
Submitted:
26 September 2025
Posted:
26 September 2025
You are already at the latest version
Abstract
The zoophilic and exophilic traits of outdoor-biting Anopheles has led to this group largely being overlooked for their role in malaria transmission, despite several species now recognized as locally important in regions of sub-Saharan Africa. Given the current limitations with identification of these understudied species, it is crucial to accurately correlate morphological features to molecular data. Here, we produced high quality reference sequence data for representative understudied anopheline species to better understand the phylogenetic relationships between under- and well-studied vectors of malaria. For mitochondrial genome assembly, shallow shotgun sequencing was implemented on single mosquito specimens and phylogenetic analyses were performed on the concatenated protein coding genes of the mitogenomes using a Bayesian approach. This study generated 10 complete mitogenomes focusing on less-studied taxa with an average length 15,383 bp and A-T content of 77.1% consistent with other anophelines containing 37 genes. Bayesian inference analysis yielded four main clades with molecular dating indicating that well-studied malaria vectors diverged from outdoor-biting species more than 65 million years ago. These findings support the taxonomic grouping of mosquitoes belonging to the Anopheles genus based on morphological characteristics and can provide molecular diagnostics for species identification enabling more precise and adept interventions for malaria control.
Keywords:
Outdoor biting
; mitochondrial genome
; malaria
; Anopheles
; understudied
1. Introduction
As African nations strive towards malaria control and elimination, insecticide re-sistance and residual malaria transmission challenge current vector-targeted malaria in-terventions [1,2,3,4]. While the primary vectors Anopheles funestus, An. coluzzii and An. gambiae are the focus of these mitigation strategies [5,6], selection pressure from indoor-focused measures such as indoor residual spraying (IRS) and insecticide treated nets (ITNs) have led to changes in mosquito behavior and populations [7,8,9]. One key change is the recog-nition that secondary Anopheles vectors such as An. rufipes [10,11], An. pharoensis [12], An. squamosus [13] and An. coustani [13,14,15], contribute to sustaining residual transmission in sub-Saharan Africa. These mosquito species are considered largely exophagic and exoph-ilic, behaviors that have allowed them to evade indoor vector control interventions [7,8,16,17].
Despite frequent collection alongside the primary malaria vectors, the genomics, ecology, biology, and behavior of these long-overlooked anopheline mosquito species are poorly understood [9]. Furthermore, the constraints of overlapping and cryptic morpho-logical features [18,19,20,21], together with the absence of available molecular data in genomic databases [17,20,22,23], have made robust morphological and molecular identification of these less studied anopheline species extremely challenging. Studies have also reported evidence of functional heterogeneity in anopheline genomes which influence their behav-ioral plasticity, a crucial characteristic for defining vectorial capacity and adaptability [24,25]. Therefore, the accurate identification and bionomic characterization of under-studied Anopheles species is now critical given their key role as local vectors in driving re-sidual malaria transmission in Zambia, Madagascar, southern Mozambique, Ethiopia and Kenya [13,15,26,27,28,29].
Although there is an extensive list of Anopheles sequences generated using molecular barcodes based on the cytochrome oxidase I (COI) and internal transcribed region 2 (ITS2) genes [12,17,18,20,26,27,28,30,31,32,33,34,35], there still remain limitations in available sequence for these understudied Anopheles species to produce robust differentiation between members of closely related taxa [17,18,31,36]. This includes cryptic species which may be incrimi-nated in residual malaria transmission but have been allotted placeholder names such as An. species 11 [17,18,20], An. species 15 [18] and An. species unknown group 1 [20], par-ticularly in the absence of comprehensive morphological identification to compliment the generated molecular barcode sequences. Furthermore, the use of the single COI gene to validate identification for less studied Anopheles has produced matches with low similari-ties (less than 80%) and weakly supported phylogenies. For instance, this has led to in-conclusive identities for members of the An. coustani group in earlier studies from Zambia [17,31] and Mozambique [26]. Recently, mitochondrial genomes were used to provide conclusive identities and differentiate the cryptic taxa of the An. coustani group into phy-logenetically well-supported taxonomic clades [37].
The acquisition of genomic datasets has become more accessible due to the expan-sion of sequencing and computational technologies, including mitochondrial genomes (mitogenomes) which have shown to be useful in the identification and resolution of phy-logenies for several mosquito species and species groups [38,39,40]. These circular, double stranded DNA molecules encode 37 genes, including 13 protein coding genes (PCGs), 22 transfer RNA (tRNA), 2 ribosomal RNA (rRNA) and a non-coding control region [40,41]. In addition to the 13 PCGs, key characteristics such as low incidence of recombination, high copy number, and maternal inheritance make the mitogenome a more effective tax-onomic tool compared to single barcodes [41,42,43,44]. Expanding mitochondrial genome re-sources to include less-studied mosquito species is essential for accurate species delinea-tion and gaining insights into mosquito ecology and systematics for public health inter-ventions. In this study, we aimed to generate high quality reference sequence data for rep-resentative understudied mosquito species and demonstrate the strength of mitogenomes compared to prior studies that were limited to the COI gene in attempts to resolve phy-logenies.
2. Materials and Methods
2.1. Mosquito collections
Specimens were collected during routine entomological surveillance in Nchelenge, Zambia in 2023-2024 as part of the Southern and Central Africa International Centers of Excellence for Malaria Research (ICEMR) investigations. Miniature CDC Light Traps (John W. Hock Co., Gainesville, Florida), were positioned both indoors and outdoors where people gather in the evening and near to animal pens.
2.2. DNA extraction and sequencing
DNA extractions using a modified extraction method [45] were performed on single mosquito specimens morphologically identified as An. rufipes, An. maculipalpis, An. preto-riensis, An. squamosus and An. pharoensis [21]. The extracted DNA and previously extracted specimens identified using the COI gene as species 11, species 15, unknown group 1, un-known group 2 and unknown group 3 from a previous study [20] were quantified using the Qubit dsDNA assay kit (Thermo Fisher Scientific, Waltham, MA) and shipped to Se-qCenter (Pittsburgh, USA) for library construction and Illumina sequencing. The libraries were 150 bp paired end sequenced to a count of 13.3 million reads per sample.
2.3. Mitochondrial genome assembly and annotation
The mitochondrial genomes were assembled using NOVOPlasty [46] (RRID:SCR_017335) version 4.3.5 with k-mer set at 39 base pairs and An. squamosus (OP_77691) as the seed sequence. Using the MITOchondrial genome annotation (MITOS) [47] galaxy tool, generated contigs were automatically annotated using the invertebrate genetic code under default settings. The start and stop codon positions of the annotated contigs were manually adjusted in Geneious Prime (RRID:SCR_010519) version 2025.1.2 (Biomatters, Auckland, New Zealand) using reference anopheline mitochondrial genomes as a guideline. The generated contigs with corresponding annotations were submitted to the GenBank database for the assignment of accession numbers
2.4. Phylogenetic analysis and tree construction
Using jModelTest (v2.1.10) [48], the best fit base pair substitution model based on the Akaike information criterion (AIC) and the Bayesian information criterion (BIC) was de-termined under default settings using an aligned sequence matrix. This alignment was generated using the MAFFT alignment tool implemented in the Geneious Prime (RRID:SCR_010519) version 2025.1.2 (Biomatters, Auckland, New Zealand from the 13 concatenated PCGs of mitogenomes generated in this study, available mitochondrial ge-nomes of understudied African anopheline species; An. marshallii (NC_064607), An. mou-cheti (NC_064608), An. gibbinsi (OR_569715), An. nili (NC_064610), An. coustani group (PQ_585798, PQ_587039, PQ_587041, PQ_587036, PP_375116), and reference mitoge-nome sequences for the well-studied species An. gambiae (MG_930894) and the An. funestus group (MG_742172, MG_742194, MT_917162, MT_917137, MT_91714, MT_917157, MT_917163). Using Bayesian Evolutionary Analysis by Sampling Trees (BEAST) 2 soft-ware [49], inference analysis was performed on the aligned sequence matrix using tree independent runs and a 20% burn-in rate for tree building purposes under default set-tings. Bayesian analysis was also performed on an alignment generated from COI se-quences available from GenBank complementary to representative species in the mito-chondrial genome tree. Trees were visualized and annotated using FigTree v.1.4.4 (http://tree.bio.ed.ac.uk/software/figtree/).
2.5. Dating time estimation
Divergence time estimations were calculated using the previously mentioned pa-rameters used for Bayesian inference in BEAST2. The frequently referenced Ae-des-Anopheles divergence time of approximately 154.7 million years ago (MYA) [50] was used as the calibration point set to normal distribution.
3. Results
The 10 mitochondrial genomes produced in this study were consistent with other Af-rican anopheline mitogenomes represented in NCBI’s GenBank database as comprised of 13 PCGs, 22 tRNAs and 2 rRNAs, with lengths ranging from 15,534 bp (Unknown group 2) to 15,346 bp (An. pharoensis) and a mean AT content of 77.1 % (Table 1).
Bayesian inference for the mitochondrial genomes resulted in a phylogenetic tree which separated specimen sequences (Figure 1) compared to COI tree which resulted in 4 weakly supported main clades (Figure S1). The most recent common ancestor (MRCA) for An. funestus and An. gambiae with the outdoor biting Anopheles included in this study was 66.45 and 69.65 MYA respectively (Figure 2).
4. Discussion
Bayesian phylogenetic analysis based on the concatenated PCGs from the mitoge-nomes was able to clearly delineate less studied African anopheline taxa compared to phylogenetic analyses using exclusively the COI gene as in earlier studies [17,20]. Several of these species are often misidentified during morphological examination or matched to an unassigned species following molecular barcoding [20,26,33]. This may be attributed at least in part to the unknown diversity of outdoor biting anophelines and relative inexpe-rience in morphological identification of these numerous taxa, some of which may yet to be fully described due to the bias placed on understanding highly anthropophilic, endophagic and endophilic species [6,9,17,28,31,31]. The uncertainty of vector richness of the exophagic anophelines together with their perceived adaptability and undetermined foraging times, habitats, and opportunistic feeding patterns have led to ambiguous spe-cies assignments [9,14,28,33,51]. This combination of characteristics emphasizes the need for the expansion of outdoor surveillance and investigation of the anophelines found in this space, as well as innovative strategies to overcome the shortcomings of in-door-focused interventions.
A growing number of understudied anophelines have been confirmed or implicated in human malaria transmission. Despite historic consideration as a secondary vector [12,17,27,52], An. coustani is now regarded as a primary local vector in regions of Mada-gascar [15,29,53]. Additionally, previous studies have identified An. pretoriensis [34], An. pharoensis [52,54], and cryptic anopheline species [17,18,20] demonstrating opportunistic feeding patterns on humans, some infected with sporozoites of human malaria parasite species. Anopheles squamosus is another species strongly implicated as a malaria vector, with a wide geographical range, and has demonstrated variable foraging behavior to-wards human blood meals [13,55,56]. Related to this are a number of ‘molecular taxa’. Examples include An. species 11 and An. species 15 which are often morphologically keyed as An. squamosus but are differentiated by the COI barcode and even more strongly by the mitogenome sequence (Fig. 1)[17,20]. Others include An. unknown groups 1 – 3 for which morphology and molecular barcoding was inconclusive [20]. Here the mitogenome data provide the most comprehensive insight into the identification of these ‘molecular taxa’, but as with prior studies, without a more extensive sequence database of recognized species, these specimens remain taxonomically unresolved. The fact that many of these exophagic taxa cluster together in the phylogenetic analysis may be an artifact of their shared ancestry and that these share behavioral adaptations may have been reinforced over millennia.
It is clear that full mitogenomes offer much more discriminatory power for a phylo-genetic approach to inquire about shared biological traits and possibly ascertain whether behaviors such as biting preference are due to recent adaptations or reflect the existence of genetically distinct lineages which may have been overlooked when restricted to morpho-logical identification. Dating time estimations from well-recognized malaria vectors fur-ther corroborate the presence of these outdoor biting Anopheles as cryptic lineages with distinct ecological niches, suggestive of understudied species that may maintain trans-mission outdoors, perhaps under certain conditions such as relative absence of non-human hosts, or human behavior that promotes high opportunistic human biting rates. Furthermore, the accurate taxonomic placement of these mosquitoes highlights the relationships between known vectors and putative vector species which may provide fur-ther insights into understanding the differences in biting, foraging, and vectorial capacity of these less studied species. Linking morphological reference specimens to genomic data is key for the accurate identification given the status of unassigned Anopheles species with sporozoites collected in the field [18,19,26,52].
5. Conclusions
Although reference sequences are available for many commonly encountered outdoor biting anopheline species, there remains a paucity of data to accurately identify and taxo-nomically place these species in the wider Anopheles genus. This study contributes valu-able genetic datasets representing exophagic species collected in Zambia and present across the African continent. The generation of mitochondrial genomes for cryptic unas-signed species that are commonly collected has given priority to the use integrative tax-onomy in future research. The linking of molecular data with morphological and type specimens can further strengthen the credibility of species delimitation for the assigned zoological nomenclature of these cryptic taxa. The analyses from this study identified the phylogenetic relationships between the primary malaria vectors and understudied species implicated in malaria transmission, assisting to close the genetic gap of what we know about these anophelines of public health importance.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/doi/s1, Figure S1: Bayesian tree showing the phylogenetic rela-tionship of less studied Anopheles mosquito species with main vectors on malaria using the cy-tochrome oxidase I gene (COI) using. The posterior probabilities supporting the tree topology are represented by the values at the nodes.
Author Contributions
R.L.M.N.A. and D.E.N. conceived and designed the study. M.E.G., L.S., K.S., W.H., H.C. and M.M. performed field collections and morphological identification of mosquito specimens. R.L.M.N.A worked on laboratory extractions and bioinformatic pipelines for generated datasets. D.E.N. and R.L.M.N.A. drafted the manuscript. C.J.M and A.C.M reviewed and approved the manuscript with all authors. W.J.M. and D.E.N. attained funding, read and approved the manuscript with all authors.
Funding
This research was funded in part by funds from the National Institute of Allergy and Infectious Diseases (NIAID) of the National Institutes of Health (U19AI089680),T32 support to M.E.G. (T32AI138953) and A.C.M. (T32AI138953), a Johns Hopkins Malaria Research Institute Postdoctoral Award to R.L.M.N.A., the Bloomberg Philanthropies, and NSF-Accelerator Project D-688: Computing the Biome (2134862).
Data Availability Statement
The mitochondrial genomes are available with accession numbers PP068255 – PP068257, PP068259, PP093765, PV943467 – PV943469, PX240906 and PX257875.
Acknowledgments
We would like to thank the Zambian study teams at the Tropical Diseases Research Centre and Macha Research Trust. We are grateful to the communities of Nchelenge and Choma Districts for their cooperation.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
COI Cytochrome Oxidase I
ITS2 Internal transcribed spacer 2 region
PCGs Protein coding genes
tRNA Transfer RNA
rRNA Ribosomal RNA
ICEMR International Centers of Excellence for Malaria Research
AIC Akaike information criterion
BIC Bayesian information criterion
BEAST Bayesian Evolutionary Analysis by Sampling Trees
MYA Million years ago
References
- Rodriguez, M.H. Residual Malaria: Limitations of Current Vector Control Strategies to Eliminate Transmission in Residual Foci. J Infect Dis 2021, 223, S55–S60. [Google Scholar] [CrossRef] [PubMed]
- Zinszer, K.; Talisuna, A.O. Fighting Insecticide Resistance in Malaria Control. The Lancet Infectious Diseases 2023, 23, 138–139. [Google Scholar] [CrossRef]
- Sherrard-Smith, E.; Winskill, P.; Hamlet, A.; Ngufor, C.; N’Guessan, R.; Guelbeogo, M.W.; Sanou, A.; Nash, R.K.; Hill, A.; Russell, E.L.; et al. Optimising the Deployment of Vector Control Tools against Malaria: A Data-Informed Mod-elling Study. The Lancet Planetary Health 2022, 6, e100–e109. [Google Scholar] [CrossRef]
- Sanou, A.; Nelli, L.; Guelbéogo, W.M.; Cissé, F.; Tapsoba, M.; Ouédraogo, P.; Sagnon, N.; Ranson, H.; Matthiopoulos, J.; Ferguson, H.M. Insecticide Resistance and Behavioural Adaptation as a Response to Long-Lasting Insecticidal Net Deployment in Malaria Vectors in the Cascades Region of Burkina Faso. Sci Rep 2021, 11, 17569. [Google Scholar] [CrossRef]
- Pryce, J.; Medley, N.; Choi, L. Indoor Residual Spraying for Preventing Malaria in Communities Using Insecticide-treated Nets - Pryce, J - 2022 | Cochrane Library.
- Sherrard-Smith, E.; Ngufor, C.; Sanou, A.; Guelbeogo, M.W.; N’Guessan, R.; Elobolobo, E.; Saute, F.; Varela, K.; Chaccour, C.J.; Zulliger, R.; et al. Inferring the Epidemiological Benefit of Indoor Vector Control Interventions against Malaria from Mosquito Data. Nat Commun 2022, 13, 3862. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.R.; Overgaard, H.J.; Abaga, S.; Reddy, V.P.; Caccone, A.; Kiszewski, A.E.; Slotman, M.A. Outdoor Host Seeking Behaviour of Anopheles gambiae Mosquitoes Following Initiation of Malaria Vector Control on Bioko Island, Equatorial Guinea. Malaria Journal 2011, 10, 184. [Google Scholar] [CrossRef]
- Musiime, A.K.; Smith, D.L.; Kilama, M.; Rek, J.; Arinaitwe, E.; Nankabirwa, J.I.; Kamya, M.R.; Conrad, M.D.; Dorsey, G.; Akol, A.M.; et al. Impact of Vector Control Interventions on Malaria Transmission Intensity, Outdoor Vector Biting Rates and Anopheles Mosquito Species Composition in Tororo, Uganda. Malaria Journal 2019, 18, 445. [Google Scholar] [CrossRef]
- Laurent, S.; Brandy Mosquito Vector Diversity and Malaria Transmission. Front. Malar. 2025, 3.
- Tabue, R.N.; Awono-Ambene, P.; Etang, J.; Atangana, J.; C, A.-N.; Toto, J.C.; Patchoke, S.; Leke, R.G.F.; Fondjo, E.; Mnzava, A.P.; et al. Role of Anopheles (Cellia) rufipes (Gough, 1910) and Other Local Anophelines in Human Malaria Transmission in the Northern Savannah of Cameroon: A Cross-Sectional Survey. Parasites & Vectors 2017, 10, 22. [Google Scholar] [CrossRef]
- Saili, K.; de Jager, C.; Sangoro, O.P.; Nkya, T.E.; Masaninga, F.; Mwenya, M.; Sinyolo, A.; Hamainza, B.; Chanda, E.; Fillinger, U.; et al. Anopheles rufipes Implicated in Malaria Transmission Both Indoors and Outdoors alongside Anopheles funestus and Anopheles arabiensis in Rural South-East Zambia. Malaria Journal 2023, 22, 95. [Google Scholar] [CrossRef] [PubMed]
- Cross, D.E.; Healey, A.J.E.; McKeown, N.J.; Thomas, C.J.; Macarie, N.A.; Siaziyu, V.; Singini, D.; Liywalii, F.; Sakala, J.; Silumesii, A.; et al. Temporally Consistent Predominance and Distribution of Secondary Malaria Vectors in the Anopheles Community of the Upper Zambezi Floodplain. Sci Rep 2022, 12, 240. [Google Scholar] [CrossRef] [PubMed]
- Fornadel, C.M.; Norris, L.C.; Franco, V.; Norris, D.E. Unexpected Anthropophily in the Potential Secondary Malaria Vectors Anopheles coustani s. l. and Anopheles squamosus in Macha, Zambia.Vector Borne Zoonotic Dis 2011, 11, 1173–1179. [Google Scholar] [CrossRef]
- Hendershot, A.L. Understanding the Role of An. Coustani Complex Members as Malaria Vector Species in the Democratic Republic of Congo. thesis, University of Notre Dame, 2021.
- Goupeyou-Youmsi, J.; Rakotondranaivo, T.; Puchot, N.; Peterson, I.; Girod, R.; Vigan-Womas, I.; Paul, R.; Ndiath, M.O.; Bourgouin, C. Differential Contribution of Anopheles Coustani and Anopheles Arabiensis to the Transmission of Plasmodium Falciparum and Plasmodium Vivax in Two Neighbouring Villages of Madagascar. Parasites & Vectors 2020, 13, 430. [Google Scholar] [CrossRef]
- Sougoufara, S.; Ottih, E.C.; Tripet, F. The Need for New Vector Control Approaches Targeting Outdoor Biting Anopheline Malaria Vector Communities. Parasites & Vectors 2020, 13, 295. [Google Scholar] [CrossRef]
- Lobo, N.F.; Laurent, B.S.; Sikaala, C.H.; Hamainza, B.; Chanda, J.; Chinula, D.; Krishnankutty, S.M.; Mueller, J.D.; Deason, N.A.; Hoang, Q.T.; et al. Unexpected Diversity of Anopheles Species in Eastern Zambia: Implications for Evalu-ating Vector Behavior and Interventions Using Molecular Tools. Sci Rep 2015, 5, 17952. [Google Scholar] [CrossRef]
- Zhong, D.; Hemming-Schroeder, E.; Wang, X.; Kibret, S.; Zhou, G.; Atieli, H.; Lee, M.-C.; Afrane, Y.A.; Githeko, A.K.; Yan, G. Extensive New Anopheles Cryptic Species Involved in Human Malaria Transmission in Western Kenya. Sci Rep 2020, 10, 16139. [Google Scholar] [CrossRef]
- Stevenson, J.C.; Norris, D.E. Implicating Cryptic and Novel Anophelines as Malaria Vectors in Africa. Insects 2017, 8, 1. [Google Scholar] [CrossRef]
- Jones, C.M.; Ciubotariu, I.I.; Muleba, M.; Lupiya, J.; Mbewe, D.; Simubali, L.; Mudenda, T.; Gebhardt, M.E.; Carpi, G.; Malcolm, A.N.; et al. Multiple Novel Clades of Anopheline Mosquitoes Caught Outdoors in Northern Zambia. Front. Trop. Dis 2021, 2. [Google Scholar] [CrossRef]
- Coetzee, M. Key to the Females of Afrotropical Anopheles Mosquitoes (Diptera: Culicidae). Malaria Journal 2020, 19, 70. [Google Scholar] [CrossRef] [PubMed]
- Moraes Zenker, M.; Portella, T.P.; Pessoa, F.A.C.; Bengtsson-Palme, J.; Galetti, P.M. Low Coverage of Species Con-strains the Use of DNA Barcoding to Assess Mosquito Biodiversity. Sci Rep 2024, 14, 7432. [Google Scholar] [CrossRef] [PubMed]
- Davidson, J.R.; Wahid, I.; Sudirman, R.; Small, S.T.; Hendershot, A.L.; Baskin, R.N.; Burton, T.A.; Makuru, V.; Xiao, H.; Yu, X.; et al. Molecular Analysis Reveals a High Diversity of Anopheles Species in Karama, West Sulawesi, Indonesia. Parasites & Vectors 2020, 13, 379. [CrossRef]
- Neafsey, Daniel E. , Robert M. Waterhouse, Mohammad R. Abai, Sergey S. Aganezov, Max A. Alekseyev, James E. Allen, James Amon et al. Highly evolvable malaria vectors: the genomes of 16 Anopheles mosquitoes. Science 2015, 347, 1258522.
- Bartilol, B.; Omuoyo, D.; Karisa, J.; Ominde, K.; Mbogo, C.; Mwangangi, J.; Maia, M.; Rono, M.K. Vectorial Capacity and TEP1 Genotypes of Anopheles gambiae sensu lato Mosquitoes on the Kenyan Coast. Parasites & Vectors 2022, 15, 448. [Google Scholar] [CrossRef]
- Máquina, M.; Opiyo, M.A.; Cuamba, N.; Marrenjo, D.; Rodrigues, M.; Armando, S.; Nhate, S.; Luis, F.; Saúte, F.; Candrinho, B.; et al. Multiple Anopheles Species Complicate Downstream Analysis and Decision-Making in a Malaria Pre-Elimination Area in Southern Mozambique. Malar J 2024, 23, 23. [Google Scholar] [CrossRef] [PubMed]
- Mustapha, A.M.; Musembi, S.; Nyamache, A.K.; Machani, M.G.; Kosgei, J.; Wamuyu, L.; Ochomo, E.; Lobo, N.F. Secondary Malaria Vectors in Western Kenya Include Novel Species with Unexpectedly High Densities and Parasite Infection Rates. Parasites & Vectors 2021, 14, 252. [Google Scholar] [CrossRef] [PubMed]
- Assa, A.; Eligo, N.; Massebo, F. Anopheles Mosquito Diversity, Entomological Indicators of Malaria Transmission and Challenges of Morphological Identification in Southwestern Ethiopia. Trop Med Health 2023, 51, 38. [Google Scholar] [CrossRef]
- Nepomichene, T.N.J.J.; Tata, E.; Boyer, S. Malaria Case in Madagascar, Probable Implication of a New Vector, Anopheles Coustani. Malar J 2015, 14, 475. [Google Scholar] [CrossRef]
- Jones, C.M.; Ciubotariu, I.I.; Gebhardt, M.E.; Lupiya, J.S.; Mbewe, D.; Muleba, M.; Stevenson, J.C.; Norris, D.E. Evalu-ation of Anopheline Diversity and Abundance across Outdoor Collection Schemes Utilizing CDC Light Traps in Nch-elenge District, Zambia. Insects 2024, 15, 656. [Google Scholar] [CrossRef]
- Ciubotariu, I.I.; Jones, C.M.; Kobayashi, T.; Bobanga, T.; Muleba, M.; Pringle, J.C.; Stevenson, J.C.; Carpi, G.; Norris, D.E. for the Southern and Central Africa International Centers of Excellence for Malaria Research Genetic Diversity of Anopheles Coustani (Diptera: Culicidae) in Malaria Transmission Foci in Southern and Central Africa. Journal of Medical Entomology 2020, 57, 1782–1792. [Google Scholar] [CrossRef]
- Salomé, G.; Riddin, M.; Braack, L. Species Composition, Seasonal Abundance, and Biting Behavior of Malaria Vectors in Rural Conhane Village, Southern Mozambique. International Journal of Environmental Research and Public Health 2023, 20, 3597. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, M.E.; Krizek, R.S.; Coetzee, M.; Koekemoer, L.L.; Dahan-Moss, Y.; Mbewe, D.; Lupiya, J.S.; Muleba, M.; Stevenson, J.C.; Moss, W.J.; et al. Expanded Geographic Distribution and Host Preference of Anopheles Gibbinsi (Anopheles Species 6) in Northern Zambia. Malaria Journal 2022, 21, 211. [Google Scholar] [CrossRef]
- Campos, M.; Patel, N.; Marshall, C.; Gripkey, H.; Ditter, R.E.; Crepeau, M.W.; Toilibou, A.; Amina, Y.; Cornel, A.J.; Lee, Y.; et al. Population Genetics of Anopheles Pretoriensis in Grande Comore Island. Insects 2022, 14, 14. [Google Scholar] [CrossRef]
- Antonio-nkondjio, C.; Kerah, C.H.; Simard, F.; Awono-ambene, P.; Chouaibou, M.; Tchuinkam, T.; Fontenille, D. Complexity of the Malaria Vectorial System in Cameroon: Contribution of Secondary Vectors to Malaria Transmission. Journal of Medical Entomology 2006, 43, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Calzolari, M.; Bellin, N.; Dottori, M.; Torri, D.; Di Luca, M.; Rossi, V.; Magoga, G.; Montagna, M. Integrated Taxon-omy to Advance Species Delimitation of the Anopheles Maculipennis Complex. Sci Rep 2024, 14, 30914. [Google Scholar] [CrossRef]
- Usmani, S.; Gebhardt, M.E.; Simubali, L.; Saili, K.; Hamwata, W.; Chilusu, H.; Muleba, M.; McMeniman, C.J.; Mar-tin, A.C.; Moss, W.J.; et al. Phylogenetic Taxonomy of the Zambian Anopheles Coustani Group Using a Mitogenomics Approach. Malaria Journal 2025, 24, 203. [Google Scholar] [CrossRef]
- Dong, Z.; Wang, Y.; Li, C.; Li, L.; Men, X. Mitochondrial DNA as a Molecular Marker in Insect Ecology: Current Status and Future Prospects. Annals of the Entomological Society of America 2021, 114, 470–476. [Google Scholar] [CrossRef]
- Campos, M.; Crepeau, M.; Lee, Y.; Gripkey, H.; Rompão, H.; Cornel, A.J.; Pinto, J.; Lanzaro, G.C. Complete Mitoge-nome Sequence of Anopheles Coustani from São Tomé Island. Mitochondrial DNA Part B 2020, 5, 3376–3378. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.-H.; He, S.-L.; Fu, W.-B.; Yan, Z.-T.; Hu, Y.-J.; Yuan, H.; Wang, M.-B.; Chen, B. Mitogenome-Based Phyloge-ny of Mosquitoes (Diptera: Culicidae). Insect Sci 2024, 31, 599–612. [Google Scholar] [CrossRef]
- Chen, D.-H.; He, S.-L.; Fu, W.-B.; Yan, Z.-T.; Hu, Y.-J.; Yuan, H.; Wang, M.-B.; Chen, B. Mitogenome-Based Phyloge-ny of Mosquitoes (Diptera: Culicidae). Insect Science 2024, 31, 599–612. [Google Scholar] [CrossRef]
- Bohmann, K.; Mirarab, S.; Bafna, V.; Gilbert, M.T.P. Beyond DNA Barcoding: The Unrealized Potential of Genome Skim Data in Sample Identification. Molecular Ecology 2020, 29, 2521–2534. [Google Scholar] [CrossRef]
- Guo, J.; Yan, Z.-T.; Fu, W.-B.; Yuan, H.; Li, X.-D.; Chen, B. Complete Mitogenomes of Anopheles peditaeniatus and Anopheles nitidus and Phylogenetic Relationships within the Genus Anopheles Inferred from Mitogenomes. Parasites & Vectors 2021, 14, 452. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Villegas, L.; Assis-Geraldo, J.; Koerich, L.B.; Collier, T.C.; Lee, Y.; Main, B.J.; Rodrigues, N.B.; Orfano, A.S.; Pires, A.C.A.M.; Campolina, T.B.; et al. Characterization of the Complete Mitogenome of Anopheles aquasalis, and Phylo-genetic Divergences among Anopheles from Diverse Geographic Zones. PLOS ONE 2019, 14, e0219523. [Google Scholar] [CrossRef] [PubMed]
- Ali, R.; Gebhardt, M.E.; Lupiya, J.S.; Muleba, M.; Norris, D.E. The First Complete Mitochondrional Genome of Anopheles Gibbinsi Using a Skimming Sequencing Approach. F1000Res 2024, 13, 553. [Google Scholar] [CrossRef] [PubMed]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De Novo Assembly of Organelle Genomes from Whole Ge-nome Data. Nucleic Acids Res 2017, 45, e18. [Google Scholar]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de Novo Metazoan Mitochondrial Genome Annotation. Mol Phylogenet Evol 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: Phylogenetic Model Averaging. Molecular Biology and Evolution 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.-H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLOS Computational Biology 2014, 10, e1003537. [Google Scholar] [CrossRef]
- Krzywinski, J.; Grushko, O.G.; Besansky, N.J. Analysis of the Complete Mitochondrial DNA from Anopheles funestus: An Improved Dipteran Mitochondrial Genome Annotation and a Temporal Dimension of Mosquito Evolution. Molecular Phylogenetics and Evolution 2006, 39, 417–423. [Google Scholar] [CrossRef]
- Bouafou, L.; Makanga, B.K.; Rahola, N.; Boddé, M.; Ngangué, M.F.; Daron, J.; Berger, A.; Mouillaud, T.; Makunin, A.; Korlević, P.; et al. Host Preference Patterns in Domestic and Wild Settings: Insights into Anopheles Feeding Behavior. Evol Appl 2024, 17, e13693. [Google Scholar] [CrossRef]
- Gebhardt, M.E.; Searle, K.M.; Kobayashi, T.; Shields, T.M.; Hamapumbu, H.; Simubali, L.; Mudenda, T.; Thuma, P.E.; Stevenson, J.C.; Moss, W.J.; et al. Understudied Anophelines Contribute to Malaria Transmission in a Low-Transmission Setting in the Choma District, Southern Province, Zambia. Am J Trop Med Hyg 2022, 106, 1406–1413. [Google Scholar] [CrossRef]
- Finney, M.; McKenzie, B.A.; Rabaovola, B.; Sutcliffe, A.; Dotson, E.; Zohdy, S. Widespread Zoophagy and Detection of Plasmodium Spp. in Anopheles Mosquitoes in Southeastern Madagascar. Malaria Journal 2021, 20, 25. [Google Scholar] [CrossRef] [PubMed]
- Aschale, Y.; Getachew, A.; Yewhalaw, D.; De Cristofaro, A.; Sciarretta, A.; Atenafu, G. Systematic Review of Sporo-zoite Infection Rate of Anopheles Mosquitoes in Ethiopia, 2001–2021. Parasites & Vectors 2023, 16, 437. [Google Scholar]
- Hoffman, J.E.; Ciubotariu, I.I.; Simubali, L.; Mudenda, T.; Moss, W.J.; Carpi, G.; Norris, D.E.; Stevenson, J.C.; on be-half of Southern and Central Africa International Centers of Excellence for Malaria Research Phylogenetic Complexity of Morphologically Identified Anopheles squamosus in Southern Zambia. Insects 2021, 12, 146. [CrossRef]
- Nguyen, V.T.; Dryden, D.S.; Broder, B.A.; Tadimari, A.; Tanachaiwiwat, P.; Mathias, D.K.; Thongsripong, P.; Reeves, L.E.; Ali, R.L.M.N.; Gebhardt, M.E.; et al. A Comprehensive Review: Biology of Anopheles squamosus, an Understudied Malaria Vector in Africa. Insects 2025, 16, 110. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Bayesian tree showing the phylogenetic relationship of 10 new mitochondrial genomes (highlighted in blue) of understudied Anopheles mosquito species with other available anophe-line sequences. The tree includes assigned accession numbers and was constructed using BEAST v2.7.6. The posterior probabilities supporting the tree topology are represented by the values at the nodes.
Figure 1.
Bayesian tree showing the phylogenetic relationship of 10 new mitochondrial genomes (highlighted in blue) of understudied Anopheles mosquito species with other available anophe-line sequences. The tree includes assigned accession numbers and was constructed using BEAST v2.7.6. The posterior probabilities supporting the tree topology are represented by the values at the nodes.
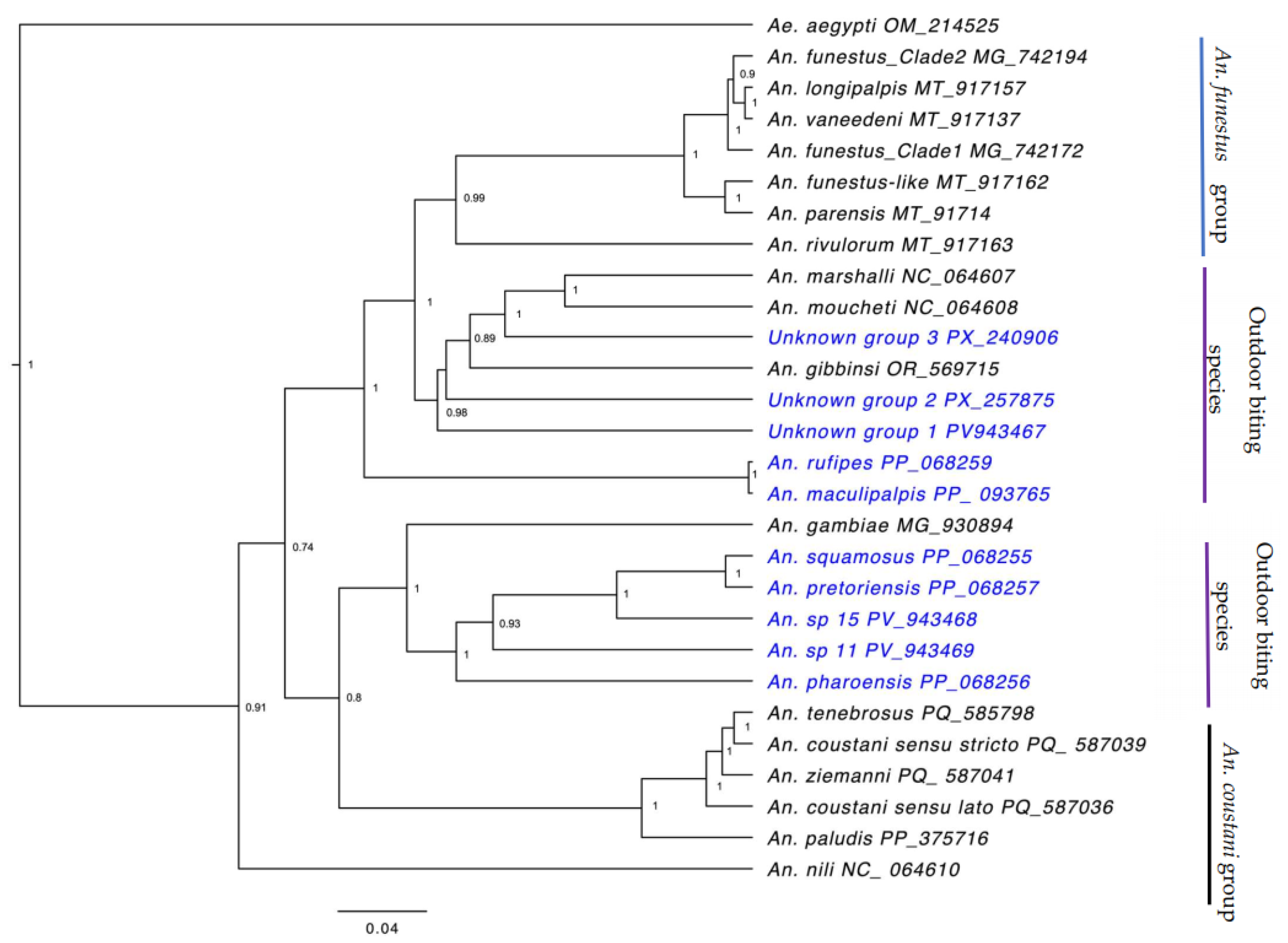
Figure 2.
Phylogenetic tree showing inferred molecular divergence estimates (MYA) for outdoor biting Anopheles using the concatenated PCGs from mitogenomes generated in this study. The mean divergence time (MYA) predicted for each event is represented by the values at the tree nodes. The bars show the values at tree nodes, 95% confidence intervals.
Figure 2.
Phylogenetic tree showing inferred molecular divergence estimates (MYA) for outdoor biting Anopheles using the concatenated PCGs from mitogenomes generated in this study. The mean divergence time (MYA) predicted for each event is represented by the values at the tree nodes. The bars show the values at tree nodes, 95% confidence intervals.
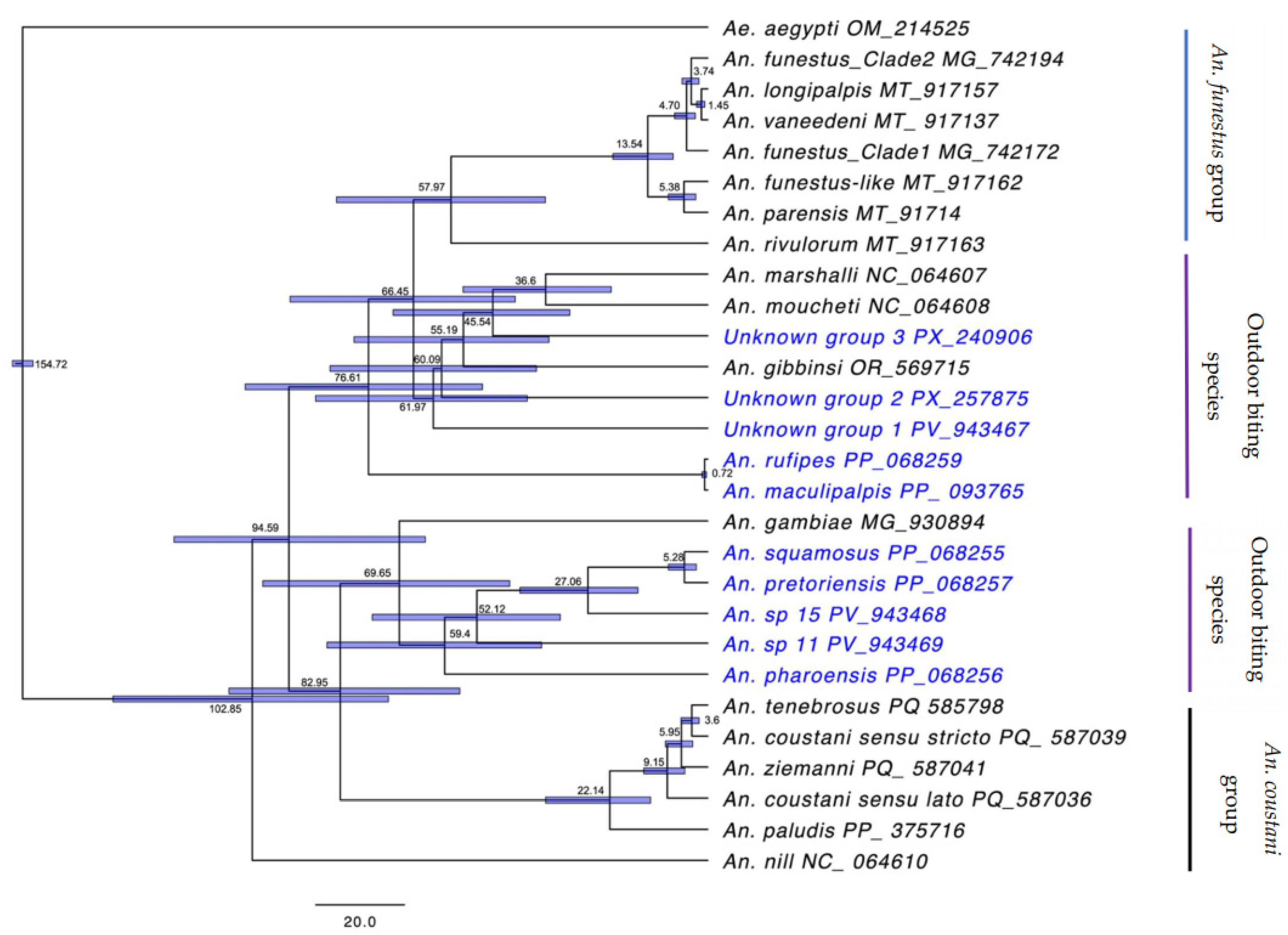

Table 1.
Genome characteristics for mitochondrial genomes of 8 understudied anopheline mos-quito species generated in this study.
Table 1.
Genome characteristics for mitochondrial genomes of 8 understudied anopheline mos-quito species generated in this study.
| Identification | Contig Size | GC % | AT % | GenBank Accession # |
| Morphological | ||||
| An. pretoriensis | 15348 | 23.0 | 77.0 | PP_068257 |
| An. pharoensis | 15346 | 23.7 | 76.3 | PP_068256 |
| An. rufipes | 15362 | 22.9 | 77.1 | PP_068259 |
| An. squamosus | 15349 | 23.1 | 76.9 | PP_068255 |
| An. maculipalpis | 15361 | 23.4 | 76.6 | PP_093765 |
| Molecular | ||||
| Species 11 | 15350 | 23.1 | 76.9 | PV_943469 |
| Species 15 | 15354 | 22.8 | 77.2 | PV_943468 |
| Unknown group 1 | 15398 | 21.1 | 78.9 | PV_943467 |
| Unknown group 2 | 15534 | 23.1 | 76.9 | PX_257875 |
| Unknown group 3 | 15436 | 20.3 | 79.7 | PX_240906 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



