
Submitted:
01 September 2025
Posted:
02 September 2025
You are already at the latest version
Abstract
Water deficit is a critical limiting factor for tomato plant development and productivity, reducing photosynthesis, membrane stability, fruit nutrient content, and yield. This study evaluated the efficacy of zinc oxide nanoparticles (ZnO NPs) and iron oxide nanoparticles (Fe₂O₃NPs), compared to their conventional forms (ZnSO₄·7H₂O and FeSO₄·7H₂O), in mit-igating water stress (50% ETc) under field conditions. Tomato (Solanum lycopersicum Mill.), hybrid HM 2798 was utilized as the experimental material. A split-plot field experiment was conducted with two irrigation levels (50% and 100% ETc) and five foliar treatments: Control (no application), FeSO₄·7H₂O (T1), Fe₂O₃NPs (T2), ZnONPs (T3), ZnSO₄·7H₂O (T4), with four replications. The water deficit significantly reduced the leaf area index, photo-synthetic rate, membrane stability, calcium and boron contents in fruits, and total and marketable yield. Foliar application of iron and zinc had a limited effect on tomato plant growth but increased the photosynthetic rate under both irrigation levels. Notable results included a 61% increase in total yield under full irrigation with ZnSO₄·7H₂O (T4), and a 117.3% increase in fruit iron content under water deficit with Fe₂O₃ NPs (T2). It is con-cluded that ZnSO₄·7H₂O was the most effective treatment in mitigating water stress, sug-gesting conventional zinc supplementation as a viable strategy to sustain tomato produc-tion under water restriction.
Keywords:
Solanum Lycopersicon
; drought stress
; fruit quality
; gas exchange
; mineral nutrients
1. Introduction
Water scarcity is one of the greatest global challenges of the 21st century, exacerbated by climate change, population growth, and unsustainable water resource use, especially in countries like Brazil, where agriculture accounts for about 70% of freshwater consumption [1,2]. Reduced water availability directly compromises plant productivity, particularly in sensitive crops like tomato (Locatelli et al. 2024; Wadood et al., 2024) [3,4].
Tomato (Solanum lycopersicum) is one of the most widely cultivated and consumed vegetables globally, notable for its nutritional value, richness in lycopene, vitamins, and antioxidants, which confer human health benefits [5,6]. In Brazil, tomato ranks among the top vegetables in economic value and is widely consumed fresh in salads, as sauces, and processed into pulps and extracts [7]. In the semi-arid Northeast region of Brazil, where edaphoclimatic conditions favor the cultivation of this vegetable, water scarcity is a major factor contributing to reduced yield and commercial quality [8].
Water deficit significantly compromises gas exchange in tomato plants, reducing photosynthetic rate due to stomatal closure and decreased leaf conductance, which limits CO2 assimilation [8-10]. These physiological alterations inhibit vegetative growth, resulting in reduced leaf area and biomass, and also affect the synthesis of photosynthetic pigments such as chlorophylls and carotenoids, essential for plant energy efficiency [11]. In fruits, water stress reduces the translocation of photoassimilates and mineral nutrients (such as calcium, potassium, magnesium, iron, and zinc) to the fruits and negatively impacts productivity, reducing the number and size of fruits per plant [12].
The decrease in mineral nutrients in fruits also occurs due to reduced diffusion and mass flow processes, which contribute significantly to the quantity of nutrients reaching the roots and their subsequent redistribution to the fruits [13]. These responses highlight the need for water management strategies to mitigate losses under water scarcity conditions.
Foliar application of zinc oxide nanoparticles (ZnONPs) and iron oxide nanoparticles (Fe₂O₃NPs) emerges as a promising strategy to mitigate the effects of water deficit in horticultural crops, as demonstrated by recent advances in agricultural nanotechnology [14-19]. Due to their high specific surface area, the primary mode of action of ZnONPs would be their stimulatory effect on plant metabolism through interaction with biomolecules, promoting increased enzymatic activity and the synthesis of compounds involved in combating plant oxidative stress [20].
In this sense, NPs appear to function as nano-enzyme activators, elevating the activity of antioxidant enzymes like superoxide dismutase (SOD) and catalase (CAT), while simultaneously inducing the synthesis of osmoprotectants (proline and glycine betaine) that maintain cellular osmotic pressure under drought conditions [20]. NPs containing iron (Fe) or zinc (Zn) can also act as efficient sources of Fe and Zn for plants, with slower release than conventional sources like zinc sulfate [11,21].
Zinc and iron are vital micronutrients in plants [13]. Zinc participates in the synthesis of various hormones, such as gibberellin, auxin, cytokinin, and abscisic acid, and is involved in chlorophyll production, chloroplast development, and cellular membrane integrity, providing protection, stability, and [22,23]. These processes, in turn, are associated with water stress tolerance mechanisms and plant growth and development, with implications for fruit productivity and quality [24-26]. Iron, in turn, acts as an enzymatic cofactor in chlorophyll biosynthesis and chloroplast maintenance [13], is a central component of electron transport chains (cytochromes and ferredoxin), crucial for the photochemical phase of photosynthesis [27], and regulates the activity of antioxidant enzymes (like catalase and peroxidase), protecting photosynthetic pigments from oxidative stress [28], with implications for fruit productivity [29].
In the present study, two main hypotheses were considered. The first hypothesis was that water deficit negatively affects tomato plant growth, physiological aspects, productivity, and fruit quality in terms of mineral composition. The second hypothesis was that foliar application of ZnONPs or Fe₂O₃NPs could mitigate the negative effects of water deficit on growth, productivity, and fruit mineral composition compared to their respective conventional nutrient sources.
The objective of this study was to evaluate the effect of applying zinc oxide and iron oxide nanoparticles and their conventional sources on growth, gas exchange, photosynthetic pigments, productivity, and mineral composition of tomato fruits cultivated under two irrigation levels.
2. Materials and Methods
2.1. Characterization of the Experimental Area
The experiment was conducted under field conditions from October 2023 to February 2024, at the Experimental Farm of the Center for Food Science and Technology (Centro de Ciências e Tecnologia Agroalimentar) of the Federal University of Campina Grande (UFCG), Pombal Campus, Paraíba State (PB). The Experimental Farm is located in the municipality of São Domingos, approximately 30 km from Pombal (PB). The municipality is situated in the Western region of Paraíba State, within the Sertão Paraibano mesoregion and the Sousa microregion. According to the Köppen classification, the predominant climate is type Aw, characterized as hot and humid. Annual precipitation averages around 800 mm, and the thermal amplitude is always less than 5°C. The Experimental Farm is situated at an altitude of 190 m, at the following coordinates: Latitude 6°50'4'' South, Longitude 37°53'9'' West. According to the Gaussen classification, the prevailing bioclimatic type is Mediterranean, also known as medium-dry northeastern, featuring a dry season lasting between 4 to 6 months. Area preparation (Figure A1) consisted of one plowing followed by one harrowing using a plow harrow. After this stage, soil sampling was performed in the area, within the 0–20 cm layer, for chemical and physical characterization (Table 1), following the methodology described in [30].
2.2. Treatments and Experimental Design
The experiment was arranged in a randomized block design with split plots in space. The main plots consisted of two irrigation levels, denoted as ID (50% and 100% of crop evapotranspiration - ETc), while the subplots comprised four treatments involving foliar application of iron and zinc sources, termed water deficit alleviators (WDA). The WDA treatments were: ferrous sulfate (FeSO₄·7H₂O = T1), iron oxide nanoparticles (Fe₂O₃NP = T2), zinc oxide nanoparticles (ZnONP = T3), zinc sulfate (ZnSO₄·7H₂O = T4), and a control (C). The design included four blocks, totaling 40 experimental plots.
The total experimental area measured 31.2 m in length by 20.1 m in width. Plants were spaced at 1.2 m between rows and 0.7 m within rows. Each subplot contained three 2.1-m-long rows with 12 plants. The useful plot area (2.1 m × 2.4 m) in the center of each subplot included 6 plants for evaluation. Subplots were separated by 1.5 m, and blocks by 2 m, resulting in a total area of 627.12 m².
2.3. Seedling Transplanting
Tomato seedlings (Solanum lycopersicum Mill.), hybrid HM 2798, were produced in expanded polystyrene trays (Isopor®), using a sterilized substrate composed of soil, sand, and cattle manure in a 1:1:1 ratio. Transplanting was carried out when the seedlings had 4 to 6 true leaves. The seedlings were planted on ridges measuring 0.4 m in width and 0.30 m in height. Based on the soil analysis (Table 1), basal and top-dressing fertilization was performed according to the recommendations of the Fertilizer Recommendation Manual for the State of Pernambuco [32]. For basal fertilization, the following were applied per planting row in each subplot: 134g of NPK fertilizer (10-10-10), 35g of potassium chloride (58% K₂O), and 93g of single superphosphate (18% P₂O₅). These fertilizers were incorporated into the soil during ridge formation. Additional nitrogen and potassium fertilization was applied via fertigation, consisting of three split applications of 30 kg ha-1 urea (45% N) and 12 kg ha-1 potassium chloride (58% K₂O), applied at 25, 50, and 75 days after transplanting (DAT).
2.4. Application of Treatments with Iron and Zinc Sources
As the ZnONP source, zinc oxide nanoparticles (ZnO) from Sigma Aldrich® were used, with 97% purity, particle size <100 nm, and a specific surface area of 10.8 m² g⁻¹. Suspensions of this product were prepared at a concentration of 0.25 g L⁻¹, equivalent to 200 mg of Zn L⁻¹. As the Fe₂O₃NP source, iron oxide nanoparticles (Fe₂O₃) from Sigma-Aldrich® were used, with 98% purity, particle dimensions <50 nm, and an average specific surface area of 50–245 m² g⁻¹. Suspensions of this product were prepared at a concentration of 0.30 g L⁻¹, equivalent to 200 mg of Fe L⁻¹. For conventional zinc and iron sources, the reagents ZnSO₄·7H₂O (analytical grade) and FeSO₄·7H₂O (analytical grade) were employed, respectively, both at a concentration of 4.5 g L⁻¹. Solutions were applied at a volume of 300 L ha⁻¹, corresponding to approximately 20 mL per plant. To optimize product adhesion to plants, a neutral detergent was used as a spreader-sticker agent at a concentration of 1 mL L⁻¹.
2.5. Irrigation Depths Application
After transplanting, plants were drip-irrigated using emitters spaced at 0.20 m intervals with a nominal flow rate of 1.6 L h⁻¹. Following seedling emergence and standardization of plant count and size within subplots, irrigation was applied according to different water depth levels starting 32 days after transplanting (DAT).
The total irrigation requirement (TIR) was calculated using the following expression:
Where:
TIR (Total Irrigation Requirement) – corresponds to the total initial water depth applied, in mm;
FC – Soil moisture at field capacity, %;
PWP – Soil moisture at permanent wilting point, %;
Z – Effective root zone depth (30 cm);
BD– Soil bulk density, g cm⁻³;
f – Water depletion factor (0.5).
Water application uniformity tests were conducted using the Christiansen Uniformity Coefficient (CUC) methodology proposed by [33].
The daily water volume for each irrigation depth was controlled at a standardized time, based on the ratio of emitter flow rate to the time required to meet reference evapotranspiration (ETc) proportions. As the scheduled time interval for each depth was reached, the corresponding drip tapes were sequentially shut off. The 100% ETc irrigation depth was calculated using the following expression [34]:
Where:
ETc– Crop evapotranspiration (mm day⁻¹);
ETo – Reference evapotranspiration (mm day⁻¹);
Kc – Crop coefficient (dimensionless).
The Kc values for each crop growth stage and daily ETo data were obtained using the FAO Penman-Monteith model [35]. Meteorological data during the experiment were collected from the automated weather station in São Gonçalo, Paraíba (nearest to the experimental site) via the Agritempo platform [31].
Daily irrigation depths were applied based on irrigation time, calculated using system and crop characteristics as per the following expression:
Where:
Ti – Irrigation time (hours);
ETo – Reference evapotranspiration (mm day⁻¹);
Kc – Crop coefficient (dimensionless);
A – Area occupied per plant (m²);
n – Number of emitters per plant;
q – Emitter flow rate (L h⁻¹);
Ea – Application efficiency (0.90).
2.6. Cultural Practices and Phytosanitary Control
Weed control was carried out mechanically, either manually or using hand hoes. Pest and disease control, when necessary, was performed using available natural products, such as neem extract, at a dose of 200 mL L⁻¹. For vertical staking, stakes measuring approximately 1.8 m in length were used, alternating between treatments. Following this, string was run from one end to the other of each row, tied horizontally to the stakes to help support the plants during their growth and fruiting.
2.7. Evaluated Variables
2.7.1. Plant Growth
At the pre-flowering stage, the leaf area index (LAI), plant height, and stem diameter of plants within the useful plot area were evaluated. Plant height and stem diameter were measured 2 cm above the ground. LAI values were obtained using a photosynthetically active radiation (PAR) meter (AccuPAR model LP-80, Decagon Devices, Inc., Pullman, USA). Four readings were taken in each useful plot.
2.7.2. Gas Exchange and Photosynthetic Pigments
Concurrent with the plant growth evaluation, measurements of gas exchange, pigment concentration, and cell membrane integrity were performed. Gas exchange (net photosynthesis, stomatal conductance, transpiration, and intercellular CO₂ concentration) was measured using an infrared gas analyzer (Infra-red Gas Analyzer, Li-Cor 6400, LCpro Analytical Development, Kings Lynn, UK) with a constant light source of 1,200 µmol photons m⁻² s⁻¹ and a CO₂ concentration of 370 µmol mol⁻¹.
Gas exchange measurements were taken between 7:00 and 9:00, determining the CO₂ assimilation rate – A (µmol CO₂ m⁻² s⁻¹), transpiration – E (mmol H₂O m⁻² s⁻¹), stomatal conductance – gs (mol H₂O m⁻² s⁻¹), and internal CO₂ concentration – Ci (µmol CO₂ mol⁻¹). Water use efficiency (WUE) was estimated by the A/E ratio, while the A/Ci ratio was used to estimate instantaneous carboxylation efficiency.
The contents of chlorophyll a (chl a), chlorophyll b (chl b), and carotenoid pigments (carotenoids and chlorophylls) were determined according to the methodology described by [36]. Pigments were extracted in pure, chilled acetone, subsequently filtered through 0.45 μm filter paper, and quantified by UV/VIS spectrophotometry, according to the expressions of Lichtenthaler [37]:
Where, FW and V correspond to the values of leaf fresh mass and extract volume, respectively. Membrane integrity was assessed by estimating electrolyte leakage (Ext) using the methodology suggested by Campos and Thi [38]. For this, eight leaf discs with an area of 113 mm² each were obtained from plants within the useful plot area of each experimental plot, using a copper hole punch. These discs were washed and placed in Petri dishes containing 20 mL of deionized water. The dishes were sealed and maintained at 25°C for 90 minutes. Immediately afterwards, the initial electrical conductivity of the medium (Ci) was measured using a bench conductivity meter (MB11, MS Techonopon®). Subsequently, the dishes containing the leaf discs were subjected to 80°C for 90 minutes in a drying oven (SL100/336, SOLAB®). After cooling, the final electrical conductivity (Cf) was measured. Electrolyte leakage was expressed as a percentage of the initial electrical conductivity relative to the final electrical conductivity, using the expression below, as described in Juzon-Sikora et al. [39].
2.7.3. Fruit Yield
During the fruit production phase, two harvests were carried out throughout the experiment, both performed early in the morning, manually, with the aid of bags and cardboard boxes for better storage and transport of the fruits. All fruits at the ripening stage were harvested from each useful plot. The fruits were placed in bags, all identified with their corresponding plot and treatment.
After harvest, the fruits were immediately transported to the Food Analysis and Food Chemistry and Biochemistry Laboratory of CCTA/UFCG. They were then sorted into marketable and non-marketable fruits. Fruits were considered marketable if they had a diameter equal to or greater than 35 mm, and were free of any abnormalities or diseases that could compromise their structure and flavor [40]. The following were evaluated: total number of fruits per plant, number of marketable fruits per plant, longitudinal diameter, yield of non-marketable fruits, yield of marketable fruits, and total yield.
2.7.4. Mineral Nutrients Content in Fruits
Six marketable fruits from each subplot were separated to obtain a composite sample of 100 g fresh fruit mass. These samples were dried in a forced-air oven (65-70 °C) until constant weight was achieved, to subsequently determine dry mass and water content. The dried material was then ground in a Wiley-type mill, followed by sulfuric acid digestion according to the methodology described by Tedesco et al.[41]. Digestion was performed in a Kjeldahl microdigestion block, using 0.2g samples of dried plant material in 3 mL of concentrated sulfuric acid and 1 mL of hydrogen peroxide, with an initial temperature of 180°C and a final temperature of 360°C. In the sulfuric acid digestate, the contents of the macronutrients calcium (Ca), potassium (K), phosphorus (P), and magnesium (Mg), and the micronutrients boron (B), manganese (Mn), iron (Fe), and zinc (Zn) were determined using ICP-OES (Inductively Coupled Plasma Optical Emission Spectrometry) at the Mineral Nutrition Laboratory of the Soil Science Department at the Federal University of Lavras (UFLA). The element contents obtained on a dry mass basis were converted to fresh mass basis contents by correcting the values for the fruit water content.
2.7.5. Radar Chart
A radar chart was created using the variables most impacted by the treatments mitigating water deficit (marketable yield, total yield, photosynthetic pigments, photosynthetic rate, transpiration rate, stomatal conductance, and fruit mineral content), to illustrate the magnitude of the positive (increase) and negative (decrease) effects, expressed as relative values compared to the control treatment, under both irrigation levels. The following expression was used:
Where:
RVi = Relative value of variable i
ViTn = Absolute value of variable i obtained in treatment Tn (n=1, 2, 3 or 4)
ViC = Absolute value of variable i in the control treatment
2.8. Statistical Analyses
The data were initially subjected to the Shapiro-Wilk normality test [42]. The obtained data were subjected to analysis of variance, followed by comparison of means using Tukey's test (p ≤ 0.05), as well as a Pearson’s linear correlation analysis (p ≤ 0.05). These statistical analyses were performed using R statistical software [43].
3. Results
3.1. Plant Growth
The irrigation depth corresponding to 50% of ETc reduced LAI (Leaf Area Index) by 12.6% compared to the 100% ETc depth (Figure 2a). Conversely, plant height (Figure 2b) and stem diameter (Figure 2c) were unaffected by irrigation depths or WDA treatments, except for stem diameter under 50% ETc, which was influenced by WDA treatments. Under these conditions (50% ETc), foliar application of ZnSO₄·7H₂O increased stem diameter by 25.7% relative to the control (C), though with statistically similar effects to other treatments (T1, T2, and T3).
3.2. Gas Exchange, Photosynthetic Pigments, and Electrolyte Leakage
Regarding gas exchange parameters, water deficit (50% ETc) promoted an average 7.6% decrease in CO₂ assimilation (A, Figure 3a), without significantly affecting transpiration rate (E, Figure 3b), stomatal conductance (Gₛ, Figure 3c), internal CO₂ concentration (Cᵢ, Figure 3d), or the A/E ratio (Figure 3e). Concerning WDA treatments, the application of iron oxide nanoparticles (Fe₂O₃NPs) and zinc oxide nanoparticles (ZnONPs), as well as their conventional sources (FeSO₄·7H₂O and ZnSO₄·7H₂O), increased CO₂ assimilation by 20.1% and 24.5% compared to the control under 50% and 100% Etc irrigation depths, respectively.
3.3. Gas Exchange Parameters
Foliar application of Fe₂O₃NPs (T2) increased transpiration rate (E, Figure 3b) by 34.8% under 50% ETc irrigation depth but did not differ from treatments T3 (ZnONPs) and T4 (ZnSO₄·7H₂O). Under 100% ETc, the highest E values occurred with Fe₂O₃NPs or ZnONPs, showing an 18% increase over the control. Treatments T2 and T4 increased stomatal conductance (Gₛ, Figure 3c) under 50% ETc, while T2 increased Gₛ under 100% ETc. Internal CO₂ concentration (Cᵢ) values, averaged across WDA treatments, were slightly higher under 50% ETc (Figure 3d). Water use efficiency (A/E ratio) remained unaffected by irrigation depths or WDA treatments (Figure 3e).
3.4. Photosynthetic Pigments and Electrolyte Leakage
Water deficit (50% ETc) significantly increased chlorophyll a (Figure 4a), chlorophyll b (Figure 4b), total chlorophyll (a+b, Figure 4c), carotenoid content (Figure 4e), and electrolyte leakage (Figure 4f), but did not alter the chlorophyll a/b ratio (Figure 4d). WDA treatments did not affect chlorophyll a, b, total chlorophyll, or carotenoid content at any irrigation level, nor the a/b ratio under 50% ETc. However, under full irrigation (100% ETc), treatments T1, T2, and T4 showed similar values—56.5% higher than the control.
Regarding electrolyte leakage, under water deficit (50% ETc), all Fe/Zn source treatments (T1-T4) reduced leakage by 39.1% versus control. Under full irrigation, T2 and T4 reduced leakage by 20.8% versus control. Water deficit increased electrolyte leakage (Ext) by 21.9% compared to 100% ETc (Figure 4f). Under 50% ETc, treatments T1-T4 all reduced Ext versus control but did not differ from each other. Under 100% ETc, treatments T2-T4 lowered Ext versus control but were statistically similar to T1.
3.5. Fruit Productivity
The total number of fruits per plant (Figure 5a) was not significantly affected by the irrigation depths. On the other hand, water deficit reduced the number of marketable fruits per plant (Figure 5b), the transverse diameter (Figure 5c), the longitudinal diameter (Figure 5d), the total yield (Figure 5e), and the marketable fruit yield (Figure 5f). Water restriction decreased total fruit production by 34.9% and marketable production by 47.9% compared to full irrigation (100% ETc). Under water deficit, treatment T4 increased the total number of fruits by 55.0% compared to the control, but did not alter total fruit yield. Under full irrigation, treatment T4 resulted in a higher number of fruits per plant compared to treatments T1 and T3 (which did not differ from the control and T2 treatments), and increased total yield by 61% compared to the control. The WDA treatments did not modify the quantity of marketable fruits per plant under the 50% ETc irrigation depth. However, under full irrigation, treatment T4 outperformed treatments T1, T2, and T3, but not the control treatment. Under both irrigation depths, the WDA treatments did not influence the transverse diameter, the longitudinal diameter of the fruits, nor the marketable fruit yield.
3.6. Mineral Element Content in Fruits
The effect of irrigation depths on boron (B, Figure 6a) content in fruits depended on the WDA treatments. Calcium (Ca, Figure 6b) content was reduced by 16.5% due to water restriction. Water deficit did not affect the content of iron (Fe, Figure 6c), potassium (K, Figure 6d), manganese (Mn, Figure 6e), magnesium (Mg, Figure 6f), phosphorus (P, Figure 6g) or zinc (Zn, Figure 6h).
The 50% ETc irrigation depth reduced boron (B) content by 32.9% in treatment T4, but did not alter the content of this nutrient in the other treatments. Under the 100% ETc irrigation depth, treatment T4 increased boron content by 39.6% and calcium (Ca) content by 24.7% compared to the control treatment. The WDA treatments particularly affected iron (Fe) content. Under water restriction, treatments T1 (FeSO₄·7H₂O) and T2 (Fe₂O₃NPs) increased the content of this nutrient by 67.5% and 117.3%, respectively, compared to the control. Under full irrigation, treatments T1 and T2 increased iron content by 24.5% and 88.8%, respectively. The WDA treatments influenced zinc (Zn) content only under the 50% ETc irrigation depth, where treatment T4 increased zinc content by 18.5% compared to the control treatment.
Figure 7 below presents the relative values (Rv) of selected variables to illustrate the magnitude of the effects of the WDA treatments under both tested irrigation depths. Iron content was excluded due to the high percentage increase in treatments T1 and T4, which would obscure visualization of effects on other treatments for this variable.
Under water deficit, treatment T1 enhanced photosynthetic rate (A), transpiration rate (E), carotenoids (Carot), and calcium (Ca) content in fruits (Figure 7a). Under full irrigation, T1 increased values of A, E, total chlorophyll content (Chlt), chlorophyll a, and carotenoids (Figure 7b). Under both irrigation depths, treatment T2 elevated values of A, E, stomatal conductance (gs), Ca content, and total (TYF) and marketable (YMF) fruit yield, in addition to increasing chlorophyll a and carotenoids under 100% ETc. Under water restriction, treatment T3 most markedly increased values of A, E, and Ca content in fruits, while under full irrigation it elevated A, E, and chlorophylls (a and b). Conversely, treatment T4 promoted increases in gs, A, E, and Ca and Zn content under water deficit. Under full irrigation, treatment T4 enhanced A, E, chlorophylls (a and b), Ca and B content, and fruit yield (TYF and YMF).
Pearson’s linear correlation analysis (Figure 8) revealed significant positive correlations between number of marketable fruits and B content (r = 0.57), Fe content and physiological parameters A/Ci (r = 0.75), A (r = 0.68), E (r = 0.46), gs (r = 0.62), and Mn content (r = 0.50). Additionally, B content correlated positively with total fruit production (r = 0.55), K content showed direct relationships with Mg (r = 0.52) and P (r = 0.54), P content strongly correlated with Mn content (r = 0.86). Conversely, electrolyte leakage exhibited negative correlations with, E (r = -0.52), A/E (r = -0.76) e A/i (r = -0.69).
4. Discussion
4.1. Plant Growth
This study evaluated the potential of iron (Fe₂O₃NPs), and zinc (Fe₂O₃NPs) nano-oxide as well as their conventional forms (FeSO₄·7H₂O and ZnSO₄·7H₂O), to mitigate the negative effects of water deficit (50% ETc) on growth traits, gas exchange, photosynthetic pigments, and nutrient content in tomato fruits. The results demonstrate that water deficit reduced leaf area index (LAI), photosynthetic rate, and tomato yield. Generally, water stress decreases cell turgor, essential for proper cellular metabolism such as photosynthesis [44], enzymatic activity [46]. Foliar application of Fe₂O₃NPs, ZnONPs, FeSO₄·7H₂O, and ZnSO₄·7H₂O partially attenuated these effects, corroborating previous studies highlighting nanoparticles potential in mitigating water stress [17,18].
The 12.6% reduction in LAI under 50% ETc is attributed to decreased cell expansion and leaf growth [47]. However, the maintenance of plant height and stem diameter suggests that ZnSO₄·7H₂O (T4) may promote osmotic adjustments or improved water-use efficiency, as observed in potatoes [9] and green peppers [12] and tomatoes [65]. The 25.7% increase in stem diameter with T4 under water deficit reinforces role of in auxin synthesis and structural integrity [48, 49].
4.2. Gas Exchange, Photosynthetic Pigments, and Electrolyte Leakage
Water deficit reduced CO₂ assimilation (A) by 7.6%, but treatments with Fe₂O₃NPs (T2) and ZnO NPs (T3) increased A by up to 24.5% under full irrigation, likely due to enhanced Rubisco activity and electron transport [9]. The 34.8% increase in transpiration (E) with T2 under 50% ETc may link to nanoparticle-mediated stomatal regulation, balancing water loss and photosynthesis [16,43]. Unchanged water-use efficiency (A/E) suggests treatments prioritized productivity over water conservation [50].
Photosynthetic pigments are key cellular components affected by water deficit [51]. Water restriction typically reduces pigment content, especially chlorophyll (Qiao et al. 2024)[44], due to oxidative damage from reactive oxygen species and increased activity of oxidases (e.g., catalase, peroxidase, SOD) [52]. However, chlorophyll levels may increase or decrease depending on stress severity and plant species [52,53]—e.g., moderate stress elevated chlorophyll in Lonicera caerulea. Here, increased chlorophyll/carotenoid content under deficit may indicate an adaptive response to maximize light capture [47], compounded by reduced leaf area concentrating pigments per unit area.
The 39.1% reduction in electrolyte leakage with T1–T4 indicates NPs and conventional Fe and Zn sources strengthened cell membranes, likely via antioxidant induction [23]. Zinc activates oxidative stress defense enzymes (e.g., SOD), aids tryptophan (IAA precursor) synthesis, and supports cell division and membrane integrity [23,26]
4.3. Yield, Fruit Nutrient Content, and Variable Correlations
Water deficit negative impact on tomato yield aligns with prior studies [9,29,53]. The 47.9% decrease in marketable yield under 50% ETc reflects stress-induced reductions in fruit number and size [29,54], consistent with lower LAI, photosynthetic rate, and higher electrolyte leakage. However, T4 (ZnSO₄·7H₂O) increased total yield by 61% under full irrigation, underscoring the zinc role in cell division and fruit development [13].
Positive correlations between fruit boron (B) content and fruit number (r = 0.57) or marketable yield (r = 0.55) highlight the B importance in cell wall integrity and postharvest quality [13]. Though fruit calcium (Ca) content did not correlate significantly with yield, deficit reduced Ca by 16.5%. Both B and Ca directly affect cell wall plasticity/elongation [55,56]. These nutrients move via diffusion and mass flow to roots water deficit impedes their uptake and phloem-immobile translocation to fruits [13,57].
Fe₂O₃ NPs (T2) increased fruit iron content by up to 117.3% under deficit, confirming Fe oxide NPs efficacy in nutrient delivery under low water availability, as seen in wheat [58]. Chlorophyll content remained unchanged with Fe treatments, but Fe correlation with photosynthetic parameters (A, gs) confirms its central role in chlorophyll synthesis/electron transport [9,28,59]. Under water stress, ZnO NPs did not alter fruit zinc content, while ZnSO₄·7H₂O increased it by 18.5%. Performance differences between NPs may stem from physical properties: Fe₂O₃ NPs (<50 nm; 50–245 m² g⁻¹ surface area) vs. ZnO NPs (<100 nm; 10.8 m² g⁻¹). The mechanisms governing foliar uptake of NPs are complex and not yet fully elucidated. It has been proposed that NP efficacy depends on cellular uptake routes: passive transport (symplastic and apoplastic pathways), endocytosis, or transmembrane protein-mediated transport [6,63]. Internalized Fe₂O₃ and ZnO NPs may reach organelles (chloroplasts, mitochondria), releasing Fe and Zn to modulate electron transport chains and associated processes (e.g., oxidative phosphorylation) [36]. However, NP entry is limited by pore size selectivity and accessibility [19,64,65].
The nutritional enhancement of fruit vegetables is particularly critical in Brazilian semi-arid region. These areas typically feature soils with inherently low availability of micronutrients such as iron and zinc, a characteristic attributed to the region's soil composition [66]. This scarcity can lead to human malnutrition, notably iron deficiency. The efficient foliar application of these two nutrients presents a highly effective method to increase their concentration in these vegetables, thereby improving the nutritional quality of the harvested produce [12].
5. Conclusions
Water deficit (50% ETc) significantly compromised tomato LAI, photosynthesis, and total/marketable yield. Foliar application of nano-oxides (Fe₂O₃ NPs, ZnO NPs) and conventional sources (FeSO₄·7H₂O, ZnSO₄·7H₂O) partially mitigated these losses, with ZnSO₄·7H₂O standing out—increasing stem diameter by 25.7% and total yield by 61% under full irrigation. NPs improved CO₂ assimilation (up to 24.5%) and reduced electrolyte leakage, indicating membrane protection. Nevertheless, ZnSO₄·7H₂O showed greater efficiency in stress mitigation, suggesting its potential for sustaining productivity under water scarcity. Thus, zinc supplementation, particularly in conventional form, is a viable strategy for tomato production in water-limited scenarios.
Author Contributions
ECA, JLAR, FHFP, KNF, ACSR, RHCRA, and JEJS, participated in the experiment setup, acquisition of data collection, data analysis and article writing. JLAR, CSGR, GL and FBN proposed the design and design of the study and writing of the article. FVSS, JSLS, LCS and RN participated in the discussed the results and contributed to the preparation of the manuscript.
Funding
This research was funded by FAPESQ (Foundation to support research in the state of Paraíba), Fapesq/PB Edital Universal 09/2021 SEECT/Fapesq. grant number 3071/2021. All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
The data used to support the findings of this study are included within the article.
Acknowledgments
We sincerely thank the Department of Soil Science at the Federal University of Lavras, lavras-MG for laboratory support for plant material analysis.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Figure A1.
Partial view of the experimental area showing the soil tillage (a), installation of the drip irrigation system (b), seedlings initial growth (c) and the plants on the pre-flowering phase (d).
Figure A1.
Partial view of the experimental area showing the soil tillage (a), installation of the drip irrigation system (b), seedlings initial growth (c) and the plants on the pre-flowering phase (d).

References
- United Nations Water (UN-Water). The United Nations World Water Development Report 2021: Valuing Water; UNESCO: Paris, France, 2021. [Google Scholar]
- Agência Nacional de Á guas e Saneamento Básico (ANA). Conjuntura dos recursos hídricos no Brasil 2023: Relatório pleno; ANA: Brasília, Brazil, 2023. Available online: https://www.snirh.gov.br/portal/snirh/centraisde-conteudos/conjuntura-dos-recursos-hidricos/conjuntura_2023.pdf (accessed on 10 June 2024).
- Locatelli, S.; Barrera, W.; Verdi, L.; Nicoletto, C.; Marta, A.D.; Mauricieri, C. Modelling the response of tomato on deficit irrigation under greenhouse conditions. Scientia Horticulturae 2024, 326, 112770. [Google Scholar] [CrossRef]
- Wadood, A.; Hameed, A.; Akram, S.; Ghaffar, M. Unraveling the impact of water deficit stress on nutritional quality and defense response of tomato genotypes. Front. Plant Sci. 2024, 15, 1403895. [Google Scholar] [CrossRef] [PubMed]
- Caseiro, A.; Ascenso, A.; Costa, A.; Creagh-Flynn, J.; Johnson, M.; Simões, S. Lycopene in human health. LWT - Food Sci Technol 2020, 127, 109323. [CrossRef]
- Li, M.; Wu, X.; Zhunag, W.; Xia, L.; Chen, Y.; Wu, C.; Rao, Z.; Du, L.; Zhao, R.; Yi, M.; et al. Tomato and lycopene and multiple health outcomes: Umbrella review. Critical Reviews Food Sci. Nutr. 2021, 61, 2063–2076. [Google Scholar] [CrossRef]
- EMBRAPA. Embrapa Hortaliças. Tomate: Produção e Mercado no Brasil; Documentos 245; Embrapa: Brasília, Brazil, 2022. [Google Scholar]
- Oliveira, E.C.; Carvalho, J.A.; Rezende, F.C.; Freitas, W.A.; Ramos, M.M. Stomatal regulation and photosynthetic performance in tomato under water deficit. Environ. Exp. Botany 2021, 187, 104465. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, W.; Kang, Y.; Shi, M.; Yang, X.; Li, H.; Yu, H.; Wang, Y.; Qin, S. Application of different foliar iron fertilizers for improving the photosynthesis and tuber quality of potato (Solanum tuberosum L.) and enhancing iron biofortification. Chem. Biological Tech. Agriculture 2022, 9, 79. [CrossRef]
- Haghpanah, M.; Hashemipetroudi, S.; Arzani, A.; Araniti, F. Drought tolerance in plants: Physiological and molecular responses. Plants 2024, 13, 2962. [Google Scholar] [CrossRef]
- Li, J.; Zafar, S.; Javaid, A.; Perveen, S.; Hasnain, Z.; Ihtisham, M.; Abbas, A.; Usman, M.; El-Sappah, A.H.; Abbas, M.; et al. Zinc nanoparticles (ZnNPs): High-fidelity amelioration in turnip (Brassica rapa L.) production under drought stress. Sustainability. 2023; 15, 6512. [Google Scholar] [CrossRef]
- Martins, B.L.R.; Ferreira, K.N.; Rocha, J.L.A.; Araujo, R.H.C.R.; Lopes, G.; Santos, L.C.d.; Bezerra Neto, F.; Sá, F.V.d.S.; Silva, T.I.d.; da Silva, W.I.; et al. Nano ZnO and Bioinoculants Mitigate Effects of Deficit Irrigation on Nutritional Quality of Green Peppers. Horticulturae 2024, 10, 969. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: London, UK, 2012. [Google Scholar]
- Uresti-Porras, J.G.; Cabrera-De La Fuente, M.; Benavidez-Mendoza, A.; Olivares-Sáenz, E.; Cabrera, R.I.; Juárez-Maldonado, A. Effect of graft and nano ZnO on nutraceutical and mineral content in bell pepper. Plants 2021, 10, 2793. [Google Scholar] [CrossRef]
- Semida, W.M.; Abdelkhalik, A.; Mohamed, G.F.; Abd El-Mageed, T.A.; Abd El-Mageed, S.A.; Rady, M.M.; Ali, E.F. Foliar application of zinc oxide nanoparticles promotes drought stress tolerance in eggplant (Solanum melongena L.)Plants. 2021; 10, 421. [Google Scholar] [CrossRef]
- Azam, M.; Bhatti, H.N.; Khan, A.K.; Zafar, L.; Iqbal, M. Zinc oxide nano-fertilizer application (foliar and soil) effect on the growth, photosynthetic pigments and antioxidant system of maize cultivar. Biocat. Agric. Biotechnology 2022, 42, 102343. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Farooq, A.; Javad, S.; Jabeen, K.; Shah, A.A.; Ahmad, A.; Shah, A.N.; Alyemeni, M.N.; Mosa, W.F.A.; Abbas, A. Effect of calcium oxide, zinc oxide nanoparticles and their combined treatments on growth and yield attributes of Solanum lycopersicum L. J. King Saud Univ- Sci. 2023; 35, 102647. [Google Scholar] [CrossRef]
- Wu, H.; Li, Z. Nano-enabled agriculture: How do nanoparticles cross barriers in plants? Plant Communications 2022, 3, 100346. [Google Scholar] [CrossRef]
- Dokoupil, L.; Mlcek, J. Foliar application of ZnO-NPS influences chlorophyll fluorescence and antioxidants pool in Capsicum annum L. under salinity. Horticulturae 2022, 8, 908. [Google Scholar] [CrossRef]
- García-López, J.I.; Niño-Medina, G.; Olivares-Sáenz, E.; Lira-Saldivar, R.H.; Barriga-Castro, E.D.; Vázquez-Alvarado, R.; Rodríguez-Salinas, P.A.; Zavala-García, F. Foliar application of zinc oxide nanoparticles and zinc sulfate boosts the content of bioactive compounds in habanero peppers. Plants 2019, 8, 254. [Google Scholar] [CrossRef] [PubMed]
- Baybordi, A. Zinc in Soils and Crop Nutrition; Parivar Press: Tehran, Iran, 2006. [Google Scholar]
- Elsheery, N.I.; Helaly, M.N.; El-Hoseiny, H.M.; Alam-Eldein, S.M. Zinc oxide and silicone nanoparticles to improve the resistance mechanism and annual productivity of salt-stressed mango trees. Agronomy 2020, 10, 558. [Google Scholar] [CrossRef]
- Ybaez, Q.E.; Sanchez, P.B.; Badayos, R.B. Synthesis and characterization of nano zinc oxide foliar fertilizer and its influence on yield and postharvest quality of tomato. Philippine Agricultural Scientist 2020, 103, 55–65. [Google Scholar] [CrossRef]
- Ganguly, R.; Sarkar, A.; Dasgupta, D.; Acharya, K.; Keswani, C.; Popova, V.; Minkina, T.; Maksimov, A.Y.; Chakraborty, N. Unravelling the efficient applications of zinc and selenium for mitigation of abiotic stresses in plants. Agriculture 2022, 12, 1551. [Google Scholar] [CrossRef]
- Ahmed, N.; Zhang, B.; Chachar, Z.; Li, J.; Xiao, G.; Wang, Q.; Hayat, F.; Deng, L.; Narejo, M.; Bozdar, B.; et al. Micronutrients and their effects on horticultural crop quality, productivity and sustainability. Scientia Horticulturae 2024, 323, 112512. [Google Scholar] [CrossRef]
- Nagao, R.; Kato, K.; Hamaguchi, U.; Ueno, Y.; Tsuboshita, N.; Shimizu, S.; Furutani, S.; Ehira, S.; Nakajima, Y.; Kawakami, K.; et al. Structure of a monomeric photosystem I core associated with iron-stress-induced-A proteins from Anabaena sp. PCC 7120. Nature Communications 2023, 14, 920. [Google Scholar] [CrossRef]
- Nasar, J.; Wang, G.Y.; Ahmad, S.; Muhammad, I.; Zeeshan, M.; Gitari, H.; Adnan, M.; Fahad, S.; Khalid, M.H.B.; Zhou, X.B.; et al. Nitrogen fertilization coupled with iron foliar application improves the photosynthetic characteristics, photosynthetic nitrogen use efficiency, and the related enzymes of maize crops under different planting patterns. Front. Plant Sci. 2022, 13, 988055. [Google Scholar] [CrossRef]
- Zahedifar, M.; Moosavi, A.A.; Gavili, E.; Ershadi, A. Tomato fruit quality and nutrient dynamics under water deficit conditions: The influence of an organic fertilizer. PLoS ONE 2025, 20, e0310916. [Google Scholar] [CrossRef]
- EMBRAPA. Manual of Soil Analysis Methods, 2nd ed.; Centro Nacional de Pesquisa em Solo: Rio de Janeiro, Brazil, 2011. [Google Scholar]
- AGRITEMPO. Agrometeorological monitoring system: meteorological stations for the state of Paraiba. 2023. Disponível em: https://www.agritempo.gov.br/agritempo/jsp/Estacao/index.jsp?siglaUF=PB. Acessado em 15 outubro 2023.
- Cavalcante, F.J.A. Fertilization Recommendations for the State of Pernambuco (Brazil), 2nd ed.; Instituto Agronômico de Pernambuco: Recife, Brazil, 2008. [Google Scholar]
- Christiansen, J.E. Irrigation by Sprinkling; University of California Agricultural Experiment Station: Berkeley, CA, USA, 1943. [Google Scholar]
- Jensen, M.E. Water consumption by agriculture plants. In Water Deficit and Plant Growth; Kozlowski, T.T., Ed.; Academic Press: New York, NY, USA,; 1968; Volume 2, pp. 1–22. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration—Guidelines for Computing Crop Water Requirements; FAO Irrigation and Drainage Paper No. 56; FAO: Rome, Italy, 1998. [Google Scholar]
- Jafari, A.; Hatami, M. Foliar-applied nanoscale zero-valent iron (nZVI) and iron oxide (Fe3O4) induce differential responses in growth, physiology, antioxidative defense and biochemical indices in Leonurus cardiaca L. Environmental Research 2022, 215, 114254. Environmental Research 2022, 215, 114254. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods in Enzymology 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Campos, P.; Thi, A. Effects of an abscisic acid pretreatment on membrane leakage and lipid composition of Vigna unguiculata leaf discs subjected to osmotic stress. Plant Science 1997, 130, 11–18. [Google Scholar] [CrossRef]
- Juzon-Sikora, K.; Lasko´s, K.; Warchoł, M.; Czyczyło-Mysza, I.M.; Dziurka, K.; Grzesiak, M.; Skrzypek,E. Water Relations and Physiological Traits Associated with the Yield Components of Winter Wheat (Triticum aestivum L.). Agriculture 2024, 14, 1887. [Google Scholar] [CrossRef]
- Gayet, J.P.; Bleinroth, E.W.; Matallo, M.; Garcia, E.E.C.; Garcia, A.E.; Ardito, E.F.G.; Bordin, M.R. Tomate para Exportação: Procedimentos de Colheita e Pós-colheita; Série Publicações Técnicas FRUPEX 13; EMBRAPA-SPI: Brasília, Brazil, 1995. [Google Scholar]
- Tedesco, M.J.; Gianello, C.; Bissani, C.A.; Bohnen, H.; Volkweiss, S.J. Analysis of Soil, Plants and Other Materials, 2nd ed.; Universidade Federal do Rio Grande do Sul: Porto Alegre, Brazil, 1995. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (Complete Samples). Biometrika 1965, 52, 591–609. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing. Viena: Austria, 2023.
- Qiao, M.; Hong, C.; Jiao, Y.; Hou, S.; Gao, H. Impacts of drought on photosynthesis in major food crops and the related mechanisms of plant responses to drought. Plants 2024, 13, 1808. [Google Scholar] [CrossRef]
- Colmenero-Flores, J.M.; Arbona, V.; Morillon, R.; Gómez-Cadenas, A. Salinity and water deficit. In The Genus Citrus; Talon, M., Caruso, M., Gmitter, F.G., Eds.; Woodhead Publishing: Cambridge, UK, 2020; pp. 291–309. [Google Scholar] [CrossRef]
- Kundel, D.; Lori, M.; Fliessbach, A.; van Kleunen, M.; Meyer, S.; Mader, P. Drought effects on nitrogen provisioning in different agricultural systems: Insights gained from a field experiment. Nitrogen 2021, 2, 1–17. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Moller, I.M.; Murphy, A. Plant Physiology and Development, 6th ed.; Sinauer Associates: Sunderland, MA, USA, 2017. [Google Scholar]
- Cakmak, I.; Marschner, H. Increase in membrane permeability and exudation in roots of zinc deficient plants. J. Plant Physiology 1988, 132, 356–361. [Google Scholar] [CrossRef]
- Hassan, M.U.; Aamer, M.; Chattha, M.U.; Haiying, T.; Shahzad, B.; Barbanti, L.; Nawaz, M.; Rasheed, A.; Afzal, A.; Liu, Y.; et al. The critical role of zinc in plants facing the drought stress. Agriculture 2020, 10, 396. [Google Scholar] [CrossRef]
- Shewangizaw, B.; Kassie, K.; Assefa, S.; Lemma, G.; Gete, Y.; Getu, D.; Getanh, L.; Shegaw, G.; Manaze, G. Tomato yield, and water use efficiency as affected by nitrogen rate and irrigation regime in the central low lands of Ethiopia. Scientific Reports 2024, 14, 13307. [Google Scholar] [CrossRef]
- Suárez-Salazar, J.C.; Guaca-Cruz, L.; Quiceno-Mayo, E.J.; Ortiz-Morea, F.A. Photosynthetic responses and protective mechanisms under prolonged drought stress in cocoa. Pesquisa Agropecuária Brasileira 2024, 59, e03543. [Google Scholar] [CrossRef]
- Yang, Y.; Nan, R.; Mi, T.; Song, Y.; Shi, F.; Liu, X.; Wang, Y.; Sun, F.; Xi, Y.; Zhang, C. Rapid and nondestructive evaluation of wheat chlorophyll under drought stress using hyperspectral imaging. Inter. J. Molec. Sciences 2023, 24, 5825. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Lu, Y.; Guo, L.; Liu, Y.; Li, M.; Zhang, B.; Zhang, B.; Zhang, L.; Qin, D.; Huo, J. Effects of drought stress on photosynthesis and chlorophyll fluorescence in blue honeysuckle. Plants 2024, 13, 2115. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Dash, P.K.; Das, D.; Das, S. Growth, yield and water productivity of tomato as influenced by deficit irrigation water management. Environmental Processes 2023, 10, 10. [Google Scholar] [CrossRef]
- Wdowiak, A.; Podgórska, A.; Szal, B. Calcium in plants: An important element of cell physiology and structure, signaling, and stress responses. Acta Physiologiae Plantarum 2024, 46, 108. [Google Scholar] [CrossRef]
- Vera-Maldonado, P.; Aquea, F.; Reyes-Díaz, M.; Cárcamo-Fincheira, P.; Soto-Cerda, B.; Nunes-Nesi, A.; Inostroza-Blancheteau, C. Role of boron and its interaction with other elements in plants. Front. Plant Sci. 2024, 15, 1332459. [Google Scholar] [CrossRef]
- Peroni, E.C.; Messoas, R.S.; Gallo, V.; Bardwosko, J.M.; Souza, E.R.; Avola, L.D.; Bamberg, A.L.; Rambo, C.V. Mineral content and antioxidant compounds in strawberry fruit submitted to drought stress. Food Science and Technology 2019, 39, 245–254. [Google Scholar] [CrossRef]
- Zia-ur-Rehman, M.; Mfarrej, M.F.B.; Usman, M.; Anayatullah, S.; Rizwan, M.; Alharby, H.F.; Zeid, I.M.A.; Alabdallah, N.M.; Ali, S. Effect of iron nanoparticles and conventional sources of Fe on growth, physiology and nutrient accumulation in wheat plants grown on normal and salt-affected soils. Journal of Hazardous Materials 2023, 458, 131861. [Google Scholar] [CrossRef]
- Gao, D.; Zhao, S.; Huang, R.; Geng, Y.; Guo, L. The effects of exogenous iron on the photosynthetic performance and transcriptome of rice under salt-alkali stress. Agronomy 2024, 14, 1253. [Google Scholar] [CrossRef]
- Maret, W.; Sandstead, H.H. Zinc requirements and the risks and benefits of zinc supplementation. J. Trace Elements in Med Biology 2006, 20, 3–18. [Google Scholar] [CrossRef]
- Jeng, S.S.; Chen, Y.H. Association of zinc with anemia. Nutrients 2022, 14, 4918. [Google Scholar] [CrossRef]
- Cançado, R.D. Iron deficiency anemia in women: Pathophysiological, diagnosis, and practical management. Rev Associação Méd. Brasileira 2023, 69 (Suppl. 1). [CrossRef]
- Wei, L.; Liu, J.; Jiang, G. Nanoparticle-specific transformations dictate nanoparticle effects associated with plants and implications for nanotechnology use in agriculture. Nature Communications 2024, 15, 7389. [Google Scholar] [CrossRef]
- Yadav, M. Nanoparticle-facilitated targeted nutrient delivery in plants: Breakthroughs and mechanistic insights. Plant Nano Biology 2025, 12, 100156. [Google Scholar] [CrossRef]
- Alordzinu, K.E.; Appiah, S.A.; AL Aasmi, A.; Darko, R.O.; Li, J.; Lan, Y.; Adjibolosoo, D.; Lian, C.; Wang, H.; Qiao, S. Evaluating the influence of deficit irrigation on fruit yield and quality indices of tomatoes grown in sandy loam and silty loam soils. Water 2022, 14, 1753. [Google Scholar] [CrossRef]
- Oliveira, M.L.J.; Valladares, G.S.; Vieira, J.S.; Coelho, R.M. Availability and spatial variability of copper, iron, manganese and zinc in soils of the State of Ceará, Brazil. Revista Ciência Agronômica 2018, 49, 371-380.
- Nascimento Júnior, A.L.; Souza, S.; Paiva, A.Q.; Souza, LD.; Souza-Filho, L.F.; Fernandes Filho, E.I.; Schaefer, C.E.G.R.; Santos, J.A.G.; Bomfim, M.R.; Silva, E.F.; Fernandes, A.C.O.; Xavier, F.A.S.bConcentration and variability of soil trace elements in an agricultural area in a semiarid region of the Irecê Plateau, Bahia, Brazil. Geoderma Regional, 2021, 24, e00351. [CrossRef]
Figure 1.
Climatological data on maximum (T max) and minimum (T min) air temperature air temperature, maximum (RH max) and minimum (RH min) air relative humidity and rainfall during the experimental period in the field [31].
Figure 1.
Climatological data on maximum (T max) and minimum (T min) air temperature air temperature, maximum (RH max) and minimum (RH min) air relative humidity and rainfall during the experimental period in the field [31].
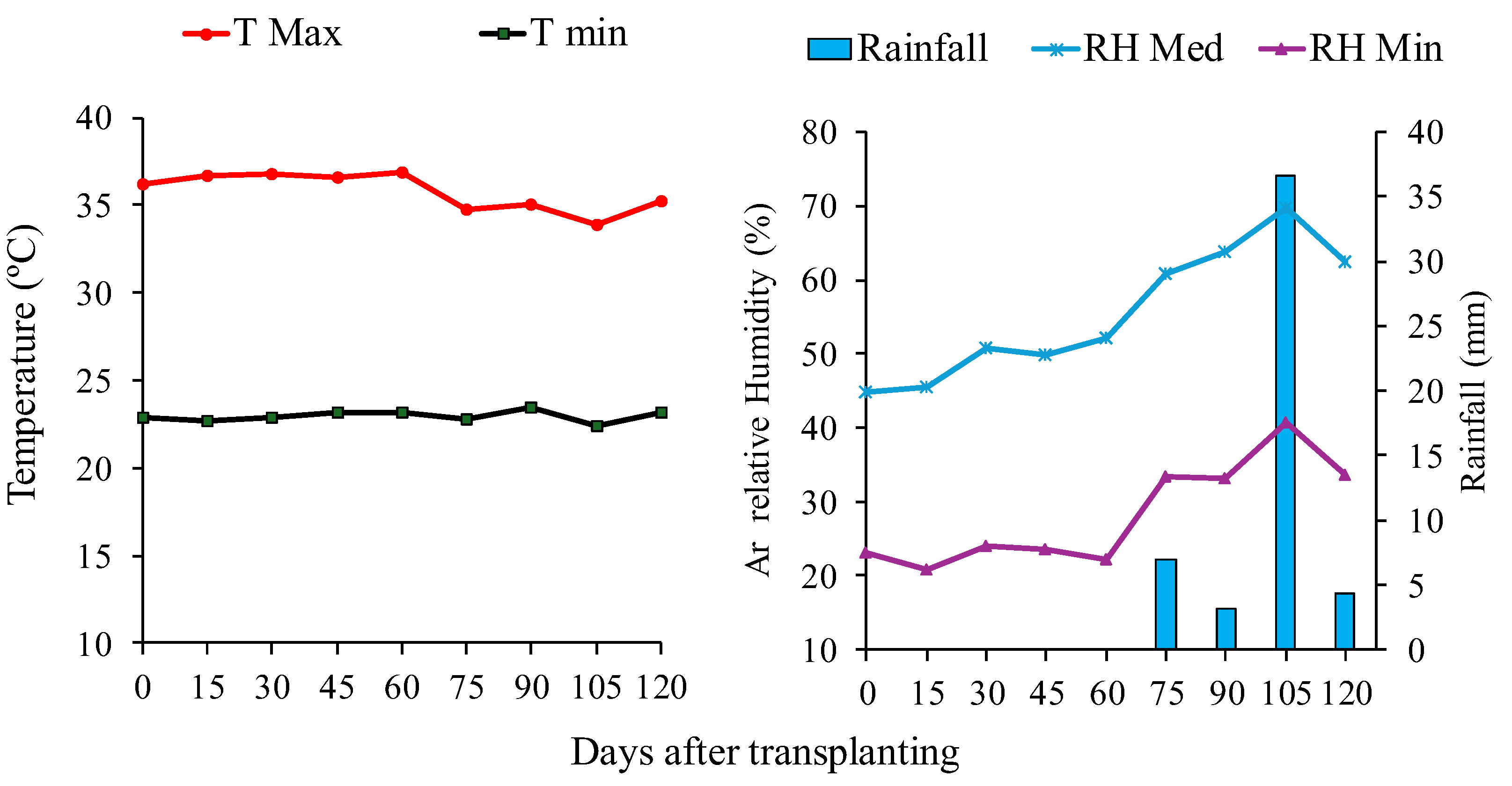
Figure 2.
Leaf area index (a), plant height (b), and stem diameter (c) of tomato hybrid 'HM' under different foliar application treatments and irrigation regimes: control (C), FeSO₄·7H₂O (T1), Fe₂O₃NPs (T2), ZnONPs (T3), and ZnSO₄·7H₂O (T4), subjected to both deficit (50% ETc) and full irrigation (100% ETc). Data represent means ± S.E. Lowercase letters compare treatments (iron and zinc sources), while uppercase letters compare irrigation levels (50% ETc and 100% ETc).
Figure 2.
Leaf area index (a), plant height (b), and stem diameter (c) of tomato hybrid 'HM' under different foliar application treatments and irrigation regimes: control (C), FeSO₄·7H₂O (T1), Fe₂O₃NPs (T2), ZnONPs (T3), and ZnSO₄·7H₂O (T4), subjected to both deficit (50% ETc) and full irrigation (100% ETc). Data represent means ± S.E. Lowercase letters compare treatments (iron and zinc sources), while uppercase letters compare irrigation levels (50% ETc and 100% ETc).
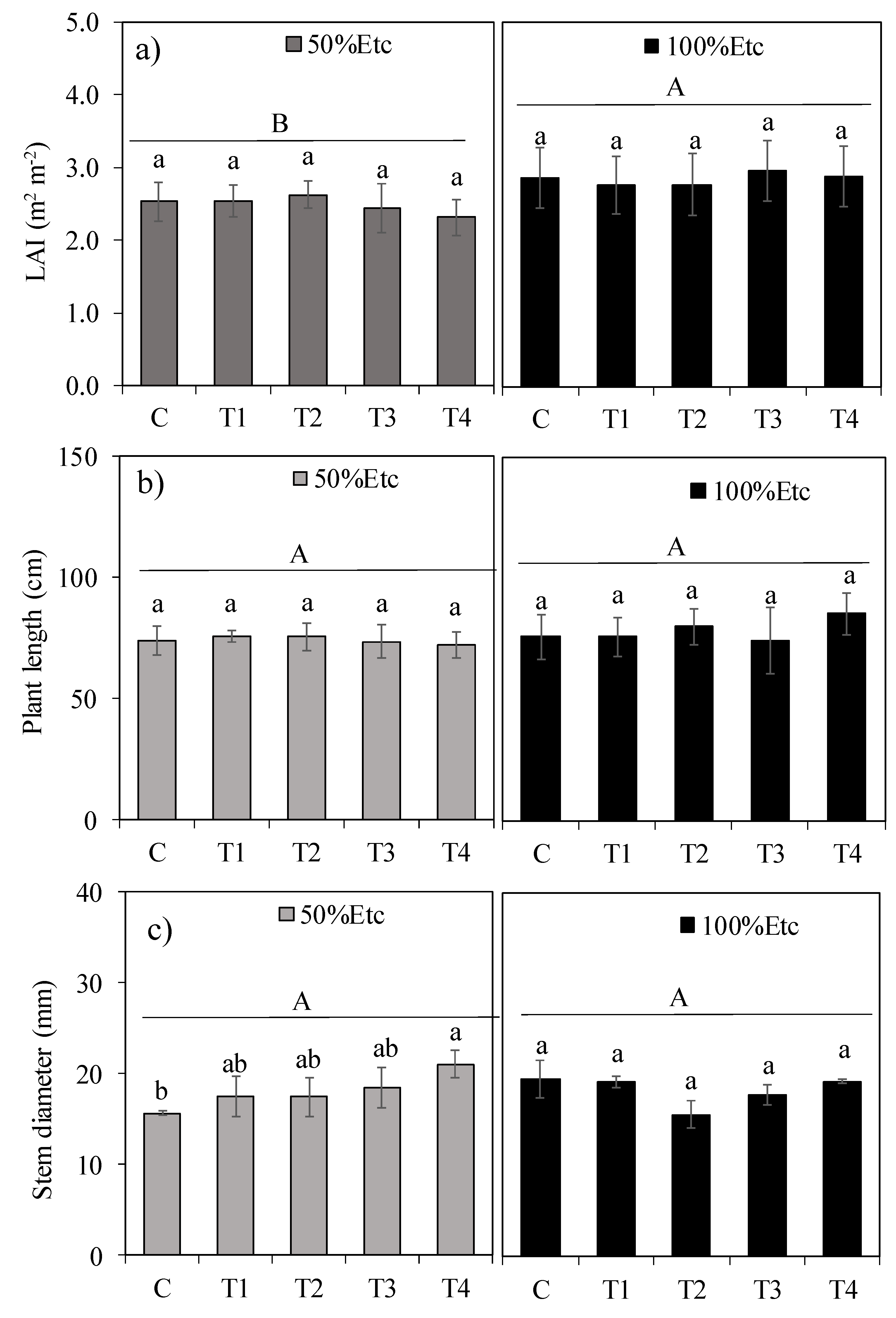
Figure 3.
CO2 assimilation rate (a), transpiration rate (b), stomatal conductance (c), internal CO2 concentration (d), water use efficiency © and sap leakage (f) of tomato hybrid ‘HM’ under different foliar application treatments and irrigation regimes: control (C), FeSO₄·7H₂O (T1), Fe₂O₃NPs (T2), ZnONPs (T3), and ZnSO₄·7H₂O (T4), subjected to both deficit (50% Etc) and full irrigation (100% Etc). Data represent means ± S.E. Lowercase letters compare treatments (iron and zinc sources), while uppercase letters compare irrigation levels (50% Etc and 100% Etc.
Figure 3.
CO2 assimilation rate (a), transpiration rate (b), stomatal conductance (c), internal CO2 concentration (d), water use efficiency © and sap leakage (f) of tomato hybrid ‘HM’ under different foliar application treatments and irrigation regimes: control (C), FeSO₄·7H₂O (T1), Fe₂O₃NPs (T2), ZnONPs (T3), and ZnSO₄·7H₂O (T4), subjected to both deficit (50% Etc) and full irrigation (100% Etc). Data represent means ± S.E. Lowercase letters compare treatments (iron and zinc sources), while uppercase letters compare irrigation levels (50% Etc and 100% Etc.
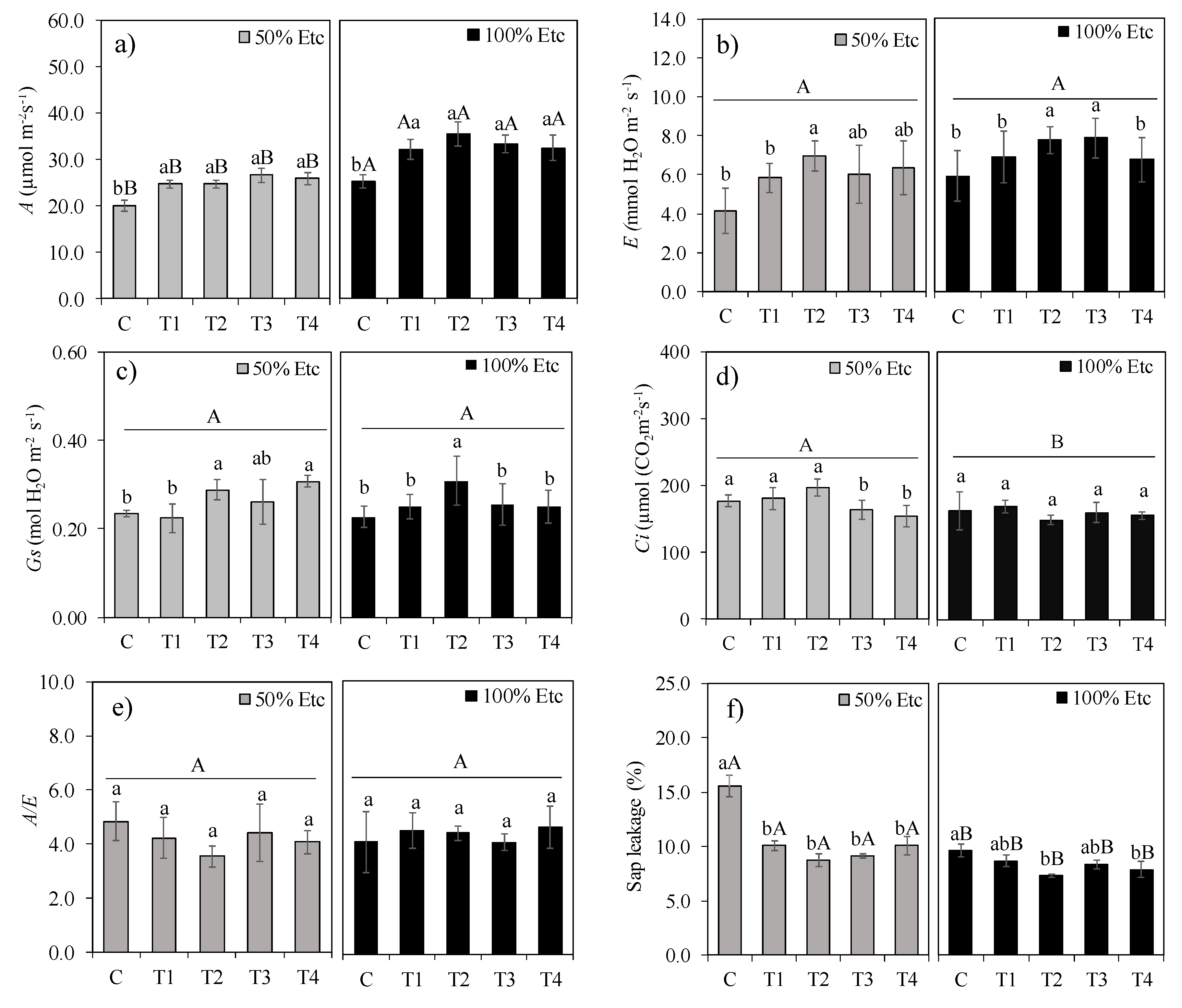
Figure 4.
Content of chlorophyll a (a), chlorophyll b (b), chlorophyll a + chlorophyll b (c), chlorophyll a/b ratio (d), carotenoids (e) and sap leakage (f) of tomato hybrid ‘HM’ under different foliar application treatments and irrigation regimes: control (C), FeSO₄·7H₂O (T1), Fe₂O₃NPs (T2), ZnONPs (T3), and ZnSO₄·7H₂O (T4), subjected to both deficit (50% ETc) and full irrigation (100% ETc). Data represent means ± S.E. Lowercase letters compare treatments (iron and zinc sources), while uppercase letters compare irrigation levels (50% ETc and 100% ETc).
Figure 4.
Content of chlorophyll a (a), chlorophyll b (b), chlorophyll a + chlorophyll b (c), chlorophyll a/b ratio (d), carotenoids (e) and sap leakage (f) of tomato hybrid ‘HM’ under different foliar application treatments and irrigation regimes: control (C), FeSO₄·7H₂O (T1), Fe₂O₃NPs (T2), ZnONPs (T3), and ZnSO₄·7H₂O (T4), subjected to both deficit (50% ETc) and full irrigation (100% ETc). Data represent means ± S.E. Lowercase letters compare treatments (iron and zinc sources), while uppercase letters compare irrigation levels (50% ETc and 100% ETc).
Figure 5.
Effects of foliar treatments and irrigation regimes on tomato ‘HM’: (a) total fruit number per plant, (b) marketable fruit number per plant, (c) fruit transverse diameter, (d) fruit longitudinal diameter, (e) total fruit yield, and (f) marketable fruit yield. Treatments: control (C), FeSO₄·7H₂O (T1), Fe₂O₃ NPs (T2), ZnO NPs (T3), and ZnSO₄·7H₂O (T4), under deficit (50% ETc) or full irrigation (100% ETc). Data represent means ± SE. Lowercase letters denote significant differences among foliar treatments (Tukey’s test, *p* < 0.05); uppercase letters compare irrigation levels.
Figure 5.
Effects of foliar treatments and irrigation regimes on tomato ‘HM’: (a) total fruit number per plant, (b) marketable fruit number per plant, (c) fruit transverse diameter, (d) fruit longitudinal diameter, (e) total fruit yield, and (f) marketable fruit yield. Treatments: control (C), FeSO₄·7H₂O (T1), Fe₂O₃ NPs (T2), ZnO NPs (T3), and ZnSO₄·7H₂O (T4), under deficit (50% ETc) or full irrigation (100% ETc). Data represent means ± SE. Lowercase letters denote significant differences among foliar treatments (Tukey’s test, *p* < 0.05); uppercase letters compare irrigation levels.
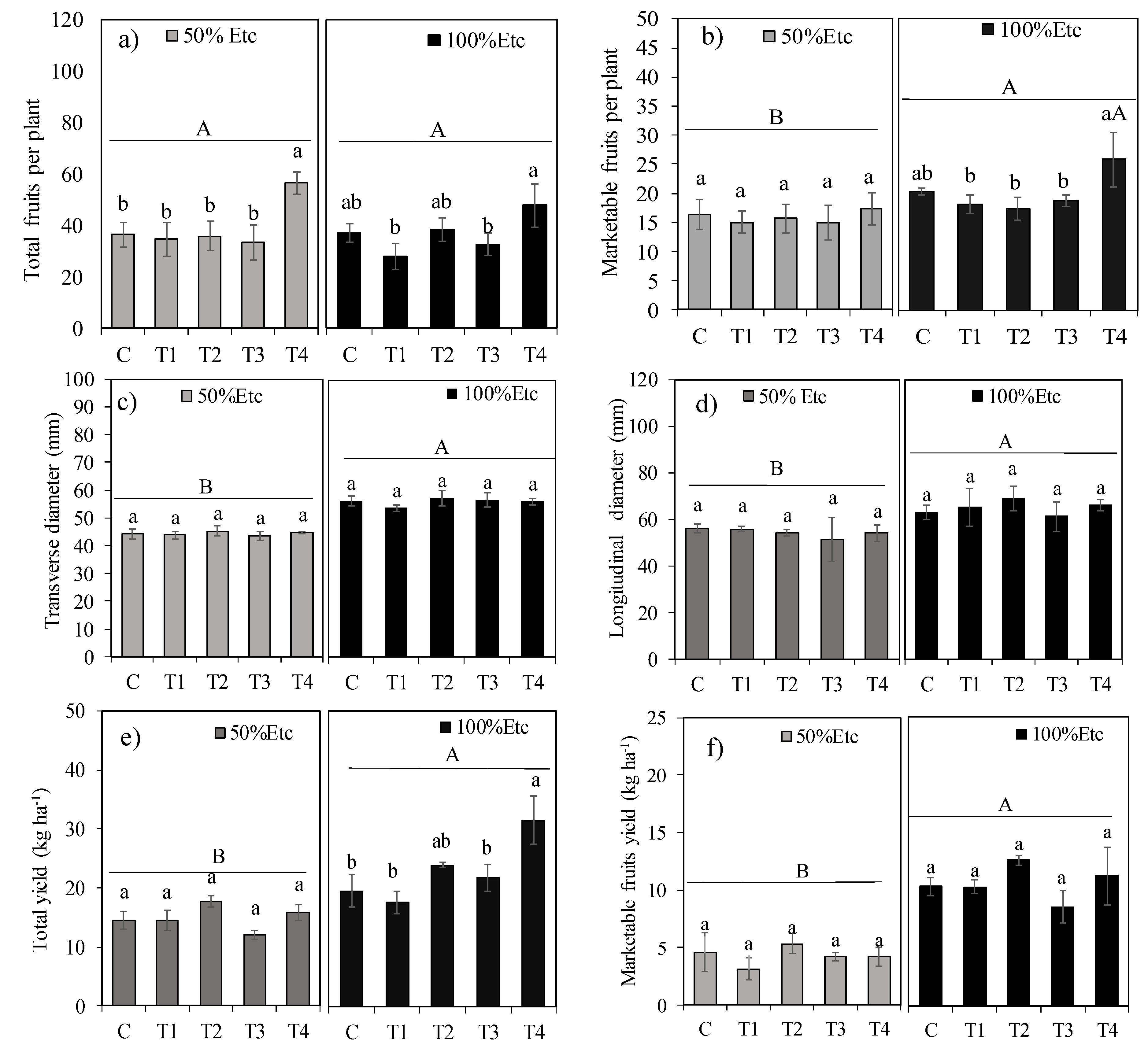
Figure 6.
Boron (a), calcium (b), iron (c), potassium (d), manganese (e), magnesium (f), phosphorus (g) and zinc (h) content in fruits of tomato hybrid ‘HM’ under different foliar application treatments and irrigation regimes: control (C), FeSO₄·7H₂O (T1), Fe₂O₃NPs (T2), ZnONPs (T3), and ZnSO₄·7H₂O (T4), subjected to both deficit (50% ETc) and full irrigation (100% ETc). Data represent means ± S.E. Lowercase letters compare treatments (iron and zinc sources), while uppercase letters compare irrigation levels (50% ETc and 100% ETc).
Figure 6.
Boron (a), calcium (b), iron (c), potassium (d), manganese (e), magnesium (f), phosphorus (g) and zinc (h) content in fruits of tomato hybrid ‘HM’ under different foliar application treatments and irrigation regimes: control (C), FeSO₄·7H₂O (T1), Fe₂O₃NPs (T2), ZnONPs (T3), and ZnSO₄·7H₂O (T4), subjected to both deficit (50% ETc) and full irrigation (100% ETc). Data represent means ± S.E. Lowercase letters compare treatments (iron and zinc sources), while uppercase letters compare irrigation levels (50% ETc and 100% ETc).
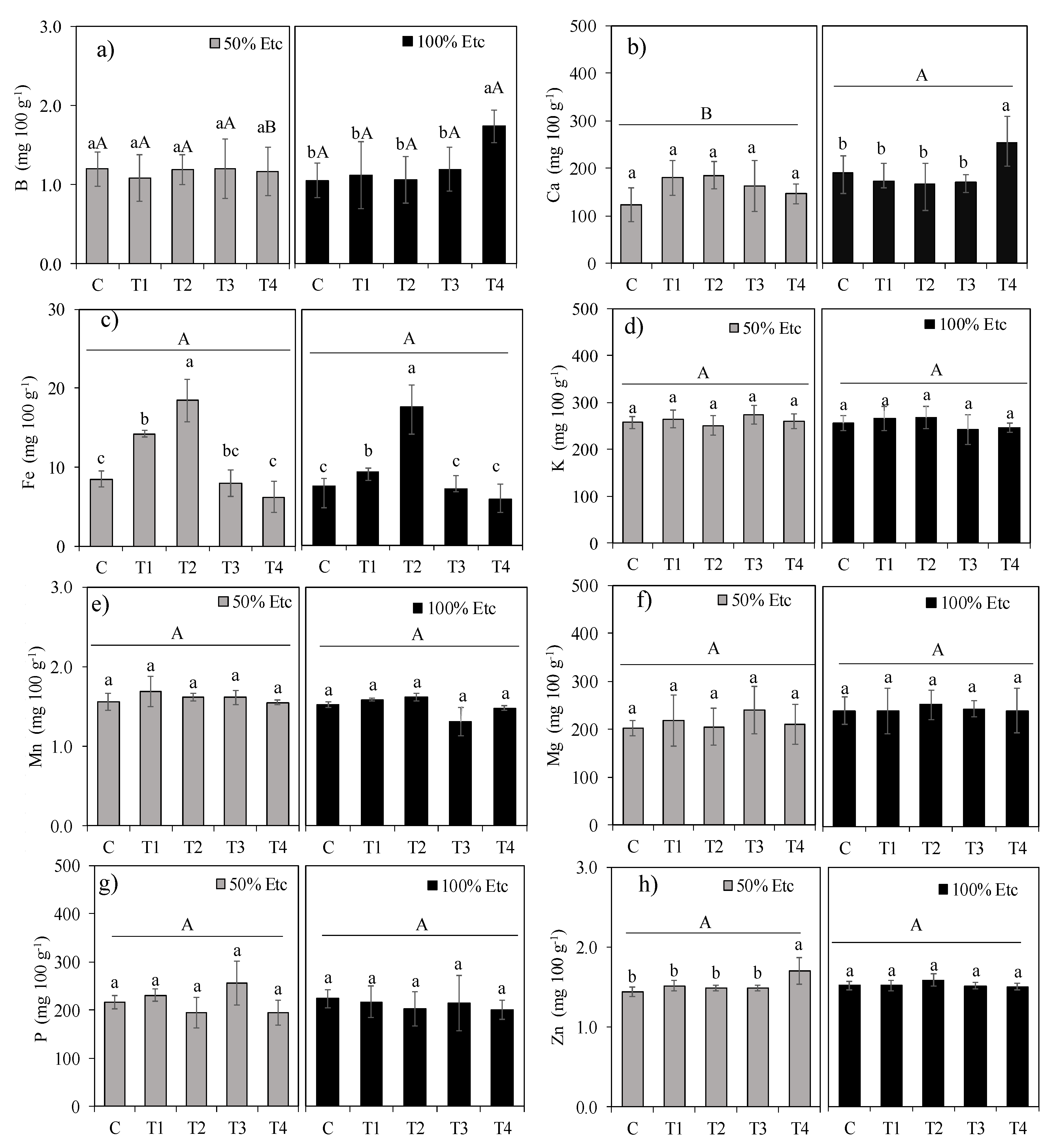
Figure 7.
Relative values (% of control) of marketable yield (YMF), total yield (TYF), photosynthetic pigments [carotenoids (Carot), chlorophyll a (Chl a), b (Chl b), total chlorophyll (Chl t)], physiological parameters [photosynthetic rate (A), transpiration (E), stomatal conductance (gₛ)], and fruit mineral content [B, Ca, Fe, K, Mg, Mn, P, Zn] in tomato 'HM' under foliar treatments: control (C), FeSO₄·7H₂O (T1), Fe₂O₃NPs (T2), ZnONPs (T3), and ZnSO₄·7H₂O (T4),) and irrigation regimes (50% ETc, 100% ETc).
Figure 7.
Relative values (% of control) of marketable yield (YMF), total yield (TYF), photosynthetic pigments [carotenoids (Carot), chlorophyll a (Chl a), b (Chl b), total chlorophyll (Chl t)], physiological parameters [photosynthetic rate (A), transpiration (E), stomatal conductance (gₛ)], and fruit mineral content [B, Ca, Fe, K, Mg, Mn, P, Zn] in tomato 'HM' under foliar treatments: control (C), FeSO₄·7H₂O (T1), Fe₂O₃NPs (T2), ZnONPs (T3), and ZnSO₄·7H₂O (T4),) and irrigation regimes (50% ETc, 100% ETc).
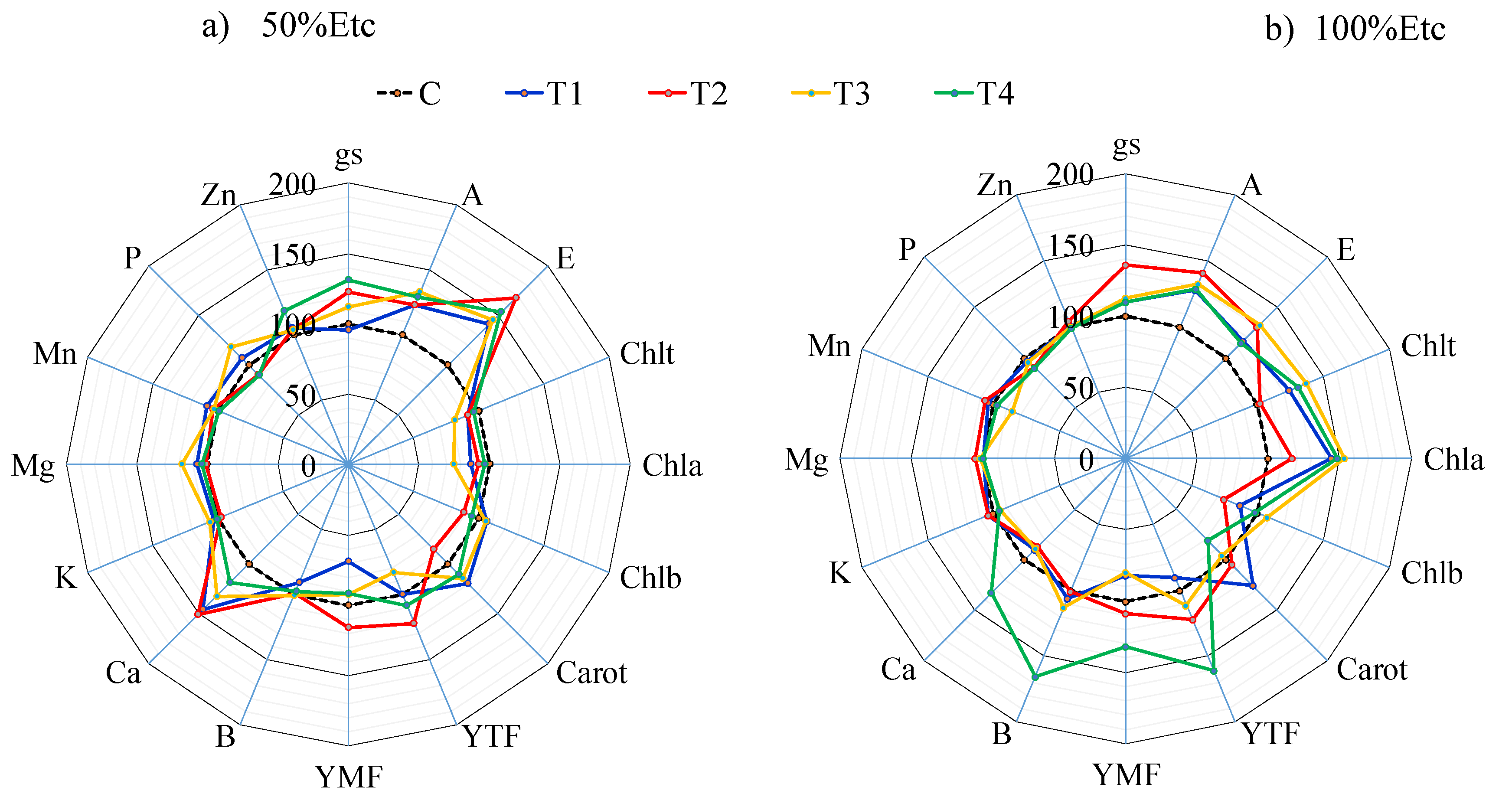
Figure 8.
Pearson’s linear correlation coefficients among the variables leaf area index (LAI), plant height (PH), stem diameter (SD), marketable (AMF) and non-marketable fruit quantity (ANMF), fruit transverse diameter (TD) and longitudinal diameter (LD), marketable (YMF) and non-marketable fruit yield (YNMF), total fruit yield (TYF), internal CO₂ concentration (Ci), photosynthetic rate (A), transpiration rate (E), stomatal conductance (gs), total chlorophyll content (Chlₜ), chlorophyll a (Chl a), chlorophyll b (Chl b), chlorophyll a/b ratio (Chl a/Chl b), carotenoids (car), electrolyte leakage (Ext), water use efficiency (WUE), A/Ci ratio, and fruit mineral content—boron (B), calcium (Ca), iron (Fe), potassium (K), magnesium (Mg), manganese (Mn), phosphorus (P), and zinc (Zn)—in tomato hybrid ‘HM’. Only significant correlations (p < 0.05) are shown.
Figure 8.
Pearson’s linear correlation coefficients among the variables leaf area index (LAI), plant height (PH), stem diameter (SD), marketable (AMF) and non-marketable fruit quantity (ANMF), fruit transverse diameter (TD) and longitudinal diameter (LD), marketable (YMF) and non-marketable fruit yield (YNMF), total fruit yield (TYF), internal CO₂ concentration (Ci), photosynthetic rate (A), transpiration rate (E), stomatal conductance (gs), total chlorophyll content (Chlₜ), chlorophyll a (Chl a), chlorophyll b (Chl b), chlorophyll a/b ratio (Chl a/Chl b), carotenoids (car), electrolyte leakage (Ext), water use efficiency (WUE), A/Ci ratio, and fruit mineral content—boron (B), calcium (Ca), iron (Fe), potassium (K), magnesium (Mg), manganese (Mn), phosphorus (P), and zinc (Zn)—in tomato hybrid ‘HM’. Only significant correlations (p < 0.05) are shown.

Table 1.
Chemical and physical attributes of the soil used in the experiment.
| Chemical | Value | Physical | Value |
|---|---|---|---|
| pH (CaCl2) | 6.45 | Sand (g kg-1) | 389 |
| OM (g kg-1) | 11.2 | Silt (g kg-1) | 430 |
| P (mg dm-3) | 180.1 | Clay (g kg-1) | 181 |
| K+ (mg dm-3) | 256.8 | BD (g cm-3) | 1.30 |
| Ca++ (cmolc dm-3) | 7.72 | PD (g cm-3) | 2.59 |
| Mg++ (cmolc dm-3) | 2.83 | TP (m3 m-3) | 0.47 |
| Na+ (cmolc dm-3) | 0.06 | FC (%) | 12.87 |
| H + Al+++ (cmolc dm-3) | 1.35 | PWP (%) | 5.29 |
| Fe (mg dm-3) | 18.0 | AWC (%) | 7.58 |
| Zn (mg dm-3) | 1.69 | - | - |
P, K+, Na+, Fe and Zn - Mehlich 1 extractant, H+ + Al+3 - 0.5 mol L-1 calcium acetate extractant at pH 7, Ca+2, Mg+2 - 1 mol L-1 KCl extractant, OM - organic matter, V - Base saturation, CEC - cationic exchange capacity, BD - bulk density, PD - soil particle density, TP - total porosity, FC - field capacity, PWP - permanent wilting point, AWC - available water content.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



