
Submitted:
15 August 2025
Posted:
18 August 2025
You are already at the latest version
Abstract
This study investigates the species composition, seasonal variation, and diversity dynamics of macroinvertebrate communities associated with Mytilus galloprovincialis (Lamarck, 1819) cultivated in offshore longline systems in the central Black Sea. Monthly sampling conducted between September 2023 and August 2024 yielded a total of 99,719 individuals representing 20 taxa. The communities were predominantly dominated by amphipods, particularly Jassa marmorata (71.3%) and Stenothoe monoculoides (27.8%). Individual abundance peaked in autumn, whereas species richness reached its highest levels in summer. ANOVA analyses revealed significant seasonal differences in diversity indices (p< 0.05). Multivariate analyses, including NMDS and PCA (RDA proxy), indicated distinct seasonal clustering, with pH (41%) and salinity (34.5%) identified as the primary environmental drivers of community composition. These findings demonstrate that offshore mussel longline systems function not only as aquaculture infrastructure but also as reef-like artificial habitats that support benthic biodiversity. The dominance of opportunistic and detritivorous amphipods, along with their sensitivity to environmental gradients, suggests their potential utility as bioindicators for ecological monitoring.
Keywords:
mussel aquaculture
; macroinvertebrates
; Jassa marmorata
; artificial reefs
; benthic biodiversity
; ecological ındicators
1. Introduction
The Black Sea, due to its limited water exchange with other marine systems, is recognized as the largest semi-enclosed inland sea in the world. This unique hydrographic structure directly shapes the region’s physical, chemical, and biological characteristics. Surface waters are dominated by freshwater inputs from rivers, resulting in salinity levels as low as approximately 17‰. With increasing depth, more saline, Mediterranean-origin waters occupy the deeper layers, leading to the formation of a pronounced halocline 1, 2]. This density stratification severely restricts vertical mixing between surface and bottom waters, rendering the layers below 150–200 meters anoxic. Beneath this anoxic zone, hydrogen sulfide (H₂S) accumulates, making the Black Sea one of the rare marine systems where aerobic life is absent in the deep-water column [3].
Such stratification directly influences phytoplankton productivity, zooplankton composition, and benthic life. In addition, high nutrient inputs particularly nitrogen and phosphorus delivered by rivers trigger eutrophication processes, leading to algal blooms, oxygen depletion, and an overall deterioration in ecosystem health [4]. These dynamics significantly shape the distribution and diversity of macrozoobenthic communities in the region.
The Black Sea, with its rich biological diversity and ecological heterogeneity, holds significant importance at both regional and global scales. Its benthic ecosystems, characterized by diverse habitats and high species richness, play a critical role in maintaining ecological balance and sustaining biological productivity [5,6,7]. In the mediolittoral and infralittoral zones of the Black Sea and the Sea of Marmara, M. galloprovincialis Lamarck, 1819 forms dense beds on hard substrates [3,8,9].
Mussel aquaculture systems are valuable not only as units of food production but also in terms of the ecosystem services they provide, including habitat provision, contribution to nutrient cycling, and enhancement of biodiversity. In recent years, there has been growing interest in the role of these systems as reef-like structures [10,11].
Macroinvertebrates are widely recognized as effective bioindicators due to their sensitivity to environmental changes and their integral roles within benthic food webs [12,13]. In the Black Sea, M. galloprovincialis beds are generally observed in shallow waters, but they can extend to depths of 30–50 meters, and in some areas, even down to 80 meters [14,15]. These mussels are capable of colonizing a wide variety of habitats, ranging from coastal hard substrates and rocky shores to artificial structures and deep-sea muds.
As suspension feeders, M. galloprovincialis consume phytoplankton, organic detritus, bacteria, and dissolved organic matter present in the water column [16,17]. The productivity of natural populations can be limited by factors such as physiological stress, food scarcity, predation, and density dependent pressures [18,19]. Macro benthic organisms associated with mussel beds are generally categorized into ecological groups such as epibenthic fauna, epiphytic fauna, infauna, and free-living fauna [20,21]. The composition of these groups is influenced by environmental conditions and by the accumulation of particulate organic matter within the mussel beds [22].
In recent years, numerous scientific studies have been conducted in the Black Sea focusing on the monitoring and conservation of benthic invertebrates. These efforts provide valuable insights into ecosystem health assessment and biodiversity conservation. Benthic invertebrates are indispensable components for maintaining ecosystem balance and water quality, and their protection is crucial for ensuring the region’s long-term ecological sustainability [23].
The Black Sea’s water temperature, ranging between 7 and 25 °C, and salinity levels between 17–20 ppt offer favorable conditions for mussel aquaculture [24,25]. Mussel farms in the region are designed as surface-deployed longline systems, based on the expectation that the high phytoplankton concentrations in surface waters will promote mussel growth. Studies on macro benthic invertebrates associated with M. galloprovincialis have identified amphipods as one of the dominant groups inhabiting these environments [26,27]. Amphipods utilize mussel beds both as shelter and as feeding grounds within these detritus- and microorganism-rich habitats. Species belonging to the families Gammaridae and Caprellidae make significant contributions to the ecological richness of mussel beds. Such symbiotic associations enhance the biodiversity and structural complexity of mussel assemblages, thereby supporting ecosystem sustainability.
This study posits that the structure and diversity of macroinvertebrate communities in offshore mussel aquaculture systems are strongly influenced by environmental gradients particularly pH and salinityand that under moderate environmental stress conditions, opportunistic amphipod species (e.g., J. marmorata Holmes, 1905) tend to dominate. Accordingly, these communities may serve as effective biological indicators for assessing environmental quality.
2. Materials and Methods
2.1. Longline System
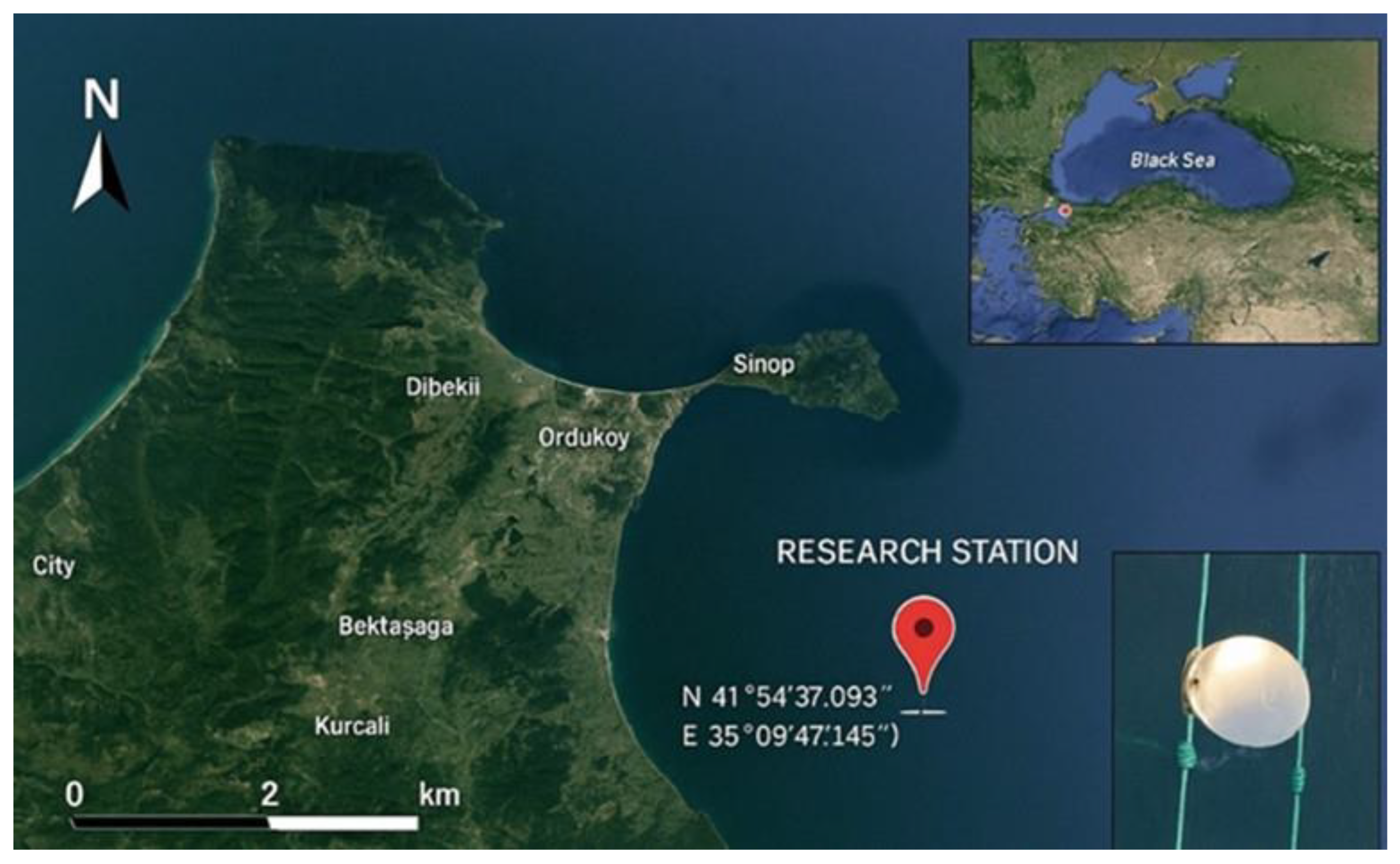
The “Mussel Aquaculture Research Facility” is located within a First-Degree Aquaculture Zone in Sinop Province, Türkiye, and consists of a surface-deployed longline system (Figure 1). The nearest point of the project site to the shore is approximately 4,490 meters, with depths ranging between 35 and 42 meters. The study was conducted in the central Black Sea, in offshore waters at depths of 25 to 27 meters, between September 2023 and August 2024. This marine area was selected due to its reduced exposure to harsh weather conditions such as strong winds, currents, and high waves. The system was designed in accordance with local environmental conditions and consists of twelve submerged longline units, each measuring 8 meters in length (Figure 2).
Figure 1.
Coordinates of the Longline System.
2.2. Environmental Parameters
Water temperature and salinity data were measured monthly between September 2023 and August 2024 using a YSI 6600 multi-parameter probe. Measurements were conducted in situ by immersing the probe directly into the water from the sampling vessel, and the data were recorded immediately. All water parameter data collected during the one-year sampling period were compiled into an Excel file and prepared for subsequent analyses.
2.3. Mussel Sample Collection
In the longline system, each 8-meter-long rope was suspended within the water column using buoys and weights. Sampling was carried out over a 12 month period, with each sampling event conducted between 09:00 and 14:00. On twelve standardized mussel ropes, uniform in length, thickness, and material, stopper pins were positioned at 90 cm intervals to prevent mussel slippage. Macroinvertebrate samples were collected from 30 cm segments located between these pins. To avoid resampling, sampled sections were marked with colored ropes. To ensure seasonal representativeness, three replicate samples were collected monthly from mussel ropes suspended at a depth of 3 meters, using standardized mesh bags.
Macroinvertebrate sampling was conducted using mesh bags measuring 30 cm in length, with a mesh size of 0.25 mm, equipped with Velcro fastenings on three sides. These bags were secured in situ around the designated rope segments. Mussels and associated organisms were gently loosened by hand into the bag. The Velcro closures were then carefully opened and resealed to remove the contents in a controlled manner [28].
Collected material was emptied into a container on board and transferred with a small scoop into plastic jars with a capacity of 500–1000 ml. Samples were preserved in 96% ethanol for subsequent processing. The total number of individuals obtained from the three replicates was divided by three to calculate the mean individual abundance per unit.
Figure 2.
Longline System.

Once transported to the laboratory, mussel samples were washed using a double-layered sieving system with mesh sizes of 2 mm and 0.5 mm. Organisms retained on the 0.5 mm sieve were included in the study, while smaller fractions were excluded. The retained material was sorted under a stereomicroscope with appropriate illumination, separated into taxonomic groups, labeled, and preserved in 75% ethanol for species identification. Each specimen was identified to the lowest possible taxonomic level. However, in cases where individuals were fragmented or lacked sufficient morphological features, taxa were classified at the genus level. Species lists and biodiversity index results were reported according to the sampling site.
For the identification of marine macro-benthic invertebrates, standard morphological keys and regional faunal references were used. For members of the class Polychaeta, [29] and [30] were consulted, while [31] and [27] served as primary references for mollusks. For Crustacea particularly Amphipoda and Decapoda [32] and [33] were used. Species identifications and current taxonomic information were further verified using digital databases such as the World Register of Marine Species [34] and the Ocean Biodiversity Information System [35].
2.4. Data Analysis
All statistical analyses were performed using R software (v4.3.1) with the vegan and ggplot2 packages. Analyses included one-way ANOVA, diversity indices (Shannon, Simpson, and Pielou’s Evenness Index), Non-Metric Multidimensional Scaling (NMDS), and Principal Component Analysis (PCA, used as a proxy for RDA).
The dominance index for each identified taxon was calculated according to [20] using the following formula:
Here,
D = Dominance percentage of species i
Ni = Number of individuals belonging to species i,
Nt = otal number of macrobenthic invertebrate individuals
To characterize the monthly macroinvertebrate communities, diversity and evenness metrics were applied. These included the Shannon-Wiener Diversity Index (H') [36], Pielou’s Evenness Index (J') [37], and Simpson’s Diversity Index (1–D). Shannon’s diversity index was calculated using the formula proposed by [36]:
where
- H' = Shannon diversity index
- Pi = Proportion of individuals in taxon i
- Ni = Number of individuals in taxon i
- N = Total number of individuals in the sample
- S = Total number of taxa
This index ranges from 0 to 5, although it rarely exceeds 1; values closer to 5 indicate higher taxonomic diversity.
Pielou’s evenness index was calculated using the formula:
Here,
S =represents the number of taxa in the sample, while
H = refers to the Shannon diversity index value [36]. This index ranges from 0 to 1, with values approaching 1 indicating that taxa are distributed relatively evenly throughout the community. The most commonly used formula for Simpson’s Diversit Index is as follows:
D = Simpson's index (measures the probability that two individuals randomly selected from a sample will belong to the same species)
S = Total number of species in the community (species richness)
n = Number of individuals of species i
N = Total number of individuals of all species
= Proportional abundance of species i
3. Results
3.1. Physicochemical Parameters and Seasonal Variations
Figure 3.
Monthly Physicochemical Parameter Data.
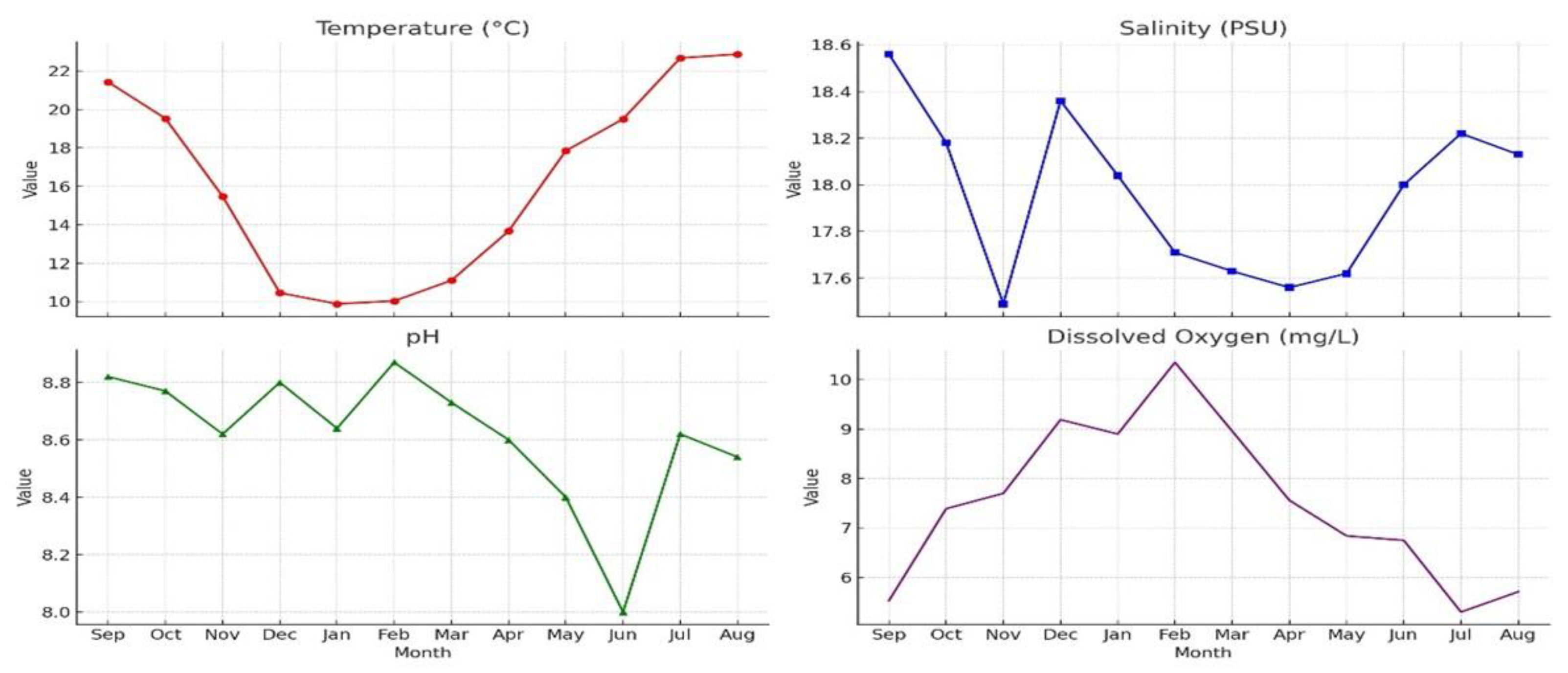
An overview of the water parameters measured throughout the one-year sampling period is presented in Figure 3:
Water Temperature (T): Water temperature exhibited a typical seasonal cycle, with the lowest value recorded in winter (9.88 °C in January) and the highest in summer (22.88 °C in August). The lowest temperatures occurred in January–February, followed by a gradual increase starting in May.
Salinity (S): Salinity fluctuated within a relatively narrow range throughout the year, reaching its minimum in autumn and maximum at the end of summer. These variations are likely influenced by environmental factors such as freshwater inputs and evaporation rates.
pH: pH values remained within a slightly alkaline range (8.00–8.87). Increases during winter may be associated with lower temperatures and higher dissolved oxygen levels, whereas decreases observed in summer could be linked to increased biological activity and the decomposition of organic matter.
Dissolved Oxygen (DO): Dissolved oxygen levels peaked in February at 10.35 mg/L and reached their lowest level in July at 5.30 mg/L. This pattern corresponds with the reduced solubility of oxygen at higher temperatures and increased biological oxygen demand during the warmer months.
3.2. Macroinvertebrate Community
During the 12-month sampling period conducted within the mussel longline system in the Black Sea, a total of 99,719 macroinvertebrate individuals representing 20 taxa were recorded (Table 1). The identified taxa encompassed a wide range of invertebrate groups, including Crustacea, Mollusca, Polychaeta, Cirripedia, Platyhelminthes, Cnidaria, and Nematoda. This diversity reflects both the ecological heterogeneity and the structural complexity of the benthic community associated with mussel aquaculture.

Table 1.
Dominant macroinvertebrate species ranked by mean relative abundance, with notes on their ecological roles.
Table 1.
Dominant macroinvertebrate species ranked by mean relative abundance, with notes on their ecological roles.
| Species | Mean Relative Abundance (%) | Ecological Role |
|---|---|---|
| Jassa marmorata | 71.36% | Opportunist, habitat-forming |
| Stenothoe monoculoides | 27.8% | Detritivore, inhabits sticky substrates |
| Nereis zonata | 0.37% | Omnivore, sediment bioturbator |
| Nematoda (general) | 0.12% | Microscopic, sensitive to organic matter |
| Hyale crassipes | 0.10% | Detritivore, shows seasonal abundance trend |
3.3. Species Distribution and Seasonal Variation
Monthly species distribution exhibited pronounced temporal (seasonal) variability, largely shaped by the dominance of J. marmorata Holmes, 1905, which accounted for approximately 71.36% of the total individuals (Table 1). These dominant taxa are considered to play key roles in the trophic structure and habitat dynamics of benthic environments associated with mussel beds.
Figure 4.
Shannon–Wiener (H′), Simpson (1–D), and Pielou (J′) diversity indices.
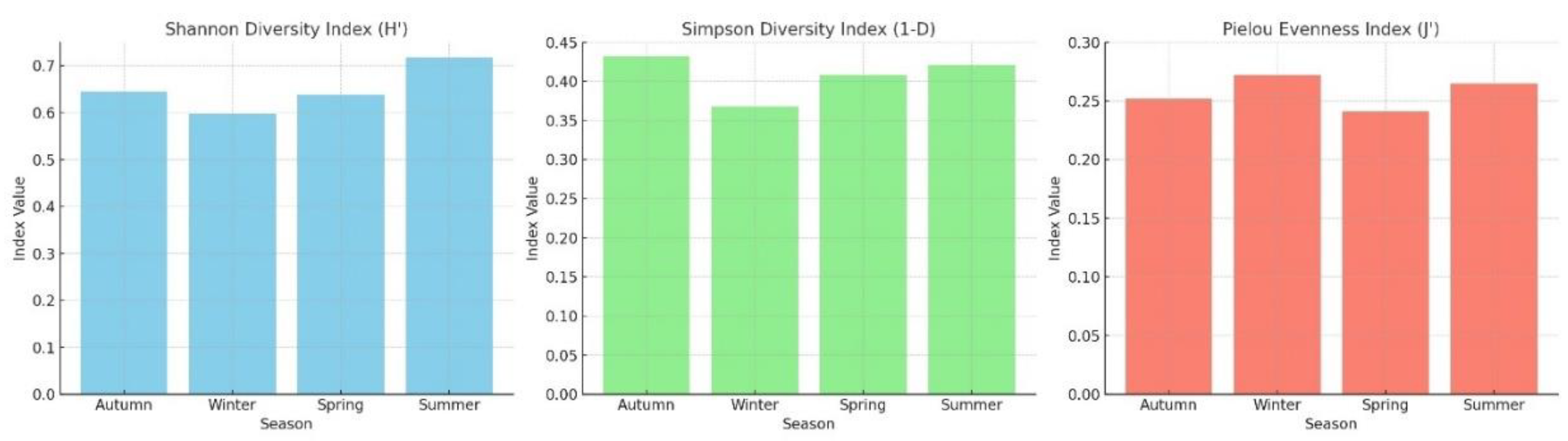
ANOVA results indicated statistically significant seasonal differences in the Shannon diversity index (p < 0.05), with the highest diversity observed during summer and the lowest values recorded in winter. To evaluate the ecological diversity of the macrozoobenthic community structure, the Shannon–Wiener (H′), Simpson (1–D), and Pielou’s Evenness (J′) indices were calculated. The index values revealed notable temporal variations in diversity and distribution patterns over the 12-month study period (Figure 4).
Although the Kruskal–Wallis test revealed no statistically significant differences (p > 0.05), species richness peaked in summer, while individual abundance reached its maximum in autumn. This pattern suggests asynchronous responses of the community to environmental variables.
NMDS analysis revealed clear seasonal clustering. The greatest Bray–Curtis dissimilarity was found between summer and autumn samples (0.499), whereas the highest similarity occurred between spring and autumn samples (0.119).
Although the Kruskal–Wallis test revealed no statistically significant differences (p > 0.05), species richness peaked in summer, while individual abundance reached its maximum in autumn. This pattern suggests asynchronous responses of macroinvertebrate communities to environmental variables.
A pronounced increase in both species richness and individual abundance was observed during summer and early autumn (June–September). This seasonal peak coincides with periods of higher temperatures and reduced hydrodynamic impact, indicating enhanced macrofaunal productivity. The consistent dominance of J. marmorata during these months reflects the species’ rapid colonization ability and competitive advantage on artificial structures. In contrast, the lower and more stable abundances recorded in winter and spring are consistent with suppressed benthic activity under low temperatures and elevated oxygen conditions (Figure 5).
Total abundance reached its highest levels in autumn, potentially reflecting a post-reproductive dispersal phase of dominant taxa following their summer breeding period. Conversely, winter was characterized by a notable decline in both richness and abundance, likely driven by stressors such as low temperatures, high dissolved oxygen levels, and limited organic matter availability.
Figure 5.
Shaded bar graphs showing the monthly macrofaunal composition recorded in the longline system.
Figure 5.
Shaded bar graphs showing the monthly macrofaunal composition recorded in the longline system.
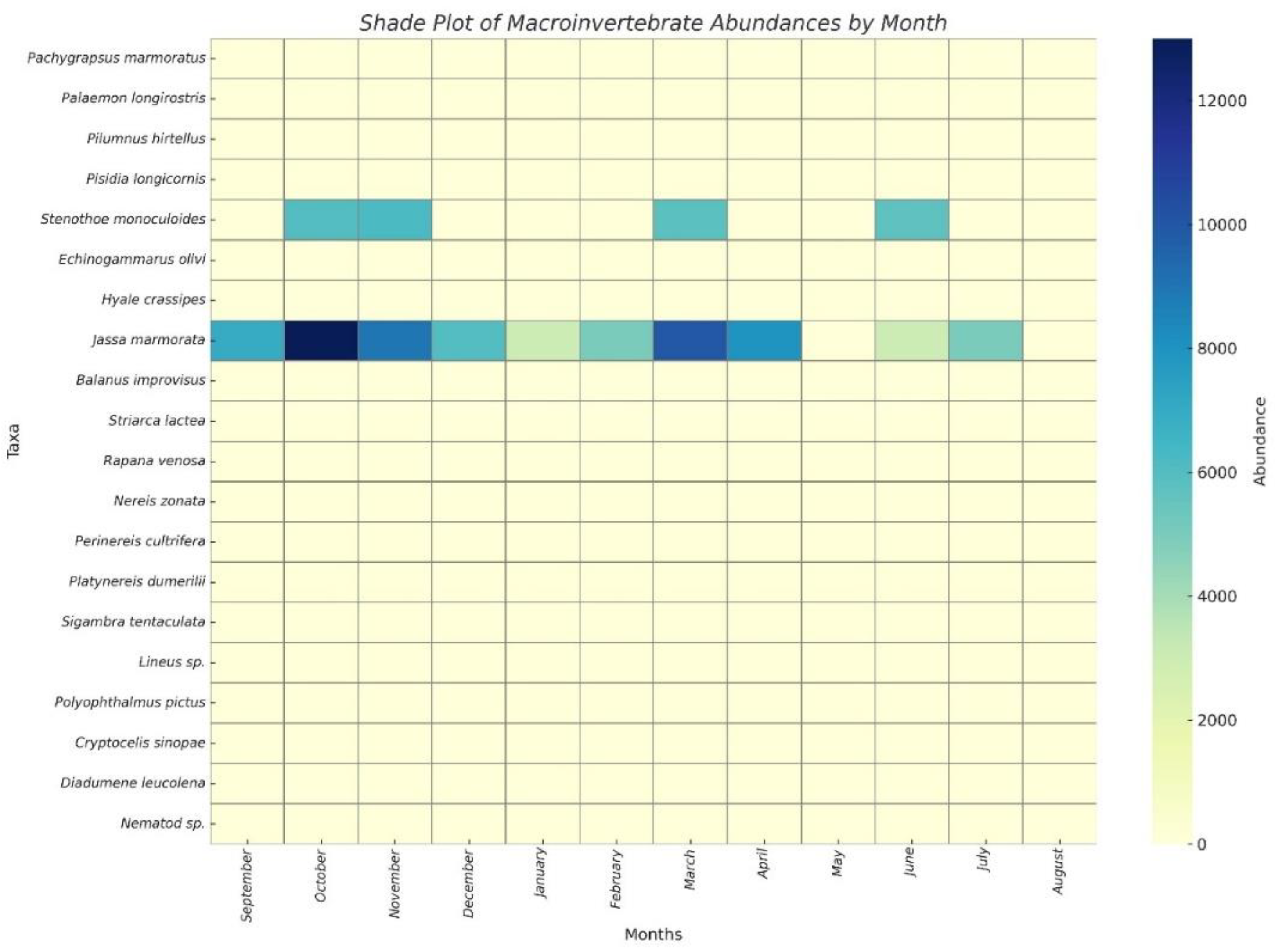
While the dominant species J. marmorata Holmes, 1905 and S. monoculoides (Montagu, 1813) largely shaped the overall temporal trends, several low-abundance taxa such as Balanus improvisus, Cryptocelis sinopae, Hyale crassipes, and Nereis zonata contributed to short-term peaks in richness during specific sampling periods (e.g., S4, S9) (Table 2). Although these episodic fluctuations had minimal influence on total abundance, they underscore the ecological variability of the community and the occurrence of sporadic settlement events.
Figure 6.
Redundancy analysis (RDA) biplot.
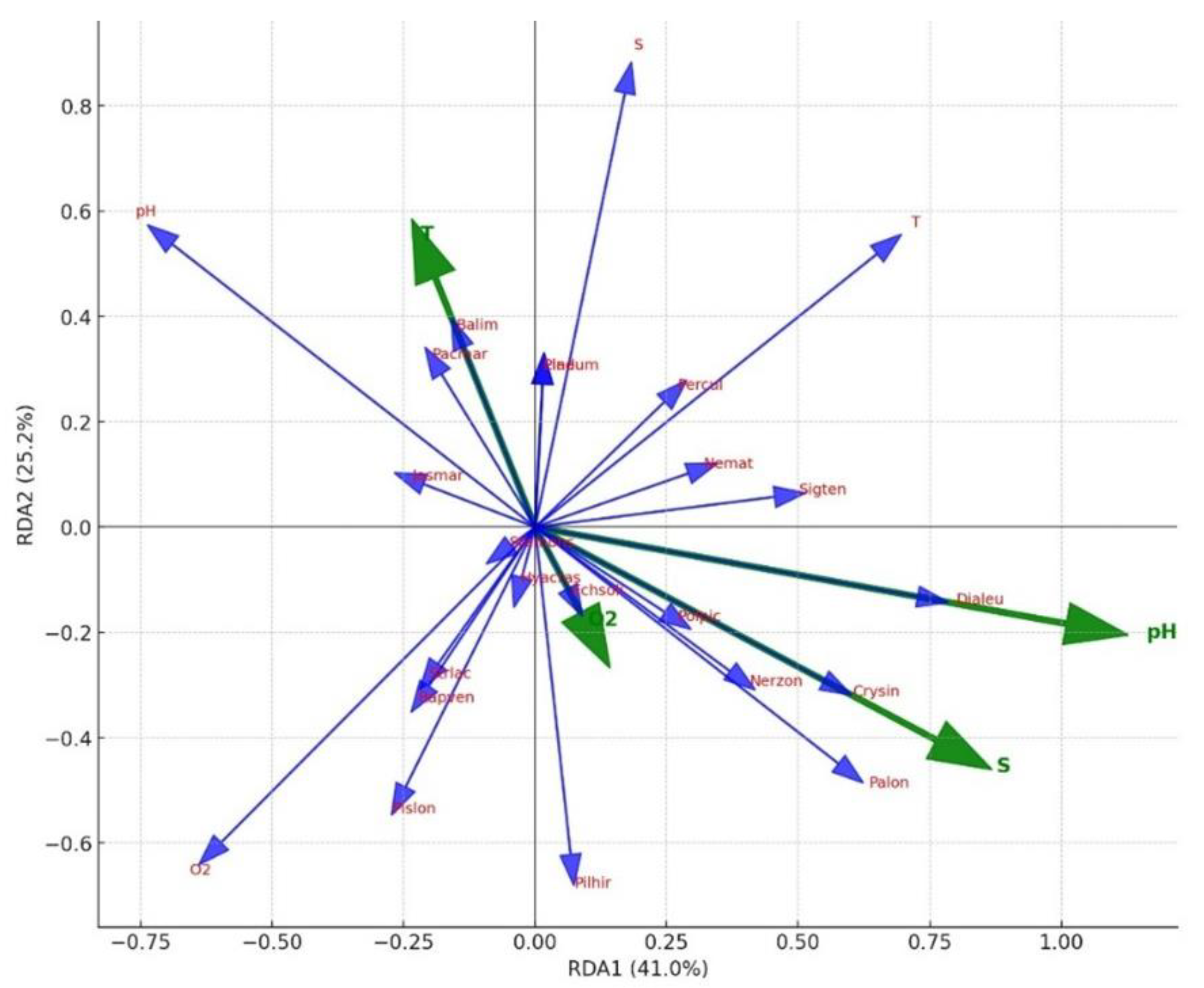
The RDA biplot illustrated the relationships between species and environmental variables (pH, dissolved oxygen, temperature, and salinity) (Fig.6). These findings are of considerable importance for understanding the influence of environmental variables on species distribution and habitat preferences.
Figure 7.
PCA Biplot Showing the Relationship Between Seasonal Distribution and Environmental Variables.
Figure 7.
PCA Biplot Showing the Relationship Between Seasonal Distribution and Environmental Variables.
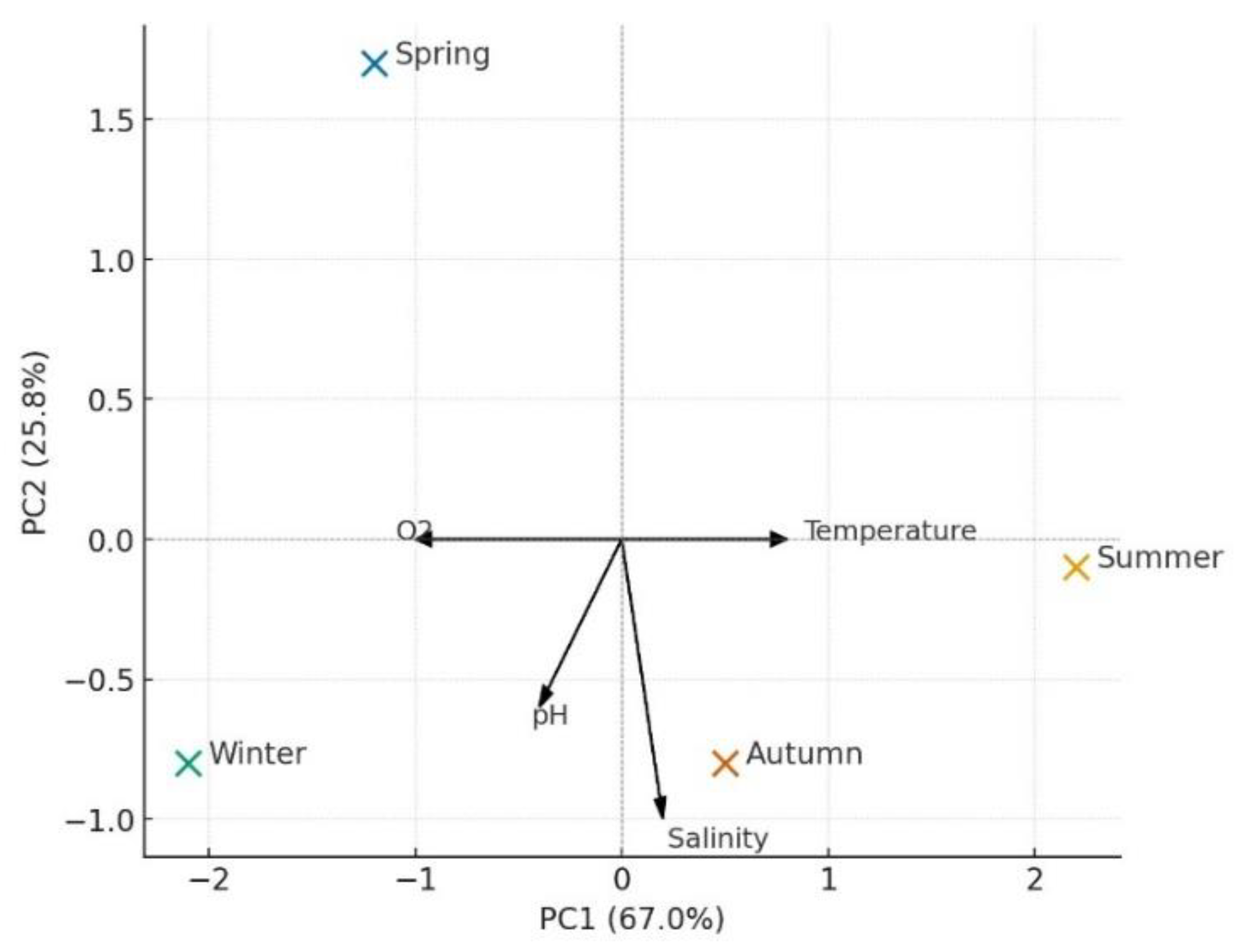
In the PCA (RDA proxy) biplot, temperature and salinity were positively correlated with summer samples, whereas pH and dissolved oxygen were associated with winter and spring samples. The biplot clearly illustrates that macroinvertebrate communities are seasonally structured in response to environmental factors. Samples collected during summer were associated with higher temperature and salinity values, whereas spring and winter samples were linked to higher dissolved oxygen and pH levels (Figure 7). This pattern indicates that community structure beneath mussel aquaculture is strongly influenced by seasonal hydrographic variations.
Overall, the offshore macrofaunal communities exhibited strong seasonality, driven primarily by a few opportunistic and structurally dominant species. Species richness and abundance were not evenly distributed throughout the year but instead followed distinct seasonal patterns shaped by environmental conditions and the life-history strategies of constituent taxa.
These findings support the hypothesis that: "Offshore longline mussel aquaculture systems create reef-like structures that promote seasonally variable yet generally enhanced species richness, evenness, and biomass within macroinvertebrate communities. These structures also influence the distribution of taxa particularly amphipods and mollusks that are sensitive to changes in environmental variables such as temperature, pH, and oxygen."
Table 2.
Species identified in the study (*RDA: Species abbreviations used in Redundancy Analysis; SUM: Total number of individuals; %D: Dominant species by ,relative abundance).
Table 2.
Species identified in the study (*RDA: Species abbreviations used in Redundancy Analysis; SUM: Total number of individuals; %D: Dominant species by ,relative abundance).
| September-2023 | October | November | December | January | February | March | April | May | June | July | August-2024 | SUM | %D | |||||||||||||||
| RDA * | S1 | S2 | S3 | S4 | S5 | S 6 | S7 | S 8 | S9 | S10 | S11 | S12 | ||||||||||||||||
| CRUSTACEA | ||||||||||||||||||||||||||||
| Decapoda | ||||||||||||||||||||||||||||
| Pachygrapsus marmoratusPacmar | 6 | 0 | 2 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 12 | 0,0120 | ||||||||||||||
| Palaemon longirostris Pallon | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 4 | 0,0040 | ||||||||||||||
| Pilumnus hirtellus Pilhir | 0 | 0 | 9 | 0 | 0 | 4 | 2 | 0 | 0 | 7 | 0 | 0 | 22 | 0,0221 | ||||||||||||||
| Pisidia longicornis Pislon | 0 | 0 | 4 | 0 | 4 | 5 | 1 | 0 | 0 | 1 | 0 | 1 | 16 | 0,0160 | ||||||||||||||
| Amphipoda | ||||||||||||||||||||||||||||
| Stenothoe monoculoides Stemon | 1900 | 5000 | 5290 | 1040 | 500 | 2000 | 2000 | 2000 | 4520 | 1000 | 1006 | 1470 | 27726 | 27,8041 | ||||||||||||||
| Echinogammarus oliviEcholi | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 4 | 0 | 0 | 5 | 13 | 0,0130 | ||||||||||||||
| Hyale crassipes Hyacra | 0 | 0 | 0 | 0 | 10 | 30 | 2 | 0 | 30 | 0 | 10 | 20 | 102 | 0,1023 | ||||||||||||||
| Jassa marmorata Jasmar | 5800 | 13250 | 8160 | 5530 | 2100 | 4000 | 6000 | 10000 | 6600 | 2500 | 2024 | 5200 | 71164 | 71,3645 | ||||||||||||||
| CİRRİPEDİA | ||||||||||||||||||||||||||||
| Balanus improvisus Balimp | 7 | 0 | 16 | 2 | 1 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 29 | 0,0291 | ||||||||||||||
| MOLLUSCA | ||||||||||||||||||||||||||||
| Striarca lactea Strlac | 0 | 0 | 2 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0,0040 | ||||||||||||||
| Rapana venosa Rapven | 0 | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 3 | 0,0030 | ||||||||||||||
| ANNELIDAE | ||||||||||||||||||||||||||||
| Polychaetes | ||||||||||||||||||||||||||||
| Nereis zonata | Nerzon | 6 | 2 | 57 | 16 | 47 | 0 | 20 | 13 | 53 | 41 | 12 | 100 | 367 | 0,3680 | |||||||||||||
| Perinereis cultrifera | Percul | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 3 | 0,0030 | |||||||||||||
| Platynereis dumerilii | Pladum | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0,0050 | |||||||||||||
| Sigambra tentaculata | Sigten | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 15 | 17 | 0,0170 | |||||||||||||
| Polyophthalmus pictus | Polpic | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 1 | 7 | 0,0070 | |||||||||||||
| Nemertea | ||||||||||||||||||||||||||||
| Lineus sp. | Lineus | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0,0030 | |||||||||||||
| Platyhelminthes | ||||||||||||||||||||||||||||
| Cryptocelis sinopae | Crysin | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 11 | 5 | 3 | 0 | 19 | 0,0191 | |||||||||||||
| CHİNİDARİA | ||||||||||||||||||||||||||||
| Anemone | ||||||||||||||||||||||||||||
| Diadumene leucolena | Diaieu | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 13 | 45 | 14 | 3 | 78 | 0,0782 | |||||||||||||
| NEMATODA | ||||||||||||||||||||||||||||
| Nematod sp. | Nemato | 0 | 15 | 0 | 0 | 0 | 0 | 0 | 0 | 68 | 0 | 0 | 42 | 125 | 0,1254 | |||||||||||||
3.4. Abundance and Species Richness
Macrofaunal individual abundance peaked in autumn, whereas the lowest values were recorded during summer. In contrast, species richness reached its maximum in summer, with a total of 15 taxa identified during this period (Figure 8).
The monthly and seasonal patterns of species richness and individual abundance (Figure 8) clearly reveal the temporal dynamics of macrofaunal communities in the investigated offshore mussel aquaculture system. These patterns appear to be shaped by both seasonal variations in environmental conditions and the life cycle characteristics of the taxa present.
Figure 8.
Monthly and seasonal distribution of species abundance.
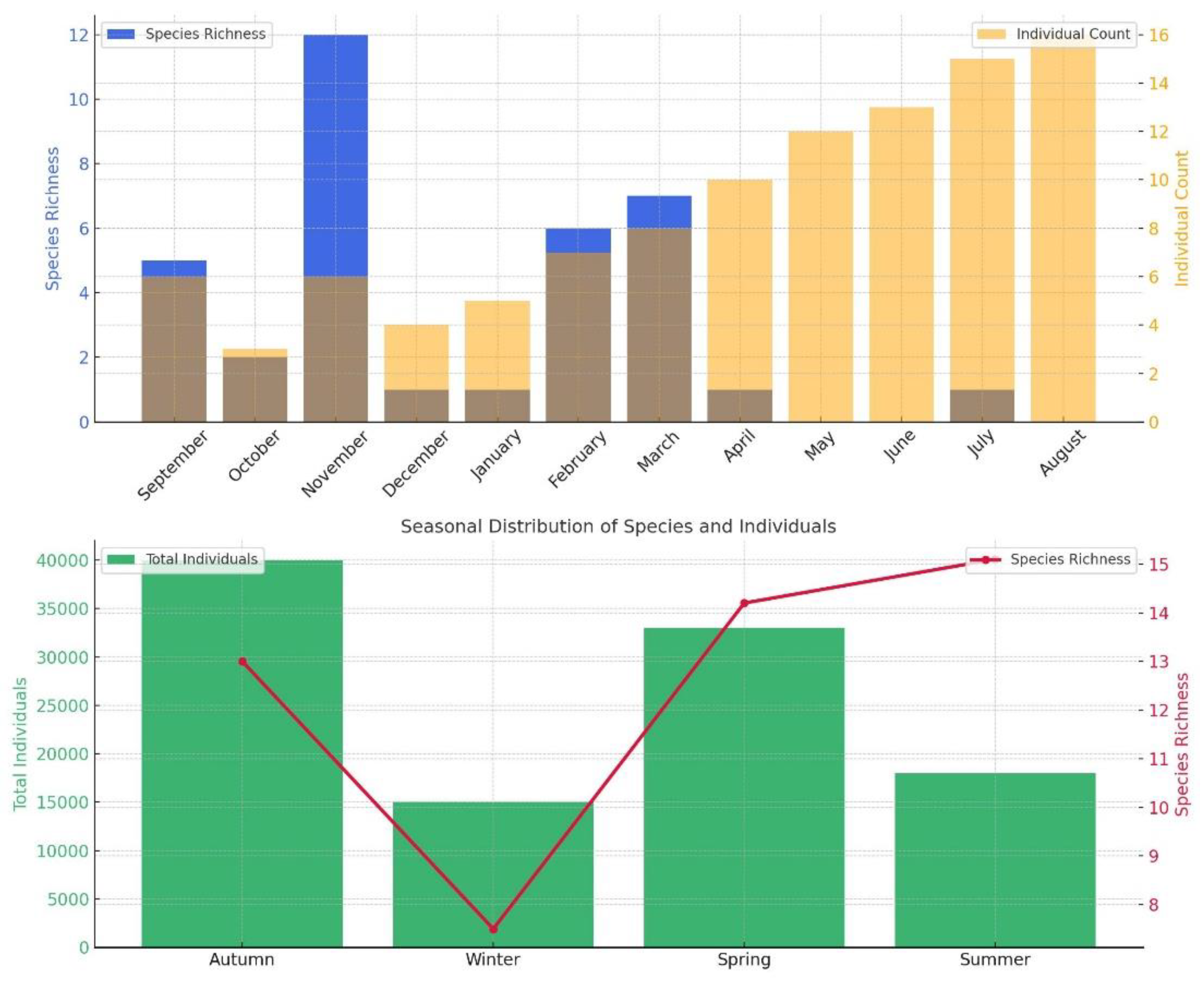
During summer and early autumn (June–September), a marked increase in both species richness and abundance was observed. This seasonal peak coincides with periods of elevated temperatures and reduced hydrodynamic impact, indicating enhanced macrofaunal productivity. The persistent dominance of J. marmorata during these months reflects the species’ rapid colonization capacity and competitive advantage on artificial structures. In contrast, the lower and more stable abundances recorded in winter and spring are consistent with suppressed benthic activity under low temperatures and high oxygen conditions.
Total abundance reached its highest level in autumn, potentially reflecting a dispersal phase of dominant taxa following their summer reproductive period. Conversely, winter was characterized by a pronounced decline in both richness and abundance, likely driven by stress factors such as low temperatures, high dissolved oxygen concentrations, and limited organic matter availability.
Figure 9.
Jassa marmorata (a) and Stenothoe monoculoides (b).

The dominant species, J. marmorata Holmes, 1905 and S. monoculoides (Montagu, 1813) (Figure 9), largely shaped the overall temporal trends, while several low-abundance taxa such as B. improvisus, C. sinopae, H. crassipes, and N. zonata contributed to short-term increases in richness during specific sampling periods (e.g., S4, S9). Although these transient fluctuations had little effect on total abundance, they highlight the ecological variability of the community and the occurrence of episodic settlement events.
In conclusion, offshore macrofaunal communities exhibited strong seasonality, driven primarily by a few opportunistic and structurally dominant species. Species richness and abundance were not evenly distributed throughout the year; rather, they followed a pronounced seasonal pattern determined by environmental conditions and the life-history strategies of the taxa.
4. Discussion
4.1. Findings on the Molluscan Fauna of the Black Sea and the Contribution of This Study
The Black Sea, a semi-enclosed inland sea, harbors a permanent anoxic layer starting from depths of 150–200 m. This condition, driven by the presence of hydrogen sulfide (H₂S), limits the development of deep-sea benthic fauna [38,39]. Despite this limitation, the phylum Mollusca, particularly the classes Bivalvia and Gastropoda, plays a significant role within the region’s macroinvertebrate communities, ranking second in abundance after Arthropoda and first in terms of species richness [40,41,42].
However, studies on the molluscan species of the Black Sea along the Türkiye coast are scarce and scattered. Much of the existing literature is outdated or focused on the northern coasts of the basin (Russia, Romania, Ukraine) [3,42]. Surveys along the Türkiye coastline are mostly based on historical records and often lack contemporary ecological context [9,24,43].
In this context, our study fills a critical gap by providing up-to-date data on molluscan fauna through systematic sampling in offshore mussel aquaculture areas. In particular, the dominance of species such as M. galloprovincialis Lamarck, 1819, Bittium reticulatum (da Costa, 1778), and Anadara inaequivalvis (Bruguière, 1789) in artificial habitats aligns with previously reported distribution patterns along the Black Sea coasts [43,44]. Among these, A. inaequivalvis, as an invasive species, can significantly influence local benthic community dynamics, a phenomenon also emphasized by [3].
RDA analyses revealed that environmental variables such as dissolved oxygen, pH, and substrate type are primary determinants of molluscan distribution [30]. These findings support the high environmental sensitivity of molluscan taxa and their potential use as indicators of habitat quality.
Beyond compositional aspects, species such as M. galloprovincialis contribute functionally by enhancing habitat complexity, stabilizing sediments, and filtering the water column [45]. However, the spread of invasive species like A. inaequivalvis requires monitoring due to potential long-term impacts on aquaculture systems.
4.2. Abundance and Dominance Patterns of Macroinvertebrates
[3]; demonstrated that mussel aquaculture does not negatively affect benthic fish and macroinvertebrates. Considering that species such as flounder, which live in close association with bottom sediments, are sensitive to pollutants, a reduction in their abundance would be expected if aquaculture operations significantly contributed to pollution [55]. However, the abundance of commercially important species such as Pseudopleuronectes americanus was found to be similar directly under mussel lines and at distant reference sites. Interestingly, these species were reported at lower densities in seagrass beds.
Previous studies have emphasized that the effects of artificial structures on sensitive ecosystems such as lagoons, bays, and estuaries depend on location and natural habitat type. Mussel farms are often established on sandy bottoms, where the addition of hard structures may prevent the full preservation of natural benthic community structures. Nevertheless, it is possible to minimize changes in species distribution and productivity.
In this study, J. marmorata (71.36%) was identified as the dominant species in offshore mussel aquaculture systems in the Black Sea, using mussel beds as shelter and food sources (Franz & Mohamed, 1989; Caine, 1983). Similarly, S. monoculoides plays a significant role in supporting amphipod diversity on biogenic substrates [49].
Macroinvertebrate abundance peaked in autumn, while species richness reached its highest level in summer. This pattern can be explained by the accumulation of organic matter in autumn supporting heterotrophic species [43,44], while more stable environmental conditions (temperature, oxygen, pH) in summer promote higher species diversity [30]. High Shannon (~0.72) and Simpson (~0.42–0.43) values during summer indicate that environmental stability supports diversity. Conversely, low Shannon (~0.60) and Simpson (~0.37) values in winter suggest environmental stress linked to lower temperature and dissolved oxygen [50].
These ecological patterns were further confirmed through multivariate analyses. NMDS revealed clear seasonal differences in macroinvertebrate composition, with summer samples clustering distinctly due to higher diversity and evenness. Notably, non-dominant species were more prevalent in summer, contributing to compositional diversity. The PCA biplot showed that summer samples were strongly associated with temperature and salinity, whereas winter samples aligned with dissolved oxygen and pH, indicating lower diversity and higher individual abundance. This supports the link between autumn abundance peaks and organic matter accumulation, and summer richness peaks with environmental stability.
The high Pielou’s evenness values observed in winter and summer suggest that dominant species (J. marmorata and S. monoculoides) possess broad environmental tolerance. The repeated appearance of these species in seasonal NMDS and PCA clusters highlights their potential as bioindicator species for ecosystem monitoring. Previous studies have also emphasized their tolerance to environmental variability in the Black Sea benthic systems [43,50].
Offshore mussel aquaculture systems function as artificial reef-like habitats, enhancing biodiversity. However, these benefits can only be sustained under appropriate environmental conditions and management practices. Low diversity and evenness values (S1 and S4 sampling periods) may indicate habitat degradation or stress. In this context, macrozoobenthic communities are used as biological quality elements in environmental monitoring frameworks such as the EU Water Framework Directive.
The observed seasonal pattern higher abundance in autumn and higher species richness in summer demonstrates that these artificial habitats support biodiversity and functional cycles year-round.
4.3. Environmental Drivers of Community Structure
Among the environmental variables studied, pH and salinity emerged as key factors influencing macroinvertebrate community composition. RDA results indicated that pH (41.0%) and salinity (34.5%) explained the largest proportion of variation in species distribution. Species such as Dialeu, Crysin, Palon, and Nerzon were associated with higher pH and salinity, indicating their stenotopic nature. Conversely, species like Pacmar, Jasmar, and Rapven were associated with lower pH and dissolved oxygen levels, suggesting tolerance to stressful conditions.
Seasonal changes in temperature influenced the presence of temperature-sensitive species such as Stemons and Pladum. J. marmorata Holmes, 1905 demonstrated eurytopic traits, being resilient to various stressors, and dominated across seasons. These patterns highlight the importance of integrating abiotic and biotic interactions in understanding benthic community dynamics.
PCA (RDA proxy) analysis indicated that increased temperature and salinity during summer had the strongest influence on species distribution. In contrast, pH and dissolved oxygen were more influential in winter and spring, shaping distinct community structures under colder and oxygen-rich conditions. NMDS analysis supported these findings, with the most pronounced compositional difference occurring between summer and autumn (Bray–Curtis= 0.499). This suggests that even in the absence of statistically significant differences in species richness, seasonal turnover is evident [13,23].
4.4. Management Implications for Mussel Aquaculture
While mussel aquaculture creates artificial habitats that enhance local biodiversity, it also introduces challenges such as space and resource competition with pecies like B. improvisus Darwin, 1854 [51]. This issue becomes particularly prominent during larval settlement periods, potentially hindering mussel growt. Therefore, larval collector deployment should be carefully timed. To prevent settlement by Balanus larvae, which peak during July–August [52], collectors should be deployed in May, and surface cleaning strategies should be implemented to reduce colonization.
High macroinvertebrate biomass and diversity have been documented in tidal mussel aggregations [53,54]. Cultivated bivalves, particularly suspended mussel farming [55,56], as well as both surface and bottom oyster culture systems [57,58], have been shown to support rich macroinvertebrate assemblages. The structures used in bivalve aquaculture provide favorable habitat conditions for macroinvertebrate species and are increasingly recognized as artificial reef systems that support not only macroinvertebrates but also larger marine fauna and fish [56,59]. Moreover, macroinvertebrate communities associated with suspended bivalve farming are generally considered a functional component of the benthic environment. Therefore, integrating data from both sediment-dwelling macroinvertebrates and those associated with mussel socks can yield a more comprehensive understanding of the effects of bivalve aquaculture on benthic ecosystems [60,61].
Although the current study recorded Rapana venosa (Valenciennes, 1846) at low densities, its known predatory behavior on mussels [62] suggests potential long-term effects on both mussel populations and associated macroinvertebrate communities. Future studies should incorporate targeted assessments of predator-prey interactions between R. venosa (Valenciennes, 1846), M. galloprovincialis Lamarck, 1819, and dominant amphipod species (e.g., J. marmorata Holmes, 1905), especially given the ecological and economic importance of mussel aquaculture in the Black Sea.
In shellfish aquaculture, ecological carrying capacity is mostly considered in terms of “negative” impacts, typically associated with issues such as water quality degradation, sediment accumulation beneath farms, benthic enrichment, and effects on surrounding habitats [63]. Conversely, there is limited research exploring aquaculture sites as potential novel habitats that could enhance the abundance and productivity of marine organisms [45,55]. In addition, macroinvertebrate based bioindicator programs should be integrated into aquaculture monitoring strategies. Species with stenotopic traits can serve as early warning indicators of environmental stress. Understanding the ecological functions of dominant species can guide habitat specific management practices.
Overall, the findings underscore the need for adaptive management that considers both production goals and ecological sustainability. Mussel aquaculture systems should be evaluated not only in terms of yield but also as dynamic ecosystems supporting diverse and responsive benthic communities.
5. Conclusions
Invertebrates inhabiting mussel beds contribute to nutrient cycling and ecological balance by consuming plankton, detritus, and microbial matter. In the Black Sea, benthic invertebrates play a critical role in maintaining ecosystem health. Mussel beds dominated by M. galloprovincialis form structured habitats that provide shelter and feeding grounds. Amphipods, in particular, find refuge among mussel shells and feed on organic matter.
This study assessed the ecological roles and biodiversity contributions of macroinvertebrates associated with M. galloprovincialis cultivated in offshore longline systems in the central Black Sea. Over a one-year monitoring period, J. marmorata emerged as the dominant species. Species richness peaked in summer, whereas individual abundance was highest in autumn.
The findings demonstrate that mussel aquaculture systems function not only as production platforms but also as habitats that support local biodiversity. However, ecological challenges such as competition with B. improvisus can be mitigated through adjustments in timing and culture depth.
In conclusion, offshore mussel aquaculture systems act as artificial reef-like habitats, enhancing benthic biodiversity and contributing to ecosystem resilience. These systems are both productive and ecologically valuable habitats in terms of species richness and sensitivity to environmental gradients. Sustaining habitat-sensitive aquaculture strategies is crucial for maintaining ecological integrity and economic viability.
Author Contributions
The theoretical framework and study design were developed by E.A.Ç. All material preparation, data collection, analysis, manuscript drafting, and final approval were completed solely by the author.
Funding
No funding: grants, or financial support were received for the preparation of this article.
Ethical Statement
The author has read, understood, and complied with the "Authors' Ethical Responsibilities" declaration.
Data Availability Statement
The datasets generated and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
The author expresses sincere gratitude to Sinop University Faculty of Fisheries and Prof. Dr. Meryem Yeşim Çelik for establishing the Mussel Research Facility, which made this study possible.
Conflicts of Interest
The author declares no conflict of interest.
References
- Murray JW, Top Z, Özsoy E. Hydrographic properties and ventilation of the Black Sea. Deep Sea Research Part A: Oceanographic Research Papers 1991, 38, S663–S689. [Google Scholar] [CrossRef]
- Özsoy E, Ünlüata Ü. Oceanography of the Black Sea: A review of some recent results. Earth-Science Reviews 1997, 42, 231–272. [Google Scholar] [CrossRef]
- Zaitsev YP, Mamaev V. Marine biological diversity in the Black Sea: A study of change and decline. New York: United Nations Publications; 1997.
- Mee, LD. The Black Sea in crisis: A need for concerted international action. Ambio 1992, 21, 278–286. [Google Scholar]
- Ivanov L, Bulatov O. Mytilus galloprovincialis populations in the Black Sea. Hydrobiologia 1990, 187, 25–33.
- Aydemir-Çil E, Birinci-Özdemir Z, Özdemir S. First find of the starfish Asterias rubens Linnaeus 1758 off the Anatolian coast of the Black Sea (Sinop). Marine Biological Journal 2023, 8, 97–101. [Google Scholar]
- Birinci-Özdemir Z, Aydemir-Çil E, Özdemir S, Duyar HA. An important species for Black Sea biodiversity: Some population characteristics of invasive sea vase (Ciona intestinalis Linnaeus, 1767) distributed on the Sinop shores. Turkish Journal of Zoology 2024, 48, 630–639. [Google Scholar] [CrossRef]
- Altuğ G, Aktan Y, Oral M, Topaloğlu B, Oğuz A. Bacterial and nutrient properties of the Golden Horn Estuary and their relationships with zooplankton species. Turkish Journal of Fisheries and Aquatic Sciences 2011, 11, 309–316. [Google Scholar]
- Çınar ME, Bakır K, Öztürk B, Doğan A, Açik Ş, Kırkım F, et al. Spatial distribution pattern of macroinvertebrates associated with the black mussel Mytilus galloprovincialis (Mollusca: Bivalvia) in the Sea of Marmara. Journal of Marine Systems 2020, 211, 103402. [Google Scholar] [CrossRef]
- Callier MD, McKindsey CW, Desrosiers G. Multi-scale spatial variations in benthic sediment geochemistry and macrofaunal communities under a suspended mussel culture. Marine Ecology Progress Series 2007, 348, 103–115. [Google Scholar] [CrossRef]
- Fabi G, Manoukian S, Spagnolo A. Impact of an open-sea suspended mussel culture on macrobenthic community (Western Adriatic Sea). Aquaculture 2009, 289, 54–63. [CrossRef]
- Borja A, Franco J, Pérez V. A marine biotic index to establish the ecological quality of soft-bottom benthos within European estuarine and coastal environments. Marine Pollution Bulletin 2000, 40, 1100–1114. [Google Scholar] [CrossRef]
- Reiss H, Kröncke I. Seasonal variability of benthic indices: An approach to test the applicability of different indices for ecosystem quality assessment. Marine Pollution Bulletin 2005, 50, 1490–1499. [Google Scholar] [CrossRef]
- Ross AH, Nisbet RM. Dynamic models of growth and reproduction of the mussel Mytilus edulis L. Functional Ecology 1990, 4, 777–787.
- Seed, R. The ecology of Mytilus edulis L. (Lamellibranchiata) on exposed rocky shores: I. Breeding and settlement. Oecologia 1969, 3, 277–316. [Google Scholar] [CrossRef]
- Thompson, RJ. The reproductive cycle and physiological ecology of the mussel Mytilus edulis in a subarctic, non-estuarine environment. Marine Biology 1984, 79, 277–288. [Google Scholar] [CrossRef]
- Koehn RK, Bayne BL. Towards a physiological and genetical understanding of the energetics of the stress response. Biological Journal of the Linnean Society 1989, 37, 157–171. [Google Scholar] [CrossRef]
- Navarrete SA, Castilla JC. Predation by Norway rats in the intertidal zone of central Chile. Marine Ecology Progress Series 1993, 92, 187–199. [Google Scholar] [CrossRef]
- Bellan-Santini, D. Contribution à l’étude des peuplement infralittoraux sur substrat rocheux (Etude qualitative et quantitative de la franch Superiere). Recherche Travaux Station Marine Endoume 1969, 63, 9–294. [Google Scholar]
- Tsuchiya, M. Faunal structures associated with patches of mussels on East Asian coasts. Helgoland Marine Research 2002, 56, 31–36. [Google Scholar] [CrossRef]
- Dittmann, S. Mussel beds: A unique habitat for intertidal fauna. Helgoländer Meeresuntersuchungen 1990, 44, 229–243. [Google Scholar] [CrossRef]
- Karayücel H, Doğan H, Aydın M. The potential for mussel aquaculture in the Black Sea: A preliminary assessment. Turkish Journal of Fisheries and Aquatic Sciences 2002, 12, 95–103. [Google Scholar]
- Çelik, MY. Türkiye’de midye yetiştiriciliği: Potansiyel ve uygulamalar. Ege Journal of Fisheries and Aquatic Sciences 2006, 23, 387–394. [Google Scholar]
- Çınar ME, Katağan T, Öztürk B, Egemen Ö, Ergen Z, Kocataş A, et al. Karadeniz bentik makroomurgasızları. Turkish Journal of Marine Science 2020, 26, 165–182. [Google Scholar]
- Öztürk B, Doğan A, Katağan T, Salman A. Marine molluscs of the Türkiye coasts: A checklist. Istanbul: Türkiye Marine Research Foundation (TUDAV), 2014.
- Robichaud L, Richard M, Gendron L, McKindsey CW, Archambault P. Influence of suspended mussel aquaculture and an associated invasive ascidian on benthic macroinvertebrate communities. Water 2022, 14, 2751. [Google Scholar] [CrossRef]
- Fauchald, K. The polychaete worms: Definitions and keys to the orders, families and genera. Washington DC: Smithsonian Institution Press; 1977.
- Çınar ME, Ergen Z, Dağlı E. Checklist and zoogeographic affinities of polychaetes from the coasts of Türkiye. Zootaxa 2006, 1168, 1–22. [Google Scholar]
- Poppe GT, Goto Y. European seashells. Wiesbaden: Verlag Christa Hemmen; 1991.
- Holthuis, LB. FAO species catalogue Vol. 1: Shrimps and prawns of the world. FAO Fisheries Synopsis No. 125, Vol. 1. Rome: FAO; 1980.
- Ruffo, S. The Amphipoda of the Mediterranean. Monaco: Mémoires de l’Institut Océanographique; 1982–1989.
- WoRMS. World Register of Marine Species. 2024. Available from: https://www.marinespecies.org/.
- OBIS. Ocean Biodiversity Information System. 2024. Available from: https://obis.org/.
- Shannon CE, Weaver W. The mathematical theory of communication, University of Illinois Press: Urbana, 1948.
- Pielou, EC. Ecological diversity. New York: Wiley-Interscience; 1975.
- Zenkevitch, L. Biology of the seas of the U.S.S.R. London: George Allen and Unwin; 1963.
- Bakan G, Büyükgüngör H. The Black Sea. Marine Pollution Bulletin 2000, 41, 24–43. [Google Scholar] [CrossRef]
- Çulha, M. Türomical and ecological characteristics of Prosobranchia (Mollusca-Gastropoda) species distributed around Sinop. [PhD dissertation]. İzmir: Ege University; 2004. 150 p.
- Çulha M, Bat L, Çulha ST, Çelik MY. Benthic mollusk composition of some facies in the upper-infralittoral zone of the southern Black Sea, Türkiye. Turkish Journal of Zoology 2010, 34, 523–532. [Google Scholar]
- Anistratenko VV, Anistratenko OY. Fauna of Ukraine. Vestnik Zoologii 2001, 29, 1–240. [Google Scholar]
- Mutlu E, Betil Ergev M. Spatio-temporal distribution of soft-bottom epibenthic fauna on the Cilician shelf (Turkey), Mediterranean Sea. Revista de Biología Tropical 2008, 56, 1919–1946. [Google Scholar]
- Bat L, et al. Long-term changes and causes of biota assemblages in the southern Black Sea coasts. Marine Biology Research 2024, 20, 251–265. [Google Scholar] [CrossRef]
- Archambault P, Grant J, Brosseau C. Secondary productivity of fish and macroinvertebrates in mussel aquaculture sites. Secondary productivity of fish and macroinvertebrates in mussel aquaculture sites. ICES CM 2008, H, 15. [Google Scholar]
- Bat L, et al. Biological diversity of the Turkish Black Sea coast. Turkish Journal of Fisheries and Aquatic Sciences 2011, 11, 683–692. [Google Scholar]
- Guerra-García JM, Ruiz-Tabares A, Baeza-Rojano E, Cabezas MP, Díaz-Pavón JJ, Pacios I, et al. Trace metals in Caprella (Crustacea: Amphipoda). A new tool for monitoring pollution in coastal areas? Ecological Indicators 2010, 10, 734–743. [Google Scholar] [CrossRef]
- Sezgin M, Çil Aydemir E. Crustacean fauna of a mussel cultivated raft system in the Black Sea. Arthropods 2013, 2, 89–97. [Google Scholar]
- Grintsov, VA. Taxonomic diversity of Amphipoda (Crustacea) from the Black Sea and the Sea of Azov. Marine Biological Journal 2022, 7, 34–45. [Google Scholar] [CrossRef]
- Kolyuchkina GA, et al. The role of abiotic environmental factors in the vertical distribution of macrozoobenthos at the northeastern Black Sea coast. Biology Bulletin 2020, 47, 1126–1141. [Google Scholar] [CrossRef]
- Çınar ME, Dağlı E, Katağan T. Seasonal changes in the soft-bottom polychaete assemblages in the Black Sea. Mediterranean Marine Science 2016, 17, 150–162. [Google Scholar]
- Karayücel İ, Karayücel S, Erdem M. Settlement and growth of the barnacle (Balanus improvisus) on ropes used in mussel cultivation. Turkish Journal of Veterinary and Animal Sciences 2002, 26, 1155–1160. [Google Scholar]
- Zaitsev Y, et al. The Black Sea. Exotic species in the Aegean, Marmara, Black, Azov and Caspian Seas. Istanbul: Turkish Marine Research Foundation; 2001. p. 73–138.
- Commito JA, Dankers N. Dynamics of spatial and temporal complexity in European and North American soft-bottom mussel beds. In: Reise K, editor. Ecological comparisons of sedimentary shores, 2001; 39–59.
- Norling P, Kautsky N. Patches of the mussel Mytilus sp. are islands of high biodiversity in subtidal sediment habitats in the Baltic Sea. Aquatic Biology 2008, 4, 75–87. [Google Scholar] [CrossRef]
- D’Amours O, Archambault P, McKindsey CW, Johnson LE. Local enhancement of epibenthic macrofauna by aquaculture activities. Marine Ecology Progress Series 2008, 371, 73–84. [Google Scholar] [CrossRef]
- McKindsey CW, Archambault P, Callier M, Olivier F. Influence of suspended and off-bottom mussel culture on the sea bottom and benthic habitats: A review. Canadian Journal of Zoology 2011, 89, 622–646. [Google Scholar] [CrossRef]
- Shumway SE, editor. Shellfish aquaculture and the environment, Wiley-Blackwell: Oxford, 2011.
- Forrest BM, Keeley NB, Hopkins GA, Webb SC, Clement DM. Bivalve aquaculture in estuaries: Review and synthesis of oyster cultivation effects. Aquaculture 2009, 298, 1–15. [Google Scholar] [CrossRef]
- Kaiser MJ, Laing I, Utting SD, Burnell GM. Environmental impacts of bivalve mariculture. Journal of Shellfish Research 1998, 17, 59–66. [Google Scholar]
- McKindsey CW, Thetmeyer H, Landry T, Silvert W. Review of recent carrying capacity models for bivalve culture and recommendations for research and management. Aquaculture 2006, 261, 451–462. [Google Scholar] [CrossRef]
- Callier MD, McKindsey CW, Desrosiers G. Influence of suspended mussel lines on the biogeochemical fluxes in the benthic boundary layer. Marine Environmental Research 2013, 83, 10–18. [Google Scholar]
- Crawford CM, Macleod CK, Mitchell IM. Effects of shellfish farming on the benthic environment. Aquaculture 2003, 224, 117–140. [Google Scholar] [CrossRef]
- Govorın, I A. The predatory marine gastropod Rapana venosa (Valenciennes, 1846) in Northwestern Black Sea: morphometric variations, imposex appearance and biphallia phenomenon. In: Molluscs. IntechOpen, 2018.
- Tıčına, V; Katavıć, I; Grubıšıć, L. Marine aquaculture impacts on marine biota in oligotrophic environments of the Mediterranean Sea–a review. Frontiers in marine science 2020, 7, 217. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



