol. Plants 2024, 13, 1733. [Google Scholar] [CrossRef]
- Karremans, A.P.; Chinchilla, I.F.; Rojas-Alvarado, G. Studies on Costa Rican Vanilloideae: The return of Cleistes rosea and a reaffirmation of Epistephium ellipticum. Lankesteriana 2023, 23, 613–622. [Google Scholar] [CrossRef]
- Azofeifa-Bolaños, J.B.; Paniagua-Vásquez, A.; García-García, J.A. Importancia y desafíos de la conservación de Vanilla spp. (Orchidaceae) en Costa Rica. Agron. Mesoam. 2014, 25, 189–202. Available online: http://www.scielo.sa.cr/scielo.php?script=sci_arttext&pid=S1659-13212014000100019 (accessed on 15 September 2025). [CrossRef]
- Leyva, V.E.; Lopez, J.M.; Zevallos-Ventura, A.; Cabrera, R.; Cañari-Chumpitaz, C.; Toubiana, D.; Maruenda, H. NMR-based leaf metabolic profiling of Vanilla planifolia and three endemic Vanilla species from the Peruvian Amazon. Food Chem. 2021, 358, 129365. [Google Scholar] [CrossRef]
- Rocha-Flores, R.G.; Herrera-Cabrera, B.E.; Velasco-Velasco, J.; Salazar-Rojas, V.M.; Delgado-Alvarado, A.; Mendoza-Castillo, M.C. Determinación preliminar de componentes de rendimiento para el cultivo de vainilla (Vanilla planifolia Jacks. ex Andrews) en la región Totonacapan, México. Agro Prod. 2018, 11, 9–14. [Google Scholar]
- Bouétard, A.; Lefuvre, P.; Gigant, R.; Bory, S.; Pignal, M.; Besse, P.; Grisoni, M. Evidence of transoceanic dispersion of the genus Vanilla based on plastid DNA phylogenetic analysis. Mol. Phylogenet. Evol. 2010, 55, 621–630. [Google Scholar] [PubMed]
- Rodríguez-Deméneghi, M.V.; Ramírez-Mosqueda, M.A.; Armas-Silva, A.A.; Aguilar-Rivera, N.; Gheno-Heredia, Y.A. Biofábricas de vainilla (Vanilla planifolia) en México como oportunidad de desarrollo agrario. Cuad. Biodivers. 2022, 63, 49–54. [Google Scholar] [CrossRef]
- Quinteros-Gómez, Y.; Zarco-González, M.; Gómez Ticerán, D.; Endara-Agramont, A.; Monroy-Vilchis, O. Effects of human disturbance on above-ground carbon stocks in north-west Amazonian Mauritia flexuosa peat swamp forests. Mires and Peat 2023, 29, 1–19. [Google Scholar]
- Humboldt, A. Essai politique sur le royaume de la Nouvelle-Espagne, 2 vols.; F. Schoell: Paris, France, 1811; p. 1–[vol. 2 end]. [Google Scholar]
- Humboldt, A.; Bonpland, A.; Kunth, C. Nova Genera et Species Plantarum, Quarto ed.; Librariae Graeco-Latino-Germaniae: Paris, France, 1815; Volume 1, pp. 1–377. [Google Scholar]
- Kraenzlin, F. Plantae novae andinae imprimis weberbauerianae I. Bot. Jahrb. Syst. 1906, 37, 373–398. [Google Scholar]
- Schlechter, R. Die Orchideenfloren der sudamerikanischen Kordillerenstaaten, IV. Perú. Repert. Spec. Nov. Regni Veg., Beihefte 1921, 9, 80–400. [Google Scholar]
- Schweinfurth, C. Orchids of Perú. Fieldiana Bot. 1958, 30, 1–260. [Google Scholar]
- Damián-Parizaca, L. Taxonomía del género Vanilla Plum. ex Mill. (Orchidaceae: Vanilleae) en el Perú. Bachelor’s Thesis, Universidad Nacional Mayor de San Marcos, Lima, Perú, 2020. [Google Scholar]
- Householder, E.; Janovec, J.; Balarezo, A.; Maceda, H.; Wells, J.; Valega, R. Diversity, natural history, and conservation of Vanilla (Orchidaceae) in Amazonian wetlands of Madre de Dios, Perú. J. Bot. Res. Inst. Tex. 2010, 4, 227–243. [Google Scholar]
- Janovec, J.; Householder, J.E.; Tobler, M. Evaluación de los actuales impactos y amenazas inminentes en aguajales y cochas de Madre de Dios; World Wildlife Fund (WWF): Lima, Perú, 2013; pp. 1–244. [Google Scholar]
- Soto, M.A.; Cribb, P. A new infrageneric classification and synopsis of the genus Vanilla Plum. ex Mill. (Orchidaceae: Vanillinae). Lankesteriana 2010, 9, 355–398. [Google Scholar]
- Herrera, X. Aprovechamiento de la fibra de coco y la hojarasca para la propagación de vainilla (Vanilla pompona subsp. grandiflora (Lindl.) y Vanilla odorata C. (Presl)) con fines de mitigación ambiental y conservación de la biodiversidad. Bachelor’s Thesis, Universidad Peruana Unión, Lima, Perú, 2019. Available online: https://repositorio.upeu.edu.pe/items/f4c465d5-6907-4fe6-adec-c15ddebe593 (accessed on 15 September 2025).
- Raffi, A.; Abdullah; Yunus, N. -S.M.; Go, R. Preliminary foliar anatomical assessment of four Vanilla species (Orchidaceae) from Perak, Malaysia. Malays. Appl. Biol. 2019, 42, 807–812. Available online: https://openurl.ebsco.com/EPDB%3Agcd%3A13%3A33252973/detailv2?sid=ebsco%3Aplink%3Ascholar&id=ebsco%3Agcd%3A136874117&crl=c&link_origin=scholar.google.com.pe (accessed on 15 September 2025).
- Martínez-Quezada, D.M.; Sandoval-Zapotitla, E.; Solís-De la Cruz, J.; Velázquez-Vázquez, D.E.; Herrera-Cabrera, E. Caracterización anatómica y análisis de variación de epidermis foliar y caulinar entre dos genotipos de Vanilla planifolia Jacks. ex Andrews. Agro Prod. 2018, 9, 1. Available online: https://revista-agroproductividad.org/index.php/agroproductividad/article/view/704 (accessed on 15 September 2025).
- Odoux, E.; Brillouet, J.-M. Anatomy, histochemistry and biochemistry of glucovanillin, oleoresin and mucilage accumulation sites in green mature vanilla pod (Vanilla planifolia; Orchidaceae): A comprehensive and critical reexamination. Fruits 2009, 64, 221–241. [Google Scholar] [CrossRef]
- Stern, W.L.; Judd, W.S. Comparative vegetative anatomy and systematics of Vanilla (Orchidaceae). Bot. J. Linn. Soc. 1999, 131, 353–382. [Google Scholar] [CrossRef]
- Flores Jiménez, Á.; López, D.R.; García, D.J.; Arenas, O.R.; Rivera, A.; Lara, M.H.; Silva, A.P. Diversidad de Vanilla spp. en México: Distribución geográfica y perfiles bioclimáticos. Rev. Mex. Cienc. For. 2017, 8, 62–86. [Google Scholar] [CrossRef]
- Quinteros-Gómez, Y.M.; Cabrera Mestanza, D.; Macedo Bedoya, J.; Santos Linares, V.; Salinas Inga, A. Propagación vegetativa de Vanilla pompona subsp. grandiflora (Orchidaceae) en territorios inundables del Valle del Alto Mayo, Perú. Acta Bot. Mex. 2024, 131. [Google Scholar] [CrossRef]
- Ahmad, H.; Khera, R.A.; Hanif, M.A.; Ayub, M.A.; Jilani, M.I. Vanilla. In Elsevier EBooks; Elsevier: Amsterdam, The Netherlands, 2019; pp. 657–669. [Google Scholar] [CrossRef]
- Nakata, P. A. (2003). Advances in our understanding of calcium oxalate crystal formation and function in plants. Plant Science 2003, 164, 901–909. [Google Scholar] [CrossRef]
- Ribeiro, J.; Paula-Souza, J.; da Silva, C. Morfoanatomia de órgãos vegetativos de duas espécies de Cattleya (Orchidaceae) nativas do Brasil. Rodriguésia 2020, 71. [Google Scholar] [CrossRef]
- Muthukumar, T.; Shenbagam, M. Vegetative anatomical adaptations of Epidendrum radicans (Epidendroideae, Orchidaceae) to epiphytic conditions of growth. Mod. Phytomorphol. 2017, 11, 117–130. [Google Scholar] [CrossRef]
- Barreda-Castillo, J.M.; Monribot-Villanueva, J.L.; Velázquez-Rosas, N.; Bayman, P.; Guerrero-Analco, J.A.; Menchaca-García, R.A. Morphological and physio-chemical responses to PEG-induced water stress in Vanilla planifolia and V. pompona hybrids. Int. J. Mol. Sci. 2023, 24, 4690. [Google Scholar] [CrossRef] [PubMed]
- Saoncella, A.L.; Marteline, M.A.; de Moraes, C.P. Anatomia dos órgãos vegetativos de Cattleya violacea (Kunth) Rolfe (Orchidaceae). Iheringia, Sér. Bot. 2017, 72, 114–126. [Google Scholar] [CrossRef]
- de Lima, J.F.; Moreira, A.S.F.P. Structural plasticity in roots of the hemiepiphyte Vanilla phaeantha Rchb.f. (Orchidaceae): A relationship between environment and function. Sci. Nat. 2022, 109, 46. [Google Scholar] [CrossRef] [PubMed]
- de Lima, J.F.; Coelho de Oliveira, D.; Coelho Kuster, V.; Franco Pinheiro Moreira, A.S. Aerial and terrestrial root habits influence the composition of the cell walls of Vanilla phaeantha (Orchidaceae). Protoplasma 2024, 262, 87–98. [Google Scholar] [CrossRef]
- Deseo, N.B.; de Guzman, C.C.; Nuevo, P.A.; Torreta, N.K. Morpho-Anatomy of Adventitious Roots of Vanilla (Vanilla planifolia Jacks. ex Andrews) during Attachment to Support Post. Thailand Nat. Hist. Mus. J. 2020, 14, 31–36. [Google Scholar]
- Botomanga, A.; Jeannoda, V.H.; Fuzzati, N.; Ramarosandratana, A.V. Morpho-anatomical responses of leafless Vanilla spp. roots to drought and habitat degradation. Flora 2024, 317, 152562. [Google Scholar] [CrossRef]
- Ravier, A.; Chalut, P.; Belarbi, S.; Santerre, C.; Vallet, N.; Nhouchi, Z. Impact of the post-harvest period on the chemical and sensorial properties of Vanilla planifolia and V. pompona. Molecules 2024, 29, 839. [Google Scholar] [CrossRef] [PubMed]
- Hickey, L.J. Classification of the architecture of dicotyledonous leaves. Am. J. Bot. 1973, 60, 17–33. [Google Scholar] [CrossRef]
- Ash, A.; Ellis, B.; Hickey, L.J.; Johnson, K.; Wilf, P.; Wing, S.L. Manual of Leaf Architecture: Morphological Description and Categorization of Dicotyledonous and Net-Veined Monocotyledonous Angiosperms; Smithsonian Institution: Washington, DC, USA, 1999. [Google Scholar] [CrossRef]
- D’Ambrogio de Argüeso, A. Manual de Técnicas en Histología Vegetal; Hemisferio Sur S.A.: Buenos Aires, Argentina, 1986. [Google Scholar]
- Lock, O. Investigación Fitoquímica: Métodos en el Estudio de Productos Naturales, 2ª ed.; Pontificia Universidad Católica del Perú: Lima, Perú, 1994. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Evans, W.C. Trease y Evans Farmacognosia, 13th ed.; Interamericana-McGraw-Hill: Madrid, Spain, 1991. [Google Scholar]
Figure 1.
Map of the collection site of Vanilla pompona subsp. grandiflora in Moyobamba, San Martín, Peru.
Figure 1.
Map of the collection site of Vanilla pompona subsp. grandiflora in Moyobamba, San Martín, Peru.

Figure 2.
Vegetative features of Vanilla pompona subsp. grandiflora. From left to right: habit with aerial roots, stem, and foliage leaves.
Figure 2.
Vegetative features of Vanilla pompona subsp. grandiflora. From left to right: habit with aerial roots, stem, and foliage leaves.

Table 2.
Quantitative parameters analyzed in the vegetative organs of Vanilla pompona subsp. grandiflora (Lindl.) Soto Arenas.
Table 2.
Quantitative parameters analyzed in the vegetative organs of Vanilla pompona subsp. grandiflora (Lindl.) Soto Arenas.
| Quantitative parameter | Measurement |
| Mesophyll thickness (µm) | 1144.88 ± 54.17 |
| Stomatal density (stomata/mm²) | 4.5 ± 0.85 |
| Stomatal index (%) | 7.92 ± 1.32 |
| Leaf cuticle thickness (µm) | 4.50 ± 0.37 |
| Stem epidermis thickness (µm) | 34.09 ± 5.92 |
| Stem cuticle thickness (µm) | 5.26 ± 1.90 |
| Stem xylem vessel diameter (µm) | 35.52 ± 4.27 |
| Root diameter (µm) | 1482.55 ± 10.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



