Submitted:
28 August 2025
Posted:
29 August 2025
You are already at the latest version
Abstract
This study evaluated the yield stability and environmental responsiveness of 42 pro-vitamin A cassava genotypes across multi-season trials using Finlay-Wilkinson regression and trait-based clustering approaches. Regression parameters-intercept and slope were used to quantify baseline yield potential and sensitivity to environmental variation, respectively. Hierarchical and k-means clustering grouped genotypes into three biologically distinct clusters with clear agronomic relevance. Cluster 2 genotypes exhibited moderate responsiveness and positive yield baselines, indicating broad adaptability and suitability for regional deployment. Cluster 3 showed high environmental sensitivity but low yield potential, suggesting limited resilience under marginal conditions. Cluster 1 comprised highly responsive genotypes with poor baseline productivity, reflecting unstable performance and strong genotype × environment interaction. One-way ANOVA confirmed significant differences among clusters for both slope (F(2,39) = 40.89, P < 0.001) and intercept (F(2,39) = 102.10, P < 0.001), validating the clustering structure. Dendrogram profiling reinforced these classifications, offering a quantitative framework for genotype prioritization. The findings support strategic breeding decisions tailored to agroecological contexts and highlight the need for multi-trait integration in future clustering models to enhance cultivar deployment precision.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Study Location and Plant Materials
2.2. Experimental Design and Field Layout
2.3. Trait Measurement
2.4. Statistical Analysis and Model Specification
2.5. Genotype Stability Analysis via Finlay–Wilkinson Regression
3. Results
3.1. High-Yielding Cassava Genotypes and Their Environmental Responsiveness
3.2. Cassava Genotypes Exhibiting Extreme Yield Sensitivity to Environmental Conditions
3.3. Stable and Predictable Cassava Genotypes with Near-Unit Environmental Response Slopes
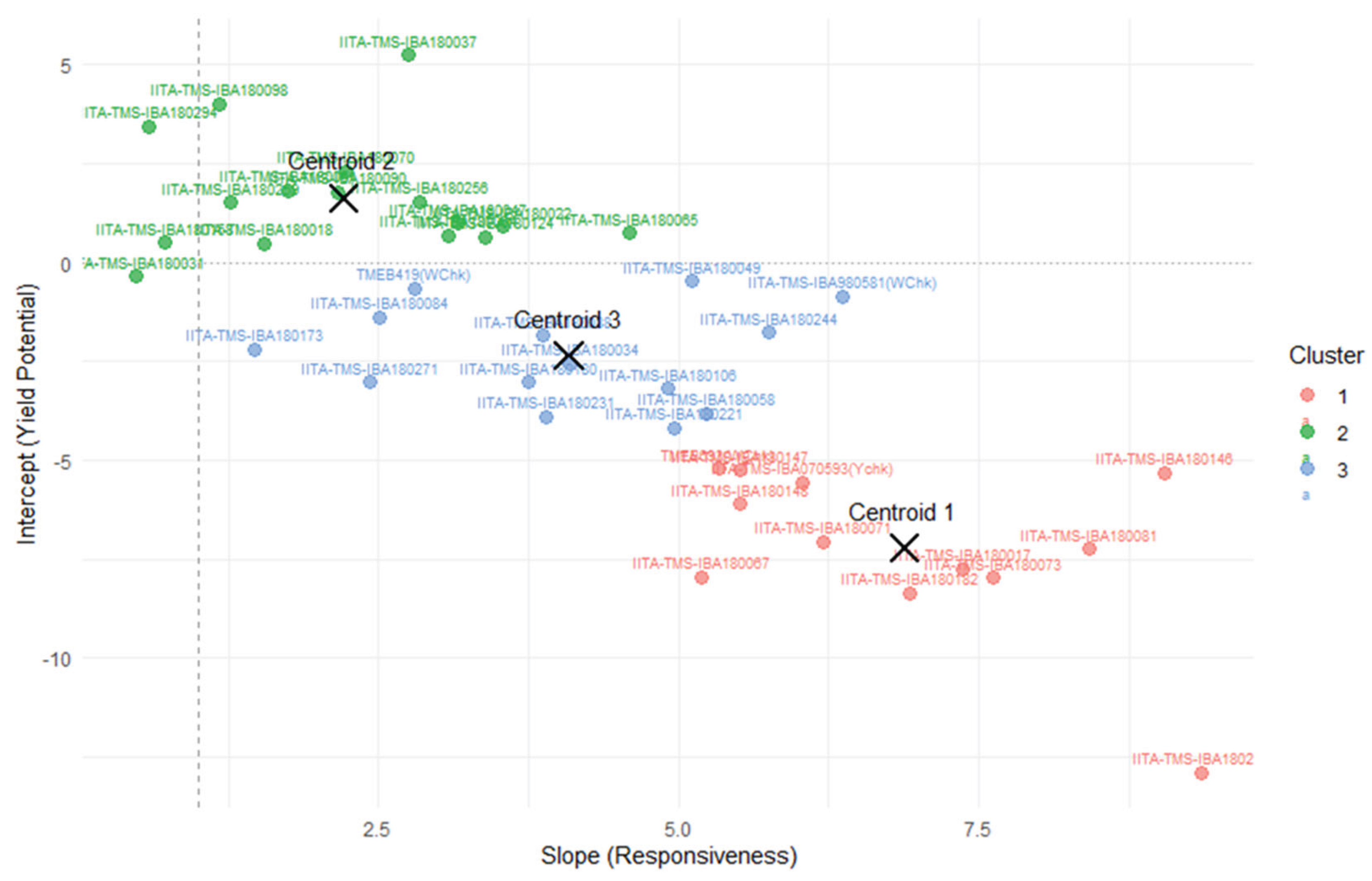
3.4. Cluster-Defined Performance Profiles from Finlay–Wilkinson Regression Traits
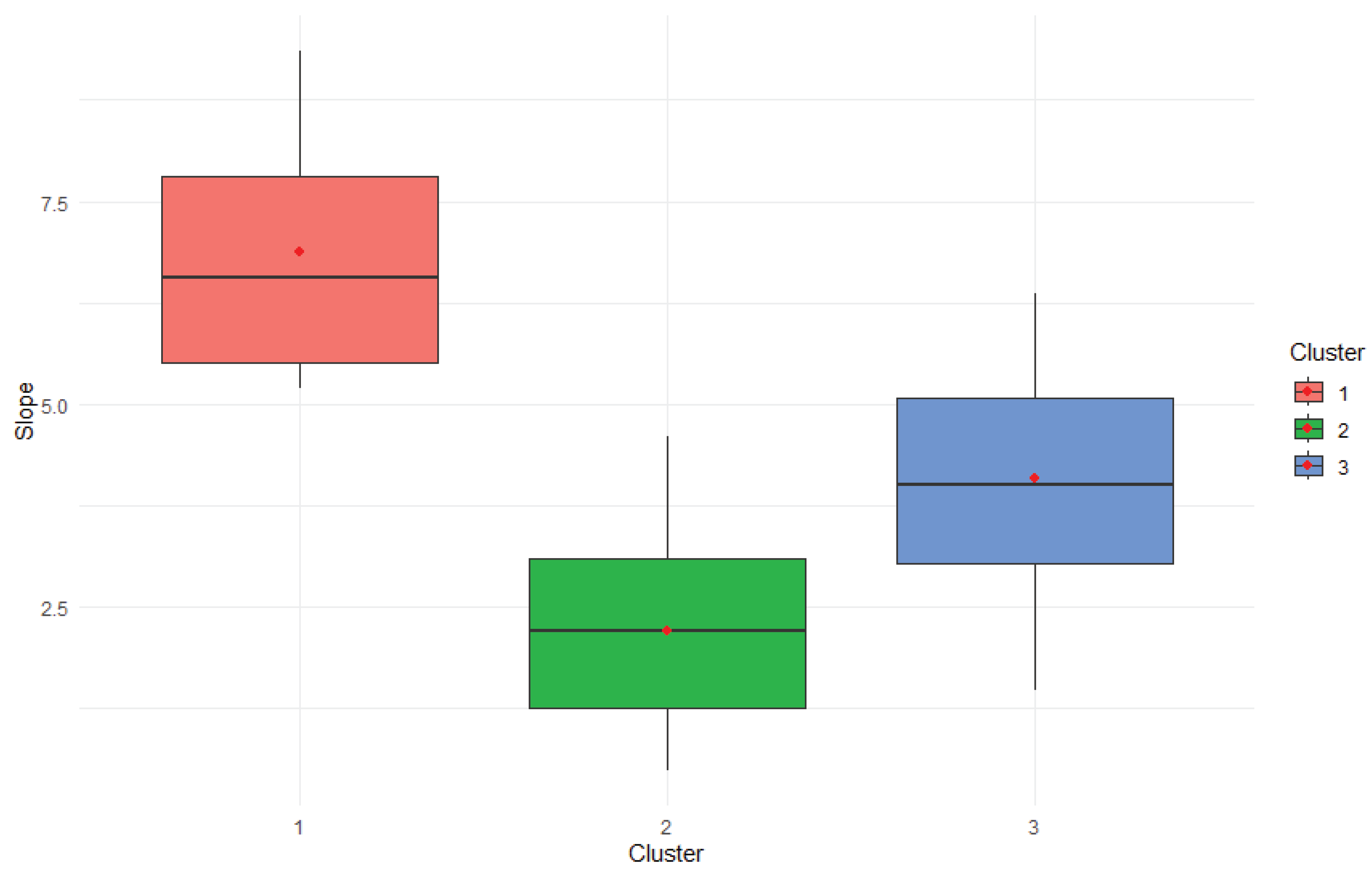
3.5. Visual Summary of Environmental Sensitivity Across Clusters
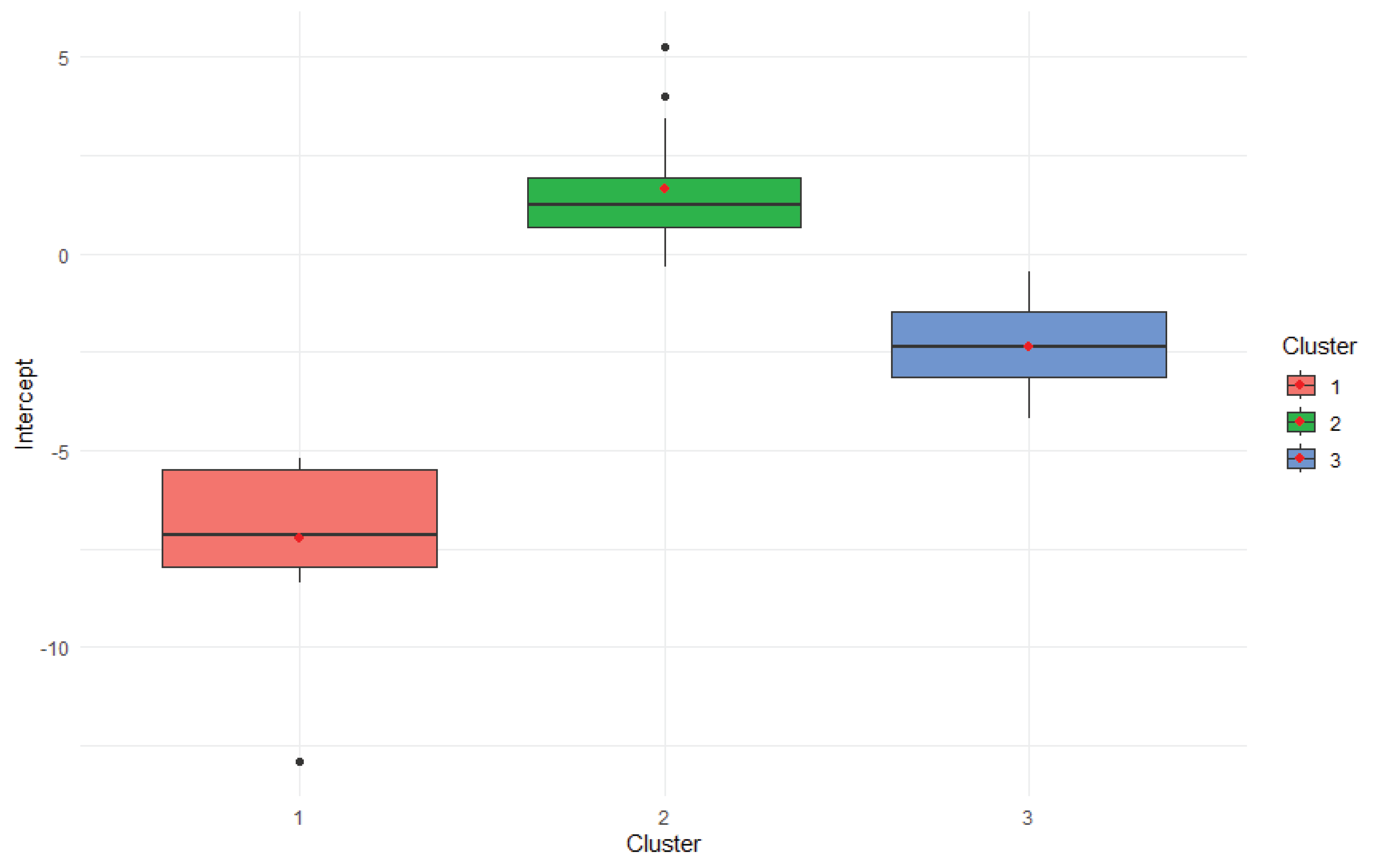
3.6. Baseline Performance Variation Across Clusters
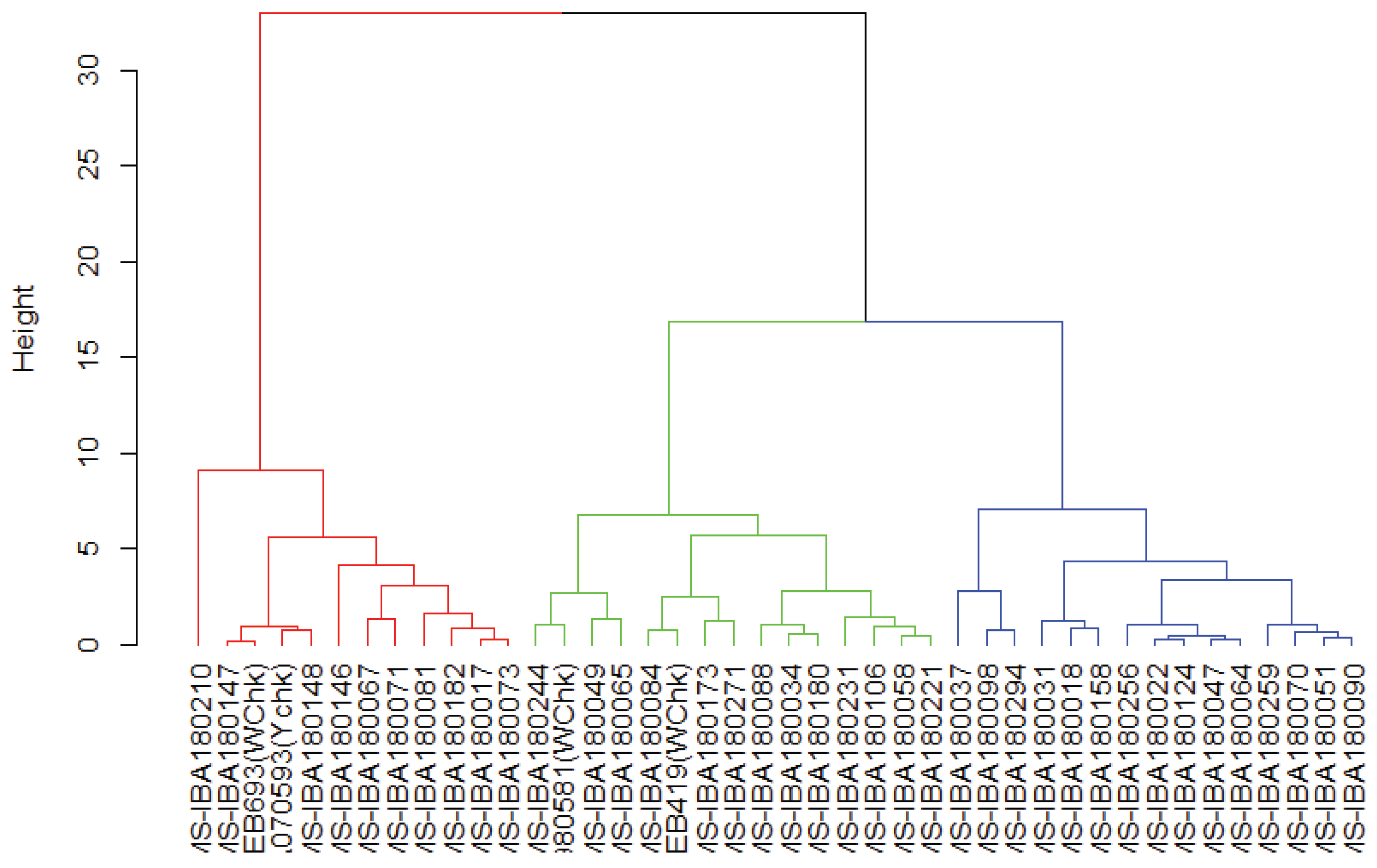
3.7. Genetic Clustering Reveals Population Structure Among Cassava Genotypes
3.8. Yield-Responsiveness Analysis Highlights Genotypic Adaptability Across Environments
4. Discussion
4.1. Genotype Performance and Stability Based on Finlay–Wilkinson Regression
4.2. Trait-Based Hierarchical Clustering of Genotypic Performance
4.3. Trait-Based Hierarchical Clustering of Genotypic Performance
4.4. Cluster-Based Variation in Yield Stability and Environmental Sensitivity
4.4.1. ANOVA-Based Validation of Cluster Differentiation
4.4.2. Centroid Analysis and Agronomic Implications
4.4.3. Convergent Performance Among Genetically Diverse Cassava Genotypes
5. Recommendation
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CMD | Cassava Mosaic Disease |
References
- Otekunrin, O. A. (2024). Cassava (Manihot esculenta Crantz): A global scientific footprint—production, trade, and bibliometric insights. Discover Agriculture, 2, Article 94. [CrossRef]
- National Root Crops Research Institute. (2024, February 27). The resilience and versatility of cassava: A staple root crop in Africa. Available online:https://nrcri.gov.ng/the-resilience-and-versatility-of-cassava-a-staple-root-crop-in-africa/ (accessed on Day15th August, 2025.
- Omoluabi, J. E., & Ibitoye, S. J. (2024). Cassava production and agricultural growth in Nigeria: Analysis of effects and forecast. GSC Advanced Research and Reviews, 21(1), 37–46. [CrossRef]
- Karim, K. Y., & Norman, P. E. (2021). Genotype × environment interaction and stability analysis for selected agronomic traits in cassava (Manihot esculenta). International Journal of Environment, Agriculture and Biotechnology, 6(8), 1–12. Retrieved from https://ijoear.com/assets/articles_menuscripts/file/IJOEAR-AUG-2021-4.pdf.
- Begna, T. (2020). The role of genotype by environmental interaction in plant breeding. International Journal of Agriculture and Biosciences, 9(5), 209–215. Retrieved from https://www.ijagbio.com/pdf-files/volume-9-no-5-2020/209-215.pdf.
- Bates, D., Mächler, M., Bolker, B., & Walker, S. (2015). Fitting Linear Mixed-Effects Models Using lme4. Journal of Statistical Software, 67(1), 1–48. [CrossRef]
- MacQueen, J. B. (1967). Some methods for classification and analysis of multivariate observations. In L. M. Le Cam & J. Neyman (Eds.), Proceedings of the Fifth Berkeley Symposium on Mathematical Statistics and Probability (Vol. 1, pp. 281–297). University of California Press.
- Ward, J. H. (1963). Hierarchical grouping to optimize an objective function. Journal of the American Statistical Association, 58(301), 236–244. [CrossRef]
- Sokal, R. R., & Sneath, P. H. A. (1963). Principles of numerical taxonomy. W.H. Freeman.
- Hair, J. F., Black, W. C., Babin, B. J., & Anderson, R. E. (2010). Multivariate data analysis (7th ed.). Pearson.
- Piepho, H. P., Möhring, J., Melchinger, A. E., & Büchse, A. (2008). Blup for phenotypic selection in plant breeding and variety testing. Euphytica, 161, 209–228. [CrossRef]
- R Core Team. (2020). R: A language and environment for statistical computing (Version 4.0.3) [Computer software]. R Foundation for Statistical Computing. https://www.r-project.org/.
- Finlay, K. W., & Wilkinson, G. N. (1963). The analysis of adaptation in a plant-breeding programme. Australian Journal of Agricultural Research, 14(6), 742–754.
- Wickham, H., François, R., Henry, L., Müller, K., Vaughan, D., & Posit Software, PBC. (2023). dplyr: A grammar of data manipulation (Version 1.1.4) [R package]. Comprehensive R Archive Network (CRAN). https://cran.r-project.org/package=dplyr.
- Wickham, H., Chang, W., Henry, L., Pedersen, T. L., Takahashi, K., Wilke, C., Woo, K., Yutani, H., Dunnington, D., van den Brand, T., & Posit, PBC. (2025). ggplot2: Create elegant data visualisations using the grammar of graphics (Version 3.5.2) [R package]. Comprehensive R Archive Network (CRAN). https://cran.r-project.org/package=ggplot2.
- Eberhart, S. A., & Russell, W. A. (1966). Stability parameters for comparing varieties. Crop Science, 6(1), 36–40. [CrossRef]
- Carvalho, H. F., Rio, S., García-Abadillo, J., & Sánchez, J. I. (2024). Revisiting superiority and stability metrics of cultivar performances using genomic data: derivations of new estimators. Plant Methods, 20, Article 85. [CrossRef]
- Egea-Gilabert, C., Pagnotta, M. A., & Tripodi, P. (2021). Genotype × environment interactions in crop breeding. Agronomy, 11(8), 1644. [CrossRef]
- Jarquin, D., Howard, R., Crossa, J., Beyene, Y., Gowda, M., Martini, J. W. R., ... & Prasanna, B. M. (2020). Genomic prediction enhanced sparse testing for multi-environment trials. G3: Genes, Genomes, Genetics, 10(9), 3023–3033. [CrossRef]
- Bančič, J., Ovenden, B., Gorjanc, G. et al. Genomic selection for genotype performance and stability using information on multiple traits and multiple environments. Theor Appl Genet 136, 104 (2023). [CrossRef]
- van Eeuwijk, F. A., Bustos-Korts, D. V., & Malosetti, M. (2010). Two-mode clustering of genotype by trait and genotype by environment data. Euphytica, 175(3), 365–379. [CrossRef]
- Ebem, E. C., Afuape, S. O., Chukwu, S. C., & Ubi, B. E. (2021). Genotype × Environment Interaction and Stability Analysis for Root Yield in Sweet Potato (Ipomoea batatas [L.] Lam). Frontiers in Agronomy, 3, Article 665564. [CrossRef]
- Dalmaijer, E. S., Nord, C. L., & Astle, D. E. (2022). Statistical power for cluster analysis. BMC Bioinformatics, 23, Article 205. [CrossRef]
- Strickert, M., Sreenivasulu, N., Villmann, T., & Hammer, B. (2008). Robust centroid-based clustering using derivatives of Pearson correlation. In Proceedings of the First International Conference on Bio-inspired Systems and Signal Processing (BIOSTEC). ScitePress. https://www.scitepress.org/Papers/2008/10626/10626.pdf.
- Sabaghnia, N., Shekari, F., Nouraein, M., & Janmohammadi, M. (2025). Cluster analysis of agronomic traits in chickpea genotypes under cool upland semi-arid region. Annals of Arid Zone, 64(1), 13–21. https://epubs.icar.org.in/index.php/AAZ/article/download/159002/59598.
- Zakir, M. (2018). Review on genotype × environment interaction in plant breeding and agronomic stability of crops. Journal of Biology, Agriculture and Healthcare, 8(12), 1–9. https://www.iiste.org/Journals/index.php/JBAH/article/view/43065.
- Lian, L., & de los Campos, G. (2016). FW: An R package for Finlay–Wilkinson regression that incorporates genomic/pedigree information and covariance structures between environments. G3: Genes|Genomes|Genetics, 6(3), 589–597. [CrossRef]
- Pour-Aboughadareh, A., Jadidi, O., Jamshidi, B., Bocianowski, J., & Niemann, J. (2025). Cross-talk between stability parameters and selection models: A new procedure for improving the identification of superior genotypes in multi-environment trials. BMC Research Notes, 18, Article 306. https://bmcresnotes.biomedcentral.com/articles/10.1186/s13104-025-07366-1.

| Genotype | Intercept | Slope | Insight |
|---|---|---|---|
| IITA-TMS-IBA180037 | 5.25 | 2.75 | High yield, responsive |
| IITA-TMS-IBA180294 | 3.44 | 0.588 | High yield, low sensitivity |
| IITA-TMS-IBA180070 | 2.29 | 2.23 | Good yield, moderately responsive |
| IITA-TMS-IBA180256 | 1.52 | 2.84 | Stable but responsive |
| IITA-TMS-IBA180259 | 1.51 | 1.27 | Balanced and modest performer |
| Genotype | Slope | Interpretation |
|---|---|---|
| IITA-TMS-IBA180081 | 8.42 | Highly sensitive, erratic under stress |
| IITA-TMS-IBA180073 | 7.61 | Responsive but potentially unstable |
| IITA-TMS-IBA180017 | 7.37 | Very sensitive, may lack reliability |
| IITA-TMS-IBA180182 | 6.93 | Possible strong G×E interaction |
| IITA-TMS-IBA180146 | 9.04 | Extreme response, risky adaptability |
| Genotype | Intercept | Slope | Stability |
|---|---|---|---|
| IITA-TMS-IBA180018 | 0.47 | 1.54 | Stable, slight responsiveness |
| IITA-TMS-IBA180051 | 1.81 | 1.74 | Responsive with modest yield |
| IITA-TMS-IBA180098 | 4.02 | 1.17 | Slightly sensitive, high yield |
| IITA-TMS-IBA180259 | 1.51 | 1.27 | Balanced performance |
| Cluster | Representative Genotypes | Mean Intercept (Yield) | Mean Slope (Sensitivity) | Trait Profile | Agronomic Interpretation |
| 1 | IITA-TMS-IBA180146, IITA-TMS-IBA180081 | -7.23 | 6.87 | Extremely sensitive, low yield | Unstable genotypes; high G×E interaction |
| 2 | IITA-TMS-IBA180073, IITA-TMS-IBA180256, IITA-TMS-IBA180244 | 1.64 | 2.20 | Moderate responsiveness, positive yield | Balanced and adaptable performers |
| 3 | IITA-TMS-IBA180037, IITA-TMS-IBA180049 | -2.35 | 4.08 | High sensitivity with low baseline yield | Inconsistent performance under stress |
| Trait | Source | Df | Sum Sq | Mean Sq | F-value | P-value | Significance |
|---|---|---|---|---|---|---|---|
| Slope | Cluster | 2 | 149.83 | 74.91 | 40.89 | <0.001 | *** |
| Residuals | 39 | 71.44 | 1.83 | ||||
| Intercept | Cluster | 2 | 539.10 | 269.57 | 102.10 | <0.001 | *** |
| Residuals | 39 | 103.00 | 2.64 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



