
Submitted:
23 January 2026
Posted:
26 January 2026
You are already at the latest version
Abstract
Cellular senescence is typically driven by DNA damage, telomere attrition, and metabolic/mitochondrial stress, resulting in a state of durable proliferative arrest accompanied by a senescence-associated secretory phenotype (SASP) that amplifies inflammation and paracrine remodeling across tissues, thereby accelerating functional decline and age-related pathologies. This review examines the molecular mechanisms and in vivo activities of procyanidin C1 (PCC1), a natural dual
mode geroprotector. This dual behavior ostensibly allows PCC1 to function in an early or low
dose phase wherein PCC1 attenuates selected NF
κB–driven SASP components while stabilizing redox and bioenergetic homeostasis; under high senescent burden or tumor-associated stress, elevated doses of PCC1 enable the selective reduction of refractory senescent cells and mitigation of pro-tumorigenic SASP outputs, concomitant with modulation of immune infiltration and metabolic reprogramming. PCC1 further exhibits pronounced anti-inflammatory and anti-tumor potential by reshaping inflammatory and chemotactic gradients within the tumor microenvironment. Reflecting broad multi-organ geroprotective potential, PCC1 can exert antioxidative, mitochondrial-supportive, and anti-fibrotic modulation in cardiovascular/metabolic tissues, skin, liver, kidney, and neural niches. Compared with BCL-2 inhibitors or multi-kinase senolytics, PCC1 shows lower cytotoxicity toward normal proliferating cells and fewer indications of platelet or hematopoietic suppression, with its staged regulatory profile and natural scaffold suggesting a wider therapeutic window. Future priorities include quantifying dose–timing transition thresholds, establishing integrated biomarker panels, and optimizing delivery strategies to define its translational potential in precision, phase-adapted geroprotective interventions.
Keywords:
cellular senescence
; geroprotector
; procyanidin C1
; senolytic
; supplement
1. Introduction
Cellular senescence represents an adaptive defense mechanism triggered under stress conditions such as telomere shortening, persistent DNA damage, oxidative stress, and mitochondrial dysfunction [1,2,3]. Key to its regulatory framework is the cascade activation of the p53/p21 signaling axis and the p16^INK4a/Rb (p16) pathway, which collectively induce cell cycle arrest, thereby halting cellular proliferation. During this process, senescent cells exhibit distinct phenotypic hallmarks, including increased cell size and flattened morphology, accompanied by metabolic reprogramming, adaptive adjustments in mitochondrial function, dysregulation of the autophagy-lysosomal system, hierarchical remodeling of chromatin three-dimensional architecture, and epigenetic landscape reprogramming [2]. The aforementioned biochemical cascades not only arrest cellular proliferation but also drive the formation of the senescence-associated secretory phenotype (SASP) through genome-wide transcriptional reprogramming. SASP factors are continuously secreted via paracrine and autocrine pathways [3,4], exhibiting remarkable heterogeneity in composition. These factors primarily include: (1) pro-inflammatory cytokines (e.g., IL-6, IL-8); (2) chemokines (e.g., MCP-1/CCL2); (3) growth and angiogenic factors (e.g., TGF-β1, VEGF); and (4) matrix-remodeling enzymes (e.g., MMPs) [3,4]. Additionally, coagulation/tissue remodeling regulators, lipid/metabolic mediators [1], and extracellular vesicles such as exosomes are also encompassed within SASP. These molecules collectively modulate immune cell recruitment, remodeling of tissue architecture and microenvironmental signaling, and reprogramming of cellular fates. However, persistent activation of SASP can amplify chronic inflammation, deplete stem cell functionality, promote tissue fibrosis and structural degeneration, and drive immunosenescence, thereby accelerating the onset and progression of various age-related diseases [1,2,4].
The core aim of current geroprotective strategies is to mitigate age-related diseases and extend healthspan by either eliminating senescent cells or modulating the SASP [5,6,7]. Among these approaches, senolytics aim to selectively induce apoptosis in senescent cells by transiently inhibiting multiple senescent cell anti-apoptotic pathways. Representative senolytic agents, such as the dasatinib-quercetin (D+Q) combination, fisetin, and the BCL-2 inhibitor Navitoclax, effectively reduce the burden of senescent cells, thereby alleviating SASP-induced tissue damage and delaying age-associated functional decline [5,8]. Alternatively, senomorphics target the suppression of SASP-driven chronic inflammation while preserving tissue repair mechanisms. Key compounds in this category include mTOR inhibitors (e.g., rapamycin), JAK-STAT pathway inhibitors, and modulators of critical inflammatory signaling pathways such as NF-κB and p38 MAPK. These agents aim to restore tissue functionality and decelerate the aging process [5,6,9]. In addition, immune-mediated strategies, including CAR-T cell therapy, vaccines, and antibody-drug conjugates, enhance the clearance of senescent cells by targeting surface markers specific to senescence. However, these approaches are often associated with high costs and risks of excessive immune activation [6,7]. Despite their promise, all these strategies face challenges related to precise targeting, dose optimization, and long-term safety. Multi-target combinatorial interventions and intermittent dosing regimens are emerging as promising directions for minimizing adverse effects and improving therapeutic efficacy [5,7,8].
In nature, certain naturally occurring polyphenols have demonstrated senolytic properties, enabling the selective clearance of senescent cells. Notably, compounds such as quercetin and fisetin have been developed as dietary supplements and are now commercially available. In recent years, a trace bioactive component derived from grape seed extract (GSE), the B-type procyanidin trimer PCC1, has attracted increasing interest due to its unique molecular mechanisms, exceptional bioactivity, and pan-organ senotherapeutic potential. Crucially, PCC1 has been reported to exhibit superior senolytic activity in vitro compared to quercetin and fisetin [10]. Currently, dietary supplements utilizing GSE and PCC1 for senolytic purposes are being introduced to the market. The bioactivity of GSE and its key active components, procyanidins, has been extensively investigated, with both efficacy and safety validated in human trials. However, PCC1 remains the only known procyanidin molecule with demonstrated senotherapeutic activity, yet its underlying molecular mechanisms and safety profile have not been comprehensively explored or summarized. This review first examines the molecular mechanisms of PCC1, its organ-specific anti-aging effects, as well as its anti-tumor and anti-inflammatory properties. Subsequently, it delves into a comparative analysis of the safety profile of PCC1 relative to other senolytics. Finally, the review highlights current limitations in the applications of PCC1 and proposes future directions for its development.
2. Multiple Mechanism of PCC1 for Diverse Application
To date, emerging evidence suggests that PCC1 exhibits a wide range of bioactivities, including anti-aging, anti-cancer, anti-inflammatory, antioxidant, and antiviral effects. These diverse mechanistic pathways can act synergistically, and their efficacy has been validated across various disease models. This section provides a comprehensive overview of these molecular mechanisms underlying PCC1’s actions, along with its in vivo outcomes.
2.1. PCC1 as a Senotherapeutic Candidate
With regard to its anti-aging properties, PCC1 has been shown in multiple in vivo studies to possess pan-organ senotherapeutic activity, encompassing the retina, brain, lungs, kidneys, skin, hematopoietic and immune systems (HIS). This broad efficacy across diverse organs rivals that of the well-established D+Q combination, suggesting PCC1 holds significant promise for human applications. Importantly, unlike the dose-dependent toxic side effects associated with D+Q or Navitoclax, the naturally derived PCC1 has demonstrated arguably fewer safety concerns across the reported dosage ranges to date.
2.1.1. Dose-Dependent Dual Mechanism
In 2021, Sun and the Kirkland Group first jointly elucidated the dose-dependent dual-action mechanism of PCC1 [10]. At low concentrations (approximately 1–20 μM), PCC1 functions as a senomorphic agent, mitigating the inflammatory damage caused by senescent cells to surrounding tissues by suppressing SASP release, while exhibiting negligible toxicity to healthy cells. Conversely, at higher concentrations (≥50 μM, approximately 43 μg/mL), PCC1 transitions into a senolytic agent, selectively inducing apoptosis in senescent cells while preserving normal tissue integrity. For certain highly resistant senescence models, such as oxidative stress-induced tubular cell senescence, effective clearance may require higher concentrations (approximately 100 μM). Subsequently, this dose-stratified dual mechanism has been validated across various cellular models in multiple studies (Table 1), demonstrating that PCC1 significantly reduces SASP such as SA-β-GAL, IL-6, and IL-1β [10,11,12]. Furthermore, findings from various animal models, such as NOD-SCID mice and BALB/c nude mice, have confirmed PCC1’s bidirectional intervention in senescence burden and inflammation amplification, further supporting its cross-organ anti-aging potential (Table 2). The following section will detail the molecular mechanisms and anti-aging efficacy of PCC1 across different organs and tissues.
The molecular basis for the biphasic dose-response is best explained by the ROS threshold theory. Senescent cells maintain a precarious redox homeostasis characterized by elevated basal oxidative stress [10]. At low concentrations, PCC1 primarily functions as a signal modulator, likely interacting with surface receptors to attenuate SASP-driving cascades, such as NF-κB and p38 MAPK, without exceeding the cytotoxic threshold. In contrast, at higher concentrations, the intracellular accumulation of PCC1 induces a significant increase in mitochondrial ROS. While healthy cells can neutralize this oxidative burst through robust antioxidant systems, senescent cells are pushed beyond their survival capacity, leading to the activation of JNK/caspase-dependent apoptotic pathways [10]. This mechanism establishes a broad therapeutic window, distinguishing PCC1 from other senolytic agents that do not exhibit such redox-dependent selectivity.
This mechanism is critical for PCC1’s ability to address the heterogeneity of senescent cell populations and achieve efficacy across different organs. Recent studies have shown that cells expressing p16 and p21 often represent distinct senotypes with divergent transcriptomic trajectories. Unlike certain senolytics that exhibit pathway dependency, PCC1 demonstrates a strong capacity to eliminate both phenotypes. Direct genetic evidence from p16-luciferase reporter mice confirmed that PCC1 effectively clears p16-driven senescent cells in vivo [10]. Furthermore, in various pathological contexts—ranging from the aged retina [12] to fibrotic kidneys [13] and skin [14]—PCC1 treatment consistently induces synchronous downregulation of both p16 and p21 at the mRNA and protein levels. This indicates that PCC1 targets a common metabolic vulnerability of senescent cells, specifically mitochondrial dysfunction and ROS intolerance, rather than relying on specific upstream cell cycle regulators, thereby ensuring comprehensive clearance of heterogeneous senescent burdens.
2.1.2. PCC1 for Anti-Fibrosis and Overcoming Senolytic Resistance
Tissue fibrosis represents a pathological process closely linked to aging, characterized by tissue thickening, hardening, reduced elasticity, and impaired organ function. Senescent cells play a pivotal role in this process, primarily through the secretion of SASP factors, such as TGF-β, IL-6, and MMPs, which alter the local microenvironment. These SASP factors can promote fibroblast proliferation and collagen deposition, thereby driving the progression of tissue fibrosis in organs such as the skin, kidneys, and lungs. Specifically, TGF-β1 induces fibroblast-to-myofibroblast transition (FMT) via Smad-dependent pathways, exacerbating collagen synthesis and tissue stiffening. In the kidneys, senescent tubular epithelial cells (TECs) secrete TGF-β1 to trigger epithelial–mesenchymal transition (EMT), leading to interstitial fibrosis. Furthermore, SASP factors like IL-6 and MMP13 amplify these signals, establishing a positive feedback loop between chronic inflammation and fibrosis. Consequently, targeting the clearance of senescent cells or blocking SASP signaling pathways holds promise as a novel therapeutic strategy for combating fibrosis [25,26,27,28].
Crucially, PCC1's anti-fibrotic potency addresses a critical limitation of first-generation senolytics like Dasatinib and Quercetin (D+Q). Recent studies indicate that a subset of 'senolytic-resistant' senescent cells can evade D+Q clearance by shifting their phenotype towards a highly fibrotic state driven by TGF-β signaling [29]. Unlike D+Q, which primarily targets anti-apoptotic pathways (SCAPs), PCC1 directly intercepts the TGF-β/Smad signaling axis [9,30]. By blocking this pathway, PCC1 effectively eliminates these refractory senescent subpopulations that would otherwise persist and drive tissue stiffening. This capacity to target the heterogeneity of senescence—clearing both inflammatory and fibrotic/resistant subpopulations—positions PCC1 as a superior candidate for reversing organ fibrosis compared to single-target agents.
In recent years, multiple studies have reported that PCC1 can alleviate fibrosis in various organs through its dual senomorphic and senolytic actions, by reducing SASP secretion from senescent cells and selectively clearing these cells (Figure 1A–C). Wang et al. demonstrated that in skin fibrosis (Figure 1A), PCC1 mitigates bleomycin-induced skin thickening and hardening by inhibiting excessive receptor phosphorylation in the EGFR/Smad pathway, thereby reducing the expression of α-SMA and type I collagen [14]. In a renal interstitial fibrosis model (Figure 1B), Yu et al. demonstrated that PCC1 significantly reduces collagen deposition and fibrosis severity by inducing apoptosis in senescent TECs and suppressing anti-apoptotic signaling mediated by ANGPTL4/NOX4 [13]. Furthermore, Xu et al. reported that PCC1 selectively eliminates senescent subpopulations in human embryonic lung fibroblasts (WI38) in vitro, reduces SASP factor levels, and inhibits TGF-β1-induced myofibroblast differentiation, highlighting its potential therapeutic effects on pulmonary fibrosis (Figure 1C) [10]. Collectively, PCC1 not only attenuates SASP-driven fibroblast proliferation and collagen deposition through the clearance of senescent cells but also directly suppresses key profibrotic signaling pathways, achieving multi-target, multi-mechanism antifibrotic effects across the skin, kidneys, and lungs.
2.1.3. PCC1 for Retina Protection
Senescent cells play a critical role in the pathogenesis of ocular fundus diseases by secreting SASP, such as IL-6, IL-8, and MMP-9, which drive chronic inflammation, extracellular matrix remodeling, and pathological neovascularization [31,32,33,34]. These factors not only disrupt the blood-retinal barrier (BRB) but also induce fibrosis, leading to damage to retinal pigment epithelial (RPE) cells and neural retinal cells. In age-related macular degeneration (AMD), senescent RPE cells progressively accumulate and secrete pro-inflammatory and pro-osteogenic factors, accelerating degenerative changes in the macular region [32,33]. In diabetic retinopathy (DR), the hyperglycemic microenvironment induces premature senescence of retinal vascular endothelial cells, resulting in microvascular rarefaction, capillary occlusion, and aberrant neovascular growth with vascular leakage [34]. Furthermore, senescent cells exacerbate the inflammation-fibrosis-angiogenesis vicious cycle by activating signaling pathways such as NF-κB, MAPK, and TGF-β, thereby promoting inflammation propagation and tissue fibrosis [31,32,33,34]. Collectively, senescent cells emerge as key drivers of chronic progression in ocular fundus diseases through mechanisms of “inflammation-vascular-neural” imbalance [31,32,33,34,35]. UBX1325, a BCL-xL inhibitor developed by the American biotechnology company Unity Biotechnology, is a senolytic drug that has demonstrated potential in early clinical studies for diabetic macular edema (DME) and neovascular AMD, showing improvements in visual acuity and macular thickness with a favorable safety profile [35,36,37].
PCC1 can offer dual senolytic and senomorphic pathways to protect the retina and delay the progression of related ocular diseases. For instance, long-term administration of PCC1 in mouse models significantly alleviates retinal senescence burden, as evidenced by reduced expression of SA-β-Gal, p16, and p21, alongside suppression of SASP factors such as IL-6, IL-1β, and TNF-α [12]. Single-cell RNA sequencing reveals that PCC1 reshapes gene expression across multiple retinal cell types, enhancing NADH regeneration and mitochondrial autophagy while downregulating inflammation-related signaling pathways such as NF-κB and MAPK (Figure 1D). PCC1 selectively induces mitochondrial-associated programmed cell death in senescent cells via enhanced ROS response and activation of apoptotic pathways, while maintaining high viability in non-senescent cells, underscoring its “minimal toxicity” advantage. Furthermore, this study demonstrates that PCC1 significantly improves retinal structure and function, including restoring retinal electrophysiological responses and reducing abnormalities in the synaptic layer.
2.1.4. PCC1 for HIS Modulation
With aging, the immune and hematopoietic systems undergo cellular functional decline and immune response imbalance, ultimately leading to immunosenescence. Immunosenescence is characterized not only by a reduction in immune cell numbers and functional efficacy but also by the exacerbation of chronic low-grade inflammation, significantly impairing the body’s resistance to infections, tumors, and autoimmune diseases [38]. Moreover, one study by Geiger et al. revealed that the functional decline of hematopoietic stem cells (HSCs) leads to an imbalance in the ratio of myeloid and lymphoid cells, placing the immune system in a dual predicament of inflammatory activation and insufficient immune defense, thereby accelerating tissue damage and disease progression [39]. While first-generation senolytics like the Dasatinib and Quercetin (D+Q) cocktail effectively clear senescent cells, they carry a risk of compromising immune integrity. Dasatinib, as a broad-spectrum tyrosine kinase inhibitor (TKI), mechanistically suppresses Src family kinases (e.g., Lck), which are critical for T-cell receptor signaling and activation [40]. Consequently, D+Q has been reported to impair immune surveillance by reducing the infiltration of cytotoxic CD8+ T cells and macrophages in certain contexts [41,42].
In stark contrast, PCC1 demonstrates a unique capacity to rejuvenate the HIS without these immunosuppressive tradeoffs. Upon systemic administration, PCC1 enters the extracellular microenvironment of the bone marrow and spleen, directly targeting HSCs, B cells, and myeloid cells(Figure 1E) [23]. PCC1 utilizes a ROS-dependent mechanism rather than kinase inhibition, allowing it to selectively induce apoptosis in senescent myeloid cells by disrupting the mitochondrial outer membrane [10,23]. Crucially, this mechanism spares healthy immune cells. In surviving B cells, PCC1 downregulates the key regulatory gene Cebpb, reducing the release of TNF-α and suppressing the accumulation of age-associated B cells (ABCs). Furthermore, PCC1 enhances the expression of Nedd4 and the CD62L-Ca2+ axis in HSCs, boosting stem cell self-renewal and differentiation capacities [23]. These findings collectively position PCC1 as a superior alternative that remodels the immune landscape toward a youthful phenotype while preserving the functional signal transduction required for active immune defense.
2.1.5. PCC1 for Alzheimer’s Disease Mitigation
The precise pathogenesis of neurodegenerative diseases, such as Alzheimer’s disease (AD), remains a topic of ongoing debate. Current therapeutic strategies for AD primarily focus on inhibiting Aβ production/clearance, Tau kinase inhibition, cholinesterase inhibitors, and modulation of neuroinflammation [44,45,46,47,48]. Recent studies have highlighted the close association between disease progression and cellular senescence. For instance, senescent cells, through mechanisms involving DNA damage, oxidative stress, and neuroinflammation, drive glial cells and neurons into a state of permanent proliferative arrest while releasing pro-inflammatory SASP factors. These factors can exacerbate Aβ aggregation, Tau protein phosphorylation, and neuroinflammation, ultimately leading to synaptic dysfunction and neuronal loss [49,50]. Therefore, the targeted clearance of senescent cells in the brain has been shown to significantly reduce Aβ and Tau pathological burden, thereby suppressing inflammatory responses and improving cognitive function. In recent years, D+Q has emerged as a promising anti-senescence therapeutic strategy. Zhang P et al. reported that D+Q effectively eliminates senescent cells with high expression of p16, reduces pathological burden, and improves cognitive performance [45]. Furthermore, a Phase I clinical trial conducted by Gonzales et al. confirmed that D+Q is safe and well-tolerated in AD patients, accompanied by a reduction in SASP factor levels, laying the groundwork for subsequent large-scale studies [51].
As a novel senotherapeutic molecule, PCC1 holds potential for the treatment of neurodegenerative diseases. For instance, when formulated as nanoparticles (e.g., Glu-BSA/C1 NPs), PCC1 can traverse the blood-brain barrier and directly target senescent cells in the brain [21]. In senescent neurons or glial cells, PCC1 disrupts the mitochondrial outer membrane and activates the BAX/BCL-2 pathway, leading to caspase protein activation and apoptosis induction, thereby facilitating the clearance of senescent cells. This process significantly reduces the release of pro-inflammatory SASP factors, alleviating neuroinflammation and oxidative stress (Figure 1). Meanwhile, PCC1 translocates into the nuclei of surviving cells, where it suppresses the expression of senescence-associated genes such as p53, p21, and p16, thereby reducing the levels of inflammatory cytokines (e.g., TNF-α, IL-18) and oxidative stress markers (e.g., MDA). Additionally, PCC1 enhances antioxidant capacity in the cytoplasm by increasing the activity of glutathione (GSH) and superoxide dismutase (SOD), while modulating the PI3K/AKT1 metabolic pathway to protect neuronal function [21]. By mitigating inflammation and cellular damage in brain tissue, PCC1 significantly improves synaptic plasticity and neuronal survival, effectively slowing the progression of AD.
2.2. PCC1 for Anti-Cancer Application
Malignant tumors remain one of the leading causes of mortality worldwide, with their high recurrence, metastasis, and drug resistance closely linked to intrinsic uncontrolled proliferation, EMT, and the cascade amplification of inflammatory and stress signals within the tumor microenvironment (TME). Long-term use of multi-target small molecules or single-target inhibitors often faces challenges such as increased tumor resistance and difficulty in controlling metastatic lesions. This highlights the urgent need for “functionally integrative” molecules capable of simultaneously intervening in malignant proliferation, blocking invasion pathways, and disrupting mechanisms of death evasion [52,53].
PCC1 has attracted interest for its multifaceted mechanisms of action, including synergistic growth inhibition, metastasis blockade, and induction of programmed cell death across multiple cancer models [15,16,18,24]. In colorectal cancer, Lv et al. reported that PCC1 suppresses tumor progression by downregulating miR-501-3p, thereby relieving its negative regulation on the tumor suppressor protein HIGD1A. This disruption impairs cellular adaptation to hypoxia and mitochondrial stress, leading to a marked inhibition of both primary tumor growth and metastatic nodule formation (Figure 2A) [15]. In the breast cancer model, Koteswari et al. demonstrated that PCC1 induces DNA damage and activates checkpoint signaling, leading to G2/M phase arrest. Concurrently, PCC1 downregulates Bcl-2, upregulates BAX, and sequentially activates caspases, triggering mitochondria-dependent apoptosis, thereby exhibiting a dual effect of “growth blockade + death induction” (Figure 2B) [18]. In melanoma, Bae et al. revealed that PCC1 binds to the 67 kDa laminin receptor (67LR), disrupting the cytoskeleton and cytokinesis, which results in mitotic arrest during the late anaphase and precise proliferation suppression (Figure 2C) [24]. Regarding the invasive and metastatic phenotypes of lung and colorectal cancers, PCC1 upregulates E-cadherin while downregulating N-cadherin and Snail, key factors of EMT (Figures 2A,D), thereby impairing tumor cell migration and invasion capabilities and effectively blocking early events in metastasis at the pathway level [15].
In addition to targeting intrinsic tumor pathways, PCC1 provides a significant advantage in reversing the aging-associated pre-metastatic niche by reshaping the tumor microenvironment (TME). The accumulation of senescent cells often creates an immunosuppressive barrier that facilitates tumor escape. A pivotal study by Duan et al. revealed that aging promotes melanoma metastasis through a specific γδT17-Neutrophil-CD8 axis{Duan, 2024 #46}. PCC1 treatment effectively disrupts this pro-tumorigenic cascade by selectively depleting senescent γδT cells and neutrophils. Importantly, unlike non-selective agents, PCC1 restores the stemness (TCF1+) and proliferative capacity (Ki67+) of tumor-infiltrating CD8+ T cells, resulting in a significant reduction in metastatic burden{Duan, 2024 #46}.
This "immune-synergistic" capability contrasts sharply with the "immune-antagonistic" risks observed with D+Q in neoplastic contexts. Luo et al. reported that D+Q treatment in a skin papilloma model unexpectedly accelerated tumor progression{Luo, 2025 #54}. Mechanistically, D+Q compromised anti-tumor immunity by upregulating PD-L1 expression and depleting CD8+ T cells and macrophages within the TME{Luo, 2025 #54}. This creates a "cold" tumor environment vulnerable to escape. Therefore, while D+Q may inadvertently foster an immunosuppressive TME, PCC1 synergistically combines senolysis with immune surveillance restoration, presenting a safer strategy for cancer prevention in the context of aging.
In summary, PCC1 establishes a comprehensive anti-tumor pathway through a multi-layered cascade mechanism, encompassing a coordinated network. It includes the regulation of gene expression through microRNA signaling pathways, receptor-mediated remodeling of the cytoskeleton, activation of cell cycle checkpoints to ensure genomic stability and proper function, induction of mitochondrial-driven apoptosis, and reversal of epithelial-to-mesenchymal transition to restore normal cell differentiation. This integrative strategy effectively suppresses tumor proliferation, invasion, and metastasis, while demonstrating pharmacological consistency across diverse cancer models and tissue types [15,16,18,24]. As previously described, PCC1 has been independently validated for its ability to eliminate or modulate senescent cells. Its senotherapeutic properties further mitigate the pro-tumorigenic inflammatory and immunosuppressive microenvironment sustained by SASP, indirectly reinforcing its direct anti-tumor effects. These preliminary findings provide a theoretical foundation for the translational exploration of PCC1 in precision stratified medicine and the development of complementary therapeutic combinations with conventional chemotherapy or targeted/immunotherapy.
2.3. PCC1 for Anti-Inflammation
As a polyphenolic molecule, PCC1 shares the prominent and multi-pathway anti-inflammatory properties characteristic of other polyphenols, such as quercetin and EGCG. The anti-inflammatory activity of PCC1 is mediated through a synergistic mechanism involving the suppression of key inflammatory signaling pathways, modulation of immune cell function, attenuation of oxidative stress, and inhibition of senescence-associated inflammation. Byun et al. demonstrated that PCC1 effectively suppresses the canonical NF-κB and MAPK pathways by blocking TLR4-mediated signaling, inhibiting the phosphorylation of p38, ERK, and JNK, and preventing IκBα degradation, thereby restraining NF-κB activation and nuclear translocation [17]. This pathway inhibition subsequently reduces the transcription of downstream pro-inflammatory genes, significantly lowering TNF-α and IL-6 production, while also suppressing iNOS expression and excessive NO generation [17,54]. Furthermore, Hua et al. revealed that PCC1 activates the SIRT3/FOXO3 signaling axis, restoring mitochondrial homeostasis, scavenging ROS, and alleviating oxidative stress-induced damage [55]. Additionally, Shi et al. demonstrated that PCC1 modulates immune cell function by inhibiting the polarization of macrophages toward the pro-inflammatory M1 phenotype, thereby alleviating tissue inflammation in colitis models [2]. As previously noted, PCC1 also exhibits unique efficacy in combating age-related chronic low-grade inflammation by significantly suppressing the SASP of senescent cells, reducing the secretion of pro-inflammatory factors such as IL-6 [10], and delaying structural and functional degeneration in the retinas of aged mice [12]. Collectively, PCC1 establishes an anti-inflammatory network through multi-target and multi-pathway mechanisms, offering a clear mechanistic basis and novel therapeutic strategies for inflammation-related diseases.
2.4. PCC1 for Glucose Metabolism Regulation
Procyanidins (PCs) have been shown to alleviate β-cell dysfunction by enhancing insulin sensitivity, promoting GLUT4 translocation, and exerting antioxidant and anti-inflammatory effects [56]. Early studies on the overall insulin-regulatory effects of procyanidins highlighted the coordinated activation of the IRS/PI3K/AKT and AMPK pathways, along with the downregulation of GSK3; however, the specific active monomer or oligomeric structure responsible for these effects remains undefined . Recent studies by Zhang et al. and El-Ashmawy et al. have highlighted the insulin-regulatory effects of PCC1 [19,57,58,59]. In 3T3-L1 adipocytes, PCC1 significantly promotes adipogenic differentiation and enhances insulin-stimulated glucose uptake through mechanisms involving the AKT-eNOS axis and NO signaling, suggesting its direct role in augmenting insulin efficacy [60]. Furthermore, Sun P et al. demonstrated that PCC1 protects pancreatic β-cells from lipotoxicity or oxidative stress, effectively reducing ROS levels and supporting glucose-stimulated secretory function, indicating its dual role in peripheral insulin sensitization and endocrine support [19]. These mechanisms align with the ability of oligomeric procyanidins to enhance insulin receptor phosphorylation, upregulate GLUT4 and PI3K, and inhibit GSK3, suggesting that PCC1 functions as a multi-pathway nodal molecule to amplify insulin signal transduction efficiency [59]. In a skeletal muscle study by Yamashita et al., trimeric and tetrameric procyanidins were shown to activate both insulin and AMPK signaling pathways, facilitating GLUT4 membrane translocation. These findings may support the cross-pathway synergistic role of PCC1 in muscle tissues [58]. Collectively, PCC1, as a multi-pathway nodal molecule, holds potential therapeutic value in peripheral insulin sensitization and pancreatic β-cell protection, offering novel strategies for the prevention and treatment of diabetes and its complications.
2.5. Potential Antivirus Application
Since its emergence in late 2019, SARS-CoV-2 has rapidly spread worldwide, yet effective and broad-spectrum small-molecule therapeutic options remain limited. Viral entry and replication are mediated by the interaction between the spike (S) protein and ACE2 receptor, as well as the coordinated actions of key enzymes such as Mpro, helicase, and RdRp. A molecular docking and dynamics simulation study revealed that procyanidins (including PCC1) can form stable hydrogen bonds and hydrophobic/van der Waals interactions with ACE2, Mpro, and the S protein. The MM-PBSA binding energies were calculated as -89.5 kcal/mol for Mpro, -50.2 kcal/mol for ACE2, and -23.1 kcal/mol for the S protein, supporting the potential for multi-target affinity. Furthermore, the biological effects of PCC1, including anti-inflammatory activity, regulation of oxidative stress, and ACE inhibition, have been demonstrated in previous studies, theoretically suggesting its ability to mitigate the inflammatory “amplification” phenomenon induced by COVID-19 [17,54]. However, current evidence for its antiviral efficacy is primarily computational, and PCC1 requires systematic validation through in vitro studies, animal models, and pharmacokinetic and safety assessments before its therapeutic potential can be fully evaluated [61].
3. Safety Considerations
The heterogeneity of cellular and tissue senescence, along with inter-individual variability, presents a significant challenge in advancing senolytic drug development to human clinical applications. A critical hurdle lies in accurately distinguishing pathological senescent cells from benign senescent cells. In the absence of precise identification methods, the most prudent strategy would be to adopt a relatively conservative therapeutic approach rather than indiscriminately eliminating all senescent cells. Recent reports have highlighted a concerning example where combined treatment with D+Q potentially promoted early tumor development in mice, highlighting the need for cautious evaluation [42]. While the likelihood of such adverse effects occurring in the general population is relatively low, given that D+Q has already undergone extensive animal and human clinical testing, heightened vigilance and long-term monitoring are warranted for its effects in pre-tumorigenic conditions. This is particularly important as Dasatinib is known to affect T cell activation and migration, with documented cytotoxic effects on immune infiltration [40]. In contrast, no direct immunosuppressive effects have been reported for PCC1. On the contrary, PCC1 exhibits multiple immunocompensatory and tumor-suppressive mechanisms that are absent in D+Q. For instance, PCC1 can inhibit tumor cell division and metabolic adaptation through pathways such as 67LR-PKA-PP2A and miR-501-3p/HIGD1A axes [15,16,18,24]. Additionally, as an EGFR inhibitor, PCC1 has been shown in multiple models to potentially decelerate clonal expansion and suppress certain PD-L1-driven STAT3/PI3K cascades [14]. These multifaceted compensatory mechanisms suggest that PCC1 may effectively clear senescent cells without compromising normal immune infiltration functions, although further in-depth validation and investigation are required.
From the perspective of dietary supplementation, PCC1 may represent the most promising short-term alternative for targeting senescent cell clearance. On one hand, its intrinsic senomorphic properties enable effective suppression of the persistent secretion of SASP by senescent cells at low doses [10]. This feature also allows for real-time monitoring and tracking, providing a safeguard for personalized therapeutic interventions. On the other hand, these naturally derived candidates have demonstrated exceptionally high safety profiles even at senolytic-effective high doses, a stark contrast to the dose-dependent toxicity challenges faced by synthetic drugs such as Navitoclax and Dasatinib [40,62]. While Quercetin and Fisetin also exhibit safety profiles comparable to PCC1 at high doses, they lack clear dose-response segmentation. For instance, low-dose Fisetin in human trials was shown to only reduce IL-8 levels, with no significant impact on other SASP components [63]. Therefore, Quercetin and Fisetin may be more suited for high-dose hit-and-run strategies to achieve senescent cell clearance and subsequently reduce SASP secretion. In contrast, PCC1 inherently possesses senomorphic effects that allow for the reduction of SASP secretion at low doses [10].
4. Unique Pharmacokinetics
PCC1 exhibits a distinct pharmacokinetic profile characterized by limited oral bioavailability of the parent molecule yet significant systemic biological activity. Due to its high molecular weight (approximately 866 Da) and hydrophilicity, PCC1 faces substantial physical barriers to passive diffusion across the intestinal epithelium. In vitro assessments utilizing Caco-2 cell monolayers—the standard model for human intestinal absorption—have consistently demonstrated negligible transepithelial transport of the intact trimer, in sharp contrast to the higher permeability observed for monomeric flavanols like epicatechin [64,65]. Furthermore, stability studies indicate that while PCC1 is relatively stable in the acidic gastric environment, it is susceptible to degradation and isomerization under the alkaline conditions of the small intestine, further reducing the concentration of the intact molecule available for absorption [66].
Despite these absorption barriers and low detectable plasma concentrations, in vivo biodistribution studies suggest alternative uptake pathways that bypass the traditional portal vein route. Wang et al. (2018) reported that following oral administration in mice, PCC1 accumulates at significantly higher concentrations in mesenteric adipose tissue compared to the liver or plasma. This specific tissue distribution supports the hypothesis of lymphatic transport, wherein lipophilic or large molecules are incorporated into chylomicrons within intestinal enterocytes and transported via the lymphatic system to the thoracic duct. This mechanism allows the intact compound to bypass hepatic first-pass metabolism, potentially explaining its bioactivity in distant tissues such as the lungs and visceral fat despite low plasma levels [67].
In addition to lymphatic transport, the unabsorbed fraction of PCC1 undergoes extensive biotransformation by the gut microbiota in the colon. Recent studies have identified that specific commensal bacteria, such as Akkermansia muciniphila, metabolize the trimeric structure into lower-molecular-weight phenolic metabolites, including valeric acid. Unlike the parent compound, these metabolites exhibit high membrane permeability and have been shown to activate systemic signaling pathways, such as the FOXO1 axis, thereby contributing to mucosal protection and anti-inflammatory effects [68]. Consequently, the therapeutic efficacy of PCC1 is likely mediated through a synergistic mechanism involving the lymphatic delivery of the intact molecule and the systemic action of bioactive microbial metabolites.
5. Future Directions
The application of PCC1 in anti-aging and combinatorial anti-tumor therapies holds immense potential; however, its precise mechanisms and in vivo metabolism may represent significant barriers to its broader clinical translation. Among the numerous molecular pathways identified, the only relatively well-characterized mechanism to date is PCC1’s ability to target the membrane protein EGFR, thereby modulating its downstream signaling pathways [14]. However, significant gaps remain in understanding how PCC1 influences mitochondrial function and key processes such as p53 nuclear translocation within cells [10]. Some molecular simulation study has highlighted the potential of PCC1’s exceptional Na+-K+-ATPase binding capacity, meriting further investigation [69]. This property is not only critical for elucidating its multifaceted anti-aging and anti-tumor activities in vivo but also has implications for determining safe dosage ranges and optimal administration cycles. Recent human studies [Clinical Trial No. NCT06641869] have preliminarily confirmed that long-term daily administration of PCC1 at doses of 2.5–5 mg is safe, providing a reference for senomorphic dosing in chronic interventions. However, the higher doses required for ‘hit-and-run’ senescent cell clearance remain to be further defined. Additionally, whether prolonged high-dose PCC1 administration induces aberrant immune cell cytotoxicity, thereby compromising immune infiltration in vulnerable microenvironments, requires rigorous experimental validation.
Another major factor limiting the clinical application of PCC1 in humans is its poor oral absorption efficiency and gastrointestinal stability. While oligomeric procyanidins can be absorbed to some extent through the gastrointestinal tract, existing reports suggest that most are metabolized by gut microbiota prior to absorption, resulting in complex metabolites such as methylated and sulfonated derivatives [70]. These intricate metabolic pathways pose a significant question: whether the observed in vivo effects of PCC1 are primarily driven by the parent compound itself or by the synergistic actions of its metabolites. Advances in formulation technologies may offer promising solutions to improve PCC1’s oral bioavailability. For instance, Indena’s phytosome technology has already been successfully applied to enhance the absorption of hydrophilic plant extracts. Phytosomal formulations of procyanidin dimers and trimers have demonstrated a 2- to 3-fold improvement in gastrointestinal absorption efficiency [71]. This hypothesis aligns with our preliminary findings (submission under peer review), where the PCC1-phytosome group showed a significant reduction in senescence markers such as SA-β-Gal, p16, and p21 in PBMCs from elderly individuals, accompanied by marked decreases in plasma levels of SASP components including IL-6, IL-8, VEGF, and TGF-β1. In contrast, the decline of some SASP were not observed in the unformulated PCC1 group, but the decrease of MMP9 was noted. These findings underscore the dual impact of formulation technologies: not only do they enhance gastrointestinal absorption efficiency, but for trimers like PCC1, they also likely induce substantial alterations in metabolic pathways. Consequently, further investigations are required to elucidate the in vivo metabolites, tissue distribution profiles, and molecular mechanisms associated with PCC1-phytosome formulations.
Emerging as a novel anti-aging strategy in recent years, senolytics have demonstrated promising preliminary clinical applications in humans. PCC1 exhibits remarkably potent senotherapeutic activity among the myriad bioactive compounds derived from nature, outperforming most known senolytics in both in vitro and in vivo studies. These initial findings provided in this work align closely with the conceptual framework for an “ideal” senescent cell clearance intervention in geriatric medicine, such as programmable dose modulation (senomorphic effects at low doses and senolytic effects at high doses), intermittent administration, multi-target therapeutic benefits, systemic anti-aging activity across organs, and non-toxic profiles. It is anticipated that with further mechanistic and metabolic studies, PCC1 holds great potential as a key nutraceutical for future health interventions and promoting healthy aging in the elderly population.
Author Contributions
Conceptualization: Ling Wang, Qinghua Lyu; Investigation (Experiments / Data Acquisition): Jiaxin Liang, Qun Wang, Jie Li, Xueying Lu; Writing: Jiaxin Liang, Qun Wang; Review & Editing: All authors.
Funding
This research was primarily supported by resources and funds from Lonvi Biosciences (Shenzhen) Co., Ltd. The company provided the necessary experimental facilities, materials, and financial support for the preparation of the B type procyanidin oligomers and the subsequent animal safety evaluations.
Conflicts of Interest
The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: Jiaxin Liang, Qun Wang, and Qinghua Lyu are affiliated with Lonvi, Inc. (USA). Jie Li and Xueying Lu are affiliated with Lonvi Biosciences (Shenzhen) Co., Ltd. (China). The work was supported by Lonvi Biosciences (Shenzhen) Co., Ltd., which provided funding, experimental facilities, and materials for the study. Ling Wang declares no competing financial interests.
References
- Guo, J.; et al. Aging and aging-related diseases: from molecular mechanisms to interventions and treatments. Signal Transduct Target Ther 2022, 7, 391. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; et al. The landscape of aging . Science China Life Sciences 2022, 65, 2354–2454. [Google Scholar] [CrossRef] [PubMed]
- Suryadevara, V.; et al. SenNet recommendations for detecting senescent cells in different tissues . Nature Reviews Molecular Cell Biology 2024, 25, 1001–1023. [Google Scholar] [CrossRef] [PubMed]
- Wong, C. How to kill the ‘zombie’cells that make you age; Nature Publishing Group, 2024. [Google Scholar]
- Chaib, S.; Tchkonia, T.; Kirkland, J.L. Cellular senescence and senolytics: the path to the clinic . Nature medicine 2022, 28, 1556–1568. [Google Scholar] [CrossRef]
- Lelarge, V.; et al. Senolytics: from pharmacological inhibitors to immunotherapies, a promising future for patients’ treatment. npj Aging 2024, 10, 12. [Google Scholar] [CrossRef]
- Gasek, N.S.; et al. Strategies for targeting senescent cells in human disease . Nature aging 2021, 1, 870–879. [Google Scholar] [CrossRef]
- Boccardi, V.; Mecocci, P. Senotherapeutics: Targeting senescent cells for the main age-related diseases . Mechanisms of ageing and development 2021, 197, 111526. [Google Scholar] [CrossRef]
- Wang, B.; et al. The senescence-associated secretory phenotype and its physiological and pathological implications. Nature Reviews Molecular Cell Biology 2024, 25, 958–978. [Google Scholar] [CrossRef]
- Xu, Q.; et al. The flavonoid procyanidin C1 has senotherapeutic activity and increases lifespan in mice. Nat Metab 2021, 3, 1706–1726. [Google Scholar] [CrossRef]
- Shao, M.; et al. Procyanidin C1 inhibits bleomycin-induced pulmonary fibrosis in mice by selective clearance of senescent myofibroblasts. FASEB J 2024, 38, e23749. [Google Scholar] [CrossRef]
- Liu, Y.; et al. Senolytic and senomorphic agent procyanidin C1 alleviates structural and functional decline in the aged retina. Proc Natl Acad Sci U S A 2024, 121, e2311028121. [Google Scholar] [CrossRef] [PubMed]
- Gan, Y.; et al. Senolytic procyanidin C1 alleviates renal fibrosis by promoting apoptosis of senescent renal tubular epithelial cells. Faseb j 2025, 39, e70362. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; et al. Procyanidin C1 ameliorates aging-related skin fibrosis through targeting EGFR to inhibit TGFbeta/SMAD pathway. Phytomedicine 2025, 142, 156787. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.L.; et al. Procyanidin C1 inhibits tumor growth and metastasis in colon cancer via modulating miR-501-3p/HIGD1A axis. J Adv Res 2024, 60, 215–231. [Google Scholar] [CrossRef]
- Kin, R.; et al. Procyanidin C1 from Cinnamomi Cortex inhibits TGF-beta-induced epithelial-to-mesenchymal transition in the A549 lung cancer cell line. Int J Oncol 2013, 43, 1901–6. [Google Scholar] [CrossRef]
- Byun, E.B.; et al. The procyanidin trimer C1 inhibits LPS-induced MAPK and NF-kappaB signaling through TLR4 in macrophages. Int Immunopharmacol 2013, 15, 450–6. [Google Scholar] [CrossRef]
- Koteswari, L.L.; et al. A comparative anticancer study on procyanidin C1 against receptor positive and receptor negative breast cancer. Nat Prod Res 2020, 34, 3267–3274. [Google Scholar] [CrossRef]
- Sun, P.; et al. Procyanidin C1, a Component of Cinnamon Extracts, Is a Potential Insulin Sensitizer That Targets Adipocytes. J Agric Food Chem 2019, 67, 8839–8846. [Google Scholar] [CrossRef]
- Song, J.H.; Lee, H.J.; Kang, K.S. Procyanidin C1 Activates the Nrf2/HO-1 Signaling Pathway to Prevent Glutamate-Induced Apoptotic HT22 Cell Death. Int J Mol Sci 2019, 20(1). [Google Scholar] [CrossRef]
- Duan, L.; et al. Glucose-modified BSA/procyanidin C1 NPs penetrate the blood-brain barrier and alleviate neuroinflammation in Alzheimer's disease models. Int J Biol Macromol 2024, 268, 131739. [Google Scholar] [CrossRef]
- Chen, J.; et al. The Relationship between Procyanidin Structure and Their Protective Effect in a Parkinson's Disease Model. Molecules 2022, 27(15). [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; et al. Single-cell profiling unveils a geroprotective role of Procyanidin C1 in hematopoietic immune system via senolytic and senomorphic effects. NPJ Aging 2025, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.; et al. Procyanidin C1 Inhibits Melanoma Cell Growth by Activating 67-kDa Laminin Receptor Signaling. Mol Nutr Food Res 2020, 64, e1900986. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.K.; Sheppard, D.; Chapman, H.A. TGF-β1 signaling and tissue fibrosis . Cold Spring Harbor perspectives in biology 2018, 10, a022293. [Google Scholar] [CrossRef]
- Sisto, M.; Ribatti, D.; Lisi, S. Organ fibrosis and autoimmunity: the role of inflammation in TGFβ-dependent EMT. Biomolecules 2021, 11, 310. [Google Scholar] [CrossRef]
- O’Reilly, S.; Tsou, P.-S.; Varga, J. Senescence and tissue fibrosis: Opportunities for therapeutic targeting . Trends in molecular medicine 2024, 30, 1113–1125. [Google Scholar] [CrossRef]
- Pohlers, D.; et al. TGF-β and fibrosis in different organs—molecular pathway imprints . Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease 2009, 1792, 746–756. [Google Scholar] [CrossRef]
- Tripathi, U.; et al. Senolytic-Resistant Senescent Cells Have a Distinct SASP Profile and Functional Impact: The Path to Developing Senosensitizers. bioRxiv 2025, 2025.08. 27.672709. [Google Scholar] [CrossRef]
- Gan, Y.; et al. Senolytic procyanidin C1 alleviates renal fibrosis by promoting apoptosis of senescent renal tubular epithelial cells. The FASEB Journal 2025, 39, e70362. [Google Scholar] [CrossRef]
- Sreekumar, P.G.; Hinton, D.R.; Kannan, R. The Emerging Role of Senescence in Ocular Disease . Oxidative Medicine and Cellular Longevity 2020, 2020, 1–19. [Google Scholar] [CrossRef]
- Li, C.; et al. Age-related macular degeneration: a disease of cellular senescence and dysregulated immune homeostasis. Clinical Interventions in Aging 2024, 939–951. [Google Scholar] [CrossRef]
- Blasiak, J. Senescence in the pathogenesis of age-related macular degeneration . Cellular and Molecular Life Sciences 2020, 77, 789–805. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.-L.; et al. Senescent endothelial cells: a potential target for diabetic retinopathy. Angiogenesis 2024, 27, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Garcia, S.; et al. Therapeutic targeting of cellular senescence in diabetic macular edema: preclinical and phase 1 trial results. Nat Med 2024, 30, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Macha, N.; et al. Multifocal Electroretinography Changes after UBX1325 (Foselutoclax) Treatment in Neovascular Age-Related Macular Degeneration. J Clin Med 2024, 13(18). [Google Scholar] [CrossRef]
- Klier, S.; et al. Safety and Efficacy of Senolytic UBX1325 in Diabetic Macular Edema. NEJM Evid 2025, 4, EVIDoa2400009. [Google Scholar] [CrossRef]
- Wrona, M.V.; et al. The 3 I's of immunity and aging: immunosenescence, inflammaging, and immune resilience. Front Aging 2024, 5, 1490302. [Google Scholar] [CrossRef]
- Geiger, H.; Denkinger, M.; Schirmbeck, R. Hematopoietic stem cell aging . Curr Opin Immunol 2014, 29, 86–92. [Google Scholar] [CrossRef]
- Cheng, F.; et al. Adverse reactions after treatment with dasatinib in chronic myeloid leukemia: Characteristics, potential mechanisms, and clinical management strategies. Front Oncol 2023, 13, 1113462. [Google Scholar] [CrossRef]
- Duan, R.; et al. Aging-induced immune microenvironment remodeling fosters melanoma in male mice via γδ17-Neutrophil-CD8 axis. Nature Communications 2024, 15, 10860. [Google Scholar] [CrossRef]
- Luo, Y.; et al. Senolytic drugs dasatinib and quercetin promote skin papilloma progression by eliminating senescent cells and impairing immune surveillance. Oral Science and Homeostatic Medicine 2025, 1(1). [Google Scholar] [CrossRef]
- Xu, Q.; et al. The flavonoid procyanidin C1 has senotherapeutic activity and increases lifespan in mice. Nature Metabolism 2021, 3, 1706–1726. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Wang, X. Alzheimer's disease: insights into pathology, molecular mechanisms, and therapy. Protein Cell 2025, 16, 83–120. [Google Scholar] [CrossRef] [PubMed]
- Reddi Sree, R.; et al. Newer Therapeutic Approaches in Treating Alzheimer's Disease: A Comprehensive Review. ACS Omega 2025, 10, 5148–5171. [Google Scholar] [CrossRef]
- Zhang, J.; et al. Recent advances in Alzheimer's disease: Mechanisms, clinical trials and new drug development strategies. Signal Transduct Target Ther 2024, 9, 211. [Google Scholar] [CrossRef]
- Peng, Y.; et al. Current and future therapeutic strategies for Alzheimer's disease: an overview of drug development bottlenecks. Front Aging Neurosci 2023, 15, 1206572. [Google Scholar] [CrossRef]
- Tiwari, S.; et al. Alzheimer's disease: pathogenesis, diagnostics, and therapeutics. Int J Nanomedicine 2019, 14, 5541–5554. [Google Scholar] [CrossRef]
- Martínez-Cué, C.; Rueda, N. Cellular Senescence in Neurodegenerative Diseases . Front Cell Neurosci 2020, 14, 16. [Google Scholar] [CrossRef]
- Liu, R.M. Aging, Cellular Senescence, and Alzheimer's Disease. Int J Mol Sci 2022, 23(4). [Google Scholar] [CrossRef]
- Gonzales, M.M.; et al. Senolytic therapy in mild Alzheimer's disease: a phase 1 feasibility trial. Nat Med 2023, 29, 2481–2488. [Google Scholar] [CrossRef]
- Srivastava, N.; Saxena, A.K. Novel small molecule inhibitors of cyclin-dependent kinases as anticancer agents. Current Medicinal Chemistry 2025. [Google Scholar] [CrossRef]
- Puxeddu, M.; Silvestri, R.; Regina, G.L. Metabolism, a Blossoming Target for Small-Molecule Anticancer Drugs . Molecules 2025, 30, 3457. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; et al. Procyanidin improves experimental colitis by regulating macrophage polarization. Biomed Pharmacother 2023, 165, 115076. [Google Scholar] [CrossRef] [PubMed]
- Hua, W.; et al. Procyanidin C1 ameliorates acidic pH stress induced nucleus pulposus degeneration through SIRT3/FOXO3-mediated mitochondrial dynamics. J Transl Med 2024, 22, 1071. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; et al. Grape seed proanthocyanidins ameliorate pancreatic beta-cell dysfunction and death in low-dose streptozotocin- and high-carbohydrate/high-fat diet-induced diabetic rats partially by regulating endoplasmic reticulum stress. Nutr Metab (Lond) 2013, 10, 51. [Google Scholar] [CrossRef]
- Zhang, Y.; et al. The application of procyanidins in diabetes and its complications: a review of preclinical studies. Frontiers in Pharmacology 2025, 16, 1532246. [Google Scholar] [CrossRef]
- Yamashita, Y.; et al. Procyanidin promotes translocation of glucose transporter 4 in muscle of mice through activation of insulin and AMPK signaling pathways. PLoS One 2016, 11, e0161704. [Google Scholar] [CrossRef]
- El-Ashmawy, N.E.; et al. Upregulation of GLUT4 and PI3K, and downregulation of GSK3 mediate the anti-hyperglycemic effects of proanthocyanidins. Medicine International 2022, 2, 14. [Google Scholar] [CrossRef]
- Sun, P.; et al. Procyanidin C1, a component of cinnamon extracts, is a potential insulin sensitizer that targets adipocytes. Journal of Agricultural and Food chemistry 2019, 67, 8839–8846. [Google Scholar] [CrossRef]
- Maroli, N.; et al. The potential role of procyanidin as a therapeutic agent against SARS-CoV-2: a text mining, molecular docking and molecular dynamics simulation approach. J Biomol Struct Dyn 2022, 40, 1230–1245. [Google Scholar] [CrossRef]
- Gandhi, L.; et al. Phase I study of Navitoclax (ABT-263), a novel Bcl-2 family inhibitor, in patients with small-cell lung cancer and other solid tumors. J Clin Oncol 2011, 29, 909–16. [Google Scholar] [CrossRef] [PubMed]
- Farsad-Naeimi, A.; et al. Effect of fisetin supplementation on inflammatory factors and matrix metalloproteinase enzymes in colorectal cancer patients. Food & Function 2018, 9, 2025–2031. [Google Scholar] [CrossRef] [PubMed]
- Hemmersbach, S.; et al. Transepithelial permeability studies of flavan-3-ol-C-glucosides and procyanidin dimers and trimers across the Caco-2 cell monolayer. Journal of agricultural and food chemistry 2013, 61, 7932–7940. [Google Scholar] [CrossRef] [PubMed]
- Ou, K.; Gu, L. Absorption and metabolism of proanthocyanidins . Journal of Functional Foods 2014, 7, 43–53. [Google Scholar] [CrossRef]
- Zhu, Y.; et al. New agents that target senescent cells: the flavone, fisetin, and the BCL-XL inhibitors, A1331852 and A1155463. Aging (Albany NY) 2017, 9, 955. [Google Scholar] [CrossRef]
- Wang, L.; et al. Absorption, metabolism, distribution and faecal excretion of B-type procyanidin oligomers in mice after a single oral administration of black soybean seed coat extract. Food & Function 2018, 9, 5362–5370. [Google Scholar] [CrossRef]
- Wang, X.; et al. Procyanidin C1 modulates the microbiome to increase FOXO1 signaling and valeric acid levels to protect the mucosal barrier in inflammatory bowel disease. Engineering 2024, 42, 108–120. [Google Scholar] [CrossRef]
- Heger, T.; et al. Procyanidin C1 from Viola odorata L. inhibits Na+, K+-ATPase . Scientific Reports 2022, 12, 7011. [Google Scholar] [CrossRef]
- Nie, F.; et al. Oligomeric proanthocyanidins: An updated review of their natural sources, synthesis, and potentials. Antioxidants 2023, 12, 1004. [Google Scholar] [CrossRef]
- Lu, M.; et al. Phyto-phospholipid complexes (phytosomes): A novel strategy to improve the bioavailability of active constituents. Asian journal of pharmaceutical sciences 2019, 14, 265–274. [Google Scholar] [CrossRef]
Figure 1.
Senotherapeutic Mechanisms of PCC1 in Different Tissues: Skin (A), Kidney (B), Lung(C), Retina(D), Hematopoietic and immune system(E) and Brain (F).
Figure 1.
Senotherapeutic Mechanisms of PCC1 in Different Tissues: Skin (A), Kidney (B), Lung(C), Retina(D), Hematopoietic and immune system(E) and Brain (F).
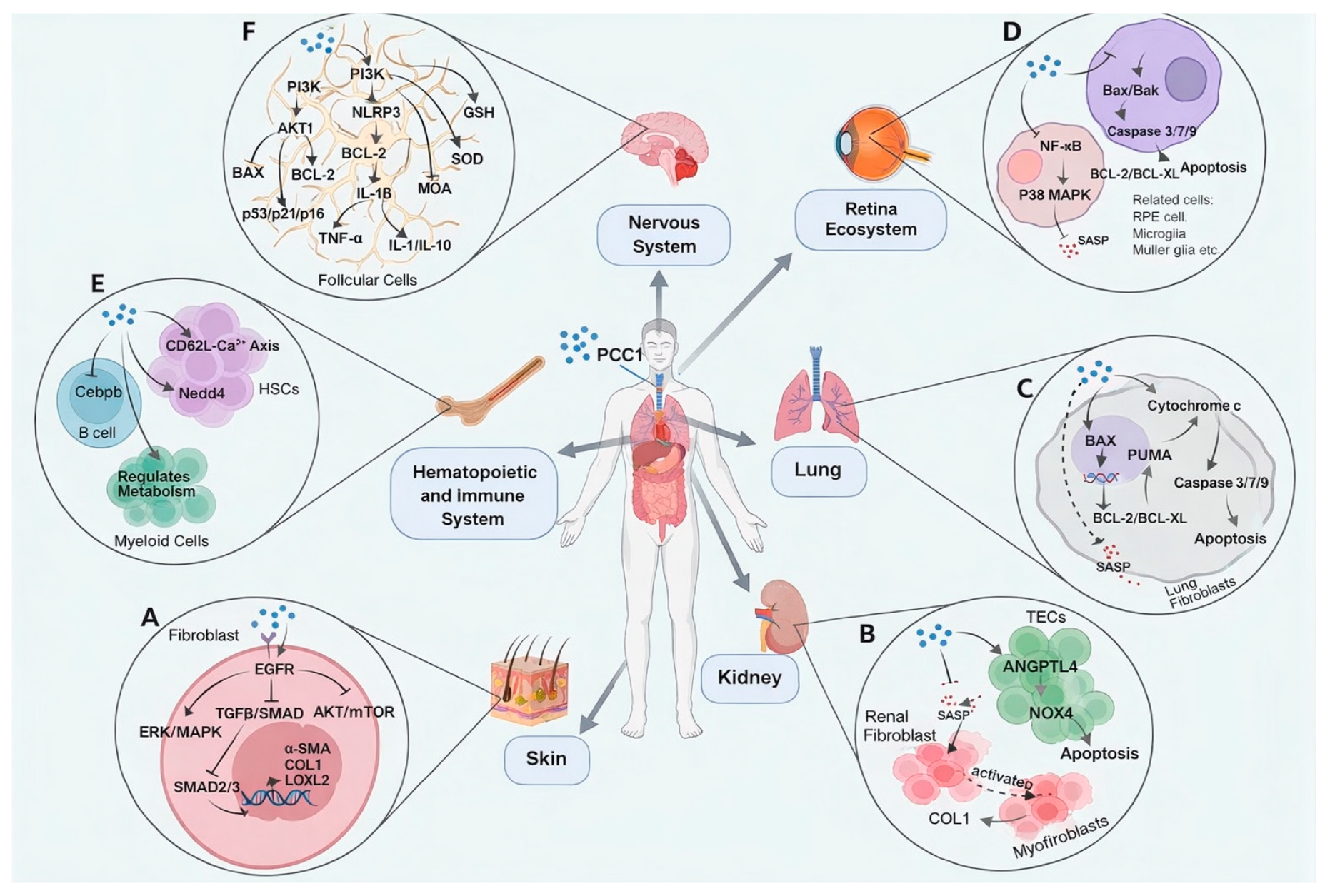
Figure 2.
Anticancer Mechanisms of PCC1 in Multiple Tumor Models: Colon Adenocarcinoma(A), Breast Cancer (B), Skin Cutaneous Melanoma (C) and Lung Cancer (D).
Figure 2.
Anticancer Mechanisms of PCC1 in Multiple Tumor Models: Colon Adenocarcinoma(A), Breast Cancer (B), Skin Cutaneous Melanoma (C) and Lung Cancer (D).
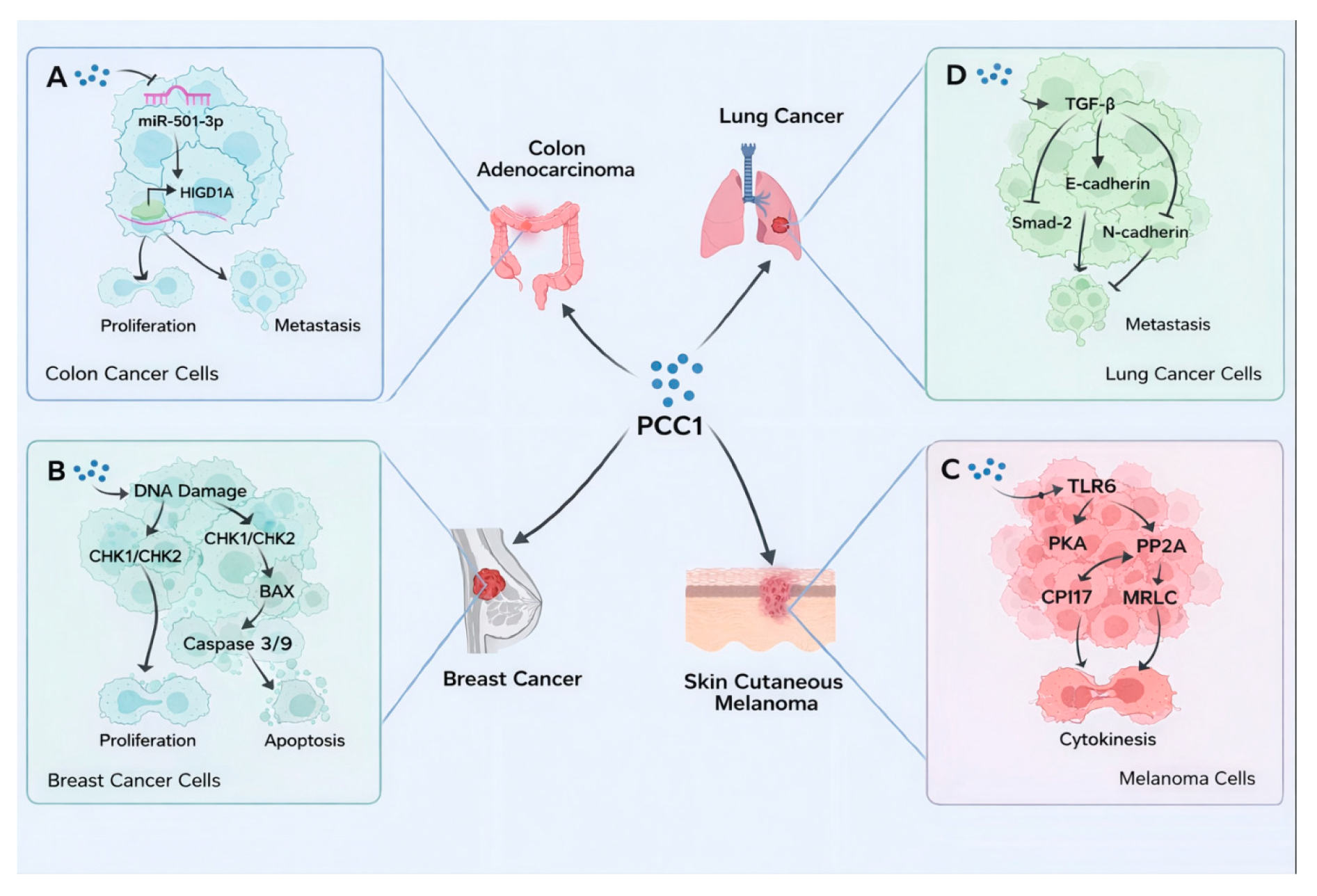

Table 1.
Molecular Mechanisms of Procyanidin C1 in Various Senescent Cells.
| Cells | Dosages | Mechanisms (index) | References |
|---|---|---|---|
| 661W, BV2, ARPE-19, HRMEC Cells | 50–200 μM | ↓:SA-β-GAL, p16/p21, IL-6, IL-1β, TNF-α, VEGF-A | [12] |
| Primary Pulmonary FibroblastsPSC27, WI-38,HUVEC, MSC Cells |
100-200 μM | ↓: SA-β-GAL, CXCL8, IL-1α ↑: NOXA, PUMA, ROS, MitoSOX |
[10] |
| 100 μM | ↓: SA-β-GAL, p16, p21, p53, ECM Deposition, Mitochondrial Membrane Potential (ΔΨm) ↑: Mitochondrial ROS levels |
[11] | |
| Colorectal Cancer Cell Lines | 5,10 μM | ↓: miR-501-3p, p-Rb, MMP-9, N-cadherin ↑: HIGD1A |
[15] |
| Lung Cancer Cell Line A549 | 5,10, 20 μM | ↓: N-cadherin, p-Smad2 ↑: E-cadherin |
[16] |
| RAW264.7, BMDMs | 29, 58, 115 μM | ↓: NO, TNF-α, IL-6, PGE2, CD80, CD86, MHC II, iNOS, COX-2 | [17] |
| MCF-7, MDA-MB-231 | 40 μM | ↑: BAX/Bcl-2, Caspase-3/9 | [18] |
| Murine Melanoma B16 Cells | 5 μM | ↑: PP2A, PKA | [18] |
| Mouse 3T3-L1 Preadipocytes | 25 μM | ↓: Glycerol Release ↑: TG, aP2/PPARγ, p-AKT, NO, GLUT4 |
[19] |
| HT22 Cells | 5,10 μM | ↓: p-ERK1/2,p-p38, ROS, Annexin V+ ↑: DPPH , Nrf2, HO-1 |
[20] |
| 58 μM | ↓: ROS, MDA, TNF-α, IL-1β, BAX ↑: GSH, SOD, Bcl-2, IL-4, IL-10 |
[21] | |
| PC12 Cells | 5 μM | ↓: ROS, MDA ↑: Nrf2, HO-1, NQO1 |
[22] |
| Murine fibroblast cell line L929 | 12.5, 25, 50 μM | ↓:p-EGFR, ERK/MARK, SA-β-Gal, LOXL2, COL1 ↑:EGFR |
[14] |
| HK2 | 100 μM | ↓:TGFB1, IL-1B, IL-6, MMP1, MMP3, FAS | [13] |
Table 2.
Anti-Aging Efficacy of Procyanidin C1 in various animal models.
| Models | Dosages | Mechanisms (index) | References |
|---|---|---|---|
| NOD-SCID Mice | 20 mg/kg | ↓: SA-β-GAL, IL-6, CXCL8, WNT16B, tumor volume ↑: caspase-3 |
[10] |
| BALB/c Nude Mice | 20 mg/kg 40 mg/kg |
↓: miR-501-3p, Ki67+, p-Rb,MMP-9,N-cadherin ↑: HIGD1A |
[15] |
| C57BL/6J Mice | 8 mg/kg | ↓:SA-β-GAL, p16/p21, IL-6,IL-1β, TNF-α,Apoe, rem2, Spp1 | [12] |
| 20 mg/kg | ↓: SA-β-GAL, IL-6, CCL2, WNT16B , MCP-1 ↑:NOXA, PUMA, Caspase-3, Median Survival, Walking Speed, Balance Performance |
[10] | |
| 8 mg/kg | ↓:TNF, IL-6, IL-1α/β, CCL, CXCL2, MAPK ↑:Nedd4, Ube2r2, Ube2k, Cul3, Adipor1, Adipor2 |
[23] | |
| 2.5 mg/kg 5 mg/kg 10 mg/kg |
↓:p-EGFR, TGF-β1, α-SMA | [14] | |
| 20 mg/kg | ↓:SA-β-Gal, p16, p21 | [13] | |
| B16 Melanoma-Bearing Mice | 30 mg/kg | ↓: CPI17, MRLC tumor volume ↑: PP2A, PKA |
[24] |
| Zebrafish Larvae | 25 μM | ↓: ROS, MDA, GSH-Px, CAT ↑: Nrf2, HO-1, NQO1, SOD |
[22] |
| 5×FAD Transgenic Mice | 5 mg/kg | ↓:Aβ1-42, Tau, p-Tau (ser396,235), AChE, Bax, P21, P16, P53, TNF-α, IL-1β, MDA, NLRP3 ↑:BDNF, Ki67, NeuN, DCX, Bcl-2, IL-4, IL-10, GSH, SOD, PI3K/AKT |
[21] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



