
Submitted:
27 August 2025
Posted:
28 August 2025
You are already at the latest version
Abstract
Human activities have driven habitat degradation and biodiversity loss in marine ecosystems, highlighting the need for strategies that reconcile food production with ecosystem restoration. Restorative aquaculture has emerged as a potential tool to enhance habitat complexity, nutrient cycling, and biodiversity. Here, we combined a systematic literature review with an experimental study in NW Spain to evaluate the ecological effects of mussel aquaculture on macroinvertebrates. The review highlighted a global imbalance between mussel production and research on its ecological benefits, with most studies emphasizing environmental impacts rather than positive ecosystem effects. Mussel farms enhance habitat complexity and provide trophic subsidies through mussel fall-off, supporting higher abundances of crustaceans and echinoderms, including commercial species such as lobsters. Ecological effects vary with spatial scale, production type (inshore vs. offshore), substrate characteristics, and farm structure. The review highlights both ecological benefits and potential risks, including ecological traps. Research gaps include the need for quantitative assessments and long-term monitoring. The experimental study quantified O. nigra abundance revealed higher densities near mussel rafts. Mussel farms act as both trophic and structural facilitators, demonstrating how low trophic aquaculture can synergistically support marine biodiversity and ecosystem resilience. Properly managed, mussel farming could transition from a food production activity to a regenerative ecological tool.
Keywords:
restorative aquaculture
; mussel production
; epibenthic fauna
; longline system
; Mytilus sp.
1. Introduction
Over recent decades, human activities and climate change have led to significant loss of habitat and biodiversity across the world’s oceans [1]. Habitat destruction, overexploitation, and pollution are the primary stressors driving these changes, emphasising the need to balance human development with ocean health. In this context, restorative aquaculture has emerged as a strategy to restore the structure and function of degraded, damaged, or destroyed habitats by leveraging the ecological benefits of cultivated aquatic species [2]. Defined as "commercial aquaculture with ecological benefits" [3] restorative aquaculture offers a pathway to provide ecosystem services such as habitat formation, nutrient cycling, and biodiversity enhancement [2,4].
In 2022, global aquaculture production surpassed capture fisheries for the first time, with 31% of marine animal harvests sourced from aquaculture. Specifically, 10.2% of this production was attributed to sea mussels [5]. This aquaculture is typically established in soft-bottom areas characterized by low abundance and biodiversity, primarily represented by infaunal communities [6]. Extensive research on the environmental impacts of bivalve aquaculture has predominantly focused on benthic organic enrichment and its associated biological and physicochemical effects [7,8,9] In addition, bivalve aquaculture has been shown to increase habitat complexity on the seabed beneath farms compared to nearby control areas [10], because mussel fall-off in farming areas enhancing shelters and suitable habitats for macrofauna [11]. Moreover, the organic enrichment associated with mussel farming fosters trophic resource availability, from detritivorous to higher trophic levels. In this way, mussel aquaculture provides new benthic habitats and additional trophic subsidy that may benefits epibenthic fauna, enhancing secondary production in the ecosystem[10,11,12,13,14].
Given its role as both a new food source and a habitat engineer, mussel aquaculture has significant restoration potential for impacted soft-bottom areas, supporting the abundance and diversity of commercial valuable species, such as crabs and lobsters, which have often been overexploited. In fact, one of the main principles of the restorative aquaculture is the development in an area where the habitat availability has declined or limited due to human stressors [15].
This restoration potential can be clearly illustrated in Galicia, on the Northwest coast of Spain (NW Iberian Peninsula), one of the world’s leading mussel-producing regions [5]. Here, production in carried out using raft systems, installed within the sheltered rías. These structures may function simultaneously as a source of food and as habitat-forming elements, fostering complex benthic communities. In such environments, echinoderms are notably abundant. In the Ría of Vigo, suspension-feeding species such as Ophiocomia nigra (Müller, 1789) can form dense aggregations beneath production structures, likely attracted by the enrichment generated from pseudofaeces deposition [16].
Therefore, the present study takes adopts two complementary approaches -a literature review and an experimental study- to gain deeper insight into the ecological effects of bivalve aquaculture, with a particular focus on its influence on macroinvertebrate populations. First, we reviewed published studies to evaluate the potential of these systems to act as restorative practices and to enhance the ecological services of soft-bottom habitats associated with aquaculture. Second, we conducted a case study in the Ría de Vigo, assessing whether mussel production influences the distribution of macrofauna by quantifying the abundance of Ophiocomina nigra beneath raft structures.
2. Materials and Methods
2.1. Literature Review
A comprehensive literature search was conducted following the PRISMA methodology, using Google Scholar and Scopus. The initial search included broad keywords such as ‘mussel’, ‘suspended bivalve aquaculture’, ‘environmental impact’, ‘macroinvertebrates’, ‘aquaculture’, ‘restoration’, ‘restorative aquaculture’. To refine and expand the scope, a more specific literature search was performed using additional keywords, including ‘spatial variation’, ‘top-down control’, ‘acoustic telemetry’, ‘offshore mussel farm’, ‘artificial reef’, ‘ecosystem services’, ‘marine protected area’, ‘habitat value’, ‘lobster’, ‘crabs’ and ‘echinoidea’.
Relevant literature was stored and classified using the Zotero reference manager. This review focused exclusively on studies examining the attraction of epibenthic fauna to suspended mussel aquaculture, primarily in English-language publications. Given the scarcity of studies on this topic, no restrictions were applied regarding publication date or geographic regions.
While some studies provide qualitative descriptions of species’ trophic interactions and reproductive success, quantitative data on abundance and biomass remain limited, precluding a meta-analysis. The few available telemetry studies, mostly on lobster and crab home range, offer a valuable insight into species’ movement and habitat use around the farms but are insufficient for robust statistical assessments. Therefore, this review adopts a qualitative and descriptive approach, synthesising existing knowledge while identifying critical gaps for future research.
To visualize the global distribution of scientific effort, a map was generated using the ggplot2 R package [17]. For clarity, only those countries with high mussel production and scientific effort were included. Furthermore, data on global Mytilidae production by country was obtained from FAO database for the year 2022, the most recent year for which data is available. A barplot depicting the main producer countries was also created using the ggplot2 R package [17].
2.2. Experimental Study
2.2.1. Sampling Approach
The experimental study focused on randomly selected rafts systems in the Ria de Vigo, one of the four Rias Baixas of the NW Iberian Peninsula (Figure 1). The northern boundary of the NW African upwelling system is located here; winds engage with the coastal topography to create upwelling and downwelling dynamics on the continental shelf. The upwelling of nutrients from the cold deep seawater, which is transported into de bay, is essential for phytoplankton production. This nutrient-rich environment creates a high primary production area, which supports extensive suspended mussel farming (Mytilus galloprovincialis) in the Rias [18]. Production is based on mussel floating rafts 500 m2 made of wooden laths, with an average of 500 hanging ropes 12m long each.
A total of 24 rafts were initially sampled, with 12 sites at difference distances from the aquaculture structure: 4 ‘Near’, 4 ‘Middle’ and 4 ‘Far’. However, due to the visibility constraints and image quality only 9 rafts were ultimately retained for analysis.
To quantify Ophiocomia nigra abundance, images were obtained using a quadrat structure (1m2) with four arms and a support to fix the GoPro HERO+3 and an artificial light. The structure was lowered to the bottom with a rope and retrieved after 10 seconds of video recording. Sites were separated 10 m, starting near the structure and extending along transects of up to 120 m. High-quality frames from of each video were extracted to quantify the abundance of O. nigra (ind/m2).
2.2.2. Data Analysis
To test whether abundance of O. nigra differed with distance from the raft structures, we fitted a Generalized Linear Mixed Model (GLMM) using the R package glmmTMB [19]. Abundance was specified as the response variable, using Distance as fixed factor with three levels: near (0-40m), middle (41-80m) and far (80-120m). To account for the hierarchical sampling, Raft identity was included as random intercept, thereby allowing for variation in mean abundance among rafts. A negative binomial distribution was selected to model the response, as the family appropriately handles overdispersion commonly observed in count data.
3. Results
3.1. Production Rates and Scientific Effort
Globally, mollusc aquaculture is dominated by mussel production (Mytilidae), with China accounting for 40% of global output, followed by Canada, Chile, New Zealand and Spain [5]. However, the geographical distribution of scientific research on its ecological benefits does not match the production rates (Figure 2A). Most studies have focused on environmental impacts and potential drawbacks of this activity, while research on its ecological benefits remain limited.
Specifically, only 18 studies were identified that assess the effects of suspended mussel aquaculture on epibenthic macrofauna. Interestingly, China and Chile, the leading global producers, have no publications addressing this topic. In contrast, Canada and New Zealand while not leading in production volume, are at the top of research effort in this field (Figure 2B).
The earliest study dates 1982, but interest remained sparse in the following decades. Since 2020, however, there has been a notable increase in attention, with nine of the seventeen studies published within the last years. This reflects the growing awareness on ecosystem restoration, particularly in the context of offshore expansion and the rising use of acoustic telemetry for behavioural and spatial variation studies.
3.2. Ecological Benefits
Non-fed mussel aquaculture provides critical ecosystem services, particularly through water filtration. Mussels act as a link between benthic and pelagic systems by filtering phytoplankton, removing organic and inorganic particles, and enhancing sediment denitrification. This process forms nitrogen-enriched pellets, which contribute to sediment fertility [20]. Furthermore, ammonia excretion by mussels stimulates phytoplankton proliferation and accelerates the remineralization of organic matter through sediment deposition [21,22]. This dynamic establishes a feedback loop, where mussels both consume and promote phytoplankton production.
However, the ecological benefits extended beyond nutrient loading and eutrophication mitigation. Mussel fall-off enriches benthic habitat with organic material, supporting trophic interactions and acting as an ecosystem engineer [13,14,23,24,25,26]. This process enriches the substrate and provides additional habitat that supports both macrobenthic and pelagic communities. These changes create synergies with commercial fisheries and aids in the restoration of marine ecosystems [11,27,28,29]. Beyond their ecological contributions, mussel aquaculture also has socio-economic benefits and has been recognised by the Marine Conservation Society as a “best choice” protein source [30], highlighting its potential a sustainable sector to address nutritional demands while simultaneously delivering Ecosystem Services [31].
3.3. Effects on Epibenthic Macroinvertebrates
The predominant macroinvertebrates associated with mussel farms include Crustacea, Asteroidea, and Gastropoda, benefiting from increased habitat complexity and food availability [6,13,24,32,33,34]. Key species include sea stars (Asterias sp.), lobsters (Homarus sp.) and rock crabs (Cancer sp.). Nevertheless, the interaction between these species and the aquaculture structures may not follow the same patters and depends on the spatial scale and the trophic position [11].
Drouin and colleagues [11] observed that rock crabs and sea stars are attracted by mussel fall-off and, therefore, by trophic interactions. This attraction arises partly from the infauna and epifauna associated with the ropes and structures provided by the farms, as well as from the live mussel fall-off. Mussels are considered the main prey of rock crabs (Cancer sp.) in coastal areas [35] and have shown an abundance up to six times higher within the farm area [11,12,13,26,34,36,37,38]. Studies on their distribution suggest that they do not interact heavily with the farm structure or the associated habitat (8,9). Nevertheless, dietary studies of C. pagurus revealed a significant increase in the proportion of mussels in their diets within the farms [39], although there was an overlap between individuals captures inside and outside the mussel farm [26]. Furthermore, Lees and colleagues [37] identified that C. irroratus (currently C. plebejus Poeppig, 1836) tended to remain within mussel farm and exhibited slower movement, potentially indicating foraging on fallen mussels and detritus beneath the cages.

Table 1.
Main macroinvertebrate groups and species attracted to suspended mussel aquaculture (longline and raft systems), their ecological role and implications for aquaculture and ecosystem functioning.
Table 1.
Main macroinvertebrate groups and species attracted to suspended mussel aquaculture (longline and raft systems), their ecological role and implications for aquaculture and ecosystem functioning.
| References | Ecological role/Implication | Response to mussel farm | Group/Specie |
|---|---|---|---|
| [12,37,39] | Predator of mussels; important prey of lobsters; enhances trophic links | Attraction to mussel fall-off; abundance up to 6 times within farm; diet shift toward mussel | Cancer, Pagurus, |
| C. irroratus | |||
| (rock crabs) | |||
| [40,41] | High value commercial species; uses anchor blocks and fall-off as habitat; diverse diet by higher prey availability | Mixed results: limited association in sandy offshore farms; sometimes higher abundances near structures; inter-stage competition | Homarus americanus (American lobster) |
| [26] | Key commercial species; Higher shelter fidelity and then behavioural shifts | Strong association with offshore farm structures; sheltering in anchor blocks and mussel-off | Homarus gammarus |
| (European lobster) | |||
| [13,42,43] | Keystone predator; causes economic losses in aquaculture; higher reproductive success and rapid growth | Major predator of mussels; classified as “pests” by producers | Asterias spp. |
| (sea star) | |||
| [24,44] | Predator; reproductive success boosted by mussel deposits | Abundance 25x higher within farms; enhanced growth and gonad production; interspecies competition | Coscinasterias muricata |
| (brittle star) | |||
| [16] | Detritivores; recycling organic matter | Attracted to organic deposits beneath farms | Holothuroidea (sea cucumber) |
| [16] | Grazers; influence benthic community composition | Recruitment on culture structures; individuals often fall to seabed | Echinoidea |
| (sea urchin) | |||
| [45] | Suspension feeder; increases local biodiversity | Dense beds under longlines due to pseudofaeces enrichment | Ophiocomia nigra (brittle star) |
Echinoderms are also significantly higher within mussel farms compared to control areas, with notable increases in sea stars, sea cucumbers and sea urchins’ abundances [43]. Sea cucumbers are primarily attracted to the organic deposits accumulating on the seabed beneath the farms, while sea urchins often fall from the culture structures, where recruitment success appears to be higher [3]. For many sea star species, mussels are a preferred prey under natural conditions [43], showing abundances 25 times higher within farms for Coscinasterias muricata [24,44]. Particularly, the genus Asterias, play a pivotal role as keystone predators, exerting top-down control over prey populations and significantly shaping ecosystem dynamics. This predatory behaviour together with the fact that frequently emerge as the most abundant predators around the farms [13], leading aquaculture producers and fisheries managers to classify them as “pests” due to the associated economic losses because mussel mortality increase [36,46].
The high predation rates by sea stars highlight their ability to exploit mussel farms as abundant food sources. Several mechanisms may drive sea stars aggregations, including: (i) active movement toward chemical cues released by mussels and other preys [24], (ii) direct asexual recruitment in the high-density prey patches [24,27,47], (iii) reduced foraging effort due to the availability of easily accessible food [24], and (iv) high recruitment rates to mussel lines [44,48]. This substantial food supply significantly increases the chance of successful reproduction and promotes rapid growth, enhanced gonad production, and prolonged proximity to conspecifics [24]. Moreover, zones with abundant food sources may limit fission due to the rapid growth, leading to a biological state in which asexual division appears to be suppressed [24].
However, this reproductive advantage also intensifies interspecies competition for resources. In Australia, studies on competition between native C. muricata and the invasive Asterias amurensis reveal interesting dynamics. Despite the invasive species shows better body and reproductive conditions within mussel farms, no significant increase in abundance within these areas has been recorded. Behavioural patterns, including predator recognition and avoidance by the invasive species, highlight the competitive interactions in food-rich habitat [44]. In contrast, in regions where Asterias sp. is native, these species benefit greatly from food subsidies provided by mussel farms, particularly in areas where the sea stars is the major predator [13,45,49].
For describe how mussel farms influence lobster’s diet (Homarus americanus), Sardenne and colleagues [41] proposed three potential trophic-pathways; (i) mussel subsidies do not enter to the lobster’s food web, with lobsters interacting only with the farm structure; (ii) lobsters benefit indirectly by consuming other organisms that feed on mussel fall-off; and (iii) lobsters directly consumed fallen mussels, enhancing their energy intake and potential growth. The results showed a larger trophic niche for organisms found within the mussel farms, which pointed to a more diverse diet maybe influenced by the greater prey availability. The proportion of mussels in the diet of adult lobsters increases from 15% in control sites to 46% in farm areas. Nevertheless, these patterns were not observed in smaller lobsters, potentially due to inter-stage competition or the mineral requirements associated with moulting requirement [50]. Smaller lobsters primarily consumed rock crabs, which comprised 99% of their diet, providing essential calcium. Although smaller lobsters can consume fallen mussels, they may avoid the farm to minimize competition with larger individuals, especially in areas with a significantly higher abundance of adults such as farm areas [41]. Moreover, rock crabs are important prey for lobsters of all sizes [51] with a key role in the growth, condition and ovary development of this specie [52]. Therefore, increase in rock crab abundance could lead to a corresponding rise in lobster populations under mussel farms, and the ecological impacts of the mussel farms scale-up the food web.
Despite trophic effects being the main driver of attraction, the provision of habitat by anchor blocks and the mussels fall-off contributes to habitat complexity and heterogeneity, enhancing habitat availability in soft-bottom areas and creating suitable shelter areas [24,25,42]. For epibenthic species, habitat selection- and therefore the substrate type- is another key factor influencing distribution, especially for lobsters. The European lobster is primarily influenced by the substrate type, whereas rock crab is also affected by substrate rugosity [17]. One possible explanation for this difference is that rock crabs may bury themselves in mud or sand, showing low fidelity to shelters, while lobsters appear to be highly dependent on physical structures, especially during the critical growth stages and during the moulting periods [11,53].
Additionally, lobsters exhibit significant shelter fidelity, with individuals occupying the same shelter for extended periods [54,55]. Similarly, American lobsters have shown behavioural shifts in response to substrate type, adopting a more nomadic behaviour in sandy benthic environment. Moreover, data also showed that lobsters were not as strongly associated with the offshore farm as reported in other studies, suggesting that, despite the availability of artificial structures, substrate type may be a determining factor in habitat selection [40].
The key drivers influencing macroinvertebrate species distribution also affect their spatial patterns within the mussel farming areas [11], a framework of three spatial-scale variations to explain these distributional trends. (i) Bay-scale variation: based on the hypothesis that macrofauna abundance is generally higher within mussel farms compared to surrounding areas; (ii) Farm-scale variation: linked to the farm’s production cycle, where zones with larger mussels support greater abundances due to the enhanced trophic interactions; and (iii) Line-scale variation, which considers the structural complexity of the production lines themselves, highlighting their role in habitat provision. Interestingly, results showed that lobsters, rock crabs and common sea stars were more abundant within farm areas. However, rock crabs and sea stars were particularly associated with areas where older mussels were cultivated, related to the increased food availability. In contrast, lobsters were most frequently observed directly beneath the lines, using anchor blocks as a shelter habitat.
Species interaction with aquaculture structures can also vary significantly between inshore and offshore mussel farms due to the differences in environmental conditions and characteristics. While both systems provide similar habitats, their impacts on the bottom may differ. Inshore aquaculture is typically located in protected embayment with softer and muddier substrates, whereas offshore operate in exposed, deeper waters with coarser sandy substrates [40]. Offshore systems feature wider-spaced structures and stronger hydrodynamic conditions, which result in reduced benthic impact compared to inshore sites [6,56]. Another key difference is the distance between the mussel socks and the seabed, which varies depending on the culture method and the depth condition. In inshore farms, mussel socks are typically suspended 1 to 3 meters above the bottom, whereas in offshore farms, they hang at approximately 10 meters above the seabed [40,57]. These structural differences influence species interactions and habitat use. Offshore systems also benefit from good oxygen conditions, reduced risk of biotoxins, diseases, parasites and terrestrial contamination [25]. These advantages, along with greater compatibility with marine spatial planning, are driving the global growth of offshore aquaculture [58].
Overall, studies on offshore aquaculture sites have shown that epibenthic macrofauna response positively to this activity and could be developed as a potential restorative aquaculture [12,14,25,26,40]. However, in Canada, research has indicated that American lobster exhibit limited association with offshore longline mussel aquaculture, possibly due to difference in farm structure and depth, as well as the predominance of sandy substrates, which provide a less suitable habitat [40]. However, other predator species, such as rock crabs (C. irroratus) and sea stars, were significantly more abundant in farm sites compared to reference areas. Interestingly, some studies also reported relatively high abundance of lobsters in farmed areas [12]. In contrast, studies conducted in the United Kingdom have shown that European lobsters display a stronger association with offshore farm structures, utilizing anchor blocks and bivalve fall-off as habitat [26].
Beyond their role as aquaculture sites, mussel farms may function as marine protected areas [59,60,61], contributing to marine conservation goals. The exclusion of fishing activities within these farms’ fosters biodiversity, provides refugee for species and creates synergies between aquaculture and commercial fisheries [25]. Nevertheless, they can also function as ecological traps. The aggregation of commercially valuable species around farms may increase vulnerability to fishing pressure, as target species concentrate within or near the farm site [62,63].
3.4. Experimental Study
The mean estimated abundance (± standard deviation) of O. nigra in the study area was 36.63 ± 62.64 ind/m2. The abundance of ophiuroids decreased with increasing distance from the aquaculture facility, from 56.36 ± 84.35 ind/m2 near of mussel farms to 24.11 ± 40.98 ind/m2 and 26.42 ± 50.36 ind/m2 at medium and far distances, respectively (Figure 3). Although the differences were not statistically significant, there was a near-significant trend towards lower abundances at medium distances compared to near positions (p = 0.099). Far distances showed a similar tendency, but the effect was not significant (p = 0.374). Substantial variability among rafts was detected (random intercept SD = 4.3), highlighting the importance of accounting for the hierarchical sampling design. In practical terms, abundance at middle distance was approximately 54% lower than at near positions, while at far distances it was reduced by 52% (Table 2; Figure 3).
4. Discussion
The extensive review on the effects of mussel farming on macrobenthic communities indicates that, although there is considerable descriptive information on the capacity of mussel beds to attract marine invertebrates, empirical data quantifying the extent of these attractions remains scarce [64]. Consequently, the potential of mussel farms as a form of restorative aquaculture remains unquantified.
Overall, studies suggest that epibenthic macrofauna response positively to suspended mussel aquaculture, with crustaceans and echinoderms being among the most commonly attracted taxa [11,13,14,23,24,25,26,37,40,57,65,66].
Our case of study suggests that abundance of Ophiocomia nigra tended to decrease with increasing distance from mussel farms (Figure 3). Although statistical support was limited due to the high heterogeneity, they tend observed at middle and at far distances indicates that proximity to raft ropes may play a role in structuring local epibenthic communities. These findings are consistent with previous observations that mussel aquaculture structures create localized habitat heterogeneity, concentrating epibenthic fauna near facilities. This result support the hypothesis that mussel farming can act as a driver of small-scale spatial variation in epibenthic assemblages, which may have implications for local biodiversity and ecosystem functioning.
The primary mechanisms driving these attractions are trophic subsidies, as mussel fall-off provide a food source that attracts crabs and echinoderms [24], while lobsters are drawn both to mussels and crabs [11,14,26,41]. Additionally, mussel farms contribute to habitat provision in soft-bottom areas, creating suitable shelter, while the increased food availability enhances reproductive success, further supporting population growth [24,25,42]. However, the extent and magnitude of this attraction differ between areas and production types.
4.1. Balancing Ecological and Socio-Economic Benefits
A key consideration for the development of its restorative potential is balancing its ecological benefits with socio-economic contributions. Restorative aquaculture should increase ecosystem productivity and provide ecosystem services while simultaneously contributing to food security and provide resilience to local economies. Mizuta and colleagues [67] define restorative aquaculture as “Commercial or subsistence aquaculture that supports initiatives to provide/or directly provides ecological benefits to the environment, leading to improved environmental sustainability and ecosystem services, in addition to the supply of seafood or other commercial products and opportunities for livelihood”. In the case of the mussel production, its restorative potential is linked to the increased abundance, growth, and reproductive success of commercially valuable species. By integrating aquaculture with marine conservation and spatial planning strategies, the industry can foster ecological recovery while providing economic resilience for coastal communities. However, achieving this balance requires a strategic approach that addresses multiple challenges, including water quality, climate change and sustainable food production [68,69].
Although the overall effect of mussel aquaculture on epibenthic macrofauna appears to be positive, some studies report no clear attraction effects, particularly in areas where the substrate is not suitable, mainly for lobster and crab species [65]. One potential management strategy to address these habitat limitations is the addition of artificial reefs around the production areas, similar to those used in Marine Protected Areas [70]. These structures could not only enhance productivity by providing shelter for species such as lobster, but also increase habitat heterogeneity and three-dimensional complexity, fostering more diverse benthic community. By introducing hard substrates into predominantly soft-bottom environments, artificial reefs could facilitate the colonization of sessile invertebrates, enhance trophic interactions, and create more complex habitats that support a wider range of species [71].
The size, diversity, and density of organisms inhabiting artificial reefs and their surrounding areas are influenced by the availability and variety of ecological niches, as well as the structural complexity of the reef itself [70,72]. A greater range of niches sizes benefits juvenile organisms, providing critical refuge and foraging opportunities. In this context, the introduction of hard substrates due to the accumulation of mussel shell may function as nursery grounds for mobile epifauna, thereby improving the carrying capacity of the ecosystem [72]. Consequently, the addition of physical structures from the aquaculture activities, along with introduction of additional trophic resoruces, could shifts aquaculture zones into potential biomass spillover areas.
4.2. Challenges and Spatial Planning Considerations
One of the main challenges in the development of the aquaculture sector is the spatial competition with other marine activities. To address that, the EU Directive on Marine Spatial Planning has proposed relocating human activities offshore to reduce environmental impact and mitigate conflicts over marine space [58]. However, expansion of offshore aquaculture requires clear regulatory frameworks and multi-sector agreements to ensure compatibility with existing marine activities.
The potential of mussel aquaculture as a restoration tool may be particularly relevant in offshore environments that have been impacted by human activities such as overfishing and habitat degradation due to extraction activities [25]. With appropriate monitoring and management, offshore aquaculture could offer an innovative strategy for marine habitat restoration [73]. In this context, a critical yet underexplored aspect is the quantification of the global impact of bivalve aquaculture on ecosystem service provision [44,74]. Addressing this knowledge gap requires long-term ecological monitoring, standardised methodologies, and the integration of ecosystem services into marine policy and spatial planning. These steps are essential for optimizing suitable aquaculture practices and ensuring their contribution to marine conservation and food-security.
While offshore aquaculture offers suitable protein production, its interactions with other offshore human activities such as offshore wind farms, remain uncertain [75]. Recent studies suggest that these artificial habitats may enhance macrofauna colonization, increase biodiversity, and support resident populations of commercially important crustacean species [76]. However, further long-term studies are necessary to evaluate the cumulative impacts and synergistic effects among offshore activities, facilitating the development of Environmental Impact Assessment and Marine Spatial Planing. This underscores the urgent need for further research to optimise offshore aquaculture development [25].
Changes in food production methods could significantly reduce resources requirements, making production more efficient and mitigating key drivers of climate change and biodiversity loss [77]. In this context, the development of restorative aquaculture represents a unique opportunity to integrate suitable food production with marine ecosystem restoration.
In that way, the implementation of the Ecosystem Approach to Aquaculture [78] can facilitate the progress toward a more dynamic and integrated understanding of the environmental interactions of aquaculture by providing a framework that balances ecological, socioeconomic, and governance objectives. This approach encourages multi-stakeholder participation in decision-making and promotes the integration of ecosystem services into aquaculture planning and management [79]. For restorative aquaculture, this means not only assessing its impact on benthic assemblages and sediment quality, but also designing strategies that maximise its ecological benefits without compromising its economic and social viability [80].
The future of restorative aquaculture will depend on our ability to integrate science, management, and policy into a development model that not only meets the growing demand for sustainable food but also actively enhances the resilience of marine ecosystems. This requires a paradigm shift in how we perceive aquaculture—not merely as an extractive activity, but as a driver of environmental regeneration. Properly managed bivalve farms can increase the abundance of macrofauna around their installations, providing additional ecological services that boost secondary production and potentially enhance local fisheries. In this way, bivalve aquaculture exemplifies how human food production can be aligned with the restoration and enrichment of marine ecosystems, transforming aquaculture from resource exploitation into a truly regenerative practice.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Example of images obtained for the assessment of Ophicomina nigra abundance, near (left) and far (right) from the mussel farming rafts. The surveyed area was 1 m².
Author Contributions
Conceptualization, E.C, A.M, L.L and P.S..; methodology, E.C, A.M, L.L. and P.D.; software, E.C, A.M., P.S.; validation, P.T, J.T. formal analysis, E.C., A.M; investigation, E.C., ; resources, P.S, J.T.; data curation, A.M.; writing—original draft preparation, E.C., P.S.; writing—review and editing, E.C. and P.S.; visualization, E.C.; supervision, P.S.; project administration, P.S; funding acquisition, P.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Project CLIMAREST, founded by the European Union’s Horizon Europe research and innovation programme under Grant Agreement No. 101093865.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data will be made available on request.
Acknowledgments
We thank Raúl Guillén Vaquero for his assistance with field work for ophiuroids sampling.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| NW | Northwest |
| GLMM | Generalised Linear Mixed Model |
| EU | European Union |
References
- Rogers, A.D.; Aburto-Oropeza, O.; Appeltans, W.; Assis, J.; Ballance, L.T.; Cury, P.; Duarte, C.; Favoretto, F.; Kumagai, J.; Lovelock, C.; et al. Critical Habitats and Biodiversity: Inventory, Thresholds and Governance. In The Blue Compendium [Internet]; Lubchenco, J., Haugan, P.M., Eds.; Springer International Publishing: Cham, 2023; pp. 333–392. [Google Scholar] [CrossRef]
- Overton, K.; Dempster, T.; Swearer, S.E.; Morris, R.L.; Barrett, L.T. Achieving conservation and restoration outcomes through ecologically beneficial aquaculture. Conserv. Biol. 2023, 38, e14065. [Google Scholar] [CrossRef]
- Theuerkauf, S.J.; Barrett, L.T.; Alleway, H.K.; Costa-Pierce, B.A.; Gelais, A.S.; Jones, R.C. Habitat value of bivalve shellfish and seaweed aquaculture for fish and invertebrates: Pathways, synthesis and next steps. Rev. Aquac. 2021, 14, 54–72. [Google Scholar] [CrossRef]
- Hassan R, Scholes R, Ash N, Condition M, Group T. Ecosystems and Human Well-Being: Current State and Trends: Findings of the Condition and Trends Working Group (Millennium Ecosystem Assessment Series). 2005.
- FAO. The State of World Fisheries and Aquaculture 2024 [Internet]. FAO; 2024 [cited 2024 Aug 28]. Available from: https://openknowledge.fao.org/handle/20.500.14283/cd0683en.
- Wilding, T.A.; Nickell, T.D.; Solan, M. Changes in Benthos Associated with Mussel (Mytilus edulis L.) Farms on the West-Coast of Scotland. PLOS ONE 2013, 8, e68313. [Google Scholar] [CrossRef]
- Grant, C.; Archambault, P.; Olivier, F.; McKindsey, C.W. Influence of ‘bouchot’ mussel culture on the benthic environment in a dynamic intertidal system. Aquac. Environ. Interactions 2012, 2, 117–131. [Google Scholar] [CrossRef]
- Chamberlain, J.; Fernandes, T.F.; Read, P.; Nickell, T.D.; Davies, I.M. Impacts of biodeposits from suspended mussel (Mytilus edulis L.) culture on the surrounding surficial sediments. ICES J. Mar. Sci. 2001, 58, 411–416. [Google Scholar] [CrossRef]
- Hartstein, N.D.; A Rowden, A. Effect of biodeposits from mussel culture on macroinvertebrate assemblages at sites of different hydrodynamic regime. Mar. Environ. Res. 2004, 57, 339–357. [Google Scholar] [CrossRef] [PubMed]
- Mascorda-Cabre, L.; Hosegood, P.; Attrill, M.J.; Bridger, D.; Sheehan, E.V. Detecting sediment recovery below an offshore longline mussel farm: A macrobenthic Biological Trait Analysis (BTA). Mar. Pollut. Bull. 2023, 195, 115556. [Google Scholar] [CrossRef]
- Drouin, A.; Archambault, P.; Clynick, B.; Richer, K.; McKindsey, C. Influence of mussel aquaculture on the distribution of vagile benthic macrofauna in îles de la Madeleine, eastern Canada. Aquac. Environ. Interactions 2015, 6, 175–183. [Google Scholar] [CrossRef]
- Sean, A.-S.; Drouin, A.; Archambault, P.; McKindsey, C.W. Influence of an Offshore Mussel Aquaculture Site on the Distribution of Epibenthic Macrofauna in Îles de la Madeleine, Eastern Canada. Front. Mar. Sci. 2022, 9. [Google Scholar] [CrossRef]
- D’aMours, O.; Archambault, P.; McKindsey, C.; Johnson, L. Local enhancement of epibenthic macrofauna by aquaculture activities. Mar. Ecol. Prog. Ser. 2008, 371, 73–84. [Google Scholar] [CrossRef]
- Bridger, D.; Attrill, M.J.; Davies, B.F.R.; Holmes, L.A.; Cartwright, A.; Rees, S.E.; Cabre, L.M.; Sheehan, E.V. The restoration potential of offshore mussel farming on degraded seabed habitat. Aquac. Fish Fish. 2022, 2, 437–449. [Google Scholar] [CrossRef]
- Alleway, H.K.; Waters, T.J.; Brummett, R.; Cai, J.; Cao, L.; Cayten, M.R.; Costa-Pierce, B.A.; Dong, Y.; Hansen, S.C.B.; Liu, S.; et al. Global principles for restorative aquaculture to foster aquaculture practices that benefit the environment. Conserv. Sci. Pr. 2023, 5, e12982. [Google Scholar] [CrossRef]
- Olaso Toca, I. Ecología de los equinodermos de la Ría de Arosa: N.o334. Boletín Instituto Español de Oceanografía [Internet]. 1982, 7, 3–30. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Figueiras, F.G.; Labarta, U.; Fernández Reiriz, M.J. Coastal upwelling, primary production and mussel growth in the Rías Baixas of Galicia. Hydrobiologia 2002, 484, 121–131. [Google Scholar] [CrossRef]
- Brooks, ME, Kristensen, K, van Benthem, KJ, Magnusson, A, Berg, CW, Nielsen, A, et al. glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. The R Journal 2017, 9, 378–400.
- Kaspar, H.F.; Gillespie, P.A.; Boyer, I.C.; MacKenzie, A.L. Effects of mussel aquaculture on the nitrogen cycle and benthic communities in Kenepuru Sound, Marlborough Sounds, New Zealand. Mar. Biol. 1985, 85, 127–136. [Google Scholar] [CrossRef]
- Shumway, S.; Davis, C.; Downey, R.; Karney, R.; Kraeuter, J.; Parsons, J.; et al. Guest Editorial Shellfish aquaculture — In praise of sustainable economies and environments. World Aquaculture 2012, 34. [Google Scholar]
- Rose, J.M.; Bricker, S.B.; Tedesco, M.A.; Wikfors, G.H. A Role for Shellfish Aquaculture in Coastal Nitrogen Management. Environ. Sci. Technol. 2014, 48, 2519–2525. [Google Scholar] [CrossRef]
- Davidson, R.J.; Scrimgeour, G.J.; Richards, L.A.; Locky, D. Multi-Spatial and Temporal Assessments of Impacts and Recovery of Epibenthic Species and Habitats Under Mussel Farms in the Marlborough Sounds, New Zealand. J. Shellfish. Res. 2024, 43, 15–28. [Google Scholar] [CrossRef]
- Inglis, G.J.; Gust, N. Potential indirect effects of shellfish culture on the reproductive success of benthic predators. J. Appl. Ecol. 2003, 40, 1077–1089. [Google Scholar] [CrossRef]
- Cabre, L.M.; Hosegood, P.; Attrill, M.J.; Bridger, D.; Sheehan, E.V. Offshore longline mussel farms: a review of oceanographic and ecological interactions to inform future research needs, policy and management. Rev. Aquac. 2021, 13, 1864–1887. [Google Scholar] [CrossRef]
- Stamp, T.; Pittman, S.J.; Holmes, L.A.; Rees, A.; Ciotti, B.J.; Thatcher, H.; Davies, P.; Hall, A.; Wells, G.; Olczak, A.; et al. Restorative function of offshore longline mussel farms with ecological benefits for commercial crustacean species. Sci. Total. Environ. 2024, 951, 174987. [Google Scholar] [CrossRef]
- McKindsey CW, Anderson MR, Courtenay S, Landry T, Skinner M. Effets off Shellfish Aquaculture on Fish Habitat. 2006.
- Powers, M.J.; Peterson, C.H.; Summerson, H.C.; Powers, S.P. Macroalgal growth on bivalve aquaculture netting enhances nursery habitat for mobile invertebrates and juvenile fishes. Mar. Ecol. Prog. Ser. 2007, 339, 109–122. [Google Scholar] [CrossRef]
- Costa-Pierce, B.A.; Bridger, C.J. The role of marine aquaculture facilities as habitats and ecosystems. Responsible Marine Aquaculture. 2002, 105–144. [Google Scholar]
- Marine Conservation Society. Good Fish Guide: Best Choice seafood [Internet]. 2018. Available from: https://www.mcsuk.org/news/why-you-should-be-eating-more-uk-shellfish.
- Food and Agriculture Organization of the United Nations. The State of World Fisheries and Aquaculture 2020: Sustainability in action. 2020.
- Callier, M.D.; McKindsey, C.W.; Desrosiers, G. Evaluation of indicators used to detect mussel farm influence on the benthos: Two case studies in the Magdalen Islands, Eastern Canada. Aquaculture 2008, 278, 77–88. [Google Scholar] [CrossRef]
- Clynick, B.; McKindsey, C.; Archambault, P. Distribution and productivity of fish and macroinvertebrates in mussel aquaculture sites in the Magdalen islands (Québec, Canada). Aquaculture 2008, 283, 203–210. [Google Scholar] [CrossRef]
- Romero, P.; González-Gurriarán, E.; Penas, E. Influence of mussel rafts on spatial and seasonal abundance of crabs in the R a de Arousa, North-West Spain. Mar. Biol. 1982, 72, 201–210. [Google Scholar] [CrossRef]
- Drummond-Davis, N.C.; Mann, K.H.; Pottle, R.A. Some Estimates of Population Density and Feeding Habits of the Rock Crab, Cancer irroratus, in a Kelp Bed in Nova Scotia. Can. J. Fish. Aquat. Sci. 1982, 39, 636–639. [Google Scholar] [CrossRef]
- Dolmer, P. The interactions between bed structure of Mytilus edulis L. and the predator Asterias rubens L. J. Exp. Mar. Biol. Ecol. 1998, 228, 137–150. [Google Scholar] [CrossRef]
- Lees, K.; Lavoie, M.; Macgregor, K.; Simard, É.; Drouin, A.; Comeau, L.; McKindsey, C. Movement of American lobsters Homarus americanus and rock crabs Cancer irroratus around mussel farms in Malpeque Bay, Prince Edward Island, Canada. Aquac. Environ. Interactions 2023, 15, 179–193. [Google Scholar] [CrossRef]
- Wong, M.C.; Barbeau, M.A. Prey selection and the functional response of sea stars (Asterias vulgaris Verrill) and rock crabs (Cancer irroratus Say) preying on juvenile sea scallops (Placopecten magellanicus (Gmelin)) and blue mussels (Mytilus edulis Linnaeus). J. Exp. Mar. Biol. Ecol. 2005, 327, 1–21. [Google Scholar] [CrossRef]
- Freire, J.; González-Gurriarán, E. Feeding ecology of the velvet swimming crab Necora puber in mussel raft areas of the Ría de Arousa (Galicia, NW Spain). Mar. Ecol. Prog. Ser. 1995, 119, 139–154. [Google Scholar] [CrossRef]
- Lavoie, M.; Simard, É.; Drouin, A.; Archambault, P.; Comeau, L.; McKindsey, C. Movement of american lobster Homarus americanus associated with offshore mussel Mytilus edulis aquaculture. Aquac. Environ. Interactions 2022, 14, 189–204. [Google Scholar] [CrossRef]
- Sardenne, F.; Forget, N.; McKindsey, C.W. Contribution of mussel fall-off from aquaculture to wild lobster Homarus americanus diets. Mar. Environ. Res. 2019, 149, 126–136. [Google Scholar] [CrossRef]
- Agüera, A.; Saurel, C.; Møller, L.F.; Fitridge, I.; Petersen, J.K. Bioenergetics of the common seastar Asterias rubens: a keystone predator and pest for European bivalve culture. Mar. Biol. 2021, 168, 1–14. [Google Scholar] [CrossRef]
- Gaymer, C.F.; Himmelman, J.H.; E Johnson, L. Use of prey resources by the seastars Leptasterias polaris and Asterias vulgaris: a comparison between field observations and laboratory experiments. J. Exp. Mar. Biol. Ecol. 2001, 262, 13–30. [Google Scholar] [CrossRef]
- Barrett, L.T.; Swearer, S.E.; Dempster, T. Native predator limits the capacity of an invasive seastar to exploit a food-rich habitat. Mar. Environ. Res. 2020, 162, 105152. [Google Scholar] [CrossRef]
- Olaso Toca, I. Biología de los equinodermos de la ría de Arosa: N.o 270. Boletín Instituto Español de Oceanografía [Internet]. 1979, 5, 81–128. [Google Scholar]
- Agüera, A. The role of sea stars (Asterias rubens L.) predation in blue mussel (Mytilus edulis L.) seedbed stability. 2015.
- Pryor, M.L. Temporal and spatial distribution of larval and post-larval blue mussels (Mytilus edulis/Mytilus trossulus) and sea stars (Asterias vulgaris) within four Newfoundland mussel culture sites [Internet]. Memorial University of Newfoundland; 2004 [cited 2025 Aug 21]. Available from: https://hdl.handle.net/20.500.
- Barrett, L.T.; Swearer, S.E.; Dempster, T. Impacts of marine and freshwater aquaculture on wildlife: a global meta-analysis. Rev. Aquac. 2018, 11, 1022–1044. [Google Scholar] [CrossRef]
- Bivalve aquaculture and exotic species: A review of ecological considerations and management issues [Internet]. [cited 2025 Aug 21]. Available from: https://www.researchgate.net/publication/232686987_Bivalve_aquaculture_and_exotic_species_A_review_of_ecological_considerations_and_management_issues.
- Zanotto, F.; Wheatly, M. Calcium balance in crustaceans: nutritional aspects of physiological regulation. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 2002, 133, 645–660. [Google Scholar] [CrossRef]
- PDF) Niche segregation between American lobster Homarus amencanus and rock crab Cancer irroratus [Internet]. [cited 2025 Aug 21]. Available from: https://www.researchgate.net/publication/250214574_Niche_segregation_between_American_lobster_Homarus_amencanus_and_rock_crab_Cancer_irroratus. 250.
- Gendron, L.; Fradette, P.; Godbout, G. The importance of rock crab (Cancer irroratus) for growth, condition and ovary development of adult American lobster (Homarus americanus). J. Exp. Mar. Biol. Ecol. 2001, 262, 221–241. [Google Scholar] [CrossRef]
- Wahle, R.A.; Steneck, R.S. Habitat restrictions in early benthic life: experiments on habitat selection and in situ predation with the American lobster. J. Exp. Mar. Biol. Ecol. 1992, 157, 91–114. [Google Scholar] [CrossRef]
- Cobb, J.S. The Shelter-Related Behavior of the Losbter, Homarus Americanus. Ecology 1971, 52, 108–115. [Google Scholar] [CrossRef]
- Karnofsky, E.B.; Atema, J.; Elgin, R.H. Field Observations of Social Behavior, Shelter Use, and Foraging in the Lobster, Homarus americanus. Biol. Bull. 1989, 176, 239–246. [Google Scholar] [CrossRef]
- Lacoste, É.; Drouin, A.; Weise, A.; Archambault, P.; McKindsey, C. Low benthic impact of an offshore mussel farm in Îles-de-la-Madeleine, eastern Canada. Aquac. Environ. Interactions 2018, 10, 473–485. [Google Scholar] [CrossRef]
- McKindsey, C.W.; Archambault, P.; Callier, M.D.; Olivier, F. Influence of suspended and off-bottom mussel culture on the sea bottom and benthic habitats: a review. Can. J. Zoöl. 2011, 89, 622–646. [Google Scholar] [CrossRef]
- European Comission. Quality Status Report (QRS) 2010 [Internet]. 2012. Available from: https://oap.ospar.org/en/ospar-assessments/quality-status-reports/qsr-2023/other-assessments/aquaculture/.
- Sala, E.; Lubchenco, J.; Grorud-Colvert, K.; Novelli, C.; Roberts, C.; Sumaila, U.R. Assessing real progress towards effective ocean protection. Mar. Policy 2018, 91, 11–13. [Google Scholar] [CrossRef]
- Zupan, M.; Fragkopoulou, E.; Claudet, J.; Erzini, K.; e Costa, B.H.; Gonçalves, E.J. Marine partially protected areas: drivers of ecological effectiveness. Front. Ecol. Environ. 2018, 16, 381–387. [Google Scholar] [CrossRef]
- e Costa, B.H.; Claudet, J.; Franco, G.; Erzini, K.; Caro, A.; Gonçalves, E.J. A regulation-based classification system for marine protected areas: A response to Dudley et al. [9]. Mar. Policy 2017, 77, 193–195. [Google Scholar] [CrossRef]
- Fernandez-Jover, D.; Sanchez-Jerez, P.; Bayle-Sempere, J.T.; Valle, C.; Dempster, T. Seasonal patterns and diets of wild fish assemblages associated with Mediterranean coastal fish farms. ICES J. Mar. Sci. 2008, 65, 1153–1160. [Google Scholar] [CrossRef]
- Dempster, T.; Uglem, I.; Sanchez-Jerez, P.; Fernandez-Jover, D.; Bayle-Sempere, J.; Nilsen, R.; Bjørn, P. Coastal salmon farms attract large and persistent aggregations of wild fish: an ecosystem effect. Mar. Ecol. Prog. Ser. 2009, 385, 1–14. [Google Scholar] [CrossRef]
- Weitzman, J. Applying the ecosystem services concept to aquaculture: A review of approaches, definitions, and uses. Ecosyst. Serv. 2019, 35, 194–206. [Google Scholar] [CrossRef]
- Lavoie, M.-F.; Lacoste, É.; Weise, A.M.; McKindsey, C.W. Benthic responses to organic enrichment under a mussel (Mytilus edulis) farm. Front. Mar. Sci. 2024, 11, 1433365. [Google Scholar] [CrossRef]
- Ysebaert, T.; Hart, M.; Herman, P.M.J. Impacts of bottom and suspended cultures of mussels Mytilus spp. on the surrounding sedimentary environment and macrobenthic biodiversity. Helgol. Mar. Res. 2008, 63, 59–74. [Google Scholar] [CrossRef]
- Mizuta, D.D.; Froehlich, H.E.; Wilson, J.R. The changing role and definitions of aquaculture for environmental purposes. Rev. Aquac. 2022, 15, 130–141. [Google Scholar] [CrossRef]
- Burg, S.v.D.; Termeer, E.; Skirtun, M.; Poelman, M.; Veraart, J.; Selnes, T. Exploring mechanisms to pay for ecosystem services provided by mussels, oysters and seaweeds. Ecosyst. Serv. 2022, 54. [Google Scholar] [CrossRef]
- Willot, P.-A.; Aubin, J.; Salles, J.-M.; Wilfart, A. Ecosystem service framework and typology for an ecosystem approach to aquaculture. Aquaculture 2019, 512. [Google Scholar] [CrossRef]
- Vivier, B.; Dauvin, J.-C.; Navon, M.; Rusig, A.-M.; Mussio, I.; Orvain, F.; Boutouil, M.; Claquin, P. Marine artificial reefs, a meta-analysis of their design, objectives and effectiveness. Glob. Ecol. Conserv. 2021, 27. [Google Scholar] [CrossRef]
- Perkol-Finkel, S.; Hadary, T.; Rella, A.; Shirazi, R.; Sella, I. Seascape architecture – incorporating ecological considerations in design of coastal and marine infrastructure. Ecol. Eng. 2018, 120, 645–654. [Google Scholar] [CrossRef]
- Langhamer, O. Artificial Reef Effect in relation to Offshore Renewable Energy Conversion: State of the Art. Sci. World J. 2012, 2012, 1–8. [Google Scholar] [CrossRef]
- Zhao, F.; Wu, J. The Role of Shellfish Aquaculture in Coastal Habitat Restoration. Int. J. Mar. Sci. 2024, 14. [Google Scholar] [CrossRef]
- Pacifico, A.M.; Brigolin, D.; Mulazzani, L.; Semeraro, M.; Malorgio, G. Managing marine aquaculture by assessing its contribution to ecosystem services provision: The case of Mediterranean mussel, Mytilus galloprovincialis. Ocean Coast. Manag. 2024, 259. [Google Scholar] [CrossRef]
- Gimpel, A.; Stelzenmüller, V.; Töpsch, S.; Galparsoro, I.; Gubbins, M.; Miller, D.; Murillas, A.; Murray, A.G.; Pınarbaşı, K.; Roca, G.; et al. A GIS-based tool for an integrated assessment of spatial planning trade-offs with aquaculture. Sci. Total. Environ. 2018, 627, 1644–1655. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, H.; Stamp, T.; Wilcockson, D.; Moore, P.J.; Birchenough, S.; Lopez, L.L. Residency and habitat use of European lobster (Homarus gammarus) within an offshore wind farm. ICES J. Mar. Sci. 2023, 80, 1410–1421. [Google Scholar] [CrossRef]
- Food and Land Use Coalition. Growing Better: Ten Critical Transitions to Transform Food and Land Use. 2019.
- Soto D, Aguilar-Manjarrez J, Hishamunda N. Building an ecosystem approach to aquaculture: FAO/Universitat de les Illes Balears expert workshop, 7-11 may 2007, Palma de Mallorca, Spain. Rome: FAO; 2008.
- Brugère, C.; Aguilar-Manjarrez, J.; Beveridge, M.C.M.; Soto, D. The ecosystem approach to aquaculture 10 years on – a critical review and consideration of its future role in blue growth. Rev. Aquac. 2018, 11, 493–514. [Google Scholar] [CrossRef]
- L, B. First Evidence of Spatial Relationships between Ecosystem Functioning and Services in the marine environment. J. Ecol. Nat. Resour. 2023, 7. [Google Scholar] [CrossRef]
Figure 1.
Study area in the Ría de Vigo (NW Iberian Peninsula) with the position of sampled rafts.
Figure 2.
(A) Global distribution of the mussel production and (B) the scientific effort on the ecological benefits of the bivalve aquaculture. The bubble chart represents the number of publications from each country that has contributed to this field. The colour coding indicates the annual production levels for each country in the year 2022.
Figure 2.
(A) Global distribution of the mussel production and (B) the scientific effort on the ecological benefits of the bivalve aquaculture. The bubble chart represents the number of publications from each country that has contributed to this field. The colour coding indicates the annual production levels for each country in the year 2022.
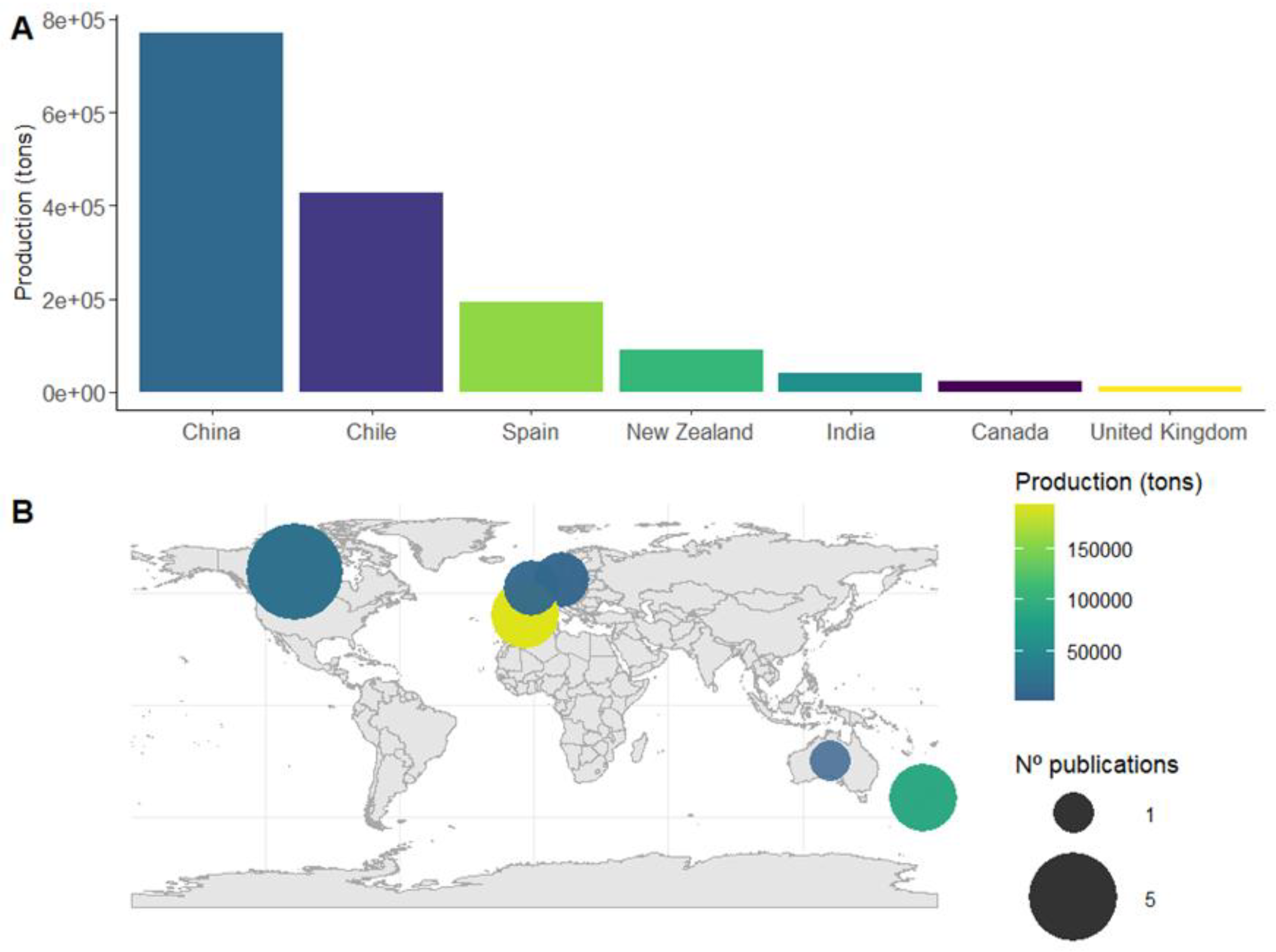
Figure 3.
Estimated abundance of O. nigra (ind/m2) in the Ría of Vigo at different Distance levels. The boxes represent the interquartile range (25th-75th percentile) , whiskers indicate the minimum and maximum values within 1.5 times the interquartile range, dots represent outliers, and the horizontal bar represents the median value. Small dots represent real abundance values.
Figure 3.
Estimated abundance of O. nigra (ind/m2) in the Ría of Vigo at different Distance levels. The boxes represent the interquartile range (25th-75th percentile) , whiskers indicate the minimum and maximum values within 1.5 times the interquartile range, dots represent outliers, and the horizontal bar represents the median value. Small dots represent real abundance values.
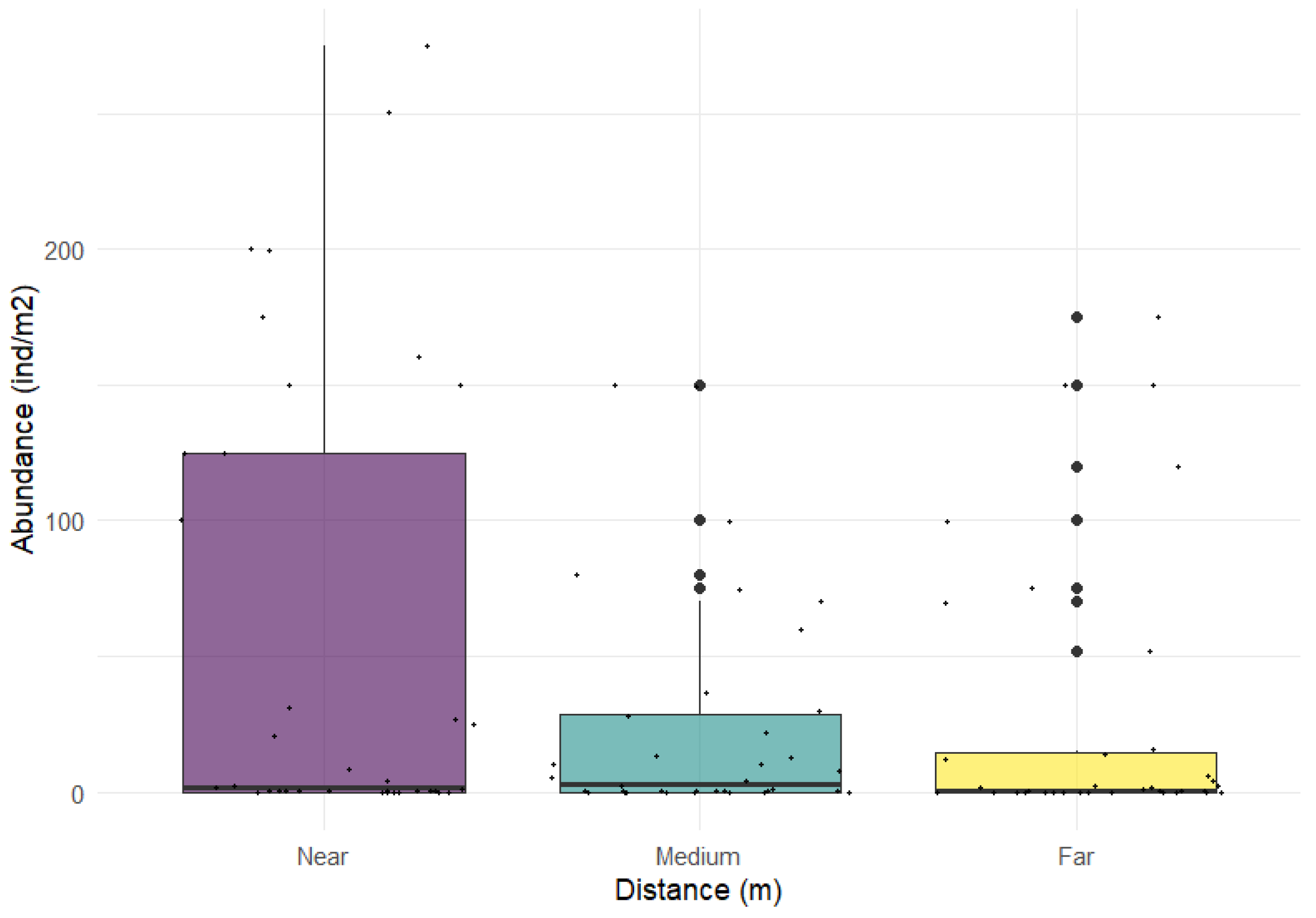
Table 2.
Results of the generalized linear mixed model (GLMM) testing the effect of distance from the rafts on Ophiocomia nigra. The model was fitted with a negative binomial distribution and included rafts as random effect.
Table 2.
Results of the generalized linear mixed model (GLMM) testing the effect of distance from the rafts on Ophiocomia nigra. The model was fitted with a negative binomial distribution and included rafts as random effect.
| P value | Z value | Std. Error | Estimate | Fixed effects |
|---|---|---|---|---|
| 0.436 | 0.779 | 1.633 | 1.272 | Near |
| 0.099 | -1.648 | 0.476 | -0.784 | Middle |
| 0.375 | -0.888 | 0.618 | -0.549 | Far |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



