Submitted:
18 August 2025
Posted:
19 August 2025
You are already at the latest version
Abstract
Host plant limitation poses a major threat to the endangered specialist butterfly Luehdorfia chinensis, whose larvae are strictly monophagous on Asarum spp. (A. sieboldii or A. forbesii). To simulate natural starvation caused by host plant scarcity, third- to fifth-instar larvae were subjected to a three-day deprivation treatment, and the effects on individual fitness traits—including larval development, pupal duration, and adult fecundity—together with population dynamics were assessed. Starvation significantly prolonged larval development, shortened the pupal stage, reduced female fecundity, and markedly decreased key population parameters, such as the intrinsic rate of increase (rₘ) and the net reproductive rate (R₀). Population projections further indicated that repeated starvation stress could reduce population size by more than 83% within two years, potentially intensifying genetic drift, inbreeding depression, and demographic instability, and ultimately increasing extinction risk. These findings provide direct evidence that host plant limitation drives population decline in L. chinensis, contribute to the broader understanding of global butterfly declines, and underscore the critical importance of conserving and restoring essential habitat resources. Moreover, they highlight the relevance of the resource-based habitat concept for the effective protection of specialist species.
Keywords:
Luehdorfia chinensis
; Larval starvation
; host plant limitation
; population dynamics
; life table
1. Introduction
Nowadays, with the intensification of global climate change and increasing frequency of human disturbances, global biodiversity is facing unprecedented pressure [1,2,3], and many taxa have shown declining diversity levels [4,5]. Butterflies, as one of the most sensitive indicators of environmental changes [6,7], are among the fastest-declining taxa globally. Long-term monitoring data from various regions consistently indicate that butterflies are undergoing population declines [8,9,10], the situation being even more severe in specialist species [11].
Butterfly decline is a complex ecological process driven by multiple factors, including natural enemies [12,13], climate change [14], and habitat degradation. As for monophagous or oligophagous butterfly species, host plant availability and limitation are among the key factors influencing population trends [15,16,17,18]. Primarily, two types of ecological pressure could be imposed by the availability of host plant in shaping larval dispersal and foraging behaviors: (1) density-dependent resource limitation, where larvae experience stress when the number of host plants per unit area is insufficient to support normal feeding; and (2) increased search costs due to greater distances between host plant patches. These two pressures jointly determine the spatial distribution patterns and local population occurrence in herbivorous butterflies [19,20]. Host plant limitation not only increases the difficulty and risk of oviposition site selection for females [21,22], but also forces larvae to disperse more frequently during development, thereby increasing their exposure to starvation stress and predation risk [15,23], ultimately potentially disrupting population establishment and persistence. Therefore, it is important to investigate the decline of monophagous butterflies by directly assessing the effects of starvation—resulting from host plant limitation—on individual fitness and population development.
L. chinensis is a monophagous butterfly endemic to China, with a narrow range primarily in mountainous and hilly areas of the Qinling Mountains and the middle to lower reaches of the Yangtze River [24,25]. Its larvae feed exclusively on the perennial herbs Asarum forbesii Maxim. and A. sieboldii Miq. [26,27]. The distribution and survival of L. chinensis populations are highly sensitive to host plant limitation, including both spatial configuration (e.g., patch distance) and quantitative characteristics (e.g., host plant abundance) of patches [23,28,29]. In particular, once larvae develop to the third instar, they abandon gregarious feeding behavior and begin to disperse individually into surrounding patches. During this stage, their success in locating food—as well as the associated time, energy expenditure, and risks (e.g., starvation, predation)—is largely determined by the spatial pattern of host plant patches, including patch size, spacing, and density [12,30]. For example, in Taohongling of Jiangxi Province, L. chinensis has only been recorded in two relatively large A. forbesii patches—Maoyingwo [23,31] and Taohongsha [29,32]—underscoring its strong dependence on large, continuous host plant patches. This dependence likely influences larval survival, as some individuals are forced to disperse over long distances due to low host plant encounter rates (e.g., low density or scattered distribution), thereby facing elevated starvation and predation risks that may reduce fitness (e.g., slower development) or result in mortality [28,29].
Importantly, the specific host plants of L. chinensis (A. forbesii and A. sieboldii) are of high medicinal value and have long been harvested for use in traditional Chinese medicine [33,34]. Overharvesting has led to severe fragmentation of host plant patches across many regions, causing drastic declines in plant abundance and even local extinctions [32,35,36]. Under such conditions, L. chinensis has likely experienced local extinctions [25], such as the population in Lushan, or drastic reductions in population size, placing its long-term persistence at risk [29]. Recognizing its high conservation value, L. chinensis was listed on the IUCN Red List as early as 1986 and designated as a Class II species in the Chinese National Key Protected Wild Animals List in 1989 [37]. Therefore, it is urgent to clarify the population dynamics of this rare butterfly under host plant limitation, identify strategies to mitigate its decline, and develop approaches for population recovery through host plant restoration. This study aims to investigate butterfly fitness (developmental duration, survival rate, reproductive capacity, etc.) under starvation stress during the dispersal larval stage (third to fifth instars), simulating the effects of host plant limitation on the occurrence of L. chinensis in Taohongling.
2. Materials and Methods
2.1. Insect Sources
In March 2021, mature eggs of L. chinensis were obtained from the Conservation Research Base of Taohongling, Jiangxi Province, China. The eggs were kept at ambient room temperature in Taohongling. Egg hatching was monitored daily. The newly hatched larvae were then individually transferred to transparent plastic containers for rearing and observation.
The Conservation Research Base is located at an elevation of 410 m, within the region of Taohongling Sika Deer National Nature Reserve. The reserve has a subtropical monsoon climate, with an average annual temperature of 16.5 °C, annual precipitation of 1,172 mm, and annual evaporation of 1,587.2 mm [38].
2.2. Larval Rearing and Starvation Treatments
In Taohongling, the butterfly larvae of L. chinensis feed exclusively on leaves of A. forbesii Therefore, fresh leaves with petioles were collected to rear larvae in experiments. The leaves were inserted into moist putty after being rinsed with clean water, and placed in transparent containers (17.2 cm in diameter, 13.2 cm in height), then the newly hatched larvae were transferred to leaves using a fine brush. The containers were placed at ambient room temperature. During rearing experiments, the frass was removed daily, and the containers were washed with tap water, disinfected with 75% ethanol, and air-dried before reuse.
Field observations find that the larvae of L. chinensis will leave the original leaves once molting into the third instar, which indicates the larvae shift to the actively-dispersal stage for searching more hostplant resource needed. However, the risk of starvation stress could increase if facing hostplant limitation. Therefore, to simulate this situation, the third-instar or older larvae were used in starvation experiments (SE). According to the method of Guo [39], newly molted larvae were returned to normal rearing after undergoing three days of starvation (without causing mortality). As such, there were three starvation treatments, the third-instar, the fourth-instar and the fifth-instar starvation groups, and the control group (CK, without starvation stress) in this study. Each group used 30 larval individuals. Once they ceased feeding and reached the pre-pupal stage, the fifth-instar larvae were transferred to rearing cages for pupation. And the pupae were labeled and kept in cool, dark conditions until emergence the following year. Throughout the rearing experiments, larval instars, survival status, and other parameters. were recorded daily, and the developmental duration and other life history parameters were calculated accordingly.
2.3. Adult Mating and Oviposition
In early March 2022, the overwintered pupae of the same group were transferred to rearing cages for adult emergence, which contained cotton balls soaked in honey water and a potted nectar plant (Viola philippica Cav.). The emergence time and sex of each butterfly were recorded daily, and a pair of newly emerged adults from the same treatment group were transferred to another mating cage, which contained cotton balls soaked in honey water, a potted nectar plant, as well as a potted host plant (A. forbesii). The mating time, individual labels, survival, egg production, and other parameters. were observed and recorded daily. During observation, water was periodically sprayed inside the cages to maintain appropriate environmental humidity.
2.4. Data analysis
2.4.1. Life Table Data Analysis
The age-stage, two-sex life table approach was used to analyze the raw life-history data for L. chinensis under control and starvation treatments [40] [41]. The age-stage-specific survival rate (sₓⱼ), defined as the probability that an individual of age x and stage j survives to that specific stage, was evaluated. The age-stage-specific fecundity (fₓⱼ), representing the daily number of eggs laid by an individual at age x and stage j, the age-specific fecundity (mₓ), and the age-specific survival rate (lₓ), referring to the probability that a newly laid egg survives to age x, were calculated accordingly [41,42,43].
The intrinsic rate of increase (r) was calculated based on the Euler–Lotka equation [44,45]. The finite rate of increase (λ) was computed as λ = eʳ [46]. The net reproductive rate (R₀) was estimated as [47]. The mean generation time (T), defined as the time required for the population to increase R₀-fold under a stable age-stage distribution, was calculated as T = ln(R₀)/r [48]. The gross reproductive rate (GRR) was calculated as GRR = ∑mₓ [43].
The raw life-history data were entered into Microsoft Excel 2013, and the TWOSEX-MSChart® software (available at http://140.120.197.173/Ecology/) was used for all life table analyses [49]. This program provides a simplified and standardized method for computing population parameters, avoiding the complexity of manual calculation. The means, standard errors, and variances of the population parameters were estimated using the bootstrap method (with 100,000 replications), which is integrated into the TWOSEX-MSChart® software [49]. All graphs were produced using Origin 2022 (OriginLab, Northampton, MA, USA).
2.4.2. Population Projection
To predict and compare the population growth and age-stage structure of L. chinensis under control and starvation treatments, we used life table data on developmental time, survival rate, and fecundity to simulate population growth using the TIMING-MSChart program [49] The data file for TIMING-MSChart was generated directly from the output file "15_For_TIMING.txt" produced by TWOSEX-MSChart, simplifying the data preparation process [50].
To assess stage-specific growth dynamics, the stage size at time t was calculated as log10(Nj, t + 1), and the daily growth rate of stage j from time t to t + 1 was computed asj, t =ln (Nj, t+1+1) – ln (Nj, t+ 1). This method avoids undefined values caused by zero individuals at a given time point and allows consistent comparisons across treatments [51]. Paired Wilcoxon rank-sum tests were used to assess statistical differences, with significance determined at P < 0.05.
3. Results and Analyses
3.1. Effects of Starvation on Developmental Duration and Survival
Starvation significantly influenced the larval developmental duration of L. chinensis. As shown in Table 1, The durations were extended by 32.4% (5th-instar starvation group) to 103.9% (3rd-instar starvation group) relative to the control group. Although subsequent instars of the starvation groups showed durations similar to or shorter than those of the control group. Overall, the larval period remained significantly prolonged, with the 3rd-instar group exhibiting the most pronounced delay. In contrast, Starvation significantly shortened pupal developmental duration (Table 1), indicating that the butterfly emergence date would occur earlier than that of non-starved individuals.
Starvation also significantly altered the age-stage-specific survival curve (lx) of L. chinensis. As shown in Figure 1, Both the starvation and control groups exhibited a Type III Deevey-type survival curve(lx), but the two curves differed significantly (P < 0.001). This discrepancy was mainly attributed to the pupal stage, during which the survival rate (sxj) in the starvation treatment was markedly lower than that of the control group.
3.2. Effects of Starvation on Adult Lifespan and Expectancy
The impact of starvation on adult lifespan varied between sexes. As shown in Figure 2, Female lifespan was not significantly affected by starvation, whereas male lifespan in the starvation treatment was significantly extended compared with the control. In contrast, the overall life expectancy (exj) did not differ significantly between groups (Figure 3). However, when assessed separately across developmental stages, the life expectancy of the larval stage exhibited a significant reduction in the starvation treatment, indicating negative effects on larval health and survival potential.
3.3. Effects of Starvation on Butterfly Fecundity
As illustrated in Figure 4, starvation significantly reduced the age-stage specific reproductive value (vx) of L. chinensis (P < 0.001). The age-specific fecundity (mx) of starvation treatment was markedly lower (mx = 0.7) than that of the control group (mx = 21). Likewise, the net reproductive rate (lxmx) declined from 3.0 in the control group to 2.7 of the starvation treatment. Additionally, females that experienced larval starvation laid approximately 40% fewer eggs than control females, indicating a substantial decline of the reproductive output.
3.4. Effects of Starvation on Butterfly Population Dynamics
Starvation had a profound impact on the population dynamics of L. chinensis. As shown in Table 2, several key population parameters declined significantly in the starvation treatment. The intrinsic rate of increase (rm) decreased by 48.4%, the net reproductive rate (R0) declined by 59.5%, and total fecundity (vx) was reduced by 53.2%. In contrast, the finite rate of increase (λ) and mean generation time (T) remained relatively stable between groups.
Based on the above population parameters, the butterfly population dynamic was projected for the future two years. As shown in Figure 5, the starvation treatment exhibited lower population growth, and the annual growth rate decreased from 1.7 in the first year to 1.3 in the second year. By the end of the second year, the predicted population size in the starvation treatment declined by more than 83% relative to the control group, suggesting a substantial risk of population decline under host-plant limitation.
4. Discussion
The larvae of the specialist butterfly L. chinensis exhibit a highly restricted diet, with different geographic populations feeding exclusively on either A. sieboldii or A. forbesii as their sole host plant [24,25]. Consequently, the occurrence and distribution patterns of this species in the wild are strongly dependent on the abundance and spatial distribution of host plants [25,32,52]. In areas where host resources are limited, adults may face oviposition constraints, while larvae are prone to starvation stress, potentially leading to greater population fluctuations and an elevated risk of local extinction [31,53,54]. In this study, we conducted a controlled starvation experiment to simulate population dynamics of L. chinensis under host plant limitation, as observed in wild populations such as those in Taohongling of Jiangxi Province. Our results showed that third- to fifth-instar larvae exposed to three days of starvation exhibited significantly prolonged developmental periods, reduced fecundity, and marked declines in multiple fitness-related parameters, including the intrinsic rate of increase (rm). Model projections further suggest that starvation stress could lead to a population decline exceeding 83% within the following two years. These findings provide direct empirical evidence supporting the hypothesis that host plant limitation drives butterfly population decline and underscore the critical importance of maintaining and restoring key habitat resources in the conservation of endangered butterflies [18,55].
The persistence of butterfly populations in the wild depends on two main categories of habitat resources: (1) consuming resources, such as host plants and nectar sources; and (2) utilities, including functional sites for courtship, mating, and pupation [18,55]. For the monophagous L. chinensis, the host plant A. sieboldii or A. forbesii are indispensable for population maintenance [25,32,52]. However, over the past decades, species of Asarum have suffered extensive harvesting and habitat destruction due to their medicinal value [32,35,36]. These perennial plants grow slowly, have strict habitat requirements (e.g., shaded, moist, fertile soils), and exhibit poor natural recovery, especially under adverse conditions such as climate warming and frequent extreme weather events [56]. As a result, remnant host plant resources remain scarce, placing many L. chinensis populations under chronic oviposition and foraging constraints. Unlike naturally occurring resource fluctuations, starvation stress driven by anthropogenic disturbance or long-term ecological change tends to be frequent and persistent, with potentially severe consequences for both individuals and populations [57].
During starvation, the balance between mass and energy flux through an animal becomes disrupted [57]. For insects such as butterflies, the imbalance of endogenous substances and energy caused by larval starvation may eventually lead to a reduction in individual fitness [58]. Generally, the response of insects such as butterflies to starvation stress is more complex. On one hand, larvae may prolong development and feeding periods to compensate for nutritional deficits and accumulate energy for metamorphosis. For example, larvae of Bicyclus anynana (Lepidoptera: Nymphalidae) significantly extend their developmental duration under dietary restriction to increase feeding time [59]. In the present study, L. chinensis larvae subjected to starvation similarly prolonged each instar, likely as an energy compensation strategy. On the other hand, the pupal period was shortened, leading to earlier adult emergence. This may reflect an energy reallocation mechanism, in which extending feeding stages enhances energy accumulation, while compressing non-feeding stages reduces mortality risk [60,61]. In addition, starvation markedly suppressed adult reproductive capacity. According to a common survival trade-off mechanism, insects tend to reduce reproductive investment under starvation in order to maintain somatic functions [62,63,64]. In L. chinensis, larval starvation significantly reduced female egg production, a pattern also reported in Pieris brassicae (Lepidoptera: Pieridae) and Hyphantria cunea (Lepidoptera: Arctiidae) [60,65]. Given that L. chinensis adults have limited feeding capacity, reproductive system development relies heavily on energy reserves acquired during the larval stage [66], suggesting that starvation may force individuals to prioritize basic survival over reproduction [64,67,68].
Host plant limitation–induced larval starvation not only reduces individual fitness but also constrains population growth potential [20,60]. In this study, the rₘ, net reproductive rate (R₀), and total fecundity (vₓ) of the starvation group were all significantly lower than those of the control group. Population modeling predicted that continued starvation could lead to a population decline exceeding 83% within two years. For endangered taxa such as L. chinensis, persistent population decline can exacerbate the effects of genetic drift and inbreeding depression, resulting in the progressive erosion of allelic diversity and a concomitant reduction in adaptive potential to environmental variability. According to the extinction vortex framework [69,70,71], these genetic processes interact synergistically with ecological stochasticity, demographic stochasticity, and maladaptive behavioral responses, creating self-reinforcing feedback loops that accelerate population decline toward extinction. For instance, Euphydryas aurinia (Lepidoptera: Nymphalidae) has experienced increased subpopulation instability and elevated extinction risk due to the loss of its larval host plant (Succisa pratensis) and habitat fragmentation [72], while western North American monarch butterflies (Danaus plexippus (Lepidoptera: Nymphalidae)) have declined from millions in the 1980s to fewer than 2,000 individuals in 2020 under the combined pressures of pesticide use, habitat loss, and climate change [73]. These cases highlight the universality and severity of multiple interacting factors accelerating butterfly population decline and further underscore the central importance of maintaining and restoring critical habitat resources in the conservation of endangered butterflies [74,75].
5. Conclusions
The survival of the endangered butterfly L. chinensis is tightly linked to the availability of its host plants (A. sieboldii or A. forbesii), making local populations highly vulnerable to host plant limitation. In the present study, we simulated such resource constraints in the wild by subjecting larvae to short-term starvation stress. Our results demonstrated that starvation not only impaired larval growth and development but also exerted delayed effects on subsequent life stages, including the pupal and adult phases. In particular, female fecundity was markedly reduced, leading to lower individual fitness and, consequently, a decline in population size. These findings provide novel evidence for the hypothesis that host plant limitation is a key driver of population decline in L. chinensis, contributing to the broader understanding of global butterfly declines [8,9,10]. Furthermore, they emphasize the necessity of conserving and restoring critical habitat resources as a core component of butterfly conservation programs, and highlight the applicability of the “resource-based habitat” concept to the protection of specialist species such as L. chinensis [11,55].
Author Contributions
Conceptualization, J.Z.; Methodology, W.Y.; Q.Z.; Software, W.Y., Y.Z., C.Y.; Validation, J.Z.; Formal analysis, J.Z.; Investigation, W.Y., W.W., J.Z.; Resources, W.Y., W.W., Q.Z., J.Z.; Data curation, W.Y.; Writing-original draft preparation, W.Y., C.Y., J.Z.; Writing-review and editing, W.Y., J.Z.; Visualization, W.Y.; Supervision, J.Z.; Project administration, J.Z.; Funding acquisition, J.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (32360272) and partly supported by “Science and Technology Innovation Project of Jiangxi Forestry Bureau (202233)” and “Jiangxi Provincial Key Laboratory of Conservation Biology(No. 2023SSY02081)” project.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
We are grateful to the Administration Bureau of Jiangxi Taohongling Sika Deer National Nature Reserve for their invaluable assistance and support during the study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Change on the Future of Biodiversity. Ecology Letters 2012, 15, 365–377. [CrossRef] [PubMed]
- Román-Palacios, C.; Wiens, J.J. Recent Responses to Climate Change Reveal the Drivers of Species Extinction and Survival. Proceedings of the National Academy of Sciences 2020, 117, 4211–4217. [Google Scholar] [CrossRef]
- Hung, K.-L.J.; Sandoval, S.S.; Ascher, J.S.; Holway, D.A. Joint Impacts of Drought and Habitat Fragmentation on Native Bee Assemblages in a California Biodiversity Hotspot. Insects 2021, 12, 135. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Further Evidence for a Global Decline of the Entomofauna. Austral Entomology 2021, 60, 9–26. [Google Scholar] [CrossRef]
- Finn, C.; Grattarola, F.; Pincheira-Donoso, D. More Losers than Winners: Investigating Anthropocene Defaunation through the Diversity of Population Trends. Biological Reviews 2023, 98, 1732–1748. [Google Scholar] [CrossRef]
- Hill, G.M.; Kawahara, A.Y.; Daniels, J.C.; Bateman, C.C.; Scheffers, B.R. Climate Change Effects on Animal Ecology: Butterflies and Moths as a Case Study. Biological Reviews 2021, 96, 2113–2126. [Google Scholar] [CrossRef]
- Chowdhury, S.; Dubey, V.K.; Choudhury, S.; Das, A.; Jeengar, D.; Sujatha, B.; Kumar, A.; Kumar, N.; Semwal, A.; Kumar, V. Insects as Bioindicator: A Hidden Gem for Environmental Monitoring. Front. Environ. Sci. 2023, 11. [Google Scholar] [CrossRef]
- Warren, M.S.; Maes, D.; van Swaay, C.A.M.; Goffart, P.; Van Dyck, H.; Bourn, N.A.D.; Wynhoff, I.; Hoare, D.; Ellis, S. The Decline of Butterflies in Europe: Problems, Significance, and Possible Solutions. Proceedings of the National Academy of Sciences 2021, 118, e2002551117. [Google Scholar] [CrossRef]
- Navarro-Cano, J.A.; Karlsson, B.; Posledovich, D.; Toftegaard, T.; Wiklund, C.; Ehrlén, J.; Gotthard, K. Climate Change, Phenology, and Butterfly Host Plant Utilization. AMBIO 2015, 44, 78–88. [Google Scholar] [CrossRef]
- Herremans, M.; Gielen, K.; Van Kerckhoven, J.; Vanormelingen, P.; Veraghtert, W.; Swinnen, K.R.R.; Maes, D. Abundant Citizen Science Data Reveal That the Peacock Butterfly Aglais Io Recently Became Bivoltine in Belgium. Insects 2021, 12, 683. [Google Scholar] [CrossRef]
- Ellis, S.; Bourn, N.; Buiman, C. Landscape-Scale Conservation for Butterflies and Moths: Lessons from the UK; Butterfly Conservation: Wareham, 2012. [Google Scholar]
- Marini, L.; Zalucki, M.P. Density-Dependence in the Declining Population of the Monarch Butterfly. Sci Rep 2017, 7, 13957. [Google Scholar] [CrossRef] [PubMed]
- Ubach, A.; Páramo, F.; Prohom, M.; Stefanescu, C. Weather and Butterfly Responses: A Framework for Understanding Population Dynamics in Terms of Species’ Life-Cycles and Extreme Climatic Events. Oecologia 2022, 199, 427–439. [Google Scholar] [CrossRef] [PubMed]
- Hunter, A.F.; Lindgren, B.S. Range of Gypsy Moth in British Columbia: A Study of Climatic Suitability. Journal of the Entomological Society of British Columbia 1995, 92, 45–56. [Google Scholar]
- Varela, L.G.; Bernays, E.A. Behavior of Newly Hatched Potato Tuber Moth Larvae, Phthorimaea Operculella Zell. (Lepidoptera: Gelechiidae), in Relation to Their Host Plants. J Insect Behav 1988, 1, 261–275. [Google Scholar] [CrossRef]
- Hanspach, J.; Schweiger, O.; Kühn, I.; Plattner, M.; Pearman, P.B.; Zimmermann, N.E.; Settele, J. Host Plant Availability Potentially Limits Butterfly Distributions under Cold Environmental Conditions. Ecography 2014, 37, 301–308. [Google Scholar] [CrossRef]
- Curtis, R.J.; Brereton, T.M.; Dennis, R.L.H.; Carbone, C.; Isaac, N.J.B. Butterfly Abundance Is Determined by Food Availability and Is Mediated by Species Traits. Journal of Applied Ecology 2015, 52, 1676–1684. [Google Scholar] [CrossRef]
- Dennis, R.L.H.; Shreeve, T.G.; Arnold, H.R.; Roy, D.B. Does Diet Breadth Control Herbivorous Insect Distribution Size? Life History and Resource Outlets for Specialist Butterflies. J Insect Conserv 2005, 9, 187–200. [Google Scholar] [CrossRef]
- Gotthard, K.; Nylin, S.; Wiklund, C. Adaptive Variation in Growth Rate: Life History Costs and Consequences in the Speckled Wood Butterfly, Pararge Aegeria. Oecologia 1994, 99, 281–289. [Google Scholar] [CrossRef]
- Crone, E.E.; Schultz, C.B. Host Plant Limitation of Butterflies in Highly Fragmented Landscapes. Theor Ecol 2022, 15, 165–175. [Google Scholar] [CrossRef]
- Jones, L.C. Insects Allocate Eggs Adaptively According to Plant Age, Stress, Disease or Damage. Proceedings of the Royal Society B: Biological Sciences 2022, 289, 20220831. [Google Scholar] [CrossRef]
- Rausher, M.D.; Papaj, D.R. Demographic Consequences of Descrimination among Conspecific Host Plants by Battus Philenor Butterflies. Ecology 1983, 64, 1402–1410. [Google Scholar] [CrossRef]
- Chen, L. Population Ecology of Luehdorfia chinensis in Taohongling, Jiangxi. Master’s Thesis, Jiangxi Agricultural University, Nanchang, China, 2023. [Google Scholar]
- Xu, X.L. A Discussion on the Distribution Boundaries of Luehdorfia chinensis. Agriculture and Technology 2014, 34, 254. [Google Scholar]
- Yuan, D.C.; Mai, G.Q.; Xue D., Y.; Hu, C.; Ye, G.Y. The Habitat Biology and Conservation Status of Luehdorfia chinensis (Lepidoptera: Papilionidae). Chinese Biodiversity 1998, 26–36. [Google Scholar] [CrossRef]
- Hu, C.; Wu, X.J.; Wang, X.M. The Biology of Luehdorfia Chinensis Leech, A Rare and Endangered Butterfly. Acta Entomologica Sinica 1992, 195–199. [Google Scholar] [CrossRef]
- Li, C.L. The Early Stages of Chinese Rhopalocera— Luehdorfia chinensis Leech (Parnassiidae: Zerynthiinae). Acta Entomologica Sinica 1978, 161, 239. [Google Scholar] [CrossRef]
- Chen, L. Wang L.; Yang W.J.; Wu W. G.; Liu X. H.; Zhang Y.; Zeng J. P. Traits Variability of Asarum forbesii and Conservation Implications to the Rare Butterfly of Luehdorfia chinensis in Taohongling, South China. Acta Agriculturae Universitatis Jiangxiensis 2022, 44, 1122–1134. [Google Scholar] [CrossRef]
- Zou, M.H. Limitation Effect and Growth Conditions of the Hostplant Asarum Forbesii on the Butterfly Population of Luehdorfia chinensis in Taohongling. Master’s Thesis, Jiangxi Agricultural University, Nanchang, China 2025. [Google Scholar]
- Zalucki, M.P.; Lammers, J.H. Dispersal and Egg Shortfall in Monarch Butterflies: What Happens When the Matrix Is Cleaned Up?
- He, G.Q.; Jia F., H.; Zhu H., B. The Species Distribution and Quantity Surveying of Luehdorfia Chinese (Leech) in JiangXi. Journal of Jiangxi University of Tcm 2011, 23, 75–76. [Google Scholar]
- Su, J. Geographical Distributions, Environmental Niches and Conservation in the Rare Butterflies of Luehdorfia Spp. Master’s Thesis, Jiangxi Agricultural University, Nanchang, China, 2020. [Google Scholar]
- Wang D. Q.; Huang S. H. Medicinal Plants of Asarumin Anhui Province. China Journal of Chinese Materia Medica 1989, 6–8, 61.
- Nie, A.Z.; Bian, M.; Zhu, C.S.; Gao, M.M. Mechanism of Asari Radix et Rhizoma water extract induced liver injury based on proteomics. Chinese Traditional and Herbal Drugs 2024, 55, 5145–5153. [Google Scholar]
- Jeong, H.J.; Kim, J.G. Small-Scale Spatial Genetic Structure of Asarum Sieboldii Metapopulation in a Valley. j ecology environ 2021, 45, 11. [Google Scholar] [CrossRef]
- Yang, Z.L. The Geographical Distribution of Asarum (Aristolochiaceae) from Sichuan Province in China. Guihaia 1988, 83–88. [Google Scholar]
- Jonathan, B.; Craig, H.-T. Red List of Threatened Species: A Global Species Assessment; IUCN-The World Conservation Union, 2004.
- Taohongling Sika Deer Reserve Jiangxi Taohongling Meihualu Baohuqu; China Forestry Publishing House: Beijing, 2000.
- Guo, H.; Jia, N.; Chen, H.; Xie, D.; Chi, D. Preliminary Analysis of Transcriptome Response of Dioryctria Sylvestrella (Lepidoptera: Pyralidae) Larvae Infected with Beauveria Bassiana under Short-Term Starvation. Insects 2023, 14, 409. [Google Scholar] [CrossRef]
- Chi, H.; Liu, H. Two New Methods for the Study of Insect Population Ecology. Bull. Inst. Zool., Acad. Sin 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-Table Analysis Incorporating Both Sexes and Variable Development Rates among Individuals. Environmental Entomology 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Iranipour, S.; Mahmoodi Arabi, S.; Michaud, J.P. Does the Two-Sex Life Table for Sexual Populations Invalidate Those Based Solely on Female Cohorts? Ann Entomol Soc Am 2025, 118, 189–205. [Google Scholar] [CrossRef]
- Zhu, Y.; Qi, F.; Tan, X.; Zhang, T.; Teng, Z.; Fan, Y.; Wan, F.; Zhou, H. Use of Age-Stage, Two-Sex Life Table to Compare the Fitness of Bactrocera Dorsalis (Diptera: Tephritidae) on Northern and Southern Host Fruits in China. Insects 2022, 13, 258. [Google Scholar] [CrossRef]
- Abbes, K.; Harbi, A.; Guerrieri, E.; Chermiti, B. Using Age-Stage Two-Sex Life Tables to Assess the Suitability of Three Solanaceous Host Plants for the Invasive Cotton Mealybug Phenacoccus Solenopsis Tinsley. Plants 2024, 13, 1381. [Google Scholar] [CrossRef] [PubMed]
- Rismayani, *!!! REPLACE !!!*; Ullah, M.S.; Chi, H.; Gotoh, T. Impact of Constant and Fluctuating Temperatures on Population Characteristics of Tetranychus Pacificus (Acari: Tetranychidae). J Econ Entomol 2021, 114, 638–651. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Bashir, N.H.; Naeem, M.; Tian, R.; Tian, X.; Chen, H. Age-Stage, Two-Sex Life Table of Atractomorpha Lata (Orthoptera: Pyrgomorphidae) at Different Temperatures. Insects 2024, 15, 493. [Google Scholar] [CrossRef] [PubMed]
- El Aalaoui, M.; Sbaghi, M.; Mokrini, F. Effect of Temperature on the Development and Reproduction of Olive Psyllid Euphyllura Olivina Costa (Hemiptera: Psyllidae). Crop Protection 2025, 190, 107131. [Google Scholar] [CrossRef]
- Bankar, D.R.; Bhamare, V.K. Comparative Biology, Life Tables and Intrinsic Rate of Increase of Spodoptera Frugiperda (J.E. Smith) Reared on Pearl Millet and Sugarcane. Journal of Entomological Research 2023, 47, 866–870. [Google Scholar] [CrossRef]
- Chi, H. TIMING-MSChart: A Computer Program for the Population Projection Based on Age-Stage, Two-Sex Life Table Available online:. Available online: http://140.120.197.173/Ecology/Download/Timing-MSChart.rar (accessed on 20 January 2020).
- Chi, H. TWOSEX-MSChart: A Computer Program for Age Stage, Two-Sex Life Table Analysis, National Chung Hsing University: Taichung, Taiwan, 2018.
- Lehmann, P.; Ammunét, T.; Barton, M.; Battisti, A.; Eigenbrode, S.D.; Jepsen, J.U.; Kalinkat, G.; Neuvonen, S.; Niemelä, P.; Terblanche, J.S.; et al. Complex Responses of Global Insect Pests to Climate Warming. Frontiers in Ecology and the Environment 2020, 18, 141–150. [Google Scholar] [CrossRef]
- Chen, A.; Liu, B.; Zhou, R.; Zhang, H.; Zhou, L.; Xie, X.; Zhuo, Z.; Xu, D. Habitat Suitability Analysis for Luehdorfia Chinensis Leech, 1893 (Lepidoptera: Papilionidae) in the Middle and Lower Yangtze River: A Study Based on the MaxEnt Model. Insects 2025, 16, 396. [Google Scholar] [CrossRef]
- Wang, R.P.; Li, L. The Extinction Vortex of Small Population. Journal of Bology 2008, 25, 14–16. [Google Scholar]
- Bin, W.; Weiping, W.; Haihua, W.; Gang, H. A Retrospective Analysis on the Population Viability of the Yangtze River Dolphin or Baiji (Lipotes Vexillifer). Indian Journal of Animal Research 2022, 56(6): 775-779.
- Dennis, R.L.H.; Shreeve, T.G.; Van Dyck, H. Towards a Functional Resource-Based Concept for Habitat: A Butterfly Biology Viewpoint. Oikos 2003, 102, 417–426. [Google Scholar] [CrossRef]
- Iutzi, I.; Crews, T.; Crews, M. Perennializing Grain Crop Agriculture: A Pathway. The Land 2020, 823–5376. [Google Scholar]
- McCue, M.D. Starvation Physiology: Reviewing the Different Strategies Animals Use to Survive a Common Challenge. Comp Biochem Physiol A Mol Integr Physiol 2010, 156, 1–18. [Google Scholar] [CrossRef] [PubMed]
- McKay, A.F.; Ezenwa, V.O.; Altizer, S. Consequences of Food Restriction for Immune Defense, Parasite Infection, and Fitness in Monarch Butterflies. Physiological and Biochemical Zoology 2016. [Google Scholar] [CrossRef] [PubMed]
- Bauerfeind, S.S.; Fischer, K. Effects of Food Stress and Density in Different Life Stages on Reproduction in a Butterfly. Oikos 2005, 111, 514–524. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, L.; Zheng, J.; Li, T.; Zhao, L. Effects of Larval Starvation Stress on the Life History and Adult Fitness of Fall Webworm, Hyphantria Cunea. Insects 2025, 16, 410. [Google Scholar] [CrossRef]
- Brueggemann, L.; Singh, P.; Müller, C. Life Stage- and Sex-Specific Sensitivity to Nutritional Stress in a Holometabolous Insect. Ecology and Evolution 2025, 15, e70764. [Google Scholar] [CrossRef]
- Elkin, C.M.; Reid, M.L. Low Energy Reserves and Energy Allocation Decisions Affect Reproduction by Mountain Pine Beetles, Dendroctonus Ponderosae. Functional Ecology 2005, 19, 102–109. [Google Scholar] [CrossRef]
- García-Roger, E.M.; Martínez, A.; Serra, M. Starvation Tolerance of Rotifers Produced from Parthenogenetic Eggs and from Diapausing Eggs: A Life Table Approach.
- Billings, A.C.; Schultz, K.E.; Hernandez, E.A.; Jones, W.E.; Price, D.K. Male Courtship Behaviors and Female Choice Reduced during Experimental Starvation Stress. Behav Ecol 2019, 30, 231–239. [Google Scholar] [CrossRef]
- Gols, R.; Croijmans, L.; Dicke, M.; van Loon, J.J.A.; Harvey, J.A. Plant Quantity Affects Development and Reproduction of a Gregarious Butterfly More than Plant Quality. Entomologia Experimentalis et Applicata 2022, 170, 646–655. [Google Scholar] [CrossRef]
- Zhao, X.; Geng, Y.; Hu, T.; Xie, C.; Xu, W.; Zuo, Z.; Xue, M.; Hao, D. Ecological Strategies of Hyphantria Cunea (Lepidoptera: Arctiidae) Response to Different Larval Densities. Front. Ecol. Evol. 2023, 11. [Google Scholar] [CrossRef]
- Zhang, Z.-J.; Zhang, S.-S.; Niu, B.-L.; Ji, D.-F.; Liu, X.-J.; Li, M.-W.; Bai, H.; Palli, S.R.; Wang, C.-Z.; Tan, A.-J. A Determining Factor for Insect Feeding Preference in the Silkworm, Bombyx Mori. PLOS Biology 2019, 17, e3000162. [Google Scholar] [CrossRef]
- Boggs, C.L. Understanding Insect Life Histories and Senescence through a Resource Allocation Lens. Functional Ecology 2009, 23, 27–37. [Google Scholar] [CrossRef]
- Fagan, W.F.; Holmes, E.E. Quantifying the Extinction Vortex. Ecology Letters 2006, 9, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Gilpin, M.E.; Soule, M.E. Minimum Viable Populations: Processes of Extinction. In Conservation Biology: The Science of Scarcity and Diversity (Ed. Soule´, M.E.). In; Sinauer Associates, MA: Sunderland, 1986; pp. 19–34. [Google Scholar]
- Nordstrom, S.W.; Hufbauer, R.A.; Olazcuaga, L.; Durkee, L.F.; Melbourne, B.A. How Density Dependence, Genetic Erosion and the Extinction Vortex Impact Evolutionary Rescue. Proceedings of the Royal Society B 2023. [Google Scholar] [CrossRef] [PubMed]
- Brunbjerg, A.K.; Høye, T.T.; Eskildsen, A.; Nygaard, B.; Damgaard, C.F.; Ejrnæs, R. The Collapse of Marsh Fritillary (Euphydryas Aurinia) Populations Associated with Declining Host Plant Abundance. Biological Conservation 2017, 211, 117–124. [Google Scholar] [CrossRef]
- James, D.G. Monarch Butterflies in Western North America: A Holistic Review of Population Trends, Ecology, Stressors, Resilience and Adaptation. Insects 2024, 15, 40. [Google Scholar] [CrossRef] [PubMed]
- Lukens, L.; Thieme, J.; Thogmartin, W.E. Milkweed and Floral Resource Availability for Monarch Butterflies (Danaus Plexippus) in the United States. Front. Ecol. Evol. 2024, 12. [Google Scholar] [CrossRef]
- Kukkonen, J.M.; von Numers, M.; Brommer, J.E. Conserving Apollo Butterflies: Habitat Characteristics and Conservation Implications in Southwest Finland. J Insect Conserv 2024, 28, 1199–1210. [Google Scholar] [CrossRef]
Figure 1.
Comparison of age-specific survival rate (lx) and age-stage specific survival rate (Sxj) between control check (CK) and starvation treatments (ST) in L. chinensis.
Figure 1.
Comparison of age-specific survival rate (lx) and age-stage specific survival rate (Sxj) between control check (CK) and starvation treatments (ST) in L. chinensis.
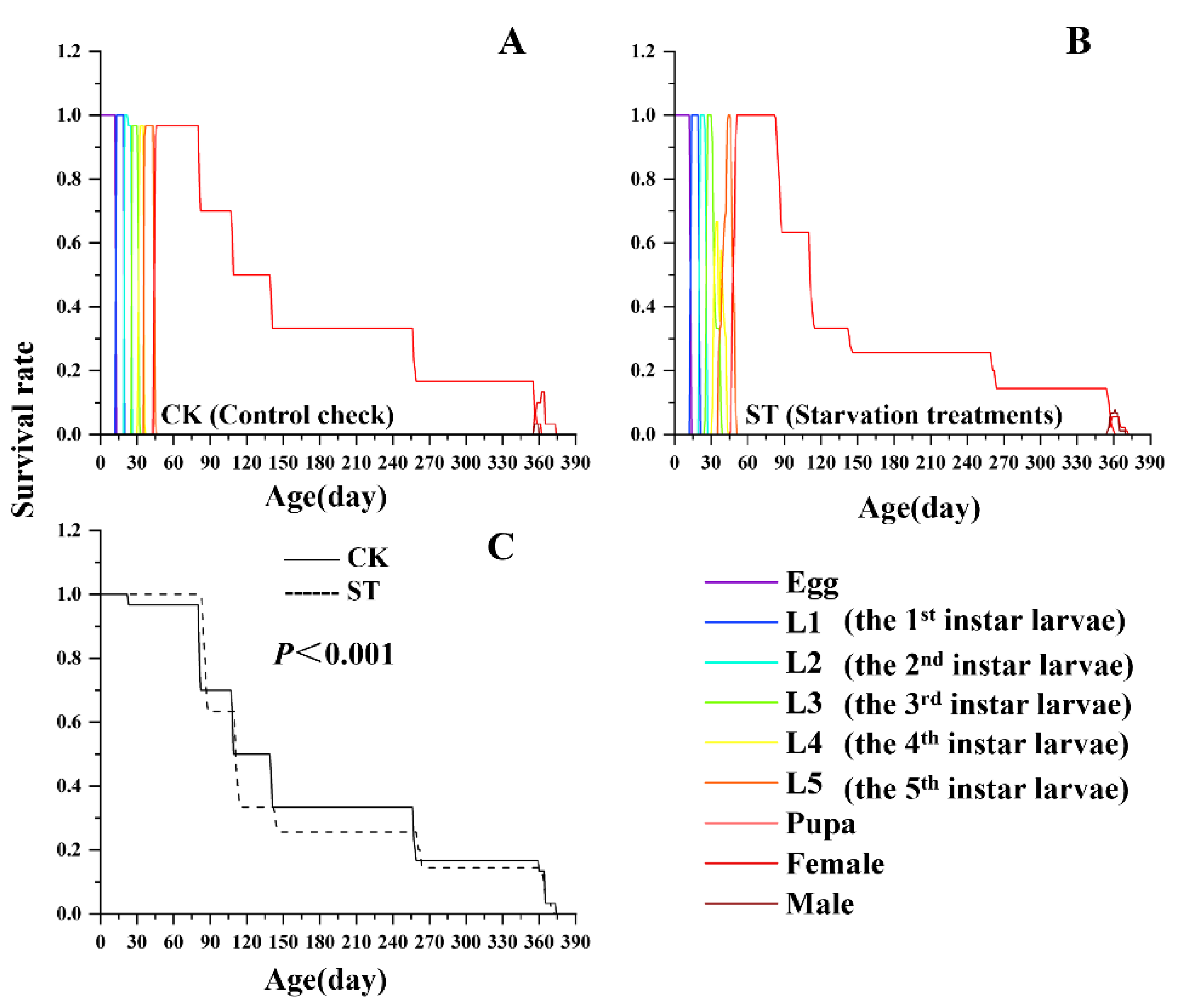
Figure 2.
Effects of starvation on the fecundity and longevity of L. chinensis.
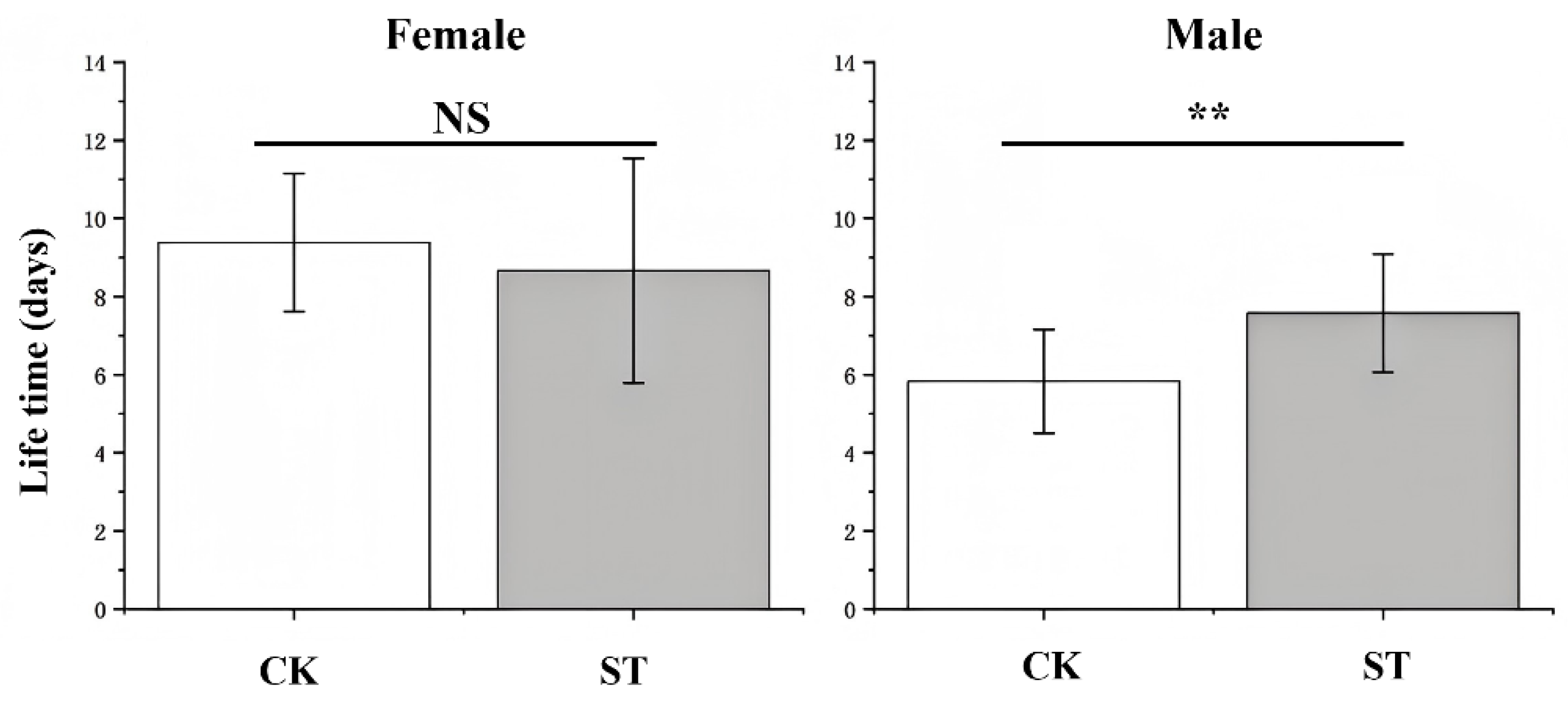
Figure 3.
Comparison of age-stage specific life expectancy(exj) between control check (CK) and starvation treatments (ST) in L. chinensis.
Figure 3.
Comparison of age-stage specific life expectancy(exj) between control check (CK) and starvation treatments (ST) in L. chinensis.
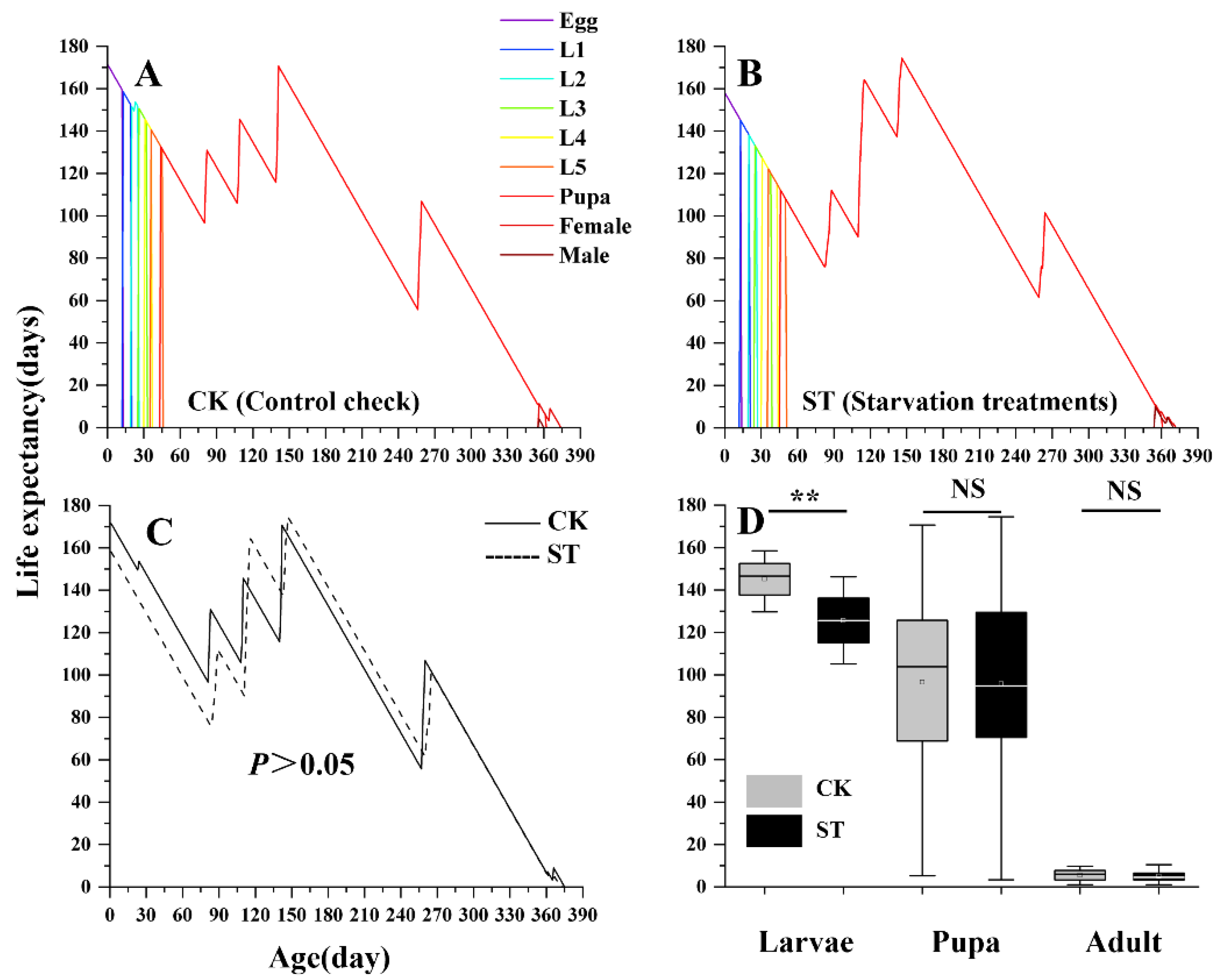
Figure 4.
Comparison of age-specific fecundity(mx,lxmx) (A, B) and age-stage specific reproductive value(vx)(C) between control check (CK) and starvation treatments (ST) in L. chinensis.
Figure 4.
Comparison of age-specific fecundity(mx,lxmx) (A, B) and age-stage specific reproductive value(vx)(C) between control check (CK) and starvation treatments (ST) in L. chinensis.
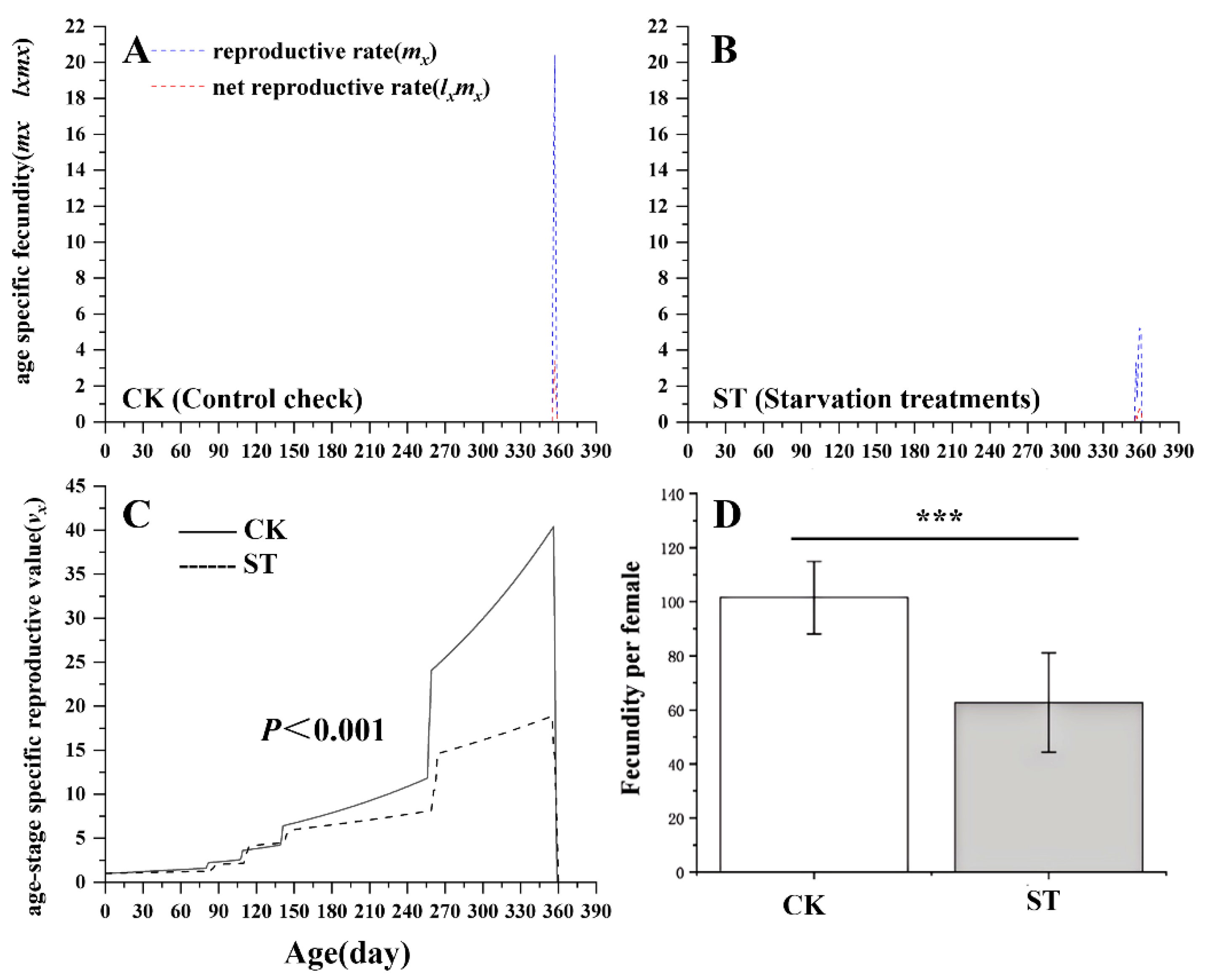
Figure 5.
Comparison of stage size (A, B) and growth rate dynamics (C, D) between control check (CK) and starvation treatments (ST) of L. chinensis.
Figure 5.
Comparison of stage size (A, B) and growth rate dynamics (C, D) between control check (CK) and starvation treatments (ST) of L. chinensis.
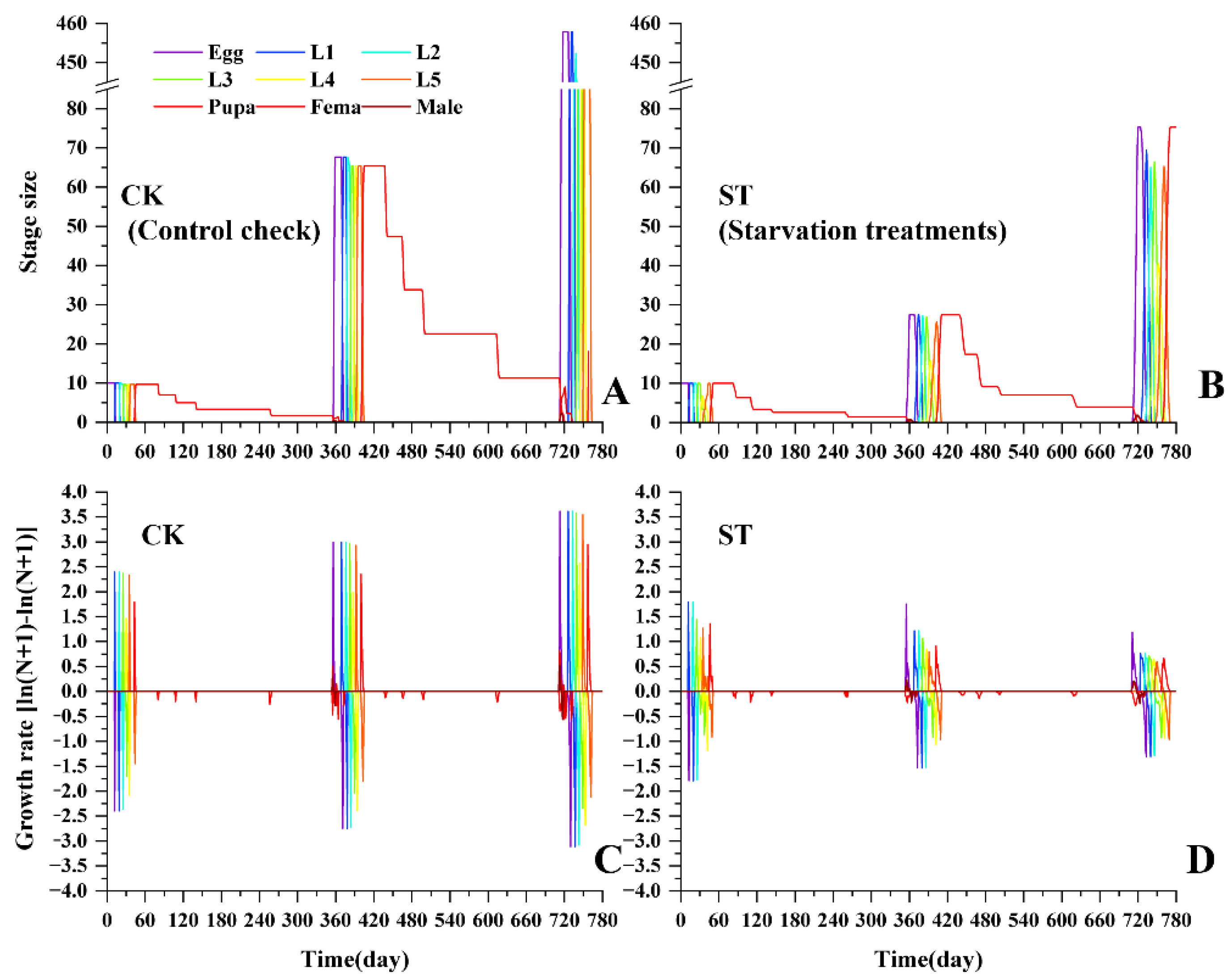

Table 1.
Effects of starvation on the developmental duration (Mean ± SD) of immature stages in L. chinensis.
Table 1.
Effects of starvation on the developmental duration (Mean ± SD) of immature stages in L. chinensis.
| Stages | CK(n) | Starvation treatments (ST) | ||
|---|---|---|---|---|
| 3rd(n) | 4th(n) | 5th(n) | ||
| egg | 13.00±0.00a (30) | 13.97±0.18a (30) | 13.33±0.48a (30) | 13.20±0.41a (30) |
| 1st instar larvae | 7.00±0.00a (30) | 7.00±0.00a (30) | 7.00±0.00a (30) | 7.00±0.00a (30) |
| 2nd instar larvae | 6.00±0.00a (30) | 6.00±0.00a (30) | 5.83±0.38a (30) | 6.00±0.00a (30) |
| 3rd instar larvae | 5.69±0.10b (30) | 11.60±0.40a (30) | 5.47±0.51b (30) | 5.73±0.57b (30) |
| 4th instar larvae | 4.34±0.09b (30) | 4.77±0.43b (30) | 7.03±0.18a (30) | 4.33±0.48b (30) |
| 5th instar larvae | 8.48±0.09b (30) | 7.20±0.41c (30) | 8.37±0.49b (30) | 11.23±0.41a (30) |
| Larval duration | 31.37±0.76c (30) | 35.93±0.64a (30) | 33.77±0.90b (30) | 33.97±0.72b (30) |
| Pupa | 313.2±0.97a (14) | 308.14±0.71b (6) | 309.00±0.00b (2) | 308.75±0.85b (5) |
Note: Data in the table are compared horizontally, with different letters representing differences between groups at a significance level of P < 0.05.
Table 2.
Comparison of population parameters (Mean ± SD) between control check (CK) and starvation treatments (ST) in L. chinensis.
Table 2.
Comparison of population parameters (Mean ± SD) between control check (CK) and starvation treatments (ST) in L. chinensis.
| Population parameters | CK (n=30) | ST (n=90) | Total (n=120) |
|---|---|---|---|
| Intrinsic rate of increase(rm) | 0.0064±0.0026a | 0.0033±0.0016b | 0.0032±0.0013b |
| Net reproductive rate(R0) | 6.77±4.72a | 2.74±1.4b | 2.83±1.19b |
| Finite rate of increase(λ) | 1.0064±0.0027a | 1.0033 ±0.0017a | 1.0032±0.0013a |
| Total fecundity(vx) | 40.6±27.53a | 19±8.92b | 25±9.42b |
| Mean generation time(T) | 357.87±0.69a | 358.66±0.84a | 356.64±1.03a |
Note: Data in the table are compared horizontally, with different letters representing differences between groups at a significance level of P<0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



