Submitted:
30 August 2025
Posted:
02 September 2025
Read the latest preprint version here
Abstract
Citronellol is a naturally occurring acyclic monoterpene alcohol found in essential oils of aromatic plants such as Pelargonium graveolens, Rosa spp., and Cymbopogon nardus. Known for its pleasant rose-like aroma, citronellol has gained attention for its diverse industrial and pharmacological applications, including use in fragrances, cosmetics, insect repellents, and as a bioactive compound with antioxidant, anti-inflammatory, antibacterial, antifungal, antidiabetic, and anticancer effects with beneficial effects in neurological and cardiovascular diseases. Due to limitations in plant extraction and variability in yield, biotechnological approaches have emerged as sustainable alternatives for citronellol production. This review provides a comprehensive overview of citronellol biosynthesis, spanning natural plant pathways and engineered microbial systems. Key enzymes such as geraniol synthase, citral reductases, and short-chain dehydrogenases are discussed in the context of cytosolic and plastidic biosynthetic routes. Advances in metabolic engineering, including enzyme cascade design, have enabled microbial hosts like E. coli, S. cerevisiae, and Y. lipolytica to produce citronellol from renewable feedstocks. Biotransformation strategies using whole-cell systems and purified enzymes have further expanded the repertoire of citronellol derivatives, including rose oxide. Fermentation optimization, scale-up strategies, and innovative bioreactor designs—such as gas-phase and two-phase systems—are highlighted for their role in enhancing monoterpene yields and mitigating volatility-related challenges. Analytical tools including GC-MS, chiral HPLC, and molecular docking are reviewed for compound characterization and bioactivity prediction. Functional studies underscore citronellol’s therapeutic potential, supported by network pharmacology and in vivo models.
Keywords:
biotransformation
; citronellol
; microbial production
; molecular docking
; synthetic biology
; therapeutic applications
1. Introduction
Citronellol, a rose-scented monoterpene alcohol, is a key component in the essential oils of Pelargonium graveolens, Rosa spp., and Cymbopogon nardus. Its biosynthesis involves diverse enzymatic pathways reflecting the evolutionary complexity of plant secondary metabolism. In plants, citronellol is biosynthesized from geranyl diphosphate (GPP) via a multistep pathway involving key enzymes such as geraniol synthase (GES), citral reductases (PhCIRs), and citronellal reductases (CARs). These enzymes belong to the PRISE and short-chain dehydrogenases/reductase (SDR) families and exhibit stereoselectivity that determines the enantiomeric composition of citronellol [1,2,3]. Notable examples include conserved monoterpene routes in Rosa spp., Lagerstroemia caudata, and Paeonia lactiflora, and alternative mechanisms in Pelargonium involving Nudix hydrolases and PRISE enzymes [2]. The orchid Caladenia plicata showcases evolutionary innovation by integrating monoterpene and phenolic biosynthesis to mimic insect pheromones. Citronellol biosynthesis is further shaped by compartmentalization and stereochemistry. Plastidic and cytosolic pathways operate in parallel, with chemotype-specific expression influencing metabolic flux. The enantiomers (R)-(+)- and (S)-(−)-citronellol differ in scent and bioactivity due to stereoselective enzyme interactions [4,5]. These insights into spatial regulation and chiral specificity offer valuable tools for metabolic engineering, fragrance optimization, and therapeutic development.
Building on the natural diversity of citronellol biosynthesis, metabolic engineering and synthetic biology have enabled its sustainable microbial production. By reconstructing plant-derived pathways in hosts like S. cerevisiae, E. coli, and Y. lipolytica, researchers convert simple carbon sources into high-value monoterpenes [6,7,8]. Key strategies include heterologous enzyme expression, modular genetic design, and compartmentalization to enhance flux and minimize bottlenecks [6]. Innovations such as “push-pull-restrain” tactics and peroxisomal targeting have significantly boosted citronellol yields and stereochemical precision, offering scalable alternatives to plant extraction [9]. The optimization of enzyme cascades—from GPP to geraniol and citronellol—marks a critical frontier. Modular pathways, fusion constructs, and cofactor balancing enhance efficiency and enantiomeric control [10]. Beyond biosynthesis, citronellol’s structural properties support its use in analytical platforms, such as mesogenic phases for chiral HPLC separations [11]. These dual roles—as a biosynthetic product and functional material—highlight its interdisciplinary value. Through enzyme engineering and systems-level regulation, citronellol emerges as a model for sustainable innovation in fragrance, therapeutics, and analytical chemistry.
Beyond engineered biosynthesis, biotransformation provides a sustainable route for converting citronellol into high-value fragrance derivatives. Whole-cell systems—including engineered yeasts, fungi, and bacteria—enable regio- and stereoselective modifications without purified enzymes [12]. These platforms efficiently produce rose oxide and other oxygenated terpenoids using surface display, cofactor recycling, and flow biocatalysis [13]. Advanced strategies such as interface bioreactors and modular enzyme cascades enhance stereochemical control, yielding (R)- and (S)-citronellol derivatives with high enantiomeric purity [14]. Enzymes like copper radical oxidases and Old Yellow Enzymes (OYE) exemplify the precision achievable in catalytic tailoring. Selective enzymatic transformation—via alcohol dehydrogenases, ketoreductases, laccases, and peroxidases—further refines citronellol into enantiopure and oxidized forms [15,16,17]. These biocatalysts, often deployed in whole-cell systems, align with green chemistry principles and offer scalable alternatives to chemical synthesis [18]. Rose oxide, a key derivative with four stereoisomers, is produced under mild conditions through microbial and chemoenzymatic cyclization, enhancing essential oil complexity and commercial value [13]. As enzyme engineering and bioprocess design advance, citronellol’s transformation into diverse aroma compounds exemplifies sustainable innovation in fragrance chemistry and natural product valorization.
Realizing citronellol’s industrial potential requires overcoming challenges related to its volatility, hydrophobicity, and cytotoxicity. Conventional aqueous bioreactors often suffer from product loss and microbial inhibition, prompting the development of advanced systems [19]. Gas-phase reactors (e.g., closed-loop designs) enhance VOC recovery and substrate contact, while two-phase partitioning bioreactors (TPPBs) improve mass transfer and reduce toxicity via selective absorption in non-aqueous phases [20]. When coupled with metabolic engineering, these platforms support high-density cultures and sustained monoterpene biosynthesis under controlled conditions [21]. Collectively, they bridge the gap between lab-scale optimization and industrial implementation, positioning fermentation technology as a cornerstone in the commercial production of citronellol.
Citronellol’s structural complexity and biological relevance necessitate precise analytical tools for its characterization and stereochemical resolution. GC–MS remains essential for volatile profiling, while chiral HPLC enables accurate enantioseparation using specialized phases and optical detection. Emerging techniques—such as supercritical fluid chromatography and integrated screening platforms—accelerate method development for fragrance and therapeutic applications [22,23]. These analytical innovations support quality control, pathway elucidation, and bioactivity prediction, forming a critical toolkit for translating molecular insights into functional outcomes. Computational tools have significantly expanded our understanding of citronellol’s bioactivity [23]. Molecular docking and dynamics simulations reveal interactions with neuroprotective, analgesic, and insecticidal targets, while scoring functions and graph-based models clarify its role in terpenoid networks [24,25,26]. Machine learning algorithms enhance bioactivity prediction, essential oil optimization, and compound synergy analysis, bridging molecular structure with application [27]. The integration of docking, ADMET profiling, and AI has transformed citronellol-based drug discovery. Platforms like SwissADME and pkCSM enable rapid screening for antifungal, anti-inflammatory, and antioxidant properties, confirming drug-likeness and safety [28]. AI-driven workflows—combining multi-omics, machine learning, and spectral analysis—accelerate phytochemical discovery and biosynthetic pathway reconstruction [29]. These scalable, predictive approaches position citronellol as a model compound for therapeutic, agricultural, and industrial innovation.
Citronellol’s broad bioactivity and industrial relevance make it a promising candidate for therapeutic innovation and eco-friendly technologies [4]. Beyond its role in fragrance, it exhibits potent antimicrobial, antifungal, and anti-inflammatory effects by targeting bacterial efflux pumps, disrupting fungal membranes, and modulating oxidative stress [4]. Structural modifications, polymer conjugation, and inclusion complexes enhance its utility in drug delivery, coatings, and preservatives [30,31,32]. Citronellol-rich chemotypes and engineered analogs show efficacy against dermatophytosis, multidrug-resistant pathogens, and gastric oxidative injury, positioning it as a versatile scaffold for natural therapeutics and functional materials [5,33].
Its pharmacological spectrum spans anti-inflammatory, neuroprotective, anticancer, analgesic, cardiovascular, and respiratory domains [34,35,36,37,38]. Citronellol modulates inflammatory mediators, stabilizes membranes, and protects neural and gastric tissues [5,39,40]. It induces apoptosis and cell cycle arrest in cancer models [36], attenuates pain via neuroimmune pathways [41], and exerts vasorelaxant and myorelaxant effects [42,43]. Citronellol-derived isothiocyanates also demonstrate anti-barnacle activity, highlighting its potential in marine antifouling [44]. Emerging studies reveal additional therapeutic roles: anti-allergic activity via mast cell inhibition [45], antifibrotic effects through MAPK modulation, and antidiabetic properties that preserve pancreatic and hepatic integrity [46]. As an anticonvulsant and antispasmodic, it regulates neuronal excitability and smooth muscle tone [47,48]. These diverse mechanisms underscore citronellol’s potential in addressing complex pathologies and advancing sustainable biomedicine.
Citronellol’s applications extend beyond pharmacology into eco-conscious consumer products, notably insect repellents and cosmetics. As a major component of citronella oil, it serves as a natural alternative to synthetic repellents like DEET by interfering with mosquito olfaction [49]. To address its volatility and limited duration, advanced formulations—such as β-cyclodextrin encapsulation, structural modification, and sustained-release systems—have improved stability and efficacy [50]. Widely incorporated into lotions, wristbands, and diffusers, citronellol demonstrates low dermal absorption and a favorable safety profile [51]. In cosmetics, it contributes floral aroma and potential soothing effects, though its allergenic potential requires careful formulation [52]. Recent green chemistry innovations have integrated citronellol into natural deep eutectic solvents (NADES), supporting eco-friendly extraction and analytical use [52]. These developments underscore its multifunctionality and expanding role in sustainable personal care, pest control, and wellness technologies.
Despite advances in metabolic engineering, microbial citronellol biosynthesis remains limited by low yields, pathway bottlenecks, and host toxicity [53]. Incomplete catabolic pathway characterization and narrow biocatalyst specificity further hinder scalability [54]. Emerging solutions focus on improving microbial tolerance, broadening substrate range, and enhancing enzyme selectivity—especially in resilient hosts like pseudomonads [55]. AI-driven metabolic design and machine learning-based fermentation control are reshaping biomanufacturing. Predictive algorithms, smart bioreactors, and physics-informed neural networks enable real-time process optimization and precision scaling. As these technologies evolve, they offer new efficiencies and pave the way for sustainable, high-yield citronellol production across industrial sectors.
This review aims to provide a multidisciplinary synthesis of citronellol’s biosynthesis, analytical characterization, biotechnological production, and functional applications. By integrating insights from plant metabolic pathways, microbial engineering, biotransformation strategies, and computational modeling, it seeks to elucidate the enzymatic logic and stereochemical specificity underlying citronellol’s natural and engineered derivation. Furthermore, the paper highlights its therapeutic potential, ecological relevance, and role in sustainable consumer products, offering a framework for future innovation in fragrance chemistry, drug development, and green biomanufacturing. Through this comprehensive analysis, the review positions citronellol as a model compound for advancing natural product research at the interface of biology, chemistry, and technology.
2. Natural Biosynthesis Pathways
Citronellol, a monoterpene alcohol with a rose-like scent, is naturally synthesized in aromatic plants such as P. graveolens, Rosa spp., and Cymbopogon nardus [2,3,56]. Its biosynthesis involves compartmentalized enzymatic steps that influence yield, stereochemistry, and biological activity.
2.1. Citronellol Production in Plants: Key Enzymes and Metabolic Steps
The biosynthesis of citronellol in plants is a multifaceted process that varies across species, reflecting both evolutionary diversity and metabolic specialization. At its core, citronellol production typically begins with the formation of GPP, a universal monoterpene precursor synthesized via either the mevalonate (MVA) or methylerythritol phosphate (MEP) pathway. GPP is then converted to geraniol by GES, followed by sequential reduction to citronellal and ultimately to citronellol. While this general framework applies broadly, individual plant species exhibit distinct enzymatic routes and compartmentalization strategies that shape their citronellol biosynthetic profiles.
In Rosa species, the biosynthesis of β-citronellol is orchestrated through a cytosolic three-step enzymatic cascade (Figure 1). GPP is synthesized by geranyl/farnesyl diphosphate synthase (RhG/FPPS1), and rather than being directly converted by terpene synthases (TPS), geraniol undergoes oxidation to geranial via geraniol dehydrogenase (RhGeDH1). Geranial is then reduced to citronellal by geranial reductase (RhGER2 or RhOPR1), followed by final reduction to β-citronellol through citronellal reductase (RhCAR2). These enzymes belong to the MDR, SDR, and OYE gene families, and their roles have been validated through transient expression in tobacco and rose tissues. This cytosolic pathway appears conserved in other horticultural species such as Lagerstroemia caudata and Paeonia lactiflora, underscoring its evolutionary significance in floral scent biosynthesis [3].
In L. caudata, citronellol biosynthesis involves dual compartmentalization and enzymatic specialization. GPP is synthesized plastidially via the MEP pathway and then processed by two TPSs: LcTPS14, localized in plastids, catalyzes the formation of linalool and citronellal, while LcTPS1, found in the cytosol, contributes to citronellal and citronellol production (Figure 1). Although the specific reductase converting citronellal to citronellol remains unidentified, functional assays confirmed that overexpression of LcTPS1 and LcTPS14 in Arabidopsis thaliana enhances citronellol accumulation, while gene silencing in L. caudata reduces it. This highlights the coordinated regulation and spatial separation of monoterpene biosynthesis in this species [57]. Similarly, P. lactiflora utilizes a plastid-localized monoterpene pathway initiated by the MEP route. The first committed step involves 1-deoxy-D-xylulose 5-phosphate synthase (DXS), leading to GPP formation. TPSs then convert GPP into monoterpene alcohols such as geraniol, linalool, and citronellol. Although the specific TPS responsible for citronellol synthesis was not individually characterized, transcriptomic data revealed strong correlations between TPS gene expression and citronellol emission during peak flowering. These findings suggest that citronellol biosynthesis in P. lactiflora is tightly regulated by developmental cues and plastidial gene expression [58]. In Cymbopogon winterianus (citronella grass), citronellol biosynthesis also follows a plastidial MEP pathway. IPP and DMAPP are condensed to form GPP via geranyl diphosphate synthase (GPPS), which serves as the substrate for monoterpene formation. Transcriptomic analysis identified TPSs and oxidoreductases likely involved in converting GPP to geraniol, then to citronellal and citronellol. Although specific enzymes such as geraniol reductase or citronellal reductase were not pinpointed, the presence of citronellol among major volatiles and the expression of relevant biosynthetic genes support a canonical monoterpene pathway with cytosolic reduction steps. This study lays the groundwork for future functional validation and metabolic engineering in citronella grass [56].
In contrast, Pelargonium species exhibit a distinct biosynthetic strategy that diverges from classical TPS-mediated routes (Figure 1). Citronellol production begins with citral (a mixture of geranial and neral), which is reduced to citronellal by PRISE-family enzymes known as PhCIRs (Pelargonium citral reductases). These enzymes display stereoselectivity, producing either (S)-citronellal or racemic mixtures depending on active site variations. Citronellal is then reduced to citronellol by endogenous reductases. Functional validation through RNAi and in vitro assays confirmed the role of PhCIRs, and phylogenetic analysis revealed multiple PRISE clades with diverse substrate affinities. This independently evolved pathway highlights the metabolic diversity of Pelargonium and its potential for fragrance bioengineering [1]. Further specialization is observed in P. graveolens, where citronellol biosynthesis bypasses conventional plastidial TPS mechanisms entirely (Figure 1). GPP, synthesized via the MEP pathway in plastids, is exported to the cytosol and hydrolyzed by PgNudx1 to form geraniol monophosphate (GP). In citronellol-rich chemotypes, GP is reduced to citronellyl monophosphate (CP), likely by an unidentified reductase, and subsequently dephosphorylated to yield (−)-citronellol. The absence of cyclic monoterpene intermediates and the presence of GDP, GP, and CP in glandular trichomes confirm the cytosolic localization and chemotype-specific regulation of this acyclic biosynthetic route. This discovery expands the known roles of Nudix hydrolases and introduces an alternative metabolic strategy for essential oil biosynthesis in Pelargonium [2,59]. Adding to this enzymatic repertoire, researchers have identified three isoforms of 12-oxophytodienoic acid reductase (PgOPR1–3) in P. graveolens, which catalyze the stereoselective reduction of citral to (S)-citronellal (Figure 1). Although these enzymes do not directly convert geraniol to citronellol, they efficiently process citral—an α,β-unsaturated aldehyde—into a key intermediate that is subsequently reduced to (−)-citronellol by endogenous reductases. PgOPRs exhibit broad substrate specificity and rely on the α,β-unsaturated carbonyl structure for catalytic activity, expanding the enzymatic toolkit available for metabolic engineering of fragrance compounds in Pelargonium [60].
In a striking example of evolutionary innovation, the sexually deceptive orchid Caladenia plicata synthesizes (S)-β-citronellol through a multistep pathway that mimics the sex pheromones of its pollinator. The process begins with plastidial GPP synthesis via the MEP pathway, followed by conversion to geraniol by geraniol synthase (GER1). Geraniol is then stereoselectively reduced to (S)-β-citronellol by alcohol dehydrogenase 3 (ADH3) (Figure 1). This monoterpene pathway operates in tandem with a phenolic biosynthetic route producing 2-hydroxy-6-methylacetophenone, both localized in glandular sepal tips. Transcriptomic analysis confirmed tissue-specific expression of key enzymes, and behavioral assays demonstrated that a 1:4 blend of citronellol and the phenolic compound effectively triggers copulation attempts in male Zeleboria wasps. This study reveals how Caladenia orchids have evolved a sophisticated chemical mimicry system by integrating distinct metabolic pathways to produce biologically active scent compounds [61]. Collectively, these diverse biosynthetic strategies underscore the evolutionary plasticity and biochemical ingenuity of citronellol production in plants. From plastidial MEP-driven routes to cytosolic reductive cascades and pheromone-mimicking systems, each species offers unique insights into the enzymatic logic and ecological roles of monoterpene biosynthesis. Understanding these pathways not only enriches our knowledge of plant secondary metabolism but also opens new avenues for metabolic engineering and sustainable fragrance innovation.
2.2. Comparative Biosynthetic Routes: Cytosolic vs Plastidic Mechanisms
Plants possess two distinct isoprenoid biosynthetic pathways that contribute to the production of terpenoids: the plastid-localized MEP pathway and the cytosolic MVA pathway [62]. Traditionally, the MEP pathway has been associated with monoterpene biosynthesis, while the MVA pathway is linked to the formation of sesquiterpenes and sterols [62]. However, recent studies have revealed that monoterpene production can also occur in the cytosol via non-canonical routes, expanding our understanding of compartmentalized terpene metabolism.
In Pelargonium species, citronellol biosynthesis exemplifies this duality, occurring through both plastidic and cytosolic pathways that are chemotype-specific and enzymatically distinct. The plastidic route begins with the MEP pathway, which generates the isoprenoid precursors isopentenyl diphosphate (IPP) and dimethylallyl diphosphate (DMAPP). These are condensed by GPPS to form GPP, the universal monoterpene precursor. Within plastids, GPP is converted to geraniol by GES, a member of the TPS-g subfamily, and subsequently reduced to citronellol through a series of enzymatic steps including alcohol dehydrogenases (ADH) and citral reductases (PhCIR). This pathway is predominant in rose-scented cultivars and correlates strongly with the expression of plastidial TPS genes across various Pelargonium × hybridum lines [63]. In contrast, the cytosolic pathway represents a non-canonical mechanism that bypasses traditional terpene synthases. Here, GPP—typically synthesized in plastids—is hydrolyzed in the cytosol by a Nudix hydrolase enzyme (PgNudx1), yielding GP. GP is then reduced to CP, likely via an unidentified cytosolic reductase, and finally dephosphorylated to produce citronellol. This route operates independently of plastidial machinery and is localized to glandular trichomes, the primary sites of essential oil storage. Metabolite profiling has shown that citronellol-rich chemotypes accumulate GPP, GP, and CP in nearly equimolar concentrations, while citronellyl diphosphate is notably absent—confirming the cytosolic origin of citronellol in these lines. Moreover, this pathway is associated with chemotypes that favor acyclic monoterpenes over cyclic p-menthane derivatives, indicating a distinct metabolic preference [2].
Together, these findings reveal a remarkable metabolic divergence within Pelargonium, where citronellol biosynthesis can proceed via either plastidic TPS-mediated conversion or a cytosolic Nudix-dependent route. This duality reflects evolutionary adaptation and chemotypic specialization, offering valuable insights for fragrance breeding and metabolic engineering. In contrast, species such as Cymbopogon rely predominantly on plastidic pathways for monoterpene biosynthesis. Interestingly, cross-compartmental transport of intermediates like IPP and GPP has been observed in several plant systems, suggesting dynamic metabolic crosstalk between cytosolic and plastidic routes [64]. Such integration allows plants to fine-tune monoterpene production in response to developmental stages and environmental stimuli, underscoring the complexity and flexibility of plant secondary metabolism.
2.3. Role of Stereoisomerism in Biological Activity
Citronellol, a naturally occurring acyclic monoterpenoid, exists as two enantiomers: (R)-(+)-citronellol and (S)-(−)-citronellol. Although these stereoisomers share the same molecular formula and connectivity, their three-dimensional arrangements differ, resulting in distinct biological and sensory properties. This phenomenon—known as stereoselectivity—reflects the inherent preference of biological systems, such as enzymes, receptors, and transport proteins, for one enantiomer over another [65]. In biological environments, stereoselectivity plays a critical role in determining molecular behavior. Because most biomolecules are chiral, they interact asymmetrically with stereoisomers. For example, one enantiomer may fit precisely into a receptor’s active site and elicit a biological response, while its mirror image may fail to bind or act as an antagonist. This principle is especially relevant in pharmacology and toxicology, where the therapeutic efficacy and safety of a compound often depend on its stereochemical compatibility with target biomolecules [66].
In the case of citronellol, stereoisomerism significantly influences both olfactory perception and bioactivity [67]. Studies have demonstrated that (R)-(+)-citronellol and (S)-(−)-citronellol differ in antifungal potency, scent profile, and interactions with other compounds [68]. These differences arise from stereoselective binding to biological targets, such as fungal membranes and olfactory receptors. A comparative evaluation of (R)-(+)-β-citronellol and (S)-(−)-β-citronellol against Candida albicans and C. tropicalis revealed notable differences in antifungal activity. Both enantiomers exhibited membrane-disruptive effects without compromising the fungal cell wall. For C. albicans, the MIC₅₀ and MFC₅₀ values were 64 µg/mL and 256 µg/mL, respectively. In contrast, C. tropicalis required higher concentrations, with MIC₅₀ at 256 µg/mL and MFC₅₀ at 1024 µg/mL. When combined with Amphotericin B, (R)-(+)-citronellol showed synergism and indifference, while (S)-(−)-citronellol exhibited synergism, additivity, or indifference depending on the strain and dosage. These findings highlight how enantiomeric configuration can modulate both potency and pharmacodynamic interactions [68]. At the enzymatic level, stereoselectivity is governed by structural features of biocatalysts. Enzymes such as PRISE homologs and carbonyl reductases (CARs) influence the enantioselective reduction of citral and citronellal, with single amino acid substitutions capable of altering stereochemical outcomes [1]. This level of control is essential for tailoring citronellol’s functional properties in fragrance design, therapeutic formulation, and ecological signaling. In summary, the stereochemistry of citronellol is not merely a structural detail—it is a defining factor in its biological performance. Understanding and harnessing stereoselectivity enables precise modulation of citronellol’s activity, offering strategic advantages in fields ranging from drug development to aroma biotechnology.
3. Metabolic Engineering and Synthetic Biology
Metabolic engineering and synthetic biology have enabled the microbial production of citronellol through the rational design of biosynthetic pathways, and enzyme optimization. This section outlines key strategies and examples of engineered organisms used to construct efficient microbial cell factories for citronellol biosynthesis.
3.1. Construction of Biosynthetic Pathways in Microbes
Microbial biosynthesis of citronellol offers a sustainable and scalable alternative to plant extraction, relying on engineered metabolic pathways that convert simple carbon sources into high-value monoterpenes (Figure 2) [69]. The process typically begins with the generation of GPP, a key precursor synthesized via either the MVA or MEP pathway. GPP is then converted to geraniol by GES, followed by stereoselective reduction to citronellol through enzymes such as OYEs or CARs. To construct efficient biosynthetic routes, researchers have employed a suite of synthetic biology tools and metabolic engineering strategies. These include heterologous expression of plant-derived enzymes (e.g., GES, OYE) in microbial hosts such as E. coli, S. cerevisiae, and Y. lipolytica, modular pathway design using Golden Gate assembly and CRISPR-based regulation, and precursor flux enhancement through overexpression of MVA pathway genes (e.g., tHMG1, IDI1) [6,7]. Additionally, engineering of GPP synthase variants (e.g., ERG20F96W) and strategic compartmentalization of enzymes within organelles such as mitochondria and peroxisomes have been implemented to improve substrate channeling and reduce metabolic bottlenecks [8,70].
Systematic chassis optimization in Saccharomyces cerevisiae has emerged as a promising strategy for enhancing monoterpene biosynthesis. A recent study addressed key metabolic bottlenecks in citronellol production, including limited precursor availability, cofactor imbalance, and compound cytotoxicity. The researchers employed a multi-pronged engineering approach, incorporating additional copies of key mevalonate pathway genes and targeting biosynthetic enzymes to peroxisomes to improve flux and compartmentalization (Figure 2). Enhancements in NADPH regeneration were achieved through modulation of the nonoxidative branch of the pentose phosphate pathway, while transporter engineering facilitated improved tolerance and export of the target compound. These interventions collectively enabled substantial improvements in production efficiency and demonstrated the scalability of the engineered yeast platform for industrial applications [71].
An integrative “push-pull-restrain” metabolic engineering framework has been employed to enhance citronellol biosynthesis in S. cerevisiae. This approach strategically modulates precursor supply, enzymatic conversion, and pathway competition. To increase flux toward geranyl pyrophosphate (GPP), a mutant variant of farnesyl diphosphate synthase was introduced, effectively redirecting metabolic flow without compromising cellular viability. Enzyme screening efforts identified iridoid synthase from Catharanthus roseus as a highly efficient catalyst for the conversion of geraniol to (S)-citronellol, contributing to improved product specificity. To mitigate losses through competing pathways, deletion of genes associated with intermediate degradation was implemented. Additionally, spatial organization of key biosynthetic enzymes via synthetic scaffolds and fusion constructs facilitated enhanced substrate channeling [9] (Figure 2).
Together, these studies underscore the effectiveness of modular design, enzyme colocalization, and pathway balancing in microbial hosts. By integrating synthetic biology with metabolic engineering, researchers have established robust platforms for the de novo production of citronellol from renewable feedstocks—paving the way for sustainable manufacturing of fragrance, flavor, and pharmaceutical compounds.
3.2. Enzyme Cascade Design: Optimizing Flux Through GPP → Geraniol → Citronellol
The efficient microbial biosynthesis of citronellol hinges on the precise orchestration of enzymatic steps converting GPP to geraniol and subsequently to citronellol. This cascade begins with GES, which catalyzes the conversion of GPP to geraniol. The subsequent reduction of geraniol or its oxidized intermediate, citronellal, to citronellol is mediated by OYEs and SDRs [1,60]. Recent advances have demonstrated that bienzymatic cascades—such as those combining alcohol oxidases (e.g., CgrAlcOx) with OYEs—can achieve high enantiomeric purity of citronellal from geraniol [10]. Moreover, the use of protein scaffolding and fusion constructs has proven effective in enhancing substrate channeling and minimizing intermediate loss [9]. These findings underscore the critical role of enzyme selection and spatial organization in optimizing metabolic flux and product yield.
Citronellol’s structural adaptability has extended its utility beyond biosynthetic applications, particularly in the development of advanced chromatographic materials. One notable innovation involves the design of mesogenic stationary phases for high-performance liquid chromatography (HPLC), wherein citronellol derivatives serve as functional components. A representative example is the LC-LCC8 phase, synthesized by grafting a liquid crystal-based ligand onto a siloxane-functionalized silica matrix [11]. The mesogenic properties of the ligand, validated through thermal and optical analyses, contribute to enhanced molecular ordering and unique chromatographic behavior. Comprehensive physicochemical characterization confirmed the structural integrity and surface morphology of the stationary phase, while performance assessments demonstrated pronounced shape selectivity and dual-mode compatibility. In both normal-phase and reversed-phase configurations, the LC-LCC8 column exhibited high resolution for isomeric and structurally diverse analytes, underscoring its potential for chiral and shape-selective separations. Temperature-dependent studies further revealed tunable selectivity, attributed to the dynamic orientation of the mesogenic ligands [11].
Returning to microbial biosynthesis, a particularly promising strategy involves the enzymatic conversion of geraniol to enantiopure (R)-citronellal—a key intermediate en route to citronellol. This transformation is achieved via a two-step cascade: oxidation of geraniol to geranial by SDRs, followed by selective reduction of geranial to (R)-citronellal by ene reductases (ERs). The high enantioselectivity of this system is especially valuable for fragrance and pharmaceutical applications, where stereochemistry critically influences bioactivity and sensory properties. Implementation in microbial hosts such as E. coli and S. cerevisiae involves synthetic biology techniques to clone and express SDR and ER genes, optimize codon usage, and fine-tune promoter strength for balanced enzyme expression. Redox cofactor management is essential to sustain enzymatic activity and prevent metabolic bottlenecks. This module is typically integrated with upstream pathways converting GPP to geraniol, forming a complete biosynthetic route from central metabolism to citronellol. Enhancing flux through this route often requires overexpression of GES, suppression of competing pathways, and dynamic regulation strategies. Collectively, these modular enzymatic cascades offer a renewable, controllable, and scalable alternative to traditional citronellol production, advancing the industrial viability of high-value terpenoids [72].
4. Biotransformation and Catalysis
Biotransformation offers a sustainable and selective route for modifying monoterpenes such as citronellol into high-value fragrance derivatives. This section explores whole-cell systems, enzyme-specific conversions, and the catalytic transformation of citronellol into rose oxide and related compounds.
4.1. Whole-Cell Conversion Systems: Engineered Yeasts and Fungi
Whole-cell biocatalysis leverages the metabolic machinery of living microorganisms to perform complex chemical transformations without requiring enzyme purification. Engineered yeasts and fungi have emerged as powerful platforms for monoterpene biotransformation, offering high selectivity, operational simplicity, and sustainability. Yeast surface display systems, for instance, enable the co-expression of multiple enzymes on the cell wall, facilitating multistep reactions and enhancing substrate accessibility. Arming technologies have been successfully applied to S. cerevisiae, Y. lipolytica, and Pichia pastoris, allowing for the functional display of catalytic proteins and improving whole-cell performance [12]. Filamentous fungi such as Penicillium and Aspergillus species have demonstrated remarkable tolerance to high terpene concentrations and the ability to perform regio- and stereoselective oxidations of citronellol into rose oxide and other aroma compounds [13,73].
Similarly, Pseudomonas spp. exhibit exceptional resistance to terpene toxicity and catalytic efficiency, converting citronellol into cis- and trans-rose oxides with yields reaching 29.67 mg/L [74]. These microbial systems exemplify the versatility of whole-cell platforms in producing structurally diverse and industrially relevant monoterpenoids. Recent advances in flow biocatalysis have further expanded the utility of whole-cell systems for continuous and scalable chemical production. Whole-cell catalysts offer intrinsic advantages in flow environments, including simplified preparation, enhanced enzyme stability, and built-in cofactor regeneration. Integration into packed-bed, microfluidic, and segmented flow reactors enables precise control over reaction parameters, improved mass transfer, and sustained productivity [75]. Importantly, whole cells can execute complex enzymatic cascades involving cofactor-dependent reactions without external supplementation, owing to their native metabolic networks. Their membrane-bound architecture also permits operation under harsh or non-aqueous conditions, broadening their applicability beyond conventional aqueous systems. The convergence of whole-cell biocatalysis with flow chemistry has enabled the synthesis of fine chemicals, APIs, and monoterpenoids such as citronellol and its derivatives through non-natural enzymatic cascades, including stereoselective reductions, oxidations, and rearrangements [75]. For instance, a metabolically engineered microbial platform was developed for the enantioselective synthesis of (R)-citronellal from renewable substrates. By introducing a cascade of heterologous enzymes and optimizing precursor availability and cofactor balance, the system achieved high yield and regioselectivity under mild conditions, demonstrating its potential for sustainable monoterpenoid production [14].
In a complementary study, E. coli was engineered to co-express geraniol dehydrogenase and an (R)-selective enoate reductase, forming a self-sufficient cascade for the bioconversion of geraniol to (R)-citronellal. Optimization of enzyme expression, cofactor recycling, and substrate uptake enabled high conversion rates and enantioselectivity without external cofactor addition, underscoring the scalability and efficiency of microbial biosynthetic platforms [76]. Further innovations in stereoselective biotransformation were demonstrated using a double coupling system within an interface bioreactor. Racemic citronellol was resolved via transacetylation using alcohol acetyltransferase from Pichia kluyveri, yielding (R)-citronellol with >98% ee and (S)-citronellyl acetate with 70–80% ee. Subsequent oxidation steps using Rhodococcus equi, Geotrichum candidum, and Candida viswanathii produced optically active (R)- and (S)-citronellol derivatives with high stereochemical purity. The interface bioreactor facilitated phase-boundary reactions, enhancing selectivity and product recovery [77].
Filamentous fungi continue to serve as valuable biocatalysts for aroma compound synthesis. Aspergillus and Penicillium species were shown to convert both (R)-(+)- and (S)-(−)-citronellol into rose oxide and other oxygenated terpenoids via regioselective hydroxylation, cyclization, and rearrangement. Solid-phase microextraction (SPME) coupled with GC-MS enabled efficient profiling of volatile biotransformation products directly from the culture headspace, preserving compound integrity and streamlining analysis [78]. The enzymatic versatility of Rhizopus oryzae was highlighted in its transformation of citronellene and citronellol into hydroxylated and rearranged products. Cultivation in potato dextrose broth yielded dihydroxycitronellene and 7-hydroxycitronellol as major metabolites, alongside minor compounds such as rose oxide, rose glycol, and isopulegol. These transformations, confirmed via GC-MS and spectral matching, underscore the potential of fungal biocatalysis for generating structurally diverse terpenoid derivatives under mild conditions [79]. S. cerevisiae has also demonstrated endogenous enzymatic capacity for oxidizing monoterpene alcohols such as geraniol, nerol, citral, and citronellal into their corresponding acids. The transformation of citral into citronellic acid was proposed to proceed via intermediate formation of citronellal, followed by oxidation. Gene deletion studies targeting ADH1, ADH3, ADH4, and SFA1 confirmed their roles in sequential dehydrogenation steps, revealing yeast’s metabolic potential for terpenoid modification [80]. Notably, this study represents the first report of yeast-mediated biosynthesis of geranic acid and citronellic acid from monoterpene alcohols, with implications for aroma compound development in fermented beverages and biotechnological applications in the flavor and fragrance industries [80].
Complementing these whole-cell approaches, a modular bienzymatic cascade was developed for the stereoselective conversion of geraniol into either (R)- or (S)-citronellal. The system employed a copper radical alcohol oxidase (CRO-AlcOx) to oxidize geraniol into geranial, followed by selective reduction using stereocomplementary variants of OYE. By choosing specific OYE isoforms, researchers achieved tunable production of either enantiomer with high optical purity under mild aqueous conditions. The enzymes were applied in sequential or co-expression formats to optimize conversion efficiency. This strategy highlights the utility of combining radical oxidases with ene-reductases for tailored monoterpene transformations and underscores the potential of enzyme-based cascades for sustainable and selective biomanufacturing of citronellol derivatives [10].
4.2. Enzyme-Specific Conversions: Reductases and Alcohol Dehydrogenases
Enzyme-specific biocatalysis provides precise control over reaction pathways, enabling highly selective transformations of monoterpenes into valuable derivatives. Among the most prominent catalysts in this domain are ADHs and ketoreductases (KREDs), which mediate stereoselective oxidation and reduction of alcohols, aldehydes, and ketones. These NAD(P)H-dependent enzymes are foundational tools in asymmetric synthesis and chiral resolution, offering high efficiency and selectivity in both laboratory and industrial settings [15,16]. Recent advances in protein engineering have expanded the substrate scope and enantioselectivity of ADHs, allowing for tailored production of (R)- or (S)-citronellol and structurally related monoterpenoids. These engineered enzymes facilitate the construction of stereochemically pure compounds under mild conditions, making them attractive for applications in pharmaceuticals, flavors, and fragrances [17].
The ability to fine-tune enzyme activity through rational design or directed evolution has significantly enhanced the versatility of ADHs in monoterpene biotransformation. Beyond reductases, oxidative enzymes such as laccases and peroxidases have gained attention for their role in the chemoenzymatic conversion of citronellol to rose oxide. These enzymes catalyze oxidative cyclization reactions in the presence of suitable mediators, offering a sustainable alternative to traditional photooxidation methods [13]. Their application in both purified form and as part of whole-cell systems broadens their utility, enabling integration into microbial hosts for streamlined biocatalytic processes. Together, these enzyme classes—reductases, dehydrogenases, laccases, and peroxidases—form a complementary toolkit for the selective modification of monoterpenes. Their integration into biotechnological workflows supports the development of efficient, scalable, and environmentally friendly routes for producing high-value terpenoid compounds.
4.3. Conversion of Citronellol to Rose Oxide and Other Fragrance Derivatives
Citronellol serves as a versatile precursor for a range of high-value fragrance molecules, including rose oxide, citronellyl esters, and glycosides [81]. Among these, rose oxide stands out as a key odor-impact compound found in rose and geranium oils. It exists as four stereoisomers, each contributing distinct olfactory notes, making its selective synthesis a focal point in fragrance chemistry [13]. The transformation of citronellol-rich essential oils into rose oxide exemplifies the convergence of natural biosynthetic pathways and targeted enzymatic or chemical engineering. In planta, citronellol is synthesized via the MVA pathway as an intermediate in monoterpenoid biosynthesis. However, the conversion to rose oxide—a compound celebrated for its potent floral aroma—requires post-biosynthetic modification, often achieved through microbial or chemical catalysis. Biotransformation using fungi and bacteria has demonstrated high selectivity in producing cis- and trans-rose oxide isomers, offering a biologically inspired route to fragrance enhancement. A notable advancement in this domain is the work by Dwivedi et al., who introduced a novel oxidative strategy for upgrading citronella essential oil. Their approach selectively converts β-citronellol into rose oxide using iodosylbenzene (PhIO) as the oxidant. This transformation proceeds via intramolecular cyclization, yielding rose oxide with enhanced selectivity and efficiency under mild conditions. The in situ enrichment of essential oil not only improves its olfactory profile but also significantly boosts its commercial value. Importantly, this single-step oxidation process minimizes byproducts and avoids harsh reagents, presenting a sustainable alternative to conventional chemical synthesis [81].
Building on this, enzymatic strategies have emerged as powerful tools for selective oxidation of citronellol. By employing monooxygenases or engineered biocatalysts, researchers have replicated nature’s precision under scalable and environmentally friendly conditions. These biocatalytic systems enable the production of enantiopure cis- and trans-rose oxides, which contribute substantially to the sensory complexity and market appeal of essential oils. The integration of enzymatic transformation into essential oil processing aligns with the principles of green chemistry—reducing waste, eliminating toxic reagents, and enabling selective synthesis. This approach not only enhances fragrance profiles but also exemplifies the potential of post-biosynthetic tailoring in natural product valorization. As such, biocatalytic routes offer eco-conscious alternatives to traditional synthesis, paving the way for sustainable innovation in perfumery and cosmetic formulation [13].
5. Fermentation and Bioprocess Development
The efficient microbial production of citronellol, a highly volatile and hydrophobic monoterpene, requires not only advanced metabolic engineering but also carefully optimized fermentation strategies and bioreactor configurations. Traditional aqueous-phase bioreactors often face significant limitations, including product evaporation, poor substrate availability, and microbial inhibition at high product concentrations [19]. To overcome these challenges, recent studies have highlighted innovative reactor designs and process intensification strategies that enhance both yield and scalability. This section outlines key approaches for enhancing fermentation performance, scaling up production, and mitigating volatility-related challenges.
5.1. Gas-Phase Bioreactors
Gas-phase bioreactor systems have emerged as effective platforms for handling volatile monoterpenes. In contrast to conventional setups, these reactors operate by exposing microbial biofilms or immobilized cells to nutrient mist or vapor-phase substrates, thereby minimizing product loss and improving oxygen transfer. A notable example is the study by Wüst et al., who applied a continuous closed-gas-loop bioreactor (CCGLB) for the S. cerevisiae (baker’s yeast)-mediated biotransformation of geraniol into citronellol. This configuration enabled efficient conversion of volatile monoterpenes in the gas phase, reduced substrate evaporation, and enhanced contact between gaseous substrates and microbial catalysts [82]. The CCGLB system demonstrated superior process stability, particularly for high-density cultures, and offered an efficient strategy for recovering volatile organic compounds (VOCs). Such designs underscore the potential of gas-phase fermentation for overcoming the inherent challenges of citronellol biosynthesis.
5.2. Two-Phase Partitioning Bioreactors (TPPBs)
Another promising strategy involves the integration of biphasic aqueous-organic systems, which provide both toxicity mitigation and improved mass transfer. These reactors incorporate a non-aqueous phase—such as silicone oil, hexadecane, or polymer beads—that selectively absorbs hydrophobic monoterpenes. By maintaining low aqueous concentrations of citronellol, these systems protect microbial cells from product inhibition while ensuring sustained activity. Doig et al. demonstrated the effectiveness of this approach in the stereoselective reduction of geraniol to citronellol by S. cerevisiae as the biocatalyst. Their two-phase partitioning bioreactor allowed for precise kinetic analysis under pseudo-steady-state conditions, leveraging the hydrophobicity of both substrate and product to achieve improved reaction control [20]. This setup not only enhanced conversion efficiency but also provided a framework for industrial-scale application, particularly in processes involving transient VOC loadings and oxygen-limited environments.
5.3. Integration and Industrial Relevance
Together, gas-phase bioreactors and TPPBs represent two complementary strategies for addressing the core challenges in citronellol fermentation—volatility, hydrophobicity, and microbial inhibition [21]. While gas-phase systems excel in substrate utilization and VOC recovery, biphasic approaches provide enhanced process control and product stability [83]. The continued integration of these bioprocess designs with metabolic engineering advances offers a promising route toward scalable, high-yield production of citronellol and related monoterpenes.
6. Analytical Tools and Characterization
The characterization of citronellol and its derivatives relies on a variety of analytical platforms, each offering unique insights into chemical composition, biosynthetic pathways, stereochemistry, assessing purity, and bioactivity. Together, these methods provide a complementary toolkit for profiling, isolating, and predicting the biological relevance of this versatile monoterpene. This section outlines key analytical techniques used for profiling, quantification, enantiomeric resolution, and computational bioactivity prediction.
6.1. GC-MS for Profiling and Quantification
Gas chromatography–mass spectrometry (GC–MS) remains the gold standard for profiling VOCs, including monoterpenes like citronellol. This technique separates analytes based on volatility and polarity, followed by mass-based identification. In qualitative analysis, GC-MS identifies compounds by retention time and fragmentation patterns, matched against spectral libraries such as NIST [84]. In quantitative analysis, peak area and height are proportional to analyte concentration. Internal standards and calibration curves improve accuracy [85]. For example, GC-MS has been used to profile citronellol and related metabolites in Trichoderma harzianum, revealing bioactive compounds such as 6-pentyl-2H-pyran-2-one and fatty acids with antifungal properties [86]. Advanced platforms like GCxGC-MS and GC-TOF-MS offer enhanced resolution and sensitivity, supporting untargeted metabolomics and complex mixture analysis [87].
6.2. Chiral HPLC for Enantiomer Separation
While GC–MS provides compositional profiles, stereochemical characterization requires chiral resolution methods. Direct separation achieved using chiral stationary phases (CSPs) such as polysaccharide derivatives or cyclodextrins. CSPs interact differently with each enantiomer, enabling stereoselective separation [88,89]. However, indirect separation involves derivatization into diastereomers using chiral reagents, followed by separation on achiral columns [90]. SUMICHIRAL OA columns and CHIRALPAK series have been used to separate citronellol enantiomers for fragrance and therapeutic studies [91].
Chiral HPLC plays a central role in separating citronellol enantiomers, which differ in olfactory and pharmacological properties [92]. The enantioseparation of citronellol and other chiral monoterpenes was investigated using reverse-phase HPLC on three distinct chiral stationary phases (CSPs): amylose tris(3,5-dimethylphenylcarbamate) (AD-H), cellulose tris(3,5-dimethylphenylcarbamate) (OD-H), and β-cyclodextrin (β-CD) phenylcarbamate (CD-Ph), with on-line optical rotatory dispersion (ORD) detection enabling real-time chiroptical analysis. Among the CSPs, the AD-H column exhibited superior enantioselectivity, successfully resolving seven out of nine tested monoterpenes, including citronellol, while the CD-Ph and OD-H columns showed limited and moderate performance, respectively. The ORD detector allowed direct determination of optical activity and absolute configuration without derivatization, with the (3S)-(+)-linalool configuration confirmed via SPME-GC-MS. The study highlighted the influence of functional groups such as hydroxyl and alkene moieties on chiral recognition, and demonstrated that preparative HPLC using AD-H could achieve high-throughput isolation with significantly greater sample capacity than GC methods. This approach provides a powerful analytical tool for the stereochemical characterization and quality control of citronellol in essential oils and related applications [93]. Recent innovations, including automated screening systems that integrate supercritical fluid chromatography (SFC) and HPLC, further accelerate method development and optimization for chiral monoterpenes like citronellol [22,23].
6.3. Molecular Docking and Computational Tools for Bioactivity Predictions
Molecular docking and computational modeling are powerful tools for predicting the bioactivity of citronellol and its derivatives [94]. Molecular docking simulates interactions between citronellol and target proteins [95]. Docking scores and binding poses help assess affinity and specificity.
6.3.1. Computational Docking and Simulation Studies of Citronellal and Citronellol: From Neuroprotection to Vector Control
Computational approaches such as molecular docking and molecular dynamics (MD) simulations have become valuable tools for elucidating the bioactivity of citronellal and citronellol, offering mechanistic perspectives that complement in vivo and in vitro findings. Citronellal, in particular, has demonstrated promising neuroprotective properties across both computational and experimental models. Docking analyses revealed strong affinity for nitric oxide synthase (NOS), suggesting a potential role in modulating oxidative stress pathways. Weaker interactions were observed with key inflammatory mediators, including interleukins (IL-6, IL-12) and tumor necrosis factor-alpha (TNF-α), indicating a selective binding profile [24]. MD simulations further validated the stability of the citronellal–NOS complex, with favorable binding free energy values supporting sustained interaction under physiological conditions. These computational predictions were substantiated in a rat model of cerebral ischemia, where citronellal administration led to reduced infarct volume, enhanced antioxidant defense (elevated glutathione levels), and decreased levels of oxidative and inflammatory biomarkers such as malondialdehyde (MDA), nitric oxide (NO), myeloperoxidase (MPO), and acetylcholinesterase (AChE). Histopathological assessments confirmed attenuation of neuronal damage, reinforcing the compound’s cerebroprotective potential [24].
Beyond its neuroprotective potential, citronellal has been implicated in mosquito repellency through disruption of olfactory signaling. In Vitex negundo L. essential oil, citronellal was identified as a principal active component. Docking simulations against odorant-binding proteins (OBPs) of Anopheles gambiae revealed strong affinities, particularly with OBP1 and OBP7, suggesting interference with host-seeking behavior. Structural validation through RMSD analysis confirmed the reliability of these models. These computational predictions were supported by chemical profiling and behavioral assays, which demonstrated rapid and significant repellency, underscoring the utility of docking tools in correlating molecular interactions with biological efficacy [26]. In addition to repellency, computational modeling has guided formulation strategies to enhance citronellal’s pharmacological performance. Encapsulation within β-cyclodextrin (β-CD) was shown to improve physicochemical stability and prolong analgesic activity. Docking studies predicted strong interactions with glutamatergic system targets, aligning with in vivo observations of reduced hyperalgesia and decreased Fos protein expression in pain-associated brain regions. These findings illustrate how docking-guided delivery systems can optimize the therapeutic potential of volatile natural products [25].
Citronellol, structurally analogous to citronellal, has also been investigated for its insecticidal properties. In Cymbopogon citratus (lemongrass) oil, citronellol was identified as a major constituent and exhibited potent larvicidal activity against Anopheles sinensis. Biochemical assays confirmed inhibition of acetylcholinesterase (AChE), while docking simulations revealed favorable binding to the enzyme’s active site via hydrogen bonding and Pi-Sigma interactions with key residues. These results support citronellol’s neuroinhibitory mechanism and its contribution to the bioactivity of lemongrass oil [96]. Overall, these studies highlight how computational docking and MD simulations serve as powerful tools for predicting and rationalizing the diverse bioactivities of citronellal and citronellol. From neuroprotection and analgesia to mosquito repellency and larvicidal activity, such integrative approaches bridge chemical interactions with biological outcomes, strengthening the foundation for their development in therapeutic and vector-control applications.
6.3.2. Scoring Functions and Theoretical Frameworks for Predicting Citronellal-Derived Reactions and Terpenoid Biosynthesis
Scoring functions—including force-field-based, empirical, and knowledge-based models—serve as essential tools for evaluating binding affinities, transition state stability, and enzymatic selectivity. These computational approaches have been applied to elucidate citronellal-derived reactions, providing mechanistic insights into biosynthetic pathways. For instance, a theoretical study investigated the regioselectivity of the Pictet–Spengler reaction between dopamine and aldehydes such as (S)-citronellal, catalyzed by (S)-norcoclaurine synthase (NCS). Using density functional theory (DFT) in combination with scoring function analysis, the reaction mechanism was modeled through transition-state structures yielding para-cyclized (salsolinol) and ortho-cyclized (isosalsolinol) products. Results revealed that para-cyclization is energetically favored in acidic media, whereas ortho-cyclization proceeds preferentially under neutral conditions through a zwitterionic phenolate–iminium intermediate. Scoring analysis of transition states further confirmed the stereoelectronic influence of NCS in directing product selectivity. Collectively, these findings highlight the predictive capacity of computational scoring tools in rationalizing enzyme-mediated citronellal transformations and alkaloid biosynthesis [97].
Complementing these mechanistic insights, more recent work has extended the application of theoretical frameworks toward large-scale terpenoid biosynthesis. A graph-theory-based approach, termed TeroGen, was developed to generate a virtual library of terpenoids and to map plausible biosynthetic mechanisms. This framework integrates physically based simulations with deep learning, mimicking the two fundamental stages of terpene biosynthesis—cyclization and decoration. Using this method, more than 30,000 sesterterpenoids were predicted and organized into a reaction network, substantially expanding the accessible chemical space. Thermodynamic and kinetic scoring functions were employed to assess both the feasibility of synthetic routes and the biological relevance of candidate structures. By systematically enumerating terpene scaffolds and their biosynthetic trajectories, TeroGen provides a versatile platform for exploring enzyme-guided transformations, designing novel terpenoid structures, and informing heterologous biosynthesis strategies. Importantly, such computational frameworks hold promise for guiding synthetic biology and drug discovery efforts involving citronellol-related terpenoids [98].
6.3.3. Machine Learning Approaches for Predicting Bioactivity and Formulation Outcomes of Citronellol-Rich Essential Oils
Machine learning (ML) has emerged as a powerful tool for predicting the biological activity of natural compounds, including monoterpenes such as citronellol, and for guiding the design of optimized formulations. Algorithms such as support vector machines and logistic regression have been successfully applied to predict antiviral and antimicrobial properties, while Quantitative Activity–Composition Relationship (QCAR) models provide a framework for linking essential oil composition to functional outcomes. A comprehensive study employing these approaches evaluated the predictive potential of ML for essential oils and natural extracts, including citronellol-rich profiles. By integrating supervised learning with QCAR modeling, researchers were able to identify constituents most strongly associated with anti-biofilm activity and predict the efficacy of different oil combinations. Importantly, unsupervised clustering techniques further allowed the grouping of bacterial strains such as Staphylococcus aureus and Pseudomonas aeruginosa according to their susceptibility profiles, enabling more targeted antimicrobial testing. These findings underscore the utility of ML tools for deconvoluting complex chemical mixtures, informing artificial blend design, and facilitating the discovery of bioactive compounds like citronellol for pharmaceutical and industrial applications [27].
Expanding on this integrative framework, another study applied ML to explore the relationship between the chemical composition of 61 essential oils and their antibiofilm activities against S. aureus and S. epidermidis. Essential oil profiles were characterized by GC–MS, and supervised models—including decision trees and support vector machines—were trained to correlate chemical composition with antibiofilm efficacy. Complementary unsupervised clustering revealed distinct patterns of bacterial response, allowing oils to be classified by their bioactivity. Notably, oils enriched in citronellol, thymol, and eugenol demonstrated pronounced antibiofilm activity. Moreover, predictive modeling identified synergistic combinations and ranked oils by their predicted potency, providing a rational basis for formulation strategies. Taken together, these studies illustrate how ML approaches can accelerate natural product research by predicting bioactivity, uncovering synergistic interactions, and enabling the targeted development of non-biocidal antibiofilm agents. This highlights the broader potential of ML-driven methodologies in advancing citronellol-based therapeutic and formulation science [99].
6.3.4. Integrating Docking with ADME/Tox Profiling to Identify Citronellol-Related Compounds as Therapeutic Candidates
The integration of molecular docking with ADME/toxicity (ADMET) profiling has become a central strategy in evaluating the therapeutic potential of citronellol-related compounds. Docking studies have revealed that derivatives of citronellol and structurally related monoterpenes possess promising antifungal and anti-inflammatory activities, while computational ADMET platforms such as SwissADME and pkCSM provide valuable insights into their pharmacokinetic and safety profiles. One study investigated the antifungal potential of Cymbopogon khasianus (palmarosa) and C. citratus (lemongrass) essential oils, where GC–MS analysis identified geraniol and geranial—monoterpenes structurally analogous to citronellol—as dominant constituents. Docking simulations using PatchDock targeted fungal enzymes involved in riboflavin biosynthesis, including riboflavin synthase (RS), RibD domain-containing protein, and 3,4-dihydroxy-2-butanone 4-phosphate synthase (DBPS). Both compounds exhibited strong binding affinities through hydrogen bonding and hydrophobic interactions, and in vitro assays confirmed antifungal activity against Aspergillus fumigatus, Aspergillus flavus, and Mucor spp. ADMET analysis supported their drug-likeness, compliance with Lipinski’s rules, and low toxicity, highlighting their therapeutic promise[28].
A complementary study focused on C. citratus oil against A. fumigatus, combining in vitro assays with docking simulations of its principal component, citral. Docking targeted enzymes essential for fungal cell wall biosynthesis—UDP-glycosyltransferase, glucosamine-6-phosphate synthase, and chitin synthase—where citral demonstrated favorable binding and stable hydrogen and hydrophobic interactions. Integrated ADMET profiling again indicated good pharmacokinetics and acceptable toxicity, while experimental validation confirmed significant growth inhibition. These findings emphasize the potential of citronellol-related aldehydes as antifungal agents and illustrate the added value of coupling docking with predictive pharmacokinetic models in natural product drug discovery [100].
Beyond antifungal activity, C. citratus essential oil has also been evaluated for antioxidant and anti-inflammatory potential. GC–MS analysis identified citral, limonene, and citronellol-related monoterpenes among its constituents. Antioxidant assays (DPPH, ABTS, FRAP) and anti-inflammatory testing (protein denaturation model) confirmed bioactivity, while docking against peroxiredoxin 5 and cyclooxygenase-2 (COX-2) revealed strong binding for caryophyllene oxide and β-caryophyllene. These results were further validated through molecular dynamics simulations, which confirmed stable ligand–protein interactions. Pharmacokinetic predictions suggested high gastrointestinal absorption, good drug-likeness, and low toxicity, reinforcing the therapeutic relevance of citronellol-related compounds in oxidative stress and inflammatory pathways [101].
At a broader scale, large-scale in silico screening of 1175 monoterpenoids from the ChEBI database has provided a systematic perspective on their drug discovery potential. By integrating drug-likeness filters (Lipinski, Veber, Ghose), ADMET predictions, docking studies, and chemoinformatics clustering, the analysis identified several promising scaffolds, with citronellol derivatives emerging as particularly favorable candidates due to their absorption, safety, and binding affinity profiles. Such computational workflows offer a strategic framework for prioritizing monoterpenoids in medicinal chemistry and for guiding future efforts in natural product-based drug discovery [102]. Together, these studies highlight how the combined use of docking, molecular dynamics, and predictive ADME/Tox profiling strengthens the evaluation of citronellol-related monoterpenes as therapeutic agents. This integrative approach not only supports the identification of antifungal, anti-inflammatory, and antioxidant leads but also accelerates the translation of natural product chemistry into rational drug design.
6.3.5. Artificial Intelligence and Systems-Level Approaches in Phytochemical and Biosynthetic Pathway Research
The integration of artificial intelligence (AI) with systems-level analyses is transforming phytochemical and biosynthetic pathway research, offering new opportunities for the discovery and optimization of bioactive natural products, including citronellol and related monoterpenes. Emerging AI-guided workflows, coupled with dynamic simulations, are enhancing the accuracy of bioactivity predictions, accelerating lead identification, and providing rational strategies for pathway engineering. A recently published review (2000–2025) emphasizes the impact of AI-driven methodologies—including ML, deep learning, natural language processing, and network-based approaches—on pathway mining, enzyme-target prediction, and biosynthetic route design [29,31]. Integrated predictive tools such as SwissADME and pkCSM are highlighted as essential for assessing pharmacokinetic and toxicity profiles, thereby supporting compound prioritization during drug development. Together, these advances illustrate the growing synergy between AI platforms and experimental biology, which collectively enhance molecular insights and streamline the rational design of complex biosynthetic pathways [103].
Complementing these developments, the study by Varghese et al, underscores the broader role of AI in metabolite discovery, structural elucidation, and functional annotation. By integrating multi-omics datasets—genomics, transcriptomics, proteomics, and metabolomics—AI facilitates the reconstruction of intricate biosynthetic pathways and regulatory networks in medicinal plants. ML models are increasingly applied to predict the bioactivity, toxicity, and pharmacokinetics of phytochemicals, while AI-assisted platforms such as CASE enhance spectral data interpretation (NMR, MS) for accurate structure determination. Moreover, AI supports chemotaxonomy and comparative metabolomics, enabling species classification and the identification of bioactive chemotypes with therapeutic potential. Despite challenges such as data quality, model interpretability, and the need for standardized benchmarks, AI is positioned as a transformative driver of phytochemical research, offering sustainable and scalable solutions for natural product innovation and drug discovery [104].
7. Applications and Functional Studies
Citronellol exhibits a wide range of biological activities and industrial applications [4]. This section highlights its pharmacological properties, eco-friendly uses, and therapeutic potential explored through network pharmacology.
7.1. Biological Activities
Citronellol is extensively serves as a precursor for the synthesis of other fragrance compounds such as rose oxide and citronellyl esters [81]. Beyond its sensory appeal, citronellol exhibits several biological activities (Figure 3), making it a promising candidate for therapeutic applications [4].
7.1.1. Antimicrobial Activity
Citronellol exhibits broad-spectrum antimicrobial activity through diverse mechanisms and formulations. It has been shown to potentiate the antibacterial efficacy of norfloxacin against S. aureus by inhibiting the NorA efflux pump, as evidenced by reduced minimum inhibitory concentrations (MICs) of ethidium bromide and increased membrane permeability. Molecular docking and ADMET modeling further confirmed its binding affinity to the NorA receptor, supporting its potential as a pharmacologically viable efflux pump inhibitor [105]. A novel class of citronellol-derived methacrylate polymers (PCHM-Xs), incorporating side chain functional groups such as thioether, sulfoxide, sulfone, and ether, was synthesized to investigate structure–activity relationships in bactericidal performance. Among these, the sulfoxide-functionalized polymer (PCHM-SO) demonstrated exceptional antibacterial efficacy (99.99%), attributed to strong interactions with bacterial membranes leading to cytoplasmic leakage and membrane disruption—representing the first systematic evaluation of functional group polarity on citronellol-derived polymer bioactivity [30]. Chemoenzymatic oxidation of citronellol and geraniol into their respective epoxide forms (CT-ox and GR-ox) was employed to enhance their reactivity for grafting onto plasma-activated polymer surfaces. Antibacterial assays against E. coli, S. aureus, and Corynebacterium glutamicum confirmed that epoxidation preserved bioactivity, with the modified terpenoids disrupting membrane integrity and altering cell envelope hydrophobicity, as supported by Zeta Potential measurements, scanning electron microscopy (SEM), and surface energy analyses [106].
Citronellol-functionalized diatomaceous earth (DtQC) achieved over 99.9% bacterial reduction and exhibited broad-spectrum activity against both fungal and bacterial strains, including S. aureus. Structural characterization confirmed successful integration of citronellol and quaternary ammonium salts into the siliceous framework, suggesting DtQC’s potential as a sustainable bioactive hybrid material for indoor coatings and preservation systems [31]. Although (+)-β-citronellol and (β-CD) alone showed limited antibacterial activity against multidrug-resistant strains, their inclusion complex (βCT/β-CD) exhibited inhibitory effects with an MIC of 1024 µg/mL. Moreover, this complex demonstrated a synergistic modulatory effect when combined with gentamicin against E. coli, indicating its potential to enhance antibiotic efficacy [32]. GC-MS analysis of two populations of Thymus pubescens from Mishov-Dagh revealed a novel citronellol-rich chemotype, with citronellol comprising over 42% of the essential oil composition—distinct from previously reported thymol- and carvacrol-dominant profiles. This chemical polymorphism supports the designation of a new chemotype, T. pubescens chemotype Citronellol, whose essential oils exhibited notable antibacterial and antioxidant activities, underscoring its potential for therapeutic and preservative applications [107].
7.1.2. Antifungal Activity
Citronellol has demonstrated broad-spectrum antifungal properties through diverse mechanisms targeting fungal growth, membrane integrity, and virulence factors. Against Colletotrichum camelliae, the causative agent of tea plant anthracnose, citronellol demonstrated strong mycelial inhibition with EC₅₀ values of 76.88 mg/L and 29.79 μL/L air. Microscopic observations and gene expression analyses revealed disruption of fungal cell wall integrity and modulation of chitin- and glucan-associated enzymatic pathways, leading to cellular deformation and growth suppression [108]. In dermatophyte models, citronellol and geraniol significantly inhibited Trichophyton rubrum by suppressing mycelial growth, conidial germination, and fungal colonization on nail fragments. Their antifungal mechanism involved disruption of membrane integrity and inhibition of ergosterol biosynthesis, with additional effects on cell wall architecture, suggesting therapeutic potential for the treatment of dermatophytosis [33]. Both enantiomers of β-citronellol—(R)-(+)- and (S)-(−)—exhibited comparable antifungal activity against C. albicans and C. tropicalis, primarily through membrane disruption without affecting the cell wall. When combined with Amphotericin B, the (R)-(+)-isomer showed interactions ranging from synergistic to indifferent, while the (S)-(−)-isomer demonstrated synergistic, additive, or indifferent effects, indicating promise for combination therapy to enhance antifungal efficacy [68].
The antifungal activity of β-citronellol against C. albicans occurs through multiple interconnected mechanisms (Figure 4). First, it disrupts the fungal cell wall and membrane by targeting key proteins such as Als2p, Rbt1p, and Pga4p. Als2p and Rbt1p are adhesion proteins essential for host cell binding, biofilm formation, and virulence, while Pga4p is an enzyme involved in elongating β-1,3-glucan chains in the fungal cell wall [109]. By suppressing Als2p and Pga4p, β-citronellol reduces adhesion, biofilm development, and membrane stability, leading to impaired cell wall biosynthesis, weakened membrane integrity, and surface wrinkling as observed in SEM studies [39]. Second, β-citronellol induces oxidative stress by suppressing Sod1p, a critical antioxidant enzyme, thereby causing the accumulation of reactive oxygen species (ROS) [39,110].
This ROS overload overwhelms antioxidant defenses, resulting in oxidative damage, apoptosis, and the activation of stress-related proteins such as Hsp30p, Ssa2p, and Sti1p [111]. At the same time, Ddr48p expression is increased as a stress sensor linked to antifungal resistance, while Gst2p, required for filamentous growth under nutrient stress, is reduced [66]. Third, β-citronellol disrupts calcium homeostasis and induces endoplasmic reticulum (ER) stress by depleting Ca²⁺, triggering the unfolded protein response (UPR). The upregulation of Cmd1p (calmodulin) suggests a compensatory response to Ca²⁺ imbalance; however, this disruption ultimately weakens protein folding and stress recovery [39,112]. Fourth, mitochondrial function and ATP synthesis are inhibited through the downregulation of ATP synthase subunits (Atp3p and Atp7p) and respiratory chain proteins (Cox1p and Cobp) [39]. This leads to reduced ATP production, energy depletion, and mitochondrial dysfunction, which are further aggravated by ROS accumulation. Finally, β-citronellol induces apoptosis through the combined effects of ROS generation, mitochondrial impairment, and stress response disruption. Although Cap1p, an AP-1-like transcription factor that protects against oxidative stress, is upregulated as a compensatory mechanism, apoptosis still occurs, indicating that β-citronellol overwhelms the fungal defense system [39]. Overall, β-citronellol exerts its antifungal effects against C. albicans by weakening the cell wall and membrane, inducing oxidative and ER stress, disrupting mitochondrial ATP production, and ultimately triggering apoptosis, thereby reducing fungal virulence and survival.
7.1.3. Anti-Inflammatory Potential
Citronellol, along with geraniol—both major constituents of geranium essential oil—exhibited significant anti-inflammatory activity through modulation of key inflammatory pathways. In LPS-stimulated RAW 264.7 macrophages, both compounds dose-dependently inhibited the production of nitric oxide (NO) and prostaglandin E₂ (PGE₂). Geraniol suppressed both mRNA and protein expression of inducible nitric oxide synthase (iNOS), whereas citronellol primarily inhibited iNOS enzymatic activity. Additionally, both agents attenuated cyclooxygenase-2 (COX-2) expression and reversed activation of the NF-κB signaling pathway, highlighting their potential in managing inflammation-associated disorders [34]. β-Citronellol has further exhibited robust anti-inflammatory, antioxidant, and gastroprotective effects across in vitro, in vivo, and in silico models. In a rat model of indomethacin-induced gastric ulceration, β-citronellol significantly reduced mucosal damage and oxidative stress. Its therapeutic efficacy was associated with upregulation of protective mediators such as COX-1, PGE₂, and endothelial nitric oxide synthase (eNOS), alongside suppression of pro-inflammatory markers including COX-2, 5-lipoxygenase (5-LOX), and intercellular adhesion molecule-1 (ICAM-1). These molecular modulations were accompanied by enhanced antioxidant enzyme activity and improved oxidative balance in gastric tissues, suggesting a dual role in inflammation control and mucosal protection [5]. These findings support citronellol’s potential as a promising natural compound for the development of anti-inflammatory therapeutics, particularly in conditions involving oxidative stress and gastrointestinal inflammation.
In a comprehensive study by Iqbal et al, β-citronellol was evaluated for its anti-inflammatory, antioxidant, and gastro-protective properties using a combination of in vitro, in vivo, and in silico approaches. The compound demonstrated strong protein stabilization effects in vitro, particularly in assays involving bovine serum albumin and red blood cell membranes, with maximum activity observed at 6,400 µg/mL. In vivo, β-citronellol significantly reduced paw edema and protected gastric tissue in rats subjected to indomethacin-induced ulcers. Mechanistic analysis revealed that β-citronellol modulated key inflammatory and oxidative stress markers: it increased levels of COX-1, PGE₂, and eNOS, while suppressing COX-2, 5-LOX, and ICAM-1. Additionally, it enhanced antioxidant defenses by boosting SOD and CAT activity and reducing MDA levels in gastric tissues. These effects were dose-dependent and most pronounced at 50 and 100 mg/kg [5].
7.1.4. Neuroprotective Effects
Citronellol has shown promising neuroprotective properties in preclinical models of neurodegenerative and cerebrovascular disorders. In a rotenone-induced Parkinson’s disease rat model, citronellol significantly attenuated oxidative stress, neuroinflammation, and neuronal apoptosis. It preserved dopaminergic neurons in both the Substantia nigra and striatum, while modulating autophagy-related markers such as LC-3 and p62. Furthermore, it regulated key inflammatory and apoptotic mediators, including COX-2, iNOS, TNF-α, IL-1β, Bax, Bcl-2, and α-synuclein, indicating a multifaceted mechanism of neuroprotection [35]. In a vagotomized ischemic stroke model, β-citronellol enhanced neuronal survival in the hippocampus and improved cognitive performance, particularly memory function. These effects were associated with reduced oxidative stress and neuroinflammation, decreased acetylcholinesterase activity, and improved cerebral blood flow and glucose metabolism. Notably, β-citronellol also modulated gut microbiota composition by increasing the abundance of beneficial bacterial strains, suggesting a contributory role of the gut–brain axis in its neuroprotective mechanism [40]. These findings highlight citronellol’s therapeutic potential in mitigating neurodegenerative processes and promoting neural resilience through antioxidant, anti-inflammatory, and microbiota-mediated pathways.
7.1.5. Anticancer Effects
Citronellol and its structural analogs have demonstrated significant anticancer activity across multiple experimental models, particularly in breast and lung cancer cell lines. Citronellol and citronellal, bioactive constituents of Citrus hystrix leaf extract, exerted potent cytotoxic effects against MDA-MB-231 Triple-Negative Breast Cancer (TNBC) cells. These compounds inhibited cellular proliferation, migration, and colony formation. Mechanistically, they induced cell cycle arrest and apoptosis via downregulation of the anti-apoptotic protein Bcl-2 and activation of the caspase-3-dependent pathway, suggesting therapeutic relevance in aggressive breast cancer subtypes [36].
To enhance anticancer efficacy, citronellol-based esters—CIB, CDB, and CC—were synthesized and evaluated for cytotoxicity. CIB and CDB exhibited strong activity against Artemia salina and MCF-7 breast cancer cells, with low IC₅₀ values of 2.82 and 4.75 μg/mL, respectively. CC was comparatively less effective, indicating that acyclic citronellol esters may serve as promising chemopreventive agents for breast cancer [113]. β-Citronellol significantly reduced cell viability in MCF-7 cells in a dose-dependent manner, with notable cytotoxic effects observed at concentrations as low as 250 µg/mL after 24 and 48 hours of treatment. These findings reinforce its potential as an anticancer agent and support further investigation into its molecular targets and pharmacodynamics [114]. In human non-small cell lung cancer NCI-H1299 cells, citronellol induced necroptosis—a regulated form of cell death—by upregulating TNF-α and RIP1/RIP3 signaling pathways, while concurrently suppressing caspase-3 and -8 activity. This was accompanied by a biphasic accumulation of reactive oxygen species (ROS), contributing to its cytotoxic efficacy in both in vitro and in vivo models [115].
7.1.6. Antinociceptive Effect
Citronellol has demonstrated robust antinociceptive activity across diverse experimental pain models, supporting its potential as a natural analgesic agent. In orofacial pain models induced by formalin, capsaicin, and glutamate, citronellol significantly reduced nociceptive behaviors in mice. Immunofluorescence analysis revealed activation of central nervous system regions, particularly the retrosplenial cortex and periaqueductal grey, suggesting that its analgesic effects are mediated through supraspinal modulation of pain pathways [116]. In inflammatory pain models, citronellol markedly attenuated mechanical hyperalgesia induced by carrageenan, tumor necrosis factor-α (TNF-α), prostaglandin E₂ (PGE₂), and dopamine. These effects were accompanied by reduced paw edema and decreased activation of spinal cord lamina I neurons, indicating a central mechanism of spinal inhibition in its antinociceptive action [41].
Further studies confirmed that citronellol exerts both peripheral and central analgesic effects without impairing motor coordination. It significantly reduced pain behaviors in acetic acid-induced writhing, formalin-induced biphasic pain, and thermal nociception in the hot-plate test. Mechanistically, citronellol suppressed neutrophil infiltration and downregulated pro-inflammatory mediators such as TNF-α and nitric oxide, while modulating inflammatory responses in both peripheral tissues and the central nervous system. These effects are likely linked to its antioxidant properties, which contribute to its overall anti-inflammatory and analgesic profile [117]. Collectively, these findings support citronellol’s potential as a multifaceted antinociceptive agent capable of modulating pain through neuroimmune, inflammatory, and oxidative pathways.
7.1.7. Hypotensive and Vasorelaxant Effects
Citronellol has demonstrated notable cardiovascular bioactivity, particularly in modulating vascular tone and systemic blood pressure. In vivo studies in rodent models revealed dose-dependent hypotensive effects that were unaffected by pharmacological blockade using atropine, hexamethonium, NG-nitro-L-arginine methyl ester (L-NAME), or indomethacin. These findings suggest that citronellol’s mechanism of action is independent of autonomic regulation, nitric oxide synthesis, and prostaglandin-mediated pathways, pointing instead to a direct effect on vascular smooth muscle [42]. Ex vivo assays further support this hypothesis: citronellol induced pronounced vasorelaxation in isolated mesenteric artery rings, primarily through inhibition of calcium influx into smooth muscle cells. This calcium channel blockade underlies its myorelaxant activity and contributes to its antihypertensive potential [42].
Additional evidence comes from studies on Rosa damascena Miller essential oil, which produced concentration-dependent vasorelaxation in rat thoracic aorta segments precontracted with phenylephrine (PE) or potassium chloride (KCl). Notably, the vasorelaxant effect persisted in endothelium-denuded preparations, reinforcing the role of direct smooth muscle action. Among the oil’s constituents, geraniol and β-citronellol were identified as key contributors: geraniol induced relaxation across all tested concentrations (0.4–3.2 µg/mL), while β-citronellol exhibited significant activity at higher doses (1.6–6.4 µg/mL), confirming their involvement in vascular modulation [37]. These findings collectively support citronellol’s potential as a natural vasorelaxant and antihypertensive agent, acting through calcium-dependent smooth muscle relaxation rather than endothelial or neurohumoral pathways.
7.1.8. Myorelaxant and Antispasmodic Effects
β-Citronellol has demonstrated pronounced myorelaxant and antispasmodic properties in preclinical models of airway smooth muscle contractility. In an investigation using isolated rat tracheal rings, β-citronellol significantly relaxed sustained contractions induced by both acetylcholine and high extracellular potassium. Notably, the compound exhibited markedly lower IC₅₀ values for potassium-elicited contractions compared to cholinergic stimuli, indicating preferential targeting of voltage-gated calcium-mediated pathways [43]. These findings suggest that β-citronellol exerts its smooth muscle relaxant effects primarily through inhibition of calcium influx mechanisms, rather than receptor-mediated pathways. The ability to modulate airway tone via direct calcium channel blockade underscores its potential utility in alleviating bronchospasm and other hypercontractile airway conditions.
7.1.9. Anti-Barnacle Activity
Citronellol-derived isothiocyanate compounds have emerged as promising candidates for marine antifouling applications, particularly in the inhibition of cypris larvae settlement in Amphibalanus amphitrite, a major biofouling species. These derivatives exhibit potent bioactivity coupled with favorable safety profiles, as evidenced by high therapeutic indices and minimal larval toxicity. Structural modifications incorporating amide or isocyano functional groups have further enhanced antifouling efficacy while reducing environmental impact, positioning these compounds as viable alternatives to conventional biocides [44]. The targeted mode of action and low ecological footprint of citronellol-based isothiocyanates align with current priorities in sustainable marine coatings and biofouling management. Their ability to disrupt larval adhesion and development without inducing broad-spectrum toxicity supports their integration into environmentally responsible antifouling strategies. These findings reinforce the broader utility of citronellol derivatives in green chemistry applications and highlight their potential for innovation in marine biotechnology [44].
7.1.10. Anti-Allergic Activity
Citronellol, the principal component of geranium (P. graveolens) essential oil, has demonstrated significant anti-allergic activity through modulation of key immunological pathways. In vitro studies revealed that citronellol inhibited mast cell degranulation and suppressed immunoglobulin E (IgE)-induced tumor necrosis factor-α (TNF-α) production in a dose-dependent manner. Notably, the L-enantiomer exhibited superior efficacy compared to the D-enantiomer, suggesting stereoselective pharmacological behavior [45]. Mechanistically, citronellol attenuated extracellular signal-regulated kinase (ERK) phosphorylation, a critical step in the inflammatory signaling cascade associated with allergic responses. These findings underscore its potential as a therapeutic agent for allergic disorders, particularly those mediated by mast cell activation and IgE-dependent cytokine release.
7.1.11. Antifibrotic Agent
β-Citronellol has demonstrated promising antifibrotic properties in hepatic stellate cell models. In transforming growth factor-β1 (TGF-β1)-stimulated LX-2 cells, β-citronellol significantly attenuated fibrogenic activation by downregulating key markers associated with hepatic fibrosis, including ACTA2 (α-smooth muscle actin), COL1A1 (collagen type I alpha 1), TIMP1 (tissue inhibitor of metalloproteinases-1), and SMAD2. Additionally, it reduced matrix metalloproteinase-9 (MMP-9) secretion and modulated mitogen-activated protein kinase (MAPK) signaling pathways, suggesting a multifactorial mechanism of action [46]. These findings support β-citronellol’s potential as a natural antifibrotic agent capable of targeting both extracellular matrix remodeling and intracellular signaling cascades involved in liver fibrosis progression.
7.1.12. Antidiabetic Effects
Citronellol has demonstrated notable antidiabetic activity in streptozotocin-induced diabetic rat models. Treatment with citronellol significantly reduced plasma glucose and glycated hemoglobin (HbA1C) levels, while concurrently enhancing serum insulin, hemoglobin concentration, and hepatic glycogen content. These metabolic improvements were accompanied by restoration of key hepatic carbohydrate metabolism enzymes, indicating a normalization of glucose homeostasis [118]. Histopathological analysis further revealed preservation of normal liver architecture and pancreatic β-cell morphology, suggesting protective effects against diabetes-induced tissue damage. Collectively, these findings support citronellol’s potential as a nutraceutical agent for diabetes management, acting through glycemic regulation, enzymatic restoration, and tissue-level protection.
7.1.13. Anticonvulsant Activity
Citronellol has demonstrated significant anticonvulsant activity in rodent models, indicating its potential for modulating neuronal excitability and seizure susceptibility. In chemically induced seizure paradigms, citronellol reduced the incidence of convulsions triggered by pentylenetetrazol and picrotoxin. Additionally, in maximal electroshock-induced seizure models, it eliminated the extensor reflex in approximately 80% of treated animals, suggesting efficacy across multiple seizure types [47]. Electrophysiological assessments further revealed that citronellol decreased the amplitude of compound action potentials in isolated nerve preparations, implicating a direct effect on neuronal excitability. These findings support citronellol’s potential as a natural anticonvulsant agent, possibly acting through modulation of ion channel activity and synaptic transmission.
7.1.14. Antispasmodic Effects
Citronellol, in conjunction with geraniol and Rosa damascena essential oil, has demonstrated significant antispasmodic activity in isolated rat ileum preparations. The compounds effectively inhibited contractions induced by potassium chloride (KCl), acetylcholine, and electrical field stimulation. Among the tested agents, citronellol exhibited the most potent spasmolytic effect, with lower IC₅₀ values than the essential oil itself—highlighting its predominant role in mediating gastrointestinal smooth muscle relaxation and supporting the traditional use of rose water for digestive relief [48]. Further investigations into β-citronellol revealed biphasic cardiovascular and respiratory effects in vivo. The initial rapid phase (P1) was characterized by hypotension, bradycardia, and apnea, mediated by chemosensitive pulmonary vagal afferents. This phase was abolished by vagotomy and perivagal capsaicin treatment, confirming its neurogenic origin. The delayed phase (P2) involved direct cardioinhibitory and vasodilatory actions, which were sensitive to purinergic signaling blockade. These findings delineate distinct neural and vascular mechanisms underlying β-citronellol’s systemic effects and suggest its broader potential in modulating smooth muscle tone across organ systems [38].
7.2. Use in Green Insect Repellents and Cosmetics
Citronellol is widely used in natural insect repellents, often in combination with citronellal and geraniol. It acts by masking human odors and interfering with mosquito olfactory receptors, offering a safer alternative to synthetic repellents like DEET [49]. Common applications include:
7.2.1. Citronella Candles and Sprays for Short-Term Mosquito Deterrence
Citronella oil and its major constituents—citronellol and citronellal—have long been recognized for their mosquito-repellent properties. However, their efficacy is highly dependent on the mode of delivery. Comparative studies with other botanical monoterpenes, such as geraniol and linalool, have revealed that conventional formats like candles offer limited protection, with markedly lower indoor repellency. In contrast, sustained-release systems, including diffusers and wearable devices, have demonstrated significantly enhanced efficacy, both in controlled environments and field conditions. These findings underscore the importance of formulation strategy in maximizing the protective potential of citronella-based products [119]. To address the inherent volatility and short duration of action associated with citronella compounds, encapsulation techniques have been explored. Inclusion complexes with β-cyclodextrin (β-CD) have shown promise in improving physicochemical stability and prolonging repellent activity [50]. Analytical characterization confirmed successful complexation, and oil-in-water formulations containing these complexes exhibited slower release rates compared to uncomplexed oils. Human-bait assays further validated the enhanced efficacy of citronella oil–β-CD and citronellol–β-CD complexes, suggesting that encapsulation not only improves longevity but also enhances dermal compatibility—an important consideration for personal care applications [50].
Beyond encapsulation, chemical modification of citronella constituents has emerged as a viable strategy for improving repellent performance. Structural derivatives, such as hydroxylated cyclic acetals of citronellal, have been synthesized to reduce volatility and odor while extending protection duration [49]. Laboratory and field evaluations demonstrated that these modified compounds can achieve repellency levels comparable to synthetic agents like DEET and Icaridin, even at reduced dosages. Given the structural similarity between citronellal and citronellol, similar optimization approaches may be applicable to citronellol, reinforcing its potential as a sustainable and effective alternative to conventional repellents [49].
7.2.2. Topical Lotions and Wristbands for Personal Protection
The effectiveness of citronella-derived repellents has been extensively evaluated across both laboratory and semi-field settings. While citronella oil is widely incorporated into topical formulations and wearable devices, its performance is highly dependent on the delivery system employed. Comparative field trials assessing commercially available mosquito control products revealed that citronella wristbands—despite containing relatively high concentrations of active oil—did not significantly reduce mosquito landings. In contrast, devices utilizing sustained-release technologies, such as metofluthrin-based clip-ons and multi-oil diffusers, demonstrated markedly superior protection, achieving over 90% efficacy against Aedes albopictus and Culex pipiens. These findings underscore the critical role of controlled volatilization in ensuring consistent repellent activity and highlight the limitations of passive delivery formats like wristbands [120].
In parallel, the safety profile of citronellol, a key component of citronella oil, has been investigated with respect to dermal absorption. Studies using radiolabeled compounds applied to human cadaver skin indicate that citronellol exhibits low percutaneous penetration, with minimal systemic uptake even at elevated concentrations. The majority of the applied dose was removed during post-application washing, suggesting limited bioavailability and favorable safety for topical use. However, its classification as a potential allergen under European cosmetic regulations necessitates careful formulation strategies and transparent labeling [51]. To mitigate dermal exposure while preserving repellent efficacy, encapsulation and controlled-release approaches have been proposed. These technologies offer the dual advantage of sustained volatilization and reduced direct skin contact, thereby enhancing both safety and performance [51].
7.2.3. Essential Oil Diffusers for Ambient Repellent Effects
Citronella oil and its constituents, citronellol and citronellal, have been incorporated into diffuser-based formulations not only for their insect-repellent activity but also for their multifunctional roles in wellness and cosmetic applications. A laboratory study on reed diffusers combining Cinnamomum verum (cinnamon) and Cymbopogon nardus (citronella) essential oils demonstrated that the blend maintained stable odor and color during storage. Hedonic evaluations identified a 2:3 cinnamon-to-citronella ratio as the most preferred by users, while behavioral assays in mice revealed reduced stress-related responses following exposure. These findings suggest that citronella-based diffuser blends may provide dual benefits by delivering both aromatherapeutic and mosquito-repellent effects [121]. Beyond ambient wellness applications, essential oil vapors have been investigated for pest management in stored products. In sealed storage trials, cinnamon and citronella vapors significantly suppressed Callosobruchus chinensis populations in green gram seeds over a two-month period. While cinnamon exhibited stronger fumigant action, citronella oil provided substantial protection without impairing seed germination or organoleptic quality. By contrast, cinnamon imparted undesirable off-flavors, underscoring citronella’s advantage in maintaining consumer acceptability. These results highlight the value of citronellol-rich essential oils as eco-friendly grain protectants with potential applications in integrated pest management and green storage systems [122].
Within cosmetic formulations, citronellol is widely utilized for its floral fragrance and possible skin-soothing properties, with applications in moisturizers, perfumes, shampoos, and bath products. Although generally regarded as safe, regulatory restrictions exist in some regions due to its potential as a contact allergen, necessitating careful formulation and labeling practices. Recent advances have also explored citronellol as a component of natural deep eutectic solvents (NADES). A study developed hydrophobic NADES using essential oil-derived compounds, including citronellol, thymol, eugenol, and 1,8-cineole, as building blocks. These NADES were optimized for dispersive liquid–liquid microextraction (DLLME) of regulated volatile compounds from water-based fragrances, followed by GC-FID and GC-MS analysis. The approach yielded high recovery rates, precision, and linearity, while reducing reliance on conventional organic solvents. Incorporating citronellol into NADES not only promotes greener analytical methodologies but also reinforces its multifunctionality across fragrance formulation, quality control, and sustainable cosmetic innovation [52].
8. Challenges and Future Prospects
Despite significant advances in metabolic engineering, synthetic biology, and bioprocess development, the microbial production of citronellol still faces several technical and physiological hurdles. This section outlines key limitations, strategies to overcome them, and emerging technologies poised to reshape the future of citronellol biosynthesis.
8.1. Limitations in Yield, Toxicity, and Pathway Bottlenecks
Microbial production of monoterpenes such as citronellol is frequently limited by natural availability, incomplete pathway characterization, low biocatalyst specificity and regulatory complexity [53]. Only a few terpenoids like citronellol are available in large quantities at reasonable cost, which restricts their industrial use [54]. The catabolic pathways for many acyclic monoterpenes, including citronellol, are not fully understood. This hampers the development of efficient biotransformation processes [55]. Naturally occurring pseudomonads may not possess high specificity or efficiency for citronellol transformation, making microbial production less predictable [123]. Gene clusters like atu and liu involved in citronellol degradation are tightly regulated, and manipulating them for production rather than degradation is challenging [124]. Collectively, these challenges underscore the need for integrated strategies that optimize both pathway engineering and host physiology to enhance monoterpene biosynthesis.
8.2. Strategies for Improving Microbial Tolerance and Efficiency
To enhance microbial robustness and productivity, a range of engineering strategies like biocatalyst engineering, synthetic biology approaches, green chemistry applications and expanding substrate range has been developed [18]. There's strong potential in engineering pseudomonads or other microbes to enhance their ability to convert abundant terpenoids into valuable derivatives like citronellol [123]. Incorporating synthetic pathways into robust microbial hosts could allow scalable citronellol biosynthesis from simple carbon sources [123]. Citronellol’s role as an aroma compound makes it a prime candidate for sustainable production in food, cosmetics, and pharmaceuticals. Understanding and modifying the substrate specificity of key enzymes (e.g., alcohol dehydrogenases, CoA ligases) could broaden the range of terpenoids that can be converted into citronellol or its derivatives. Collectively, these strategies strengthen microbial resilience and support scalable monoterpene production under industrial conditions.
8.3. Emerging Technologies: AI-Guided Metabolic Design and Machine Learning in Fermentation Control
AI and ML are revolutionizing biomanufacturing by enabling predictive modeling, real-time optimization, and intelligent control systems. AI-guided metabolic design platforms such as TeselaGen and AI4b.io employ predictive algorithms to simulate enzyme kinetics, pathway fluxes, and host responses, thereby accelerating strain development and minimizing experimental burden [125,126]. In fermentation control, ML models trained on historical process data can forecast yields, detect anomalies, and recommend dynamic adjustments; neural networks and support vector machines have been successfully applied to optimize ethanol production and microbial growth dynamics [127,128]. Smart fermentation technologies, integrating biosensors, Internet of Things (IoT) devices, and ML algorithms, allow real-time monitoring of key parameters such as pH, dissolved oxygen, and metabolite concentrations, enhancing the consistency and scalability of citronellol production [129]. Furthermore, physics-informed neural networks (PINNs), which embed physical laws (e.g., Navier–Stokes equations) into ML architectures, offer computationally efficient alternatives to conventional computational fluid dynamics (CFD) simulations for modeling fluid flow and microbial behavior in large-scale bioreactors [130]. As these technologies continue to evolve, they hold significant promise for overcoming current limitations and advancing precision biomanufacturing.
9. Conclusions
Over the past decade, significant progress has been made in understanding and engineering the biosynthesis of citronellol. Natural biosynthetic pathways have been elucidated in plant species such as P. graveolens and Rosa spp., revealing both cytosolic and plastidic routes mediated by Nudix hydrolases, citral reductases, and SDR enzymes. Advances in metabolic engineering and synthetic biology have enabled microbial production of citronellol through heterologous expression of key enzymes—including GPPS, GES, and OYE homologs—in microbial hosts such as E. coli, S. cerevisiae, and Y. lipolytica. Biotransformation strategies employing whole-cell systems and purified enzymes have facilitated the selective conversion of citronellol into high-value fragrance compounds such as rose oxide. Innovations in fermentation and bioprocessing—including gas-phase bioreactors, biphasic systems, and adaptive control mechanisms—have addressed key challenges in scalability and process efficiency. Analytical platforms such as GC-MS, chiral HPLC, and molecular docking have supported compound characterization, enantiomeric resolution, and bioactivity prediction. Functional studies have further demonstrated citronellol’s antioxidant, anti-inflammatory, and neuroprotective properties, with network pharmacology revealing its potential as a multi-target therapeutic agent. Collectively, these developments underscore citronellol’s promise as a bio-based compound for applications spanning fragrance, pharmaceuticals, and agrochemicals.
Looking ahead, emerging technologies are poised to catalyze the next phase of innovation. Systematic strain engineering has already achieved record production titers in S. cerevisiae through integration of mevalonate pathway genes, NADPH-enhancing modules, and transporter proteins. AI and ML are enabling predictive metabolic modeling, real-time optimization, and intelligent bioprocess control. PINNs and digital twins offer scalable, computationally efficient frameworks for simulating bioreactor dynamics and microbial performance. In parallel, principles of green chemistry and the circular bioeconomy are driving the valorization of agro-industrial residues and renewable feedstocks for sustainable citronellol production. Emerging approaches such as synthetic ecology and microbial consortia may unlock novel co-culture strategies for monoterpene biosynthesis and VOC mitigation. As these technologies mature, they hold the potential to overcome current limitations in yield, toxicity, and scalability—paving the way for industrial-scale production of high-purity citronellol through environmentally responsible and economically viable bioprocesses.
Data availability Statement
All data generated or analyzed during this study are included in this article.
Conflicts of Interest
There is no conflict of interest.
References
- Martinelli, L.; Bihanic, C.; Bony, A.; Gros, F.; Conart, C.; Fiorucci, S.; et al. Citronellol biosynthesis in pelargonium is a multistep pathway involving progesterone 5β-reductase and/or iridoid synthase-like enzymes. Plant Physiology. 2024, 194, 1006–1023. [Google Scholar] [CrossRef]
- Bergman, M.E.; Bhardwaj, M.; Phillips, M.A. Cytosolic geraniol and citronellol biosynthesis require a Nudix hydrolase in rose-scented geranium (Pelargonium graveolens). The Plant Journal. 2021, 107, 493–510. [Google Scholar] [CrossRef]
- Li, H.; Li, Y.; Yan, H.; Bao, T.; Shan, X.; Caissard, J.-C.; et al. The complexity of volatile terpene biosynthesis in roses: Particular insights into β-citronellol production. Plant Physiology. 2024, 196, 1908–1922. [Google Scholar] [CrossRef]
- Santos, P.L.; Matos, J.P.S.; Picot, L.; Almeida, J.R.; Quintans, J.S.; Quintans-Junior, L.J. Citronellol, a monoterpene alcohol with promising pharmacological activities-A systematic review. Food and Chemical Toxicology. 2019, 123, 459–469. [Google Scholar] [CrossRef]
- Iqbal, U.; Malik, A.; Sial, N.T.; Mehmood, M.H.; Nawaz, S.; Papadakis, M.; et al. β-Citronellol: a potential anti-inflammatory and gastro-protective agent-mechanistic insights into its modulatory effects on COX-II, 5-LOX, eNOS, and ICAM-1 pathways through in vitro, in vivo, in silico, and network pharmacology studies. Inflammopharmacology. 2024, 32, 3761–3784. [Google Scholar] [CrossRef]
- Cravens, A.; Payne, J.; Smolke, C.D. Synthetic biology strategies for microbial biosynthesis of plant natural products. Nature communications. 2019, 10, 2142. [Google Scholar] [CrossRef]
- Li, Y.; Pfeifer, B.A. Heterologous production of plant-derived isoprenoid products in microbes and the application of metabolic engineering and synthetic biology. Current opinion in plant biology. 2014, 19, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Vavitsas, K.; Fabris, M.; Vickers, C.E. Terpenoid metabolic engineering in photosynthetic microorganisms. Genes. 2018, 9, 520. [Google Scholar] [CrossRef]
- Jiang, G.; Yao, M.; Wang, Y.; Xiao, W.; Yuan, Y. A “push-pull-restrain” strategy to improve citronellol production in Saccharomyces cerevisiae. Metabolic Engineering. 2021, 66, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Ribeaucourt, D.; Hofler, G.T.; Yemloul, M.; Bissaro, B.; Lambert, F.; Berrin, J.-G.; et al. Tunable production of (R)-or (S)-citronellal from geraniol via a bienzymatic cascade using a copper radical alcohol oxidase and old yellow enzyme. ACS catalysis. 2022, 12, 1111–1116. [Google Scholar] [CrossRef] [PubMed]
- Mermat, N.; Ferroukhi, O.; Peulon-Agasse, V.; Bayle, J.P.; Guermouche, M.H.; Cardinael, P. Original mesogenic citronellol-based stationary phase for both normal-and reversed-phase HPLC modes: properties and applications. Chromatographia. 2020, 83, 1495–1508. [Google Scholar] [CrossRef]
- Ye, M.; Ye, Y.; Du, Z.; Chen, G. Cell-surface engineering of yeasts for whole-cell biocatalysts. Bioprocess and biosystems engineering. 2021, 44, 1003–1019. [Google Scholar] [CrossRef] [PubMed]
- Lecourt, M.; Chietera, G.; Jullien, F.; Blerot, B.; Antoniotti, S. From biosynthesis in plants to post-biosynthetic enzymatic conversion. Generation of odor-impact rose oxides from citronellol-rich essential oils. Botany Letters. 2023, 170, 15–27. [Google Scholar]
- Zhang, B.; Du, H.; Zheng, Y.; Sun, J.; Shen, Y.; Lin, J.; et al. Design and engineering of whole-cell biocatalyst for efficient synthesis of (R)-citronellal. Microbial Biotechnology. 2022, 15, 1486–1498. [Google Scholar] [CrossRef] [PubMed]
- de Miranda, A.S.; Milagre, C.D.; Hollmann, F. Alcohol dehydrogenases as catalysts in organic synthesis. Frontiers in Catalysis. 2022, 2, 900554. [Google Scholar] [CrossRef]
- Chadha, A.; Padhi, S.K.; Stella, S.; Venkataraman, S.; Saravanan, T. Microbial alcohol dehydrogenases: recent developments and applications in asymmetric synthesis. Organic & Biomolecular Chemistry. 2024, 22, 228–251. [Google Scholar]
- Rudzka, A.; Zdun, B.; Antos, N.; Montero, L.M.; Reiter, T.; Kroutil, W.; et al. Biocatalytic characterization of an alcohol dehydrogenase variant deduced from Lactobacillus kefir in asymmetric hydrogen transfer. Communications Chemistry. 2023, 6, 217. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A.; Woodley, J.M. Role of biocatalysis in sustainable chemistry. Chemical reviews. 2018, 118, 801–838. [Google Scholar] [CrossRef]
- Malinowski, J.J. Two-phase partitioning bioreactors in fermentation technology. Biotechnology advances. 2001, 19, 525–538. [Google Scholar] [CrossRef]
- Doig, S.; Boam, A.; Leak, D.; Livingston, A.; Stuckey, D. Optimisation of the kinetics of the stereoselective reduction of geraniol to citronellol in a two liquid phase system. Biocatalysis and Biotransformation. 1998, 16, 27–44. [Google Scholar] [CrossRef]
- Li, T.; Liu, X.; Xiang, H.; Zhu, H.; Lu, X.; Feng, B. Two-phase fermentation systems for microbial production of plant-derived terpenes. Molecules. 2024, 29, 1127. [Google Scholar] [CrossRef]
- Subash, P.; Kumar, K.S.; Rao, K.S.; Khute, S. Advanced Chromatography Analytical Methods for the Isolation and Identification of Natural Drug Molecules. 2025.
- Chen, L.; Dean, B.; Liang, X. A technical overview of supercritical fluid chromatography-mass spectrometry (SFC-MS) and its recent applications in pharmaceutical research and development. Drug Discovery Today: Technologies. 2021, 40, 69–75. [Google Scholar] [CrossRef]
- Vamshi, G.; DSNBKP; Sampath, A.; Dammalli, M.; Kumar, P.; BS G; et al. Possible cerebroprotective effect of citronellal: molecular docking, MD simulation and in vivo investigations. Journal of Biomolecular Structure and Dynamics. 2024, 42, 1208–1219. [Google Scholar]
- Santos, P.L.; Brito, R.G.; Oliveira, M.A.; Quintans, J.S.; Guimaraes, A.G.; Santos, M.R.; et al. Docking, characterization and investigation of β-cyclodextrin complexed with citronellal, a monoterpene present in the essential oil of Cymbopogon species, as an anti-hyperalgesic agent in chronic muscle pain model. Phytomedicine. 2016, 23, 948–957. [Google Scholar] [CrossRef]
- Okoli, B.J.; Ladan, Z.; Mtunzi, F.; Hosea, Y.C. Vitex negundo L. Essential oil: odorant binding protein efficiency using molecular docking approach and studies of the mosquito repellent. Insects. 2021, 12, 1061. [Google Scholar] [CrossRef] [PubMed]
- Patsilinakos, A. Machine learning applications to essential oils and natural extracts. 2020.
- Sharma, A.D.; Kaur, I.; Chauhan, A. In vitro anti-mucormycosis and anti-aspergillosis potential of essential oils and molecular docking of principal components from Cymbopogon khasianus and Cymbopogon citratus. Chemistry Africa. 2023, 6, 2835–2848. [Google Scholar] [CrossRef]
- Reinhardt, J.K.; Craft, D.; Weng, J.-K. Toward an integrated omics approach for plant biosynthetic pathway discovery in the age of AI. Trends in Biochemical Sciences. 2025. [Google Scholar] [CrossRef] [PubMed]
- Yook, J.; Jeong, D.; Lee, J.-C. Synthesis of citronellol-derived antibacterial polymers and effect of thioether, sulfoxide, sulfone, and ether functional groups on their bactericidal activity. Macromolecules. 2023, 56, 3406–3420. [Google Scholar] [CrossRef]
- Lopez, G.P.; Roque, L.B.; Igal, K.; Espinosa, E.G.; Bellotti, N. Citronellol-functionalized natural silica: a biogenic approach for antifungal and antibacterial material applications. Frontiers in Chemistry. 2025, 13, 1535787. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.L.S.; Campina, F.F.; do Socorro Costa, M.; da Cruz, R.P.; de Freitas, T.S.; Dos Santos, A.T.L.; et al. Antibacterial and modulatory activities of β-cyclodextrin complexed with (+)-β-citronellol against multidrug-resistant strains. Microbial Pathogenesis. 2021, 156, 104928. [Google Scholar] [CrossRef]
- Pereira, F.d.O.; Mendes, J.M.; Lima, I.O.; Mota, K.S.d.L.; Oliveira WAd Lima, E.d.O. Antifungal activity of geraniol and citronellol, two monoterpenes alcohols, against Trichophyton rubrum involves inhibition of ergosterol biosynthesis. Pharmaceutical biology. 2015, 53, 228–234. [Google Scholar] [CrossRef]
- Su, Y.-W.; Chao, S.-H.; Lee, M.-H.; Ou, T.-Y.; Tsai, Y.-C. Inhibitory effects of citronellol and geraniol on nitric oxide and prostaglandin E2 production in macrophages. Planta medica. 2010, 76, 1666–1671. [Google Scholar] [CrossRef]
- Jayaraj, R.L.; Azimullah, S.; Parekh, K.A.; Ojha, S.K.; Beiram, R. Effect of citronellol on oxidative stress, neuroinflammation and autophagy pathways in an in vivo model of Parkinson's disease. Heliyon 2022, 8(. [Google Scholar] [CrossRef]
- Ho, Y.; Suphrom, N.; Daowtak, K.; Potup, P.; Thongsri, Y.; Usuwanthim, K. Anticancer effect of Citrus hystrix DC. leaf extract and its bioactive constituents citronellol and, citronellal on the triple negative breast cancer MDA-MB-231 cell line. Pharmaceuticals. 2020, 13, 476. [Google Scholar]
- Demirel, S. Geraniol and β-citronellol participate in the vasorelaxant effects of Rosa damascena Miller essential oil on the rat thoracic aorta. Fitoterapia. 2022, 161, 105243. [Google Scholar] [CrossRef]
- Ribeiro-Filho, H.V.; de Souza Silva, C.M.; de Siqueira, R.J.; Lahlou, S.; dos Santos, A.A.; Magalhães, P.J.C. Biphasic cardiovascular and respiratory effects induced by β-citronellol. European journal of pharmacology. 2016, 775, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Buakaew, W.; Pankla Sranujit, R.; Noysang, C.; Krobthong, S.; Yingchutrakul, Y.; Thongsri, Y.; et al. Proteomic analysis reveals proteins involved in the mode of action of β-Citronellol identified from Citrus hystrix DC. Leaf against Candida albicans. Frontiers in microbiology. 2022, 13, 894637. [Google Scholar]
- Yue, Y.; Li, C.; Zhang, T.; Park, S. Neuroprotective Effects of SELFormer-Selected β-Citronellol and β-Caryophyllene in Vagotomized Ischemic Stroke Model Through Direct Brain Protection and Gut Microbiota Modulation. BioFactors. 2025, 51, e70031. [Google Scholar] [CrossRef] [PubMed]
- Brito, R.G.; Dos Santos, P.L.; Quintans, J.S.; de Lucca Júnior, W.; Araújo, A.A.; Saravanan, S.; et al. Citronellol, a natural acyclic monoterpene, attenuates mechanical hyperalgesia response in mice: Evidence of the spinal cord lamina I inhibition. Chemico-Biological Interactions. 2015, 239, 111–117. [Google Scholar] [CrossRef]
- Bastos, J.F.; Moreira, Í.J.; Ribeiro, T.P.; Medeiros, I.A.; Antoniolli, A.R.; De Sousa, D.P.; et al. Hypotensive and vasorelaxant effects of citronellol, a monoterpene alcohol, in rats. Basic & Clinical Pharmacology & Toxicology. 2010, 106, 331–337. [Google Scholar]
- Vasconcelos, T.; Ribeiro-Filho, H.; Lucetti, L.; Magalhães, P. β-Citronellol, an alcoholic monoterpene with inhibitory properties on the contractility of rat trachea. Brazilian Journal of Medical and Biological Research. 2015, 49, e4800. [Google Scholar] [CrossRef]
- Tanikawa, A.; Fujihara, T.; Nakajima, N.; Maeda, Y.; Nogata, Y.; Yoshimura, E.; et al. Anti-Barnacle Activities of Isothiocyanates Derived from β-Citronellol and Their Structure–Activity Relationships. Chemistry & Biodiversity. 2023, 20, e202200953. [Google Scholar]
- Kobayashi, Y.; Sato, H.; Yorita, M.; Nakayama, H.; Miyazato, H.; Sugimoto, K.; et al. Inhibitory effects of geranium essential oil and its major component, citronellol, on degranulation and cytokine production by mast cells. Bioscience, Biotechnology, and Biochemistry. 2016, 80, 1172–1178. [Google Scholar] [CrossRef] [PubMed]
- Buakaew, W.; Krobthong, S.; Yingchutrakul, Y.; Potup, P.; Thongsri, Y.; Daowtak, K.; et al. Investigating the antifibrotic effects of β-citronellol on a TGF-β1-stimulated LX-2 hepatic stellate cell model. Biomolecules. 2024, 14, 800. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, D.P.; Gonçalves, J.C.R.; Quintans-Júnior, L.; Cruz, J.S.; Araújo, D.A.M.; de, A.l.m.e.i.d.a.RN. Study of anticonvulsant effect of citronellol, a monoterpene alcohol, in rodents. Neuroscience letters. 2006, 401, 231–235. [Google Scholar] [CrossRef]
- Sadraei, H.; Asghari, G.; Emami, S. Inhibitory effect of Rosa damascena Mill flower essential oil, geraniol and citronellol on rat ileum contraction. Research in Pharmaceutical Sciences. 2013, 8, 17. [Google Scholar]
- Iovinella, I.; Caputo, B.; Cobre, P.; Manica, M.; Mandoli, A.; Dani, F.R. Advances in mosquito repellents: effectiveness of citronellal derivatives in laboratory and field trials. Pest Management Science. 2022, 78, 5106–5112. [Google Scholar] [CrossRef]
- Songkro, S.; Hayook, N.; Jaisawang, J.; Maneenuan, D.; Chuchome, T.; Kaewnopparat, N. Investigation of inclusion complexes of citronella oil, citronellal and citronellol with β-cyclodextrin for mosquito repellent. Journal of Inclusion Phenomena and Macrocyclic Chemistry. 2012, 72, 339–355. [Google Scholar] [CrossRef]
- Gilpin, S.; Hui, X.; Maibach, H. In vitro human skin penetration of geraniol and citronellol. DERM. 2010, 21, 41–48. [Google Scholar] [CrossRef]
- Cagliero C, Bechis G, Marengo A, Sgorbini B, Bicchi C, Rubiolo P, editors. Essential oil components as GC-compatible hydrophobic deep eutectic solvents to extract regulated compounds from water-based fragrances. 53rd INTERNATIONAL SYMPOSIUM ON ESSENTIAL OILS Book of Abstract; 2023.
- Wang, X.; Zhang, X.; Zhang, J.; Zhou, Y.; Wang, F.; Wang, Z.; et al. Advances in microbial production of geraniol: From metabolic engineering to potential industrial applications. Critical Reviews in Biotechnology. 2025, 45, 727–742. [Google Scholar] [CrossRef]
- Singh, K.; Kaloni, D.; Sehgal, K.; Pan, S.; Sarethy, I.P. Essential oils: An update on their biosynthesis and genetic strategies to overcome the production challenges. Plant-derived bioactives: production, properties and therapeutic applications. 2020:33-60.
- Förster-Fromme, K.; Jendrossek, D. Catabolism of citronellol and related acyclic terpenoids in pseudomonads. Applied microbiology and biotechnology. 2010, 87, 859–869. [Google Scholar] [CrossRef]
- Devi, K.; Mishra, S.K.; Sahu, J.; Panda, D.; Modi, M.K.; Sen, P. Genome wide transcriptome profiling reveals differential gene expression in secondary metabolite pathway of Cymbopogon winterianus. Scientific reports. 2016, 6, 21026. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhang, H.; Yijing, W.; Wan, X.; Ming, C.; Huitang, P.; et al. Functional characterization of LcTPS1 and LcTPS14 explains the biosynthesis of citronellol, citronellal and linalool in Lagerstroemia caudata. Industrial Crops and Products. 2024, 209, 118033. [Google Scholar]
- Zhao, Q.; Zhang, M.; Gu, L.; Yang, Z.; Li, Y.; Luo, J.; et al. Transcriptome and volatile compounds analyses of floral development provide insight into floral scent formation in Paeonia lactiflora ‘Wu Hua Long Yu’. Frontiers in Plant Science. 2024, 15, 1303156. [Google Scholar] [CrossRef] [PubMed]
- Bergman, M.E.; Chávez, Á.; Ferrer, A.; Phillips, M.A. Distinct metabolic pathways drive monoterpenoid biosynthesis in a natural population of Pelargonium graveolens. Journal of Experimental Botany. 2020, 71, 258–271. [Google Scholar] [CrossRef]
- Iijima, M.; Kenmoku, H.; Takahashi, H.; Lee, J.-B.; Toyota, M.; Asakawa, Y.; et al. Characterization of 12-oxophytodienoic acid reductases from rose-scented geranium (Pelargonium graveolens). Natural Product Communications. 2016, 11, 1934578X1601101201. [Google Scholar] [CrossRef]
- Xu, H.; Bohman, B.; Wong, D.C.; Rodriguez-Delgado, C.; Scaffidi, A.; Flematti, G.R.; et al. Complex sexual deception in an orchid is achieved by co-opting two independent biosynthetic pathways for pollinator attraction. Current Biology. 2017, 27, 1867–1877. [Google Scholar] [CrossRef] [PubMed]
- Vranová, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annual review of plant biology. 2013, 64, 665–700. [Google Scholar] [CrossRef]
- Blerot, B.; Martinelli, L.; Prunier, C.; Saint-Marcoux, D.; Legrand, S.; Bony, A.; et al. Functional analysis of four terpene synthases in rose-scented Pelargonium cultivars (Pelargonium× hybridum) and evolution of scent in the Pelargonium genus. Frontiers in Plant Science. 2018, 9, 1435. [Google Scholar] [CrossRef]
- Bohrer, A.-S.; Kopriva, S.; Takahashi, H. Plastid-cytosol partitioning and integration of metabolic pathways for APS/PAPS biosynthesis in Arabidopsis thaliana. Frontiers in Plant Science. 2015, 5, 751. [Google Scholar] [CrossRef]
- Morcelli, S.R.; Bull, É.S.; Terra, W.S.; Moreira, R.O.; Borges, F.V.; Kanashiro, M.M.; et al. Synthesis, characterization and antitumoral activity of new cobalt (II) complexes: Effect of the ligand isomerism on the biological activity of the complexes. Journal of Inorganic Biochemistry. 2016, 161, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, N.; Aseri, M.L.; Padmanabhan, D. A review of drug isomerism and its significance. International journal of applied and basic medical research. 2013, 3, 16–18. [Google Scholar] [CrossRef] [PubMed]
- Elsharif, S.A.; Buettner, A. Influence of the chemical structure on the odor characters of β-citronellol and its oxygenated derivatives. Food Chemistry. 2017, 232, 704–711. [Google Scholar] [CrossRef]
- Silva, D.; Diniz-Neto, H.; Cordeiro, L.; Silva-Neta, M.; Silva, S.; Andrade-Júnior, F.; et al. (R)-(+)-β-Citronellol and (S)-(−)-β-Citronellol in Combination with Amphotericin B against Candida spp. International journal of molecular sciences. 2020, 21, 1785. [Google Scholar] [CrossRef]
- Li, L.; Jiang, H.; Xu, T.; Wang, X. Engineering Bienzymatic Cascade for Efficient Biosynthesis of Citronellal and Citronellol. Biotechnology and Bioengineering. 2025. [Google Scholar] [CrossRef]
- Lu, Z.; Evans, S.; McDonnell, L.; Bandari, N.C.; Weng, Y.; Jin, W.; et al. Exploiting the Geranylgeranyl-Pyrophosphate-Sensing N-Terminal Domain of HMG-CoA Reductase 2 to Regulate Farnesyl Pyrophosphate Synthase (Erg20p) for Improved Monoterpene Production in Saccharomyces cerevisiae. Yeast. 2025. [Google Scholar] [CrossRef]
- Gao, Q.; Wang, H.; Shan, M.; Wu, F.; Jiang, G.; Yao, M.; et al. Systematic Engineering To Enhance Citronellol Production in Yeast. Journal of Agricultural and Food Chemistry. 2025. [Google Scholar] [CrossRef]
- Ferrer-Carbonell, C.; Villa, R.; Viskaal, I.; Opperman, D.J.; Paul, C.E. Enzymatic Synthesis of Enantiopure (R)-Citronellal from Geraniol via a Short-Chain Dehydrogenase and Ene Reductase. Advanced Synthesis & Catalysis 2025, 2500060. [Google Scholar]
- Maróstica Jr, M.R.; Pastore, G.M. Biotransformation of citronellol in rose-oxide using cassava wastewater as a medium. Food Science and Technology. 2006, 26, 690–696. [Google Scholar] [CrossRef]
- Pimentel, M.; Molina, G.; Bertucci, T.; Pastore, G. Biotransformation of citronellol in rose oxide by Pseudomonas spp. Chemical Engineering Transactions. Chemical Engineering Transactions. 2012, 27, 295–300. [Google Scholar]
- Pinto, A.; Contente, M.L.; Tamborini, L. Advances on whole-cell biocatalysis in flow. Current Opinion in Green and Sustainable Chemistry. 2020, 25, 100343. [Google Scholar] [CrossRef]
- Ma, G.; Zhu, X.; Zhang, D.; Li, H.; Lin, J.; Wei, D. Design of a Self-Sufficient Whole-Cell Cascade for the Production of (R)-Citronellal from Geraniol. Journal of Agricultural and Food Chemistry. 2024, 72, 26305–26315. [Google Scholar] [CrossRef]
- Oda, S.; Sugai, T.; Ohta, H. Syntheses of Optically Active Citronellol, Citronellal, and Citronellic Acid by Microbial Oxidation and Double Coupling System in an Interface Bioreactor. Bulletin of the Chemical Society of Japan. 2000, 73, 2819–2823. [Google Scholar] [CrossRef]
- Demyttenaere, J.C.; Vanoverschelde, J.; De Kimpe, N. Biotransformation of (R)-(+)-and (S)-(−)-citronellol by Aspergillus sp. and Penicillium sp., and the use of solid-phase microextraction for screening. Journal of Chromatography A. 2004, 1027, 137–146. [Google Scholar] [CrossRef]
- Bhat, S.V.; Mestry, S.J.; Gupta, M.O. Biotechnological transformation of citronellene and citronellol by fungus Rhizopus oryzae (ATCC 9363) leading to interesting conversions through oxidations and rearrangements. 2022.
- Ohashi, Y.; Huang, S.; Maeda, I. Biosyntheses of geranic acid and citronellic acid from monoterpene alcohols by Saccharomyces cerevisiae. Bioscience, Biotechnology, and Biochemistry. 2021, 85, 1530–1535. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, P.; Singh, M.; Singh, U.; Jatav, S.; Sangwan, R.S.; Mishra, B.B. Iodosylbenzene (PhIO) mediated synthesis of rose oxide from β-citronellol and its application for in situ rose oxide enrichment led valorization of citronella essential oil. Journal of Cleaner Production. 2018, 172, 1765–1771. [Google Scholar] [CrossRef]
- Arifin, A.A.; Don, M.M.; Uzir, M.H. Baker's yeast mediated biotransformation of geraniol into citronellol using a continuous-closed-gas-loop bioreactor (CCGLB) system. Biochemical engineering journal. 2011, 56, 219–224. [Google Scholar] [CrossRef]
- Muñoz, R.; Daugulis, A.J.; Hernández, M.; Quijano, G. Recent advances in two-phase partitioning bioreactors for the treatment of volatile organic compounds. Biotechnology Advances. 2012, 30, 1707–1720. [Google Scholar] [CrossRef]
- Vinaixa, M.; Schymanski, E.L.; Neumann, S.; Navarro, M.; Salek, R.M.; Yanes, O. Mass spectral databases for LC/MS-and GC/MS-based metabolomics: State of the field and future prospects. TrAC Trends in Analytical Chemistry. 2016, 78, 23–35. [Google Scholar] [CrossRef]
- Chen, B.-G.; Chang, C.D.; Wang, C.-T.; Chen, Y.-J.; Chang, W.-T.; Wang, S.-M.; et al. A novel approach to evaluate the extent and the effect of cross-contribution to the intensity of ions designating the analyte and the internal standard in quantitative GC-MS analysis. Journal of the American Society for Mass Spectrometry. 2011, 19, 598–608. [Google Scholar] [CrossRef]
- Kaur, J.; Goswami, D.; Saraf, M. Response surface methodology: a comparative optimization of antifungal metabolite production by Trichoderma viride and Trichoderma harzianum using solid-state fermentation. Biomass Conversion and Biorefinery 2025, 1–24. [Google Scholar] [CrossRef]
- Stilo F, Franchina FA, Cordero C, Focant J-F, editors. Comprehensive two-dimensional gas chromatography coupled to mass spectrometry (GC× GC-TOF MS): discrimination of Italian extra virgin olive oils (EVOOs) from different regions and exploration of high resolution (HR) mass spectrometry information. 12th Multidimensional Chromatography Workshop-Guidebook; 2021.
- Scriba, G.K. Chiral recognition in separation sciences. Part I: Polysaccharide and cyclodextrin selectors. TrAC Trends in Analytical Chemistry. 2019, 120, 115639. [Google Scholar]
- Ikai, T.; Okamoto, Y. Structure control of polysaccharide derivatives for efficient separation of enantiomers by chromatography. Chemical Reviews. 2009, 109, 6077–6101. [Google Scholar] [CrossRef]
- Ilisz, I.; Berkecz, R.; Péter, A. Application of chiral derivatizing agents in the high-performance liquid chromatographic separation of amino acid enantiomers: a review. Journal of pharmaceutical and biomedical analysis. 2008, 47, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Wang, D.; Wu, Y.; Lin, Z.; Wang, L.; Wang, L.; et al. Analysis of monoterpenol Isomers in hops and beers: comparison of methods with a chiral column and a nonchiral column. Journal of the American Society of Brewing Chemists. 2017, 75, 149–155. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, W.; Sun, L.; Liang, A.; Duan, J.; Zhang, Y.; et al. Preparation of novel chiral stationary phases based on chiral metal-organic cages enable extensive HPLC enantioseparation. Analytica Chimica Acta. 2025, 1337, 343541. [Google Scholar] [CrossRef] [PubMed]
- Mookdasanit, J.; Tamura, H. Comparative enantioseparation of monoterpenes by HPLC on three kinds of chiral stationary phases with an on-line optical rotatory dispersion under reverse phase mode. Food Science and Technology Research. 2002, 8, 367–372. [Google Scholar] [CrossRef]
- de Brito, G.A.; Rocha de Oliveira, P.F.; de Andrade Silva, C.M.; de Araújo Neto, M.F.; Leite, F.H.A.; Mesquita, P.R.R.; et al. Identification of bioactive compounds against Aedes aegypti (Diptera: Culicidae) by bioassays and in silico assays. Chemistry & Biodiversity. 2021, 18, e2100242. [Google Scholar]
- Dakpa, G.; Chiang, Y.-T.; Lin, L.-Y.; Tsao, N.-W.; Wang, C.-H.; Pérez-Sánchez, H.; et al. Essential oil-derived compounds target core fatigue-related genes: A network pharmacology and molecular Docking approach. PLoS One. 2025, 20, e0314125. [Google Scholar] [CrossRef]
- Tang, Y.; Li, H.; Song, Q. Lemongrass essential oil and its major component citronellol: evaluation of larvicidal activity and acetylcholinesterase inhibition against Anopheles sinensis. Parasitology Research. 2024, 123, 315. [Google Scholar] [CrossRef]
- de Souza, B.o.z.z.i. A. Theoretical investigation of the Pictet-Spengler reaction between dopamine and (S)-citronellal catalyzed by the enzyme (S)-norcoclaurine synthase. 2021.
- Dendera, W. Generation of a virtual library of terpenes using graph theory, and its application in exploration of the mechanisms of terpene biosynthesis: RHODES UNIVERSITY, SOUTH AFRICA; 2018.
- Patsilinakos, A.; Artini, M.; Papa, R.; Sabatino, M.; Božović, M.; Garzoli, S.; et al. Machine learning analyses on data including essential oil chemical composition and in vitro experimental antibiofilm activities against Staphylococcus species. Molecules. 2019, 24, 890. [Google Scholar] [CrossRef]
- Sharma, A.D.; Kaur, I. Essential oil from Cymbopogon citratus exhibits “anti-aspergillosis” potential: in-silico molecular docking and in vitro studies. Bulletin of the National Research Centre. 2022, 46, 23. [Google Scholar] [CrossRef] [PubMed]
- Salaria, D.; Rolta, R.; Sharma, N.; Patel, C.N.; Ghosh, A.; Dev, K.; et al. In vitro and in silico antioxidant and anti-inflammatory potential of essential oil of Cymbopogon citratus (DC.) Stapf. of North-Western Himalaya. Journal of Biomolecular Structure and Dynamics. 2022, 40, 14131–14145. [Google Scholar] [CrossRef]
- Cardeal dos Santos, A.N.; Oliveira PEGd da Cruz Freire, J.E.; Araújo dos Santos, S.; Júnior, J.E.R.H.; Andrade CRd et, a.l. Computational Profiling of Monoterpenoid Phytochemicals: Insights for Medicinal Chemistry and Drug Design Strategies. International Journal of Molecular Sciences. 2025, 26, 7671. [Google Scholar] [CrossRef]
- Liao, L.; Xie, M.; Zheng, X.; Zhou, Z.; Deng, Z.; Gao, J. Molecular insights fast-tracked: AI in biosynthetic pathway research. Natural Product Reports. 2025. [Google Scholar] [CrossRef] [PubMed]
- Varghese, R.; Shringi, H.; Efferth, T.; Ramamoorthy, S. Artificial intelligence driven approaches in phytochemical research: trends and prospects. Phytochemistry Reviews 2025, 1–16. [Google Scholar] [CrossRef]
- da Silva, A.J.A.; de Sousa Silveira, Z.; Macêdo, N.S.; dos Santos Barbosa, C.R.; da Silva Sousa, Â.E.; da Silva, T.F.; etal., *!!! REPLACE !!!*. Evaluation of the antibacterial and inhibitory activity of the NorA efflux pump in Staphylococcus aureus by citronellol. Biologia 2025, 1–15. [Google Scholar] [CrossRef]
- Wei, T.; Regeard, C.; Barroca-Aubry, N.; Roger, P.; Aymes-Chodur, C. Chemoenzymatic oxidation of citronellol and geraniol: Synthesis and antibacterial activity assessment. Colloids and Surfaces B: Biointerfaces. 2025, 253, 114723. [Google Scholar] [CrossRef]
- Nazemiyeh, H.; Lotfipoor, F.; Delazar, A.; Razavi, S.M.; Asnaashari, S.; Kasebi, N.; et al. Chemical composition, and antibacterial and free-radical-scavenging activities of the essential oils of a citronellol producing new chemotype of Thymus pubescens Boiss. & Kotschy ex Celak. Records of Natural Products. 2011, 5, 184. [Google Scholar]
- Zhang, J.; Liu, H.; Yao, J.; Ma, C.; Yang, W.; Lei, Z.; et al. Plant-derived citronellol can significantly disrupt cell wall integrity maintenance of Colletotrichum camelliae. Pesticide Biochemistry and Physiology. 2024, 204, 106087. [Google Scholar] [CrossRef] [PubMed]
- Klis, F.M.; Sosinska, G.J.; De Groot, P.W.; Brul, S. Covalently linked cell wall proteins of Candida albicans and their role in fitness and virulence. FEMS yeast research. 2009, 9, 1013–1028. [Google Scholar] [CrossRef] [PubMed]
- Perrone, G.G.; Tan, S.-X.; Dawes, I.W. Reactive oxygen species and yeast apoptosis. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research. 2008, 1783, 1354–1368. [Google Scholar] [CrossRef] [PubMed]
- Cleary, I.; MacGregor, N.; Saville, S.; Thomas, D. Investigating the function of Ddr48p in Candida albicans. Eukaryotic Cell. 2012, 11, 718–724. [Google Scholar] [CrossRef]
- Liu, S.; Hou, Y.; Liu, W.; Lu, C.; Wang, W.; Sun, S. Components of the calcium-calcineurin signaling pathway in fungal cells and their potential as antifungal targets. Eukaryotic cell. 2015, 14, 324–334. [Google Scholar] [CrossRef]
- Widiyarti G, Handayani S, Hanafi M, editors. Synthesis and cytotoxic activity of citronellol esters. AIP Conference Proceedings; 2018: AIP Publishing LLC.
- Mamur, S. Investigation of Cytotoxic Effect of Monoterpenes Beta-Citronellol and (-)-Menthone in Human Breast Cancer (MCF-7) Cell Line. Adnan Menderes Üniversitesi Sağlık Bilimleri Fakültesi Dergisi. 2019, 3, 111–119. [Google Scholar]
- Yu, W.-N.; Lai, Y.-J.; Ma, J.-W.; Ho, C.-T.; Hung, S.-W.; Chen, Y.-H.; et al. Citronellol induces necroptosis of human lung cancer cells via TNF-α pathway and reactive oxygen species accumulation. in vivo. 2019, 33, 1193–1201. [Google Scholar] [CrossRef]
- Brito, R.G.; Santos, P.L.; Prado, D.S.; Santana, M.T.; Araújo, A.A.; Bonjardim, L.R.; et al. Citronellol reduces orofacial nociceptive behaviour in mice–evidence of involvement of retrosplenial cortex and periaqueductal grey areas. Basic & clinical pharmacology & toxicology. 2013, 112, 215–221. [Google Scholar]
- Brito, R.G.; Guimarães, A.G.; Quintans, J.S.; Santos, M.R.; De Sousa, D.P.; Badaue-Passos Jr, D.; et al. Citronellol, a monoterpene alcohol, reduces nociceptive and inflammatory activities in rodents. Journal of natural medicines. 2012, 66, 637–644. [Google Scholar] [CrossRef]
- Srinivasan, S.; Muruganathan, U. Antidiabetic efficacy of citronellol, a citrus monoterpene by ameliorating the hepatic key enzymes of carbohydrate metabolism in streptozotocin-induced diabetic rats. Chemico-biological interactions. 2016, 250, 38–46. [Google Scholar] [CrossRef]
- Müller, G.C.; Junnila, A.; Butler, J.; Kravchenko, V.D.; Revay, E.E.; Weiss, R.W.; et al. Efficacy of the botanical repellents geraniol, linalool, and citronella against mosquitoes. Journal of Vector Ecology. 2009, 34, 2–8. [Google Scholar] [CrossRef]
- Revay, E.E.; Junnila, A.; Xue, R.-D.; Kline, D.L.; Bernier, U.R.; Kravchenko, V.D.; et al. Evaluation of commercial products for personal protection against mosquitoes. Acta tropica. 2013, 125, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Awaluddin, N.; Awaluddin, S.W.; Bachri, N.; Mointi, S.S. The Formulation of Reed Diffuser is A Combination of Cinnamon (Cinnamomomum Verum) and Citronella (Cymbopogon Nardus) Essential Oil as An Anti-Stress Aromatheraphy. Jurnal Penelitian Pendidikan IPA. 2023, 9, 1960–1967. [Google Scholar] [CrossRef]
- Ratnesakera, D.; Nayanthara, K. Efficacy of cinnamon and citronella oil vapours in the control of Callosobruchus chinensis L. in bulk stored green gram. Journal of Food and Agriculture 2013, 3. [Google Scholar] [CrossRef]
- Soares-Castro, P.; Soares, F.; Santos, P.M. Current advances in the bacterial toolbox for the biotechnological production of monoterpene-based aroma compounds. Molecules. 2020, 26, 91. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, J.; Zavala, A.; Diaz-Perez, C.; Cervantes, C.; Diaz-Perez, A.; Campos-Garcia, J. The atu and liu clusters are involved in the catabolic pathways for acyclic monoterpenes and leucine in Pseudomonas aeruginosa. Applied and environmental microbiology. 2006, 72, 2070–2079. [Google Scholar] [CrossRef]
- Jang, W.D.; Kim, G.B.; Kim, Y.; Lee, S.Y. Applications of artificial intelligence to enzyme and pathway design for metabolic engineering. Current Opinion in Biotechnology. 2022, 73, 101–107. [Google Scholar] [CrossRef]
- Vindman, C.; Trump, B.; Cummings, C.; Smith, M.; Titus, A.J.; Oye, K.; et al. The convergence of AI and synthetic biology: the looming deluge. arXiv preprint 2024, arXiv:240418973. [Google Scholar]
- Bolmanis, E.; Galvanauskas, V.; Grigs, O.; Vanags, J.; Kazaks, A. Leveraging Historical Process Data for Recombinant P. pastoris Fermentation Hybrid Deep Modeling and Model Predictive Control Development. Fermentation. 2025, 11, 411. [Google Scholar]
- Hjort, C.L.; Heum, H.E.B. Fermentation Prediction Through Machine Learning and Its Potential Use in Production Planning and Control: NTNU; 2023.
- Yee, C.S.; Zahia-Azizan, N.A.; Abd Rahim, M.H.; Mohd Zaini, N.A.; Raja-Razali, R.B.; Ushidee-Radzi, M.A.; et al. Smart Fermentation Technologies: Microbial Process Control in Traditional Fermented Foods. Fermentation. 2025, 11, 323. [Google Scholar] [CrossRef]
- Cai, S.; Mao, Z.; Wang, Z.; Yin, M.; Karniadakis, G.E. Physics-informed neural networks (PINNs) for fluid mechanics: A review. Acta Mechanica Sinica. 2021, 37, 1727–1738. [Google Scholar] [CrossRef]
Figure 1.
General process of synthesizing citronellol in different odorant species. Key abbreviations: ADH, alcohol dehydrogenase; PhCIR, P. graveolens citral reductases; PgOPR, P. graveolens12-oxophytodienoic acid reductase; GER, geranial reductase; TPS, terpene synthases; GES, geraniol synthase; PgNUDX, P. graveolens Nudix hydrolase enzyme; GP Reductase, geraniol monophosphate reductase; RhNUDX, Rosa × hybrida Nudix hydrolase enzyme; RhGeDH, Rosa × hybrida geraniol dehydrogenase; RhGER, Rosa × hybrida geraniol Reductase; RhOPR, Rosa × hybrida 12-oxophytodienoic acid reductase; RhCAR, Rosa × hybrida citronellal reductases.
Figure 1.
General process of synthesizing citronellol in different odorant species. Key abbreviations: ADH, alcohol dehydrogenase; PhCIR, P. graveolens citral reductases; PgOPR, P. graveolens12-oxophytodienoic acid reductase; GER, geranial reductase; TPS, terpene synthases; GES, geraniol synthase; PgNUDX, P. graveolens Nudix hydrolase enzyme; GP Reductase, geraniol monophosphate reductase; RhNUDX, Rosa × hybrida Nudix hydrolase enzyme; RhGeDH, Rosa × hybrida geraniol dehydrogenase; RhGER, Rosa × hybrida geraniol Reductase; RhOPR, Rosa × hybrida 12-oxophytodienoic acid reductase; RhCAR, Rosa × hybrida citronellal reductases.
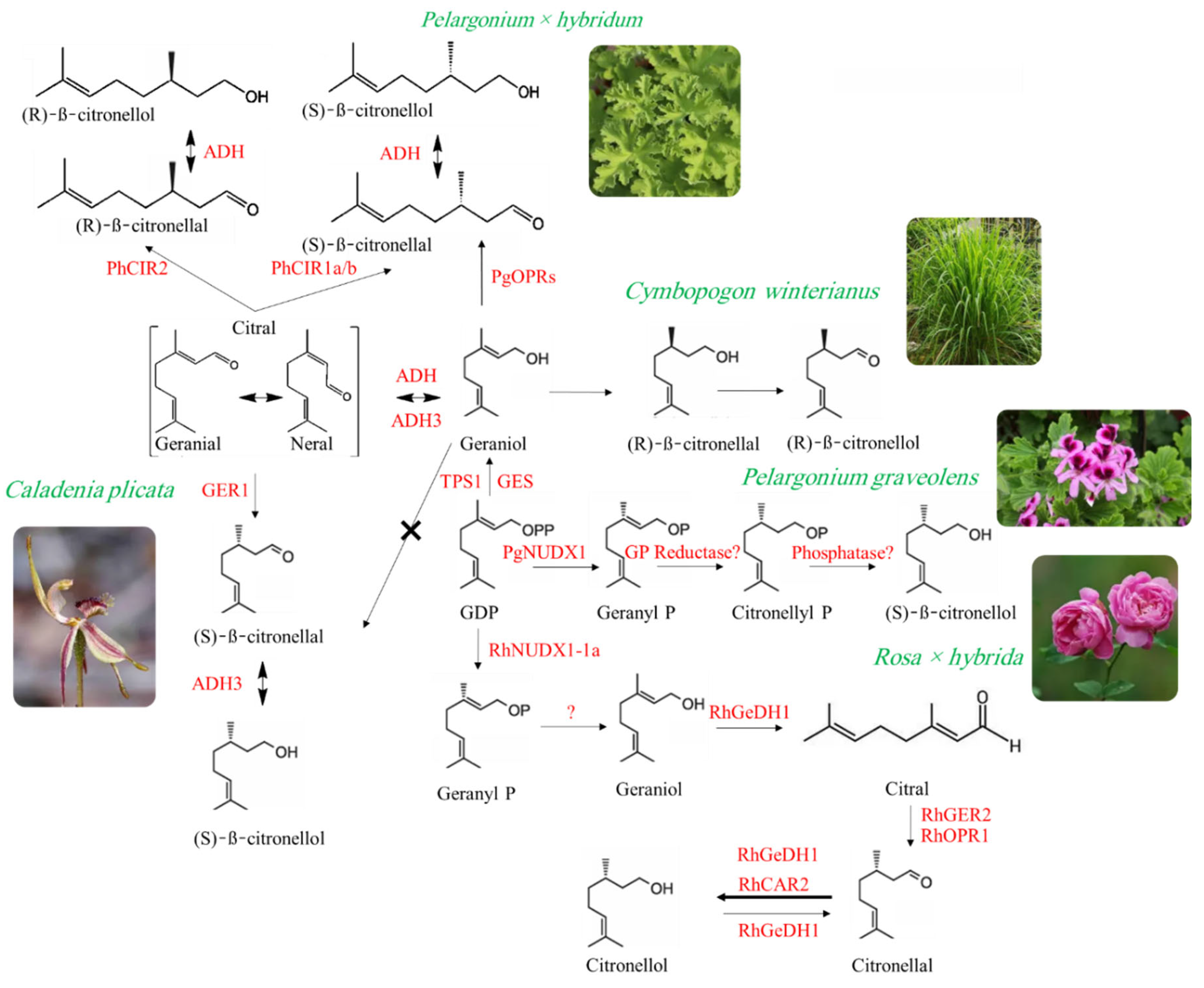
Figure 2.
Schematic Representation of the Citronellol Biosynthetic Pathway and Associated Engineering Strategies. (a) Depiction of the engineered citronellol biosynthetic pathway in S. cerevisiae [9]. Enzymes are color-coded: red for native (endogenous) enzymes, pink for mutated endogenous enzymes, and green for heterologous (non-native) enzymes. Thick arrows indicate enhanced pathway flux, while crossed arrows denote gene deletions. Key abbreviations: HMG-CoA – 3-hydroxy-3-methylglutaryl coenzyme A; IPP – Isopentenyl pyrophosphate; DMAPP – Dimethylallyl pyrophosphate; GPP – Geranyl diphosphate; tHmgr – Truncated HMG-CoA reductase; Idi1 – Isopentenyl diphosphate isomerase; GES – Geraniol synthase; Atf1 – Alcohol acetyltransferase; Erg20mut – Mutated farnesyl diphosphate synthase, including Erg20F96W and Erg20F96W,N127W variants used in synthetic protein scaffolds; (b) Enzymes incorporated from external sources as described in reference [69]: AtoB – Acetoacetyl-CoA synthase (E. coli); HMGS – HMG-CoA synthase (S. cerevisiae); HMGR – HMG-CoA reductase (S. cerevisiae); MK – Mevalonate kinase; PMK – Phosphomevalonate kinase; PMD – Phosphomevalonate decarboxylase (S. cerevisiae); IDI – Isopentenyl diphosphate isomerase (E. coli); GPPS – Geranyl pyrophosphate synthase (Abies grandis); GES – Geraniol synthase (Ocimum basilicum); GLDA – Glycerol dehydrogenase (E. coli); AHR – NADPH-dependent aldehyde reductase (E. coli); CrIS – Iridoid synthase (C. roseus); (c) Enzymes utilized in the pathway described in reference [71]: ERG8 – Phosphomevalonate kinase; ERG20ww – Modified farnesyl diphosphate synthase; tCrGES – Truncated C. roseus geraniol synthase.
Figure 2.
Schematic Representation of the Citronellol Biosynthetic Pathway and Associated Engineering Strategies. (a) Depiction of the engineered citronellol biosynthetic pathway in S. cerevisiae [9]. Enzymes are color-coded: red for native (endogenous) enzymes, pink for mutated endogenous enzymes, and green for heterologous (non-native) enzymes. Thick arrows indicate enhanced pathway flux, while crossed arrows denote gene deletions. Key abbreviations: HMG-CoA – 3-hydroxy-3-methylglutaryl coenzyme A; IPP – Isopentenyl pyrophosphate; DMAPP – Dimethylallyl pyrophosphate; GPP – Geranyl diphosphate; tHmgr – Truncated HMG-CoA reductase; Idi1 – Isopentenyl diphosphate isomerase; GES – Geraniol synthase; Atf1 – Alcohol acetyltransferase; Erg20mut – Mutated farnesyl diphosphate synthase, including Erg20F96W and Erg20F96W,N127W variants used in synthetic protein scaffolds; (b) Enzymes incorporated from external sources as described in reference [69]: AtoB – Acetoacetyl-CoA synthase (E. coli); HMGS – HMG-CoA synthase (S. cerevisiae); HMGR – HMG-CoA reductase (S. cerevisiae); MK – Mevalonate kinase; PMK – Phosphomevalonate kinase; PMD – Phosphomevalonate decarboxylase (S. cerevisiae); IDI – Isopentenyl diphosphate isomerase (E. coli); GPPS – Geranyl pyrophosphate synthase (Abies grandis); GES – Geraniol synthase (Ocimum basilicum); GLDA – Glycerol dehydrogenase (E. coli); AHR – NADPH-dependent aldehyde reductase (E. coli); CrIS – Iridoid synthase (C. roseus); (c) Enzymes utilized in the pathway described in reference [71]: ERG8 – Phosphomevalonate kinase; ERG20ww – Modified farnesyl diphosphate synthase; tCrGES – Truncated C. roseus geraniol synthase.
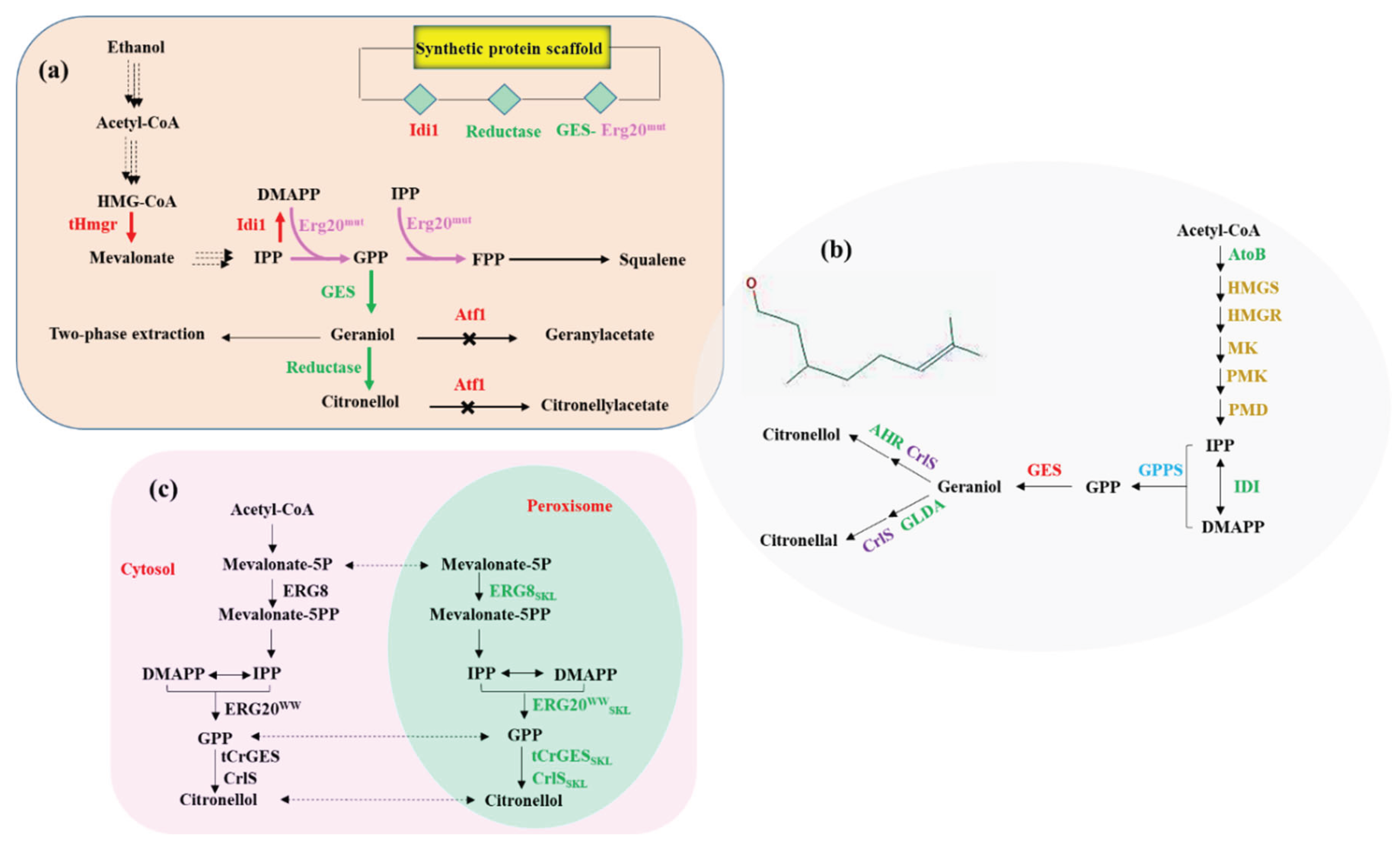
Figure 3.
A multi-layered infographic mapping the full pharmacological universe of citronellol.
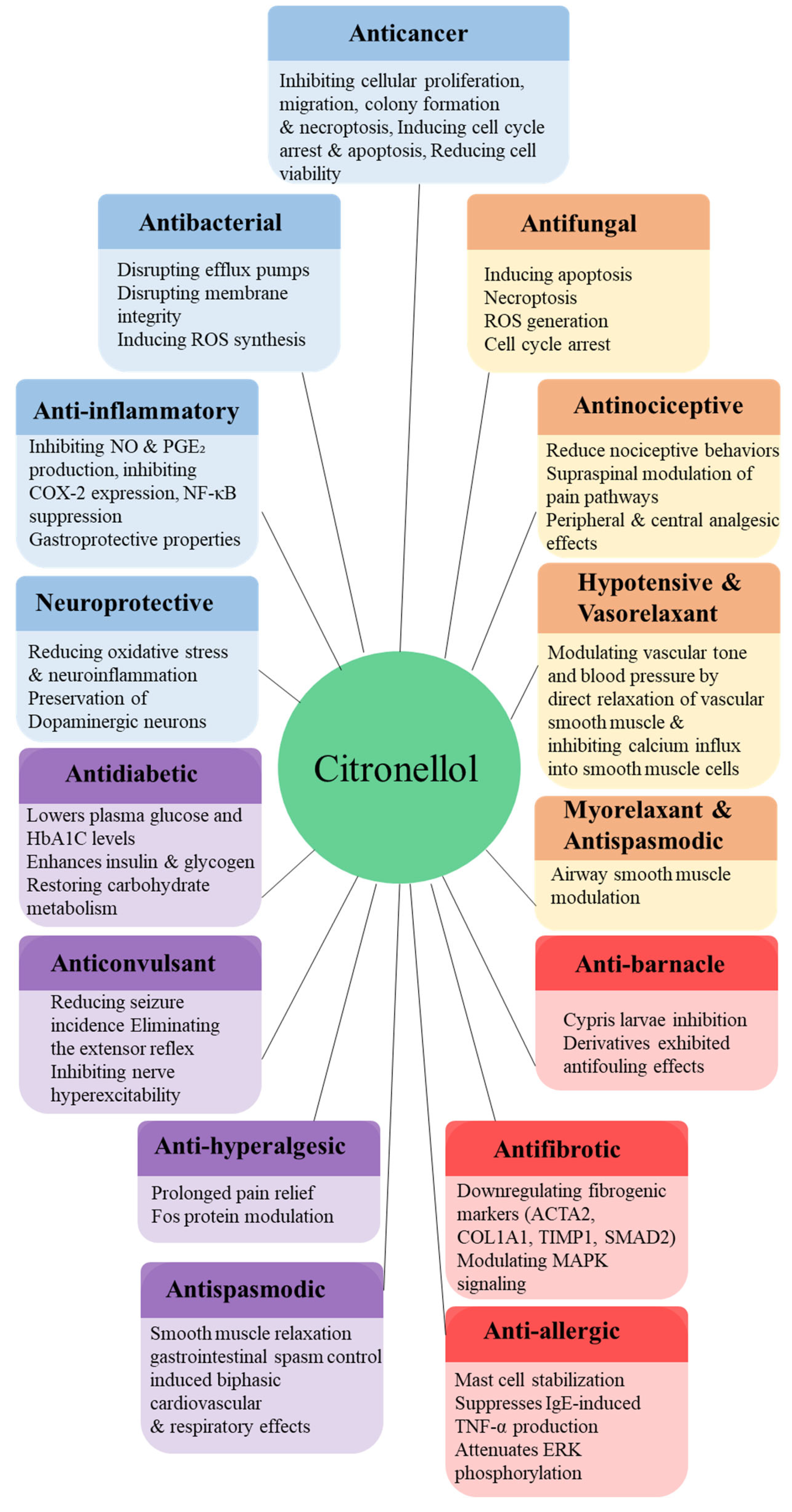
Figure 4.
Flowchart summarizing the mechanisms of action of β-citronellol against C. albicans. Abbreviations: Als2p, Agglutinin-like sequence protein 2; Rbt1p, Repressed by Tup1 protein 1, Pga4p, Putative glycosylphosphatidylinositol-anchored protein 4; Sod1p, Superoxide dismutase 1 protein; ROS, Reactive oxygen species; Ssa2p, Stress-seventy subfamily A member 2; Hsp30p, Heat shock protein 30; Sti1p, Stress-inducible protein 1; Gst2p, Glutathione S-transferase 2; ATP, Adenosine triphosphate; Cap1p, Candida AP-1-like transcription factor 1; Ca²⁺, Calcium ion; ER, Endoplasmic reticulum; Cmd1p, Calmodulin 1; Atp3p, ATP synthase subunit gamma; Atp7p, ATP synthase subunit epsilon; Cox1p, Cytochrome c oxidase subunit 1; Cobp, Cytochrome b protein.
Figure 4.
Flowchart summarizing the mechanisms of action of β-citronellol against C. albicans. Abbreviations: Als2p, Agglutinin-like sequence protein 2; Rbt1p, Repressed by Tup1 protein 1, Pga4p, Putative glycosylphosphatidylinositol-anchored protein 4; Sod1p, Superoxide dismutase 1 protein; ROS, Reactive oxygen species; Ssa2p, Stress-seventy subfamily A member 2; Hsp30p, Heat shock protein 30; Sti1p, Stress-inducible protein 1; Gst2p, Glutathione S-transferase 2; ATP, Adenosine triphosphate; Cap1p, Candida AP-1-like transcription factor 1; Ca²⁺, Calcium ion; ER, Endoplasmic reticulum; Cmd1p, Calmodulin 1; Atp3p, ATP synthase subunit gamma; Atp7p, ATP synthase subunit epsilon; Cox1p, Cytochrome c oxidase subunit 1; Cobp, Cytochrome b protein.
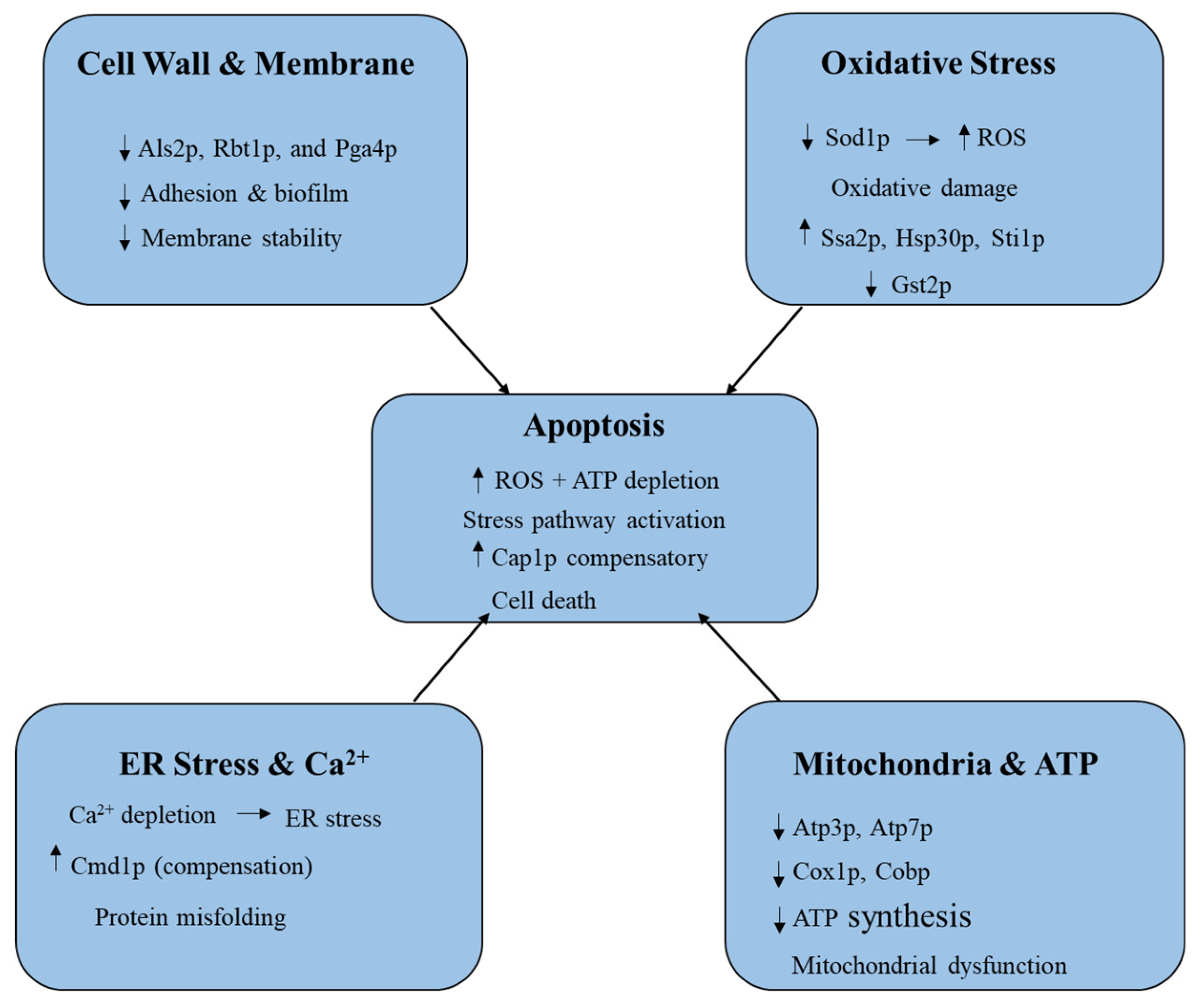
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



