
Submitted:
26 August 2025
Posted:
27 August 2025
You are already at the latest version
Abstract
[Background] Nettle is a potential non-conventional feed resource due to its high level of crude protein content, and ensiling is better for utilization in the animal industry. Previous integrated analysis (microbiome and metabolome) suggested that 4-pentenoic acid and malic acid in ensiled nettle may inhibit harmful microorganisms within the system. The present study investigated the effects of these two acids on nettle silage quality through the addition of 1% fresh weight of these acids then analyzed the characteristics and bacterial community during 60 days of nettle ensiling. [Results] Addition with 4-pentenoic acid increased the content of crude protein (CP) and acetic acid (AA) when compared with both control and malic acid treated during 30 to 60 days of ensiling (p<0.05). Lactic acid (LA) content was highest in malic acid treated (4.21 %, dry matter, DM based) when compared with control and 4-pentenoic acid treated after 7 days of nettle ensiling (p<0.05), but lower when compared with 4-pentenoic acid treated after 30 days of nettle ensiling (p<0.05), and was not detected in all groups after 60 days of silage. The content of butyric acid (BA) and ammonia (AN) was lowest (2.92%—4.39% of DM and 9.94 % —24.28% of total nitrogen, respectively) in 4-pentenoic treated when compared with both control and malic acid treated during 30 to 60 days of ensiling (p<0.05). Both acids treated increased relative abundance of Weissella after 30 days of nettle ensiling, especially for 4-pentenoic acid showed the higher capacity. Both acids treated showed trend to inhibited relative abundance of Clostridium sensu stricto 15 after 30 days of nettle ensiling. Clostridium sensu stricto 15 showed significant positive correlation with BA and AN (p<0.05). [Conclusion] Present study results suggested that addition of 4-pentenoic could improve quality of silage due to reduction level of protein degradation probably resulting from its inhibited activity against to Clostridium spp. However, malic acid was less effective than 4-pentenoic acid in suppressing Clostridium spp. activity and the associated production of BA and AN, resulting in inferior preservation of CP.
Keywords:
nettle
; 4-pentenoic acid
; malic acid
; silage
; bacteria
1. Introduction
The rapid expansion of the livestock feed industry in recent years, has caused the critical shortages of conventional protein sources(e.g., corn and soybean meal), which now fail to meet market demand. Developing non-conventional feed sources now constitutes an imperative necessity for ensuring sustainable livestock production systems [1].
Nettles (Urtica spp.) are annual or perennial herbaceous plants belonging to the family Urticaceae. Characterized by high nutritional value, robust environmental adaptability, and strong stress resistance, they exhibit widespread global distribution [2]. Furthermore, nettles are rich in essential nutrients including proteins, carbohydrates, lipids, and amino acids, as well as biologically active compounds such as carotenoids terpenoids, α-linolenic acid, 12-linoleic acid, vitamins, cis-9, and polyphenolic compounds [3]. In addition, these plants possess demonstrate antioxidant, antibacterial, and anti-inflammatory properties, positioning them as a highly promising alternative feed resource. Recent research demonstrates the significant potential of nettle species as functional feed ingredients for optimizing animal production, enhancing stress resilience, and improving the quality of animal-derived products across livestock and poultry species. Specifically, partial alfalfa (Medicago sativa) replacement with Nettles enhanced antioxidant status in dairy cows [4], dietary supplementation ameliorated heat stress in broilers [5], and inclusion improved egg yolk carotenoid deposition in laying hens [6]. Similarly, Boehmeria nivea hay substitution boosted rabbit productivity [7]."
Similar to alfalfa, nettles exhibit constrained silage characteristics by several intrinsic factors [8]. Their elevated crude protein content and high buffering capacity, coupled with a concomitantly low water-soluble carbohydrate (WSC) content, result in insufficient fermentable substrate, thereby restricting the proliferation of essential lactic acid bacteria (LAB) during the silage process [9]. Furthermore, the abundant protein serves as a readily available nitrogen source promoting the growth of undesirable Clostridia species. Subsequent proteolysis by Clostridium spp. generates ammonia (AN), which neutralizes lactic acid (LA) and impedes the critical decline in system pH [10]. This disruption of the acidification process causes ineffective silage fermentation.
Our previous study showed that malic acid enhances silage Lactobacillus growth through specialized metabolic utilization and competitive dominance [3]. Lactobacillus spp. express mleS and mic genes encoding malolactic enzyme and malic enzyme, enabling direct decarboxylation of malic acid to LA via malolactic fermentation (MLF) [3]. Concurrently, malic acid is funneled into pyruvate metabolism (pdhB, ldh upregulation) and glycolysis, sustaining energy production and biosynthetic precursors [11]. Additionally, naturally occurring antimicrobial compounds within nettles, such as 4-pentenoic acid, formic acid, and phenolic compounds, exhibit broad-spectrum inhibitory activity against microbial growth. 4-Pentenoic acid may act as a potential spoilage factor in nettle silage, as it demonstrated inhibitory effects against beneficial LAB (e.g., Pediococcus pentosaceus) in vitro through impairing fatty acid synthesis and cell membrane integrity, which could contribute to the observed quality deterioration [11]. Whereas prior microbiome analyses identified 4-pentenoic acid and malic acid as key fermentation inhibitors in nettle silage [3,11], their targeted application as antibacterial additives against Clostridium remains unexplored. To address this knowledge gap, we added these two acids in nettle silage in order to determined how these acids are involved in the fermentation process in the present study.
2. Materials and Methods
2.1. Silage Prepartion
Nettle (Urtica cannabina) was harvested on 10th September 2023 from the wild in Shawan County, central mountains of Tianshan, Xinjiang, China (E 84°58′—86°24′; N 43°26′—45°20′). After air-drying to 394.2g·kg−1 fresh weight, plants were cut into 2 cm segments using a fodder chopper. Approximately 1.0 kg of homogenized material was vacuum-sealed in polyethylene bags (23 cm× 30 cm) fitted with one-way valves. Established two treatments, addition with 1% of fresh weight for malic acid (MA, BioUltra grade, Macklin Biochemical Co., Ltd., Shanghai, China; Product No. M815619) and 4-pentenoic acid (4PA, ≥ 97 % purity, Aladdin Biochemical Technology Co., Ltd., Shanghai, China; Product No. P112354), respectively, based on our previous research demonstrating the efficacy of 1% of fresh weight in modulating microbial activity and fermentation in nettle silage, particularly for suppressing undesirable Clostridial [11]. Control groups were also prepared. Each treatment comprised nine replicates, totaling 45 sample bags stored at 24 ℃ for 60 days.
2.2. Characteristics Analysis of Nettle Silage
After 7, 30 and 60 days of ensiling, 200 g silage samples were dried at 65℃ for 48 h and ground to pass through a 1.0-mm sieve, and then used to determine DM content. Total nitrogen (TN) was determined using an automatic Kjeldahl nitrogen analyzer (K9840, Hanon Co. Ltd, Shandong, China), and crude protein was calculated according to the method of the Association of Official Agricultural Chemists (AOAC). WSC content was determined according to methods describe above [12].
A 20-g silage sample from each treatment was combined with 180 mL of deionized water to assess fermentation characteristics. The mixture (1:9 for v/v) was filtered through four layers of cheesecloth. The pH was immediately measured with a portable pH meter (PHS-3C, Instrument and Electrical Science Instrument Co., Ltd., Shanghai, China), and the supernatant was retained for ammonia and organic acid analyses following established protocols [3,13]. Organic acids (actic acid, LA; acetic acid, AA; propionic acid, PA; butyric acid, BA) were quantified via HPLC employing a C18 column (150 × 4.6 mm, 94 FMF-5559-EONU, FLM Scientific Instrument Co., Ltd., Guangzhou, China). The Na2HPO4 (1 mM, pH 2.7) and methanol were used for mobile phase A and B, respectively, with a flow rate of 0.6 mL·min−1. Separation used a 20 μL injection volume at 50 ℃ with the gradient: 0 min, 100 % A; 5—40 min, 98 % A.
2.3. Sequencing Analysis of Bacterial Community in Nettle Silage
Total DNA was extracted from triplicate samples per treatment group after 7, 30, and 60 ensiling days using the commercial DNA Kit (FastDNA® Spin Kit for Soil, MP Biomedicals, USA). Primers (338F: ACTCCTACGGGAGGCAGCAG; 806R: GGACTACHVGGGTWTCTAAT) targeting the V3—V4 regions of 16S rDNA were used for PCR amplification. The amplicons were extracted, purified, and the raw sequences analyzed [3]. The sequences were uploaded to the National Center for Biotechnology Information (NCBI, https://www.ncbi.nlm.nih.gov/): PRJNA1233857 (accessed on March 13, 2025) for addition with 4-pentenoic acid and PRJNA1236718 (accessed on March 16, 2025) for addition with malic acid into nettle silage, respectively.
2.4. Statistical Analysis
Data from the three silage groups underwent one-way ANOVA in SPSS 22 (SPSS Inc., Chicago, IL, USA), with Tukey's test (p < 0.05) determining treatment differences. Microbiota bioinformatics utilized the Majorbio Cloud platform (https://cloud.majorbio.com).
3. Results
3.1. Effect of 4-Pentenoic Acid and Malic on Characteristics of Nettle Silage
The characteristics of nettle after wilting were 39.42 % fresh weight of dry matter, 16.89 % DM of crude protein, 7.50 % DM of WSC and 8.04 of pH, respectively. As show in table 1, DM was highest in MA treated than control and 4PA after 30 days of nettle ensiling (p < 0.05). CP was lowest in malic acid treated than control and 4-pentenoic acid after 7 days of nettle ensiling (p < 0.05); highest in 4PA treated, followed by control during 30—60 days of nettle ensiling. 4PA treated showed highest pH compared with control and MA treated after 7 days of nettle ensiling (p < 0.05), while it showed lowest pH after 30—60 days of ensiling (p < 0.05).
As shown in , DM was highest in MA treated when compared with control and 4PA treated after 30 days of ensiling (p<0.05). The content of CP was higher in 4PA treated than MA treated after 7 days of ensiling (p<0.05), and showed highest content in 4PA treated than other two groups during 30—60 days of ensiling (p<0.05). The content of WSC showed none difference between control and each two treated groups during entire ensiling process (p>0.05). The pH value (7.98) was highest in 4PA treated after 7 days of ensiling, but lower during 30—60 days of ensiling when compared with control and MA treated (p<0.05). The content of LA was highest in MA treated after 7 days of ensiling when compared with control and 4PA treated (p<0.05), but lower after 30 days of ensiling when compared with 4PA treated (p<0.05), and lowest in control after 30 days of ensiling when compared with other two treated groups (p<0.05). The content of AA was highest in 4PA treated during 30—60 days of ensiling when compared with control and MA treated (p<0.05). The content of PA was highest in control when compared with two treated groups after 60 days of ensiling (p<0.05). The content of BA was lowest in 4PA treated during 30—60 days of ensiling when compared with control and MA treated (p<0.05). Ammonia was lower in 4PA treated compared with control and MA treated (p<0.05) after 30 days of ensiling, lowest in 4PA treated compared with control and MA treated (p<0.05) after 60 days of ensiling.
3.2. Effect of 4-Pentenoic Acid and Malic on Bacteria Community of Nettle Silage
As shown in Figure 1A,B, both chao 1 and ace index showed none difference between each treated groups and control (p>0.05). Shannon index was highest in MA treated after 60 days of nettle ensiling when compared with 7 days (p<0.05, Figure 1C).
As shown in Figure 2, there are clear distance between control and other acids treated groups after 30 days of nettle ensiling.
As shown in Figure 3A, Firmicutes were the most dominant bacteria among control and each treated groups, followed by Cyanobacteria and Proteobacteria during 60 days of nettle ensiling, and none significant difference was observed among each groups (p>0.05).
As shown in Figure 3B, Aerococcus (50.62 %—61.52 %) was the dominant bacteria both control and two acids treated groups after 7 days of nettle ensiling, followed by Enterococcus (5.66 %—12.32 %) and Jeotgalibaca (2.51 %—9.25 %), no significant difference was observed for these bacteria among each groups (p>0.05). Aerococcus (20.56 %—41.07%) was dominant bacteria in both control and two acids treated groups after 30 days of nettle ensiling, followed by Irregularibacter (2.50 %—39.78 %) and Atopostipes (8.04 %—11.67 %), none difference was observed for these bacteria among each groups (p>0.05). Desemzia showed the significant highest relative abundance in 4PA treated when compared with control and MA treated, and control was higher than malic treated after 30 days of nettle ensiling (relative abundant were 3.50 %, 2.43 % and 4.16% for control, MA treated and 4-pentenoic acid, respectively, p<0.05). Weissella showed significant highest relative abundant in 4PA treated when compared with control and MA treated, and MA treated was higher than control after 30 days of nettle ensiling (relative abundant were 0.64 %, 1.41 % and 2.55 % for control, MA treated and 4-pentenoic acid, respectively, p<0.05). Aerococcus (22.94 %—24.62 %) was dominant bacteria in both control and two acids treated groups after 60 days of nettle ensiling, followed by Atopostipes (15.53 %—22.45 %) and Irregularibacter (9.37 %—21.30 %), none difference was observed for these bacteria among each groups (p>0.05). Tissierella showed significant highest relative abundant in control than other two treated groups, MA treated was higher than 4PA treated after 60 days of nettle ensiling (relative abundant were 6.85 %, 2.30 % and 0.08 % for control, MA treated and 4-pentenoic acid, respectively, p<0.05).
As shown in Figure 4, Jeotgalibaca, Lactococcus and Aerococcus were the most significant difference bacteria in control, 4-pentenoic acid and MA treated groups, respectively, after 7 days of nettle ensiling (p<0.05). Facklamia and Rubellimicrobium were the most significant difference bacteria in 4-pentenoic acid and MA treated groups, respectively, after 30 days of nettle ensiling (p<0.05). Atopostipes and Tissierella were the most significant difference bacteria in control and MA treated groups, respectively, after 60 days of nettle ensiling (p<0.05).
3.3. Correlation Analysis Between Bacteria Community and Fermentation Characteristics in Nettle Silage
As shown in Figure 5, Tissierella, Anaerosporobacter, Irregularibacter, Tepidimicrobium, Atopostipes and Clostridium sensu stricto 15 showed significant positively correlation with ammonia nitrogen, BA and pH; while Aerococcus, Lactococcus, Carnobacterium and Enterococcus showed significant negative correlation with each characteristics (p<0.05). Tissierella, Anaerosporobacter, Irregularibacter, Tepidimicrobium, Atopostipes and Clostridium sensu stricto 15 showed significant negatively correlation with WSC, CP, LA and AA; while Aerococcus, Lactococcus, Carnobacterium and Enterococcus showed significant positive correlation with each characteristics (p<0.05).
4. Discussion
4.1. Effect of 4-Pentenoic Acid and Malic on Characteristics of Nettle Silage
While our prior research [14] focused on optimizing nettle ensiling using LAB inoculants. This experimental design builds directly on previous work, which identified 4PA and MA as key limiting factors in nettle silage fermentation. Critically, 4PA actively inhibits LAB activity [3, 11]; the current study extends these findings by evaluating plant-derived organic acids (specifically 4PA and MA) as targeted antimicrobial additives against spoilage microorganisms, particularly Clostridium spp. This experimental design builds directly on previous work, which identified 4PA and MA as key limiting factors in nettle silage fermentation. Critically, 4PA actively inhibits LAB activity [11]; Consequently, this work represents a mechanistically justified progression toward understanding acid-mediated preservation strategies in high-protein silages.
DM content was highest attributed to malic acid after 30 days of ensiling, but this effect was disappeared as the silage fermentation time was prolonged. After 60 days of ensiling, 4PA treatment reduced crude protein loss by 43% compared to control (CK: 21.4% DM loss vs. 4PA: 12.2% DM loss) and by 39.0% versus malic acid (MA) treatment (2.85% DM loss), demonstrating superior preservation of protein integrity. The persistently high pH (>8.0 after 60 days) and complete disappearance of LA in all groups (Table 1) align with established characteristics of failed nettle ensiling [3]. These phenomena were caused by clostridial dominance—Clostridium sensu stricto 15 degrades proteins into AN, neutralizing organic acids and maintaining high pH. Accumulation of LA mainly responds for pH reduction in silage [15]. As present study showed that LA content was decreased until disappeared after 60 days of ensiling, which respond to pH level. LAB mainly convert WSC into organic acids such as lactic acid and acetic acid [16]. Data revealed that content of WSC was stabilized during 30—60 days of nettle ensiling, indicate that LAB fermentation process probably stopped during this period. However, both ammonia and BA were increased with ensiling time prolonged, indicate that Clostridial fermentation occurred during nettle ensiling. There are two types of Clostridium fermentation that cause silage to spoil and deteriorate, the type of glycolysis such as C. butyricum that generates hydrogen, carbon dioxide and BA through the fermentation of sugar and LA, and the type of protein decomposition such as C. sporogenes that produces ammonium nitrogen and amines by breaking down free amino acids and proteins [17, 18]. WSC content was decreased 72.26 % after 30 days of nettle ensiling and stabilized with ensiling time prolonged, same as LA content except it disappear with ensiling time prolonged to 60 days. The results probably suggest that these glycolysis type of Clostridium spp. prefer utilized LA as carbon source during nettle ensiling. C. sporogenes indicated strong capacity to degradation protein, especially when WSC is limited. As present study demonstrated that content of ammonia increased 3-fold after 60 days of ensiling when compared with 7 days of ensiling, indicate that activity of protein decomposition type of Clostridium was extremely higher during ensiling.
4-pentenoic acid is not a conventional organic acid additive of silage, it mainly found in nettle silage [3]. As a plant-specialized metabolite, 4-pentenoic acid uniquely targets proteolytic Clostridial in nettle silage—unlike conventional additives like malic acid. Little know about inhibit activity of 4-pentenoic acid against to bacteria, only one study observed that 4-pentenoic acid showed certainly level of capacity to inhibited relative abundance of C. botulinum [19]. Our previous studies identified 4-pentenoic acid as an endogenous metabolite in naturally fermented nettle silage, exhibits differential antibacterial activity: it strongly inhibits the beneficial lactic acid bacterium Pediococcus pentosaceus (MIC = 16 mg·mL−1) by disrupting cell membrane integrity and suppressing fatty acid biosynthesis (fabG) and acid-tolerance proteins (accA). Conversely, 4PA indirectly suppresses harmful bacteria (e.g., Clostridium sensu stricto 15) by promoting antagonistic LAB genera (e.g., Enterococcus), evidenced by significantly reduced AN production (p<0.05) [3,11]. The present study supplemented 4-pentenoic acid at 1% fresh weight—a concentration exceeding typical endogenous levels (0.2—0.5 mg·g−1 DM) observed in natural fermentation. Results confirmed that exogenous 4-pentenoic acid significantly suppressed proteolytic Clostridium sensu stricto 15, reducing BA and AN production by 42.35% and 25.95%, respectively, after 60 days of ensiling. This demonstrates 4-pentenoic acid’s capacity to inhibit relative abundance of Clostridium spp, and particularly glycolytic types, though the pharmacological dosage used here may not reflect its physiological function in natural systems.
Malic acid effectively suppresses harmful microorganisms (e.g., Clostridial and molds) through rapid acidification, without significantly inhibiting beneficial LAB, while synergistically enhancing silage quality when combined with homofermentative LAB. As a conventional organic acid additive in silage production, studies demonstrate that malic acid supplementation at 0.6%—1% (fresh weight basis) reduces AN content in silages with high CP levels (16.86%—22.20% of dry matter, DM) [20-22]. Unexpectedly, in nettle silage ( CP levels is 16.89% of DM), MA treatment increased AN production at the late stage of ensiling (day 60). This discrepancy likely stems from complex inhibitory interactions among endogenous biologically active compounds and microorganisms, necessitating further mechanistic investigation.
4.2. Effect of 4-Pentenoic Acid and Malic On Bacteria Community of Nettle Silage
Aerococcus belong to Lactobacillales, often consider as “more peripheral” LAB, usually observed in fermented food [23]. Only a few studies have found this bacterium exists in silage [24-26]. As present study results demonstrated that, Aerococcus was most dominant bacteria during entire nettle ensiling process, and acids treated revealed none impact for it. Same result was observed in paper mulberry silage as Aerococcus was dominant during ensiling, but dropped 55.79 % activity when inoculation with some strain of Lactobacillus [24]. However, relative abundant of this bacteria were usually below 10 % in alfalfa and pennisetum, even below 1 % after Lactobacillus was inoculated [25,26]. Clearly, Lactobacillus had capacity to inhibited Aerococcus activity. Thus, high level relative abundance of Aerococcus probably due to both nettle and paper mulberry contain lots of bioactivity compounds (polyphenol, flavonoids, organic acids, etc.) such as morin and naringin which impact Lactobacillus activity [3,27].
Both acids treated increased relative abundance of Weissella after 30 days of nettle ensiling, especially for 4-pentenoic acid showed the higher capacity. Weissella is obligate heterofermentative LAB which initiating early fermentation of silage, and activity could highly reduction when pH decline (usually < 5.0) with ensiling prolonged [28]. One study observed that malic acid decreased Weissella activity in silage system, it was mainly due to pH rapidly declined reach to 4.0 in Moringa oleifera leaves (MOL) silages [29]. Present study indicated that pH was over 8, indicate 4-pentenoic acid and malic acid probably was the mainly reason for this bacteria growth, specific mechanism of this need further study. Furthermore, both acids treated inhibited relative abundance of Tissierella after 60 days of nettle ensiling, especially for 4-pentenoic acid showed the higher capacity. Tissierella could growth at pH range from 6.5 to 8.5 (optimum pH of 8.3) in sludge fermentation and produce butyric acid [30].
Additionally, present study indicated that 4PA treated showed trend to inhibited relative abundance of Clostridium sensu stricto 15 after 30 days of nettle ensiling. It is the first time to found that 4-pentenoic acid probably showed the capacity to inhibited Clostridium in silage. Same studies were found that addition with malic acid (5 g·kg−1 of FM) into alfalfa and cassava silage could decreased relative abundances of Clostridium ( BA and AN were significantly decreased.) [21,22].
Regarding the dual effects on LAB and Clostridia, both acids exhibited distinct patterns. MA transiently stimulated LAB activity early in ensiling (e.g., increased LA by 57.1% vs. control at day 7; Table 1), likely via its metabolic utilization by Lactobacillus [3,11]. However, this LAB-promoting effect diminished at day 30, coinciding with resurgence of Clostridium sensu stricto 15 (Figure 3B) and elevated BA/AN in MA silages. In contrast, 4PA initially suppressed LAB (e.g., reduced LA by 8.3% vs. MA at day 7), consistent with its inhibitory role against Pediococcus in nettle silage [3,11]. Crucially, 4PA demonstrated superior and sustained inhibition of Clostridium sensu stricto 15 , correlating with 42.4% lower BA and 26.0% lower AN vs. MA at day 60 . This indicates that while both acids may partially inhibit LAB, 4PA’s selective and potent anti-clostridial activity (evidenced by its direct suppression of C. botulinum in vitro [11,19]) overrides its early LAB suppression, ultimately preserving protein integrity and fermentation quality. MA treated’s weaker clostridial inhibition likely stems from its primary role as a metabolic substrate rather than a dedicated antimicrobial agent.
Notably, based on LEfSe analysis, present study showed that Jeotgalibaca were potential important bacteria involved in early-fermentation stage (7 days) of nettle silage, however, this result changed to Lactococcus and Aerococcus when addition with malic acid and 4-pentenoic acid, respectively. Facklamia and Rubellimicrobium were potential important bacteria involved in middle-fermentation stage (30 days) of nettle silage when addition with 4-pentenoic acid and malic acid, respectively. Atopostipes was potential important bacteria involved in later-fermentation stage (60 days) of nettle silage, however, this result changed to Tissierella when addition with malic acid. Overall, the results suggested that both acids inhibited harmful microorganisms (such as Clostridium) throughout the entire fermentation period, with 4-pentenoic acid showing superior efficacy.
4.3. Correlation Analysis Between Bacteria Community and Fermentation Characteristics in Nettle Silage
Clostridium sensu stricto 15 was positive correlated with BA and AN accumulation, which explains the substantial protein degradation (evidenced by elevated AN content in present study) and BA production in control and malic acid-treated silages after 30—60 days of ensiling. This genus likely drove proteolysis and saccharolytic activity, converting proteins and residual organic acids into ammonia and butyrate, thereby elevating pH and compromising silage quality [31].
In contrast, genera such as Aerococcus, Lactococcus, Carnobacterium, and Enterococcus demonstrated negative correlations with BA, AN, and pH , while positively correlating with WSC, CP, LA, and AA. Although Aerococcus dominated during entire ensiling process of nettle, its result in preserving CP and WSC (positive correlation) suggests potential metabolic activity that limited protein degradation. While Aerococcus showed positive correlation with CP, its persistent dominance in high-pH silage suggests environmental tolerance rather than proteolytic suppression [27]. Notably, Lactococcus and Enterococcus—typically associated with early-stage LAB fermentation—showed positive links to LA and AA but were insufficient to counteract clostridial bacteria activity in control groups [24].
Critically, the suppression of Clostridium sensu stricto 15 and Tissierella by organic acid treatments underpins their efficacy in improving silage quality. The 4-pentenoic acid treatment most strongly inhibited these genera (decreased relative abundance of Tissierella for 98.80%, CK 6.85% vs. 4PA 0.08%) , corresponding to its decreased production of 42.35 % and 25.95% for BA and AN, respectively, after 60 days of ensiling . This microbial inhibition activity in silage probably aligns with the mechanism of 4-pentenoic acid showed antibacterial activity in vitro [11]. Malic acid also reduced Tissierella (decreased for 66.42%) but was less effective against Clostridium sensu stricto 15, consistent with its weaker suppression of BA and AN production.
The inverse correlation between Weissella and clostridial genera further supports Weissella as a beneficial heterofermenter in this high-pH system [32]. Enhanced Weissella activity in 4-pentenoic acid treatment silages (2.55% vs. 0.64% in control after 30 days of ensiling) likely contributed to AA accumulation (1.37% DM vs. 0.63% DM in control), partially offsetting the lack of LA acidification [33].
5. Conclusions
This study confirms that 4-pentenoic acid (addition with 1% FW) effectively enhances nettle silage quality by specifically inhibiting proteolytic Clostridium sensu stricto 15, thereby reducing BA and AN production, while preserving crude protein. Concurrently, it promotes Weissella activity and AA accumulation, partially compensating for LA deficiency. Compared to malic acid, 4-pentenoic acid demonstrates superior efficacy in controlling clostridial spoilage, establishing it as a novel promising plant-derived preservative for high-protein silages.
Author Contributions
Y.S., Conceptualization, Writing, Reviewing, and Editing; Y.C., Resources, Validation, Methodology; S.L., Y.C., Investigation, Resources; C.M., Validation, Supervision; R.H., Writing, Reviewing, Validation Supervision and Editing; F.Z., Resources, Investigation, Validation Supervision, Review, and Editing.
Funding
This research was funded by the High-Level Talent Research Initiation Project of Shihezi University (RCZK202575); China Agriculture Research System of MOF and MARA (CARS-34); the National Natural Science Foundation of China (NSFC) (grant number: 32060399).
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Data Availability Statement
The datasets supporting the conclusions of this article are available in the National Center for Biotechnology (NCBI) Sequence Read Archive (SRA) of bacteria under accession numbers “PRJNA1233857” (accessed on March 13, 2025) for addition with 4-pentenoic acid and “PRJNA1236718” (accessed on March 16, 2025) for addition with malic acid into nettle silage, respectively. http://www.ncbi.nlm.nih.gov.
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Abbreviations
The following abbreviations are used in this manuscript:
| AA | Acetic acid |
| AN | Ammonia-N |
| BA | Butyric acid |
| CP | Crude protein |
| DM | Dry matter |
| LA | Lactic acid |
| LAB | Lactic acid bacteria |
| MA | malic acid |
| PA | Propionic acid |
| WSC | Water-soluble carbohydrate |
| 4PA | 4-pentenoic acid |
References
- Ding, Z. Z.; Xiao, C.F.; Xu, Y. Y.; Lu, W. W.; Zhu, L. H. . Research progress on the application of alternative unconventional feed ingredients in poultry breeding. Acta Agric. Shanghai. 2024, 40, 142–152. [Google Scholar] [CrossRef]
- Bhusal, K. K.; Magar, S. K.; Thapa, R.; Lamsal, A.; Bhandari, S.; Maharjan, R.; Shrestha, S.; Shrestha, J. . Nutritional and pharmacological importance of stinging nettle (Urtica dioica L.): A review. Heliyon. 2022, 8, e09717. [Google Scholar] [CrossRef]
- Huang, R. Z.; Chen, Y. C.; Ma, C. H.; Chai, Y. X.; Jia, S. A.; Zhang, F. F. Potential Factors Causing Failure of Whole Plant Nettle (Urtica Cannabina) Silages. Front. Microbiol. 2023, 13, 1113050. [Google Scholar] [CrossRef]
- Dai, Q. Z.; Hou, Z. P.; Gao, S.; Li, Z. C.; Wei, Z. S.; Wu, D. Q. . Substitution of fresh forage ramie for alfalfa hay in diets affects production performance, milk composition, and serum parameters of dairy cows. Trop. Anim. Health Prod. 2019, 51, 469–472. [Google Scholar] [CrossRef]
- Mirsaiidi Farahani, M.; Hosseinian, S. A. . Effects of dietary stinging nettle (Urtica dioica) on hormone stress and selected serum biochemical parameters of broilers subjected to chronic heat stress. Vet. Med. Sci. 2022, 8, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Pirgozliev, V. R.; Kljak, K.; Whiting, I. M.; Mansbridge, S. C.; Atanasov, A.G.; Enchev, S. B.; Tukša, M.; Rose, S. P. . Dietary stinging nettle (Urtica dioica) improves carotenoids content in laying hen egg yolk. Br. Poult. Sci. 2025, 66, 275–280. [Google Scholar] [CrossRef] [PubMed]
- De Toledo, G. S. P.; da Silva, L. P.; de Quadros, A. R. B.; Retore, M.; Araújo, I. G.; Brum, H. S.; Melchior, R. . Productive performance of rabbits fed with diets containing ramie (Boehmeria nivea) hay in substitution to alfalfa (Medicago sativa) hay. Verona, Italy: In 9th World Rabbit Congress 2008, 827–830.
- Liu, J. Y.; Zhao, M.; Hao, J. F.; Yan, X. Q.; Fu, Z. H.; Zhu, N.; Jia, Y. S.; Wang Z., J.; Ge, G. T. . Effects of temperature and lactic acid Bacteria additives on the quality and microbial community of wilted alfalfa silage. BMC Plant Biol. 2024, 24, 844. [Google Scholar] [CrossRef]
- Jia, S. A.; Chai, Y. X.; Yang, H. J. , Zhang, F. F.; Ma, C. H.. Isolation and identification of dominant lactic acid bacteria from Urtica cannabina silage. Pratac. Sci. 2023, 40, 1410–1419. [Google Scholar] [CrossRef]
- Chai, Y. X.; Jia, S. A.; Zhang, T.; Li, X.; Chen, Y. C.; Huang, R. Z.; Zhang, F. F. . Effects of Lactiplantibacillus plantarum on Silage Quality of Urtica cannabina and Rumen Degradation Characteristics of Sheep. Acta Agrestia Sin. 2024, 32, 2283–2289. [Google Scholar] [CrossRef]
- Huang, R. Z.; Chai, Y. X.; Li, S. M.; Chen, Y. C.; Jia, S. A.; Ma, C. H.; Zhang, F. F. . Involvement of 4-pentenoic acid in causing quality deterioration of nettle silage: study of antibacterial mechanism. Microbiol. Spectr. 2025, 13, e0266724. [Google Scholar] [CrossRef] [PubMed]
- McDonald, P.; Henderson, A. R. Determination of Water-Soluble Carbohydrates in Grass. J. Sci. Food Agric. 1964, 15, 395–398. [Google Scholar] [CrossRef]
- Weatherburn, M. W. Phenol-Hypochlorite Reaction for Determination of Ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- Chen, Y. C.; Li, S. M.; Sun, Y. C.; Chai, Y. X.; Jia, S. A.; Ma, C. H.; Zhang, F. F. The Effects of Lactococcus garvieae and Pediococcus pentosaceus on the Characteristics and Microbial Community of Urtica cannabina Silage. Microorganisms. 2025, 13, 1453. [Google Scholar] [CrossRef]
- Ali, N.; Wang, S. R.; Zhao, J.; Dong, Z. H.; Li, J. F.; Nazar, M.; Shao, T. Microbial Diversity and Fermentation Profile of Red Clover Silage Inoculated with Reconstituted Indigenous and Exogenous Epiphytic Microbiota. Bioresour. Technol. 2020, 314, 123606. [Google Scholar] [CrossRef]
- Ren, H.; Feng, Y.; Pei, J.; Li, J.; Wang, Z.; Fu, S.; Zheng, Y.; Li, Z.; Peng, Z. Effects of Lactobacillus Plantarum Additive and Temperature on the Ensiling T Quality and Microbial Community Dynamics of Cauliflower Leaf Silages. Bioresour. Technol. 2020, 307, 123238. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M. Y.; Abdullah, N.; Magaji, U.; Hussin, G.; Ramli, A.; Miah, G. Fermentation Quality and Additives: A Case of Rice Straw Silage. Biomed Res. Int. 2016, 7985167. [Google Scholar] [CrossRef]
- Driehuis, F.; Elferink, S. The Impact of the Quality of Silage on Animal Health and Food Safety: A Review. Vet. Q. 2000, 22, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Huhtanen, C. N.; Trenchard, H.; Milnes-McCaffery, L. Inhibition of Clostridium-Botulinum in Comminuted Bacon by Short-Chain Alkynoic and Alkenoic Acids and Esters. J. Food Prot. 1985, 48, 570–573. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q. X.; Shi, W. J.; Zhou, J. Addition of Organic Acids and Lactobacillus Acidophilus to the Leguminous Forage Chamaecrista Rotundifolia Improved the Quality and Decreased Harmful Bacteria of the Silage. Animals. 2022, 12, 2260. [Google Scholar] [CrossRef]
- Ke, W. C.; Ding, Z. T.; Li, F. H.; Xu, D. M.; Bai, J.; Muhammad, I.; Zhang, Y. X.; Zhao, L. S.; Guo, X. S. Effects of Malic or Citric Acid on the Fermentation Quality, Proteolysis and Lipolysis of Alfalfa Silage Ensiled at Two Dry Matter Contents. J. Anim. Physiol. Anim. Nutr. 2021, 106, 988–994. [Google Scholar] [CrossRef]
- Li, M.; Zhang, L. D.; Zhang, Q.; Zi, X. J.; Lv, R. L.; Tang, J.; Zhou, H. L. Impacts of Citric Acid and Malic Acid on Fermentation Quality and Bacterial Community of Cassava Foliage Silage. Front. Microbiol. 2020, 11, 595622. [Google Scholar] [CrossRef]
- Petrova, P.; Petrov, K. Lactic Acid Fermentation of Cereals and Pseudocereals: Ancient Nutritional Biotechnologies with Modern Applications. Nutrients. 2020, 12, 1118. [Google Scholar] [CrossRef]
- Zhang, Y. L.; Yang, H. J.; Huang, R. Z.; Wang, X. Z.; Ma, C. H.; Zhang, F. F. Effects of Lactiplantibacillus Plantarum and Lactiplantibacillus Brevis on Fermentation, Aerobic Stability, and the Bacterial Community of Paper Mulberry Silage. Front. Microbiol. 2022, 13, 1063914. [Google Scholar] [CrossRef]
- Li, Z. Q.; Li, F. H.; Xie, D. M.; Zhang, B. B.; Kharazian, Z. A.; Guo, X. S. Effects of Bacteriocin-Producing Lactiplantibacillus Plantarum on Fermentation, Dynamics of Bacterial Community, and Their Functional Shifts of Alfalfa Silage with Different Dry Matters. Fermentation. 2022, 8, 690. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, H.; Li, J.; Dong, Z.; Wang, S.; Shao, T. Relationships between Microbial Community, Functional Profile and Fermentation Quality during Ensiling of Hybrid Pennisetum. J. Anim. Feed Sci. 2023, 32, 447–458. [Google Scholar] [CrossRef]
- Gutierrez-Venegas, G.; Gomez-Mora, J. A.; Meraz-Rodriguez, M. A.; Flores-Sanchez, M. A.; Ortiz-Miranda, L. F. Effect of Flavonoids on Antimicrobial Activity of Microorganisms Present in Dental Plaque. Heliyon. 2019, 5, e03013. [Google Scholar] [CrossRef] [PubMed]
- Mu, L.; Xie, Z.; Hu, L. X.; Chen, G. H.; Zhang, Z. F. Cellulase Interacts with Lactobacillus Plantarum to Affect Chemical Composition, Bacterial Communities, and Aerobic Stability in Mixed Silage of High-Moisture Amaranth and Rice Straw. Bioresour. Technol. 2020, 315, 123775. [Google Scholar] [CrossRef]
- Tian, H. C.; Wang, Y.; Liu, Z. C.; Hu, Z. Y.; Guo, Y. Q.; Deng, M.; Liu, G. B.; Sun, B. L. Effects of Malic Acid and Sucrose on the Fermentation Parameters, CNCPS Nitrogen Fractions, and Bacterial Community of Moringa Oleifera Leaves Silage. Microorganisms. 2021, 9, 2102. [Google Scholar] [CrossRef]
- Maspolim, Y.; Zhou, Y.; Guo, C. H.; Xiao, K. K.; Ng, W. J. The Effect of pH on Solubilization of Organic Matter and Microbial Community Structures in Sludge Fermentation. Bioresour. Technol. 2015, 190, 289–298. [Google Scholar] [CrossRef]
- Driehuis, F.; Elferink, S.O.; Van Wikselaar, P.G. . Fermentation characteristics and aerobic stability of grass silage inoculated with Lactobacillus buchneri, with or without homofermentative lactic acid bacteria. Grass Forage Sci. 2001, 56, 330–343. [Google Scholar] [CrossRef]
- Yan, X,; Ai, L. Z.; Xia, Y. J.; Song, X.; Zhang, H.; Ni, B.; Yang, D. J. Research progress of safety, probiotic potential and functional properties of Weissella confusa. Ind. Microbiol. 2022, 52, 37–48. [Google Scholar]
- Guo, L. N.; Wang, X. K.; Chen, H. L.; Li, X. M.; Xiong, Y.; Zhou, H. Z.; Xu, G.; Yang, F. Y.; Ni, K. K. . Exploring the fermentation quality, bacterial community and metabolites of alfalfa ensiled with mugwort residues and Lactiplantibacillus pentosus. Chem. Biol. Technol. Agric. 2023, 10, 107. [Google Scholar] [CrossRef]
Figure 1.
Alpha diversity of bacterial community after addition with malic and 4-pentenoic acid during nettle ensiling. (A) chao 1 index; (B) ace index; (C) Shannon index. CK7, 4PA7 and MA7: control group, addition with 4-pentenoic acid and addition with malic acid after 7 days of nettle ensiling, respectively. CK30, 4PA30 and MA30: control group, addition with 4-pentenoic acid and addition with malic acid after 30 days of nettle ensiling, respectively. CK60, 4PA60 and MA60: control group, addition with 4-pentenoic acid and addition with malic acid after 60 days of nettle ensiling, respectively. “*” means p<0.05. Same as below.
Figure 1.
Alpha diversity of bacterial community after addition with malic and 4-pentenoic acid during nettle ensiling. (A) chao 1 index; (B) ace index; (C) Shannon index. CK7, 4PA7 and MA7: control group, addition with 4-pentenoic acid and addition with malic acid after 7 days of nettle ensiling, respectively. CK30, 4PA30 and MA30: control group, addition with 4-pentenoic acid and addition with malic acid after 30 days of nettle ensiling, respectively. CK60, 4PA60 and MA60: control group, addition with 4-pentenoic acid and addition with malic acid after 60 days of nettle ensiling, respectively. “*” means p<0.05. Same as below.
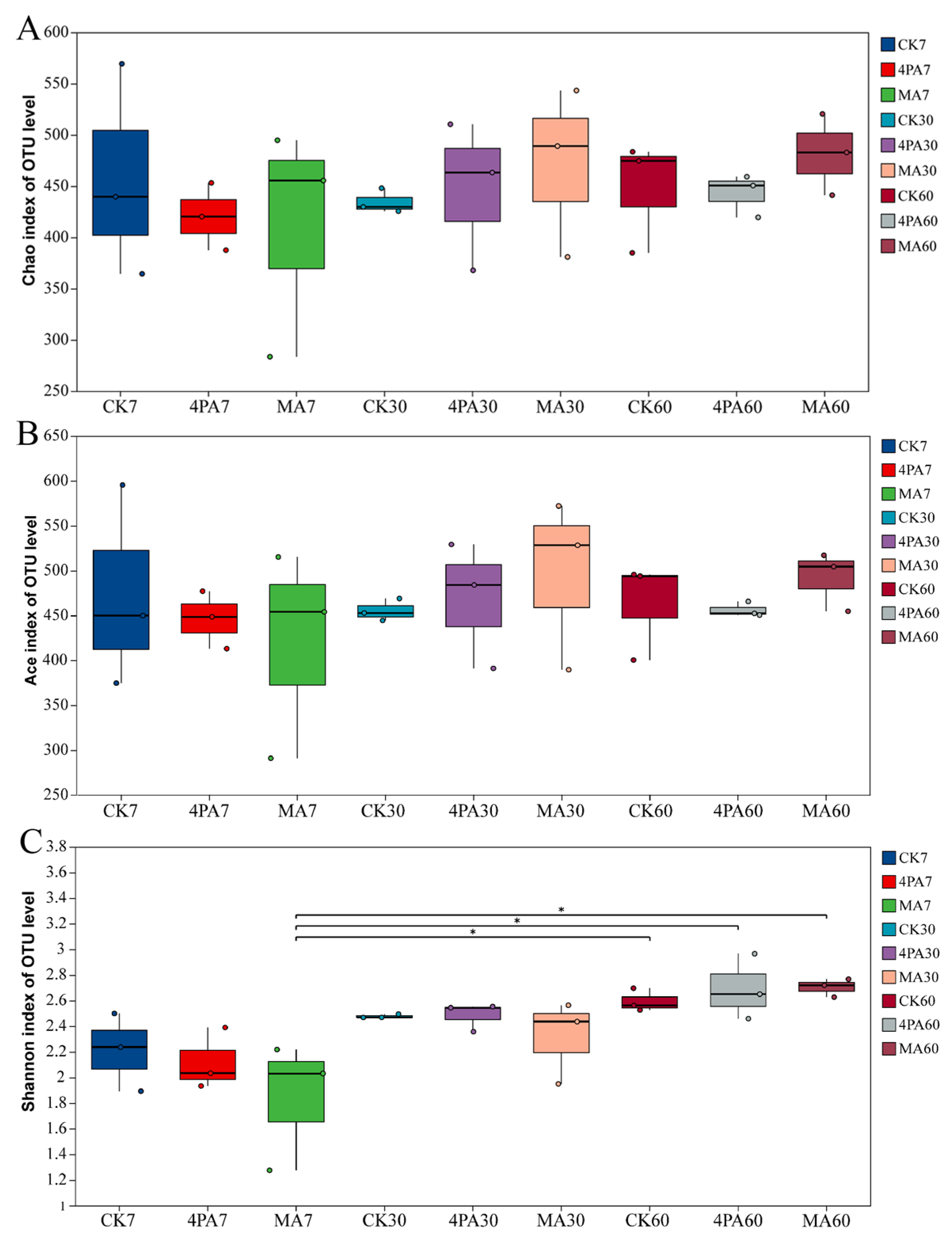
Figure 2.
Principal-coordinate analysis (PCoA) plots based on weighted UniFrac distance for bacterial community and treatment (R=0.5606, p=0.00100). CK7, 4PA7 and MA7: control group, addition with 4-pentenoic acid and addition with malic acid after 7 days of nettle ensiling, respectively. CK30, 4PA30 and MA30: control group, addition with 4-pentenoic acid and addition with malic acid after 30 days of nettle ensiling, respectively. CK60, 4PA60 and MA60: control group, addition with 4-pentenoic acid and addition with malic acid after 60 days of nettle ensiling, respectively.
Figure 2.
Principal-coordinate analysis (PCoA) plots based on weighted UniFrac distance for bacterial community and treatment (R=0.5606, p=0.00100). CK7, 4PA7 and MA7: control group, addition with 4-pentenoic acid and addition with malic acid after 7 days of nettle ensiling, respectively. CK30, 4PA30 and MA30: control group, addition with 4-pentenoic acid and addition with malic acid after 30 days of nettle ensiling, respectively. CK60, 4PA60 and MA60: control group, addition with 4-pentenoic acid and addition with malic acid after 60 days of nettle ensiling, respectively.
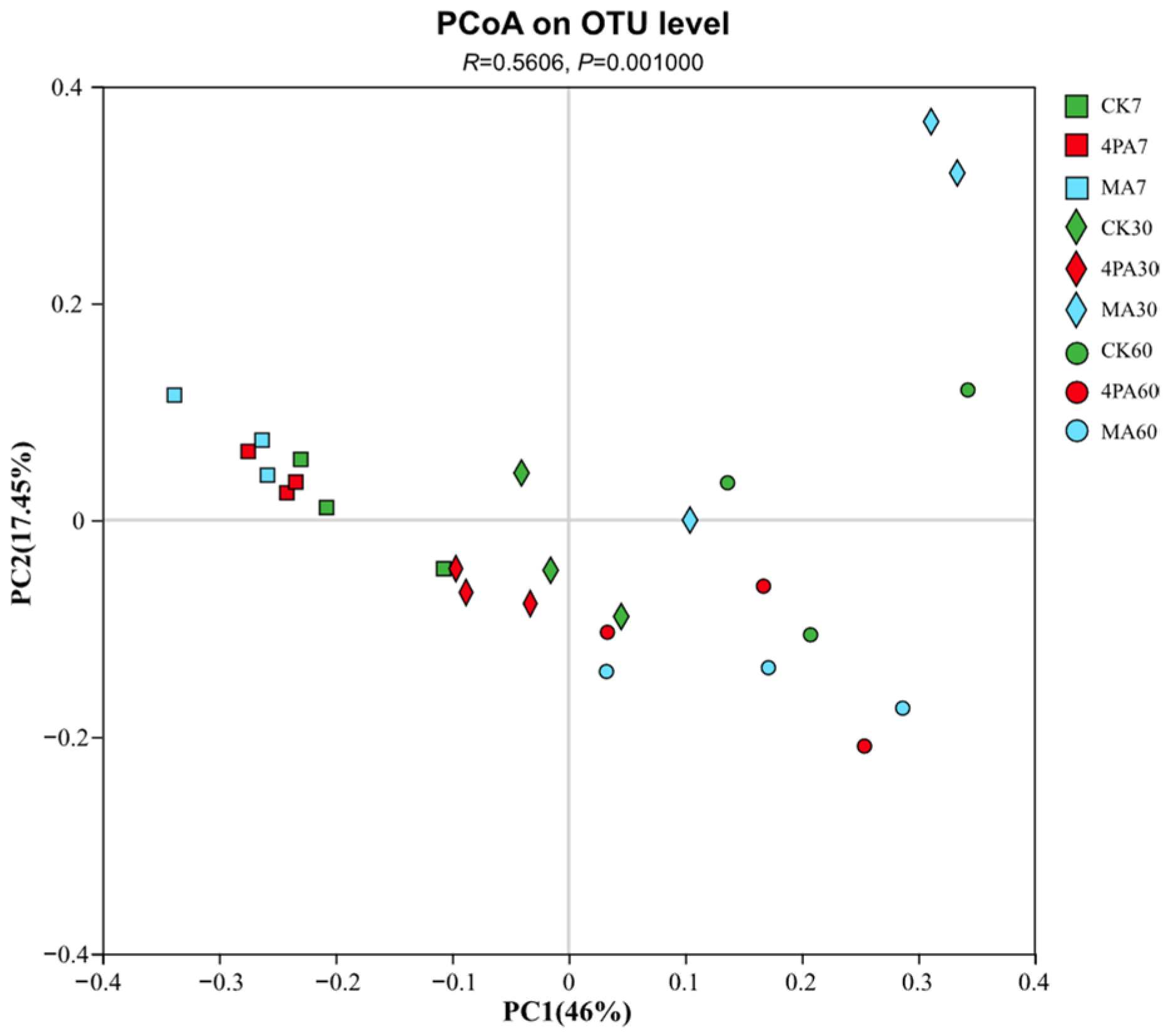
Figure 3.
Bacterial community analysis on phylum (A) and genus level (B) after addition with malic and 4-pentenoic acid during nettle ensiling. CK7, 4PA7 and MA7: control group, addition with 4-pentenoic acid and addition with malic acid after 7 days of nettle ensiling, respectively. CK30, 4PA30 and MA30: control group, addition with 4-pentenoic acid and addition with malic acid after 30 days of nettle ensiling, respectively. CK60, 4PA60 and MA60: control group, addition with 4-pentenoic acid and addition with malic acid after 60 days of nettle ensiling, respectively.
Figure 3.
Bacterial community analysis on phylum (A) and genus level (B) after addition with malic and 4-pentenoic acid during nettle ensiling. CK7, 4PA7 and MA7: control group, addition with 4-pentenoic acid and addition with malic acid after 7 days of nettle ensiling, respectively. CK30, 4PA30 and MA30: control group, addition with 4-pentenoic acid and addition with malic acid after 30 days of nettle ensiling, respectively. CK60, 4PA60 and MA60: control group, addition with 4-pentenoic acid and addition with malic acid after 60 days of nettle ensiling, respectively.
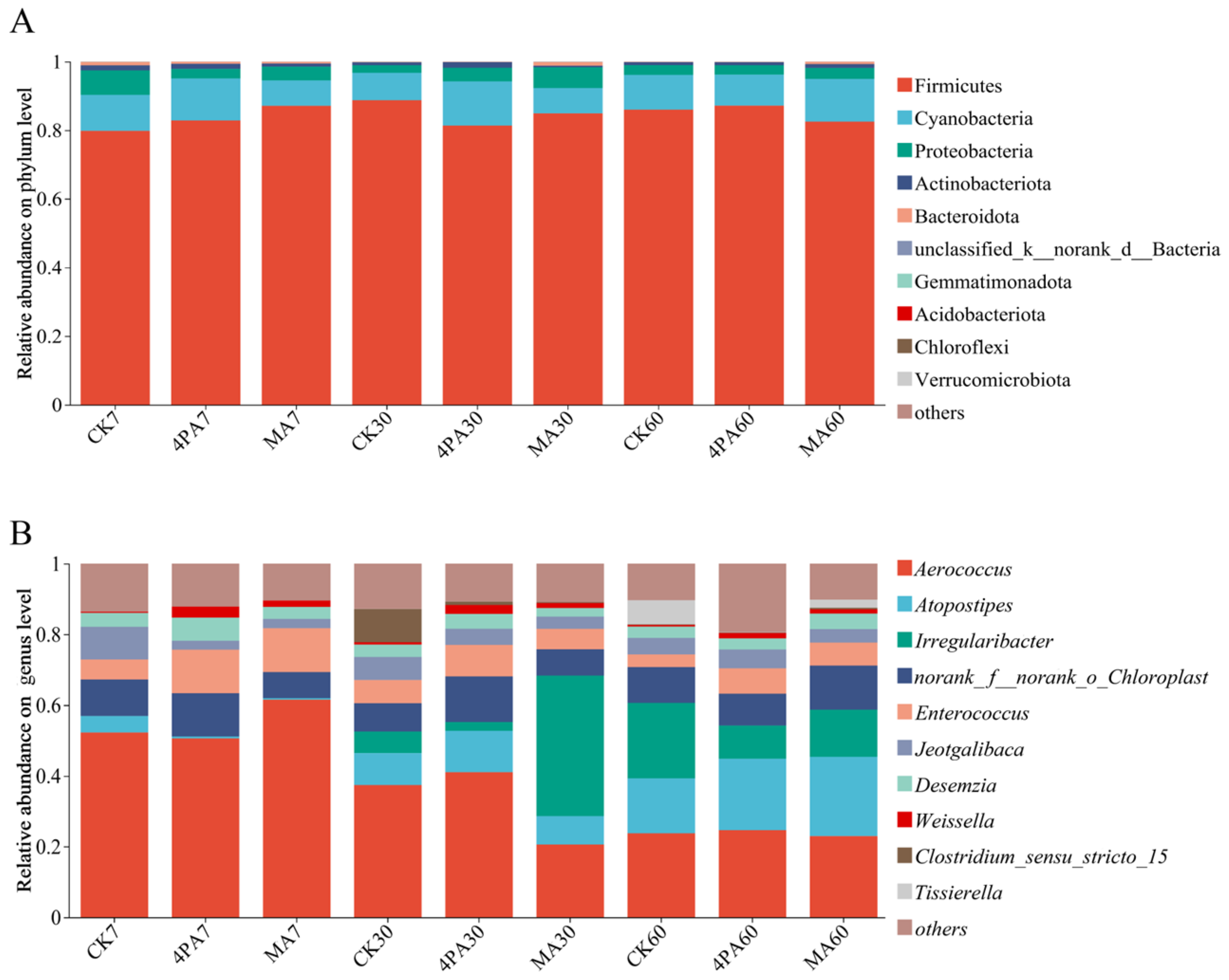
Figure 4.
Comparison of microbial variations using LEfSe (LDA effect size) analysis of nettle silage after addition with malic and 4-pentenoic acid. CK7, 4PA7 and MA7: control group, addition with 4-pentenoic acid and addition with malic acid after 7 days of nettle ensiling, respectively. CK30, 4PA30 and MA30: control group, addition with 4-pentenoic acid and addition with malic acid after 30 days of nettle ensiling, respectively. CK60, 4PA60 and MA60: control group, addition with 4-pentenoic acid and addition with malic acid after 60 days of nettle ensiling, respectively.
Figure 4.
Comparison of microbial variations using LEfSe (LDA effect size) analysis of nettle silage after addition with malic and 4-pentenoic acid. CK7, 4PA7 and MA7: control group, addition with 4-pentenoic acid and addition with malic acid after 7 days of nettle ensiling, respectively. CK30, 4PA30 and MA30: control group, addition with 4-pentenoic acid and addition with malic acid after 30 days of nettle ensiling, respectively. CK60, 4PA60 and MA60: control group, addition with 4-pentenoic acid and addition with malic acid after 60 days of nettle ensiling, respectively.
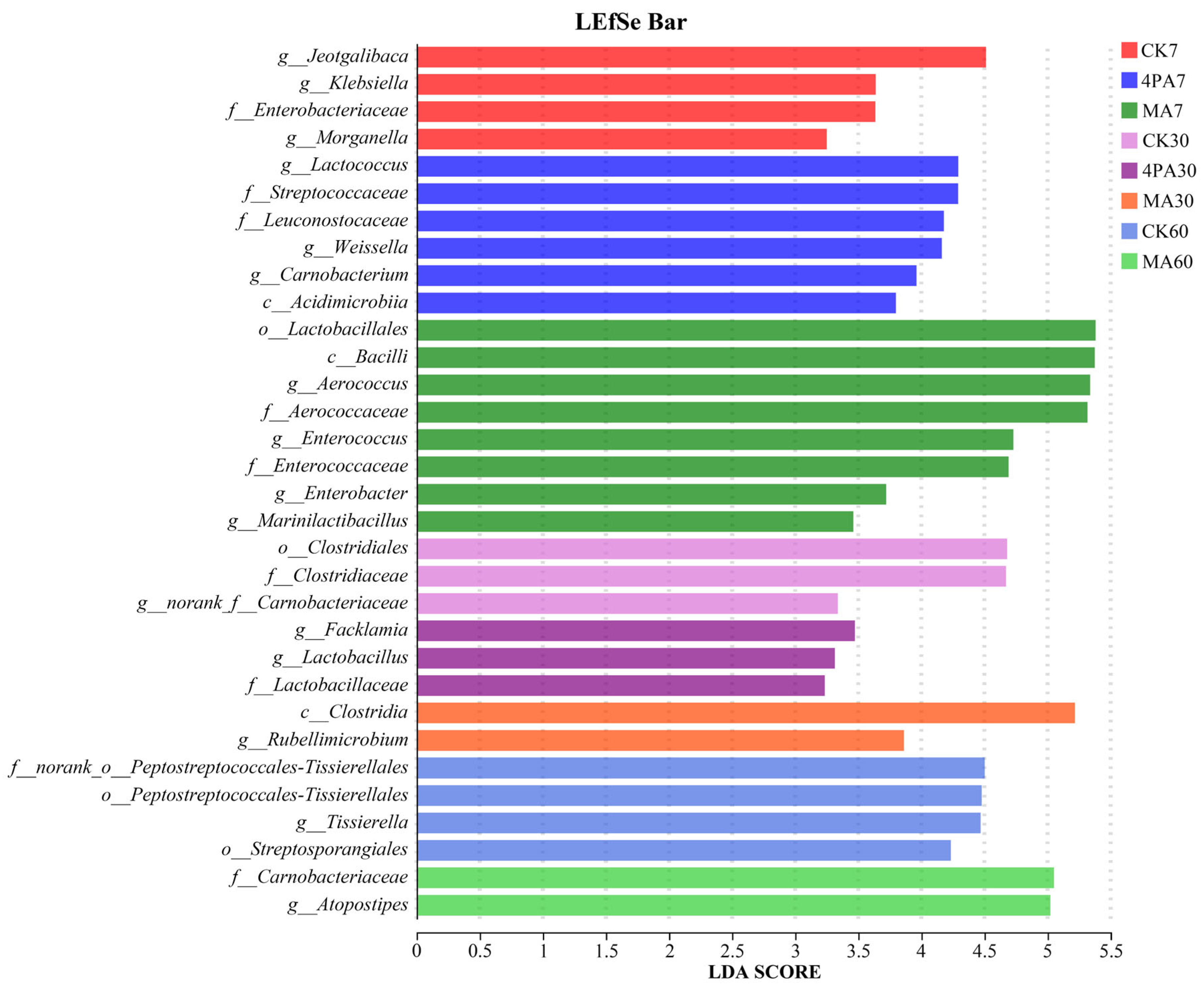
Figure 5.
Heatmap of correlation analysis between bacteria and characteristic in nettle silage. Red/blue indicate positive/negative correlations. “*” means p<0.05; “**” means 0.01<p<0.05; “***” means p<0.001. Note: DM, dry matter; WSC, water-soluble carbohydrate; LA, lactic acid; CP, crude protein; AA, acetic Acid; PA, propanoic acid; BA, butyric acid; AN, ammonia; “*” indicates significant correlation (P<0.05); “**” indicates highly significant correlation (P<0.01).
Figure 5.
Heatmap of correlation analysis between bacteria and characteristic in nettle silage. Red/blue indicate positive/negative correlations. “*” means p<0.05; “**” means 0.01<p<0.05; “***” means p<0.001. Note: DM, dry matter; WSC, water-soluble carbohydrate; LA, lactic acid; CP, crude protein; AA, acetic Acid; PA, propanoic acid; BA, butyric acid; AN, ammonia; “*” indicates significant correlation (P<0.05); “**” indicates highly significant correlation (P<0.01).
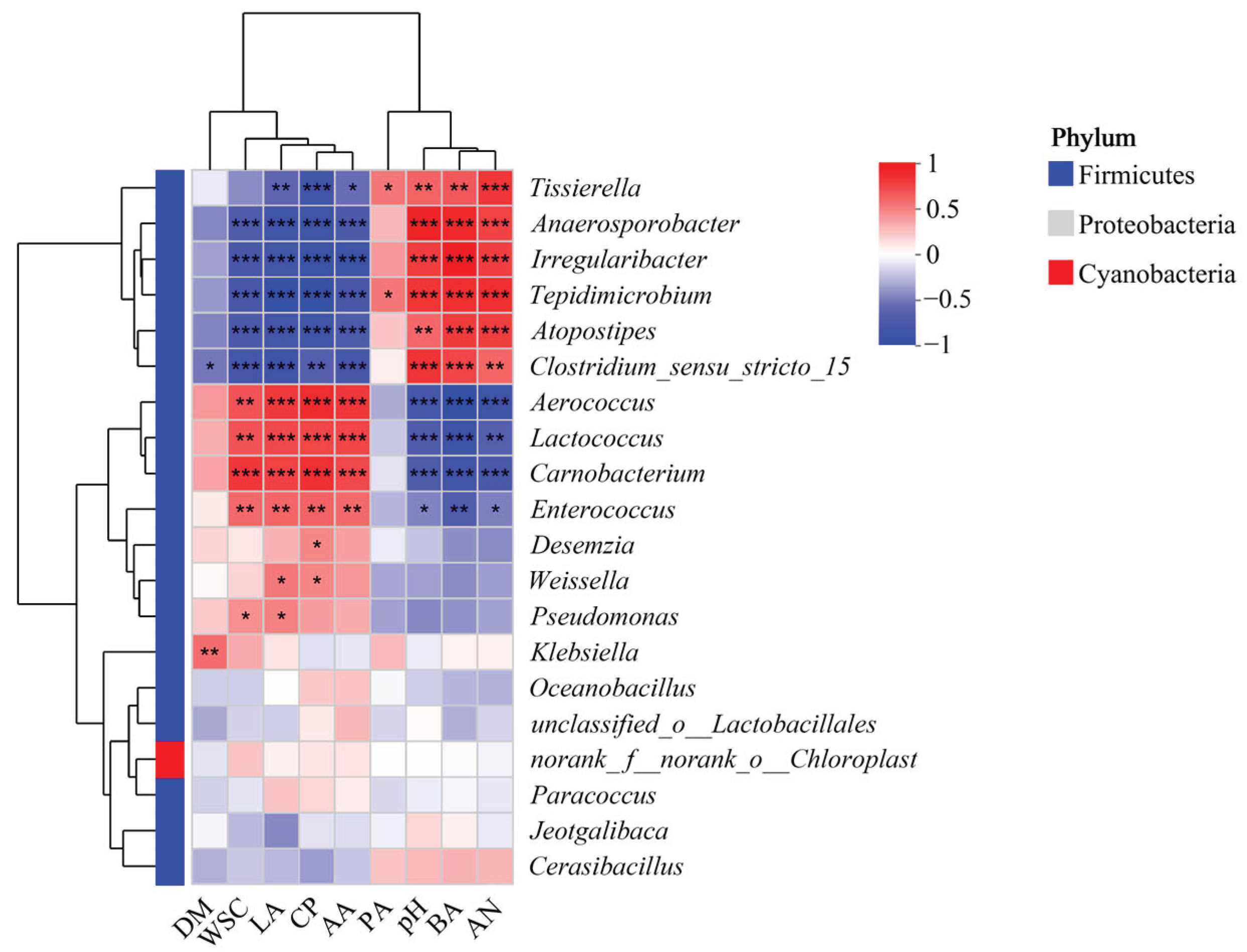

Table 1.
Characteristics during nettle ensiling after addition with relevant organic acids % DM
| Index | Time/day | Groups | SEM | pvalue | ||
| CK | MA | 4PA | ||||
| DM (% fresh weight) | 7 | 37.72 | 37.23 | 36.58 | 1.245 | 0.675 |
| 30 | 35.45 b | 36.66 a | 35.39 b | 0.038 | 0.027 | |
| 60 | 36.33 | 36.78 | 35.08 | 0.871 | 0.211 | |
| CP% DM | 7 | 16.23 a | 15.89 b | 16.32 a | 0.039 | <0.001 |
| 30 | 15.27 c | 15.60 b | 15.96 a | 0.067 | <0.001 | |
| 60 | 13.27 c | 13.72 b | 14.83 a | 0.062 | <0.001 | |
| WSC% DM | 7 | 2.92 | 3.30 | 2.95 | 0.230 | 0.263 |
| 30 | 0.68 | 0.73 | 0.71 | 0.045 | 0.606 | |
| 60 | 0.69 | 0.74 | 0.68 | 0.041 | 0.386 | |
| pH | 7 | 7.65 c | 7.80 b | 7.98 a | 0.024 | <0.001 |
| 30 | 8.35 a | 8.15 b | 8.11 b | 0.064 | 0.019 | |
| 60 | 8.47 a | 8.24 a | 8.10 b | 0.096 | 0.023 | |
| LA/(g·kg−1 DM) | 7 | 2.68 b | 4.21 a | 3.86 b | 0.425 | 0.026 |
| 30 | 0.016 c | 0.17 b | 0.40 a | 0.048 | <0.001 | |
| 60 | ND | ND | ND | - | - | |
| AA/(g·kg−1 DM) | 7 | 1.74 | 1.74 | 1.77 | 0.251 | 0.990 |
| 30 | 0.94 b | 0.93 b | 1.57 a | 0.070 | <0.001 | |
| 60 | 0.63 b | 0.50 b | 1.37 a | 0.166 | 0.004 | |
| PA/(g·kg−1 DM) | 7 | 0.054 | 0.068 | 0.076 | 0.016 | 0.413 |
| 30 | 0.082 | 0.078 | 0.050 | 0.019 | 0.215 | |
| 60 | 0.19 a | 0.083 b | 0.065 b | 0.026 | 0.005 | |
| BA/(g·kg−1 DM) | 7 | ND | ND | ND | - | - |
| 30 | 4.99 a | 5.59 a | 2.92 b | 0.287 | <0.001 | |
| 60 | 8.31 a | 7.56 a | 4.79 b | 0.367 | <0.001 | |
| AN (% of TN) DM | 7 | 9.81 | 14.30 | 9.94 | 2.280 | 0.160 |
| 30 | 12.90 a | 12.47 a | 10.86 b | 0.190 | <0.001 | |
| 60 | 32.79 b | 39.33 a | 24.28 c | 1.150 | <0.001 | |
Note: CK: control group; MA: addition with 1% (fresh weight) malic acid; 4PA: addition with 1 % (fresh weight) 4-pentenoic acid. Different lowercase letters on the same row indicate significant differences (p<0.05). DM, dry matter basis; CP, crude protein; WSC, water-soluble carbohydrate; LA, lactic acid; AA, acetic Acid; PA, propanoic acid; BA, butyric acid; AN, ammonia. Data are presented as the means ± standard error. ND: not detected. SEM: standard error of the mean.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



