
Submitted:
23 September 2025
Posted:
24 September 2025
You are already at the latest version
Abstract
Globally, the growing pressure on elasmobranchs presents a significant threat. The Z-13 Fisher Colony, located in Rio de Janeiro’s metropolitan region, is composed of an urban-artisanal fishing fleet operating in a macro-area adjacent to the “Hope Spot - MoNa Cagarras” in “Rio de Janeiro Shelf ISRA”. Locally, elasmobranch fishing has been carried out for over a century. Artisanal fishery poses a potential threat to the populations of this taxon, with more than 1/3 listed globally in IUCN Red List categories. As an attempt to inform fisheries management, a Productivity Susceptibility Risk Analysis (PSA) was applied, as a locally unprecedented approach, to assess the Vulnerability of 31 landed elasmobranch species by the mentioned urban-artisanal fisheries. Analyses indicated that most small to medium-sized elasmobranch species (n = 25; 80.6%) had medium to high Productivity; while 11 (35.5%) species showed high Susceptibility to artisanal fishing. Five species showed higher Vulnerability, and were comprised by 03 sharks (S. lewini, S. zygaena, and I. oxyrinchus) and 02 rays (A. castelnaui and P. horkelii). Seven species showed medium Vulnerability being 05 sharks (S. occulta, R. porosus, C. falciformes, C. obscurus, and S. argentina) and 02 rays (D. hypostigma and P. percellens). The other 19 species (61.3%) presented lower Vulnerability and were comprised by 08 (25.8%) species of sharks and 11 (35.5%) of rays. The species identified as having high Vulnerability or potential risk should be immediately prioritized for fisheries management and administration by the Brazilian responsible agencies, to protect the potential of the taxon for ecosystem functions and prevent further extinctions.
Keywords:
Hope Spot - MoNa Cagarras
; rays
; sharks
; small-scale fishery
; Southwestern Atlantic
; stock assessment
; vulnerability
1. Introduction
According to Nelson et al. (2016), the subclass Elasmobranchii comprises a group of well-adapted and wide-ranging cartilaginous fish: Selachii (sharks) and Batomorphi (rays and relatives; batoids). Most elasmobranchs, being cosmopolitan vertebrates, are predators that occupy higher trophic levels, exercising a crucial ecological key role in maintaining cascading effects along food webs and balance in marine ecosystems (Toussaint et al., 2021), preserving the ecological equilibrium and maintaining healthy ecosystems. However, this taxon holds economic importance as a haliotic resource and provides financial benefits to fishing communities and industries worldwide (FAO, 2022). Over-exploitation is considered one of the main threats to elasmobranch populations and has led to a significant decline in the global abundance (~71.0%) of their populations over the past few decades (IUCN, 2024). The taxon is known to be highly vulnerable to overfishing due to its k-strategist life history traits (e.g., low intrinsic rates of population growth and recruitment; Dulvy et al., 2017). The most recent evaluation of the global conservation status of elasmobranchs conducted by the International Union for Conservation of Nature revealed that the proportion of threatened species with extinction increased from 17.4% to 37.5% in less than ten years (IUCN, 2024).
Sharks and rays are found along the entire coast of Brazil (Santos et al., 2024), with approximately 203 marine elasmobranch species recorded to date (Kotas et al., 2023). The state of Rio de Janeiro, located in the Southeast of Brazil, is one of the priority areas for the global conservation of elasmobranchs (Lucifora et al., 2011), with approximately 636 km of coastline (23º 00' 00'' to 21º 15' 00'' S and 45º 00' 00'' to 41º 00' 00'' W), which marks the change in orientation and geo-oceanographic conformation of the Brazilian coast from north-south to east-west (Cumplido, pers. obs.). It is home to incredible biodiversity and sustains essential fishing grounds for Brazilian sardines, penaeid shrimp, and demersal fish (MPA, 2014). According to MPA (2014), extractive marine fisheries in Rio de Janeiro account for about 2.5% of the total landings of sharks and rays captured per year in Brazil, and the state ranks fourth in economic importance and third in population according to the national ranking (IBGE, 2019), which endorses its importance in the Brazilian socio-economic context. In Rio de Janeiro, gillnets are a significant component of artisanal fishing (FIPERJ, 2020) and target various population strata, competing for the same species as an industrial fleet (Begot and Vianna, 2014; PMAP-RJ, 2020). This type of fishing has low operating costs and involves various forms of use, especially in shallower and estuarine regions of the continental shelf. Although sharks and rays may not be the official target species, these multispecies opportunistic coastal fisheries generate a significant bycatch of elasmobranchs.
Based on this overview, the Brazilian government implemented regulations prohibiting the capture of threatened species classified under any of the following categories: Vulnerable (VU); Endangered (EN); or Critically Endangered (CR) on the National Red List (ICMBio, 2018a,b). These protections are outlined in a series of Normative Instructions and Ordinances (e.g., IN 5 MMA 21/05/2004; IN 52 MMA 08/05/2005; IN 5 MPA/MMA 15/04/2011; MMA no 445 17/12/2014; MMA no 148 07/06/2022; GM/MMA no 300 14/12/2022; MMA no 354 27/01/2023), covering several species present in this study. Additionally, the National Plan of Action for the Conservation of Threatened Marine Sharks and Rays (“PAN-Tubarões”) offers further conservation guidelines (ICMBio, 2023; Kotas et al., 2023).
Opportunistic and multispecies fishing are important factors influencing the spatial-temporal and co-occurrence of catches (PMAP-RJ, 2020). Size selectivity is an essential aspect of gillnet fishing and the main target of management actions (Shester and Micheli, 2011). Although its underreported dynamics make it an unprofitable activity when compared to industrial fisheries, gillnet fishing is essential for local actors, who often find themselves in situations of socioeconomic vulnerability and strive to use all possible bycatch (Vasconcellos et al., 2011).
Stock assessment plays a crucial role in understanding the health and sustainability of fish populations and fisheries, optimizing resource utilization. Fish stocks with little biological and fishing information (e.g., elasmobranch populations) are challenging to assess by using traditional quantitative analyses. Fishing stocks with deficient data can be evaluated through alternative risk assessment methods (Stobutzki et al., 2002; Cortes et al., 2015). The Productivity and Susceptibility Analysis (PSA) is a semi-quantitative approach that can be used to evaluate the vulnerability of a stock to becoming overfished (e.g., bycatch and/or targeted stocks), based on its biological productivity (e.g., life-history patterns theory: the evolution of individual characteristics of organisms and your biological attributes (see Thorson et al., 2017; Pauly and Cheung, 2018) and susceptibility to the fishery (Cortes et al., 2010). This approach, ideal for data-limited fisheries or populations, is a valuable tool to identify vulnerable species to fishing exploitation and their risk levels, essentially ranks the relative vulnerability of a fishery based principally on the spatial overlap between a stock and its fishery, to define research priorities and to suggest management measures scenarios (Hobday et al., 2011). The input attributes are assigned categorical qualitative values on the same scale and then combined as either standard or geometric averages depending on the approach (Hordyk and Carruthers, 2018).
Over the last 20-odd years, the PSA has been progressively applied to evaluate the vulnerability or potential risk of exploitation of elasmobranch populations, based on their biological productivity and fishing susceptibility (Gallagher et al., 2012; McCully et al., 2013; Carreon-Zapiain et al., 2020; Bravo-Zavala et al., 2022; Tavares et al., 2024). The gap of information on target and non-target species (e.g., bycatch) in commercial fisheries regarding catch resilience increases the vulnerability of species and can lead to their collapse or extinction (Le Grix et al., 2023). As a result, the conservation of taxon has become increasingly pressing in recent years.
Nowadays, on the coast of Rio de Janeiro, southeastern Brazil, there is uncertainty regarding the statuses of elasmobranch populations due to the gap of monitoring by the responsible agencies. Furthermore, the socioeconomic and political situations in the country hinder fisheries management and research. The goal of this study was to estimate the vulnerability of the elasmobranch species caught by urban-artisanal fisheries through bycatch, focusing on gillnet landings, calculating productivity and susceptibility using a semi-quantitative PSA. In addition to taxonomic composition, insights into fishing selectivity can provide a guide for management efforts, which is particularly helpful for assessing multispecies fisheries with data-poor. Therefore, prioritizing species for effective management measures is crucial to improving conservation strategies to help mitigate bycatch and protect vulnerable shark and ray populations. The enforcement of existing legislation for threatened species is fundamental to a sustainable and effective fishing chain. This type of species-specific study is crucial, as it not only highlights the economic importance of haliotic resources but also supports the real sustainability of artisanal fishing.
2. Materials and Methods
2.1. Study Area
The present study took place at the Z-13 Fisher Colony, a fish market located on Copacabana Beach, in Rio de Janeiro, Southeastern Brazil. Founded on June 29, 1923, Z-13 has already brought together approximately 1,000 fishers, who are spread from Urca Beach to Pontal da Praia do Recreio dos Bandeirantes. Today, the headquarters has 50 to 60 member fishermen. These professionals primarily work in the coastal waters of Rio de Janeiro, with the insular systems of Cagarras Islands serving as a key fishing area and, locally, encompassing approximately 20 km2 of water's edge where the urban-artisanal fishery of the colony takes place and carries out the fishing activity daily. Part of these islands forms the Natural Monument of the Archipelago of the Cagarras Islands and Surrounding Waters (MoNa Cagarras), an important no-take Marine Protected Area (MPA), classified as a full-protection Conservation Unit (CU) where fishing is restricted, established in 2010, and located 05 km offshore (ICMBio, 2010). According to Machado et al. (2022), the MoNa Cagarras, together with its surrounding areas (including Rasa and Cotunduba islands), was designated as a Hope Spot by the international nonprofit organization Mission Blue and officially launched on April 16, 2021 as “Hope Spot - MoNa Cagarras”, and is recognized as one of the world´s most important sites for marine conservation and recovery, providing a refuge for abundant biodiversity and serving as an essential habitat for several rare, threatened, and endemic species along the coast of Brazil (Figure 1).
This monitored fishing macro-area is part of one that we have studied in detail and recently was recognized as an Important Shark and Ray Area (ISRA) (see Hyde et al., 2022), with an area of 549.2 km2 and a depth interval of 0-50 meters (ISRA, 2025). The study area is influenced by oceanographic features such as coastal orientation, seasonally wind-driven upwelling, and the limited width of the continental shelf, which allow for the proximity of warm tropical water masses (e.g., average of 23-25 ºC at summer/February) and cold waters (e.g., average of 21-23 ºC at winter/July) from the South Atlantic Waters Current (SAWC) (Carbonel and Galeão, 2007), with the temperature varying throughout the water column (e.g., thermoclines). During the summer there is a greater presence of upwelling (Calado et al., 2010), the outcrop or rise of nutrient-rich deep cold waters from the South Atlantic Central Water (SACW), whose assumed thermal threshold is 18 ºC, and during the winter it is less intense or even non-existent (Valentin, 1994), boosting regional primary production (Valentin and Coutinho, 1990). Coastal input from the associated estuary, accompanied by a massive influx of organic matter (Carreira et al., 2012), increases benthic biomass. Due to these combined conditions, primary production reaches up to 1.0 mg/m³, while the surrounding waters maintain the oligotrophic levels of 0.2 mg/m³ (Carreira et al., 2012). Continental output from the connected hypereutrophic estuary also provides organic matter that boosts benthic biomass, with up to 16.8 mg/l of suspended organic matter and 0.42 mg/l of particulate matter within the proposed area (Martins et al., 2016), values higher than those in the surrounding regions. The mixing of water masses, combined with a mosaic of soft and hard bottoms and access to a nearby mangrove, makes the region remarkably diverse, with 282 fish species (Machado et al., 2022). The elevated biomass supports a productive artisanal fishing community concentrated between the coastal island chains that border the area, up to a depth of -50 m, and the shoreline (Loto et al., 2018). Fisheries have been practiced in the area since the native populations established a presence (Lopes et al., 2016), with archaeological evidence of the presence of the same shark species still in the region today. Fishing remains a vital tradition for small-scale fishing communities; however, elasmobranchs face serious threats not only from population declines due to overfishing (Wosnick et al., 2024) but also from the heavily polluted waters of Guanabara Bay (Neves et al., 2023).
2.2. Data Source
A total of 31 elasmobranch species were recorded, which were landed in the Z-13 Fisher Colony (see Table S1.). Monitoring was conducted from June 1, 2021, to July 31, 2023, with a sampling period of two complete seasonal cycles, continuously from Monday to Friday, from 08:00 to 14:00 hours (h), when fishing landings are concentrated. The monitoring of fish landings was restricted to the colony, characterized by the local fishing profile as a fishing warehouse composed of an urban-artisanal fishing fleet with daily activity, mainly using gillnets of various sizes, threads, and meshes, capturing, via bycatch, diversified species of elasmobranchs that utilize the area for reproduction, birth, feeding, and growth. The primary fishing gear used by local fishers is the gillnet, a passive fishing method that targets fish living on the bottom, mid-water, and sub-surface. The gillnets remain in the water for approximately 24 to 48 h. Fishers at the Z-13 Fisher Colony go out daily with pre-established crews along the shallow inner continental shelf (< 20 km offshore). Fishers employ opportunistic strategies, which involves adjusting gear, strands and/or mesh sizes in response to seasonal variation in fishery resource availability.
2.3. Fishery Data and Biological Information
The primary data was obtained in loco, by a single researcher, the first author of this study, in order to minimize sampling bias, through informal semi-structured interviews with fishermen and other stakeholders in the fishing production chain, using a form designed to collect the following biological and fishing information: numerical identification of the form; date (e.g., for fishing seasonality); vessel and captain or person in charge; crew; fishing area; fisheries ground(s); fishing gear and configuration/setup; fishing effort; the approximate bathymetry, in meters, of the gillnet sets and the type of substrate found; period that the nets remained in the water; number of harvests; time spent fishing; landed species and the species-specific composition (n and kilograms, in kg); biometrics and percentage of importance per group (%) according to the total of specimens/species landed; value and destination of production; personal observations; and, when possible, discards. The local urban-artisanal fishing fleet was briefly characterized based on its modi operandi, including the types of materials used, gear configurations, and methods of operation (Table 1).
During the recording of landed elasmobranchs, the specimens were identified to the lowest possible taxonomic level, macroscopically sexed, and measured for the Total Length (TL, cm) according to Compagno (1984). In cases where multiple individuals of a given species were landed, biometrics were performed by selecting the largest, intermediate, and smallest specimens, with the total number of individuals in each size class recorded, always maintaining a standard sample size. Sexual maturation stages were determined through a combination of internal, during the gutting process for cleaning and preparation for marketing, and external characters according to Yokota and Lessa (2006) and Andrade et al. (2008), based on the length at maturity data.
For the biological parameters of the species, the productive attributes commonly used in PSA (see methodological section related to Productivity and Susceptibility Analysis) were considered. Data on life-history traits were obtained from published literature and an additional compilation from "grey literature" for each species. Only studies conducted in the natural environment (in situ) were selected for inclusion. For that purpose, the following sites as leading search platforms were used: Google Scholar; CAPES Journal Portal; SciELO; and Redalyc. The key terms applied were: life-history traits > population dynamics > age and growth > reproductive biology/cycle > mean fecundity > trophic level and MTL >> Southeastern Brazil > Southwest Atlantic Ocean > subtropical waters of the Southwestern Atlantic. The preference was given to studies carried out in the Southeastern Brazilian Bight (SBB), which is a crescent-shaped ‘semi-enclosed’ continental shelf of the Santos Basin (between 23º-28º S and 42º-48º W) that sustains important fishing grounds (Gasalla et al., 2010). The information gaps were complemented with data reported from adjacent areas or other marine regions. For some species, this parameter has not been determined, and in these cases, a value for the genus or conspecific species was used instead (for sources, see Supplementary Materials - S1) (Table 2).
For susceptibility attributes commonly used in PSA (see methodological section related to Productivity and Susceptibility Analysis), the primary data from the fishery (e.g., empirical from the Local-traditional Ecological Knowledge - LtEK) were acquired in the field during in loco fisheries monitoring. The proportion of immature individuals caught in the fishery was also incorporated into the PSA. Current conservation statuses were obtained from the IUCN Red List (IUCN, 2025).
2.4. Productivity and Susceptibility Analysis (PSA)
The PSA was performed to assess the vulnerability of the 31 species of elasmobranchs captured and landed by artisanal fishery in the study area (Supplementary Materials - S1). The Vulnerability (V) of a stock is defined by two dimensions: Productivity (P), which reflects the species’ capacity to recover after depletion, and Susceptibility (S), which represents the likelihood of capture and mortality resulting from the fishery (Stobutzki et al., 2001).
To calculate Productivity (P), eight attributes proposed by Patrick et al. (2010) were used: maximum age (Tmax, years); maximum size (Lmax, cm); von Bertalanffy’s growth coefficient K (cm year-1); esteemed natural mortality (M, year-1); Length at maturity (L50, cm); Reproductive Cycle (RC, periodicity); Fecundity (Fec., mid-point of the number of oocytes or embryos); and Trophic level (Tlevel, no unit) (Table 3).
For Susceptibility (S), a total of ten attributes originally proposed by Patrick et al. (2010) were considered: vertical overlap (VO); areal overlap (or Encounterability - E); morphology affecting capture (or Selectivity - S); fishing rate in relation to M; food items; seasonal migrations; schooling/aggregation/geographic concentration and behavioral responses; fishery impact; valued or desired of the fishery; and management strategy. Additionally, two attributes proposed by Tavares et al. (2024) were included in the analysis: proportion of immatures caught about the total catches sampled, indicating the general low selectivity of the fishing gear (e.g., bycatch); and the IUCN statuses, evidencing high contradiction between the precarious current commercial fishing monitoring and legislation and, unfortunately, the negative and recurrent local-regional catch of critically endangered elasmobranch species via bycatch (or even intentional capture). Susceptibility (S) attributes were primarily scored based on the authors’ local knowledge, and secondarily on information from online databases (Froese and Pauly, 2023; IUCN, 2024; Table 4).
The P and S attributes were scored using a three-point scale: 1 = low, 2 = medium, and 3 = high (Hobday et al., 2006). Species with low P and high S scores are considered at high risk of overfishing, whereas species with high P and low S scores are considered at low risk of overfishing. We used a default weight of 2 (ranging from 4 for high relevance to 1 for low relevance), as a precautionary value, for the productivity and susceptibility attributes (Duffy and Griffiths, 2019). Additionally, because data-poor stocks can lead to errors in risk, a data quality index was developed to estimate the uncertainty associated with individual vulnerability scores. This index is based on five tiers, where 1 represents the highest data quality and 5 the lowest (Patrick et al., 2010). We prioritized the information by regions, as follows: the coast of the state of Rio de Janeiro (study area) and the Southeastern Brazilian Bight (or Basin) (SBB) > Southeastern Brazil > Southern Brazil > Northeast Brazil > Northern Brazil > other regions of the Atlantic Ocean > Global.
After the range (r) and weight (w) values of each attribute (P and S) were defined for each species, the average values of P and S were estimated using the following equations:
where, r and w are the range and weight of Productivity and Susceptibility, respectively. Subsequently, the overall Vulnerability score (V) of a stock was calculated as the Euclidean distance from the origin of the x-y scatter plot (Patrick et al., 2010) by applying the following formula:
Vulnerability scores were categorized into three levels based on Cope et al. (2011): High (V ≥ 2.0); Medium (1.8 ≤ V < 2.0); and Low (V < 1.8). Stocks with low Productivity and high Susceptibility scores are considered most vulnerable to overfishing, while those with high Productivity and low Susceptibility scores are considered least vulnerable (Patrick et al., 2010). Even with strong fishing pressure, these species continue to generate catches due to their high productivity rates (Patrick et al., 2009; Cope et al., 2011). The advantage of the PSA scatter plots is that managers can easily use them to apply or adjust management measures to suitable levels based on the stock’s level of vulnerability (Cortes et al., 2015). These analyses will identify the species most impacted by fishing activities (e.g., percentage of gear overlap) and highlight those that should be prioritized for targeted fisheries management measures (e.g., specific fisheries and stocks). Data organization and calculations for this analysis were carried out by constructing PSA matrices using MS Excel (Microsoft® Office Excel 2021).
3. Results
3.1. Fishery Data and Species Composition
During the study period, in general at the Z-13 Fisher Colony, throughout the year, “Corvineira” type gillnets presented the highest results regarding totals elasmobranch caught and landed (n = 147; 58.3%), followed by “Linguadeiras” (n = 111; 33.3%) and “Come-dormes” (n = 38; 8.4%), the type of gear least used (Table 1). These fishing gear indicate which ones are more efficient and, consequently, less selective and more predatory regarding elasmobranch captures. Elasmobranchs were incidentally captured in teleost fisheries across all fishing grounds. Unfortunately, due to overlapping fishing areas, gear and interspecific habitats, this also results in the capture of sharks and rays (e.g., non-target stocks), especially species with similar coastal habits to the target teleosts (e.g., scienids and carangids).
A total of 31 elasmobranch species, belonging to 16 families, eight shark and eight ray families, comprising 16 species of sharks (51.6%) and 15 of rays (48.4%), were recorded in the urban-artisanal fishery conducted in the coastal waters of Rio de Janeiro, Southeastern Brazil (Supplementary Materials - S1). A total of 296 specimens (60 sharks (20.3%) and 236 rays (79.7%), and approximately 1.3 ton was counted, while the total-final production of teleosts and relatives was about 46.0 tons for the same period, which represented merely 2.7% of the total biomass landed locally. Among sharks, the most frequently caught species by percentage of numbers were Rhizoprionodon porosus (23.2%), R. lalandii (20.0%), Sphyrna lewini (18.3%) and S. zygaena (10.0%). Other relatively important shark species in the catches were Carcharhinus brevipinna (5.0%), C. falciformis (3.3%), Squatina guggenheim (3.3%), and C. obscurus (3.3%). Regarding rays, the most frequently observed species were Pseudobatos horkelii (55.7%), Dasyatis hypostigma (25.8%), Atlantoraja castelnaui (10.2%). Additionally, P. percellens (3.0%) and Hypanus guttatus (1.3%) were also relatively common in the study area. Regarding Total Length (TL, cm), both smaller (≥ 35 cm) and larger (≥ 66 cm) specimens were relatively common throughout the entire sampling period and present in all fishing grounds. When smaller, elasmobranchs were consumed by the fishers themselves and their families, while larger ones were traded separately, although at varied prices (in USD (1.00 USD:5.00 R$ per kg) - range: 2.00-5.00; average: 3.00-3.50 and ±SD: 2.00-5.00 (minimum of 2.00 and maximum of 5.00).
According to the National Red List, of the 31 species recorded in the study area, 81.3% of shark species and 86.7% of ray species are threatened with extinction, falling into one of the following categories: Vulnerable (VU), Endangered (EN) and Critically Endangered (CR). Similarly, the global IUCN Red List indicates that the shark and ray species found in the study area are at risk, with 93.6% and 100.0% listed as threatened, respectively (Supplementary Materials - S1). This finding is particularly concerning due to the gap of solid fishing regulations in Brazil and their implementation.
Reinforcing the importance of this study, oceanic or epipelagic species such as Galeocerdo cuvier, S. zygaena, C. brevipinna, or species associated with deeper waters and/or around islands, such as S. guggenheim, A. castelnaui, were also landed by the Z-13 Fisher Colony fisheries, mainly smaller size classes and/or juveniles, with some larger animals and subadults-adults. The landed elasmobranch species, most of which are classified as endangered, were captured at different life stages when associated with shallower waters, ranging from large pregnant females (or breeding matrices) (115 ≤ TL ≤ 100, cm) that kill not only the mother, but also the embryos, to small neonates and juveniles (35 ≥ TL ≤ 66, cm), with a predominance of adults. This is supported by the record of three pregnant P. horkelli females (115 ≤ TL ≤ 100, cm), a more coastal species, one containing oocytes, a postpartum S. lewini female (TL = 110 cm), a pregnant Hypanus berthalutzae female (TL = 47 cm), two pregnant A. castelnaui females (80 ≤ TL ≤ 110, cm) and a postpartum S. zygaena female (TL = 139 cm). Assuming that the landing of adult specimens reflects local catches, we can assume that overfishing of juveniles is not an imminent risk regarding the exploited elasmobranch populations at the study site. However, the presence of juveniles (n = 113; 38.2%) indicates that the area where the urban-artisanal fleet operates is a possible development area (used for growth and maturation) for the landed elasmobranch species.
Sharks such as Rhizoprionodon spp. and Sphyrna spp., and rays, as P. horkelii, D. hypostigma, and A. castelnaui were intensely caught and landed, regardless of seasonality, in general, with higher numbers of captures during the winter, the most critical period that requires greatest significant immediate attention concerning sustainable fisheries management.
3.2. Productivity and Susceptibility Analysis (PSA)
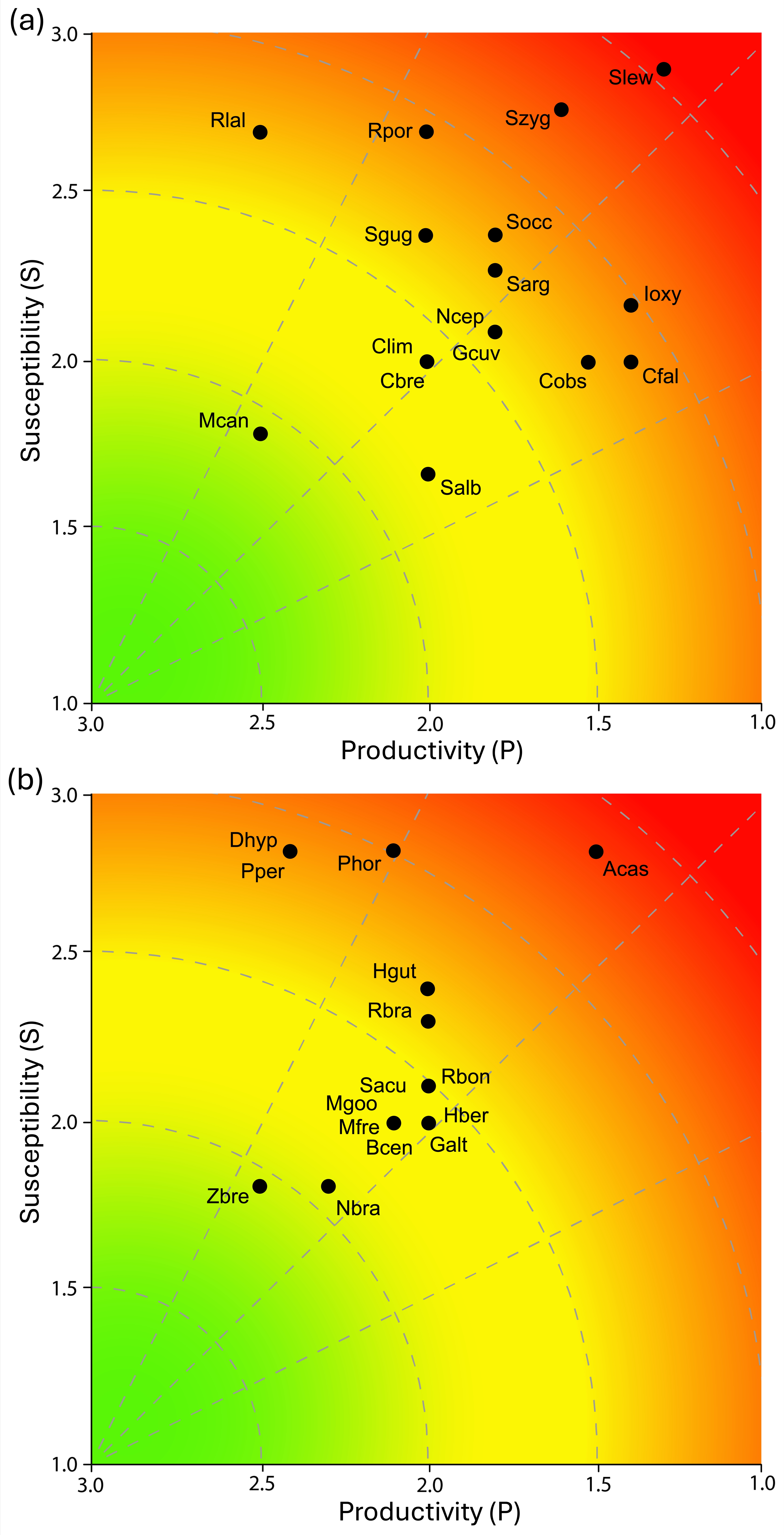
Regarding productivity attributes, of the 31 elasmobranch species evaluated, 05 (16.1%; P = 2.4-3.0) were classified as having high productivity, 20 (64.5%; P = 1.7-2.3) as moderate productivity, and 06 (19.4%; P = 1.0-1.6) as low productivity (Table 5). The 06 species with low productivity were the sharks S. lewini, Isurus oxyrinchus, C. falciformis, C. obscurus, S. zygaena, and the ray A. castelnaui. These elasmobranchs are commonly characterized by low growth rates, limited fecundity and long periods of gestation, which makes them particularly vulnerable to overfishing. Although most shark and ray species exhibited moderate (64.5%) and low (19.4%) productivity, the majority are classified as threatened on the IUCN Red List, indicating signs of potential overexploitation over time (Table 5).
Elasmobranchs were captured incidentally in teleost fisheries throughout the year and in all fishing grounds due to the relatively low selectivity of the gillnets used by the Z-13 Fisher Colony. For susceptibility attributes (Table 5), the assigned scores showed that 11 species (35.5%; S = 2.4-3.0) had high susceptibility values. These included the sharks S. lewini, S. zygaena; R. porosus, R. lalandii, Squatina occulta, and S. guggenheim, as well as the rays A. castelnaui, P. horkelii, D. hypostigma, P. percellens, and H. guttatus. All of these species are categorized as threatened on the IUCN Red List. Furthermore, several of these species are also heavily exploited in the southeastern coast of Brazil, including Pseudobatos spp., S. lewini, R. porosus, R. lalandii, D. hypostigma, and A. castelnaui. The remaining species (n = 20; 19.4%; S = 1.7-2.3) were classified as having moderate susceptibility, and no species was classified as low susceptibility (Table 5).
The assessment of the vulnerability of the 31 species showed values ranged from 1.0 to 2.3 (Table 5). Most species (n = 19; 61.3%) had low vulnerability (Figure 2); however, all of these species except one are categorized as threatened on the IUCN Red List. This group comprises relevant commercial species such as the sharks R. lalandii, C. brevipinna, and S. guggenheim, as well as the ray H. guttatus. The second group, with moderate vulnerability, consisted of 07 elasmobranch species (22.6%) that presented varying degrees of threat (Figure 2). The sharks S. argentina and S. occulta are Critically Endangered (CR), while the 02 ray species, D. hypostigma and P. percellens, are listed as Vulnerable (VU) on the IUCN Red List. The third group, corresponding to high vulnerability elasmobranchs, included 05 species (16.1%): the sharks I. oxyrinchus, S. zygaena, and S. lewini, and the rays P. horkelii and A. castelnaui (Figure 2). All of these species, also categorized as threatened on the IUCN Red List, represent considerable importance in elasmobranch catches and require special attention in conservation strategies. Within this group, S. lewini, P. horkelii, and A. castelnaui are classified as Critically Endangered (CR) and should be prioritized under national and international conservation measures.
4. Discussion
The study of the vulnerability of elasmobranch species is fundamental when considering sustainable fishery management. Overfishing puts pressure on marine ecosystems (FAO, 2021). Regarding global fisheries production, the current trend is not towards stability, but rather, declines (Pauly and Zeller, 2016), with at least 1/3 of global fisheries being overexploited (Duarte et al., 2020).
Artisanal fishing is generally multispecific and multifaceted (Batista et al., 2014). This creates potential spatial overlaps (Komoroske and Lewison, 2015). The role of artisanal fisheries in the exploitation of fishery resources is associated with concentrated efforts and high fish production in coastal areas, with this fisheries modality accounting for > 50.0% of global production (FAO, 2020). In Brazil, artisanal fisheries are practiced along the entire coast (Zamboni and Martin-Dias, 2020). Estimates suggest that 40-60+% of marine fisheries production in Brazil originates from artisanal fishing (Freire et al., 2021).
In this sense, coastal fisheries and bycatches due to gillnets comprise relevant elasmobranch population decline sources, as 99.6% of all elasmobranch species are currently categorized as under threat (Dulvy et al., 2021), encompassing higher catch rates than those achieved by industrial fleets (Peckham et al., 2008). Concerning elasmobranchs in particular, most populations in Brazil are depleted (Bornatowski et al., 2018).
This is an emblematic study of the vulnerability of elasmobranch species landed by a resilient urban-artisanal fishing fleet in a critical metropolitan region of Southeastern Brazil, in Rio de Janeiro State. Based on secondary biological data, we demonstrate a wide variation in the life-history traits parameters of 31 species caught in gillnets with various threads and meshes sizes. The broad taxonomic richness of elasmobranchs reflects the multiplicity of environments explored by fishers, such as unconsolidated (e.g., sand and mud) and consolidated (e.g., slabs, plots, rocky shores and artificial areas, such as the “Ipanema Submarine Outfall”) substrate bottoms, and the employed gillnets, whose height is sufficient to capture species presenting both epi-pelagic and demersal habits, which include sharks and rays (as bycatch), and the incidental catch compositions are highly specific to each type of fishery (e.g., bottom, mid-water, or (sub)surface). Although the diversity of elasmobranchs varied across geographic areas, we highlight that 1/3 of these species are endemic to the Southeastern Atlantic Ocean and are classified as at high risk of extinction or “Data Deficient” (ICMBio, 2025). Our PSA approach is consistent with the methods of extinction risk assessment used in conservation biology (Davies and Baum, 2012; IUCN, 2022).
It is crucial to ascertain the presence of keystone species in fishing locations. Keystone species are defined as: species with an extremely high impact on a given ecosystem relative to their population, critical to the overall structure and function of a certain ecosystem and influencing which other types of organisms that make up that ecosystem (Humphries et al., 2017). Six keystone elasmobranch species (G. cuvier, C. obscurus, C. taurus, S. lewini, S. zygaena and Zapteryx brevirostris) are observed along the southeastern coast of Brazil (Bornatowski et al., 2014). The present study, in a regionally isolated manner, recorded 05 of them, namely G. cuvier (also see Aximoff et al., 2022); C. obscurus, S. lewini, S. zygaena and Z. brevirostris, that are at the most significant potential risk from overfishing and require prioritization for continued fisheries monitoring and comprehensive assessment actions on the southeastern coast of Brazil.
The fishing area is situated in a region of intense artisanal fishing activity, with over 20,000 fishing days recorded, with gillnets accounting for nearly half of this effort (FIPERJ, 2020). Artisanal gillnets also account for the most significant number of productive units (n = 1,307) but represent just over 5.0% of the total biomass of fish landed (94,472.4 tons in the state of Rio de Janeiro between 2018 and 2019) (FIPERJ, 2020). Data from the “Rio de Janeiro state Fishery Monitoring Program” (PMAP-RJ) indicate that in 2019, artisanal gillnets were responsible for approximately 89 tons of elasmobranch production, accounting for a yield of USD4,500 (USD2.00 per kg, average at first sale value). However, despite the efforts of the “Rio de Janeiro Fishing Institute Foundation” (FIPERJ) to monitor 186 landing points scattered throughout the region, the production landed at Z-13 Fisher Colony remains unmonitored in official state statistics.
In general, knowledge of coastal elasmobranch species catches is practically non-existent (Bornatowski et al., 2018). Since 2011, consolidated national statistics on fish production are, however, unavailable in the country (Freire et al., 2021). According to FIPERJ (2015 and 2016), the artisanal production of elasmobranchs is not available for several fishing areas, such as the metropolitan region of Rio de Janeiro. Previous efforts have recorded significant elasmobranch fisheries in adjacent areas of the state of Rio de Janeiro (Silva-Junior, 2008). However, there are uncertainties due to catches frequently being illegal, underreported, or discarded as bycatch at sea during fishing activities (Freire et al., 2015). In addition, fisheries statistics also face taxonomic inaccuracy, with many species being grouped into a single fisheries management category during monitored landings (e.g., sharks, rays, angelfish, and guitarfish). The opposite is also true, with a single species receiving several reference names. It hinders the accuracy of production estimates and prevents effective fisheries management. For example, in the state of Rio de Janeiro, the “Cação” category groups 36 reference names, relating to 33 species of sharks. Meanwhile, the “Raia or Emplastro” category groups 25 reference names for 19 species of rays (FIPERJ, 2020). This considerable taxonomic loss is compounded by the fact that a significant portion of landings consists solely of carcasses, making it impossible to identify the species. Limited data for many species is one of the biggest challenges to compiling estimates of threats to elasmobranchs (Jacquet and Pauly, 2008).
Historically, the obsolescence of the management model, high fishing effort, and low resilience to biomass removal have contributed to the vulnerability of elasmobranchs. In recent decades, some species that occur in continental shelf and slope waters have been depleted by industrial fishing on the south-southeast Brazilian coast (Vooren and Klippel, 2005; Barreto et al., 2016; Oddone et al., 2019; Rigby et al., 2019; Pollom et al., 2020a,b). Despite knowledge about the biology, distribution, and conservation status of these species, historical catch series still gap details at the species level (Schroeder et al., 2024). Despite international recommendations and national legislation prohibiting the capture, landing, and trade of many species, landings of endangered species continue to occur freely in Rio de Janeiro and South-southeastern Brazil (Silva-Junior et al., 2008; Barreto et al., 2017).
Contrary to global efforts to protect elasmobranchs, Brazil ranks 11th globally as a producer and first as an importer of “shark” meat (Cruz et al., 2021). Many tropical developing countries, such as Brazil, have coastal communities that rely on sharks and rays as a significant source of protein for subsistence (FAO, 2020). However, due to it being considered “lower-quality meat”, the illegal practice of intentionally mislabeling is often used to attract consumers (Bornatowski et al., 2015). In general, people are usually unable to distinguish between species, and most do not associate the shark meat sold as “cação” with sharks and rays (Bornatowski et al., 2013). Although “cação” meat is not part of the traditional Brazilian diet, there is great interest in encouraging people to eat “cação” meat and keeping the Brazilian consumer market a perfect port for discarded meat (Bornatowski et al., 2018).
The 02 most landed species of small coastal sharks, of the genus Rhizoprionodon (R. porosus and R. lalandii), which occupy an intermediate position in the water column (Compagno, 2001), are frequently captured during fishing activities throughout the entire Brazilian coast and are significantly consumed by humans, especially in the southeastern region, where it is abundant (Andrade et al., 2008; Barreto et al., 2016). However, there is still a gap of population data and insufficient knowledge to assess fishing pressures or the effects of habitat degradation (Barreto et al., 2016). Consequently, it hinders effective management and conservation actions. They are classified as “Data Deficient” by a National Red List (ICMBio, 2023; Kotas et al., 2023). These sharks captures in Rio de Janeiro, SE Brazil, are significantly abundant, with an estimated total of 35.9 tons landed by gillnet fishing between 2019 and 2021 (PMAP-RJ, 2022). Their diet, which is primarily based on sciaenid fish and decapod crustaceans, as well as clupeid fish (neonates and adults, respectively), explains the dominance as bycatch of the main target species in southeastern fisheries (Bornatowski et al., 2012).
The southeastern portion of the Brazilian coast also represents an ecologically important area for other Carcharhiniformes, such as S. lewini, S. zygaena, C. brevipinna and C. limbatus, as also noted by Andrade et al. (2008), Silva-Junior et al. (2008) and Marques et al. (2019), in a similar fishing community located in the Recreio dos Bandeirantes neighborhood, Rio de Janeiro, and locally (Machado et al., 2022). Adult females and juveniles represented a significant portion of the local capture, indicating the significance of the study region. In addition to species commonly caught with bottom, mid-water or (sub)surface gillnets, cosmopolitan, predominantly oceanic and with epipelagic habits, such as I. oxyrinchus and the G. cuvier, were also caught, all juveniles, reinforcing the potential of the area for growth and maturation. This area may be included in a large coastal nursery which, in turn, may be subdivided into reproduction, parturition, recruitment, development (growth and maturation) areas. The capture of juveniles in Brazil, as well as the capture of large pregnant females, comprising breeding matrices with greater reproductive potential and fecundity, and adult individuals, suggest a potential impact on the recruitment and renewal rates of these populations, thus compromising the regional gene pool of the taxon (Lutz et al., 2024).
The significant ray composition captured by artisanal fishing in the state of Rio de Janeiro, as well as that of sharks is diverse and originates mainly from gillnets fisheries. The high vulnerability of demersal-coastal species (e.g., Pseudobatos spp.; Sphyrna spp.; and Squatina spp.) increases as their capture ceases to be incidental and becomes targeted, due to high first-sale values. Some elasmobranchs, such as the rays Pseudobatos spp., D. hypostigma and A. castelnaui and the sharks Rhizoprionodon spp. and Sphyrna spp. display a first-sale value equivalent to that of medium to large-sized Sciaenidae, the target fish group of the Z-13 Fisher Colony. Several threatened elasmobranchs occur in the same habitats as commercial species whose capture is permitted or regulated, such as Micropogonias furnieri, Paralichthys spp., Centropomus spp., Cynoscion spp., leading to their bycatch. As the fishing gear used to capture demersal species exhibits low selectivity, non-target species, like rays and/or Sphyrna spp. and Squatina spp. are occasionally and incidentally captured. The pressure suffered by these species, combined with the environmental vulnerability of these resources, can culminate in an unsustainable situation.
In terms of biological productivity attributes, elasmobranchs are characterized by being more susceptible to overfishing than teleosts (Walker, 1998; Myers and Worm, 2005; Cortes et al., 2015). The PSA approach used in this study demonstrated a range of sensitivity among species and highlighted those that are most sensitive. Life-history traits characteristics that influence population recovery (e.g., fecundity, reproductive cycle, growth rate) have often been used to assess stocks with limited data (King and McFarlane, 2003; Claireaux et al., 2018). For example, species with high P values included small-to-medium-sized sharks with rapid growth and annual reproductive cycles (i.e., Rhizoprinodon spp. and Mustelus canis) and rays (D. hypostigma and Pseudobatos spp.). The lowest P values were recorded among large, slow-growing, long-lived, and late-maturing oceanic species (e.g., Sphyrna spp., I. oxyrinchus, Carcharhinus spp., and A. castelnaui). These results are comparable to values found for other regions of the Atlantic and Pacific (Visintin and Perez, 2016; Furlong-Estrada et al., 2017; Mejia-Falla et al., 2019; Carreon-Zapian et al., 2020; Bravo-Zavala et al., 2022).
In this study, elasmobranchs showed high susceptibility to gillnet fishing. According to Vizintin and Perez (2016), this was particularly true among benthic predators with a flattened body shape and large pectoral fins (e.g., Squatina spp.; Sphyrna spp.; A. castelnaui; D. hypostigma; Pseudobatos spp.). These groups are often caught in highly productive coastal regions (Andrade et al., 2008; Barreto et al., 2016; Santos et al., 2022). Pregnant females, neonates, and juveniles registered in the Z-13 Fisher Colony fisheries landing confirm the importance of these shallow and protected waters. The number of elasmobranch species landed at Z-13 Fisher Colony comprises around 60.0% of sharks and rays reported in the fishing production of the state of Rio de Janeiro (FIPERJ, 2020). Multispecies coastal fisheries, such as those practiced by the miscellaneous gillnet of Z-13 Fisher Colony, generate a recurrently and relevant elasmobranch bycatch despite the fishing effort primarily targeting coastal teleosts as mentioned above. The opportunistic strategy of this urban-artisanal fishing fleet has contributed to the increasing harvesting of these catches. In general, elasmobranchs accounted for 13.0% and 22.0% of the bycatch species that landed in Rio de Janeiro state (Souza et al., 2025). The opportunistic strategy of this urban-artisanal fishing fleet has contributed to the increasing harvesting of these catches, which were initially treated as bycatch. The increased interest in large rays (e.g., D. hypostigma; Atlantoraja spp.; Hypanus spp.) has also been reported for artisanal fisheries in other traditional communities in southeastern Brazil (Santos et al., 2022). A similar historical evolution of landings has been recorded for this taxon in the South Atlantic (Martins and Schwingel, 2003; Silva-Junior et al., 2008; Perez et al., 2009).
The PSA analysis revealed that elasmobranch species with high vulnerability values exhibit low to moderate productivity. The high values recorded for oceanic top predator species (e.g., Sphyrna spp. and I. oxyrinchus) suggest that different population strata may be affected by intense and diverse fishing efforts during their life cycle. These key species influence both community structure and fishery productivity (Tavares et al., 2024). In addition, the group of rays (e.g., P. horkelli and A. castelnaui) is also highly vulnerable to various types of trawling (Vianna and Almeida, 2005), which may generate a cumulative risk of overfishing. The overlap of fisheries in the same fishing area, sharing the same resources, is recognized as a potential cause of stock depletion (McGoodwin, 2002). The PSA ranking system provides a helpful tool for rapidly identifying groups of interest for either research emphasis or management attention (Cope et al., 2011). In general, management influences the susceptibility of the stocks, while productivity attributes are usually static in the short term and are most affected by the quality of input data. Therefore, different levels of vulnerability can be verified by projecting different management action scenarios.
Regarding species vulnerability, reduced catches or total bans may be considered for the Pseudobatos spp. (CR, IUCN), most often caught by “Linguadeiras”, as post-release mortalities are lower for large specimens because they exhibit greater biological resilience. Regarding hammerheads, all six species are currently listed as prohibited for capture and sale (ICMBio, 2018a; IUCN, 2025). The present study noted two of these species (S. lewini and S. zygaena), indicating the need for immediate actions. For hammerheads and other large Carcharhinids, it may be appropriate to control age class captures to protect breeding stocks. Squatina occulta and S. guggenheim, mostly caught by “Linguadeiras”, are classified, respectively, as Critically Endangered and Endangered, by the IUCN and require attention. Population studies of these animals should be prioritized, considering their ecological and economic importance and new categorizations, such as “Near Threatened” according to the IUCN Red List.
Despite the limitations of the present study, we verify equivalent degree of vulnerability of elasmobranchs caught throughout Brazil and borders (Freire et al., 2015; Visintin and Perez, 2016; Freire et al., 2021; Souza et al., 2025), and at adjacent areas in the Atlantic Ocean, such as: Venezuelan Caribbean (Tavares et al., 2024); Colombian Caribbean (Mejia-Falla et al., 2019); and in the Gulf of Mexico (Bravo-Zavala et al., 2022; Pérez-Jiménez and Mendoza-Carranza, 2023). This confirms the high vulnerability to which the taxon is subject in the face of excessive increases in global fishing pressure. With this PSA study, as a locally unprecedented approach, we highlight a starting point for future research and data collection needs.
5. Conclusions
Given the fragility of the model due to its subjectivity in quali-quantitative analysis, it is clear that PSA should not replace formal quantitative stock assessments. However, for the case of data-poor fisheries, this approach appears to be a useful tool for identifying which species face higher risk based on their biological characteristics and their population interaction with the fishing pressure. The present study provided information regarding the catch elasmobranch composition and the level of vulnerability of the most frequently caught species from the Rio de Janeiro Coast. Considering the long periods of fishing pressure experienced by elasmobranchs with poor management strategy, there is an urgent need to formulate and implement new and appropriate fishing regulations. Among the fishing regulations, we recommend the revision of the fishing gear, assigning fishing quotas, and seasonal fishing bans. These measures should be considered, not only for the species with high vulnerability, but also for those with medium and lower levels of risk. Particular attention must be given to endangered and prohibited species captured (e.g., Sphyrna spp.; I. oxyrinchus; Squatina spp.; Carcharhinus spp.; A. castelnaui; and Pseudobatos spp.) or those not currently included in existing fishing regulations (e.g., Rhizoprionodon spp. and D. hypostigma), but considered globally at risk of extinction and/or highly threatened. This taxon should be prioritised for immediate management strategies by the Brazilian agencies responsible for fisheries administration, and it is urgent to endorse the importance of continuous fisheries monitoring programs. Finally, this type of species-specific study is crucial, as it not only highlights the economic importance of haliotic resources but also supports the real sustainability of artisanal fishing.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Table S1.: The 31 elasmobranch species landed by the commercial urban-artisanal gillnets fisheries at the Z-13 Fisher Colony, Copacabana Beach, Rio de Janeiro, RJ, Southeastern Brazil, from June 2021 to July 2023.; Supplementary dataset. A list of 31 elasmobranch species life-history traits references (Table 2).; and Supplementary Information (SI).
Author Contributions
Conceptualization, Rodrigo Cumplido, Rafael Tavares, Pablo Mendonça and Luís Gustavo Cardoso; Methodology, Rodrigo Cumplido, Rafael Tavares, Pablo Mendonça and Luís Gustavo Cardoso; Software, Rodrigo Cumplido, Rafael Tavares and Pablo Mendonça; Validation, Rodrigo Cumplido, Rafael Tavares and Pablo Mendonça; Formal analysis, Rodrigo Cumplido, Rafael Tavares and Pablo Mendonça; Investigation, Rodrigo Cumplido, Rafael Tavares and Pablo Mendonça; Resources, Rodrigo Cumplido; Data curation, Rodrigo Cumplido, Rafael Tavares, Pablo Mendonça and Luís Gustavo Cardoso; Writing – original draft, Rodrigo Cumplido, Rafael Tavares and Pablo Mendonça; Writing – review & editing, Rodrigo Cumplido, Rafael Tavares, Pablo Mendonça and Luís Gustavo Cardoso; Visualization, Rodrigo Cumplido, Rafael Tavares, Pablo Mendonça and Luís Gustavo Cardoso; Supervision, Rodrigo Cumplido; Project administration, Rodrigo Cumplido; Funding acquisition, Rodrigo Cumplido.
Acknowledgments
Special thanks are due to José Manoel P. Rebouças and his son, fisher Manasí da S. Rebouças, as well as to the interviewed fishers and other actors in the Z-13 Fisher Colony production chain for granting access to the landed elasmobranchs, kindly providing the requested data and assisting with biometric sampling. To the Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), and PPG-OCN/UERJ. We also thank Bárbara Piovani Luz Aieta Afonso for the map plotting. To the BioTecPesca’s Lab. team for their help with the bibliographic survey on the life-history traits of the elasmobranch species analyzed. To Prof. Marcelo Vianna, for his guidance, remote support, and suggestions for the manuscript; and the reviewers for their support during the review phase.
References
- Andrade, A.C.; Silva-Junior, L.C.; Vianna, M. Reproductive biology and population variables of the Brazilian sharpnose shark Rhizoprionodon lalandii (Müller & Henle, 1839) captured in coastal waters of south-eastern Brazil. J. Fish Biol. 2008, 72, 473–484. [Google Scholar] [CrossRef]
- Aximoff, I.; Cumplido, R.; Rodrigues, M.T.; de Melo, U.G.; Netto, E.B.F.; Santos, S.R.; Hauser-Davis, R.A. New Occurrences of the Tiger Shark (Galeocerdo cuvier) (Carcharhinidae) off the Coast of Rio de Janeiro, Southeastern Brazil: Seasonality Indications. Animals 2022, 12, 2774. [Google Scholar] [CrossRef]
- Barreto, R.; Ferretti, F.; Flemming, J.M.; Amorim, A.; Andrade, H.; Worm, B.; Lessa, R. Trends in the exploitation of South Atlantic shark populations. Conserv. Biol. 2016, 30, 792–804. [Google Scholar] [CrossRef]
- Barreto, R.; Bornatowski, H.; Motta, F.; Santander-Neto, J.; Vianna, G.; Lessa, R. Rethinking use and trade of pelagic sharks from Brazil. Mar. Policy 2017, 85, 114–122. [Google Scholar] [CrossRef]
- Batista, V.S.; Fabré, N.N.; Malhado, A.C.M.; Ladle, R.J. Tropical Artisanal Coastal Fisheries: Challenges and Future Directions. Rev. Fish. Sci. Aquac. 2014, 22, 1–15. [Google Scholar] [CrossRef]
- Begot, L.H.; Vianna, M. Frota pesqueira costeira do Estado do Rio de Janeiro. Bol. Inst. de Pesca 2014, 40, 79–94. [Google Scholar]
- Bornatowski, H.; Heithaus, M.R.; Abilhoa, V.; Corrêa, M.F.M. Feeding of the Brazilian sharpnose shark Rhizoprionodon lalandii (Müller & Henle, 1839) from southern Brazil. J. Appl. Ichthyol. 2012, 28, 623–627. [Google Scholar] [CrossRef]
- Bornatowski, H.; Braga, R.R.; Vitule, J.R.S. Shark Mislabeling Threatens Biodiversity. Science 2013, 340, 923–923. [Google Scholar] [CrossRef] [PubMed]
- Bornatowski, H.; Navia, A.F.; Braga, R.R.; Abilhoa, V.; Corrêa, M.F.M. Ecological importance of sharks and rays in a structural foodweb analysis in southern Brazil. ICES J. Mar. Sci. 2014, 71, 1586–1592. [Google Scholar] [CrossRef]
- Bornatowski, H.; Braga, R.R.; Kalinowski, C.; Vitule, J.R.S. “Buying a Pig in a Poke”: The Problem of Elasmobranch Meat Consumption in Southern Brazil. Ethnobiol. Lett. 2015, 6, 196–202. [Google Scholar] [CrossRef]
- Bornatowski, H.; Braga, R.R.; Barreto, R.P. Elasmobranch Consumption in Brazil: Impacts and 4 Consequences - Chapter 10. In Advances in Marine Vertebrate Research in Latin America: Technological Innovation and Conservation; Rossi-Santos, M.R., Finkl, C.W., Eds.; New York, United States of America: Springer International Publishing, Coastal Research Library: 2018; pp. 251–262. [Google Scholar] [CrossRef]
- Brasil., 2022a. Ministério do Meio Ambiente. Portaria MMA nº 148, de 07 de junho de 2022. Altera os Anexos da Portaria nº 443, de 17 de dezembro de 2014, da Portaria nº 444, de 17 de dezembro de 2014, e da Portaria nº 445, de 17 de dezembro de 2014, referentes à atualização da Lista Nacional de Espécies Ameaçadas de Extinção. Diário Oficial da União: seção 1, Brasília, DF, ano 160, n. 108, p. 74, 08 jun. Available online: https://www.in.gov.br/web/dou/-/portaria-mma-n-148-de-7-de-junho-de-2022-406272733 (accessed on 29 July 2025).
- Brasil., 2022b. Ministério do Meio Ambiente. Portaria GM/MMA nº 300, de 13 de dezembro de 2022. Revoga os Atos da Portaria nº 443, de 17 de dezembro de 2014, da Portaria nº 444, de 17 de dezembro de 2014, da Portaria nº 445, de 17 de dezembro de 2014, da Instrução Normativa nº 1, de 12 de fevereiro de 2015, da Portaria nº 98, de 28 de abril de 2015, da Portaria nº 162, de 08 de junho de 2015, da Portaria nº 163, de 08 de junho de 2015, da Portaria nº 395, de 1º de setembro de 2016, da Portaria nº 161, de 20 de abril de 2017, da Portaria nº 201, de 31 de maio de 2017, da Portaria nº 217, de 19 de junho de 2017, da Portaria nº 73, de 26 de março de 2018, da Portaria nº 148, de 7 de junho de 2022 e da Portaria nº 229, de 5 de setembro de 2022, referentes à atualização da Lista Nacional de Espécies Ameaçadas de Extinção. Diário Oficial da União: seção 1, Brasília, DF, ano 160, n. 234, p. 75-118, 14 dez. 2022. ISSN 1677-7042. Available online: https://www.in.gov.br/en/web/dou/-/portaria-gm/mma-n-300-de-13-de-dezembro-de-2022-450425464 (accessed on 29 July 2025).
- Brasil., 2023. Ministério do Meio Ambiente. Portaria nº 354, de 27 de janeiro de 2023. Revoga os Atos da Portaria MMA nº 299, de 13 de dezembro de 2022, e nº 300, de 13 de dezembro de 2022. Repristinados os seguintes atos do MMA da Portaria nº 443, de 17 de dezembro de 2014; Portaria nº 444, de 17 de dezembro de 2014; Portaria nº 445, de 17 de dezembro de 2014; Instruçao Normativa nº 1, de 12 de fevereiro de 2015; Portaria nº 98, de 28 de abril de 2015; Portaria nº 162, de 08 de junho de 2015; Portaria nº 163, de 08 de junho de 2015; Portaria nº 395, de 1º de setembro de 2016; Portaria nº 161, de 20 de abril de 2017; Portaria nº 201, de 31 de maio de 2017; Portaria nº 217, de 19 de junho de 2017; Portaria nº 73, de 26 de março de 2018; Portaria nº 148, de 7 de junho de 2022; Portaria nº 229, de 5 de setembro de 2022; Portaria nº 43, de 31 de janeiro de 2014; Portaria nº 162, de 11 de maio de 2016; e Portaria nº 444, de 26 de novembro de 2018, com inclusão de espécies na Portaria nº 148, de 7 de junho de 2022, da Lista Nacional de Espécies Ameaçadas de Extinçao. Diário Oficial da União: seção 1, Brasília, DF, ano 160, n. 21, p. 72-73, 27 jan. 2023. ISSN 1677-7042. Available online: https://www.in.gov.br/en/web/dou/-/portaria-mma-n-354-de-27-de-janeiro-de-2023-460770327 (accessed on 29 July 2025)ISSN 1677-7042.
- Bravo-Zavala, F.G.; Pérez-Jiménez, J.C.; Tovar-Ávila, J.; Arce-Ibarra, A.M. Vulnerability of 14 elasmobranchs to various fisheries in the southern Gulf of Mexico. Mar. Freshw. Res. 2022, 73, 1064–1082. [Google Scholar] [CrossRef]
- Calado, L.; Da Silveira, I.C.A.; Gangopadhyay, A.; De Castro, B.M. Eddy-induced upwelling off Cape São Tomé (22ºS, Brazil). Continental Shelf Research 2010, 30, 1181–1188. [Google Scholar] [CrossRef]
- Carbonel, C.A.A.H.; Galeão, A.C.N. A stabilized finite element model for the hydrothermodynamical simulation of the Rio de Janeiro coastal ocean. Commun. Numer. Methods Eng. 2007, 23, 521–534. [Google Scholar] [CrossRef]
- Carreón-Zapiain, M.T.; Tavares, R.; Favela-Lara, S.; Oñate-González, E.C. Ecological Risk Assessment with integrated genetic data for three commercially important shark species in the Mexican Pacific. Reg. Stud. Mar. Sci. 2020, 39, 101431. [Google Scholar] [CrossRef]
- Carreira, R.d.S.; Canuel, E.A.; Macko, S.A.; Lopes, M.B.; Luz, L.G.; Jasmim, L.N. On the accumulation of organic matter on the southeastern Brazilian continental shelf: a case study based on a sediment core from the shelf off Rio de Janeiro. Braz. J. Oceanogr. 2012, 60, 75–87. [Google Scholar] [CrossRef]
- Claireaux, M.; Jørgensen, C.; Enberg, K. Evolutionary effects of fishing gear on foraging behavior and life-history traits. Ecol. Evol. 2018, 8, 10711–10721. [Google Scholar] [CrossRef] [PubMed]
- Compagno, L.J.V. Checklist of living elasmobranchs. Pp. 471-498. In Sharks, Skates, and Rays - The Biology of Elasmobranch Fishes. 1a Ed.; Hamllet, W.C., Ed.; Baltimore, United States of America: John Hopkins University Press, 1984. [Google Scholar]
- Compagno, L.J.V. 2001. Sharks of the world - An annotated and illustrated catalogue of shark species known to date. Volume 2. Bullhead, Mackerel and Carpet sharks (Heterodontiformes, Lamniformes and Orectolobiformes). FAO, Species Catalogue for Fishery Purposes., 2001. [CrossRef]
- Cope, J.M.; DeVore, J.; Dick, E.J.; Ames, K.; Budrick, J.; Erickson, D.L.; Grebel, J.; Hanshew, G.; Jones, R.; Mattes, L.; et al. An Approach to Defining Stock Complexes for U.S. West Coast Groundfishes Using Vulnerabilities and Ecological Distributions. North Am. J. Fish. Manag. 2011, 31, 589–604. [Google Scholar] [CrossRef]
- Cortés, E.; Arocha, F.; Beerkircher, L.; Carvalho, F.; Domingo, A.; Heupel, M.; Holtzhausen, H.; Santos, M.N.; Ribera, M.; Simpfendorfer, C. Ecological risk assessment of pelagic sharks caught in Atlantic pelagic longline fisheries. Aquat. Living Resour. 2009, 23, 25–34. [Google Scholar] [CrossRef]
- Cortés, E.; Brooks, E.N.; Shertzer, K.W. Risk assessment of cartilaginous fish populations. ICES J. Mar. Sci. 2014, 72, 1057–1068. [Google Scholar] [CrossRef]
- Cruz, M.M.; Szynwelski, B.E.; de Freitas, T.R.O. Biodiversity on sale: The shark meat market threatens elasmobranchs in Brazil. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 3437–3450. [Google Scholar] [CrossRef]
- Davies, T.D.; Baum, J.K. Extinction Risk and Overfishing: Reconciling Conservation and Fisheries Perspectives on the Status of Marine Fishes. Sci. Rep. 2012, 2, 561. [Google Scholar] [CrossRef]
- Duarte, C.M.; Agusti, S.; Barbier, E.; Britten, G.L.; Castilla, J.C.; Gattuso, J.-P.; Fulweiler, R.W.; Hughes, T.P.; Knowlton, N.; Lovelock, C.E.; et al. Rebuilding marine life. Nature 2020, 580, 39–51. [Google Scholar] [CrossRef]
- Duffy, L.M.; Griffiths, S.P. Assessing attribute redundancy in the application of productivity-susceptibility analysis to data-limited fisheries. Aquat. Living Resour. 2019, 32, 20. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Simpfendorfer, C.A.; Davidson, L.N.; Fordham, S.V.; Bräutigam, A.; Sant, G.; Welch, D.J. Challenges and Priorities in Shark and Ray Conservation. Curr. Biol. 2017, 27, R565–R572. [Google Scholar] [CrossRef] [PubMed]
- Dulvy, N.K.; Pacoureau, N.; Rigby, C.L.; Pollom, R.A.; Jabado, R.W.; Ebert, D.A.; Finucci, B.; Pollock, C.M.; Cheok, J.; Derrick, D.H.; et al. Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Curr. Biol. 2021, 31, 4773–4787. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture: Sustainability in action; FAO: Rome, Italy, 2020. [Google Scholar]
- FAO. Fisheries operations: Guidelines to prevent and reduce bycatch of marine mammals in capture fisheries; FAO: Rome, Italy, 2021. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture: Sustainability in action; FAO: Rome, Italy, 2022. [Google Scholar]
- FIPERJ. Fundação Instituto de Pesca do Estado do Rio de Janeiro., 2015. Available online: http://www.fiperj.rj.gov.br/fiperj_imagens/arquivos/revistarelatorios2015.pdf (accessed on 15 January 2025).
- FIPERJ. Fundação Instituto de Pesca do Estado do Rio de Janeiro., 2016. Relatório 2016. Available online: http://www.fiperj.rj.gov.br/fiperj_imagens/arquivos/revistarelatorios2015.pdf (accessed on 15 January 2025).
- FIPERJ. Fundação Instituto de Pesca do Estado do Rio de Janeiro., 2020. Estatística Pesqueira do Estado do Rio de Janeiro. PMAP-RJ, Projeto de Monitoramento da Atividade Pesqueira no Estado do Rio de Janeiro (2018-2019). In: Projeto de Monitoramento da Atividade Pesqueira no Estado do Rio de Janeiro., 2020. Relatório Técnico Consolidado Final-RTF. 1. Available online: http://www.fiperj.rj.gov.br/index.php/publicacao/index/1 (accessed on 15 January 2025).
- Freire, K.M.F.; Aragão, J.A.N.; Araújo, A.R.R.; et al. Reconstruction of catch statistics for Brazilian marine waters (1950-2010): fisheries catch reconstructions for Brazil's mainland and oceanic islands. Fish. Centre Res. Rep. 2015, 23, 3–30. [Google Scholar]
- Freire, K.M.F.; Almeida, Z.d.S.d.; Amador, J.R.E.T.; Aragão, J.A.; Araújo, A.R.d.R.; Ávila-Da-Silva, A.O.; Bentes, B.; Carneiro, M.H.; Chiquieri, J.; Fernandes, C.A.F.; et al. Reconstruction of Marine Commercial Landings for the Brazilian Industrial and Artisanal Fisheries From 1950 to 2015. Front. Mar. Sci. 2021, 8. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. FishBase: world wide web electronic publication. Available online: https://www.fishbase.org (accessed on 10 July 2025).
- Furlong-Estrada, E.; Galván-Magaña, F.; Tovar-Ávila, J. Use of the productivity and susceptibility analysis and a rapid management-risk assessment to evaluate the vulnerability of sharks caught off the west coast of Baja California Sur, Mexico. Fish. Res. 2017, 194, 197–208. [Google Scholar] [CrossRef]
- Gallagher, A.J.; Kyne, P.M.; Hammerschlag, N. Ecological risk assessment and its application to elasmobranch conservation and management. J. Fish Biol. 2012, 80, 1727–1748. [Google Scholar] [CrossRef] [PubMed]
- Gasalla, M.A.; Rodrigues, A.R.; Duarte, L.F.; Sumaila, U.R. A comparative multi-fleet analysis of socio-economic indicators for fishery management in SE Brazil. Prog. Oceanogr. 2010, 87, 304–319. [Google Scholar] [CrossRef]
- Hobday, A.; Smith, A.; Stobutzki, I.; Bulman, C.; Daley, R.; Dambacher, J.; Deng, R.; Dowdney, J.; Fuller, M.; Furlani, D.; et al. Ecological risk assessment for the effects of fishing. Fish. Res. 2011, 108, 372–384. [Google Scholar] [CrossRef]
- Hobday, A.J.; Smith, A.; Webb, H.; Daley, R.; Wayte, S.; Bulman, C.; Dowdney, J.; Williams, A.; Sporcic, M.; Dambacher, J.; Fuller, M.; Walker, T. Ecological Risk Assessment for the Effects of Fishing: Methodology. Report R04/1072 for the Australian Fisheries Management Authority, Canberra. 2006. [Google Scholar]
- Hordyk, A.R.; Carruthers, T.R. A quantitative evaluation of a qualitative risk assessment framework: Examining the assumptions and predictions of the Productivity Susceptibility Analysis (PSA). PLOS ONE 2018, 13, e0198298. [Google Scholar] [CrossRef]
- Humphries, M.M.; Studd, E.K.; Menzies, A.K.; Boutin, S. To Everything There Is a Season: Summer-to-Winter Food Webs and the Functional Traits of Keystone Species. Integr. Comp. Biol. 2017, 57, 961–976. [Google Scholar] [CrossRef]
- Hyde, C.A.; di Sciara, G.N.; Sorrentino, L.; Boyd, C.; Finucci, B.; Fowler, S.L.; Kyne, P.M.; Leurs, G.; Simpfendorfer, C.A.; Tetley, M.J.; et al. Putting sharks on the map: A global standard for improving shark area-based conservation. Front. Mar. Sci. 2022, 9, 968853. [Google Scholar] [CrossRef]
- IBGE. Instituto Brasileiro de Geografia e Estatística., 2019. Available online: www.ibge.gov.br (accessed on 20 October 2024).
- ICMBIO. Instituto Chico Mendes de Conservação da Biodiversidade., 2010. MoNa do Arquipélago das Ilhas Cagarras. Available online: https://www.icmbio.gov.br/portal/mona-do-arquipelago-das-ilhas-cagarras (accessed on 27 October 2024).
- ICMBio. Instituto Chico Mendes de Conservação da Biodiversidade., 2018a. Livro Vermelho da Fauna Brasileira Ameaçada de Extinção: Volume VI - Peixes. ICMBio, Brasília - DF, Brazil, p. 1.235.
- ICMBio. Instituto Chico Mendes de Conservação da Biodiversidade., 2018b. Livro Vermelho da Fauna Brasileira Ameaçada de Extinção: Volume I. ICMBio / MMA, Brasília - DF, Brazil.
- ICMBio. Instituto Chico Mendes de Conservação da Biodiversidade., 2023. Plano de Ação Nacional para a Conservação dos Tubarões e Raias Marinhos Ameaçados de Extinção. Available online: https://pt.scribd.com/document/682264893/Livro-Pan-Tubaroes-2023-Vfinal-23-Digital-Compacto-Compressed-1 (accessed on 25 July 2025).
- ICMBio. Instituto Chico Mendes de Conservação da Biodiversidade., 2025. Sistema de Avaliação do Risco de Extinção da Biodiversidade - SALVE. Available online: https://salve.icmbio.gov.br/ (accessed on 31 August 2025).
- ISRA. Important Shark and Ray Area., 2025. IUCN SSC Shark Specialist Group. Rio de Janeiro Shelf ISRA Factsheet. Dubai: IUCN SSC Shark Specialist Group., 2025. Available online: https://sharkrayareas.org/factsheets/rio-de-janeiro-shelf-isra/ (accessed on 11 June 2025).
- IUCN. International Union for Conservation of Nature., 2022. Standards and Petitions Committee. Guidelines for Using the IUCN Red List Categories and Criteria. Version 15.1. Prepared by the Standards and Petitions Committee., 2022. Available online: https://www.iucnredlist.org/documents/RedListGuidelines.pdf.
- IUCN. International Union for Conservation of Nature., 2024. The IUCN Red List of Threatened Species. Version 2022-2., 2024. Available online: https://www.iucnredlist.org (accessed on 20 December 2024).
- IUCN. International Union for Conservation of Nature., 2025. The IUCN Red List of Threatened Species. Version 2025-1., 2025. Available online: https://www.iucnredlist.org (accessed on 25 July 2025).
- Jacquet, J.L.; Pauly, D. Trade secrets: Renaming and mislabeling of seafood. Mar. Policy 2008, 32, 309–318. [Google Scholar] [CrossRef]
- King, J.R.; McFarlane, G.A. Marine fish life history strategies: applications to fishery management. Fish. Manag. Ecol. 2003, 10, 249–264. [Google Scholar] [CrossRef]
- Komoroske, L.M.; Lewison, R.L. Addressing fisheries bycatch in a changing world. Frontiers in Marine Science 2015, 2, 1–11. [Google Scholar] [CrossRef]
- Kotas, J.E.; Vizuete, E.P.; Santos, R.A.; Baggio, M.R.; Salge, P.G.; Barreto, R. 2023. PAN Tubarões: Primeiro Ciclo do Plano de Ação Nacional para a Conservação dos Tubarões e Raias Marinhos Ameaçados de Extinção., 2023. ICMBio/CEPSUL, Brasília, DF, Brazil, p. 384.
- Le Grix, N.; Cheung, W.L.; Reygondeau, G.; Zscheischler, J.; Frölicher, T.L. Extreme and compound ocean events are key drivers of projected low pelagic fish biomass. Glob. Chang. Biol. 2023, 29, 6478–6492. [Google Scholar] [CrossRef]
- Lopes, M.S.; Bertucci, T.C.P.; Rapagnã, L.; Tubino, R.d.A.; Monteiro-Neto, C.; Tomas, A.R.G.; Tenório, M.C.; Lima, T.; Souza, R.; Carrillo-Briceño, J.D.; et al. The Path towards Endangered Species: Prehistoric Fisheries in Southeastern Brazil. PLOS ONE 2016, 11, e0154476. [Google Scholar] [CrossRef] [PubMed]
- Loto, L.; Monteiro-Neto, C.; Martins, R.R.M.; Tubino, R.d.A. Temporal changes of a coastal small-scale fishery system within a tropical metropolitan city. Ocean Coast. Manag. 2018, 153, 203–214. [Google Scholar] [CrossRef]
- Lucifora, L.O.; García, V.B.; Worm, B. Global Diversity Hotspots and Conservation Priorities for Sharks. PLOS ONE 2011, 6, e19356. [Google Scholar] [CrossRef]
- Lutz, Í.; Santos, P.E.; Campos, R.; De Oliveira, C.A.C.R.; Wosnick, N.; Evangelista-Gomes, G.; Petrere Jr, M.; Bentes, B. Fishing profile and commercial landings of shark and batoids in a global elasmobranchs conservation hotspot. Anais da Academia Brasileira de Ciências, Rio de Janeiro, Brazil 2024, 96, e20231083. [Google Scholar] [CrossRef]
- Machado, A.A.; de Moraes, F.C.; Aguiar, A.A.; Hostim-Silva, M.; Santos, L.N.; Bertoncini, Á.A. Rocky reef fish biodiversity and conservation in a Brazilian Hope Spot region. Neotropical Ichthyol. 2022, 20. [Google Scholar] [CrossRef]
- Marques, R.A.; Julio, T.G.; Sole-Cava, A.M.; Vianna, M. A new strategy proposal to monitor ray fins landings in south-east Brazil. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 30, 68–85. [Google Scholar] [CrossRef]
- Martins, J.M.A.; Silva, T.S.M.; Fernandes, A.M.; Massone, C.G.; Carreira, R.S. Characterization of particulate organic matter in a Guanabara Bay coastal ocean transect using elemental, isotopic and molecular markers. PANAMJAS 2016, 11, 276–291. [Google Scholar]
- Martins, R.R.; Schwingel, P.R. Variação espaço-temporal da CPUE para o gênero Rhinobatos (Rajiformes, Rhinobatidae) na costa sudeste e sul do Brasil. Braz. J. Aquat. Sci. Technol. 2003, 7, 119–129. [Google Scholar] [CrossRef]
- McCully, S.R.; Scott, F.; Ellis, J.R.; Pilling, G.M. . Productivity and susceptibility analysis: Application and suitability for data poor assessment of elasmobranchs in northern European seas. ICCAT, Collect. Vol. Sci. Pap. 2013, 69, 1679–1698. [Google Scholar]
- McGoodwin, J.R. Comprender las culturas de las comunidades pesqueras: clave para la ordenación pesquera y la seguridad alimentaria. FAO, Doc. Téc. Pesca, Roma 2002, 401, 1–301. [Google Scholar]
- Mejia-Falla, P.A.; Castro, E.R.; Ballesteros, C.A.; Bent-Hooker, H.; Caldas, J.P.; Rojas, A.; Navia, A.F. Effect of a precautionary management measure on the vulnerability and ecological risk of elasmobranchs captured as target fisheries. Reg. Stud. Mar. Sci. 2019, 31, 100779. [Google Scholar] [CrossRef]
- MPA. Ministério da Pesca e Aquicultura. Pesca Artesanal., 2014. Available online: http://www.mpa.gov.br/pesca/artesanal (accessed on 21 October 2024).
- MPF. Ministério Público Federal., 2017. Pesca artesanal legal: pescador da região sul / sudeste - conheça seus direitos e deveres!., 2017. MPF, 6ª Câmara de Coordenação e Revisão, Populações Indígenas e Comunidades Tradicionais, Brasília, DF, Brazil, p. 59.
- A Myers, R.; Worm, B. Extinction, survival or recovery of large predatory fishes. Philos. Trans. R. Soc. B: Biol. Sci. 2005, 360, 13–20. [Google Scholar] [CrossRef]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the world, 5th ed.; John Wiley & Sons: New Jersey, 2016. [Google Scholar]
- Neves, R.A.; Miralha, A.; Guimarães, T.B.; Sorrentino, R.; Calderari, M.R.M.; Santos, L.N. Phthalates contamination in the coastal and marine sediments of Rio de Janeiro, Brazil. Mar. Pollut. Bull. 2023, 190, 114819. [Google Scholar] [CrossRef] [PubMed]
- Oddone, M.; Awruch, C.A.; Barreto, R.R.; Charvet, P.; Chiaramonte, G.E.; Cuevas, J.M.; Dolphine, P.; Faria, V.V.; Paesch, L.; Rincon, G.; et al. guggenheim. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2019. [Google Scholar]
- Patrick, W.S.; et al. 2009. Use of productivity and susceptibility indices to determine the vulnerability of a stock: with example applications to six U.S. fisheries., 2009. NOAA Technical Memorandum, NMFS-F/SPO-101, Silver Spring, p. 104.
- Patrick, W.S.; Spencer, P.; Link, J.; Cope, J.; Field, J.; Kobayashi, D.; Lawson, P.; Gedamke, T.; Cortes, E.; Ormseth, O.; Bigelow, K.; Overholtz, W. Using productivity and susceptibility indices to assess the vulnerability of United States fish stocks to overfishing. Fish. Bull. 2010, 108, 305–322. [Google Scholar]
- Pauly, D.; Cheung, W.W.L. Sound physiological knowledge and principles in modeling shrinking of fishes under climate change. Glob. Chang. Biol. 2017, 24, E15–E26. [Google Scholar] [CrossRef]
- Pauly, D.; Zeller, D. Catch reconstructions reveal that global marine fisheries catches are higher than reported and declining. Nat. Commun. 2016, 7, 1–9. [Google Scholar] [CrossRef]
- Peckham, S.H.; Maldonado-Dias, D.; Koch, V.; Mancini, A.; Gaos, A.; Tinker, M.T.; Nichols, W.J. High mortality of loggerhead turtles due to bycatch, human consumption and strandings at Baja California Sur, Mexico, 2003 to 2007. Endangered Species Research 2008, 5, 171–183. [Google Scholar] [CrossRef]
- Perez, J.A.A.; Pezzuto, P.R.; Wahrlich, R.; Soares, A.L.S. Deep-water fisheries in Brazil: history status and perspectives. Latin American Journal of Aquatic Resources 2009, 37, 513542. [Google Scholar]
- Pérez-Jiménez, J.C.; Mendoza-Carranza, M. Occurrence of immature sharks in artisanal fisheries of the southern Gulf of Mexico. Arquivos de Ciências do Mar, Fortaleza, Brazil 2023, 56, 1–12. [Google Scholar] [CrossRef]
- PMAP-RJ. Projeto de Monitoramento da Atividade Pesqueira na Bacia de Santos., 2020. Relatório Técnico consolidado: 2018-2019, p. 1943.
- PMAP-RJ. Projeto de Monitoramento da Atividade Pesqueira na Bacia de Santos., 2022. Relatório Técnico consolidado: 2019-2021.
- Pollom, R.; Barreto, R.R.; Charvet, P.; Chiaramonte, G.E.; Cuevas, J.M.; Faria, V.V.; Herman, K.; Montealegre-Quijano, S.; Motta, F.; Paesch, L.; et al. Atlantoraja platana. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2020. [Google Scholar]
- Pollom, R.; Barreto, R.R.; Charvet, P.; Chiaramonte, G.E.; Cuevas, J.M.; Faria, V.V.; Herman, K.; Montealegre-Quijano, S.; Motta, F.; Paesch, L.; et al. Dasyatis hypostigma. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2020. [Google Scholar]
- Rigby, C.L.; Dulvy, N.K.; Barreto, R.R.; Carlson, J.; Fernando, D.; Fordham, S.; Francis, M.P.; Herman, K.; Jabado, R.W.; Liu, K.M.; et al. 2019. Sphyrna lewini. The IUCN Red List of Threatened Species., 2019. Available online: https://www.iucnredlist.org/species/pdf/2921825/attachment.
- Santos, P.R.; Balanin, S.; Gadig, O.B.; Garrone-Neto, D. The historical and contemporary knowledge on the elasmobranchs of Cananeia and adjacent waters, a coastal marine hotspot of southeastern Brazil. Reg. Stud. Mar. Sci. 2022, 51, 102224. [Google Scholar] [CrossRef]
- Santos, L.; Veloso, J.V.; Santos, T.B.; Bezerra, N.P.A.; Oliveira, P.; Hazin, F.H.V. An equatorial mid-Atlantic Ocean archipelago as nursery area for the cookiecutter shark: Investigating foraging strategies of neonates through bite mark inferences. J. Fish Biol. 2024, 104, 1290–1298. [Google Scholar] [CrossRef]
- Schroeder, R.; Cardoso, L.G.; Fischer, L.G.; Mourato, B.L.; Monteiro, D.S.; Sant’ana, R. Opening Pandora’s Box: Reconstruction of Catches in Southeast-South Brazil Revealed Several Threatened Elasmobranch Species under One Umbrella Name. Coasts 2024, 4, 552–567. [Google Scholar] [CrossRef]
- Shester, G.G.; Micheli, F. Conservation challenges for small-scale fisheries: Bycatch and habitat impacts of traps and gillnets. Biol. Conserv. 2011, 144, 1673–1681. [Google Scholar] [CrossRef]
- Silva-Junior, L.C.; Andrade, A.C.; Vianna, M. Caracterização de uma pescaria de pequena escala em uma área de importância ecológica para elasmobrânquios, no Recreio dos Bandeirantes, Rio de Janeiro. Arquivos de Ciências do Mar, Fortaleza, Brazil 2008, 41, 47–57. [Google Scholar] [CrossRef]
- de Souza, G.M.; Fernades, I.; Monteiro-Neto, C.; da Costa, M.R. Life-history traits, capture dynamics, and conservation status of key species landed by the artisanal gillnet fleet in the southwest Atlantic Ocean. Rev. Fish Biol. Fish. 2025, 35, 461–486. [Google Scholar] [CrossRef]
- Stobutzki, I.; Miller, M.; Brewer, D. Sustainability of fishery bycatch: a process for assessing highly diverse and numerous bycatch. Environ. Conserv. 2001, 28, 167–181. [Google Scholar] [CrossRef]
- Stobutzki, I.C.; Miller, M.J.; Heales, D.S.; Brewer, D.T. Sustainability of elasmobranchs caught as bycatch in a tropical prawn (shrimp) trawl fishery. Fish. Bull. 2002, 100, 800–821. [Google Scholar]
- Suguio, K. 2003. Geologia Sedimentar. Ed. Edgard Blucher., 2003. São Paulo, Brazil.
- Tavares, R.; Carreon-Zapiain, M.T.; Perez-Jimenez, J.C. Vulnerability of elasmobranchs caught by artisanal fishery in the Southeastern Caribbean. Reg. Stud. Mar. Sci. 2024, 79. [Google Scholar] [CrossRef]
- Thorson, J.T.; Munch, S.B.; Cope, J.M.; Gao, J. Predicting life history parameters for all fishes worldwide. Ecol. Appl. 2017, 27, 2262–2276. [Google Scholar] [CrossRef]
- Toussaint, A.; Brosse, S.; Bueno, C.G.; Pärtel, M.; Tamme, R.; Carmona, C.P. Extinction of threatened vertebrates will lead to idiosyncratic changes in functional diversity across the world. Nat. Commun. 2021, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Valentin, J.L. A ressurgência: Fonte de vida nos oceanos. Ciência Hoje 1994, 18, 18–25. [Google Scholar]
- Valentin, J.L.; Coutinho, R. Modelling maximum chlorophyll in the Cabo Frio (Brazil) upwelling: a preliminary approach. Ecol. Model. 1990, 52, 103–113. [Google Scholar] [CrossRef]
- Vasconcellos, M.; Diegues, A.C.; Kalikoski, D.C. Coastal fisheries of Brazil. In Coastal fisheries of Latin America and the Caribbean; Salas, S., Chuenpagdee, R., Charles, A., Seijo, J.C., Eds.; FAO Fisheries and Aquaculture Technical Paper, 544; FAO: Rome, 2011; pp. 73–116. [Google Scholar]
- Vianna, M.; Almeida, T. Bony fish bycatch in the southern Brazil pink shrimp (Farfantepenaeus brasiliensis and F. paulensis) fishery. Braz. Arch. Biol. Technol. 2005, 48, 611–623. [Google Scholar] [CrossRef]
- Visintin, M.R.; Perez, J.A.A. Vulnerabilidade de espécies capturadas pela pesca de emalhe-de-fundo no Sudeste-Sul do Brasil: produtividade-suscetibilidade (PSA). Bol. Do Inst. de Pesca 2016, 42, 119–133. [Google Scholar] [CrossRef]
- Vooren, C.M.; Klippel, S. Ações para a Conservação de Tubarões e Raias no sul do Brasil. Eds.; Igaré: Porto Alegre, 2005; p. 161. [Google Scholar]
- Walker, T.I. Can shark resources be harvested sustainably? A question revisited with a review of shark fisheries. Mar. Freshw. Res. 1998, 49, 553–572. [Google Scholar] [CrossRef]
- Wosnick, N.; Martins, A.P.B.; Leite, R.D.; Giareta, E.P.; Faria, V.V.; Charvet, P. Brazil. In The global status of sharks, rays, and chimaeras; Jabado, R.W., Morata, A.Z.A., Bennett, R.H., Finucci, B., Ellis, J.R., Fowler, S.L., Grant, M.I., Barbosa Martins, A.P., Sinclair, S.L., Eds.; IUCN: Gland, Switzerland, 2024; pp. 441–456. [Google Scholar]
- Yokota, L.; Lessa, R.P. A nursery Area for Sharks and Rays in Northeastern Brazil. Environ. Biol. Fishes 2006, 75, 349–360. [Google Scholar] [CrossRef]
- Zamboni, A.; Dias, M. , 2020. Auditoria da pesca: Brasil 2020: uma avaliação integrada da governança, da situação dos estoques e das pescarias. Brasília - DF, Brazil: Oceana Brasil.
Figure 1.
Study area: geographical location and detail of southeastern coastal region off Brazil, and the Rio de Janeiro State. Depicting the monitored location of the Z-13 Fisher Colony, Copacabana Beach, Rio de Janeiro, RJ (georeferenced location highlighted in yellow, 22º 59' 10” S/43º 11' 18” W); “Hope Spot - MoNa Cagarras” (georeferenced central-point highlighted in green, 23º 01' 49” S/43º 12' 01” W), a no-take Marine Protected Area (MPA); “Rio de Janeiro Shelf ISRA” encompassing the fishing macro-area used by local fishermen (highlighted in red by dashed lines indicating the suggested buffer for use in the development of appropriate place-based conservation measures, the area of “Praia Vermelha ISRA” is not included); and the 0 to -20 meters (m), -20 to -50 m and the -50 to -70 m isobaths in continuous black lines, according to Suguio (2003). Note: map lines delineate study areas and do not necessarily depict accepted national boundaries. Source: Bárbara Piovani Luz Aieta Afonso.
Figure 1.
Study area: geographical location and detail of southeastern coastal region off Brazil, and the Rio de Janeiro State. Depicting the monitored location of the Z-13 Fisher Colony, Copacabana Beach, Rio de Janeiro, RJ (georeferenced location highlighted in yellow, 22º 59' 10” S/43º 11' 18” W); “Hope Spot - MoNa Cagarras” (georeferenced central-point highlighted in green, 23º 01' 49” S/43º 12' 01” W), a no-take Marine Protected Area (MPA); “Rio de Janeiro Shelf ISRA” encompassing the fishing macro-area used by local fishermen (highlighted in red by dashed lines indicating the suggested buffer for use in the development of appropriate place-based conservation measures, the area of “Praia Vermelha ISRA” is not included); and the 0 to -20 meters (m), -20 to -50 m and the -50 to -70 m isobaths in continuous black lines, according to Suguio (2003). Note: map lines delineate study areas and do not necessarily depict accepted national boundaries. Source: Bárbara Piovani Luz Aieta Afonso.
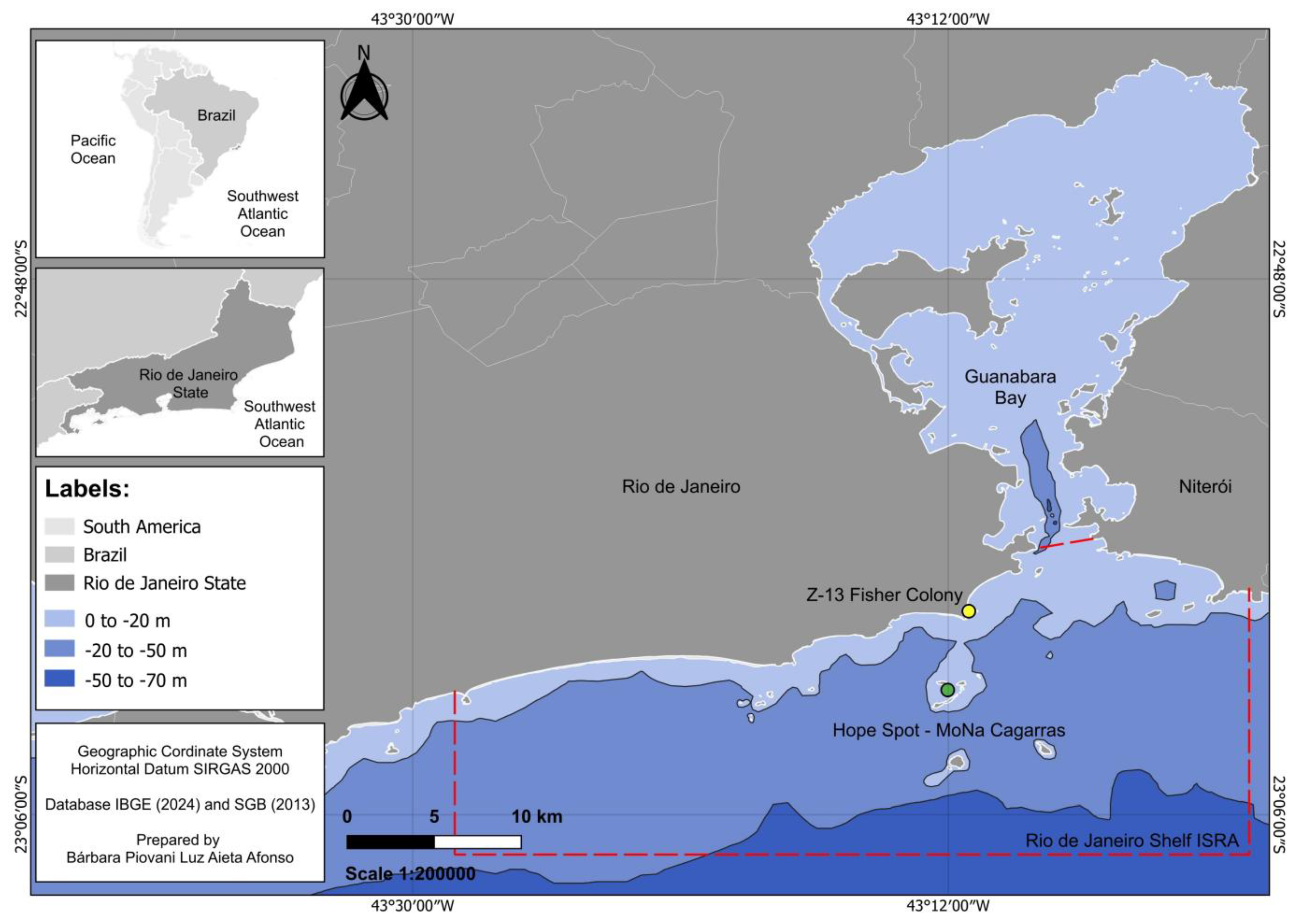
Figure 2.
Productivity (P) and Susceptibility (S) plot for the (a) 16 shark species and for the (b) 15 rays species landed by urban-artisanal gillnets fishing from the Z-13 Fisher Colony, Copacabana Beach, Rio de Janeiro, RJ, Southeastern Brazil.
Figure 2.
Productivity (P) and Susceptibility (S) plot for the (a) 16 shark species and for the (b) 15 rays species landed by urban-artisanal gillnets fishing from the Z-13 Fisher Colony, Copacabana Beach, Rio de Janeiro, RJ, Southeastern Brazil.
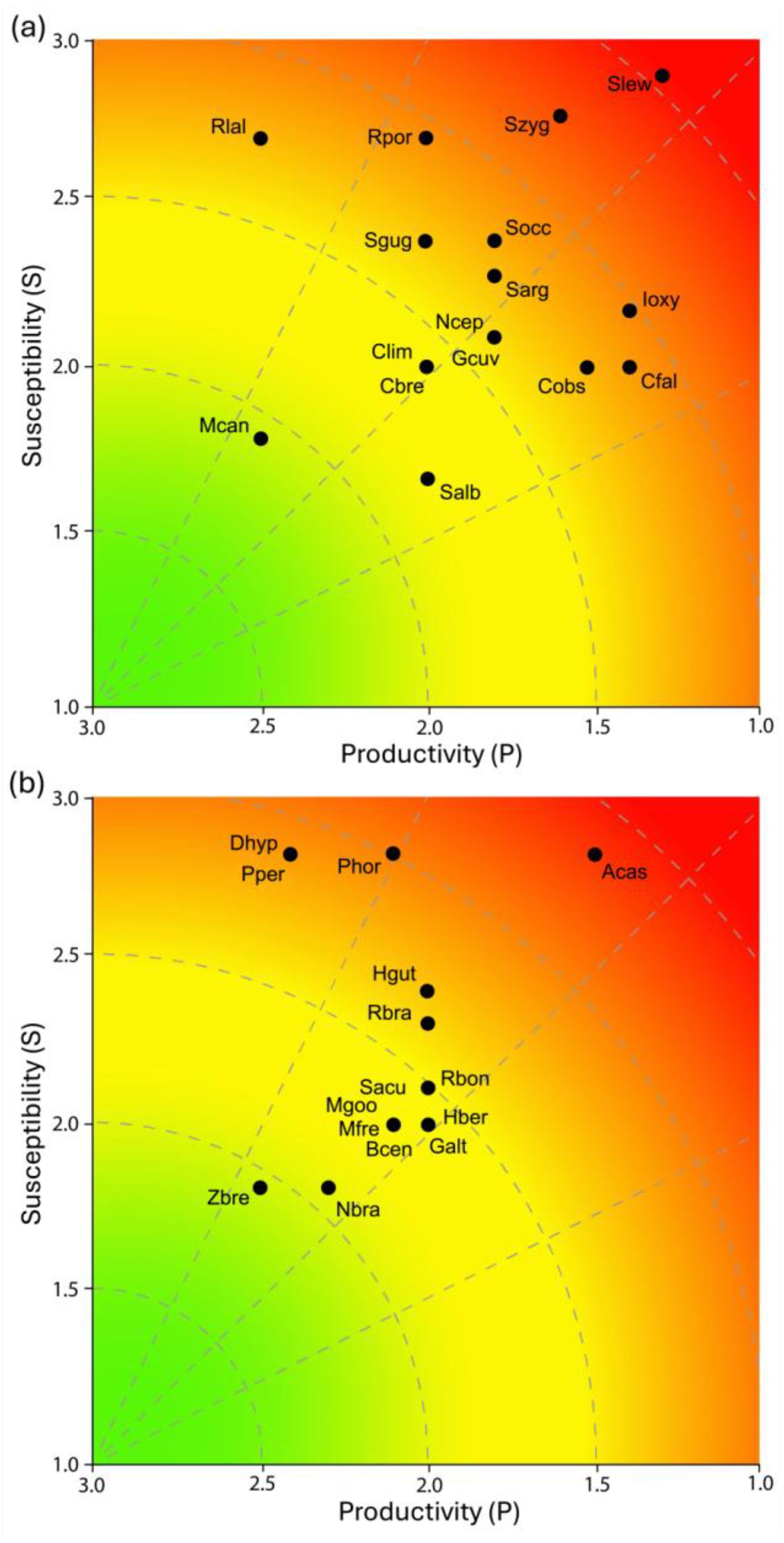

Table 1.
Characterization of the commercial urban-artisanal gillnets fishing from the Z-13 Fisher Colony, Copacabana Beach, Rio de Janeiro, RJ, Southeastern Brazil.
Table 1.
Characterization of the commercial urban-artisanal gillnets fishing from the Z-13 Fisher Colony, Copacabana Beach, Rio de Janeiro, RJ, Southeastern Brazil.
| Z-13 FISHER COLONY (C. P. Z-13): n, average, range and ±SD of the fleet, fishing and gear | |
| C. P. Z-13- DIVERSIFIED COASTAL MODE | LEGISLATION* |
| . Number of vessels (n (total): 19 (average of 10 active vessels/day (range: 00-14; average: 05-07 and; ±SD4-8 | |
| (minimum of 00 and maximum of 14); | |
| . Approx. total length: 05 m (traditional-artisanal boats) and 08 m (speedboats). With beams of 1.4-1.8 m; respectively; | √ |
| . Construction material: fiberglass (with wooden keel); | |
| . GT: ≤ 20 (artisanal fishing); | √ |
| . "Ages": traditional-artisanal boats 20+ years old and speedboats 8-16 years old; | |
| . Engine type: stern drive; | |
| . Power (HP) of diesel engines: "NSB" 75 and 90 / speedboats (gasoline and/or alcohol): 20-40 and 2-4 T; | |
| . Complements: Ursa 40 oil for diesel engines (traditional-artisanal boats) and 2 T oil for gasoline engines | |
| (speedboats); | |
| . Average consumption (sea autonomy): directly proportional to the distance from fishing grounds. | |
| Approximately 03 operational days with a full tank; | |
| . Number of crew/vessel: between 02-03 to 04 (range: 01-04; average: 03 and; ±SD2-3 (minimum of 01 and | |
| maximum of 04); | √ |
| . Materials: the most used gear are gillnets (passive fishing gear with varying dimensions, threads and | |
| meshes, with different buoys and weights - 100.0% of total-local elasmobranch landings during this study); | |
| . Gillnets height: ~02 m; | √ |
| . Configurations - Threads (mm)-meshes (mm): "Corvineiras" (mid-water and surface fishing): 40/60-50/60 | |
| (n = 147; 58.3%); "Linguadeiras" (for bottom fishing only): 50/110 (n = 111; 33.3%) and, "Come-dormes" | |
| (subsurface and mid-water fishing): 40/45-45/55 (n = 38; 8.4%); | √ |
| . Monofilament nylon threads: 0.5-1 mm, gauge (medium thickness); | √ |
| . Opening: 90-120 mm between opposite nodes; | √ |
| . Covers: 10-25 or up to 2,500 m for midwater and surface gillnets. When with two gillnets (or for bottom), | |
| reaching 30 (or up to 3,000 m). Each 100 m longitudinal long and ~1.85 km, on average, from the coast and; | √ |
| . Gillnets permanence time in water: 24-48 h | |
| Where: n (quantitative), ±SD (Standard Deviation), m (meters), GT (Gross Tonnage), T ("times") and, h (hours), and; | |
| *Interministerial Normative Instruction MPA/MMA nº 12 (Art. 2, IV, of NI 12/2012), of 08/22/2012 and MPA/MMA Ordinance | |
| nº 04, of 05/14/2015 (MPF, 2017). | |
Table 2.
Biological data for 31 species of elasmobranchs landed at Z-13 Fisher Colony, Copacabana Beach, Rio de Janeiro, RJ, Southeastern Brazil. The eight Productivity attributes (P) evaluated: maximum age (Tmax, years); maximum size (Lmax, cm); von Bertalanffy’s growth coefficient K (cm year-1); esteemed natural mortality (M, year-1); Length at maturity (L50, cm); Reproductive Cycle (RC, periodicity); Fecundity (Fec., mid-point of the number of oocytes or embryos); and Trophic level (Tlevel, no unit). References data source: see Supplementary Materials - S1.
Table 2.
Biological data for 31 species of elasmobranchs landed at Z-13 Fisher Colony, Copacabana Beach, Rio de Janeiro, RJ, Southeastern Brazil. The eight Productivity attributes (P) evaluated: maximum age (Tmax, years); maximum size (Lmax, cm); von Bertalanffy’s growth coefficient K (cm year-1); esteemed natural mortality (M, year-1); Length at maturity (L50, cm); Reproductive Cycle (RC, periodicity); Fecundity (Fec., mid-point of the number of oocytes or embryos); and Trophic level (Tlevel, no unit). References data source: see Supplementary Materials - S1.
| PRODUCTIVITY PARAMETERS-P | |||||||||
| GROUP/SPECIES | Tmax | Lmax | K | M | L₅₀ | RC | Fec. | Tlevel | SOURCES |
| Sharks | |||||||||
| Carcharhinus brevipinna (Müller & Henle, 1839) (Nrd) | 16.30 | 208.00 | 0.21 | 0.48 | 180.00 | Biennial | 9.00 | 4.20 | Tavares et al. (2024); ** |
| Carcharhinus falciformis (Müller & Henle 1839) (Nrd) | 27.20 | 272.00 | 0.10 | 0.18 | 207.50 | Biennial | 11.70 | 5.48 | Estupiñán-Montaño et al. (2017) |
| Santander-Neto et al. (2021); ** | |||||||||
| Carcharhinus limbatus (Müller & Henle, 1839) | 14.40 (Nrd) | 199.40 (Nrd) | 0.24 (Nrd) | 0.40 (Nrd) | 149.30 (Nrd) | Biennial | 5.50 (Nrd) | 4.30 | Bornatowski et al. (2014) |
| Tavares et al. (2024); ** | |||||||||
| Carcharhinus obscurus (LeSueur, 1818) | 45.00 (Nrd) | 371.00 (Nrd) | 0.04 (Nrd) | 0.90 (Nrd) | 284.00 (Nrd) | Biennial-Triennial | 8.07 (Nrd) | 4.30 | Cortés (2000) |
| Romine et al. (2009) | |||||||||
| Bornatowski et al. (2014); ** | |||||||||
| Rhizoprionodon lalandii (Müller & Henle 1839) | 6.00 (Nrd) | 80.00 | 0.30 (Nrd) | 0.35 (Nrd)* | 62.10 | Annual | 3.50 | 4.28 | Ferreira (1988) |
| Motta et al. (2007) | |||||||||
| Andrade et al. (2008) | |||||||||
| Lessa et al. (2009); ** | |||||||||
| Rhizoprionodon porosus (Poey, 1861) | 5.00 (Nrd) | 115.00 | 0.17 (Nrd) | 0.35 (Nrd) | 85.00 | Annual | 3.00 | 4.00 | Ferreira (1988) |
| Lessa et al. (2009) | |||||||||
| Mattos and Maynou (2009) | |||||||||
| Silva et al. (2023); ** | |||||||||
| Galeocerdo cuvier (Péron & LeSueur, 1822) | 13.50 (Nrd) | 393.00 | 0.25 (Nrd) | 0.36 | 310.00 (Nrd) | Biennial | 32.60 (Nrd) | 4.40 | Whitney and Crow (2007) |
| Driggers et al. (2008) | |||||||||
| Bornatowski et al. (2014) | |||||||||
| Aximoff et al. (2022) | |||||||||
| Santana da Silva et al. (2024); ** | |||||||||
| Notorynchus cepedianus (Perón, 1807) | 32.00 (Nrd) | 300.00 | 0.11 (Nrd) | 0.51 (Nrd) | 242.00 | Annual | 86.70 (Nrd) | 4.43 (Nrd) | Cortés (2000) |
| Lucifora et al. (2005) | |||||||||
| Barnett and Braccini (2019) | |||||||||
| Lewis et al. (2020) | |||||||||
| Lopes et al. (2021) | |||||||||
| Funes et al. (2023); ** | |||||||||
| Isurus oxyrinchus (Rafinesque, 1810) | 23.00 | 330.00 | 0.04 | 0.15 (Nrd) | 278.00 (Nrd) | Triennial | 11.50 | 4.38 | Costa et al. (1996) |
| Bishop et al. (2006) | |||||||||
| Doño et al. (2015) | |||||||||
| Barreto et al. (2016) | |||||||||
| Cabanillas-Torpoco et al. (2024); ** | |||||||||
| Sphyrna lewini (Griffith & Smith, 1834) | 31.50 | 344.00 | 0.05 | 0.13 | 240.00 (Nrd) | Annual | 11.50 (Nrd) | 4.44 | Hazin et al. (2001) |
| Kotas (2004) | |||||||||
| Galina and Vooren (2005) | |||||||||
| Vooren et al. (2005) | |||||||||
| Kotas et al. (2011); ** | |||||||||
| Sphyrna zygaena (Linnaeus, 1758) | 18.00 (Nrd) | 226.00 (Nrd) | 0.07 | 0.13 (Nrd) | 198.00 (Nrd) | Biennial | 35.00 | 4.30 | Vooren et al. (2005) |
| Coelho et al. (2011) | |||||||||
| CMFRI (2016) | |||||||||
| Bezerra et al. (2017) | |||||||||
| Huynh and Tsai (2023); ** | |||||||||
| Squalus albicaudus Viana, Carvalho & Gomes, 2016* | 32.00 (Nrd) | 89.00 (Nrd) | 0.20 | 0.34 | 59.00 (Nrd) | Annual | 4.50 (Nrd) | 4.17 | Compagno (1984) |
| Pajuelo et al. (2011) | |||||||||
| Viana et al. (2016); ** | |||||||||
| Squatina argentina (Marini, 1930) (Nrd) | 46.50 | 138.00 | 0.20* | 0.27* | 70.00 | Biennial-Triennial | 9.00 | 4.39* | Silva (1996) |
| Cortés (2000) | |||||||||
| Vooren and Klippel (2005) | |||||||||
| Cuevas et al. (2020); ** | |||||||||
| Squatina guggenheim Marini, 1936 | 12.00 | 95.50 | 0.27 (Nrd) | 0.36 | 72.00 (Nrd) | Biennial | 7.00 (Nrd) | 4.39 | Silva (1996) |
| Vieira (1996) | |||||||||
| Sunyé and Vooren (1997) | |||||||||
| Vögler et al. (2003) | |||||||||
| Vooren and Klippel (2005) | |||||||||
| Cardoso and Haimovici (2014) | |||||||||
| Della-Fina et al. (2020); ** | |||||||||
| Squatina occulta (Vooren & Silva, 1991) | 21.00 | 125.00 | 0.13 (Nrd) | 0.19 | 110.00 (Nrd) | Biennial | 10.00 | 4.39* | Silva (1996) |
| Vieira (1996) | |||||||||
| Vooren and Klippel (2005) | |||||||||
| Cardoso and Haimovici (2014) | |||||||||
| Della-Fina et al. (2020); ** | |||||||||
| Mustelus canis (Mitchell, 1815) (Nrd) | 11.90 | 132.00 | 0.29 | 0.49 | 102.00 | Annual | 12.00 | 3.60 | Tavares et al. (2024); ** |
| Rays | |||||||||
| Atlantoraja castelnaui (Ribeiro, 1907) | 29.00 | 116.00 | 0.05 | 0.08 | 105.00 | Annual | 5.00 | 4.50 | Casarini (2006) |
| Oddone and Amorim (2007) | |||||||||
| Oddone et al. (2008) | |||||||||
| Silva et al. (2020a) | |||||||||
| Silva et al. (2020b); ** | |||||||||
| Sympterygia acuta Garman, 1877 | 40.50 | 62.00 | 0.12 (Nrd)* | 0.05 | 44.70 (Nrd) | Annual | 52.00 (Nrd) | 3.87 (Nrd) | Basallo and Oddone (2014) |
| Hozbor and Massa (2015) | |||||||||
| Mabragaña et al. (2015) | |||||||||
| Barbinil and Lucifora (2016) | |||||||||
| Pollom et al. (2020); ** | |||||||||
| Bathytoshia centroura (Mitchill, 1815) (Nrd) | 11.00* | 300.00 | 0.28* | 0.48* | 116.50 | Annual | 3.00 | 3.62* | Jacobsen and Bennett (2013); ** |
| Dasyatis hypostigma Santos & Carvalho, 2004 | 11.00 | 58.00 | 0.29 | 0.41 | 49.70 | Annual | 2.00 | 3.70 | Santos and Carvalho (2004) |
| Ribeiro et al. (2006) | |||||||||
| Torres (2024); ** | |||||||||
| Hypanus berthalutzae Petean, Naylor & Lima, 2020 (Nrd) | 14.00* | 68.00 | 0.11* | 0.20* | 67.10* | Annual | 2.50* | 3.62* | Jacobsen and Bennett (2013); ** |
| Hypanus guttatus (Bloch & Schneider, 1801) (Nrd) | 14.00 | 166.60 | 0.11 | 0.20 | 67.10 | Annual | 2.50* | 3.60 | Da Silva et al. (2018) |
| Gianeti et al. (2019) | |||||||||
| Oliveira et al. (2020) | |||||||||
| Tavares et al. (2024); ** | |||||||||
| Gymnura altavela (Linnaeus, 1758) | 18.00 (Nrd) | 143.00 | 0.12 | 0.24 | 71.70 | Annual | 5.50 | 3.91 | Bauchot (1987) |
| Gonçalves-Silva and Vianna (2018) | |||||||||
| Parsons et al. (2018); ** | |||||||||
| Myliobatis freminvillei LeSueur, 1824 (Nrd) | 23.00* | 129.00 | 0.12* | 0.21* | 65.00 | Annual | 4.50* | 3.37* | Jacobsen and Bennett (2013); ** |
| Myliobatis goodei Garman, 1885 (Nrd) | 23.00 | 115.00 | 0.12 | 0.21 | 68.30 | Annual | 4.50 | 3.20 | Molina and Cazorla (2015) |
| Araújo et al. (2016); ** | |||||||||
| Narcine brasiliensis (Olfers, 1831) | 22.55 | 47.00 | 0.12 | 0.20 | 31.89 | Annual | 9.50 (Nrd) | 3.66 (Nrd)* | Rudlow (1989) |
| Jacobsen and Bennett (2013) | |||||||||
| Rolim et al. (2015) | |||||||||
| Rolim et al. (2020); ** | |||||||||
| Pseudobatos horkelii (Müller & Henle, 1841) | 8.00 | 138.00 | 0.18 | 0.41 | 79.60 | Annual | 8.00 (Nrd) | 3.83 | Lessa et al. (1986) |
| Caltabellotta (2014) | |||||||||
| Martins et al. (2017) | |||||||||
| Caltabellotta et al. (2019); ** | |||||||||
| Pseudobatos percellens (Walbaum, 1792) | 11.00 | 76.20 (Nrd) | 0.16 | 0.38 | 58.30 | Annual | 7.50 | 3.83* | Rocha and Gadig (2013) |
| Caltabellotta (2014) | |||||||||
| Tagliafico et al. (2017) | |||||||||
| Caltabellotta et al. (2019); ** | |||||||||
| Rhinoptera bonasus (Mitchill, 1815) | 21.00 (Nrd)* | 120.00 (Nrd) | 0.19 (Nrd)* | 0.26* | 65.30 (Nrd) | Annual | 3.00 (Nrd) | 3.43 (Nrd)* | Nerr and Thompson (2005) |
| Pérez-Jiménez (2011) | |||||||||
| Fisher et al. (2013) | |||||||||
| Jacobsen and Bennett (2013) | |||||||||
| Jones and Driggers (2015); ** | |||||||||
| Rhinoptera brasiliensis Müller, 1836 | 21.00 (Nrd)* | 94.00 | 0.19 (Nrd)* | 0.26 | 65.30 (Nrd)* | Annual | 1.00 | 3.49 (Nrd)* | Gómez-Canchong et al. (2004) |
| Domingues et al. (2009) | |||||||||
| Fisher et al. (2013); ** | |||||||||
| Zapteryx brevirostris (Müller & Henle 1839) | 10.00 (Nrd) | 56.00 (Nrd) | 0.22 | 0.43 | 42.30 | Triennial | 3.75 | 3.55 | Batista (1991) |
| Caltabellotta (2014) | |||||||||
| Carmo (2015) | |||||||||
| Carmo et al. (2018) | |||||||||
| Caltabellotta et al. (2019); ** | |||||||||
| Where: (Nrd), non-regional data (reported outside of the Southeastern Brazilian Bight - SBB), values in bold are estimated or from FishBase, data compiled from congeneric species*, and; | |||||||||
| Sources: "Supplementary dataset." and "Supplementary Information (SI).** at the Supplementary Materials - S1. | |||||||||
Table 3.
Productivity attributes (P), definition and classification of thresholds to determine the Vulnerability (V) off elasmobranchs caught and landed by the urban-artisanal fleet's multispecies fishery (e.g., gillnets) at the monitored fishing macro-area along the continental shelf of the Southeastern Brazilian Bight (SBB), in the Southwestern Atlantic, the Z-13 Fisher Colony, Copacabana Beach, Rio de Janeiro, RJ, Southeastern Brazil.
Table 3.
Productivity attributes (P), definition and classification of thresholds to determine the Vulnerability (V) off elasmobranchs caught and landed by the urban-artisanal fleet's multispecies fishery (e.g., gillnets) at the monitored fishing macro-area along the continental shelf of the Southeastern Brazilian Bight (SBB), in the Southwestern Atlantic, the Z-13 Fisher Colony, Copacabana Beach, Rio de Janeiro, RJ, Southeastern Brazil.
| PRODUCTIVITY ATTRIBUTES | DEFINITION | HIGH (3) | MEDIUM (2) | LOW (1) |
| Maximum age (Tmax, years) | Maximum observed age reported | <11.90 | 11.90-27.20 | >27.20 |
| Maximum size (Lmax, cm) | Maximum Total Length (TL) observed or Fork Length (FL) and Disc Diametric (DD) reported | <89.00 | 89.00-226.00 | >226.00 |
| von Bertalanffy's growth coefficient (K, cm year¯¹) | How rapidly an elasmobranch reaches its maximum size | >0.22 | 0.11-0.22 | <0.11 |
| Esteemed natural mortality (M, year¯¹) | Instantaneous natural mortality rate | >0.41 | 0.20-0.41 | <0.20 |
| Length at maturity (L₅₀, cm) | Length at which 50.0% of the individuals attain gonadal maturity for the first time | <65.00 | 65.00-180.00 | >180.00 |
| Reproductive Cycle (RC, periodicity) | Reproductive periodicity (or time frequency) | Biannual | Annual | Biennial-Triennial |
| Fecundity (Fec., mid-point of the number of oocytes or embryos) | Mid-point of the reported range of the number of oocytes (or embryos) per individual | >11.50 | 3.50-11.50 | <3.50 |
| Trophic level (Tlevel, no unit) | Species position in the food web | <3.62 | 3.62-4.39 | >4.39 |
Table 4.
Susceptibility attributes (S), definition and classification of thresholds to determine the Vulnerability (V) off elasmobranchs caught and landed by the urban-artisanal fleet's multispecies fishery (e.g., gillnets) at the monitored fishing macro-area along the continental shelf of the Southeastern Brazilian Bight (SBB), in the Southwestern Atlantic, the Z-13 Fisher Colony, Copacabana Beach, Rio de Janeiro, RJ, Southeastern Brazil.
Table 4.
Susceptibility attributes (S), definition and classification of thresholds to determine the Vulnerability (V) off elasmobranchs caught and landed by the urban-artisanal fleet's multispecies fishery (e.g., gillnets) at the monitored fishing macro-area along the continental shelf of the Southeastern Brazilian Bight (SBB), in the Southwestern Atlantic, the Z-13 Fisher Colony, Copacabana Beach, Rio de Janeiro, RJ, Southeastern Brazil.
| SUSCEPTIBILITY ATTRIBUTES | DEFINITION | LOW (1) | MEDIUM (2) | HIGH (3) |
| Vertical overlap (VO) | Location of the stock within the water column (i.e., demersal versus pelagic) in relation to the fishing gear |
<25% of the stock occurs at the depths of the fishery | ≥25% and ≤50% of the stock occurs at the depths of the fishery | >50% of the stock occurs at the depths of the fishery |
| Extent of geographic overlap between the | Low probability that an organism encounters a | Medium probability that an organism encounters a | High probability that an organism encounters a | |
| Areal overlap (or Encounterability - E) | known distribution of the stock and that of | fishing gear (e.g., a species of benthic habits caught | fishing gear (e.g., a species of pelagic habits, mainly on the surface | fishing gear (e.g., a species of bento-pelagic habits, caught |
| the fishery | with surface gillnet) | caught with bottom gillnet) | with gillnets) | |
| The ability of the fishing gear to captutre fish | No morphological characteristics or habits | Has few characteristics and habits | Has morphological characteristics and habits | |
| Morphology affecting capture (or Selectivity - S) | based on their morphological characteristics (e.g., body shape, spiny versus soft rayed fins, etc.) | that influence catchability (Low selectivity of the gillnet) | that influence catchability (Moderate selectivity of the gillnet) | that influence catchability (High selectivity of the gillnet) |
| Fishing rate in relation to esteemed natural mortality (M) | Fishing pressure (direct and/or indirect) | 0.00-0.50 | 0.51-0.99 | 1.00 |
| Food items | Forging site versus fishing ground | Spatial distribution of the prey does not overlap | Spatial distribution of prey partially overlapping | Spatial distribution of the prey overlaps completely |
| with the area of action of the fishing mechanism | with the fishing mechanism's area of operation | with the area of action of the fishing mechanism | ||
| Seasonal migrations | Increase or decrease in fishery-species interaction | Reduces overlap with fishing | Does not substantially affect the overlap with fishing | Increases overlap with fishing |
| when any seasonal migrations occur | ||||
| Schooling/aggregation/geographic concentration and behavioral responses | Aggregations of organisms for feeding or reproduction | Reduces the catchability of the fishing gear | Does not substantially affect the catchability of the fishing gear | Increases the catchability of the fishing gear |
| Fishery impact | Direct and/or indirect impacts to essential fish habitat | Adverse effects absent, minimal or temporary | Adverse effects more than minimal or temporary but are mitigated | Adverse effects more than minimal or temporary and are not mitigated |
| Valued or desired of the fishery | Value of fish to the final consumer | Stock is not highly valued or desired by the fishery (low or no price) | Stock is moderately valued or desired by the fishery (medium price) | Stock is highly valued or desired by the fishery (high price) |
| Management strategy | Current fisheries legislation | Targeted stocks have catch limits and proactive accountability measures; Non-target stocks are closely monitored | Targeted stocks have catch limits and reactive accountability measures | Targeted stocks do not have catch limits or accountability measures; Non-target stocks are not closely monitored |
| Immatures caugth (%) | Capture of juveniles/species (%) in relation to the selectivity | 0.0-30.0% | 31.0-60.0% | 61.0-100.0% |
| off the fishing gear in the catches sampled | ||||
| IUCN status | Conservation status | NE-DD | LC-NT | VU-EN-CR |
Table 5.
Productivity (P), Susceptibility (S), and Vulnerability (V) scores for 31 species of elasmobranchs landed by commercial urban-artisanal gillnets fishing at the Z-13 Fisher Colony, Copacabana Beach, Rio de Janeiro, RJ, Southeastern Brazil. The eight species indicated in bold (n = 05, Selachii (sharks) and n = 03, Batomorphi (rays and relatives; batoids) are those with ≥ 5.0 percentage of importance per respective group according to the total of specimens/species landed. Also shown is the determination of quality input data for P, S, and overall values, and the classification of risk category (High: V ≥ 2.0; Medium: 1.8 ≤ V < 2.0; and Low: V < 1.8).
Table 5.
Productivity (P), Susceptibility (S), and Vulnerability (V) scores for 31 species of elasmobranchs landed by commercial urban-artisanal gillnets fishing at the Z-13 Fisher Colony, Copacabana Beach, Rio de Janeiro, RJ, Southeastern Brazil. The eight species indicated in bold (n = 05, Selachii (sharks) and n = 03, Batomorphi (rays and relatives; batoids) are those with ≥ 5.0 percentage of importance per respective group according to the total of specimens/species landed. Also shown is the determination of quality input data for P, S, and overall values, and the classification of risk category (High: V ≥ 2.0; Medium: 1.8 ≤ V < 2.0; and Low: V < 1.8).
| GROUP/SPECIES | iD SPECIES | PRODUCTIVITY | PRODUCTIVITY | SUSCEPTIBILITY | SUSCEPTIBILITY | OVERALL | VULNERABILITY | RISK |
| DATA QUALITY | DATA QUALITY | DATA QUALITY | ||||||
| Sharks | ||||||||
| Sphyrna lewini | Slew | 1.3 | 4.8 | 2.9 | 2.0 | 3.1 | 2.6 | High |
| Sphyrna zygaena | Szyg | 1.6 | 5.3 | 2.8 | 2.0 | 3.3 | 2.3 | High |
| Isurus oxyrinchus | Ioxy | 1.4 | 4.3 | 2.2 | 2.0 | 2.9 | 2.0 | High |
| Squatina occulta | Socc | 1.8 | 4.5 | 2.4 | 2.0 | 3.0 | 1.9 | Moderate |
| Rhizoprionodon porosus | Rpor | 2.0 | 5.0 | 2.7 | 2.0 | 3.2 | 1.9 | Moderate |
| Carcharhinus falciformis | Cfal | 1.4 | 6.0 | 2.0 | 2.0 | 3.6 | 1.9 | Moderate |
| Carcharhinus obscurus | Cobs | 1.5 | 5.5 | 2.0 | 2.0 | 3.4 | 1.8 | Moderate |
| Squatina argentina | Sarg | 1.8 | 6.0 | 2.3 | 2.0 | 3.6 | 1.8 | Moderate |
| Rhizoprionodon lalandii | Rlal | 2.5 | 4.8 | 2.7 | 2.0 | 3.1 | 1.7 | Low |
| Notorynchus cepedianus | Ncep | 1.8 | 5.0 | 2.1 | 2.0 | 3.2 | 1.7 | Low |
| Galeocerdo cuvier | Gcuv | 1.8 | 5.0 | 2.1 | 2.0 | 3.2 | 1.7 | Low |
| Squatina guggenheim | Sgug | 2.0 | 5.3 | 2.4 | 2.0 | 3.3 | 1.7 | Low |
| Carcharhinus limbatus | Clim | 2.0 | 5.5 | 2.0 | 2.0 | 3.4 | 1.4 | Low |
| Carcharhinus brevipinna | Cbre | 2.0 | 6.3 | 2.0 | 2.0 | 3.7 | 1.4 | Low |
| Squalus albicaudus | Salb | 2.0 | 6.0 | 1.7 | 2.0 | 3.6 | 1.2 | Low |
| Mustelus canis | Mcan | 2.5 | 6.3 | 1.8 | 2.0 | 3.7 | 1.0 | Low |
| Rays | ||||||||
| Atlantoraja castelnaui | Acas | 1.5 | 3.8 | 2.8 | 2.0 | 2.7 | 2.3 | High |
| Pseudobatos horkelii | Phor | 2.1 | 4.3 | 2.8 | 2.0 | 2.9 | 2.0 | High |
| Dasyatis hypostigma | Dhyp | 2.4 | 3.8 | 2.8 | 2.0 | 2.7 | 1.9 | Moderate |
| Pseudobatos percellens | Pper | 2.4 | 4.0 | 2.8 | 2.0 | 2.8 | 1.9 | Moderate |
| Hypanus guttatus | Hgut | 2.0 | 6.0 | 2.4 | 2.0 | 3.6 | 1.7 | Low |
| Rhinoptera brasiliensis | Rbra | 2.0 | 5.0 | 2.3 | 2.0 | 3.2 | 1.6 | Low |
| Rhinoptera bonasus | Rbon | 2.0 | 6.0 | 2.1 | 2.0 | 3.6 | 1.5 | Low |
| Gymnura altavela | Galt | 2.0 | 5.0 | 2.0 | 2.0 | 3.2 | 1.4 | Low |
| Hypanus berthalutzae | Hber | 2.0 | 6.3 | 2.0 | 2.0 | 3.7 | 1.4 | Low |
| Sympterygia acuta | Sacu | 2.1 | 5.0 | 2.0 | 2.0 | 3.2 | 1.3 | Low |
| Myliobatis goodei | Mgoo | 2.1 | 6.0 | 2.0 | 2.0 | 3.6 | 1.3 | Low |
| Myliobatis freminvillei | Mfre | 2.1 | 6.5 | 2.0 | 2.0 | 3.8 | 1.3 | Low |
| Bathytoshia centroura | Bcen | 2.1 | 6.8 | 2.0 | 2.0 | 3.9 | 1.3 | Low |
| Narcine brasiliensis | Nbra | 2.3 | 4.0 | 1.8 | 2.0 | 2.8 | 1.1 | Low |
| Zapteryx brevirostris | Zbre | 2.5 | 4.8 | 1.8 | 2.0 | 3.1 | 1.0 | Low |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



