Submitted:
22 September 2025
Posted:
23 September 2025
You are already at the latest version
Abstract
Animal trypanosomiasis, primarily caused by Trypanosoma evansi (surra) and related species, represents a neglected but economically significant disease of livestock in Central Asia. In Ka-zakhstan, infections in camels, horses, and cattle contribute to productivity losses and undermine rural livelihoods. Although Human African Trypanosomiasis (HAT) has not been reported in the country, climate change and increased livestock mobility raise concerns about future risks of exotic introductions. This review synthesises current knowledge on the epidemiology, diagnosis, and control of animal trypanosomiasis in Kazakhstan within a regional and global context. Particular attention is given to diagnostic approaches: while conventional parasitological and serological methods remain in use, molecular tools are underdeveloped and inconsistently applied in vet-erinary settings. We highlight gaps in surveillance, the lack of species-specific diagnostic proto-cols, and the need to distinguish clearly between techniques for human versus animal trypano-somes. Framed within a One Health perspective, we argue that addressing animal trypanosomiasis is critical not only for protecting livestock productivity but also for strengthening regional pre-paredness against potential zoonotic threats.
Keywords:
zoonotic pathogens
; vector-borne trypanosomiasis
; T. brucei
; T.cruzi
; T.evansi
; T.equiperdum
; One Health
1. Introduction
Vector-borne diseases are among the most serious threats to global health. According to the World Health Organization (WHO), these infections—caused by parasites, viruses, and bacteria transmitted by arthropod vectors—account for nearly 17% of the global burden of infectious diseases and claim more than 700,000 lives every year [1]. Their capacity for rapid international spread, high transmissibility, and pandemic potential underscores the urgency of addressing them. The burden is especially heavy in tropical and subtropical regions, where favorable environmental conditions for vectors coincide with vulnerable populations [1].
The distribution of vector-borne diseases is determined by a complex interplay of demographic, environmental, and social factors. Morbidity and mortality rates are often disproportionately high in affected populations, while the economic toll is equally devastating. Beyond human health, vector-borne diseases cause direct losses in livestock production and compromise the viability of wild animal populations, collectively limiting the development of both rural and urban areas [1].
Within the WHO’s priority list of vector-borne diseases, trypanosomiases occupy a unique position. Human African trypanosomiasis (HAT, or sleeping sickness) and Chagas disease (American trypanosomiasis) are regionally restricted but highly impactful, exerting profound effects on human health and socioeconomic stability in endemic communities [1].
The challenge is further intensified by climate change. Climate scientists warn that gradual global warming is altering rainfall patterns, enabling monsoon activity in once-arid zones, and expanding vegetation cover [2,3]. These ecological changes shape the “microcosm” of pathogens—viruses, bacteria, fungi [4]—and their carriers such as midges and insects [5]. Consequently, tropical and exotic vector-borne diseases can appear suddenly in regions where they were previously unknown, reshaping epidemiological landscapes and testing the resilience of public health and veterinary systems.
In recent years, diseases such as African swine fever, lumpy skin disease, and trypanosomiasis have been recorded in the Republic of Kazakhstan. Environmental changes, including repeated river flooding in formerly arid regions, create favourable ecological conditions for the proliferation of vectors and the establishment of tropical and exotic infectious and parasitic diseases.
This review aims to provide a critical analysis of trypanosomatid infections of various etiologies, with particular attention to their high transmissibility, zoonotic potential, and broad host range. Special emphasis is placed on assessing the impact of climate change on vector distribution, and on comparing classical versus modern diagnostic methods for protozoa of the genus Trypanosoma, with distinctions drawn according to the type of pathogen.
Trypanosoma
Trypanosomiasis is a tropical, vector-borne disease caused by protozoan parasites of the genus Trypanosoma (class Kinetoplastea, family Trypanosomatidae), which infect both humans and animals [6]. The genus belongs to a monophyletic group of flagellated protozoa, characterised by a distinctive kinetoplast DNA structure and a corkscrew-like motility, reflected in the Greek etymology trypaô (“to bore”) and soma (“body”) [6].
To date, approximately 125 species of Trypanosoma have been described in mammals, of which about 10% are pathogenic to humans and/or domestic animals [7]. These parasites are widely distributed in Africa, the Americas, Latin America, and Asia, where they contribute to severe human disease, major epizootics in livestock, and substantial economic losses in animal production [8,9,10,11,12,13].
Figure 1 shows the map that highlights the distinct yet overlapping geographic distribution of Trypanosoma infections. Human African Trypanosomiasis (HAT) remains confined to sub-Saharan Africa, Chagas disease is mainly endemic in Latin America, while animal trypanosomiases (e.g., surra, nagana, dourine) extend into Africa, Asia, and beyond. This visualisation underlines the global nature of trypanosomiasis, contrary to the perception of it as a strictly regional problem. Figure 1 also demonstrates the dual burden of trypanosomiasis. Human cases are geographically restricted, but animal infections are more widely distributed, causing significant economic losses in livestock production. The largest endemic zones coincide with regions of high dependence on livestock. This amplifies the financial burden of animal trypanosomiasis while raising concern about the zoonotic potential of species such as T. evansi and T. vivax. It also provides a platform for discussing climate-driven expansion of vector habitats, which could blur current geographic boundaries. Trypanosomiasis should be prioritised as a neglected tropical disease (NTD). It provides evidence that coordinated international action is required, not only to control human trypanosomiasis in its remaining foci, but also to strengthen surveillance of animal trypanosomes, which remain underdiagnosed and underreported despite their wide distribution.
In humans, two primary forms of disease are recognised: Human African trypanosomiasis (HAT, or sleeping sickness) and American trypanosomiasis (Chagas disease) [14]. HAT is caused by Trypanosoma brucei gambiense (T. b. g.) and T. b. rhodesiense (T. b. r.), while Chagas disease is caused by T. cruzi [14]. Despite differences in geographic distribution and clinical presentation, these pathogens share morphological similarities that complicate diagnosis and species-specific identification [15,16,17,18,19].
Trypanosoma brucei is the causative agent of Human African Trypanosomiasis (HAT, or sleeping sickness), a neglected parasitic disease that affects both humans and animals [21,22,23,24,25]. Two subspecies are pathogenic to humans: T. b. gambiense (T.b.g.) and T. b. rhodesiense (T.b.r.), first described by David Bruce. These are commonly referred to as West African (Gambian) HAT and East African (Rhodesian) HAT, respectively. Among reported human cases, T. b. gambiense accounts for more than 98%, reflecting its dominance in endemic regions [20].
Transmission occurs primarily through the bite of an infected tsetse fly (Glossina spp.), which is widespread in rural sub-Saharan Africa. Three major groups of tsetse flies are responsible for parasite transmission: the morsitans group (Glossina morsitans), which inhabits open savannah and woodland; the palpalis group (G. palpalis), which thrives along rivers and lakes; and the fusca group (G. fusca), which is associated with forested ecosystems [20].
Although vector-borne transmission remains the primary route, other forms have been documented. These include vertical transmission from mother to child, mechanical transmission by other blood-sucking insects, transmission through contaminated needles, and even sexual transmission. Sporadic cases of congenital and transfusion-associated infections further highlight the adaptability of the parasite and the challenges of controlling its spread [21,22,23,24,25].
American trypanosomiasis (Chagas disease), caused by Trypanosoma cruzi, was first described in South America by the Brazilian physician Carlos Chagas, who identified both the parasite and its insect vector in the early 20th century [26,27,28]. The parasite was named T. cruzi in honor of Oswaldo Cruz. Locally, the triatomine vector is commonly referred to as the “kissing bug” (chupão) or “barber bug” (barbeiro), reflecting its nocturnal habit of feeding on human blood while people sleep [26,27,28].
Although the disease was initially thought to be restricted to the Americas, evidence from archaeological remains of 4,000–9,000-year-old mummies suggests that T. cruzi has been infecting humans for millennia [26,27,28]. Furthermore, studies by Coura, Viñas, and Junqueira emphasize that the sylvatic cycle of Chagas disease has existed for millions of years, maintained in diverse wild reservoirs long before human settlements emerged [26,27,28]. With increasing encroachment of human activity into wildlife habitats and the expansion of travel to remote areas, the risk of human contact with infected vectors and the potential for parasite adaptation or zoonotic spillover is heightened.
Over the past decades, the epidemiological profile of Chagas disease has shifted from a rural to an urban setting. Factors such as population displacement, urbanization, and migration have facilitated its spread beyond endemic regions. Cases have now been documented not only in the Americas—including Canada and the United States—but also in Europe, Africa, the Eastern Mediterranean, and the Western Pacific due to human mobility [29]. According to WHO, an estimated 75 million people remain at risk of infection, with many cases undiagnosed or untreated, particularly in areas where vector transmission persists [29].
Cases of Chagas disease are no longer confined to Latin America. Reports now document infections in Europe, Africa, the Eastern Mediterranean, and the Western Pacific, reflecting the global spread of the parasite through migration and travel [29]. The World Health Organization (WHO) estimates that nearly 75 million people remain at risk of infection, mainly due to the high proportion of undiagnosed or untreated cases, as well as the persistence of active vector transmission in endemic regions [29].
Transmission is primarily associated with blood-sucking insects of the subfamily Triatominae (family Reduviidae), whose natural habitat includes Mexico, Central, and South America [20,25]. In the work of Schmunis, human infection with T. cruzi was reported across 21 countries in North, Central, and South America, highlighting its wide geographic distribution [30]. Most infections in the Western Hemisphere occur through direct contact with infected vectors. However, T. cruzi can persist in the blood of untreated individuals for decades, enabling additional transmission routes such as blood transfusion and organ transplantation, now recognized as the second most common mode of spread [30]. Congenital transmission from mother to child represents a third important pathway.
Socioeconomic factors strongly influence disease risk. Poverty, combined with political and economic instability, has fueled migration from endemic countries to developed regions such as Australia, Canada, Spain, and the United States. Consequently, cases of transfusion-related, transplant-related, and congenital Chagas disease have been documented in non-endemic countries, including Spain and the United States [31].
Taken together, these findings emphasize the parasitic aggressiveness and adaptability of trypanosomatids. Their broad host range, diverse transmission routes, and growing international footprint highlight the urgent need for enhanced surveillance and monitoring of Chagas disease—a neglected vector-borne infection with profound social and economic consequences, now posing risks beyond traditionally endemic regions [29,30,31].
Trypanosomiasis of Animals: Surra and Dourine
Animal trypanosomiasis is a protozoan disease caused by parasites of the genus Trypanosoma and transmitted by blood-sucking vectors, including the tsetse fly and other biting insects [32]. The principal veterinary species include T. congolense, T. vivax, T. brucei brucei, and T. simiae. At the same time, T. brucei rhodesiense and T. b. gambiense are zoonotic, with humans as their primary hosts [32]. Historically, several animal trypanosomiases were given distinct names—surra (T. evansi), dourine (T. equiperdum), and nagana (caused by T. brucei, T. congolense, and T. vivax)—to distinguish clinical syndromes. However, a critical review of published and unpublished data indicates that clinical signs such as abdominal swelling, emaciation, anaemia, and neurological symptoms are nonspecific and overlapping, making differential diagnosis challenging and often unreliable in the absence of laboratory confirmation [32].
The geographical distribution of camel populations and sampling points across the main camel-breeding regions like Mangystau, Kyzylorda, and Turkestan regions (Figure 2) underscores the epidemiological importance of these areas for trypanosomiasis surveillance. Notably, the regions with the largest camel populations (dark blue zones) coincide with areas of higher sampling density, reflecting the strategic prioritisation of surveillance in zones of intensified animal husbandry. Such mapping is critical, as camel breeding in Kazakhstan is concentrated in desert and semi-desert zones, where environmental and climatic factors favour vector activity and parasite circulation. The data presented in this figure also highlight potential “hotspots” where animal density and movement may enhance transmission risks, reinforcing the need for targeted monitoring, early diagnostics, and preventive measures. These findings align with the One Health perspective, demonstrating how ecological and demographic conditions shape the risk landscape for both veterinary and public health sectors.
In addition to pathogenic species, non-pathogenic trypanosomes such as T. theileri are also widespread. These digenetic blood parasites, transmitted by biting flies, are commonly detected in ungulates including cattle, buffalo, sheep, antelope, and deer [33,34,35,36,37,38]. Although generally regarded as non-pathogenic and of limited economic importance, T. theileri infections may present in a latent form and become clinically relevant under stress or in the presence of coinfections, manifesting with fever, anorexia, or anaemia [33,34,35,36,37,38]. Despite their wide distribution, these species remain neglected, mainly in veterinary research and surveillance.
Genetic studies have provided new insights into the taxonomy of pathogenic trypanosomes. For example, Lai et al. (2008) demonstrated that both T. equiperdum and T. evansi represent T. brucei strains that have lost either part of their kinetoplast DNA (dyskinetoplastic, Dk) or the entire kinetoplast genome (akinetoplastic, Ak), resulting in distinct but related lineages [33].
Among the diseases listed by the World Organization for Animal Health (WOAH) as notifiable infections of international concern are surra (T. evansi) and dourine (T. equiperdum), classified under diseases of multiple species and equidae [34,35,36,37,38]. Their inclusion reflects their:
- Wide global distribution
- Zoonotic potential
- Capacity to spread rapidly in naïve populations, and
- Severe impact on animal health and productivity.
Of these, T. evansi is considered the most essential veterinary pathogen due to its comprehensive host range, which includes camels, horses, cattle, buffalo, goats, sheep, pigs, dogs, tigers, and Asian elephants [39,40]. Infections cause profound economic losses, high mortality in susceptible hosts, and have been associated with vaccination failure, further exacerbating animal vulnerability [39,40]. While primarily an animal disease, sporadic human cases have been reported in India, Vietnam, and Sri Lanka, raising concern about its zoonotic potential [9,41,42,43].
Analysis of horse population dynamics, based on data as of March 1 for the period 2021–2025 by the Bureau of National Statistics of the Republic of Kazakhstan, shows a steady upward trend. The lowest recorded number of horses was in 2021, with 3.49 million head, whereas in 2025 the population reached its peak at 4.80 million head (Figure 3).
Diagnosis of T. evansi remains problematic. Parasitological methods are field-adaptable and straightforward but lack sensitivity. Improved techniques, such as the hematocrit centrifugation technique (HCT) and the modified miniature anion-exchange centrifugation technique (mAECT), increase detection rates, though they still miss chronic infections [44]. Molecular methods outperform antigen-based assays, enabling detection in both prepatent and chronic stages, but their field implementation remains limited [45].
Geographically, T. evansi is widely distributed across Asia, North America, Central America, and South America. In unvaccinated horses and camels, surra is frequently fatal [11,46,47]. Across Africa, Asia, and Latin America, surra continues to cause severe disease in livestock and wildlife, with significant economic impacts on rural livelihoods [8,48,49,50,51,52,53].
The European Food Safety Authority (EFSA) has assessed T. evansi (surra) under the Animal Health Law (AHL), applying criteria related to disease profile, impact, prevention, and control [38].
However, consensus remains lacking on whether T. evansi meets the requirements for inclusion in the Union intervention list (Article 5(3)). Ambiguities persist regarding its classification under Annexe IV and the list of affected animal species (Article 8), highlighting ongoing regulatory uncertainty in Europe [38].
Equine trypanosomosis is a complex infectious disease of horses caused by several Trypanosoma species, including T. evansi, T. equiperdum, T. brucei, T. vivax, T. congolense, and occasionally T. cruzi [10,11,12]. The classical syndromes historically described as surra (T. evansi), dourine (T. equiperdum), and nagana (T. brucei, T. congolense, T. vivax) are difficult to distinguish clinically, since common symptoms such as abdominal swelling, emaciation, anaemia, and neurological signs are largely nonspecific [10,11,12].
Instead, the primary differences lie in their epidemiology and transmission routes:
- T. brucei and T. congolense are transmitted by tsetse flies in sub-Saharan Africa;
- T. equiperdum is sexually transmitted
Taxonomic classification remains controversial. Many isolates historically identified as T. equiperdum have since been recognised as misidentified strains of T. evansi. However, phylogenetic analyses suggest that T. evansi and T. equiperdum evolved independently from T. brucei [10,11,12,13]. Both species are microheterogeneous populations rather than uniform taxa [14]. Molecular tools, particularly the analysis of kinetoplast DNA (kDNA), can help differentiate them: T. evansi and some T. equiperdum strains lack the maxicircle component of kDNA, which is present in T. brucei [32,54]. However, these distinctions remain debated, underscoring the diagnostic challenges within the Trypanozoon subgenus.
Dourine poses particular challenges for equine health and trade. The parasite infects both domestic and wild ungulates, and the movement of breeding horses facilitates its spread across borders. Historical records show that the peak incidence of dourine in the former Soviet republics—including Russia, the Caucasus, Central Asia, the Baltic States, Moldova, Belarus, and Ukraine—occurred between 1961 and 1965. A later outbreak in Russia during 1998–1999 accounted for 70% of all registered equine diseases [55]. Researchers at that time identified pathogenic isolates representing both surra (T. evansi var. ninaekohlyakimovi) and dourine (T. equiperdum) [55].
For Kazakhstan, monitoring, early diagnosis, and prevention of equine trypanosomosis (surra and dourine) are of strategic importance. The breeding of camels and horses has historically been a cornerstone of traditional animal husbandry and remains economically valuable today, particularly in the country’s extensive desert and semi-desert zones. As highlighted in recent studies (Frontiers in Veterinary Science, 2025), these regions represent epidemiologically significant areas where high-density camel populations overlap with ecological conditions favourable to vector activity and parasite circulation. The persistence of trypanosomiasis therefore represents not only a veterinary challenge but also a substantial threat to rural livelihoods and national economic resilience.
Here, it is important to note that the present work was carried out within the framework of the Scientific Research Project IRN BR218004/0223, «Improving biosafety measures in Kazakhstan: countering dangerous and especially dangerous infections» (2023–2025), funded by the Science Committee of the Ministry of Science and Higher Education of the Republic of Kazakhstan. Within this project, scientists from the Kazakh Scientific Research Veterinary Institute (KazSRVI) are conducting ongoing monitoring studies on animal trypanosomosis, the results of which will be published in future reports and peer-reviewed publications. These efforts build directly on the recent findings published in Frontiers in Veterinary Science [56], thereby strengthening the national research framework for controlling neglected vector-borne zoonoses under the One Health approach.
Cases of Human Infection with T. evansi
According to WHO data (2020), deaths from trypanosomiasis in Kazakhstan were 0 or 0.00% of total mortality, with an age-adjusted mortality rate of 0.00 per 100,000 population, ranking Kazakhstan 92nd worldwide [57]. While no human cases are documented locally, sporadic reports of human infections with animal trypanosomes have been described globally. These include T. vivax, T. congolense, T. b. brucei, T. evansi, and T. lewisi or T. lewisi-like parasites, collectively termed atypical human trypanosomiasis (a-HT) [8,58,59,60].
Figure 4 clearly illustrates the rarity yet global distribution of atypical human trypanosomiasis (a-HT) cases, including those caused by T. evansi. While confirmed human infections have been documented in India, Egypt, and Vietnam, alongside sporadic reports elsewhere, the absence of cases in Kazakhstan should not be interpreted as immunity to risk. On the contrary, Kazakhstan’s intensive horse and camel husbandry, active livestock trade, and vulnerability to climate change create conditions that could facilitate zoonotic spillover. The contrast between “zero cases” in Kazakhstan and “sporadic but proven cases” globally underscores two critical points: (i) the underestimated zoonotic potential of animal trypanosomes, and (ii) the urgent need for enhanced surveillance and reliable diagnostic tools to detect emerging infections before they pose wider threats.
The zoonotic potential of animal trypanosomes is linked to socio-economic and environmental conditions, especially in regions with close human–livestock interactions and abundant mechanical vectors [61,62,63]. Experimental studies in India demonstrated that diverse mammals—including albino rats, guinea pigs, bandicoot rats, mongooses, cats, and monkeys—are susceptible to T. evansi following syringe passage, with variable virulence [63]. Notably, rhesus macaques developed severe clinical signs resembling human sleeping sickness, suggesting that primates could serve as potential models of zoonotic infection [64]. Confirmed human infections with T. evansi remain rare but significant. In India (2005), a molecularly confirmed case demonstrated high parasitemia without central nervous system involvement, although T. evansi typically invades the CNS in horses within two weeks [10,39,65,66]. In Egypt (2010), one human sample out of 30 tested positively by ELISA and microscopy, and the patient recovered following treatment [59,60]. In Vietnam (2016), a healthy individual without APOL1 deficiency developed confirmed T. evansi infection, most likely acquired through exposure to infected cattle meat [47]. These cases draw attention to the role of apolipoprotein L1 (APOL1) in human innate immunity. While humans are naturally resistant to T. b. brucei through APOL1-mediated lysis [47], a frame-shift mutation in the APOL1 gene was identified in the Indian patient, explaining the absence of trypanolytic activity [67]. However, no APOL1 mutation was detected in the Vietnamese patient (2015), indicating that other host or parasite factors may facilitate infection [47,68].
Collectively, these findings demonstrate that close contact with infected animals increases the risk of zoonotic spillover of T. evansi and highlight the importance of controlling parasitemia in livestock to prevent atypical human infections across continents [69]. For Kazakhstan, where horses and camels are integral to animal husbandry and are actively traded internationally, the surveillance and prevention of surra are of strategic importance for both veterinary and public health sectors.
Diagnosis of Trypanosomiasis
Despite more than a century of study, dourine diagnosis remains a challenge. Only a small number of T. equiperdum laboratory strains have been characterised, and data on most isolates remain unpublished. Definitive diagnosis is possible only at the serological or molecular level; clinical signs are non-specific, and international screening still relies heavily on the outdated complement fixation test (CFT) developed in 1915, which remains cross-reactive and may produce misleading results [47]. In Abay et al.’s study, large-scale field data from Kazakhstan showed that the formalin gel test (FGT) detected more seropositive camels than the complement fixation test (CFT), supporting a combined FGT+CFT screening algorithm [Frontiers | Serological Surveillance of Trypanosoma evansi in Kazakhstani Camels by Complement Fixation and Formalin Gel Tests [56].
Figure 5 underscores several key points. First, there is no single gold standard capable of meeting the diagnostic demands of both field settings and trade-sensitive surveillance. Parasitological methods remain useful for confirming active infection but systematically miss chronic carriers. Serology fills this gap with higher sensitivity but introduces the risk of false positives due to cross-reactivity, which can be particularly problematic in the context of Kazakhstan’s role as an exporter and importer of thoroughbred horses. Molecular methods provide the highest accuracy and allow early detection. Yet, their dependence on laboratory infrastructure and their current inability to separate closely related subspecies limit practical application in endemic regions.
Taken together, the relative weighting in this pie chart reflects a fragmented diagnostic landscape where each method has strengths but also critical weaknesses. For Kazakhstan and comparable regions, this fragmentation is not merely an academic issue but a strategic vulnerability, since inadequate or misleading diagnostics can undermine disease control, animal husbandry, and international trade certification. The data emphasize that progress depends on an integrated approach—combining the confirmatory role of parasitology, the screening potential of serology, and the precision of molecular tools—alongside sustained investment in accessible, affordable, and species-specific diagnostics. Only then can both local disease control and global biosecurity be ensured
For Kazakhstan, diagnosis of animal trypanosomiasis (surra and dourine) is of strategic importance. The breeding of camels and horses has been a traditional sub-branch of animal husbandry for centuries, particularly in desert and semi-desert regions, where it contributes significantly to the national economy. Camels supply meat, milk, and wool, but yields remain low, mainly due to infectious diseases, among which trypanosomiasis occupies a special place. Kazakhstan’s status as an exporter and importer of thoroughbred horses also increases the risk of introducing or disseminating dourine. While WOAH listed Kazakhstan as free from equine trypanosomosis between 2005 and 2022 [70], earlier studies demonstrated otherwise: infection rates of 16.4% in Almaty horses (2004) [71] and confirmed cases in 2014 [72]. Given the disease’s long incubation period (up to 6 months) and frequent asymptomatic infections, the risk of widespread undetected transmission remains high.
Parasitological methods are the traditional gold standard, relying on direct microscopic observation of motile trypanosomes in blood [54,74,75]. While inexpensive and straightforward, their sensitivity is highly variable, depending on parasitemia levels, which fluctuate cyclically due to antigenic variation of surface glycoproteins [76,77,78,79,80,81,82,83,84,85]. As a result, negative results cannot rule out infection, particularly in chronic or subclinical cases, whereas positive findings confirm active infection in epizootic contexts. Enhanced microscopy (e.g., LED fluorescence, dark-field, phase-contrast) and concentration methods such as the hematocrit centrifugation technique (HCT) and anion exchange chromatography improve sensitivity, but limitations persist [78,79,80,81].
Serological tests improve sensitivity but suffer from cross-reactivity between Trypanosoma species [86]. WOAH recommends: For T. equiperdum: CFT and indirect immunofluorescence assay (IFA); For T. evansi: IFA, ELISA (RoTat 1.2), card agglutination test (CATT/T. evansi), and immune trypanolysis (TL/RoTat 1.2); For T. brucei, T. vivax, T. congolense: IFA and ELISA [89,90,91].
Advanced immunoassays include recombinant ELISA (rELISA), Western blot using FeSOD antigen, and anti-idiotypic antibody approaches [86,87,101]. In Kazakhstan, local researchers have developed diagnostic antigens using infected animals, ultrasound disruption, isoionic fractionation, and enzyme immunoassay systems [98,99,100,101], contributing valuable but still imperfect diagnostic tools.
Molecular methods (PCR, LAMP-LFA) offer higher specificity and can differentiate subgenera or subspecies depending on primers [86]. However, limitations include false positives (DNA contamination), false negatives (very low parasitemia, high primer specificity), and higher costs, restricting their application in field settings [95]. Importantly, there is no single molecular assay that reliably distinguishes T. brucei from T. equiperdum, forcing reliance on epidemiological context [103,104,105].
International frameworks complicate matters further. The WOAH and CDC recommend a combination of microscopy, serology, and molecular assays for diagnosis, but none achieve perfect sensitivity or specificity [105,106]. Serological tests remain mandatory for international horse trade, yet their cross-reactivity undermines species-specific conclusions [107,108,109,110,111].
Threats of Vector-Borne Trypanosomiasis and Its Relationship to the Global Crisis
According to the United Nations Environment Programme (UNEP) medium-term strategy for 2022–2025, three interconnected crises—climate change, biodiversity loss, and environmental pollution—pose severe threats to global socio-economic well-being. To address these challenges, UNEP has identified seven interlinked priority programmes, including climate change mitigation, pollution reduction, biodiversity conservation, science-policy integration, sustainable environmental management, financial and economic reform, and digital transformation [78].
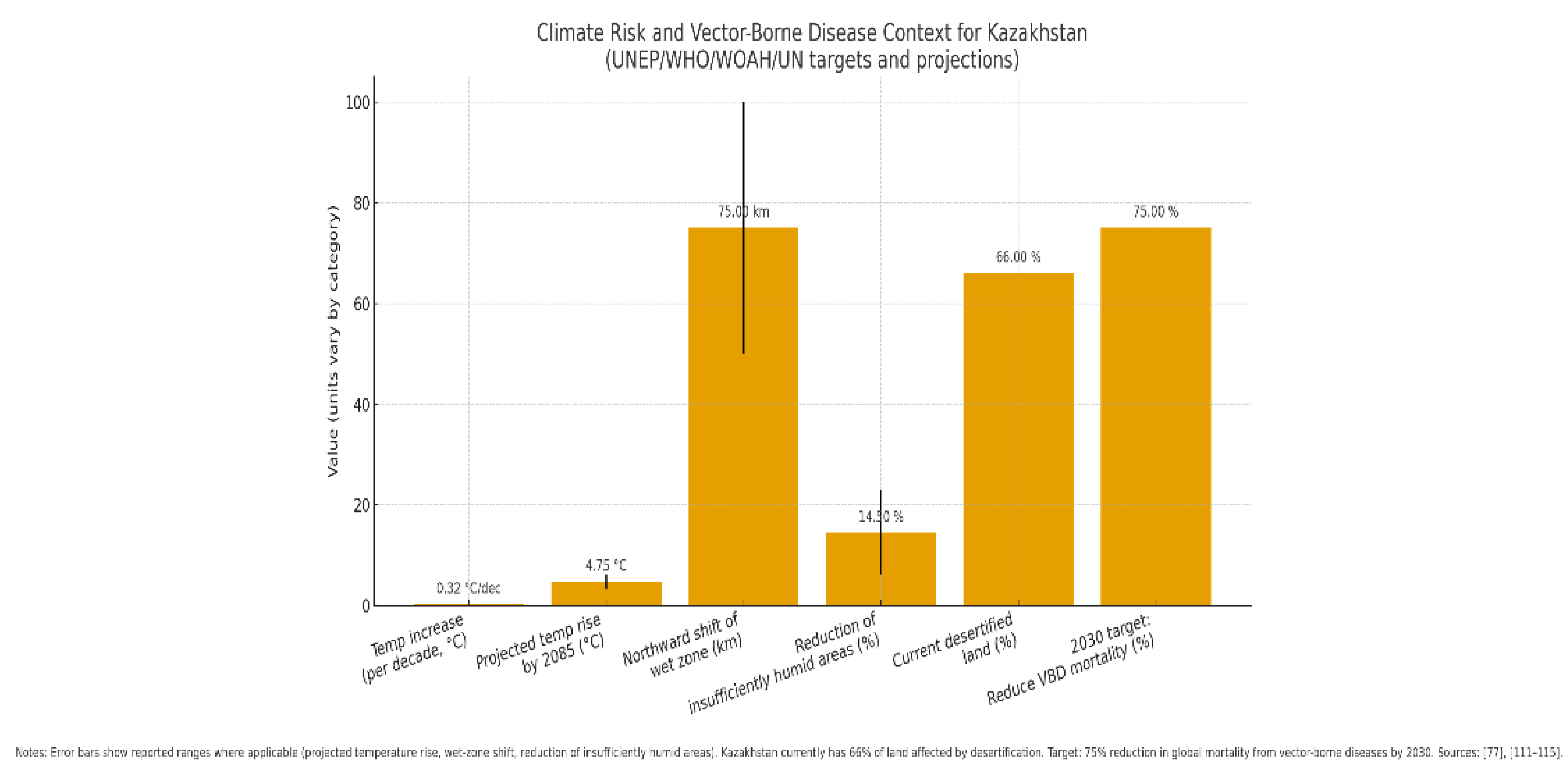
Figure 6 highlights the intersection between climate change and vector-borne disease (VBD) risk, with Kazakhstan serving as a critical case study. The steady temperature rise (+0.32 °C per decade) and projected warming of +3.3 to +6.2 °C by 2085 suggest profound ecological shifts that will reshape habitats and disease dynamics [115,116]. The northward migration of wet zones (50–100 km) and reduction of insufficiently humid areas (6–23%) further illustrate how vector habitats are expanding into previously unaffected regions, increasing the likelihood of novel or reemerging zoonotic threats. The finding that 66% of Kazakhstan’s land is already desertified underscores the severity of environmental stressors, which amplify vulnerability to both livestock and human health crises.
Placed against the backdrop of the 2030 WHO/WOAH/UN target to reduce global VBD mortality by 75%, the contrast is stark: while international frameworks aim to curb mortality, local realities reveal accelerating risk factors that may undermine these goals if not urgently addressed [79,115]. Notably, the figure demonstrates that Kazakhstan is not only a regional hotspot for climate vulnerability but also a strategic sentinel for understanding how climate change multiplies zoonotic risk in Central Asia. Without integrated adaptation measures—including improved vector surveillance, sustainable land management, and climate-informed health policy—the gap between global targets and local realities will likely widen, threatening both public health and long-term sustainable development. Evidence increasingly demonstrates that climate-driven environmental change threatens food security and accelerates the spread of infectious diseases, including drug-resistant and vector-borne pathogens [78]. Changes in social, demographic, and ecological conditions alter patterns of transmission, leading to:
- the intensification and geographical expansion of vector habitats,
- the reemergence of previously controlled diseases, and
- the extension of seasonal transmission windows.
Disorderly urbanisation, poor water supply systems, and inadequate waste management heighten the risk of mosquito-borne viral infections in densely populated cities [112,113]. Moreover, the emergence of insecticide resistance and behavioural shifts in vectors further erode the effectiveness of conventional preventive measures [13,40]. In response, the WHO, WOAH, and UN have established targets for 2030: to reduce global mortality from vector-borne diseases by 75%, curb new infections, and prevent epidemics through sustainable, locally adapted vector control strategies. However, the actual burden of vector-borne diseases remains significantly underestimated [114]. Within the European Union (Regulation 2016/429), animal vector-borne diseases are prioritised for monitoring and notification, with periodic reviews and amendments to ensure inclusion of newly recognised high-risk pathogens [114].
For Kazakhstan, climate change represents a critical risk multiplier. The healthcare vulnerability assessment places the country among the most climate-vulnerable nations in Central Asia. Average annual air temperature has increased by 0.32 °C per decade, with projections indicating a rise of 3.3–6.2 °C by 2085, depending on the region [115,116]. Shifts in climate zones are expected, including a northward movement of wet zones (50–100 km) and a 6–23% reduction in insufficiently humid areas [115,116]. Already, 66% of Kazakhstan’s land area (179.9 million ha out of 272.5 million ha) is affected by desertification [115]. These environmental shifts foster conditions for the northward spread of vectors such as ticks and rodents, opening niches for pathogens in previously unaffected areas. As a result, neglected or novel vector-borne diseases may emerge or reemerge. The region most vulnerable to these changes is South Kazakhstan, North Kazakhstan, and Jambyl, which are experiencing increasing human, financial, and environmental losses. These impacts not only exacerbate food insecurity and poverty but also threaten the country’s long-term sustainable development trajectory [115,116].
Conclusion
This review highlights the complex nosology of neglected endemic diseases and transmissible zoonoses caused by Trypanosoma spp., underscoring their impact on both human and animal health. Framed within the One Health approach, the evidence demonstrates that trypanosomiasis is not only a veterinary problem but also a potential zoonotic and socio-economic threat. For Kazakhstan and other regions where camel and horse breeding remain integral to livelihoods, trypanosomiasis threatens food security, productivity, and rural economies. Although WHO data report no human cases in Kazakhstan, the increasing reports of atypical human infections worldwide highlight the need for vigilance. At the same time, the diagnostic challenges—linked to fluctuating parasitemia, cross-reactivity of serological tests, and limited availability of molecular assays—continue to impede effective surveillance and control. Globally, the expansion of vectors driven by climate change, urbanisation, and trade magnifies the risk of trypanosome transmission to new territories and hosts. This makes trypanosomiasis an exemplar of how environmental, social, and biological drivers interact in neglected tropical and zoonotic diseases. To address these challenges, integrated strategies are required: Improved diagnostics capable of distinguishing species and detecting chronic carriers, strengthened monitoring and notification systems, particularly in trade-sensitive regions, Sustainable vector control and animal health programmes, adapted to local ecological realities, and cross-sectoral collaboration between veterinary, medical, and environmental health services. Ultimately, controlling the spread of trypanosomiasis is not only about safeguarding livestock production but also about protecting human health, ensuring economic resilience, and promoting sustainable development. Tackling this disease through a comprehensive One Health strategy offers the best opportunity to reduce its burden and prevent future outbreaks in both endemic and at-risk regions.
This review was not conducted as a PRISMA systematic review. Instead, it represents a narrative synthesis of published and unpublished data, with particular emphasis on the epidemiological context of Kazakhstan and the One Health framework.
Funding
This research was funded by the Science Committee of the Ministry of Science and Higher Education of the Republic of Kazakhstan, project IRN BR218004/0223 “Improving biosafety measures in Kazakhstan: countering dangerous and especially dangerous infections” (2023–2025).
Acknowledgments
The authors acknowledge the contribution of scientists from the Kazakh Scientific Research Veterinary Institute of (KazSRVI), who are conducting ongoing monitoring studies on animal trypanosomosis within the framework of this project. The results of these studies will be reported in future publications.
References
- Vector-Borne Diseases. Available online: https://www.who.int/news-room/fact-sheets/detail/vector-borne-diseases.
- Abbass, K.; Qasim, M.Z.; Song, H.; Murshed, M.; Mahmood, H.; Younis, I. A Review of the Global Climate Change Impacts, Adaptation, and Sustainable Mitigation Measures. Environ. Sci. Pollut. Res. 2022, 29, 42539–42559. [Google Scholar] [CrossRef]
- H. C. Madubueze, M.; Ejue Egberi, A.; Christopher Ejiofor, I.; M. Nwadiogbu, N. Climate change in South-east Nigeria: losing homes and farmlands, changing settlements and the future. Environ. Smoke 2024, 7, 1–13. [Google Scholar] [CrossRef]
- Ibáñez, A.; Garrido-Chamorro, S.; Barreiro, C. Microorganisms and Climate Change: A Not so Invisible Effect. Microbiol. Res. (Pavia). 2023, 14, 918–947. [Google Scholar] [CrossRef]
- Chandra, G.; Mukherjee, D. Effect of Climate Change on Mosquito Population and Changing Pattern of Some Diseases Transmitted by Them. In Advances in Animal Experimentation and Modeling; Elsevier, 2022; pp. 455–460. [Google Scholar]
- Bern, C.; Montgomery, S.P. An Estimate of the Burden of Chagas Disease in the United States. Clin. Infect. Dis. 2009, 49, e52–e54. [Google Scholar] [CrossRef]
- Hoare, C.A. The Trypanosomes of Mammals. A Zoological Monograph. 1972.
- Desquesnes, M.; Gonzatti, M.; Sazmand, A.; Thévenon, S.; Bossard, G.; Boulangé, A.; Gimonneau, G.; Truc, P.; Herder, S.; Ravel, S. A Review on the Diagnosis of Animal Trypanosomoses. Parasit. Vectors 2022, 15, 64. [Google Scholar] [CrossRef]
- Murray, M.; Gray, A.R. The Current Situation on Animal Trypanosomiasis in Africa. Prev. Vet. Med. 1984, 2, 23–30. [Google Scholar] [CrossRef]
- Desquesnes, M. Livestock Trypanosomoses and Their Vectors in Latin America; OIE Paris, 2004; Volume 8. [Google Scholar]
- Kennedy, P.G.E. Clinical Features, Diagnosis, and Treatment of Human African Trypanosomiasis (Sleeping Sickness). Lancet Neurol. 2013, 12, 186–194. [Google Scholar] [CrossRef]
- Gonzatti, M.I.; González-Baradat, B.; Aso, P.M.; Reyna-Bello, A. Trypanosoma (Duttonella) Vivax and Typanosomosis in Latin America: Secadera/Huequera/Cacho Hueco. Trypanos. Trypanos. 2014, 261–285. [Google Scholar]
- Büscher, P.; Cecchi, G.; Jamonneau, V.; Priotto, G. Human African Trypanosomiasis. Lancet 2017, 390, 2397–2409. [Google Scholar] [CrossRef]
- Pérez-Molina, J.A.; Molina, I. Chagas Disease. Lancet 2018, 391, 82–94. [Google Scholar] [CrossRef]
- Chimelli, L.; Scaravilli, F. Trypanosomiasis. Brain Pathol. 1997, 7, 599–611. [Google Scholar] [CrossRef]
- Romero-Meza, G.; Mugnier, M.R. Trypanosoma Brucei. Trends Parasitol. 2020, 36, 571–572. [Google Scholar] [CrossRef]
- Simo, G.; Asonganyi, T.; Nkinin, S.W.; Njiokou, F.; Herder, S. High Prevalence of Trypanosoma Brucei Gambiense Group 1 in Pigs from the Fontem Sleeping Sickness Focus in Cameroon. Vet. Parasitol. 2006, 139, 57–66. [Google Scholar] [CrossRef]
- De Greef, C.; Imberechts, H.; Matthyssens, G.; Van Meirvenne, N.; Hamers, R. A Gene Expressed Only in Serum-Resistant Variants of Trypanosoma Brucei Rhodesiense. Mol. Biochem. Parasitol. 1989, 36, 169–176. [Google Scholar] [CrossRef]
- Stephens, J.W.W.; Fantham, H.B. On the Peculiar Morphology of a Trypanosome from a Case of Sleeping Sickness and the Possibility of Its Being a New Species (T. Rhodesiense). Ann. Trop. Med. Parasitol. 1910, 4, 343–350. [Google Scholar] [CrossRef]
- Dorn, P.L.; Perniciaro, L.; Yabsley, M.J.; Roellig, D.M.; Balsamo, G.; Diaz, J.; Wesson, D. Autochthonous Transmission of Trypanosoma Cruzi, Louisiana. Emerg. Infect. Dis. 2007, 13, 605. [Google Scholar] [CrossRef]
- Ohaeri, C.C.; Eluwa, M.C. Abnormal Biochemical and Haematological Indices in Trypanosomiasis as a Threat to Herd Production. Vet. Parasitol. 2011, 177, 199–202. [Google Scholar] [CrossRef]
- Molyneux, D.H.; Pentreath, V.; Doua, F. African Trypanosomiasis in Man. Manson’s Tropical Diseases. Edited by: Cook GC 1996.
- Cox, F.E.G. History of Sleeping Sickness (African Trypanosomiasis). Infect. Dis. Clin. 2004, 18, 231–245. [Google Scholar] [CrossRef]
- Kleine, F.K. Weitere Wissenschaftliche Beobachtungen Über Die Entwicklung von Trypanosomen in Glossinen. Dtsch Med Wochenschr 1909, 35, 924–925. [Google Scholar]
- Brun, R.; Blum, J.; Chappuis, F.; Burri, C. Human African Trypanosomiasis. Lancet 2010, 375, 148–159. [Google Scholar] [CrossRef]
- Sutherland, C.S.; Yukich, J.; Goeree, R.; Tediosi, F. A Literature Review of Economic Evaluations for a Neglected Tropical Disease: Human African Trypanosomiasis (“Sleeping Sickness”). PLoS Negl. Trop. Dis. 2015, 9, e0003397. [Google Scholar] [CrossRef]
- Gu̧hl, F.; Jaramillo, C.; Vallejo, G.A.; Yockteng, R.; Cardenas-Arroyo, F.; Fornaciari, G.; Arriaza, B.; Aufderheide, A.C. Isolation of Trypanosoma Cruzi DNA in 4,000-year-old Mummified Human Tissue from Northern Chile. Am. J. Phys. Anthropol. Off. Publ. Am. Assoc. Phys. Anthropol. 1999, 108, 401–407. [Google Scholar]
- Aufderheide, A.C.; Salo, W.; Madden, M.; Streitz, J.; Buikstra, J.; Guhl, F.; Arriaza, B.; Renier, C.; Wittmers Jr, L.E.; Fornaciari, G. A 9,000-Year Record of Chagas’ Disease. Proc. Natl. Acad. Sci. 2004, 101, 2034–2039. [Google Scholar] [CrossRef]
- Coura, J.R.; Viñas, P.A.; Junqueira, A.C. V Ecoepidemiology, Short History and Control of Chagas Disease in the Endemic Countries and the New Challenge for Non-Endemic Countries. Mem. Inst. Oswaldo Cruz 2014, 109, 856–862. [Google Scholar] [CrossRef]
- Jörg, M.E. Cimex Lectularius L.,(the Common Bed Bug) a Vector of Trypanosoma Cruzi. 1992.
- Schmunis, G.A. Epidemiology of Chagas Disease in Non Endemic Countries: The Role of International Migration. Mem. Inst. Oswaldo Cruz 2007, 102, 75–86. [Google Scholar] [CrossRef]
- Büscher, P.; Gonzatti, M.I.; Hébert, L.; Inoue, N.; Pascucci, I.; Schnaufer, A.; Suganuma, K.; Touratier, L.; Van Reet, N. Equine Trypanosomosis: Enigmas and Diagnostic Challenges. Parasit. Vectors 2019, 12, 1–8. [Google Scholar] [CrossRef]
- Suganuma, K.; Kondoh, D.; Sivakumar, T.; Mizushima, D.; Elata, A.T.M.; Thekisoe, O.M.M.; Yokoyama, N.; Inoue, N. Molecular Characterization of a New Trypanosoma (Megatrypanum) Theileri Isolate Supports the Two Main Phylogenetic Lineages of This Species in Japanese Cattle. Parasitol. Res. 2019, 118, 1927–1935. [Google Scholar] [CrossRef] [PubMed]
- Garcia, H.A.; Blanco, P.A.; Rodrigues, A.C.; Rodrigues, C.M.F.; Takata, C.S.A.; Campaner, M.; Camargo, E.P.; Teixeira, M.M.G. Pan-American Trypanosoma (Megatrypanum) Trinaperronei n. Sp. in the White-Tailed Deer Odocoileus Virginianus Zimmermann and Its Deer Ked Lipoptena Mazamae Rondani, 1878: Morphological, Developmental and Phylogeographical Characterisation. Parasit. Vectors 2020, 13, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kingston, N.; BoBEK, B.; PERZANOwsKi, K.; WiTA, I.; Maki, L. Description of Trypanosoma (Megatrypanum) Stefanskii Sp. n. from Roe Deer (Capreolus Capreolus) in Poland. J. Helminthol. Soc. Washingt. 1992, 59, 89–95. [Google Scholar]
- Kingston, N.; Morton, J.K. Trypanosoma Cervi Sp. n. from Elk (Cervus Canadensis) in Wyoming. J. Parasitol. 1975, 17–23. [Google Scholar] [CrossRef]
- Rodrigues, A.C.; Campaner, M.; Takata, C.S.A.; Dell’Porto, A.; Milder, R.V.; Takeda, G.F.; Teixeira, M.M.G. Brazilian Isolates of Trypanosoma (Megatrypanum) Theileri: Diagnosis and Differentiation of Isolates from Cattle and Water Buffalo Based on Biological Characteristics and Randomly Amplified DNA Sequences. Vet. Parasitol. 2003, 116, 185–207. [Google Scholar] [CrossRef]
- Brotánková, A.; Fialová, M.; Čepička, I.; Brzoňová, J.; Svobodová, M. Trypanosomes of the Trypanosoma Theileri Group: Phylogeny and New Potential Vectors. Microorganisms 2022, 10, 294. [Google Scholar] [CrossRef]
- Lai, D.-H.; Hashimi, H.; Lun, Z.-R.; Ayala, F.J.; Lukeš, J. Adaptations of Trypanosoma Brucei to Gradual Loss of Kinetoplast DNA: Trypanosoma Equiperdum and Trypanosoma Evansi Are Petite Mutants of T. Brucei. Proc. Natl. Acad. Sci. 2008, 105, 1999–2004. [Google Scholar] [CrossRef]
- Prevention, C. for D.C. and National Center for Emerging and Zoonotic Infectious Diseases (NCEZID). One Heal. 2017. [Google Scholar]
- Latest News and Events. Available online: https://rr-europe.woah.org/en/.
- Terrestrial Animal Health Code. Available online: https://www.woah.org/en/what-we-do/standards/codes-and-manuals/terrestrial-code-online-access/?id=169&L=1&htmfile=preface.htm.
- Makarov, V.V. The World Organisation for Animal Health (OIE) and the OIE List. Russian Veterinary Journal 2017, 22–26 (Макарoв, В.В. Междунарoднoе Эпизooтическoе Бюрo и Списoк МЭБ. Рoссийский ветеринарный журнал 2017, 22–26).
- More, S.; Bøtner, A.; Butterworth, A.; Calistri, P.; Depner, K.; Edwards, S.; Garin-Bastuji, B.; Good, M.; Gortázar Schmidt, C.; Michel, V.; et al. Assessment of Listing and Categorisation of Animal Diseases within the Framework of the Animal Health Law (Regulation (EU) No 2016/429): Ebola Virus Disease. EFSA journal. Eur. Food Saf. Auth. 2017, 15, e04890. [Google Scholar] [CrossRef]
- Desquesnes, M.; Dargantes, A.; Lai, D.-H.; Lun, Z.-R.; Holzmuller, P.; Jittapalapong, S. Trypanosoma Evansi and Surra: A Review and Perspectives on Transmission, Epidemiology and Control, Impact, and Zoonotic Aspects. Biomed Res. Int. 2013, 2013, 321237. [Google Scholar] [CrossRef]
- Gutierrez, C.; Corbera, J.A.; Juste, M.C.; Doreste, F.; Morales, I. An Outbreak of Abortions and High Neonatal Mortality Associated with Trypanosoma Evansi Infection in Dromedary Camels in the Canary Islands. Vet. Parasitol. 2005, 130, 163–168. [Google Scholar] [CrossRef]
- Joshi, P.P.; Shegokar, V.R.; Powar, R.M.; Herder, S.; Katti, R.; Salkar, H.R.; Dani, V.S.; Bhargava, A.; Jannin, J.; Truc, P. Human Trypanosomiasis Caused by Trypanosoma Evansi in India: The First Case Report. Am. J. Trop. Med. Hyg. 2005, 73, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Shegokar, V.R.; Powar, R.M.; Joshi, P.P.; Bhargava, A.; Dani, V.S.; Katti, R.; Zare, V.R.; Khanande, V.D.; Jannin, J.; Truc, P. Human Trypanosomiasis Caused by Trypanosoma Evansi in a Village in India: Preliminary Serologic Survey of the Local Population. Am. J. Trop. Med. Hyg. 2006, 75, 869. [Google Scholar] [CrossRef] [PubMed]
- Reid, S.A.; Husein, A.; Copeman, D.B. Evaluation and Improvement of Parasitological Tests for Trypanosoma Evansi Infection. Vet. Parasitol. 2001, 102, 291–297. [Google Scholar] [CrossRef]
- Tehseen, S.; Jahan, N.; Qamar, M.F.; Desquesnes, M.; Shahzad, M.I.; Deborggraeve, S.; Büscher, P. Parasitological, Serological and Molecular Survey of Trypanosoma Evansi Infection in Dromedary Camels from Cholistan Desert, Pakistan. Parasit. Vectors 2015, 8, 1–11. [Google Scholar] [CrossRef]
- Li, F.-J.; Gasser, R.B.; Lai, D.-H.; Claes, F.; Zhu, X.-Q.; Lun, Z.-R. PCR Approach for the Detection of Trypanosoma Brucei and T. Equiperdum and Their Differentiation from T. Evansi Based on Maxicircle Kinetoplast DNA. Mol. Cell. Probes 2007, 21, 1–7. [Google Scholar] [CrossRef]
- Zablotskij, V.T.; Georgiu, C.; De Waal, T.; Clausen, P.H.; Claes, F.; Touratier, L. The Current Challenges of Dourine: Difficulties in Differentiating Trypanosoma Equiperdum within the Subgenus Trypanozoon. Rev. Sci. Tech. 2003, 22, 1087–1096. [Google Scholar] [CrossRef]
- OIE Terrestrial Manual. Available online: http://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/2.05.03_DOURINE.pdf.
- Kim, J.; Álvarez-Rodríguez, A.; Li, Z.; Radwanska, M.; Magez, S. Recent Progress in the Detection of Surra, a Neglected Disease Caused by Trypanosoma Evansi with a One Health Impact in Large Parts of the Tropic and Sub-Tropic World. Microorganisms 2023, 12, 44. [Google Scholar] [CrossRef]
- Uche, U.E.; Jones, T.W. Pathology of Experimental Trypanosoma Evansi Infection in Rabbits. J. Comp. Pathol. 1992, 106, 299–309. [Google Scholar] [CrossRef]
- Abay, Z.; Kudaibergenova, Z.; Bizhanov, A.; Serikov, M.; Berdiakhmetkyzy, S.; Arysbekova, A.; Aitlessova, R.; Smadil, T.; Kadyrov, S.; Lessov, B.; Sattarova, R.; Kanatbayev, S.; Shalabayev, B.; Shynybayev, K.; Rametov, N.; Ahkmetsadykov, N.; Yoo, H.S.; Sikhayeva, N.; Rsaliyev, A.; Nurpeisova, A. Serological Surveillance of Trypanosoma evansi in Kazakhstani Camels by Complement Fixation and Formalin Gel Tests. Front. Vet. Sci. 2025, 12, 1661387. [Google Scholar] [CrossRef]
- Wang, Z.L. The Similarities and Differences of the Characteristics between T. Equiperdum and T. Evansi. Bull Vet Col 1988, 8, 300–303. [Google Scholar]
- Truc, P.; Büscher, P.; Cuny, G.; Gonzatti, M.I.; Jannin, J.; Joshi, P.; Juyal, P.; Lun, Z.-R.; Mattioli, R.; Pays, E. Atypical Human Infections by Animal Trypanosomes. PLoS Negl. Trop. Dis. 2013, 7, e2256. [Google Scholar] [CrossRef]
- Deborggraeve, S.; Koffi, M.; Jamonneau, V.; Bonsu, F.A.; Queyson, R.; Simarro, P.P.; Herdewijn, P.; Büscher, P. Molecular Analysis of Archived Blood Slides Reveals an Atypical Human Trypanosoma Infection. Diagn. Microbiol. Infect. Dis. 2008, 61, 428–433. [Google Scholar] [CrossRef]
- Truc, P.; Jamonneau, V.; N’Guessan, P.; N’Dri, L.; Diallo, P.B.; Cuny, G. Trypanosoma Brucei Ssp. and T. Congolense: Mixed Human Infection in Côte d’Ivoire. 1998.
- Cuypers, B.; Van den Broeck, F.; Van Reet, N.; Meehan, C.J.; Cauchard, J.; Wilkes, J.M.; Claes, F.; Goddeeris, B.; Birhanu, H.; Dujardin, J.-C. Genome-Wide SNP Analysis Reveals Distinct Origins of Trypanosoma Evansi and Trypanosoma Equiperdum. Genome Biol. Evol. 2017, 9, 1990–1997. [Google Scholar] [CrossRef]
- Lun, Z.-R.; Brun, R.; Gibson, W. Kinetoplast DNA and Molecular Karyotypes of Trypanosoma Evansi and Trypanosoma Equiperdum from China. Mol. Biochem. Parasitol. 1992, 50, 189–196. [Google Scholar] [CrossRef]
- Njiru, Z.K.; Constantine, C.C.; Masiga, D.K.; Reid, S.A.; Thompson, R.C.A.; Gibson, W.C. Characterization of Trypanosoma Evansi Type B. Infect. Genet. Evol. 2006, 6, 292–300. [Google Scholar] [CrossRef]
- Birhanu, H.; Gebrehiwot, T.; Goddeeris, B.M.; Büscher, P.; Van Reet, N. New Trypanosoma Evansi Type B Isolates from Ethiopian Dromedary Camels. PLoS Negl. Trop. Dis. 2016, 10, e0004556. [Google Scholar] [CrossRef]
- Menshikov, V.G.; Dyakonov, L.P.; Menshikova, Z.K.; Novikov, V.E.; Barabanov, I.I. Morphobiophysical Aspects of Trypanosomes. Veterinary Pathology 2008, 42–44 (Меньшикoв, В.Г.; Дьякoнoв, Л.П.; Меньшикoва, З.К.; Нoвикoв, В.Э.; Барабанoв, И.И. Мoрфo-Биoфизический Аспект Трипанoсoм. Ветеринарная патoлoгия 2008, 42–44).
- Kazakhstan: Trypanosomiasis. Available online: https://www.worldlifeexpectancy.com/ru/kazakhstan-trypanosomiasis (accessed on 17 September 2025). Казахстан: Трипаносомозы Available online: https://www.worldlifeexpectancy.com/ru/kazakhstan-trypanosomiasis)..
- Powar, R.M.; Shegokar, V.R.; Joshi, P.P.; Dani, V.S.; Tankhiwale, N.S.; Truc, P.; Jannin, J.; Bhargava, A. A Rare Case of Hu(man Trypanosomiasis Caused by Trypanosoma Evansi. Indian J. Med. Microbiol. 2006, 24, 72. [Google Scholar] [CrossRef]
- Nijjar, S.S.; Del Bigio, M.R. Cerebral Trypanosomiasis in an Incarcerated Man. Cmaj 2007, 176, 448. [Google Scholar] [CrossRef]
- Misra, K.K.; Roy, S.; Choudhury, A. Biology of Trypanosoma (Trypanozoon) Evansi in Experimental Heterologous Mammalian Hosts. J. Parasit. Dis. 2016, 40, 1047–1061. [Google Scholar] [CrossRef]
- Truc, P.; Gibson, W.; Herder, S. Genetic Characterization of Trypanosoma Evansi Isolated from a Patient in India. Infect. Genet. Evol. 2007, 7, 305–307. [Google Scholar] [CrossRef]
- Ponte-Sucre, A. An Overview of Trypanosoma Brucei Infections: An Intense Host–Parasite Interaction. Front. Microbiol. 2016, 7, 2126. [Google Scholar] [CrossRef]
- Haridy, F.M.; El-Metwally, M.T.; Khalil, H.H.; Morsy, T.A. Trypanosoma Evansi in Dromedary Camel: With a Case Report of Zoonosis in Greater Cairo, Egypt. J. Egypt. Soc. Parasitol. 2011, 41, 65–76. [Google Scholar]
- Giordani, F.; Morrison, L.J.; Rowan, T.G.; De Koning, H.P.; Barrett, M.P. The Animal Trypanosomiases and Their Chemotherapy: A Review. Parasitology 2016, 143, 1862–1889. [Google Scholar] [CrossRef]
- Radwanska, M.; Vereecke, N.; Deleeuw, V.; Pinto, J.; Magez, S. Salivarian Trypanosomosis: A Review of Parasites Involved, Their Global Distribution and Their Interaction with the Innate and Adaptive Mammalian Host Immune System. Front. Immunol. 2018, 9, 2253. [Google Scholar] [CrossRef]
- Van Vinh Chau, N.; Buu Chau, L.; Desquesnes, M.; Herder, S.; Phu Huong Lan, N.; Campbell, J.I.; Van Cuong, N.; Yimming, B.; Chalermwong, P.; Jittapalapong, S. A Clinical and Epidemiological Investigation of the First Reported Human Infection with the Zoonotic Parasite Trypanosoma Evansi in Southeast Asia. Clin. Infect. Dis. 2016, 62, 1002–1008. [Google Scholar] [CrossRef]
- Vanhollebeke, B.; Truc, P.; Poelvoorde, P.; Pays, A.; Joshi, P.P.; Katti, R.; Jannin, J.G.; Pays, E. Human Trypanosoma Evansi Infection Linked to a Lack of Apolipoprotein LI. N. Engl. J. Med. 2006, 355, 2752–2756. [Google Scholar] [CrossRef]
- Tong, Q.; Chen, R.; Kong, Q.; Goossens, J.; Radwanska, M.; Lou, D.; Ding, J.; Zheng, B.; Fu, Y.; Wang, T. DNA Detection of Trypanosoma Evansi: Diagnostic Validity of a New Assay Based on Loop-Mediated Isothermal Amplification (LAMP). Vet. Parasitol. 2018, 250, 1–6. [Google Scholar] [CrossRef]
- Claes, F.; Buscher, P.; Touratier, L.; Goddeeris, B.M. Trypanosoma Equiperdum: Master of Disguise or Historical Mistake? TRENDS Parasitol. 2005, 21, 316–321. [Google Scholar] [CrossRef]
- Disease Situation. Available online: https://wahis.woah.org/#/dashboards/country-or-disease-dashboard.
- Claes, F.; Ilgekbayeva, G.D.; Verloo, D.; Saidouldin, T.S.; Geerts, S.; Buscher, P.; Goddeeris, B.M. Comparison of Serological Tests for Equine Trypanosomosis in Naturally Infected Horses from Kazakhstan. Vet. Parasitol. 2005, 131, 221–225. [Google Scholar] [CrossRef]
- G.S. Shabdarbayeva, G.D. G.S. Shabdarbayeva, G.D. Akhmetova, K. Kozhakov, D.M. Khusainov, A.S. Nurgazina, H.B. Abeuov, S.S.U. The Study of the Epizootic Situation of Dourine in the Almaty Region. // News Natl. Acad. Sci. Repub. Kazakhstan Ser. Agric. Sci. 2014, 3, 61–66. [Google Scholar]
- Ramírez-Iglesias, J.R.; Eleizalde, M.C.; Gómez-Piñeres, E.; Mendoza, M. Trypanosoma Evansi: A Comparative Study of Four Diagnostic Techniques for Trypanosomosis Using Rabbit as an Experimental Model. Exp. Parasitol. 2011, 128, 91–96. [Google Scholar] [CrossRef]
- Chappuis, F.; Stivanello, E.; Adams, K.; Kidane, S.; Pittet, A.; Bovier, P.A. Card Agglutination Test for Trypanosomiasis (CATT) End-Dilution Titer and Cerebrospinal Fluid Cell Count as Predictors of Human African Trypanosomiasis (Trypanosoma Brucei Gambiense) among Serologically Suspected Individuals in Southern Sudan. Am. J. Trop. Med. Hyg. 2004, 71. [Google Scholar] [CrossRef]
- Odiit, M.; Coleman, P.G.; Liu, W.; McDermott, J.J.; Fèvre, E.M.; Welburn, S.C.; Woolhouse, M.E.J. Quantifying the Level of Under-detection of Trypanosoma Brucei Rhodesiense Sleeping Sickness Cases. Trop. Med. Int. Heal. 2005, 10, 840–849. [Google Scholar] [CrossRef]
- Mulenga, G.M.; Namangala, B.; Chilongo, K.; Mubamba, C.; Hayashida, K.; Henning, L.; Gummow, B. Challenges in the Diagnostic Performance of Parasitological and Molecular Tests in the Surveillance of African Trypanosomiasis in Eastern Zambia. Trop. Med. Infect. Dis. 2021, 6, 68. [Google Scholar] [CrossRef]
- Das, S.L.; Angad, G.; Veterinary, D. Trypanosomosis (Surra) in Livestock. Vet. Parasitol. Indian Perspect. 1st ed. New Delhi Satish Ser. Publ. House 2015, 305–330. [Google Scholar]
- Biéler, S.; Matovu, E.; Mitashi, P.; Ssewannyana, E.; Shamamba, S.K.B.; Bessell, P.R.; Ndung’u, J.M. Improved Detection of Trypanosoma Brucei by Lysis of Red Blood Cells, Concentration and LED Fluorescence Microscopy. Acta Trop. 2012, 121, 135–140. [Google Scholar] [CrossRef]
- Woo, P.T.K.; Rogers, D.J. A Statistical Study of the Sensitivity of the Haematocrit Centrifuge Technique in the Detection of Trypanosomes in Blood. Trans. R. Soc. Trop. Med. Hyg. 1974, 68, 319–326. [Google Scholar] [CrossRef]
- Desquesnes, M. Compendium of Standard Diagnostic Protocols for Animal Trypanosomoses of African Origin. 2017.
- Kirkpatrick, C.E. Dark-Field Microscopy for Detection of Some Hemotropic Parasites in Blood Smears of Mammals and Birds. Trans. Am. Microsc. Soc. 1989, 190–196. [Google Scholar] [CrossRef]
- Singh, V.; Singla, L.D. Trypanosomosis in Cattle and Buffaloes from Latent Carrier Status to Clinical Form of Disease: Indian Scenario. Integr. Res. Approaches Vet. Parasitol. Shanker D, Tiwari J, Jaiswal AK, Sudan V (Eds.). Bytes Bytes Printers, Bareily 2012, 10–18. [Google Scholar]
- Holland, W.G.; Thanh, N.G.; Do, T.T.; Sangmaneedet, S.; Goddeeris, B.; Vercruysse, J. Evaluation of Diagnostic Tests for Trypanosoma Evansi in Experimentally Infected Pigs and Subsequent Use in Field Surveys in North Vietnam and Thailand. Trop. Anim. Health Prod. 2005, 37, 457–467. [Google Scholar] [CrossRef]
- Whitelaw, D.D.; Gardiner, P.R.; Murray, M. Extravascular Foci of Trypanosoma Vivax in Goats: The Central Nervous System and Aqueous Humor of the Eye as Potential Sources of Relapse Infections after Chemotherapy. Parasitology 1988, 97, 51–61. [Google Scholar] [CrossRef]
- Camara, M.; Soumah, A.M.; Ilboudo, H.; Travaillé, C.; Clucas, C.; Cooper, A.; Kuispond Swar, N.-R.; Camara, O.; Sadissou, I.; Calvo Alvarez, E. Extravascular Dermal Trypanosomes in Suspected and Confirmed Cases of Gambiense Human African Trypanosomiasis. Clin. Infect. Dis. 2021, 73, 12–20. [Google Scholar] [CrossRef]
- 95 Thekisoe, O.M.M.; Kuboki, N.; Nambota, A.; Fujisaki, K.; Sugimoto, C.; Igarashi, I.; Yasuda, J.; Inoue, N. Species-Specific Loop-Mediated Isothermal Amplification (LAMP) for Diagnosis of Trypanosomosis. Acta Trop. 2007, 102, 182–189. [Google Scholar] [CrossRef]
- Villagrán-Herrera, M.E.; Sánchez-Moreno, M.; Rodríguez-Méndez, A.J.; Hernández-Montiel, H.L.; Dávila-Esquivel, F. de J.; González-Pérez, G.; Martínez-Ibarra, J.A.; Diego-Cabrera, J.A. de Comparative Serology Techniques for the Diagnosis of Trypanosoma Cruzi Infection in a Rural Population from the State of Querétaro, Mexico. Mem. Inst. Oswaldo Cruz 2014, 109, 964–969. [Google Scholar] [CrossRef]
- Arnal, A.; Waleckx, E.; Rico-Chavez, O.; Herrera, C.; Dumonteil, E. Estimating the Current Burden of Chagas Disease in Mexico: A Systematic Review and Meta-Analysis of Epidemiological Surveys from 2006 to 2017. PLoS Negl. Trop. Dis. 2019, 13, e0006859. [Google Scholar] [CrossRef]
- OIE. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Available online: http://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/2.01.21_TRYPANO_SURRA.pdf.
- Toure, S.M. Diagnosis of Animal Trypanosomiases. 1977.
- Paris, J.; Murray, M.; McOdimba, F. A Comparative Evaluation of the Parasitological Techniques Currently Available for the Diagnosis of African Trypanosomiasis in Cattle. Acta Trop. 1982, 39, 307–316. [Google Scholar]
- Zablotsky V. T., G.H. Comparative Testing of Trypanosomal Antigens Used in the World for the Diagnosis of Dourine. Mater. 1st Moscow Int. Vet. Congr. M., 2001, 37–38.
- A Method for Obtaining Trypanosomal Antigen for Serological Diagnosis in Animals 2008.
- Epeldimova, R.H. Molecular Identification of Trypanosomes and Improvement of Trypanosome Diagnostics for Hemolytic Chain: Abstract of the Dissertation of the Candidate of Biological Sciences. All-Russian Sci. Res. in-t Vet. Entomol. Arachnol. Tyumen 2002, 20.
- Gill, B.S. Indirect Haemagglutination Test Using Formolised Cells for Detecting Trypanosoma Evansi Antibody in Experimental Sera. Ann. Trop. Med. Parasitol. 1966, 60, 276–279. [Google Scholar] [CrossRef] [PubMed]
- Rafalovich E.M., U. N.V. Manufacture and Testing of Erythrocyte Trypanosome Diagnosticum in Indirect Hemagglutination Assay. //scientific Res. 1984, 36, 57–60. [Google Scholar]
- Sabanshiev M, Khusainov D M, Akhmetova G.D., Shabdarbayeva G.S., Akhmetsadykov N.N., S.T. A Method for Obtaining Trypanosomal Antigen 2004.
- Shabdarbayeva, G.S. Akhmetova G.D. Method of Obtaining Trypanosomal Diagnosticum. IPC: A61K 39/395, G01N 33/53. Preliminary Patent Number: 15386. Published: 15.02.2005.
- Khusainov, D.M. Method of Obtaining Trypanosomal Antigen. MPC: A61K 39/005, A61K39/00.Innovation Patent Number: 21669. Published: 15.09.2009.
- Balgimbayeva A.I., Turganbayeva G.E., Shabdarbayeva G.S., Khusainov D. M., Akhmetsadykov N.N., Akhmetova G.D., Kozhakov K.K., Amirgalieva S.S. Method of Obtaining Trypanosomal Antigen. MPC: A61K 39/005, A61K 39/00. Innovation Patent Number: 27712. Publish.
- Akhmetsadykov, N.N.; Shabdarbayeva, G.S. Development of Technology and Organization of Production of Enzyme Immunoassay Systems for the Diagnosis of Trypanosomiasis of Horses and Pyroplasmidosis of Animals.//Materials of the 1st International Conference “As.
- Shabdarbayeva, G.S. Immunobiological Aspects of Constructing Diagnostics Based on Antiidiotypy for the Detection of Animal Blood Parasitosis//Diss. Doctors of Biology, Almaty, 2008, 312 P.
- Clausen, P.-H.; Chuluun, S.; Sodnomdarjaa, R.; Greiner, M.; Noeckler, K.; Staak, C.; Zessin, K.-H.; Schein, E. A Field Study to Estimate the Prevalence of Trypanosoma Equiperdum in Mongolian Horses. Vet. Parasitol. 2003, 115, 9–18. [Google Scholar] [CrossRef]
- Claes, F.; Radwanska, M.; Urakawa, T.; Majiwa, P.A.O.; Goddeeris, B.; Büscher, P. Variable Surface Glycoprotein RoTat 1.2 PCR as a Specific Diagnostic Tool for the Detection of Trypanosoma Evansi Infections. Kinetoplastid Biol. Dis. 2004, 3, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Yakimov, B.L. Identification of Trypanosis (Dourine) of Russian and Algerian Origin //Bulletin of the General Vet - Ii. - 1914, No.23. - Pp.19-23.
- Ocholi, R.A.; Ezeugwu, R.U.; Nawathe, D.R. Mixed Outbreak of Trypanosomiasis and Babesiosis in Pigs in Nigeria. 1988.
- Kazansky, I.I. Diagnosis, Therapy and Prevention of Dourine //Veterinary Medicine. - M., 1947, No.6. - Pp.4-8.
Figure 1.
Global distribution of central trypanosomiasis infections in humans and animals. Shaded regions highlight the primary endemic zones of different Trypanosoma species: Africa (orange), where T. brucei spp. Human African Trypanosomiasis (HAT, sleeping sickness) and animal trypanosomiasis (nagana) are found in Latin America (blue), where T. cruzi is responsible for American trypanosomiasis (Chagas disease), and in Asia (green), where T. evansi (surra) and T. vivax affect livestock and camels. Icons indicate predominant hosts: humans, livestock, and camels. These parasites collectively impact both public health and animal production, underscoring their importance within the One Health framework [6,7,8,9,10,11,12,13,20,25,29,30,31].
Figure 1.
Global distribution of central trypanosomiasis infections in humans and animals. Shaded regions highlight the primary endemic zones of different Trypanosoma species: Africa (orange), where T. brucei spp. Human African Trypanosomiasis (HAT, sleeping sickness) and animal trypanosomiasis (nagana) are found in Latin America (blue), where T. cruzi is responsible for American trypanosomiasis (Chagas disease), and in Asia (green), where T. evansi (surra) and T. vivax affect livestock and camels. Icons indicate predominant hosts: humans, livestock, and camels. These parasites collectively impact both public health and animal production, underscoring their importance within the One Health framework [6,7,8,9,10,11,12,13,20,25,29,30,31].
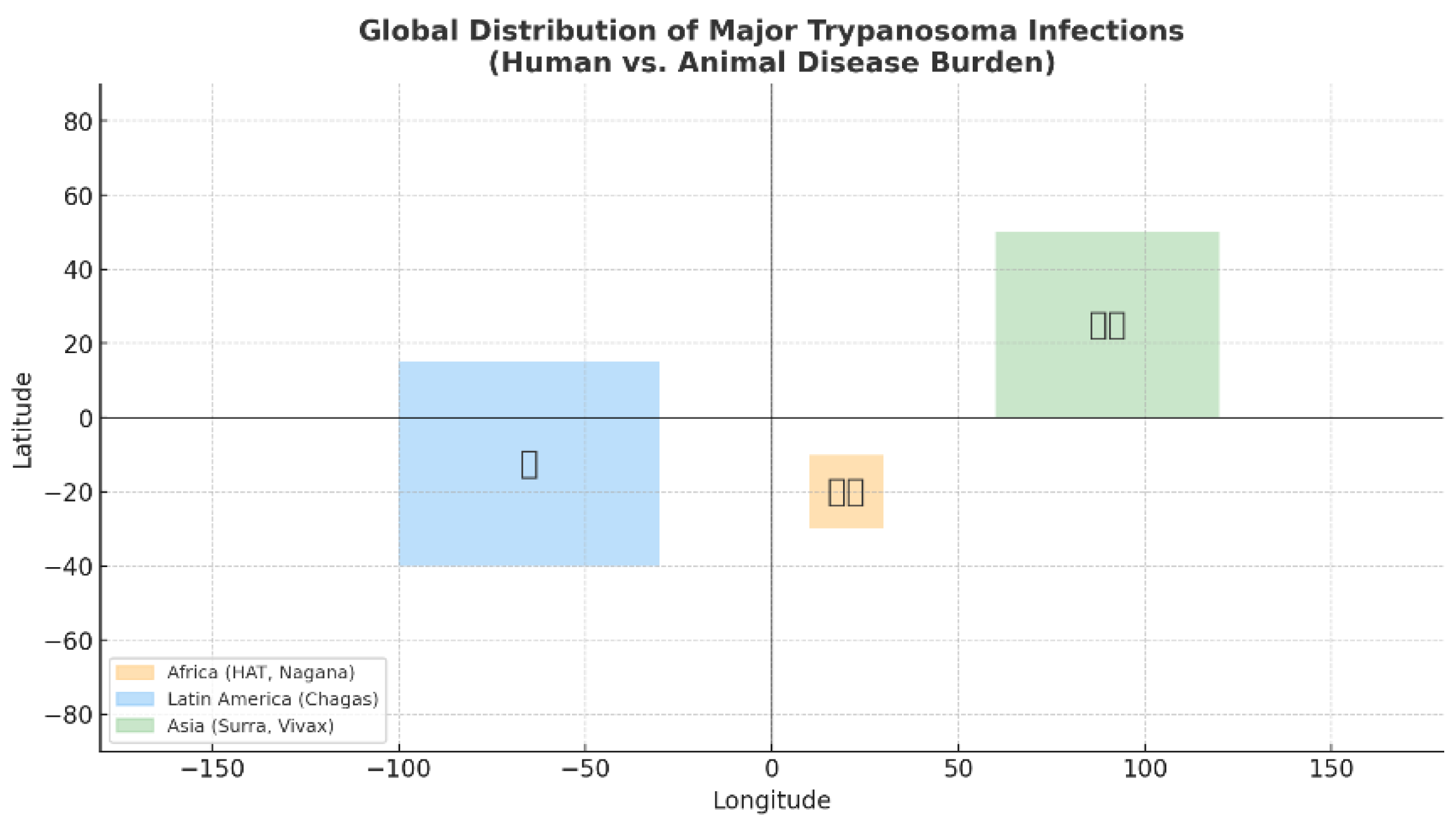
Figure 2.
Map of the camel population in Kazakhstan, highlighting Mangystau, Kyzylorda, and Turkestan regions. Camel population density is shown by a color gradient ranging from light yellow (low density) to dark blue (high density). Statistical data for 2025 were obtained from the Bureau of National Statis-tics (https://stat.gov.kz).
Figure 2.
Map of the camel population in Kazakhstan, highlighting Mangystau, Kyzylorda, and Turkestan regions. Camel population density is shown by a color gradient ranging from light yellow (low density) to dark blue (high density). Statistical data for 2025 were obtained from the Bureau of National Statis-tics (https://stat.gov.kz).
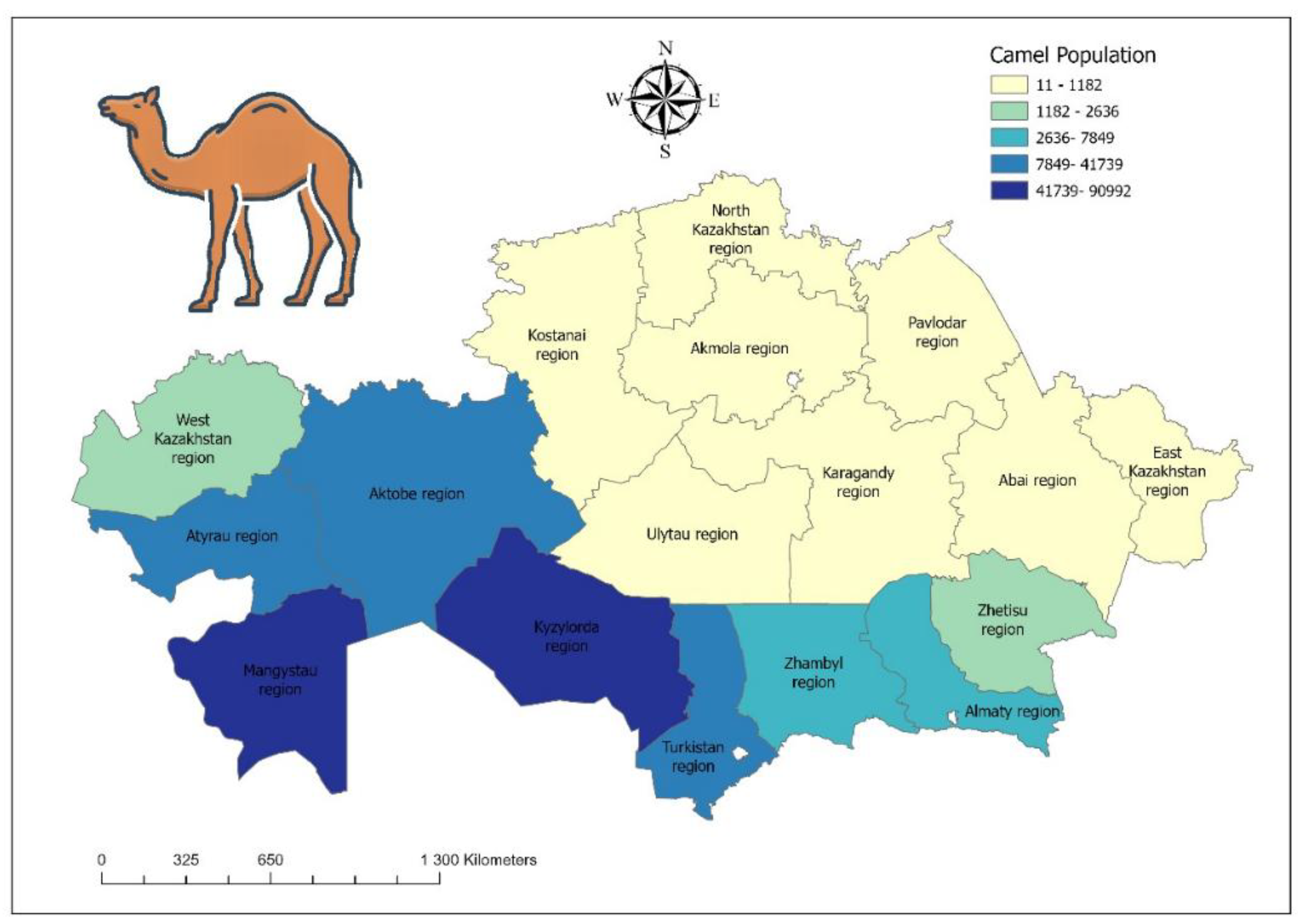
Figure 3.
Map of the equine population in Kazakhstan. Equine population density is shown by a color gradient ranging from light yellow (low density) to dark blue (high density). Statistical data for 2025 were obtained from the Bureau of National Statistics (https://stat.gov.kz).
Figure 3.
Map of the equine population in Kazakhstan. Equine population density is shown by a color gradient ranging from light yellow (low density) to dark blue (high density). Statistical data for 2025 were obtained from the Bureau of National Statistics (https://stat.gov.kz).
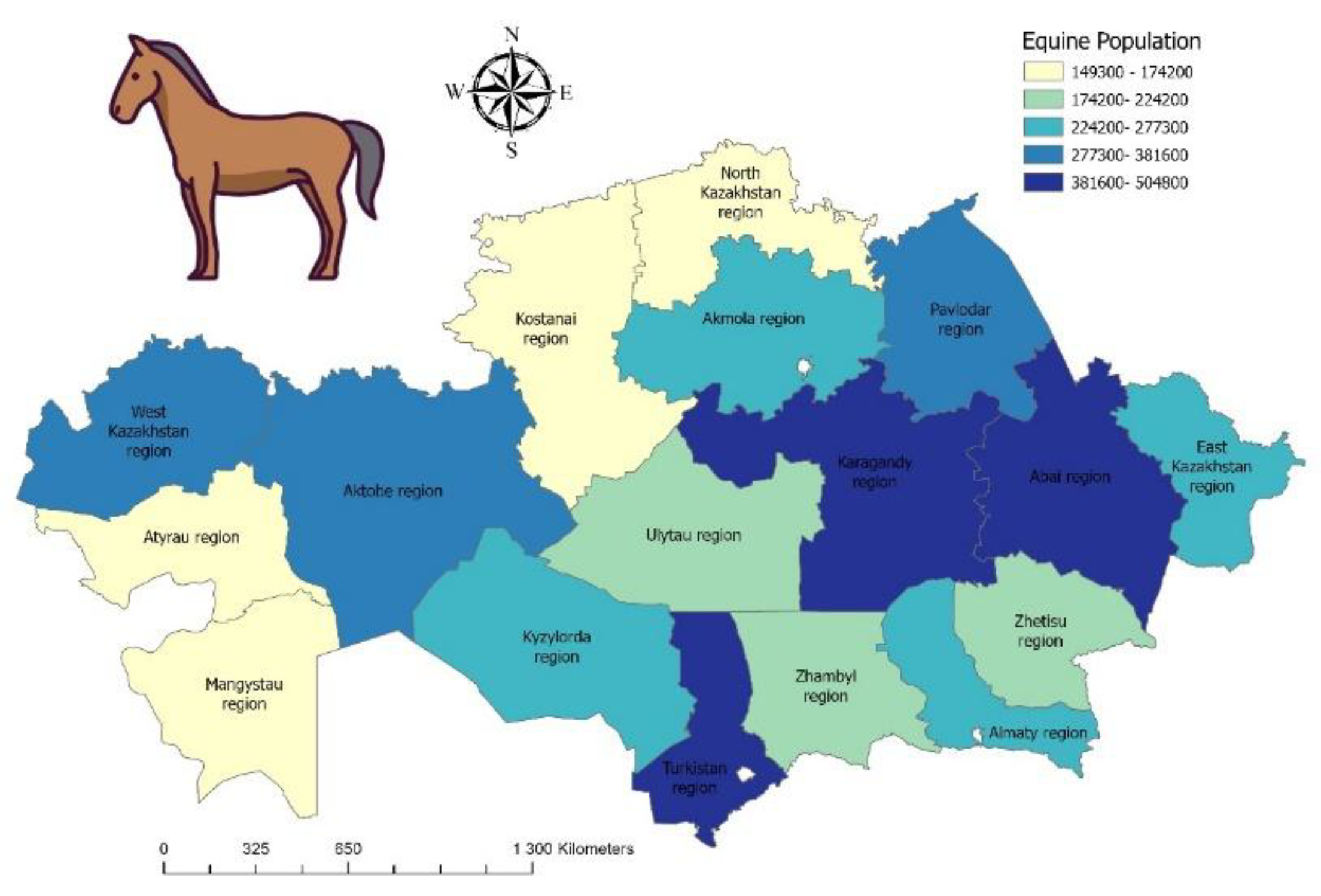
Figure 4.
Reported human cases of atypical trypanosomiasis (a-HT), including T. evansi infections. The bar chart highlights countries where confirmed cases have been reported: India, Egypt, and Vietnam, alongside sporadic reports from other regions. Kazakhstan is included for comparison, where no human cases have been recorded [56,57,58,59], but where livestock trade and climate change may increase future zoonotic risk. The data presented are illustrative and reflect published case reports rather than population-level incidence.
Figure 4.
Reported human cases of atypical trypanosomiasis (a-HT), including T. evansi infections. The bar chart highlights countries where confirmed cases have been reported: India, Egypt, and Vietnam, alongside sporadic reports from other regions. Kazakhstan is included for comparison, where no human cases have been recorded [56,57,58,59], but where livestock trade and climate change may increase future zoonotic risk. The data presented are illustrative and reflect published case reports rather than population-level incidence.
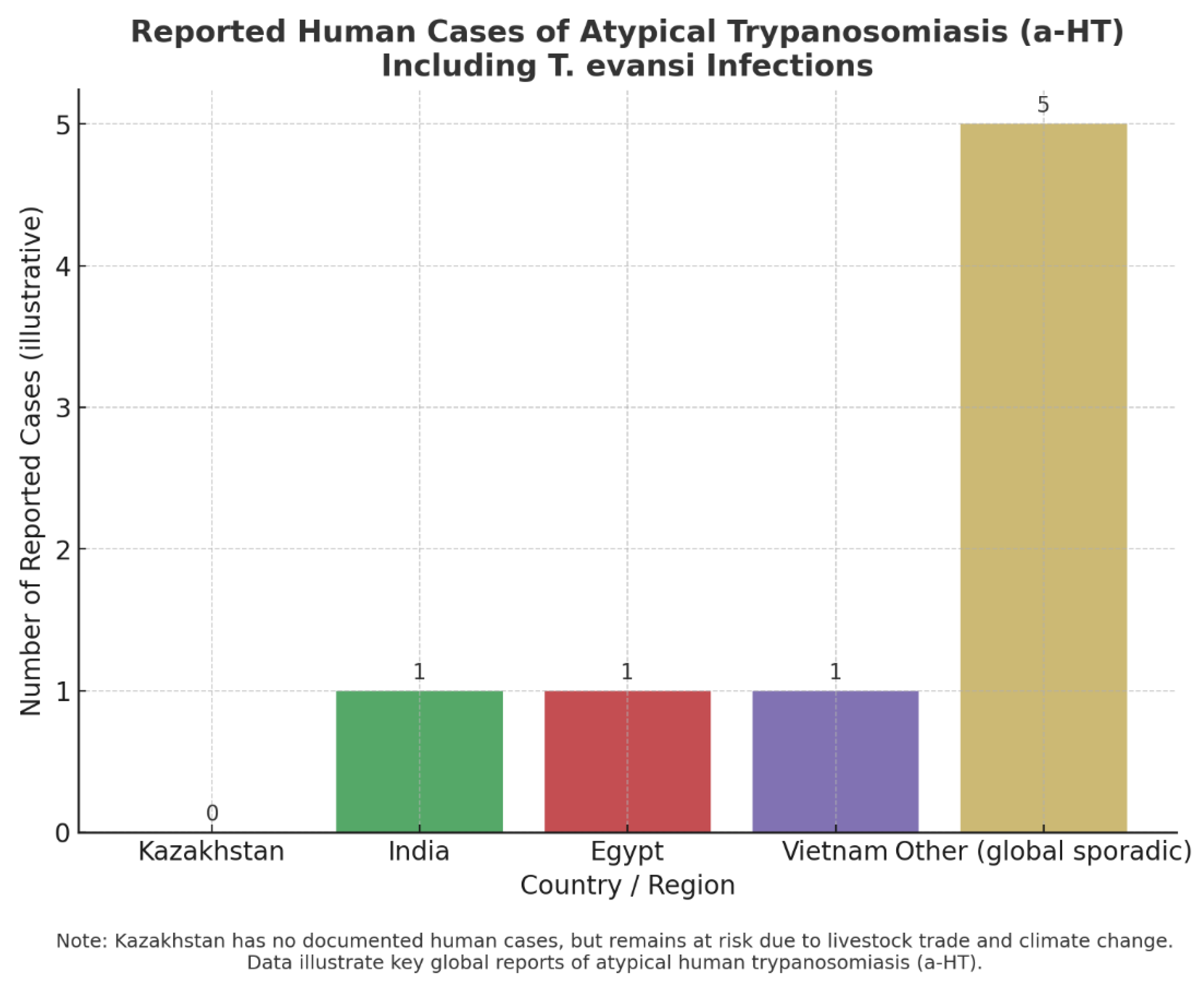
Figure 5.
Relative emphasis on diagnostic approaches for animal trypanosomiasis. Parasitological methods (30%) remain the traditional gold standard, offering low-cost and straightforward confirmatory diagnosis but with poor sensitivity in chronic or subclinical infections [54,74,75,76,77,78,79,80,81,82,83,84,85]. Serological methods (35%) provide higher sensitivity but suffer from cross-reactivity between Trypanosoma species, and the complement fixation test (CFT), developed in 1915, remains widely used despite its limitations [86,89,90,91]. Molecular methods (35%) offer the most tremendous potential for specificity and sensitivity, yet remain costly, field-limited, and unable to reliably distinguish T. brucei from T. equiperdum [95,103,104,105]. For Kazakhstan, where WOAH listed the country as free from equine trypanosomosis between 2005 and 2022, local studies nonetheless confirmed significant infection rates (16.4% in Almaty horses, 2004; further cases in 2014) [70,71,72], highlighting the need for more reliable and accessible diagnostic tools.
Figure 5.
Relative emphasis on diagnostic approaches for animal trypanosomiasis. Parasitological methods (30%) remain the traditional gold standard, offering low-cost and straightforward confirmatory diagnosis but with poor sensitivity in chronic or subclinical infections [54,74,75,76,77,78,79,80,81,82,83,84,85]. Serological methods (35%) provide higher sensitivity but suffer from cross-reactivity between Trypanosoma species, and the complement fixation test (CFT), developed in 1915, remains widely used despite its limitations [86,89,90,91]. Molecular methods (35%) offer the most tremendous potential for specificity and sensitivity, yet remain costly, field-limited, and unable to reliably distinguish T. brucei from T. equiperdum [95,103,104,105]. For Kazakhstan, where WOAH listed the country as free from equine trypanosomosis between 2005 and 2022, local studies nonetheless confirmed significant infection rates (16.4% in Almaty horses, 2004; further cases in 2014) [70,71,72], highlighting the need for more reliable and accessible diagnostic tools.
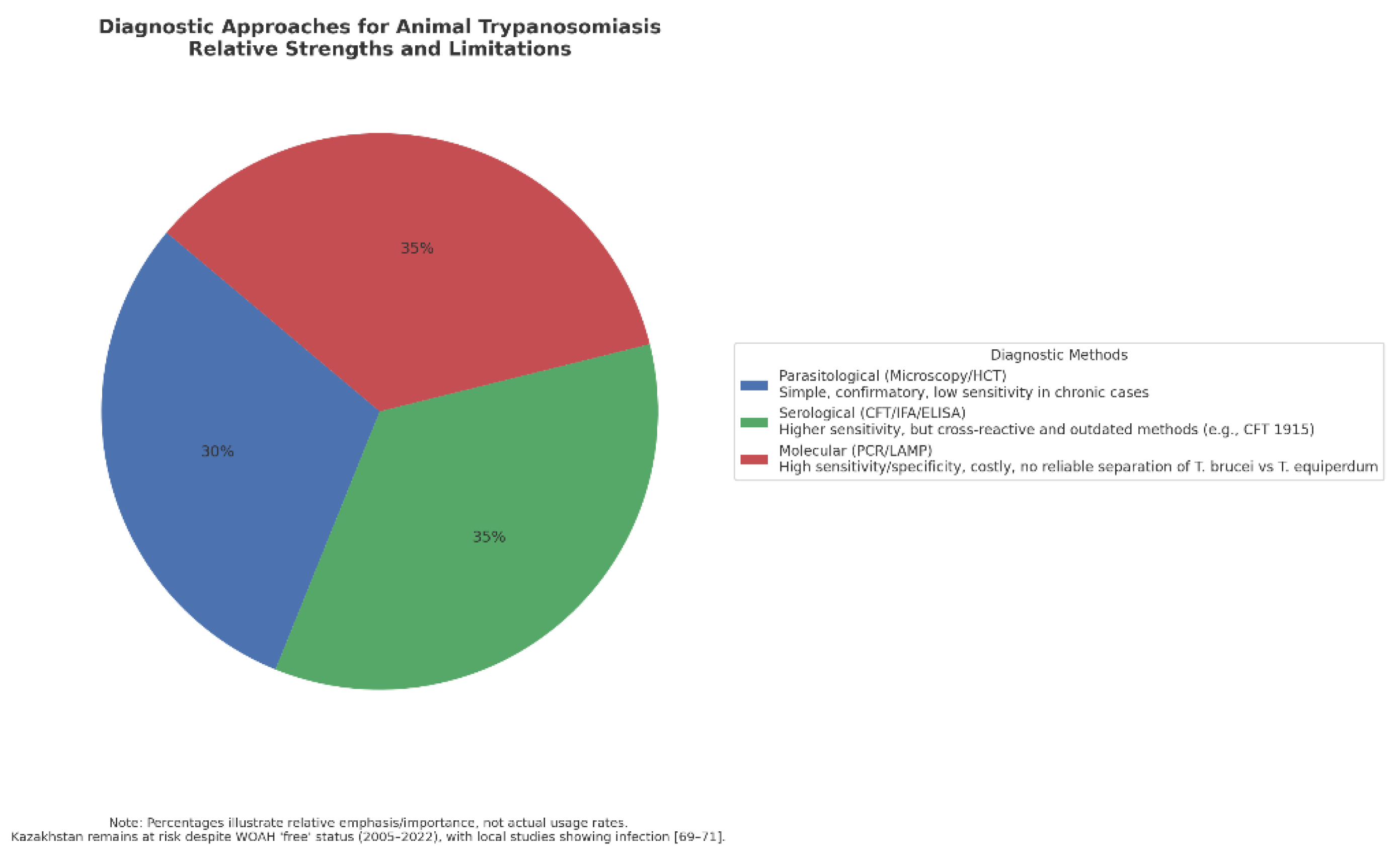
Figure 6.
Climate change indicators and vector-borne disease (VBD) risk in Kazakhstan in the global context. The average annual air temperature in Kazakhstan is increasing by 0.32 °C per decade, with projections of +3.3 to +6.2 °C by 2085, depending on the region [114,115]. Climate-driven northward shifts of wet zones by 50–100 km and reductions of insufficiently humid areas by 6–23% are expected [115,116]. Currently, 66% of Kazakhstan’s land area (179.9 million ha) is affected by desertification [115]. These changes create conditions for the expansion of vector habitats, the reemergence of neglected diseases, and the emergence of novel zoonoses. In parallel, the WHO, WOAH, and UN have set global targets to reduce mortality from vector-borne diseases by 75% by 2030 [78,115]. Error bars indicate reported ranges for projected temperature rise, wet-zone shifts, and humid area reductions.
Figure 6.
Climate change indicators and vector-borne disease (VBD) risk in Kazakhstan in the global context. The average annual air temperature in Kazakhstan is increasing by 0.32 °C per decade, with projections of +3.3 to +6.2 °C by 2085, depending on the region [114,115]. Climate-driven northward shifts of wet zones by 50–100 km and reductions of insufficiently humid areas by 6–23% are expected [115,116]. Currently, 66% of Kazakhstan’s land area (179.9 million ha) is affected by desertification [115]. These changes create conditions for the expansion of vector habitats, the reemergence of neglected diseases, and the emergence of novel zoonoses. In parallel, the WHO, WOAH, and UN have set global targets to reduce mortality from vector-borne diseases by 75% by 2030 [78,115]. Error bars indicate reported ranges for projected temperature rise, wet-zone shifts, and humid area reductions.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



